doi:10.1152/ajpcell.00287.2002 284:1362-1373, 2003. First published Jan 8, 2003; Am J Physiol Cell Physiol Henry Lambert Niels Ørtenblad, Jette Feveile Young, Niels Oksbjerg, Jacob Holm Nielsen and Ian taurine release from skeletal muscle cells Reactive oxygen species are important mediators of You might find this additional information useful... 43 articles, 16 of which you can access free at: This article cites http://ajpcell.physiology.org/cgi/content/full/284/6/C1362#BIBL 3 other HighWire hosted articles: This article has been cited by [PDF] [Full Text] [Abstract] , August 1, 2004; 97 (2): 661-668. J Appl Physiol L. Zuo, F. L. Christofi, V. P. Wright, S. Bao and T. L. Clanton Lipoxygenase-dependent superoxide release in skeletal muscle [PDF] [Full Text] [Abstract] , May 1, 2005; 98 (5): 1753-1760. J Appl Physiol V. P. Wright, P. F. Klawitter, D. F. Iscru, A. J. Merola and T. L. Clanton diaphragm Superoxide scavengers augment contractile but not energetic responses to hypoxia in rat [PDF] [Full Text] [Abstract] , September 1, 2005; 90 (5): 703-714. Exp Physiol A. Fredsted, U. R. Mikkelsen, H. Gissel and T. Clausen longus muscle Anoxia induces Ca2+ influx and loss of cell membrane integrity in rat extensor digitorum including high-resolution figures, can be found at: Updated information and services http://ajpcell.physiology.org/cgi/content/full/284/6/C1362 can be found at: AJP - Cell Physiology about Additional material and information http://www.the-aps.org/publications/ajpcell This information is current as of October 23, 2006 . http://www.the-aps.org/. American Physiological Society. ISSN: 0363-6143, ESSN: 1522-1563. Visit our website at a year (monthly) by the American Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. Copyright © 2005 by the is dedicated to innovative approaches to the study of cell and molecular physiology. It is published 12 times AJP - Cell Physiology on October 23, 2006 ajpcell.physiology.org Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

doi:10.1152/ajpcell.00287.2002 284:1362-1373, 2003. First published Jan 8, 2003;Am J Physiol Cell Physiol
Henry Lambert Niels Ørtenblad, Jette Feveile Young, Niels Oksbjerg, Jacob Holm Nielsen and Iantaurine release from skeletal muscle cells Reactive oxygen species are important mediators of
You might find this additional information useful...
43 articles, 16 of which you can access free at: This article cites http://ajpcell.physiology.org/cgi/content/full/284/6/C1362#BIBL
3 other HighWire hosted articles: This article has been cited by
[PDF] [Full Text] [Abstract]
, August 1, 2004; 97 (2): 661-668. J Appl PhysiolL. Zuo, F. L. Christofi, V. P. Wright, S. Bao and T. L. Clanton
Lipoxygenase-dependent superoxide release in skeletal muscle
[PDF] [Full Text] [Abstract], May 1, 2005; 98 (5): 1753-1760. J Appl Physiol
V. P. Wright, P. F. Klawitter, D. F. Iscru, A. J. Merola and T. L. Clanton diaphragm
Superoxide scavengers augment contractile but not energetic responses to hypoxia in rat
[PDF] [Full Text] [Abstract], September 1, 2005; 90 (5): 703-714. Exp Physiol
A. Fredsted, U. R. Mikkelsen, H. Gissel and T. Clausen longus muscle
Anoxia induces Ca2+ influx and loss of cell membrane integrity in rat extensor digitorum
including high-resolution figures, can be found at: Updated information and services http://ajpcell.physiology.org/cgi/content/full/284/6/C1362
can be found at: AJP - Cell Physiologyabout Additional material and information http://www.the-aps.org/publications/ajpcell
This information is current as of October 23, 2006 .
http://www.the-aps.org/.American Physiological Society. ISSN: 0363-6143, ESSN: 1522-1563. Visit our website at a year (monthly) by the American Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. Copyright © 2005 by the
is dedicated to innovative approaches to the study of cell and molecular physiology. It is published 12 timesAJP - Cell Physiology
on October 23, 2006
ajpcell.physiology.orgD
ownloaded from

Reactive oxygen species are important mediators oftaurine release from skeletal muscle cells
Niels Ørtenblad,1 Jette Feveile Young,1 Niels Oksbjerg,1
Jacob Holm Nielsen,1 and Ian Henry Lambert2
1Danish Institute of Agricultural Sciences, Research Center Foulum, DK-8830, Tjele; and2The August Krogh Institute, Biochemical Department, DK-2100 Copenhagen Ø, Denmark
Submitted 12 June 2002; accepted in final form 3 January 2003
Ørtenblad, Niels, Jette Feveile Young, Niels Oksb-jerg, Jacob Holm Nielsen, and Ian Henry Lambert.Reactive oxygen species are important mediators of taurinerelease from skeletal muscle cells. Am J Physiol Cell Physiol284: C1362–C1373, 2003. First published January 8, 2003;10.1152/ajpcell.00287.2002.—The present study illustrateselements of the signal cascades involved in the activation oftaurine efflux pathways in myotubes derived from skeletalmuscle cells. Exposing primary skeletal muscle cells, loadedwith 14C-taurine, to 1) hypotonic media, 2) the phospholipaseA2 (PLA2) activator melittin, 3) anoxia, or 4) lysophosphati-dyl choline (LPC) causes an increase in 14C-taurine releaseand a concomitant production of reactive oxygen species(ROS). The antioxidants butulated hydroxy toluene and vi-tamin E inhibit the taurine efflux after cell swelling, anoxia,and addition of LPC. The muscle cells possess two separatetaurine efflux pathways, i.e., a swelling- and melittin-in-duced pathway that requires 5-lipoxygenase activity for ac-tivation and a LPC-induced pathway. The two pathways aredistinguished by their opposing sensitivity toward the anionchannel blocker DIDS and cholesterol. These data provideevidence for PLA2 products and ROS as key mediators of thesignal cascade leading to taurine efflux in muscle.
C2C12; calcium; cell volume regulation; 5-lipoxygenase;melittin; anoxia; secretory phospholipase A2
THE ABILITY TO REGULATE cell volume is a crucial constit-uent of mammalian cell homeostasis and has an im-portant impact on cell function (metabolism, secretion)and cell fate (proliferation, apoptosis) (22). Cells ex-posed to hypotonic solutions respond by rapid cellswelling due to a high water permeability, followed bya regulatory volume decrease due to net loss of intra-cellular inorganic ions and organic osmolytes, e.g., freeamino acids, polyols, and methylamines. Taurine, asulphur-containing amino acid, is abundant in mostmammalian cells, and the cellular taurine concentra-tion in, e.g., skeletal muscle is reported to vary be-tween 10 and 60 mM, which is �500 times higher thanthe taurine concentration in plasma (1, 4). The cellulartaurine concentration is kept high by a Na�-dependenttaurine transporter, which is highly expressed in skel-etal muscle (37). Osmotic perturbation, as well as hor-
monal stimulation and pathological factors such asischemia, have been shown to lead to the release oftaurine from various cells, including cardiac myocytes(15), brain neurons (25, 38), and skeletal muscle (19).In skeletal muscle, there is considerable evidence thatcontractile activity leads to loss of taurine from themuscle cell (36) and increase in plasma taurine (4).Furthermore, we have recently demonstrated that ad-dition of the phospholipase A2 (PLA2) product lyso-phosphatidyl choline (LPC) leads to taurine loss frommuscle cells (20). Although taurine is an importantconstituent in the volume regulatory mechanism inmuscle, it has also been shown to modulate varioussteps in the excitation-contraction coupling (1) andpromote cellular membrane stabilization (12). How-ever, the signal cascades involved in activation of theosmolyte releasing pathways after swelling, ischemia,or contraction remain incompletely understood (20, 22).
The swelling-induced activation of taurine release invarious cells involves PLA2-mediated release of arachi-donic acid from phospholipids and sequential forma-tion of a lipoxygenase product by the 5-lipoxygenase(5-LO) (17, 21, 40). On the other hand, the LPC-in-duced taurine release seems to involve a calmodulin-/calmodulin-dependent kinase II (CaMKII)-modulatedproduction of reactive oxygen species (ROS), (18). How-ever, in spite of apparent differences in the signalingcascades, both the swelling-induced, as well as theLPC-induced, pathways comprise PLA2 activity, butthe type of fatty acid and lysophospholipids formed bythe PLA2 and the subsequent physiological conse-quences depend on the isoform and the substrate spec-ificity of the PLA2, as well as the subcellular site ofaction.
An increase in ROS production has been demon-strated under osmotic cell swelling, muscle contrac-tion, anoxia, and sepsis (16, 25, 33). However, it is atpresent not unequivocal whether ROS production isupstream or downstream of PLA2 activation. One sug-gested mechanism for ROS production involves an in-crease in the cytosolic Ca2� concentration ([Ca2�]i),reduction in ATP, and a subsequent activation of PLA2(secretory type, sPLA2), whereupon the released fatty
Address for reprint requests and other correspondence: I. H.Lambert, The August Krogh Institute, Biochemical Dept., Uni-versitetsparken 13, DK-2100 Copenhagen Ø, Denmark (E-mail:[email protected]).
The costs of publication of this article were defrayed in part by thepayment of page charges. The article must therefore be herebymarked ‘‘advertisement’’ in accordance with 18 U.S.C. Section 1734solely to indicate this fact.
Am J Physiol Cell Physiol 284: C1362–C1373, 2003.First published January 8, 2003; 10.1152/ajpcell.00287.2002.
0363-6143/03 $5.00 Copyright © 2003 the American Physiological Society http://www.ajpcell.orgC1362
on October 23, 2006
ajpcell.physiology.orgD
ownloaded from

acids interfere with the electron transport chain in themitochondria, resulting in increased ROS production(32). On the other hand, anoxia and ROS have beenreported to directly increase PLA2 activity, either dueto peroxidation of lipids, reduction in membrane fluid-ity, and/or oxidation of protein side chains or sulfhy-dryl formation (25, 27). Additionally, ROS may directlymodulate the activity of arachidonic acid oxidation byactivation of, e.g., 5-LO (42).
It follows that activation of the osmolyte-releasingpathways potentially comprise PLA2 activity, polyun-saturated fatty acids, lysophospholipids, ROS, and[Ca2�]. In the present study, we have used myotubes,derived from porcine primary satellite cells, and C2C12myotubes as model systems for skeletal muscle cells to1) describe the taurine efflux pathways after hypoto-nicity, anoxia, and exogenous addition of the lipaseactivator melittin or LPC and 2) to analyze the role ofPLA2, ROS, and [Ca2�]i in the signaling cascades in-volved in the activation of the taurine efflux. Cellcultures have a number of important advantages in thestudy of cell signaling, i.e., 1) release of osmolytes canbe examined with a relatively high time resolution, 2)quantitative estimation of variations in the intracellu-lar level of free Ca2� and ROS production are easilyobtained, 3) osmotic or ischemic perturbation, or addi-tion of various drugs or metabolites, has nearly instanteffects, and 4) relatively pure muscle cell systems canbe obtained compared with intact preparations. Basedon the present results and previously published data,we present a model (Fig. 7) illustrating the role of PLA2and ROS in release of the organic osmolyte taurinefrom muscle.
MATERIALS AND METHODS
Chemicals. Antibiotics, fetal bovine serum (FCS), horseserum (HS), Dulbecco’s modified Eagle’s medium (DMEM;high glucose) with L-glutamine, and PBS were from LifeTechnologies (Naperville, IL). [14C]taurine was from NENLife Science Products, (Boston, MA). Fura 2-AM, 5-(and-6)-carboxy-2�,7�,-dichlorodihydrofluorescin diacetate (DCFH-DA), pluoronic F-127, and ionomycin were purchased fromMolecular Probes (Eugene, OR). Laminin and collagen werefrom (Becton Dickinson, Denmark). ETH 615–139 was do-nated by Dr. I. Ahnfelt-Rønne (Løvens Kemiske Fabrik, Den-mark). LPC (with palmitic acid in sn-1) and all other com-pounds were from Sigma Chemical (St. Louis, MO). Choles-terol was added from a 10-mM stock solution in Etoh.
Inorganic media. Isosmotic KCl solution contained in mM150 KCl, 1.3 CaCl2, 0.5 MgCl2, and 10 HEPES. HyposmoticKCl solution was obtained by reduction of the KCl in theisosmotic KCl solution to 95 mM, with the other componentsremaining unchanged, giving an osmolarity of 210 mosM.Krebs-HEPES buffer (KHB) contained in mM 118.0 NaCl,4.7 KCl, 1.2 KH2PO4, 4.2 NaHCO3, 1.3 CaCl2, 1.2 MgSO4,10.0 HEPES, and 10 D-glucose. pH was in all solutionsadjusted to 7.40.
Myotube cell cultures. Measurements were performed onmyotube cultures derived from porcine primary satellitecells, isolated from M. semimembranosus from 6- to 7-wk-oldpigs using a modified version of Bischoff’s original method(3), unless otherwise stated. The muscle tissue was excised,placed in ice-cold PBS buffer (Ca2� free and with 1,000 IU/ml
of penicillin), and transferred to a laminar flow bench. Themuscle tissue was finely chopped with a pair of scissors anddigested (3� 20 min) in 20 ml of PBS (Ca2� free) containing1% glucose, 1.5 mg/ml collagenase II, 0.25% trypsin, and0.01% DNAse. After digestion, the cells were transferred to aprimary growth medium (PGM; DMEM with 10% FCS and10% HS supplemented with 100 IU/ml penicillin and 100�g/ml streptomycin sulfate, 3 �g/ml amphotericin B, and 20�g/ml gentamycin), triturated 10 times (glass pipette), cen-trifuged at 150 g for 10 min at 4°C), resuspended, and filteredthrough a 200-�m and then a 50-�m Nytex filter. To enrichthe relative proportion of satellite cells in the cell suspen-sions, we optimized the procedure, before seeding and growthof the cells, with respect to 1) dispase treatment, 2) pre-platening, and 3) Percoll-gradient centrifugation. After alltreatments (except the dispase treatment), the subsequentcell suspensions were seeded at a density of 2,000 cells percm2 in 25-cm2 Nunc flasks coated with matrigel (1:200 vol/vol) and grown in PGM until near confluence (95% air-5%CO2 at 37°C). Initial studies showed that matrigel was supe-rior to laminin or collagen coating and uncoated Nunc plasticor primaria wells (data not shown). At near confluence, cellswere left to fuse in DMEM containing 2% FCS in the absenceor presence of 1 �M insulin for 5 days. Fusion percentage wasdetermined as the number of myotube nuclei in relation tothe total number of nuclei. A myotube was defined as three ormore nuclei within a cell membrane.
From Fig. 1, it is seen that direct seeding of the filtered cellsuspension on the matrigel-coated surface (control) resultedin fusion percentages of 36 and 19% in the presence andabsence of insulin, respectively. Treatment of cells near con-fluence with dispase (0.025%), to release/remove nonmyo-genic cells (fibroblasts), did not improve the final fusionpercentage (Fig. 1). Preplatening for 30 min on either prima-ria or uncoated plates, followed by retrieval of nonattachedcells, e.g., myogenic satellite cells, was also not effective (Fig.1), which is in accordance with the findings of Doumit andMerkel (9). Spinning the cells for 20 min at 1,250 g througha density gradient of 35/50/70% Percoll did not improve thefusion percentage (Fig. 1). However, spinning the cells for 5min at 20,000 g through a 20% Percoll density gradient layerresulted in populations with fusion percentages of 58 and50% in the presence and absence of insulin, respectively. Inthe present investigations, we have accordingly used 20%Percoll centrifugation, seeding density of 15,000 cells percm2, and insulin (1 �M) in the fusion medium.
For the intracellular Ca2� and ROS measurements, cellswere cultured on matrigel-coated (1:10 vol/vol) glass cover-slips (10 �43 mm) in four-well multidishes (Nunc 366148)with two coverslips in each well. For the taurine effluxmeasurements, cells were seeded in six-well Primaria Multi-well plates (35-mm diameter; Becton Dickinson Labware). ForWestern blots, cells were grown in Nunc 25-cm2 flasks (Nunc156340). Rat myotube cultures derived from primary satellitecells were prepared according to the method used for porcinemyotubes, with the following exceptions: 1) rat satellite cellswere preplated for 30 min before being seeded, 2) fusion me-dium was DMEM with 10% HS, and 3) insulin was omitted. Themyoblast cell line (C2C12), originally derived from the thighmuscle of the mouse (Ref. 44; American Type Culture Collec-tion, Manassas, VA), was prepared, grown, and differentiatedfor experiments as described previously (34).
Estimation of the free cytoplasmic Ca2� concentration andROS production. For the Ca2� measurements, the myotubeswere grown on coverslips, washed in KHB, and incubated inKHB containing 3 �M of fura 2-AM and 0.04% pluoronicF-127 for 1 h at 25°C, washed three times with KHB, and
C1363ROLE OF PLA2 AND ROS ON TAURINE RELEASE
AJP-Cell Physiol • VOL 284 • JUNE 2003 • www.ajpcell.org
on October 23, 2006
ajpcell.physiology.orgD
ownloaded from

incubated 15 min at room temperature to allow deesterificationof the fura 2-AM. For the ROS measurements, the coverslipswere incubated with dichlorofluorescein DCFH-DA (10 �M) for2 h and washed in KHB. The coverslips were placed vertically ina 10-mm path length cuvette, and analysis was performed on aPerkin-Elmer LS50B fluorimeter (Beaconsfield). For the Ca2�
measurements, the emission wavelength was 510 nm and thesubsequent ratiometric data (the ratio R at excitation 340 nm/excitation 380 nm ratio) were collected every 2 s. For ROSexperiments, the excitation and emission wavelengths were 490and 515 nm, respectively, and data were collected every 2 s. Thebackground signal (cells without fura 2) was subtracted fromthe fluorescence data before the ratio values were converted tofree Ca2� concentrations according to the equation [Ca2�] �Kd �(R � Rmin)/(Rmax � R) �(Sf,2/Sb,2) (10). For each series of eightexperiments, the ratio signal was calibrated by adding 5 �Mionomycin (Rmax) or 8 mM EGTA (Rmin), and a Kd of 224 nMwas experimentally determined using a standard Ca2� calibra-tion buffer kit (Molecular Probes, Eugene, OR). Sequentialdegassing by suction and bubbling the assay buffer with N2,before use, obtained anoxia.
Efflux measurements–estimation of rate constants. Taurineefflux from the cells was measured at room temperature(�20°C), as described previously (11, 19). Myotubes plated insix-well dishes were loaded with [14C]taurine (30 nCi/ml) for12 h at 37°C. Preliminary loading experiments on porcinemyotubes revealed that 12 h were necessary for optimalloading. For the experiments with C2C12 cells and rat myo-tubes, cells were loaded with 14C-labeled taurine for 3 h,unless otherwise stated. The preincubation solution was as-pirated, and the cells were washed five times with isosmoticKCl solution to remove excess extracellular [14C]taurine.Experimental solution (1 ml) was added to the dish, left for 2min, and transferred to a scintillation vial for estimation of
14C activity (�-scintillation counting; Ultima Gold). This pro-cedure was repeated for 30 min. At the end of the experiment,the amount of [14C]taurine remaining inside the cells wasestimated by adding 1 ml of 0.5 M NaOH for 1 h, washing thedishes twice with distilled water, and estimating the 14Cactivity in the NaOH, as well as in water washouts. Thenatural logarithm to the fraction of 14C activity remaining inthe cells at a given time was plotted vs. time (see Fig. 2A),and the rate constant for the taurine efflux at all time pointswas estimated as the negative slope of the graph betweentime point and its proceeding time point and plotted vs. time(see Fig. 2B). Efflux experiments under anoxic conditionswere performed with deoxygenated buffer in a closed cham-ber with a constant flow of N2 (glove box environmentalchamber 34790-10, Cole-Parmer).
Western immunoblots. Cells, lysed in 3 ml of buffer con-taining Tris [50 mM/NP-40 (1%)/SDS (0.1%)/NaCl (150 mM)/Na-azide (0.02%)/Na-deoxycholate (0.5%)], were removedfrom the flask and sonicated (two times for 10 min) on ice andcentrifuged at 600 g for 10 min. The supernatant was trans-ferred to new Eppendorf tubes and centrifuged in two stepsat 120,000 g for 45 min at 4°C. The membrane fractions werediluted 1:1 with solubilization buffer containing 62.5 mMTris, 4% SDS, 18% glycerol, 200 mM dithiothreitol, 1 mMphenylmethanesulfonyl fluoride, 1 �M pepstatin A, 10 �ME-64, and 1 mM leupeptin before being loaded on SDS/polyacrylamide gels. Twenty five-microgram membranepreparations were separated on a 10% SDS-polyacrylamidegel and transferred to a nitrocellulose membrane. Mem-branes were blocked with dry milk and incubated in rabbitpolyclonal antibodies raised against native purified humanleukocyte 5-LO (Merck Frosst Centre for Therapeutic Re-search, Pointe Claire-Dorval, QC, Canada) (1:250) for 2.5 h.The blot was subsequently incubated in alkaline phospha-
Fig. 1. Validation of preparation procedures to enrich the relative number of satellite cells, isolated from M.semimembranosus. After filtration of the digested muscle tissue, the cells were subjected to 1) dispase treatmentof confluent cells to release fibroblasts, 2) Percoll-gradient centrifugation, or 3) preplatening, as described inMATERIALS AND METHODS. After all treatments, the subsequent cell suspensions were seeded in 25-cm2 Nunc flaskscoated with matrigel (1:200 vol/vol) and grown in primary growth medium (PGM) until near confluence. Atconfluence, cells were left to fuse in Dulbecco’s modified Eagle’s medium (DMEM) containing 2% FCS in theabsence or presence of 1 �M insulin for 5 days. The fusion percentage was estimated after staining withhematoxilin. There was a significant effect of both treatment (P 0.005) and insulin (P 0.001).
C1364 ROLE OF PLA2 AND ROS ON TAURINE RELEASE
AJP-Cell Physiol • VOL 284 • JUNE 2003 • www.ajpcell.org
on October 23, 2006
ajpcell.physiology.orgD
ownloaded from
tase-conjugated anti-rabbit antibody (1:500) for 1 h. Proteinbands were detected by the phosphatase activity.
Statistical analysis. Comparison of measurements madeon a single parameter (i.e., taurine release or ROS produc-tion) during two or more experimental conditions (i.e., differ-ent agents or release taurine modes) was made by usingone-way analysis of variance (ANOVA). When the ANOVAresulted in a significant F value (P 0.05), the significantdifference between means was located by the Fisher’s pro-tected least significant difference post hoc test. Statisticalcomparisons of Ca2� content and ROS production duringanoxia were evaluated with a Student’s t-test. For all statis-tical evaluations, significance was established at the 0.05level; n is the number of experiments. All values are given asmeans SE.
RESULTS
Osmotic cell swelling and exogenous addition ofmelittin or LPC stimulate taurine release from porcinemyotubes. We have recently demonstrated that expos-ing myotube cultures derived from porcine satellite
cells to hypotonic KCl medium induces taurine releasevia a transport pathway that requires 5-LO activity foractivation and that is inhibited by the anion channelblocker DIDS (20). This is confirmed in Fig. 2, A and B,and now quantified in Table 1. Dilution of the isotonicsolution to 66% of the normal osmolarity (210 mosM)leads to a 20-fold increase in the rate constant fortaurine efflux, and the maximal rate constant for tau-rine efflux, obtained by the osmotic cell swelling, issignificantly decreased by 80 and 70% in the presenceof the 5-LO inhibitor ETH 615–139 and DIDS, respec-tively (Table 1). It is noted that taurine efflux is carriedout in KCl medium to avoid the regulatory volumereduction after the osmotic cell swelling and the con-comitant inactivation of the volume-sensitive taurineefflux pathway (14). However, cells depolarize whenexposed to KCl medium and as the volume-sensitivetaurine efflux from, e.g., HeLa cells is reduced underdepolarized conditions (40), it seems reasonable to as-
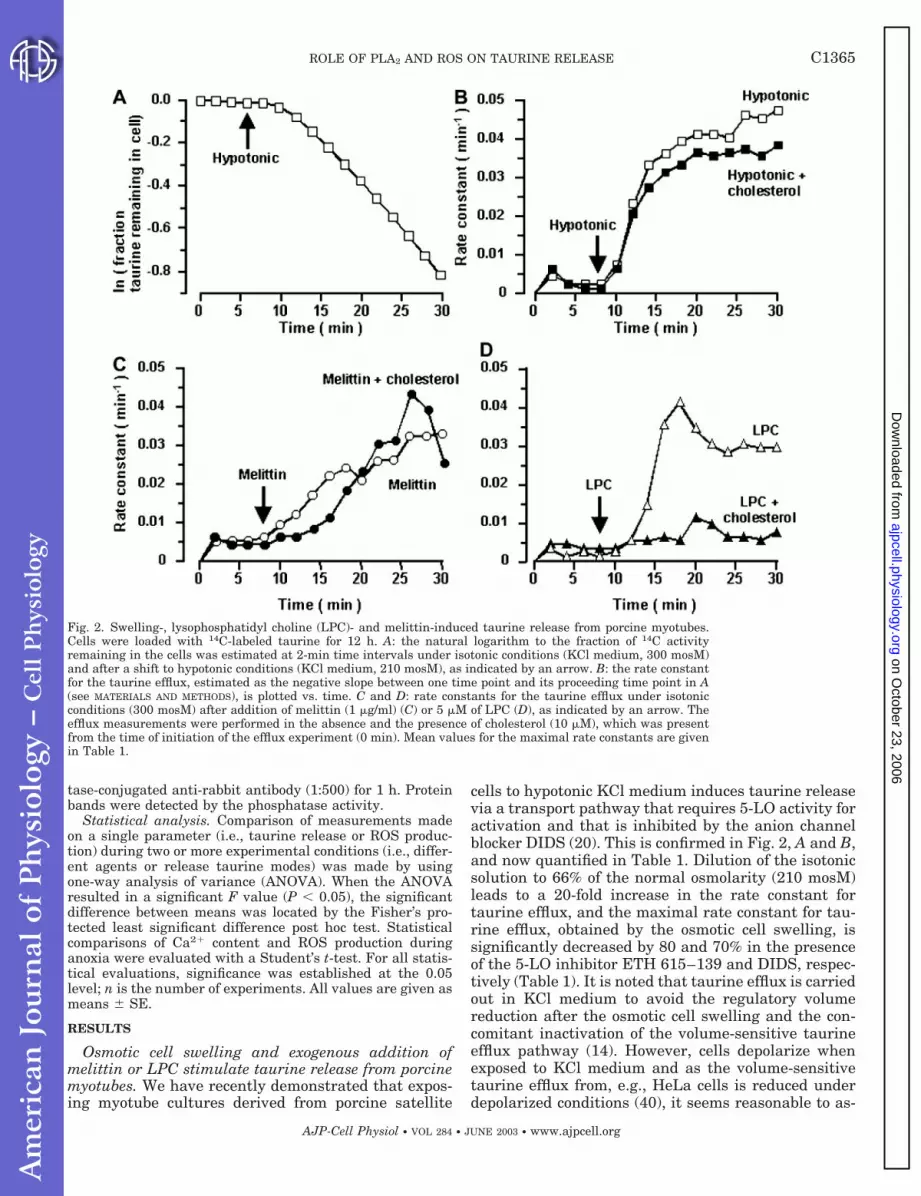
Fig. 2. Swelling-, lysophosphatidyl choline (LPC)- and melittin-induced taurine release from porcine myotubes.Cells were loaded with 14C-labeled taurine for 12 h. A: the natural logarithm to the fraction of 14C activityremaining in the cells was estimated at 2-min time intervals under isotonic conditions (KCl medium, 300 mosM)and after a shift to hypotonic conditions (KCl medium, 210 mosM), as indicated by an arrow. B: the rate constantfor the taurine efflux, estimated as the negative slope between one time point and its proceeding time point in A(see MATERIALS AND METHODS), is plotted vs. time. C and D: rate constants for the taurine efflux under isotonicconditions (300 mosM) after addition of melittin (1 �g/ml) (C) or 5 �M of LPC (D), as indicated by an arrow. Theefflux measurements were performed in the absence and the presence of cholesterol (10 �M), which was presentfrom the time of initiation of the efflux experiment (0 min). Mean values for the maximal rate constants are givenin Table 1.
C1365ROLE OF PLA2 AND ROS ON TAURINE RELEASE
AJP-Cell Physiol • VOL 284 • JUNE 2003 • www.ajpcell.org
on October 23, 2006
ajpcell.physiology.orgD
ownloaded from

sume that the maximal values for the rate constants inTable 1 are slightly underestimated. Subjecting wholecell lysates from pig myotubes to SDS/PAGE and usingpolyclonal antibodies, raised against human leukocyte5-LO, revealed two protein bands in the molecularrange of 55–60 kDa (data not shown). Thus the 5-LO ispresent in the porcine myotubes and required for acti-vation of the volume-sensitive taurine efflux pathway.
Melittin is reported to activate various lipases in-cluding sPLA2 isoform (24). From Fig. 2C and Table 1,it can be seen that exposing myotubes to melittin (1�g/ml) under isotonic conditions leads to a release oftaurine and that the rate constant for the melittin-induced taurine efflux within 15 min of stimulationbecomes similar in magnitude to the rate constant forthe swelling-induced taurine efflux. Furthermore, themaximal rate constant for the melittin-induced taurinerelease is decreased to the same degree as the maximalrate constant for the swelling-induced taurine efflux inthe presence of DIDS and ETH 615–139 (Table 1).Thus stimulation of PLA2 activity in porcine myotubes,either indirectly by osmotic swelling or directly underisotonic conditions by addition of melittin, induces tau-rine release via activation of signaling cascades whichinvolve 5-LO activity, and DIDS-sensitive efflux path-ways.
From Fig. 2D and Table 1, it can be seen that expo-sure of porcine myotube cultures to isotonic mediumcontaining LPC at a low concentration (5 �M) inducestaurine release. The maximal rate constant for theLPC-induced taurine efflux is decreased by 90% in thepresence of cholesterol and increased by DIDS. Choles-terol, on the other hand, only causes a 5-min delay ofthe onset of the melittin-induced taurine release (Fig.2C and Table 1) and a minor reduction in the maximalrate constant for the swelling-induced taurine efflux(Fig. 2B and Table 1). From the data presented in Fig.2 and Table 1, it is suggested that myotubes have twoseparate taurine efflux pathways, i.e., a pathway thatis activated by osmotic cell swelling or exposure tomelittin and a pathway that is activated by exogenousaddition of LPC. The two pathways are distinguishedby their opposing sensitivity toward the anion channelblocker DIDS and cholesterol.
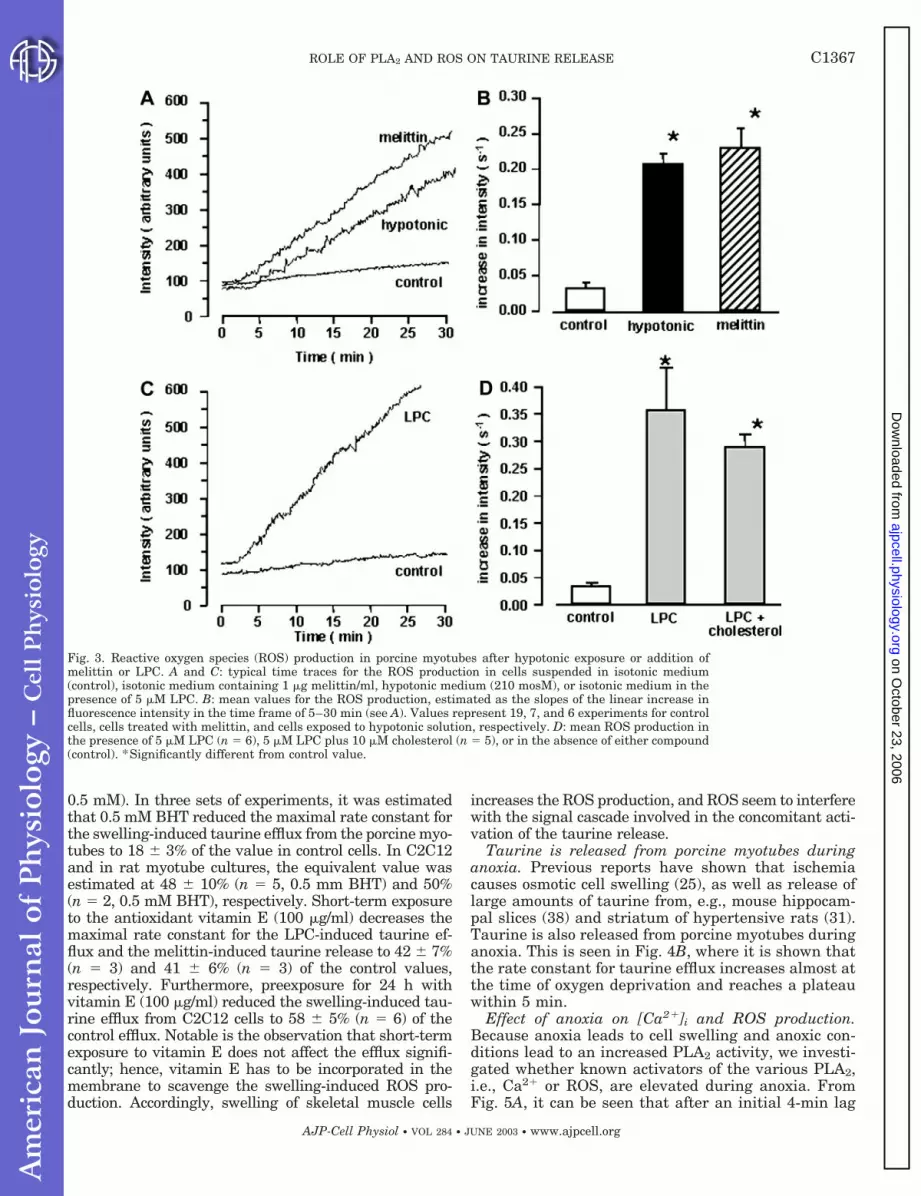
ROS production increases after osmotic cell swellingand after exposure to melittin and LPC. DCFH is rap-idly oxidized to the fluorescent compound dehydrodi-chlorofluorescein (DCF) by various ROS, as well asperoxynitrite (23). An increase in the DCF fluorescenceis accordingly taken as an indication of formation of theentire group of ROS. With the fluorescent probe DCFH/DCF used as an indicator of ROS production, it isdemonstrated that after an initial lag phase of about3–4 min, the ROS production in porcine myotubesincreases linearly with time after osmotic cell swellingand after treatment with melittin (Fig. 3A). The ROSproduction, estimated from the linear increase in thefluorescence between 5–30 min in Fig. 3A, indicatesthat there is a small background ROS production inmyotubes incubated in isotonic KHB (0.03 0.02 s�1,n � 19, Fig. 3, A and B) and that the ROS productionincreases significantly after hypotonic exposure(0.21 0.01 s�1, n � 7, Fig. 3B) and by the addition ofmelittin (0.24 0.01 s�1, n � 6, Fig. 3B).
Exogenous addition of LPC also increases ROS pro-duction in the porcine myotube cultures. This is seen inFig. 3, C and D, where it is demonstrated that thefluorescence of DCFH/DCF after a 1–3-min lag phaseincreases linearly with time. The ROS production afterLPC addition (5 �M) is estimated at 0.36 0.03 s�1
(n � 6, Fig. 3D), which is in the same range as the ROSproduction after melittin addition (Fig. 3B). There is amodest 19% decrease (P 0.05) in the LPC-inducedROS production in the presence of 10 �M cholesterol(0.29 0.01 s�1, n � 6, Fig. 3, C and D). Consequently,the inhibitory effect of cholesterol on the LPC-inducedtaurine release (Table 1) can only to a minor extent bedue to an antioxidative or substrate inhibitory effect ofthe cholesterol.
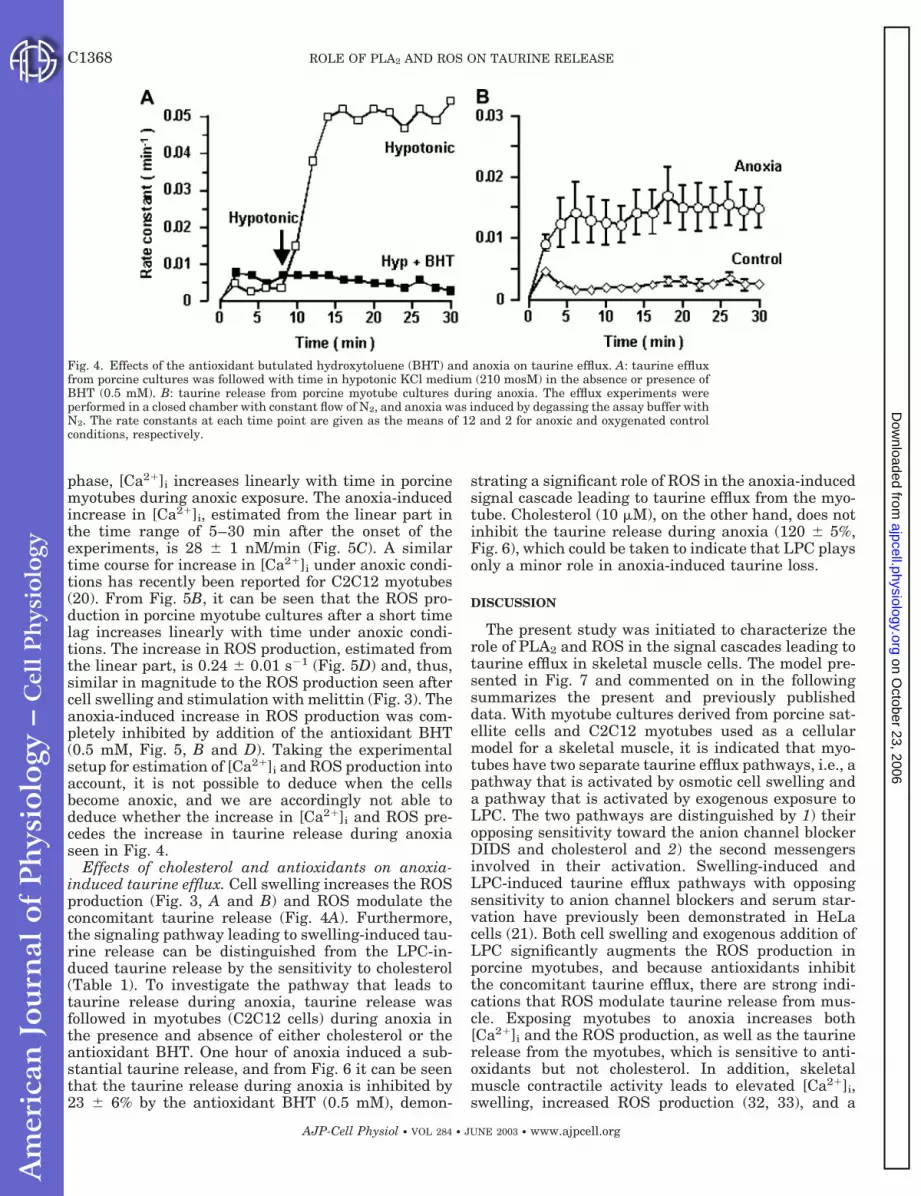
Effect of antioxidants on the swelling-induced tau-rine efflux. Cell swelling induces taurine release, aswell as an increased ROS production (Figs. 2B and 3, Aand B); however, it is not known whether there is acausal relationship between the two events. Both theswelling-induced increase in ROS production in por-cine myotubes (data not shown), as well as the concom-itant taurine efflux (Fig. 4A), are inhibited in the pres-ence of the antioxidant BHT (butulated hydroxytoluene,
Table 1. Effect of DIDS, cholesterol, and ETH 615–139 on taurine release in porcine myotube cultures
Hypotonic Melittin LPC
Maximal rate constant absolute value, min�1
0.0590.006(6) 0.0380.005(4) 0.0430.003(7)
Maximal rate constant relative to control, %
Control 100 100 100DIDS 282* (6) 167* (4) 20724* (6)Cholesterol 842* (3) 1074 (3) 122* (3)ETH 615-139 183* (3) 216* (3)
The maximal rate constant for taurine release following osmotic cell swelling in hypotonic solution or stimulation with melittin (1 �g/ml)or lysophosphatidyl choline (LPC) (5 �M) under isotonic conditions was estimated as indicated in Fig 2. DIDS (100 �M), cholesterol (10 �M),and ETH 615–139 (10 �M) were present during the whole experimental period. Values for taurine release in the presence of drugs are themaximal rate constants given relative to the respective control with no drug present. Number of experiments is given in parentheses.Asterisks indicate significant difference from the control value.
C1366 ROLE OF PLA2 AND ROS ON TAURINE RELEASE
AJP-Cell Physiol • VOL 284 • JUNE 2003 • www.ajpcell.org
on October 23, 2006
ajpcell.physiology.orgD
ownloaded from
0.5 mM). In three sets of experiments, it was estimatedthat 0.5 mM BHT reduced the maximal rate constant forthe swelling-induced taurine efflux from the porcine myo-tubes to 18 3% of the value in control cells. In C2C12and in rat myotube cultures, the equivalent value wasestimated at 48 10% (n � 5, 0.5 mm BHT) and 50%(n � 2, 0.5 mM BHT), respectively. Short-term exposureto the antioxidant vitamin E (100 �g/ml) decreases themaximal rate constant for the LPC-induced taurine ef-flux and the melittin-induced taurine release to 42 7%(n � 3) and 41 6% (n � 3) of the control values,respectively. Furthermore, preexposure for 24 h withvitamin E (100 �g/ml) reduced the swelling-induced tau-rine efflux from C2C12 cells to 58 5% (n � 6) of thecontrol efflux. Notable is the observation that short-termexposure to vitamin E does not affect the efflux signifi-cantly; hence, vitamin E has to be incorporated in themembrane to scavenge the swelling-induced ROS pro-duction. Accordingly, swelling of skeletal muscle cells
increases the ROS production, and ROS seem to interferewith the signal cascade involved in the concomitant acti-vation of the taurine release.
Taurine is released from porcine myotubes duringanoxia. Previous reports have shown that ischemiacauses osmotic cell swelling (25), as well as release oflarge amounts of taurine from, e.g., mouse hippocam-pal slices (38) and striatum of hypertensive rats (31).Taurine is also released from porcine myotubes duringanoxia. This is seen in Fig. 4B, where it is shown thatthe rate constant for taurine efflux increases almost atthe time of oxygen deprivation and reaches a plateauwithin 5 min.
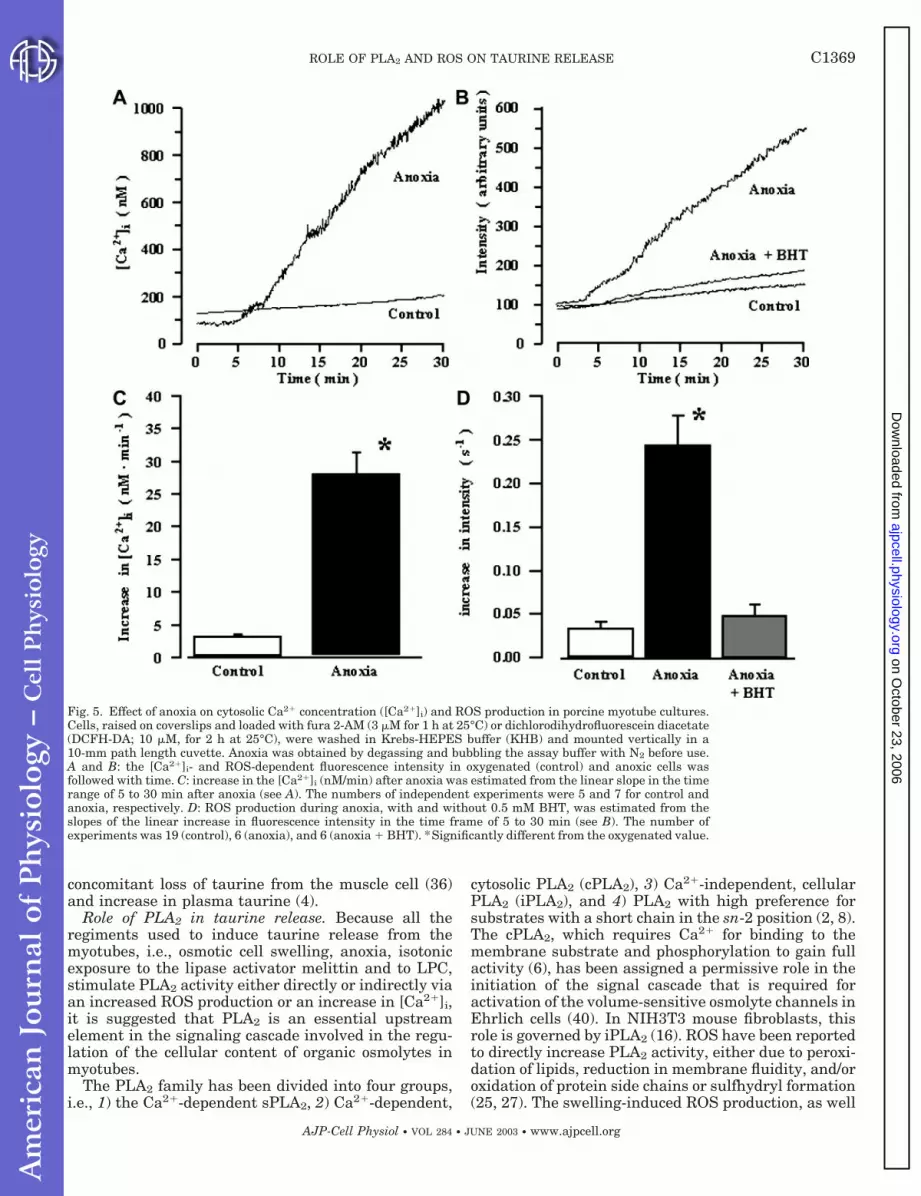
Effect of anoxia on [Ca2�]i and ROS production.Because anoxia leads to cell swelling and anoxic con-ditions lead to an increased PLA2 activity, we investi-gated whether known activators of the various PLA2,i.e., Ca2� or ROS, are elevated during anoxia. FromFig. 5A, it can be seen that after an initial 4-min lag
Fig. 3. Reactive oxygen species (ROS) production in porcine myotubes after hypotonic exposure or addition ofmelittin or LPC. A and C: typical time traces for the ROS production in cells suspended in isotonic medium(control), isotonic medium containing 1 �g melittin/ml, hypotonic medium (210 mosM), or isotonic medium in thepresence of 5 �M LPC. B: mean values for the ROS production, estimated as the slopes of the linear increase influorescence intensity in the time frame of 5–30 min (see A). Values represent 19, 7, and 6 experiments for controlcells, cells treated with melittin, and cells exposed to hypotonic solution, respectively. D: mean ROS production inthe presence of 5 �M LPC (n � 6), 5 �M LPC plus 10 �M cholesterol (n � 5), or in the absence of either compound(control). *Significantly different from control value.
C1367ROLE OF PLA2 AND ROS ON TAURINE RELEASE
AJP-Cell Physiol • VOL 284 • JUNE 2003 • www.ajpcell.org
on October 23, 2006
ajpcell.physiology.orgD
ownloaded from
phase, [Ca2�]i increases linearly with time in porcinemyotubes during anoxic exposure. The anoxia-inducedincrease in [Ca2�]i, estimated from the linear part inthe time range of 5–30 min after the onset of theexperiments, is 28 1 nM/min (Fig. 5C). A similartime course for increase in [Ca2�]i under anoxic condi-tions has recently been reported for C2C12 myotubes(20). From Fig. 5B, it can be seen that the ROS pro-duction in porcine myotube cultures after a short timelag increases linearly with time under anoxic condi-tions. The increase in ROS production, estimated fromthe linear part, is 0.24 0.01 s�1 (Fig. 5D) and, thus,similar in magnitude to the ROS production seen aftercell swelling and stimulation with melittin (Fig. 3). Theanoxia-induced increase in ROS production was com-pletely inhibited by addition of the antioxidant BHT(0.5 mM, Fig. 5, B and D). Taking the experimentalsetup for estimation of [Ca2�]i and ROS production intoaccount, it is not possible to deduce when the cellsbecome anoxic, and we are accordingly not able todeduce whether the increase in [Ca2�]i and ROS pre-cedes the increase in taurine release during anoxiaseen in Fig. 4.
Effects of cholesterol and antioxidants on anoxia-induced taurine efflux. Cell swelling increases the ROSproduction (Fig. 3, A and B) and ROS modulate theconcomitant taurine release (Fig. 4A). Furthermore,the signaling pathway leading to swelling-induced tau-rine release can be distinguished from the LPC-in-duced taurine release by the sensitivity to cholesterol(Table 1). To investigate the pathway that leads totaurine release during anoxia, taurine release wasfollowed in myotubes (C2C12 cells) during anoxia inthe presence and absence of either cholesterol or theantioxidant BHT. One hour of anoxia induced a sub-stantial taurine release, and from Fig. 6 it can be seenthat the taurine release during anoxia is inhibited by23 6% by the antioxidant BHT (0.5 mM), demon-
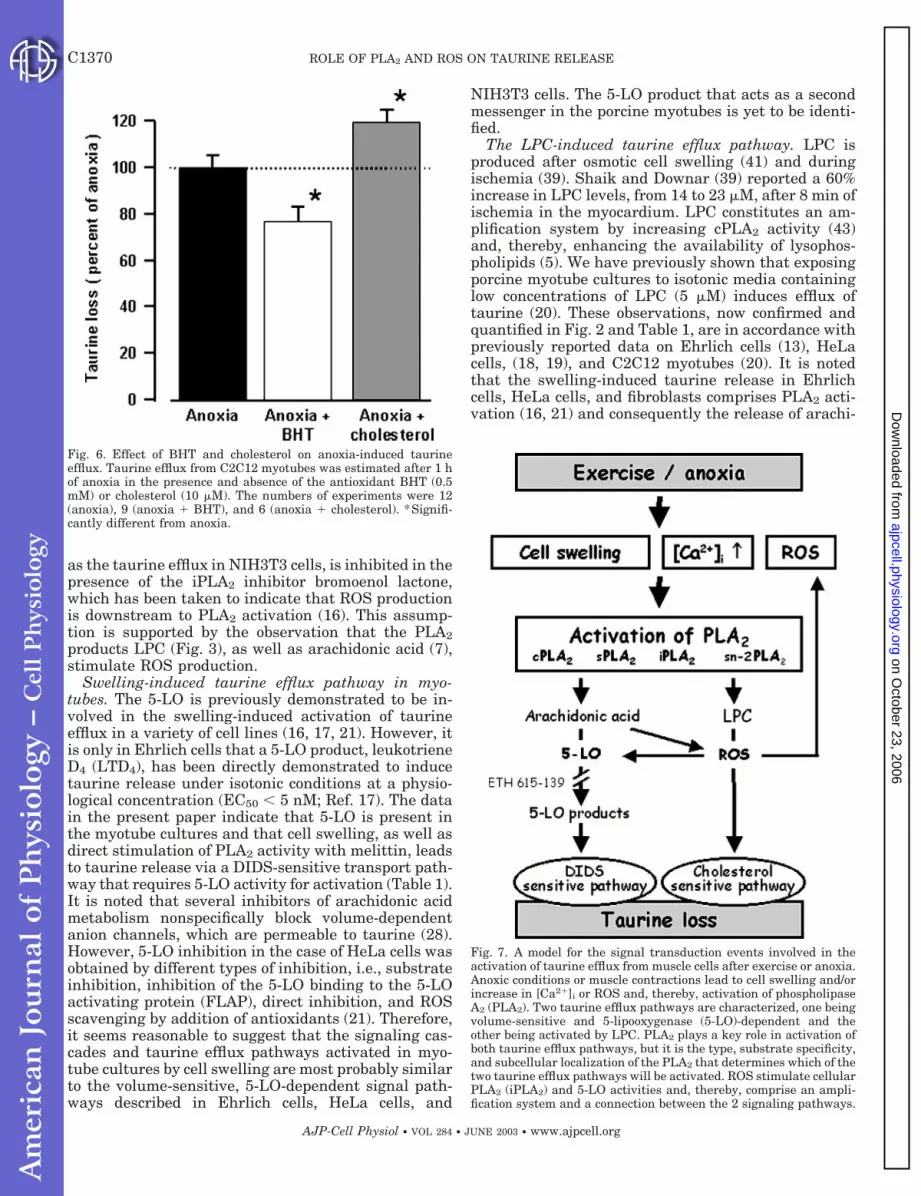
strating a significant role of ROS in the anoxia-inducedsignal cascade leading to taurine efflux from the myo-tube. Cholesterol (10 �M), on the other hand, does notinhibit the taurine release during anoxia (120 5%,Fig. 6), which could be taken to indicate that LPC playsonly a minor role in anoxia-induced taurine loss.
DISCUSSION
The present study was initiated to characterize therole of PLA2 and ROS in the signal cascades leading totaurine efflux in skeletal muscle cells. The model pre-sented in Fig. 7 and commented on in the followingsummarizes the present and previously publisheddata. With myotube cultures derived from porcine sat-ellite cells and C2C12 myotubes used as a cellularmodel for a skeletal muscle, it is indicated that myo-tubes have two separate taurine efflux pathways, i.e., apathway that is activated by osmotic cell swelling anda pathway that is activated by exogenous exposure toLPC. The two pathways are distinguished by 1) theiropposing sensitivity toward the anion channel blockerDIDS and cholesterol and 2) the second messengersinvolved in their activation. Swelling-induced andLPC-induced taurine efflux pathways with opposingsensitivity to anion channel blockers and serum star-vation have previously been demonstrated in HeLacells (21). Both cell swelling and exogenous addition ofLPC significantly augments the ROS production inporcine myotubes, and because antioxidants inhibitthe concomitant taurine efflux, there are strong indi-cations that ROS modulate taurine release from mus-cle. Exposing myotubes to anoxia increases both[Ca2�]i and the ROS production, as well as the taurinerelease from the myotubes, which is sensitive to anti-oxidants but not cholesterol. In addition, skeletalmuscle contractile activity leads to elevated [Ca2�]i,swelling, increased ROS production (32, 33), and a
Fig. 4. Effects of the antioxidant butulated hydroxytoluene (BHT) and anoxia on taurine efflux. A: taurine effluxfrom porcine cultures was followed with time in hypotonic KCl medium (210 mosM) in the absence or presence ofBHT (0.5 mM). B: taurine release from porcine myotube cultures during anoxia. The efflux experiments wereperformed in a closed chamber with constant flow of N2, and anoxia was induced by degassing the assay buffer withN2. The rate constants at each time point are given as the means of 12 and 2 for anoxic and oxygenated controlconditions, respectively.
C1368 ROLE OF PLA2 AND ROS ON TAURINE RELEASE
AJP-Cell Physiol • VOL 284 • JUNE 2003 • www.ajpcell.org
on October 23, 2006
ajpcell.physiology.orgD
ownloaded from
concomitant loss of taurine from the muscle cell (36)and increase in plasma taurine (4).
Role of PLA2 in taurine release. Because all theregiments used to induce taurine release from themyotubes, i.e., osmotic cell swelling, anoxia, isotonicexposure to the lipase activator melittin and to LPC,stimulate PLA2 activity either directly or indirectly viaan increased ROS production or an increase in [Ca2�]i,it is suggested that PLA2 is an essential upstreamelement in the signaling cascade involved in the regu-lation of the cellular content of organic osmolytes inmyotubes.
The PLA2 family has been divided into four groups,i.e., 1) the Ca2�-dependent sPLA2, 2) Ca2�-dependent,
cytosolic PLA2 (cPLA2), 3) Ca2�-independent, cellularPLA2 (iPLA2), and 4) PLA2 with high preference forsubstrates with a short chain in the sn-2 position (2, 8).The cPLA2, which requires Ca2� for binding to themembrane substrate and phosphorylation to gain fullactivity (6), has been assigned a permissive role in theinitiation of the signal cascade that is required foractivation of the volume-sensitive osmolyte channels inEhrlich cells (40). In NIH3T3 mouse fibroblasts, thisrole is governed by iPLA2 (16). ROS have been reportedto directly increase PLA2 activity, either due to peroxi-dation of lipids, reduction in membrane fluidity, and/oroxidation of protein side chains or sulfhydryl formation(25, 27). The swelling-induced ROS production, as well
Fig. 5. Effect of anoxia on cytosolic Ca2� concentration ([Ca2�]i) and ROS production in porcine myotube cultures.Cells, raised on coverslips and loaded with fura 2-AM (3 �M for 1 h at 25°C) or dichlorodihydrofluorescein diacetate(DCFH-DA; 10 �M, for 2 h at 25°C), were washed in Krebs-HEPES buffer (KHB) and mounted vertically in a10-mm path length cuvette. Anoxia was obtained by degassing and bubbling the assay buffer with N2 before use.A and B: the [Ca2�]i- and ROS-dependent fluorescence intensity in oxygenated (control) and anoxic cells wasfollowed with time. C: increase in the [Ca2�]i (nM/min) after anoxia was estimated from the linear slope in the timerange of 5 to 30 min after anoxia (see A). The numbers of independent experiments were 5 and 7 for control andanoxia, respectively. D: ROS production during anoxia, with and without 0.5 mM BHT, was estimated from theslopes of the linear increase in fluorescence intensity in the time frame of 5 to 30 min (see B). The number ofexperiments was 19 (control), 6 (anoxia), and 6 (anoxia � BHT). *Significantly different from the oxygenated value.
C1369ROLE OF PLA2 AND ROS ON TAURINE RELEASE
AJP-Cell Physiol • VOL 284 • JUNE 2003 • www.ajpcell.org
on October 23, 2006
ajpcell.physiology.orgD
ownloaded from
as the taurine efflux in NIH3T3 cells, is inhibited in thepresence of the iPLA2 inhibitor bromoenol lactone,which has been taken to indicate that ROS productionis downstream to PLA2 activation (16). This assump-tion is supported by the observation that the PLA2products LPC (Fig. 3), as well as arachidonic acid (7),stimulate ROS production.
Swelling-induced taurine efflux pathway in myo-tubes. The 5-LO is previously demonstrated to be in-volved in the swelling-induced activation of taurineefflux in a variety of cell lines (16, 17, 21). However, itis only in Ehrlich cells that a 5-LO product, leukotrieneD4 (LTD4), has been directly demonstrated to inducetaurine release under isotonic conditions at a physio-logical concentration (EC50 5 nM; Ref. 17). The datain the present paper indicate that 5-LO is present inthe myotube cultures and that cell swelling, as well asdirect stimulation of PLA2 activity with melittin, leadsto taurine release via a DIDS-sensitive transport path-way that requires 5-LO activity for activation (Table 1).It is noted that several inhibitors of arachidonic acidmetabolism nonspecifically block volume-dependentanion channels, which are permeable to taurine (28).However, 5-LO inhibition in the case of HeLa cells wasobtained by different types of inhibition, i.e., substrateinhibition, inhibition of the 5-LO binding to the 5-LOactivating protein (FLAP), direct inhibition, and ROSscavenging by addition of antioxidants (21). Therefore,it seems reasonable to suggest that the signaling cas-cades and taurine efflux pathways activated in myo-tube cultures by cell swelling are most probably similarto the volume-sensitive, 5-LO-dependent signal path-ways described in Ehrlich cells, HeLa cells, and
NIH3T3 cells. The 5-LO product that acts as a secondmessenger in the porcine myotubes is yet to be identi-fied.
The LPC-induced taurine efflux pathway. LPC isproduced after osmotic cell swelling (41) and duringischemia (39). Shaik and Downar (39) reported a 60%increase in LPC levels, from 14 to 23 �M, after 8 min ofischemia in the myocardium. LPC constitutes an am-plification system by increasing cPLA2 activity (43)and, thereby, enhancing the availability of lysophos-pholipids (5). We have previously shown that exposingporcine myotube cultures to isotonic media containinglow concentrations of LPC (5 �M) induces efflux oftaurine (20). These observations, now confirmed andquantified in Fig. 2 and Table 1, are in accordance withpreviously reported data on Ehrlich cells (13), HeLacells, (18, 19), and C2C12 myotubes (20). It is notedthat the swelling-induced taurine release in Ehrlichcells, HeLa cells, and fibroblasts comprises PLA2 acti-vation (16, 21) and consequently the release of arachi-
Fig. 6. Effect of BHT and cholesterol on anoxia-induced taurineefflux. Taurine efflux from C2C12 myotubes was estimated after 1 hof anoxia in the presence and absence of the antioxidant BHT (0.5mM) or cholesterol (10 �M). The numbers of experiments were 12(anoxia), 9 (anoxia � BHT), and 6 (anoxia � cholesterol). *Signifi-cantly different from anoxia.
Fig. 7. A model for the signal transduction events involved in theactivation of taurine efflux from muscle cells after exercise or anoxia.Anoxic conditions or muscle contractions lead to cell swelling and/orincrease in [Ca2�]i or ROS and, thereby, activation of phospholipaseA2 (PLA2). Two taurine efflux pathways are characterized, one beingvolume-sensitive and 5-lipooxygenase (5-LO)-dependent and theother being activated by LPC. PLA2 plays a key role in activation ofboth taurine efflux pathways, but it is the type, substrate specificity,and subcellular localization of the PLA2 that determines which of thetwo taurine efflux pathways will be activated. ROS stimulate cellularPLA2 (iPLA2) and 5-LO activities and, thereby, comprise an ampli-fication system and a connection between the 2 signaling pathways.
C1370 ROLE OF PLA2 AND ROS ON TAURINE RELEASE
AJP-Cell Physiol • VOL 284 • JUNE 2003 • www.ajpcell.org
on October 23, 2006
ajpcell.physiology.orgD
ownloaded from

donic acid and LPC (40). However, the observation thatcholesterol inhibits the LPC-induced taurine efflux(Fig. 2D and Table 1), whereas cholesterol has only aminor effect on the swelling-induced taurine efflux(Fig. 2B and Table 1), could be taken to indicate thatonly a minor amount of LPC is released from themuscle upon osmotic swelling. This is in accordancewith previous consideration (19). LPC is a bioactivelipid molecule involved in a broad variety of biologicalprocesses. LPC at low concentrations (10 �M) in-duces release of osmolytes in various cells (18, 19), butthe structural requirement for the lysophospholipidsis, in the case of the HeLa cells, strict, and only lyso-phospholipids with choline as the polar head group andoleic, palmitic, or stearic acid in the sn-1 position areable to elicit taurine release (19). Furthermore, LPCbinding receptors have recently been identified (13,46). On the other hand, LPC at high concentrations(�10 �M) induces the release of creatine kinase fromcardiomyocytes (5), as well as the release of nucleotidesfrom HeLa cells (19). Thus LPC at high concentrationsseems to break down the membrane permeability bar-rier. It is not possible to deduce whether the effects of5 �M LPC, used in the present paper, reflect activationof specific osmolyte transporting systems or a generalpermeabilization of the plasma membrane. Cholesterolis an integral component of the eukaryotic cell mem-branes, with a highly nonuniform distribution amongthe membranes, and it is estimated that about 80–90%of the total cell membrane cholesterol is in the surfacemembrane (45). As cholesterol has only a minor effecton the LPC-induced ROS production (Fig. 3), whereasit decreases the LPC-induced taurine efflux (Fig. 2C), itseems reasonable to exclude the possibility that theeffect of cholesterol on the flux reflects an antioxidativeeffect. However, cholesterol affects membrane fluidityand it is possible that cholesterol affects either LPCbinding to a putative receptor or the interaction be-tween the plasma membrane and the cone-shaped LPC(cross-sectional area of the polar head group is largerthan that of the area of the acyl chain) and, thereby,the action of LPC.
Role of ROS in the regulation of osmolyte-releasingsystems. Cell swelling and stimulation with melittin isaccompanied by an increase in the taurine efflux,which is evident after about 2 min (Fig. 2) and anincrease in ROS production that appears after a 3- to4-min delay (Fig. 3). Thus measurable increase in ROSproduction appears after the onset of the taurine efflux.Swelling-induced ROS production in NIH3T3 cells in-volves the NAD(P)H oxidase (16), oxidation of polyun-saturated fatty acids (arachidonic acid), and formationof lipoxygenase products by the 5-LO. H2O2 is requiredfor 5-LO activity in mammalian cells (30), and becausethe 5-LO is present in the myotube cultures (see RE-SULTS), it seems plausible to assume that ROS, pro-duced by cell swelling, modulate the activity of thevolume-sensitive taurine efflux pathway via interfer-ence with the 5-LO. This suggestion is supported bythe observation that exogenous addition of H2O2 toHeLa cells and NIH3T3 cells potentiates the swelling-
induced taurine efflux and that this effect of H2O2 isimpaired in the presence of the 5-LO inhibitor ETH615–139 (16, 19). H2O2 inhibits protein tyrosine phos-phatases and, because the swelling-induced taurineefflux in, e.g., NIH3T3 cells is regulated by tyrosinephosphorylation, it has alternatively been suggestedthat the potentiating effect of ROS on swelling inducedtaurine efflux reflect oxidation and a subsequent inhi-bition of protein tyrosine phosphatase activity (16).ROS are also produced after LPC exposure (Fig. 3), andbecause the LPC-induced taurine efflux from porcinemyotubes is inhibited by antioxidants (16), it seemsreasonable to assume that ROS production is requiredfor the LPC-induced activation of taurine efflux. Thesubcellular site of production and action of ROS in theLPC-induced signaling cascade is at present notknown; however, it seems to be modulated by a calmod-ulin-/CaMKII (18).
Role of PLA2 in anoxia-induced release of osmolytes.Anoxia and exercise lead, according to the model in Fig.7, to activation of one or several types of PLA2. Theincreased PLA2 activity subsequently leads to genera-tion of free fatty acids (arachidonic acid) and LPC.Arachidonic acids and LPC then directly or indirectlyfunction as second messengers in the activation oftaurine efflux pathways. The data in Figs. 4 and 5reveal that anoxia is accompanied by a rise in [Ca2�]i,ROS production, and taurine release. Both cPLA2 andsPLA2 are stimulated by a rise in [Ca2�]i, i.e., sPLA2requires Ca2� in the millimolar range for activation,whereas cPLA2 requires Ca2� in the micromolar rangefor translocation to the nuclear membrane and subse-quent activation (29). On the other hand, iPLA2 hasrecently been demonstrated to be involved in the mech-anism by which ROS increases the availability of freearachidonic acid in macrophages RAW 264.7 (27). Cellsswell during anoxia, and, because cPLA2 and iPLA2have already been assigned a permissive role in theswelling-induced release of osmolytes in Ehrlich cells(35) and fibroblasts (16), it seems reasonable to assumethat cPLA2 activation, together with iPLA2, is involvedin anoxia-induced taurine release. Direct stimulationof PLA2 activity in porcine myotube cultures withmelittin under isotonic conditions leads to taurine re-lease that is sensitive to the 5-LO inhibitor ETH 615–139 (Fig. 2C and Table 1). Because both LPC-inducedand melittin-induced taurine release are sensitive tovitamin E, it seems reasonable to assume that LPC isproduced by the action of melittin. The selectivity ofmelittin has been tested, and it appears that melittinat high concentrations (�1 �g/ml) activates otherlipases than sPLA2, resulting in extensive release ofvarious saturated, mono-, and polyunsaturated fattyacids from various cell lines and consequently leads toalteration of the lipid composition in cellular mem-branes, as well as initiation of diverse effects on cellphysiology (24). There are several extracellular sPLA2isozymes (type I, IIA, IIC, IID, V, and X), and they haveall been assigned a role in stimulus-induced release ofarachidonic acid (2, 29). sPLA2-X releases arachidonicacid in the absence of costimulators and is known to
C1371ROLE OF PLA2 AND ROS ON TAURINE RELEASE
AJP-Cell Physiol • VOL 284 • JUNE 2003 • www.ajpcell.org
on October 23, 2006
ajpcell.physiology.orgD
ownloaded from

hydrolyze phosphatidyl choline to LPC in the outerleaflet of the plasma membrane (29). Activation ofcPLA2 (osmotic swelling) and sPLA2 (melittin addition)stimulate ROS production (Fig. 3). As DIDS depresstaurine release after anoxia (data not shown) and theROS production is concomitantly increased, it is sug-gested that 1) PLA2 activity increases during anoxia inmuscle and 2) the 5-LO-dependent taurine efflux path-way contributes to the anoxia-induced release of tau-rine. There seems to be a functional cooperation be-tween cPLA2 and signaling through sPLA2 (IIA),whereas sPLA2 (X)-mediated release is independent ofcPLA2 activity (29). It is noted that the volume-sensi-tive taurine efflux pathways in C2C12 myoblasts aredownregulated during differentiation (26), i.e., differ-entiation most probably add to the complexity of theactivation of the taurine-releasing systems.
In conclusion, the results of this study indicate thatROS is produced during regiments that induce taurinerelease in myotubes, i.e., swelling, anoxia, and additionof LPC or melittin. Furthermore, the present findingsindicate that ROS potentiate, whereas antioxidantsattenuate, the taurine efflux after swelling, anoxia,and the addition of LPC. PLA2 is a key mediator of thetaurine efflux because its products provide upstreamelements for two separate taurine efflux pathways, i.e.,a swelling-induced pathway that requires arachidonicacid and 5-LO activity for activation and an LPC-induced pathway. The two pathways are distinguishedby their opposing sensitivity toward the anion channelblocker DIDS and cholesterol. It appears that the iden-tity, substrate specificity, and subcellular localizationof the PLA2 determine the second messengers pro-duced by the PLA2 and, subsequently, the type oftaurine efflux pathway to be activated.
This work was supported by the Danish Bacon and Meat Counciland the Danish Natural Sciences Research Council.
REFERENCES
1. Bakker AJ and Berg HM. Effect of taurine on sarcoplasmicreticulum function and force in skinned fast-twitch skeletalmuscle fibres of the rat. J Physiol 538: 185–194, 2002.
2. Balsinde J, Balboa MA, Insel PA, and Dennis EA. Regula-tion and inhibition of phospholipase A2. Annu Rev PharmacolToxicol 39: 175–189, 1999.
3. Bischoff R. Enzymatic liberation of myogenic cells from adultrat muscle. Anat Rec 180: 645–661, 1974.
4. Blomstrand E and Saltin B. Effect of muscle glycogen onglucose, lactate and amino acid metabolism during exercise andrecovery in human subjects. J Physiol 514: 293–302, 1999.
5. Chen M, Xiao CY, Hashizume H, and Abiko Y. Differentialeffects of Ca2� channel blockers on Ca2� overload induced bylysophosphatidylcholine in cardiomyocytes. Eur J Pharmacol333: 261–268, 1997.
6. Clark JD, Schievella AR, Nalefski EA, and Lin LL. Cytosolicphospholipase A2. J Lipid Mediat Cell Signal 12: 83–117, 1995.
7. Cocco T, Di Paola M, Pappa S, and Lorusso M. Arachidonicacid interaction with the mitochondrial electron transport chainpromotes reactive oxygen species generation. Free Radic BiolMed 27: 51–59, 1999.
8. Cummings BS, McHowat J, and Schnellmann RG. Phospho-lipase A2s in cell injury and death. J Pharmacol Exp Ther 294:793–799, 2000.
9. Doumit ME and Merkel RA. Conditions for isolation andculture of porcine myogenic satellite cells. Tissue Cell 24: 253–262, 1992.
10. Grynkiewicz G, Poenie M, and Tsien RY. A new generationof Ca2� indicators with greatly improved fluorescence properties.J Biol Chem 260: 3440–3450, 1985.
11. Hall JA, Kirk JA, Potts JR, Rae C, and Kirk K. Anionchannel blockers inhibit swelling-activated anion, cation, andnonelectrolyte transport in HeLa cells. Am J Physiol Cell Physiol271: C579–C588, 1996.
12. Huxtable R and Bressler R. Effect of taurine on a muscleintracellular membrane. Biochim Biophys Acta 323: 573–583,1973.
13. Kabarowski JHS, Zhu K, Le LQ, Witte ON, and Xu Y.Lysophosphatidylcholine as a ligand for the immunoregulatorreceptor G2A. Science 293: 702–705, 2001.
14. Kirk K and Kirk J. Volume-regulatory taurine release from ahuman lung cancer cell line. Evidence for amino acid transportvia a volume-activated chloride channel. FEBS Lett 336: 153–158, 1993.
15. Kramer JH, Chovan JP, and Schaffer SW. Effect of taurineon calcium paradox and ischemic heart failure. Am J PhysiolHeart Circ Physiol 240: H238–H246, 1981.
16. Lambert IH. Reactive oxygen species regulate swelling-inducedtaurine efflux in NIH3T3 mouse fibroblasts. J Membr Biol. Inpress.
17. Lambert IH. Regulation of the taurine content in Ehrlich as-cites tumour cells. In: Taurine 3: Cellular and Regulatory Mech-anisms, edited by Schaffer S, Lombardini JB, and Huxtable RJ.New York: Plenum, 1998, p. 269–276.
18. Lambert IH and Falktoft B. Lysophosphatidylcholine-inducedtaurine release in HeLa cells involves kinase activity. CompBiochem Physiol Part A: 577–584, 2001.
19. Lambert IH and Falktoft B. Lysophosphatidylcholine inducestaurine release from HeLa cells. J Membr Biol 176: 175–185,2000.
20. Lambert IH, Nielsen JH, Andersen HJ, and Ørtenblad N.Cellular model for induction of drip loss in meat. J Agric FoodChem 49: 4876–4883, 2001.
21. Lambert IH and Sepulveda FV. Swelling-induced taurineefflux from HeLa cells. In: Taurine 4: Taurine and ExcitableTissues, edited by Della Corte L, Huxtable RJ, Sgaragli GP, andTipton KF. New York: Kluwer, Academic/Plenum, 2000, p. 487–496.
22. Lang F, Busch GL, Ritter M, Volkl H, Waldegger S, Gul-bins E, and Haussinger D. Functional significance of cellvolume regulatory mechanisms. Physiol Rev 78: 247–306, 1998.
23. LeBel CP, Ischiropoulos H, and Bondy SC. Evaluation of theprobe 2�,7�-dichlorofluorescein as an indicator of reactive oxygenspecies formation and oxidative stress. Chem Res Toxicol 5:227–231, 1992.
24. Lee SY, Park HS, Lee SJ, and Choi MU. Melittin exertsmultiple effects on the release of free fatty acids from L1210cells: Lack of selective activation of phospholipase A2 by melittin.Arch Biochem Biophys 389: 57–67, 2001.
25. Lipton P. Ischemic cell death in brain neurons. Physiol Rev 79:1432–1568, 1999.
26. Manolopoulos VG, Droogmans G, and Nilius B. Hypotonic-ity and thrombin activate taurine efflux in BC3H1 and C2C12myoblasts that is down regulated during differentiation. Bio-chem Biophys Res Commun 232: 74–79, 1997.
27. Martinez J and Moreno JJ. Role of Ca2�-independent phos-pholipase A2 on arachidonic acid release induced by reactiveoxygen species. Arch Biochem Biophys 392: 257–262, 2001.
28. McManus M, Serhan C, Jackson P, and Strange K. Keto-conazole blocks organic osmolyte efflux independently of itseffect on arachidonic acid conversion. Am J Physiol Cell Physiol267: C266–C271, 1994.
29. Murakami M, Nakatani Y, Kuwata H, and Kudo I. Cellularcomponents that functionally interact with signalling phospho-lipase A2s. Biochim Biophys Acta 1488:159–166, 2000.
30. Musser JH and Kreft AF. 5-lipoxygenase: properties, pharma-cology, and the quinolinyl(bridged)aryl class of inhibitors. J MedChem 35: 2501–2524, 1992.
C1372 ROLE OF PLA2 AND ROS ON TAURINE RELEASE
AJP-Cell Physiol • VOL 284 • JUNE 2003 • www.ajpcell.org
on October 23, 2006
ajpcell.physiology.orgD
ownloaded from

31. Nakane H, Yao H, Ibayashi S, Kitazono T, Ooboshi H,Uchimura H, and Fujishima M. Protein kinase C modulatesischemia-induced amino acids release in the striatum of hyper-tensive rats. Brain Res 26: 290–296, 1998.
32. Nethery D, Callahan LA, Stofan D, Mattera R, DiMarco A,and Supinski G. PLA2 dependence of diaphragm mitochondrialformation of reactive oxygen species. J Appl Physiol 89: 72–80,2000.
33. Nethery D, Stofan D, Callahan L, DiMarco A, and SupinskiG. Formation of reactive oxygen species by the contracting dia-phragm is PLA2 dependent. J Appl Physiol 87: 792–800, 1999.
34. Oksbjerg N, Petersen JS, Sørensen IL, Henckel P, Vester-gaard M, Ertbjerg P, Møller AJ, Bejerholm C, and Støier S.Long-term changes in performance and meat quality of DanishLandrace pigs: a study on a current compared with an unim-proved genotype. Animal Sci 71: 81–92, 2000.
35. Pedersen S, Lambert IH, Thoroed SM, and Hoffmann EK.Hypotonic cell swelling induces translocation of the alpha iso-form of cytosolic phospholipase A2 but not the � isoform inEhrlich ascites tumor cells. Eur J Biochem 267: 5531–5539,2000.
36. Quesada O, Lu P, and Sturman JA. Taurine distribution indifferent cat muscles as visualized by immunohistochemistry:changes with stimulus state. Cytobios 73: 143–154, 1993.
37. Ramamoorthy S, Leibach FH, Mahesh VB, Han H, Yang-Feng T, Blakely RD, and Ganapathy V. Functional charac-terization and chromosomal localization of a cloned taurinetransporter from human placenta. Biochem J 300: 893–900,1994.
38. Saransaari P and Oja SS. Mechanisms of ischemia-inducedtaurine release in mouse hippocampal slices. Brain Res 807:118–124, 1998.
39. Shaikh NA and Downar E. Time course of changes in porcinemyocardial phospholipid levels during ischemia. A reassessmentof the lysolipid hypothesis. Circ Res 49: 316–325, 1981.
40. Stutzin A, Torres R, Oporto M, Pacheco P, Eguiguren AL,Cid LP, and Sepulveda FV. Separate taurine and chlorideefflux pathways activated during regulatory volume decrease.Am J Physiol Cell Physiol 277: C392–C402, 1999.
41. Thoroed SM, Lauritzen L, Lambert IH, Hansen HH, andHoffmann EK. Cell swelling activates phospholipase A2 inEhrlich ascites tumour cells. J Membr Biol 160: 47–58, 1997.
42. Werz O, Szellas D, and Steinhilber D. Reactive oxygen spe-cies released from granulocytes stimulate 5-lipoxygenase activ-ity in a B-lymphocytic cell line. Eur J Biochem 267: 1263–1269,2000.
43. Wong JT, Tran K, Pierce GN, Chan AC, OK, and Choy PC.Lysophosphatidylcholine stimulates the release of arachidonicacid in human endothelial cells. J Biol Chem 273: 6830–6836,1998.
44. Yaffe D and Saxel O. Serial passaging and differentiation ofmyogenic cells isolated from dystrophic mouse muscle. Nature270: 725–727, 1977.
45. Yeagle PL. Cholesterol and the cell membrane. Biochim Bio-phys Acta 822: 267–287, 1985.
46. Zhu K, Baudhuin LM, Hong G, Williams FS, Cristina KL,Kabarowski JH, Witte ON, and Xu Y. Sphingosylphosphoryl-choline and lysophosphatidylcholine are ligands for the G protein-coupled receptor GPR4. J Biol Chem 276: 41325–41335, 2001.
C1373ROLE OF PLA2 AND ROS ON TAURINE RELEASE
AJP-Cell Physiol • VOL 284 • JUNE 2003 • www.ajpcell.org
on October 23, 2006
ajpcell.physiology.orgD
ownloaded from
Related Documents











