Eur. J. Biochem. YI, 303-310 (1978) Rapid Purification of Plasmid DNAs by Hydroxyapatite Chromatography Alan COLMAN, Michael J. BYERS, Sandy B. PRIMROSE, and Alan LYONS Department of Biological Sciences, University of Warwick (Received May 26, 1978) A method is described for the rapid preparation of plasmid DNAs of molecular weight up to 14 x lo6. This method involves the chromatography, at room temperature, of bacterial cleared lysates on hydroxyapatite in the presence of high concentrations of phosphate and urea. All detectable protein and RNA contamination of plasmid DNA is removed by this procedure and the con- formation of the plasmid DNA is unaffected. Less than 0.5 % chromosomal DNA is present in the purified preparation and even this can be removed if necessary by a simple extention of the procedure to include a heat-denaturation step. The method is extremely rapid and amenable to large-scale plasmid preparation; 5 mg ColEl DNA have been purified within 40 min. The yield of plasmid DNA is similar to that obtained with the conventional dye-centrifugation technique, however the purity is greater. The advent of recombinant DNA technology has facilitated the routine isolation of specific nucleic acid sequences of both prokaryotes and eukaryotes. This technology relies heavily on the use of vectors and for most cloning experiments bacterial plasmids are the vectors of choice. Plasmid DNA rarely constitutes as much as 50% of the total DNA present in a bac- terial cell, even under growth conditions most con- ducive to plasmid replication [l]. Purification of plasmid DNA therefore requires the selective removal of chromosomal DNA. A very effective initial pmifica- tion is achieved by the preparation of a cleared lysate [2]. Methods for the further purification of plasmid DNA take advantage of the closed-circular nature of the DNA [3,4]. The most widely used method relies on the greater density of the closed-circular DNA relative to sheared chromosomal DNA in ethidium bromide/caesium chloride density gradients [5]. While this technique provides a high degree of purification it suffers from a number of disadvantages: it is ex- pensive in both centrifuge time and materials, has limited capacity, and complete removal of the ethidium bromide is both difficult and tedious. In this paper we describe a purification procedure which does not depend on CsCl centrifugation and which is rapid, inexpensive and yields DNA of high purity. Our method takes advantage of the selective and reversible retention by hydroxyapatite of small double-stranded DNA molecules under conditions known to prevent the binding of all RNA and pro- tein [6]. MATERIALS AND METHODS Muter iuls Chemicals were obtained from the following sources : thymine, 2-deoxyadenosine, chlorampheni- col, ethidium bromide, Tris (Sigma Ltd) ; casamino acids, Bacto-tryptone, yeast extract (Difco Ltd) ; all other chemicals (Analar grade, British Drug Houses). [methyl-3H]Thymidine (22 Ci/mmol) was obtained from the Radiochemical Centre (Amersham). Escherichia coli Strains and Plasmids Table 1 shows the bacterial strains used, their source, the plasmids they contain, and the molecular weights of these plasmids. Growth of Bacteria Colonies of each of the above strains (except the HBlOl strains) were picked from a freshly streaked agar plate into 10 ml of nutrient broth and grown overnight at 37°C. Each culture was then used to inoculate 1 1 of either L broth or M-9 broth [I31 supplemented with 1.5 casamino acids (Difco Ltd) and 2% glucose (20 pg/ml thymine was included during routine growth of the 581 strain) and grown at 37 "C to an absorbance of 0.6 at 650 nm (3 x lo8 cells/ml). Solid chloramphenicol was then added to a final concentration of 100 pg/ml and incubation continued for a further 20 h. When [3H]thymidine-

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Eur. J. Biochem. Y I , 303-310 (1978)
Rapid Purification of Plasmid DNAs by Hydroxyapatite Chromatography Alan COLMAN, Michael J . BYERS, Sandy B. PRIMROSE, and Alan LYONS
Department of Biological Sciences, University of Warwick
(Received May 26, 1978)
A method is described for the rapid preparation of plasmid DNAs of molecular weight up to 14 x lo6.
This method involves the chromatography, at room temperature, of bacterial cleared lysates on hydroxyapatite in the presence of high concentrations of phosphate and urea. All detectable protein and RNA contamination of plasmid DNA is removed by this procedure and the con- formation of the plasmid DNA is unaffected. Less than 0.5 % chromosomal DNA is present in the purified preparation and even this can be removed if necessary by a simple extention of the procedure to include a heat-denaturation step. The method is extremely rapid and amenable to large-scale plasmid preparation; 5 mg ColEl DNA have been purified within 40 min. The yield of plasmid DNA is similar to that obtained with the conventional dye-centrifugation technique, however the purity is greater.
The advent of recombinant DNA technology has facilitated the routine isolation of specific nucleic acid sequences of both prokaryotes and eukaryotes. This technology relies heavily on the use of vectors and for most cloning experiments bacterial plasmids are the vectors of choice. Plasmid DNA rarely constitutes as much as 50% of the total DNA present in a bac- terial cell, even under growth conditions most con- ducive to plasmid replication [l]. Purification of plasmid DNA therefore requires the selective removal of chromosomal DNA. A very effective initial pmifica- tion is achieved by the preparation of a cleared lysate [2]. Methods for the further purification of plasmid DNA take advantage of the closed-circular nature of the DNA [3,4]. The most widely used method relies on the greater density of the closed-circular DNA relative to sheared chromosomal DNA in ethidium bromide/caesium chloride density gradients [5]. While this technique provides a high degree of purification it suffers from a number of disadvantages: it is ex- pensive in both centrifuge time and materials, has limited capacity, and complete removal of the ethidium bromide is both difficult and tedious.
In this paper we describe a purification procedure which does not depend on CsCl centrifugation and which is rapid, inexpensive and yields DNA of high purity. Our method takes advantage of the selective and reversible retention by hydroxyapatite of small double-stranded DNA molecules under conditions known to prevent the binding of all RNA and pro- tein [6].
MATERIALS AND METHODS Muter iuls
Chemicals were obtained from the following sources : thymine, 2-deoxyadenosine, chlorampheni- col, ethidium bromide, Tris (Sigma Ltd) ; casamino acids, Bacto-tryptone, yeast extract (Difco Ltd) ; all other chemicals (Analar grade, British Drug Houses). [methyl-3H]Thymidine (22 Ci/mmol) was obtained from the Radiochemical Centre (Amersham).
Escherichia coli Strains and Plasmids Table 1 shows the bacterial strains used, their
source, the plasmids they contain, and the molecular weights of these plasmids.
Growth of Bacteria Colonies of each of the above strains (except the
HBlOl strains) were picked from a freshly streaked agar plate into 10 ml of nutrient broth and grown overnight at 37°C. Each culture was then used to inoculate 1 1 of either L broth or M-9 broth [I31 supplemented with 1.5 casamino acids (Difco Ltd) and 2% glucose (20 pg/ml thymine was included during routine growth of the 581 strain) and grown at 37 "C to an absorbance of 0.6 at 650 nm (3 x lo8 cells/ml). Solid chloramphenicol was then added to a final concentration of 100 pg/ml and incubation continued for a further 20 h. When [3H]thymidine-

304 Rapid Purification of Plasmid DNAs by Hydroxyapatite Chromatography
Table 1. Bacterial strains used to obtain plasmids Strains 581, OS410 and C6H27 were a kind gift of D. Sheratt. The molecular weights of their plasmids were determined to be approxi- mately 1.07 times their published values (D. Sheratt, personal communication)
E. coli strain Plasmid x M , Refer- ence
581 (thy-, strR) ColEl 4.5 [I1 OS410 (strR, thi-) A 362 3.1 171 OS410 (strR, thi-) ColEl ::Tn7 13.8 [8 I
[91 C6H27 (trpP, Bl-, strs) pML21 7.9 [I01 HBlOl pXlrl 01 10.4 [ I l l HBlOl pX1108 12.7 - HBlOl px1212 11.5 -
- SPA-0 none
a
a P. G. Boseley, T. Moss, and M. L. Birnsteil, unpublished results.
labelled DNA was required, 500 pCi/1, [3H]thymidine and 2 mg/l 2-deoxyadenosine were added with the chloramphenicol. The plasmids pX1212, pX1108 and XlrlOl all contain inserts of Xenopus laevis ribosomal DNA. Their host bacterial strains were grown and lysed (see below) under category I1 containment con- ditions (Williams Report, H.M.S.O., U.K.) by Dr P. Rigby (XlrlOl) and Dr P. Boseley (pX1108, pX1212).
Preparation of the Cleared Lysate
Bacterial pellets were prepared from 1000-ml cul- tures by centrifugation for 30 min at 2500 rev./min in an M.S.E. Mistral 6L centrifuge at 4 "C. Each pellet was resuspended in 4 m10.25 M sucrose, 2 mM MgC12 and a cleared lysate prepared by the methods of Clewell and Helinski [2]. Essentially each resuspended pellet was successively incubated with 0.8 ml of 20 mg/ ml lysozyme (Sigma Ltd) in 10 mM Tris/HCl, pH 8.0, for 5 min at O'C, and then 1.6 ml 0.25 M EDTA, pH 8.0, for 5 min at 0 "C and finally 6.4 ml of 0.5 % (w/v) Triton X-100, 0.0625 M EDTA, 50 mM Tris/ HCl, pH 8.0, for 20 rnin at 0°C. After this final ad- dition, the sample was gently swirled at 5-min intervals. The EDTA concentrations in the above solutions were modified in later experiments to 40 mM and 10 mM respectively. The processing of smaller cultures (1 - 500 ml) can be accomplished using reduced quantities of the above solutions.
Hydroxyapatite Chromatography
Hydroxyapatite columns, first developed for pro- tein chromatography by Tiselius et al. [14], were later extended to the chromatography of nucleic acids by Bernardi [15]. We use a method originally described by Britten et al. [6] and further characterized by Meinke
et al. [16] which enables DNA to be purified on hy- droxyapatite from a mixture of RNA, DNA, and protein. Hydroxyapatite powder (Biorad, DNA grade) was resuspended in 8 M urea, 0.24 M sodium phos- phate buffer pH 6.8 (the indicated molarity refers to the phosphate moeity) using 1 g hydroxyapatite for every litre of bacterial culture processed. The slurry was then poured into a 3-cm-diameter glass column and washed with 2 vol. of the 8 M urea, 0.24 M phos- phate buffer pH 6.8. This, and all subsequent chro- matographic steps were done under a positive air pressure of 3000 Pa. 1.1 vol. of cleared lysate were diluted with 8.9 vol. of 9 M urea, 0.27 M phosphate buffer, 0.9% sodium dodecyl sulphate, pH 6.8, and the mixture then loaded on to the hydroxyapatite column. The column was washed with fresh 8 M urea, 0.24 M phosphate buffer, pH 6.8, until the absorbance at 260 nm of the column effluent was approximately zero. Two column volumes of 0.01 M phosphate buffer, pH 6.8, were then washed through the column. The hydroxyapatite was then resuspended in a further 2 vol. of this low-phosphate buffer before a final 2 vol. of buffer were washed through. Addition of 0.3 M phosphate buffer, pH 6.8, caused the plasmid DNA to elute. DNA recovery was monitored either spectro- photometrically or chemically using the diphenylamine assay as described by Burton [17]. The capacity of different batches of hydroxyapatite for DNA was found to be highly variable (see also [6]). At best we could recover at least 5 mg plasmid DNA from a column containing 3 g hydroxyapatite at worst 0.6 mg could be recovered from a similar column containing a different batch of hydroxyapatite.
Density GradientlDye Centrifugation
Cleared lysates were prepared for gradient cen- trifugation by the addition of 1 g caesium chloride and 0.1 ml ethidium bromide (10 mg/ml) to each ml of lysate. Solutions were centrifuged for 48 - 60 h at 40000 rev./min and 20 "C in an M.S.E. 8 x 50-ml angle rotor. After centrifugation, the denser fluorescent band which contains the superhelical DNA [5] was collected dropwise, the ethidium bromide removed by four successive extractions with isoamyl alcohol (saturated with CsC1) and the solution was then dialysed against 10 mM Tris, 10 mM NaC1,l mM EDTA, pH 7.6.
Gel Electrophoresis
Electrophoresis of DNA samples was carried out in 0.8% agarose (Sigma Type 11) gels on a 20x 15 x 0.4-cm flat-bed apparatus. The electrophoresis buffer used was either 18 mM Tris, 18 mM boric acid, 3 mM EDTA, pH 8.3 [18] or 40 mM Tris, 20 mM sodium acetate, 2 mM EDTA, pH 8.3 [19]. Electrophoresis was performed at either 120V for 2 h or 30V for

A. Colman, M. J. Byers, S. B. Primrose, and A. Lyons 305
15 h. After electrophoresis gels were soaked for 15 min in electrophoresis buffer containing 0.5 pg/ml ethi- dium bromide.
Protein and R N A Estimations
Protein and RNA contents of cleared lysates and purified DNA samples were estimated by the methods of Lowry et al. [20] and Fleck and Munro [21] re- spectively, as described previously [22].
RESULTS
Purfication of Plasmid DNA by the Hydroxyapatite Procedure
In order to establish the usefulness of the hydroxy- apatite procedure described in Materials and Methods, a variety of plasmids ranging in molecular weight from 3.1 x lo6 to 13.8 x lo6 were purified from cleared lysates. Fig.1 shows the result obtained by electro- phoresing samples of cleared lysate taken before and after purification by the hydroxyapatite procedure in 0.8 agarose gels. Not only could plasmids be purified by the hydroxyapatite procedure but the proportion of superhelical to nicked plasmid DNA was also un- affected by it. Therefore nicks were not introduced into the DNA during hydroxyapatite chromatography despite the use of positive air pressure to facilitate passage of solutions through the hydroxyapatite. The introduction of one single-strand break into a super- helical DNA molecule results in a total relaxation of that molecule with a concomitant slower migration during agarose gel electrophoresis. We found that the relative proportion of superhelical to nicked plasmid molecules in these cleared lysates was always high for the smaller plasmid molecules (e.g. ColE1, A392; Fig. 1). This predominance of superhelical molecules was not always found with the larger plasmids grown in L broth, although it was favoured by growth in defined M-9 medium (see Materials and Methods) (Colman, unpublished observations). Plasmid DNA purified by the hydroxyapatite procedure gave perfect cleavage on treatment with restriction endonucleases (data not shown) and the transformation efficiencies were identical to those obtained with plasmids purified by caesium/ethidium centrifugation (Primrose, un- published results).
Recovery of Plasmid D N A by Hydroxyapatite Chromatography
Assessment of the recovery of DNA after hydroxy- apatite chromatography is complicated by the pres- ence of large amounts of RNA, and possibly some chromosomal DNA, in cleared lysates from plasmid-
Fig. 1. Purijication of plasmid DNA by hydroxyapatite chromatog- raphy. Cleared lysates prepared from 100-ml cultures (L broth) of plasmid-containing strains of E. coli, were subjected to hydroxy- apatite Chromatography as described in Materials and Methods. 10-pI aliquots of each sample before and after hydroxyapatite chromatography were diluted with 40 p1 Tris/acetate electrophoresis buffer containing 0.2 % sodium dodecyl sulphate and 25 % (v/v) gly- cerol and electrophoresed on a 0.8% agarose gel for 2 h at 120 V. Tracks a - e contain the following hydroxyapatite-purified plasmid DNAs: (a) ColEl . TN7; (b) pX1108; (c) pML21; (d) ColEl; (e) 4362; tracks f-j contain the same plasmid DNAs respectively, before chromatography
bearing strains of E. coli. For this reason the following protocol was adopted. 10 pl 3H-labelled ColEl DNA (1 mgiml), purified by caesium/ethidium centrifuga- tion, was diluted with 0.55 ml cleared lysate from a plasmid-free strain of E. coli and subjected to the hydroxyapatite procedure. Aliquots from the urea/ phosphate wash, the low-salt wash and the 0.3 M phosphate eluant were precipitated with trichloroacetic acid and subsequently counted. Fig. 2 shows that over 95 % of the applied radioactivity was recovered in the 0.3 M phosphate eluant. A similar degree of recovery was obtained when 25 pg unlabelled pX1212 DNA ( M , 11.5 x lo6) was subjected to the hydroxyapatite procedure, absorbance at 260 nm being used to esti- mate recovery. These observations indicate that plas- mids of molecular weight as high as 11.5 x lo6 can be quantitatively recovered by our procedure.
The results shown in Table 2 demonstrate the im- pressive degree of DNA purification achieved in the one-step hydroxyapatite chromatographic step.

Rapid Purification of Plasmid DNAs by Hydroxyapatite Chromatography 306
25 r
Fraction number
Fig. 2. Recovery ofplusmid DNA by J~ydruxyupatite chrornatugrup!iy. 10 pg [3H]thymidine-labelled ColEl DNA (1 mg/ml) was diluted into 0.55 ml of a cleared lysate prepared from a culture of E. coli, strain SPA-0 (see Table 1). The mixture was loaded on to 0.3 g hydroxyapatite, washed in succession with 8 M urea, 0.24 M phos- phate buffer (I), 0.01 M phosphate buffer (11) before elution with 0.3 M phosphate buffer (111) all operations performed as described in Materials and Methods. l-ml fractions were collected, pooled in the case of fractions 1-10 and 11-15 and then diluted with an equal volume of 200 pg/ml yeast RNA (BDH Chemicals Ltd). To each sample approximately 5000 dis./min 32P-labelled mouse DNA were added before the addition of 2 volumes of ice-cold 20% trichloroacetic acid. Precipitates were collected on Whatman GF/C filters and counted in a PPO/POPOP/toluene-based scintillator. The inclusion of the [3ZP]DNA during the precipitation step allowed correction to be made for the non-quantitative (approx. 50 %) precipitation which occurred in the urea-containing sample. No radioactivity was left on the column
Table 2. Protein and RNA contents during purification ColEl DNA was purified by hydroxyapatite chromatography of the cleared lysate from a 1-1 culture as described in Materials and Methods. Aliquots of the cleared lysate and the purified DNA were assayed for RNA and protein content (see Materials and Methods)
Constituent Concentration in ~~
cleared lysate DNA purified from 1 ml cleared lysate
mg/ml Pg RNA 1.56 < I Protein 3.5 < 2 ColEl DNA not estimated 200
Isolation of the Plasmid DNA in Bulk
When 1 - 5 1 cultures of plasmid-containing bac- teria were subjected to the clearing procedure the resultant lysates were of necessity large (15-75 ml) to obtain a good release of plasmid DNA from the bacteria. In order to process these cleared lysates rapidly while using a minimal amount of hydroxy-
apatite, the nucleic acids were first precipitated by ad- dition of 2 vol. of cold ethanol. The precipitate was resuspended in 4 of the original volume of 10 mM Tris, 10 mM NaC1, 1 mM EDTA, pH 7.6, and mixed with lysing buffer prior to loading on the hydroxy- apatite column. Since there were large amounts of DNA and RNA in the cleared lysate which effectively acted as a carrier, precipitation took less than 10 min and did not affect the yield (as high as 2 mg plasmid/l culture) of plasmid obtained.
Absence of Significant Contamination of Plasmid DNA by Bacterial DNA
In most experiments agarose gel electrophoresis of hydroxyapatite-purified plasmid DNA never re- vealed the presence of any detectable chromosomal DNA and there are three possible explanations for this observation. First, chromosomal DNA might not be released by the clearing procedure. Secondly, any large fragments of chromosomal DNA might bind irre- versibly to the hydroxyapatite. Finally, any fragments of chromosomal DNA might be so heterogenous in size that they migrate as a single continuous band from the dye front to the origin. Resolution of these three possibilities was clearly important in evaluating the hydroxyapatite method and several experimental approaches were used.
First 50 pg I DNA (Mr 30.8 x lo6) was mixed with < 1 pg 3H-labelled ColE1 DNA and the mixture chromatographed through hydroxyapatite. No i. DNA was recovered in the 0.3 M phosphate eluant, as deter- mined by absorbance at 260 nm, whilst over 95% of the radioactivity was eluted. This experiment demon- strates that large linear DNA molecules are selectively retained by hydroxyapatite. This observation is in agreement with the findings of Britten et al. [6]; how- ever, Bernardi [ 151, using hydroxyapatite chromatog- raphy in the absence of urea, reported the elution of DNA molecules with Mr as large as 10'.
Small amounts of high-molecular-weight chromo- somal DNA could be detected in cleared lysates by grossly overloading the gel (Fig. 3A, slots c and e) but we estimate (see legend, Fig. 4) that this represents less than 0.5 %of the total DNA present. This chromo- somal DNA was also found in cleared lysates from chloramphenicol enriched plasmid-free strains of E. coli (Fig. 3A, slot b) as well as in lysates from un- enriched ColEl-carrying strains (Fig. 3 A, slot d). Although this chromosomal DNA co-migrated with 2 DNA its molecular weight is unclear, since all DNA with M , > 20 x lo6 will migrate at the same rate in 0.8 agarose gels. This very small amount of chromo- somal DNA co-purified after hydroxyapatite chro- matography with plasmid DNA (data not shown) and so must have a molecular weight less than that of
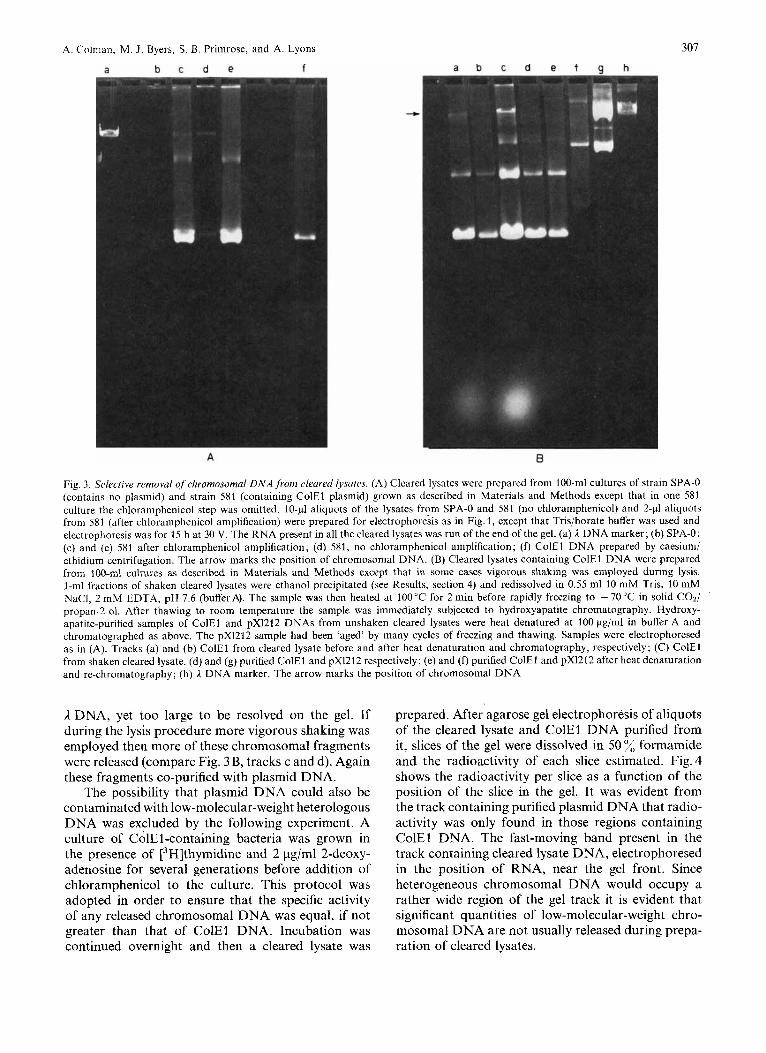
A . C'olman, M J. Byers, S. B. Primrose, and A. Lyons 307
Fig. 3 . Selective removal ojchromosomal D N A from cleared lysates. (A) Cleared lysates were prepared from 100-ml cultures of strain SPA-0 (contains no plasmid) and strain 581 (containing ColEl plasmid) grown as described in Materials and Methods except that in one 581 culture the chloramphenicol step was omitted. 10-pl aliquots of the lysates from SPA-0 and 581 (no chloramphenicol) and 2 4 aliquots from 581 (after chloramphenicol amplification) were prepared for electrophoresis as in Fig. 1, except that Tris/borate buffer was used and electrophoresis was for 15 h at 30 V. The RNA present in all the cleared lysates was run of the end of the gel. (a) I z DNA marker; (b) SPA-0; (c) and (e) 581 after chloramphenicol amplification; (d) 581, no chloramphenicol amplification; (f) ColEl DNA prepared by caesium/ ethidium centrifugation. The arrow marks the position of chromosomal DNA. (B) Cleared lysates containing ColEl DNA were prepared from 100-ml cultures as described in Materials and Methods except that in some cases vigorous shaking was employed during lysis. 1-ml fractions of shaken cleared lysates were ethanol precipitated (see Results, section 4) and redissolved in 0.55 ml 10 mM Tris, 10 mM NaCI, 2 mM EDTA, pH 7.6 (buffer A). The sample was then heated at 100 "C for 2 min before rapidly freezing to - 70 "C in solid CO*/ propan-2-01, After thawing to room temperature the sample was immediately subjected to hydroxyapatite chromatography. Hydroxy- apatite-purified samples of ColEl and pX1212 DNAs from unshaken cleared lysates were heat denatured at 100 pg/ml in buffer A and chromatographed as above. The pX1212 sample had been 'aged' by many cycles of freezing and thawing. Samples were electrophoresed as in (A). Tracks (a) and (b) ColEl from cleared lysate before and after heat denaturation and chromatography, respectively; (C) ColEl from shaken cleared lysate. (d) and (g) purified ColEl and pX1212 respectively; (e) and (f) purified ColEl and pX1212 after heat denaturation and re-chromatography; (h) 1 DNA marker. The arrow marks the position of chromosomal DNA
ADNA, yet too large to be resolved on the gel. If during the lysis procedure more vigorous shaking was employed then more of these chromosomal fragments were released (compare Fig. 3 B, tracks c and d). Again these fragments co-purified with plasmid DNA.
The possibility that plasmid DNA could also be contaminated with low-molecular-weight heterologous DNA was excluded by the following experiment. A culture of ColEl-containing bacteria was grown in the presence of t3H]thymidine and 2 &ml 2-deoxy- adenosine for several generations before addition of chloramphenicol to the culture. This protocol was adopted in order to ensure that the specific activity of any released chromosomal DNA was equal, if not greater than that of ColEl DNA. Incubation was continued overnight and then a cleared lysate was
prepared. After agarose gel electrophoresis of aliquots of the cleared lysate and ColE1 DNA purified from it, slices of the gel were dissolved in 50% formamide and the radioactivity of each slice estimated. Fig.4 shows the radioactivity per slice as a function of the position of the slice in the gel. It was evident from the track containing purified plasmid DNA that radio- activity was only found in those regions containing ColEl DNA. The fast-moving band present in the track containing cleared lysate DNA, electrophoresed in the position of RNA, near the gel front. Since heterogeneous chromosomal DNA would occupy a rather wide region of the gel track it is evident that significant quantities of low-molecular-weight chro- mosomal DNA are not usually released during prepa- ration of cleared lysates.
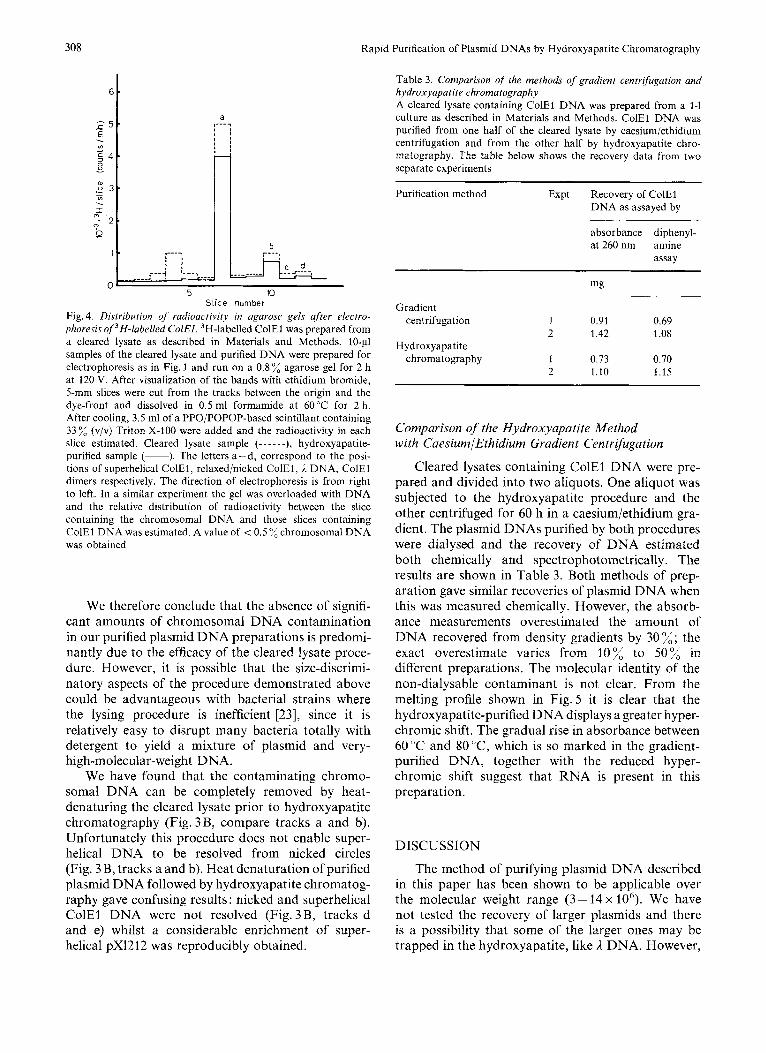
308 Rapid Purification of Plasmid DNAs by Hydroxyapatite Chromatography
I----
a
b --__ . .
Slice number Fig. 4. Distribution of radioactivity in agarose gels gfter electro- phoresis of 'H-labelled ColEI. 3H-labelled ColEl was prepared from a cleared lysate as described in Materials and Methods. 10-p1 samples of the cleared lysate and purified DNA were prepared for electrophoresis as in Fig. 1 and run on a 0.8 % agarose gel for 2 h at 120 V. After visualization of the bands with ethidium bromide, 5-mm slices were cut from the tracks between the origin and the dye-front and dissolved in 0.5 ml formamide at 60°C for 2 h. After cooling, 3.5 ml of a PPOiPOPOP-based scintillant containing 33 % (v/v) Triton X-100 were added and the radioactivity in each slice estimated. Cleared lysate sample (-- ----), hydroxyapatite- purified sample (-). The letters a-d, correspond to the posi- tions of superhelical ColE1, relaxedinicked ColE1, I DNA, ColEl diiners respectively. The direction of electrophoresis is from right to left. In a similar experiment the gel was overloaded with DNA and the relative distribution of radioactivity between the slice containing the chromosomal DNA and those slices containing ColEl DNA was estimated. A value of < 0.5 "/,chromosomal DNA was obtained
We therefore conclude that the absence of signifi- cant amounts of chromosomal DNA contamination in our purified plasmid DNA preparations is predomi- nantly due to the efficacy of the cleared lysate proce- dure. However, it is possible that the size-discrimi- natory aspects of the procedure demonstrated above could be advantageous with bacterial strains where the lysing procedure is inefficient [23], since it is relatively easy to disrupt many bacteria totally with detergent to yield a mixture of plasmid and very- high-molecular-weight DNA.
We have found that the contaminating chromo- somal DNA can be completely removed by heat- denaturing the cleared lysate prior to hydroxyapatite chromatography (Fig. 3B, compare tracks a and b). Unfortunately this procedure does not enable super- helical DNA to be resolved from nicked circles (Fig. 3 B, tracks a and b). Heat denaturation of purified plasmid DNA followed by hydroxyapatite chromatog- raphy gave confusing results : nicked and superhelical ColEl DNA were not resolved (Fig. 3B, tracks d and e) whilst a considerable enrichment of super- helical pX1212 was reproducibly obtained.
Table 3. Comparison of the methods of gradient centrifirgation and hydroxyapatite chromatography A cleared lysate containing ColEl DNA was prepared from a 1-1 culture as described in Materials and Methods. ColEl DNA was purified from one half of the cleared lysate by caesiumiethidium centrifugation and from the other half by hydroxyapatite chro- matography. The table below shows the recovery data from two separate experiments
Purification method Expt Recovery of ColEl DNA as assayed by
absorbance diphenyl- at 260 nni amine
assay
mg
Gradient centrifugation 1 0.91 0.69
2 1.42 1.08
chromatography 1 0.73 0.70 2 1.10 1.15
Hydroxyapatite
Comparison of the Hydroxyupatite Method with Cuesium 1 Et hidium Gradient Centrifugation
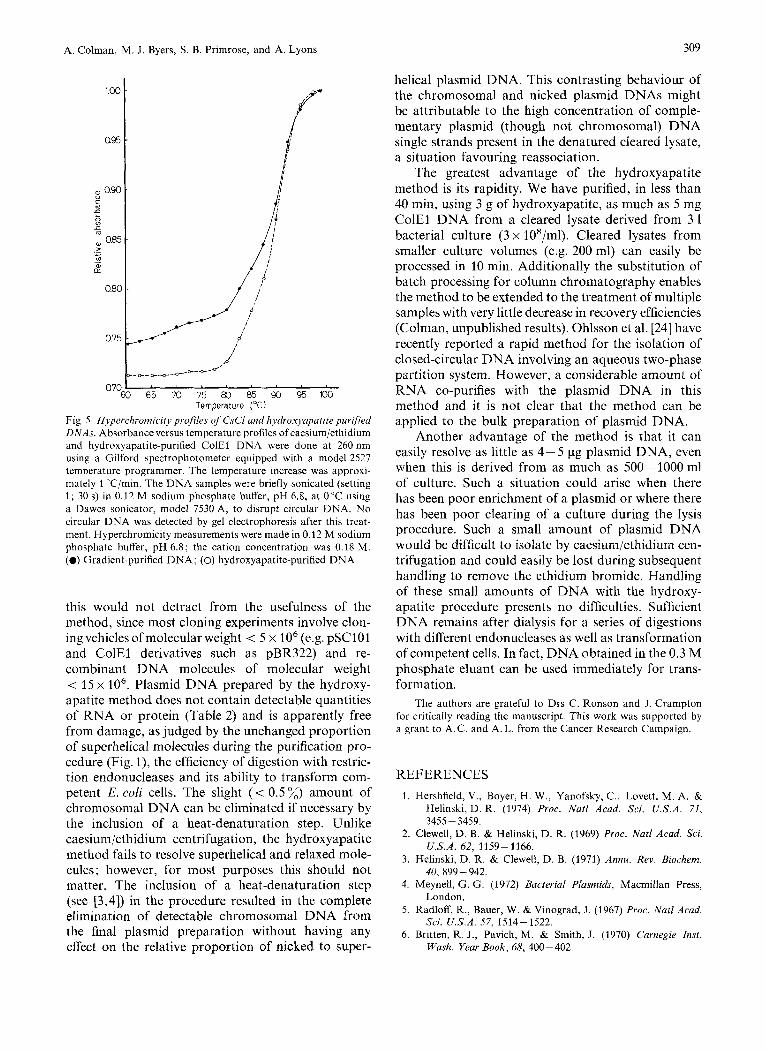
Cleared lysates containing ColEl DNA were pre- pared and divided into two aliquots. One aliquot was subjected to the hydroxyapatite procedure and the other centrifuged for 60 h in a caesium/ethidium gra- dient. The plasmid DNAs purified by both procedures were dialysed and the recovery of DNA estimated both chemically and spectrophotometrically. The results are shown in Table 3. Both methods of prep- aration gave similar recoveries of plasmid DNA when this was measured chemically. However, the absorb- ance measurements overestimated the amount of DNA recovered from density gradients by 30 % ; the exact overestimate varies from 10% to 50% in different preparations. The molecular identity of the non-dialysable contaminant is not clear. From the melting profile shown in Fig.5 it is clear that the hydroxyapatite-purified DNA displays a greater hyper- chromic shift. The gradual rise in absorbance between 60°C and 80°C, which is so marked in the gradient- purified DNA, together with the reduced hyper- chromic shift suggest that RNA is present in this preparation.
DISCUSSION
The method of purifying plasmid DNA described in this paper has been shown to be applicable over the molecular weight range (3- 14 x lo6). We have not tested the recovery of larger plasmids and there is a possibility that some of the larger ones may be trapped in the hydroxyapatite, like A DNA. However,
A. Colman, M. J. Byers, S. B. Primrose, and A. Lyons 309
070 I 60 65 70 75 80 85 90 95 100
Temperature ("C)
Fig. 5 . Hyperchromicity profiles of CsCl and hydroxyapatite purified DNAs. Absorbance versus temperature profiles of caesium/ethidium and hydroxyapatite-purified ColEl DNA were done at 260 nm using a Gilford spectrophotometer equipped with a model 2527 temperature programmer. The temperature increase was approxi- mately 1 "C/min. The DNA samples were briefly sonicated (setting 1 ; 30 s) in 0.12 M sodium phosphate buffer, pH 6.8, at 0°C using a Dawes sonicator, model 7530 A, to disrupt circular DNA. No circular DNA was detected by gel electrophoresis after this treat- ment. Hyperchromicity measurements were made in 0.12 M sodium phosphate buffer, pH 6.8; the cation concentration was 0.18 M. (0) Gradient-purified DNA; (0) hydroxyapatite-purified DNA
this would not detract from the usefulness of the method, since most cloning experiments involve clon- ing vehicles of molecular weight < 5 x lo6 (e.g. pSClOl and ColEl derivatives such as pBR322) and re- combinant DNA molecules of molecular weight < 15 x lo6. Plasmid DNA prepared by the hydroxy- apatite method does not contain detectable quantities of RNA or protein (Table 2) and is apparently free from damage, as judged by the unchanged proportion of superhelical molecules during the purification pro- cedure (Fig. l), the efficiency of digestion with restric- tion endonucleases and its ability to transform com- petent E. coli cells. The slight (< 0.5 %) amount of chromosomal DNA can be eliminated if necessary by the inclusion of a heat-denaturation step. Unlike caesium/ethidium centrifugation, the hydroxyapatite method fails to resolve superhelical and relaxed mole- cules; however, for most purposes this should not matter. The iiiclusion of a heat-denaturation step (see [3,4]) in the procedure resulted in the complete elimination of detectable chromosomal DNA from the final plasmid preparation without having any effect on the relative proportion of nicked to super-
helical plasmid DNA. This contrasting behaviour of the chromosomal and nicked plasmid DNAs might be attributable to the high concentration of comple- mentary plasmid (though not chromosomal) DNA single strands present in the denatured cleared lysate, a situation favouring reassociation.
The greatest advantage of the hydroxyapatite method is its rapidity. We have purified, in less than 40 min, using 3 g of hydroxyapatite, as much as 5 mg ColEl DNA from a cleared lysate derived from 3 1 bacterial culture (3 x 1O8/m1). Cleared lysates from smaller culture volumes (e.g. 200 ml) can easily be processed in 10 min. Additionally the substitution of batch processing for column chromatography enables the method to be extended to the treatment of multiple samples with very little decrease in recovery efficiencies (Colman, unpublished results). Ohlsson et al. [24] have recently reported a rapid method for the isolation of closed-circular DNA involving an aqueous two-phase partition system. However, a considerable amount of RNA co-purifies with the plasmid DNA in this method and it is not clear that the method can be applied to the bulk preparation of plasmid DNA.
Another advantage of the method is that it can easily resolve as little as 4 - 5 pg plasmid DNA, even when this is derived from as much as 500 - 1000 ml of culture. Such a situation could arise when there has been poor enrichment of a plasmid or where there has been poor clearing of a culture during the lysis procedure. Such a small amount of plasmid DNA would be difficult to isolate by caesium/ethidium cen- trifugation and could easily be lost during subsequent handling to remove the ethidium bromide. Handling of these small amounts of DNA with the hydroxy- apatite procedure presents no difficulties. Sufficient DNA remains after dialysis for a series of digestions with different endonucleases as well as transformation of competent cells. In fact, DNA obtained in the 0.3 M phosphate eluant can be used immediately for trans- formation.
The authors are grateful to Drs C. Ronson and J. Crampton for critically reading the manuscript This work was supported by a grant to A.C. and A. L. from the Cancer Research Campaign.
REFERENCES 1. Hershfield, V., Boyer, H. W., Yanofsky, C., Lovett, M. A. &
Helinski, D. R. (1974) Proc. Nut1 Acad. Sci. U.S.A. 71, 3455 - 3459.
2. Clewell, D. B. & Helinski, D. R. (1969) Proc. Natl Acad. Sci. U.S.A. 62, 1159-1166.
3. Helinski, D. R. & Clewell, D. B. (1971) Annu. Rev. Biochem.
4. Meynell, G. G. (1972) fiactrriai Plasinids, Macmillan Press,
5. Radloff, R., Bauer, W. & Vinograd, J. (1967) Proc. Natl Acad.
6. Britten, R. J . , Pavich, M. & Smith, J. (1970) Curnegie Inst.
40, 899 - 942.
London.
Sci. U.S.A. 57, 1514- 1522.
Wash. Year Book, 68, 400-402.

310 A. Colman, M. J. Byers, S. B. Primrose, and A. Lyons: Rapid Purification of Plasmid DNAs by Hydroxyapatite Chromatography
7. Heffron, F., Bedinger, P., Champoux, J. J. & Falkow, S. (1977)
8. Dougan, G., Saul, M., Warren, G. & Sheratt, D. (1978) Mol.
9. Primrose, S. B. (1976) J . Gen. Microbiol. 95, 159- 165.
Proc. Natl Acad. Sci. U.S.A. 74, 707-710.
Gen. Genet. 158, 325-327.
10. Inselburg, J . & Oka, A. (1975) J. Bacteriol. 123, 739-742. 11. Reeder, R. H., Higashinakagawa, T. & Miller, 0 . (1976) Cell,
12. Reference deleted. 13. Horiuchi, T. & Ohshima, Y. (1966) J . Mol. Biol. 20, 517-526. 14. Tiselius, A,, Hjerten, S. & Levin, 0. (1956) Arch. Biochem.
15. Bernardi, G. (1969) Biochim. Biophys. Acta, 174, 423-434. 16. Meinke, W., Goldstein, D. A. & Hall, M. R. (1974) Anal. Bio-
8,449 - 454.
Biophys. 65,132-145.
chem. 58, 82-88.
17. Burton, K. (1956) Biochem. J. 62, 315-323. 18. Peacock, A. D. & Dingman, C. W. (1968) Biochemistry, 7,
19. Johnson, P. H. & Grossman, L. J. (1977) Biochemistry, 16,
20. Lowry, D. H., Rosebrough, N. J., Farr, A. L. & Randall, R. J .
21. Fleck, A. & Munro, H. N. (1962) Biochim. Biophys. Arta, 55,
22. Colman, A. & Cook, P. R. (1 977) Eur. J . Biochcm. 76, 63 - 78. 23. Grindley, N. D. F., Humphreys, G. 0. &Anderson, E. S . (1973)
24. Ohlsson, R., Hentschel, C. C. & Williams, J. G. (1978) Nuctric
668 - 674.
421 7 -4225.
(1951) J . Biol. Chem. 193, 265-275.
571 - 583.
J. Bacteriol. 115, 387- 398.
Acids Res. 5, 583 - 590.
A. Colman, S. B. Primrose. and A. Lyons, Department of Biological Sciences, University of Warwick, Coventry, Warwickshire, Great Britain, CV4 7AL
M. J . Byers, La Jolla Cancer Research Foundation, La Jolla, California, U.S.A. 92037
Note Added in Proof. Our technique has now been successfully used for the isolation of plasmids from cleared lysates of Bacillus subtilis ( S . D. Ehrlich, personal communication) and Streptococcus luctis (F. L. Davies, personal communication). It has also proved possible to isolate the large plasmids RP4 and RP4: Mp by this method (S. B. Primrose, unpublished observations).
Related Documents











