- Rapid detection of bacterial infections with nuclease-activatable molecular imaging probes Bright-Smith, Franklin Oluwole https://iro.uiowa.edu/discovery/delivery/01IOWA_INST:ResearchRepository/12841202010002771?l#13841202000002771 Bright-Smith. (2019). Rapid detection of bacterial infections with nuclease-activatable molecular imaging probes [University of Iowa]. https://iro.uiowa.edu/discovery/fulldisplay/alma9984210642702771/01IOWA_INST:ResearchRepository Downloaded on 2022/06/04 00:46:56 -0500 Copyright 2019 Franklin Bright https://iro.uiowa.edu -

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

-
Rapid detection of bacterial infections withnuclease-activatable molecular imaging probesBright-Smith, Franklin Oluwolehttps://iro.uiowa.edu/discovery/delivery/01IOWA_INST:ResearchRepository/12841202010002771?l#13841202000002771
Bright-Smith. (2019). Rapid detection of bacterial infections with nuclease-activatable molecular imagingprobes [University of Iowa].https://iro.uiowa.edu/discovery/fulldisplay/alma9984210642702771/01IOWA_INST:ResearchRepository
Downloaded on 2022/06/04 00:46:56 -0500Copyright 2019 Franklin Bright https://iro.uiowa.edu
-

RAPID DETECTION OF BACTERIAL INFECTIONS WITH NUCLEASE-ACTIVATABLE MOLECULAR IMAGING PROBES
by
Franklin Oluwole Bright-Smith
A thesis submitted in partial fulfillment of the requirements for the Doctor of Philosophy degree in Molecular and Cellular Biology in the
Graduate College of The University of Iowa
December 2019
Thesis Supervisor: Associate Professor James O. McNamara

Copyright by
Franklin Bright
2019
All Rights Reserved

ii
In loving memory of my mother Monica Jobodee Smith and to my family and friends.

iii
What good is science to a man if he can’t apply it?
R. Marciano Street Religion

iv
ACKNOWLEDGEMENTS
I would like to take the time to thank everyone who has been a part of this long journey at
any capacity. The relationships that I have developed while at the University of Iowa have
impacted me in a profound manner. I would also like to thank my late mother, Monica, who has
been a major support system throughout my life. I am honored to have the privilege of being the
son of the kindest soul I’ve known. I’ll see you once my mission is complete.

v
ABSTRACT
Currently, bacterial infections are diagnosed symptomatically or by the screening of a
sample (tissue, urine, blood, etc.) collected in both hospital and clinical settings. Samples are
used to generate a microbial culture of the invading microbe followed by secondary culture-
based techniques including microscopy. Cultures can also be used to run biochemical tests, such
as the Gram stain or the acid-fast stain for identification. Additionally, collected samples can be
used for molecular diagnostics such as PCR. Although these methods have been the gold
standard for infectious disease diagnostics, they still have some significant drawbacks. When a
patient presents with a focal infection, a rapid diagnosis may be essential to survival. However,
current diagnostic methods can take days to weeks or more if there is a fastidious organism
involved in the infection. Current methods are also subject to false-negative test results, which
can further delay correct diagnosis, losing valuable time. To address these issues, this study
investigates a new method of identifying Staphylococcus aureus (S. aureus) which is the leading
cause of various types of focal infections in humans. Our group has carried out a previous study
using a molecular imaging technology that rapidly detects S. aureus infections in a mouse
pyomyositis infection model. This was accomplished with an activatable probe fluorescently
labeled in the near infrared (NIR) wavelength, to which a fluorophore and a quencher were
attached on opposite ends to a synthetic modified RNA oligonucleotide which is selectively
cleaved by the S. aureus micrococcal nuclease (MN). With this probe, we were able to rapidly
detect and localize the pathogen (via specific MN digestion of the oligonucleotide sequence) in
mouse thigh tissue. Despite the promising results from this study, a critical need for rapid
clinically relevant pathogenic infections remains. In this dissertation, we investigated whether a
second-generation probe could be used to rapidly detect a skin and catheter-associated biofilm

vi
infection by S. aureus in a noninvasive manner. We discovered that the DyLight800- P2 & P3
NIR probe could be used to detect a catheter-associated biofilm infection by S. aureus within 20
minutes of intravenous administration. However, the DyLight800 probe was not activated (with
significance) in the presence of a S. epidermidis, a nuclease-knockout S. aureus (mutant), or a
sham-infected catheter-associated biofilm infection. We discovered that the second-generation
probe labeled with fluorescein could be used to rapidly detect a skin infection by S. aureus.
Additionally, we report development and use of first-generation probe to rapidly detect a skin
infection by S. pyogenes (2nd leading cause of skin and soft tissue infections worldwide). Using
the two probes, we have also developed a multiplex imaging approach to detect a skin infection
by either S. aureus or S. pyogenes, the two most common pathogens in North America. The
findings from this study provide a novel approach to rapidly diagnosing two common classes of
S. aureus infections seen worldwide. Additionally, we believe this technology has great potential
to serve as a platform for other relevant classes of pathogenic infections.

vii
PUBLIC ABSTRACT
We previously described a novel molecular imaging technology capable of rapidly detecting S.
aureus (one of the most clinically significant pathogens worldwide) infections in mice. This was
accomplished with an activatable near infrared (NIR) fluorescent probe consisting of a Cy5.5
fluorophore and a quencher attached to opposite ends of a synthetic oligonucleotide (RNA)
sequence which is selectively cleaved by a secreted nuclease of S. aureus. Intravenous
administration of this probe enabled the rapid detection of S. aureus infections in mouse thighs
with noninvasive imaging. In order to apply this technology to more clinically relevant classes of
bacterial infections we optimized an approach with clinical applicability using a second-
generation probe. This included generating and testing a second-generation probe with a longer-
wavelength fluorophore (i.e., that absorb and emit light of >750 nanometers). Near infrared
wavelengths of >750 nanometers are known to penetrate tissues to a greater extent than the ~700
nanometer light required to image the Cy5.5 fluorophore, and thus enables deeper imaging of
tissues. However, the ability to robustly quench such “800 nm” fluorophores in vivo (a valuable
endpoint for this approach) has not been developed. In this dissertation, we describe the
DyLight800- P2 & P3 TT second generation quenched NIR fluorescent probe that can be used
instead of Cy5.5. Preliminary data showed quenching and nuclease-mediated activation in vitro
both in buffer and anticoagulated blood. We followed this with initial imaging experiments
which demonstrated fluorescence increases at S. aureus infection sites and sites of purified MN
injections in mice were observed. Here we report a second-generation probe to rapidly detect and
image animal models of two clinically significant forms of S. aureus infections (catheter-
associated biofilm and skin) in a noninvasive manner within 20 minutes of probe application.
Additionally, we report a multiplex imaging technology that allows rapid detection of S. aureus

viii
(leading cause of skin and soft tissue infections worldwide) and S. pyogenes (second leading
cause) skin infections. Rapid detection of each infection type is accomplished with high
specificity using these probes. Our imaging technologies address an unmet need and have
clinical translatability.

ix
TABLE OF CONTENTS
LIST OF TABLES ......................................................................................................................... xi
LIST OF FIGURES ...................................................................................................................... xii
CHAPTER I: THE CURRENT STATE OF DIAGNOSTIC MICROBIOLOGY AND FUTURE PERSPECTIVES .............................................................................................................1
Abstract ......................................................................................................................................1
Introduction ................................................................................................................................1
CHAPTER II: RAPID DETECTION OF S. AUREUS AND S. PYOGENES SKIN INFECTIONS WITH NUCLEASE-ACTIVATABLE IMAGING PROBES .................................9
Abstract ......................................................................................................................................9
Introduction ................................................................................................................................9
Materials and Methods .............................................................................................................11
Results ......................................................................................................................................17
Discussion ................................................................................................................................22
CHAPTER III: NIR PROBE CHARACTERISTICS IN WHOLE BLOOD...........................35
Abstract ....................................................................................................................................35
Introduction ..............................................................................................................................35
Materials and Methods .............................................................................................................36
Results ......................................................................................................................................38
Conclusion ...............................................................................................................................40
CHAPTER IV: NONINVASIVE IMAGING OF BIOFILM INFECTIONS BY S. AUREUS WITH SECOND-GENERATION NIR PROBE .....................................................48
Abstract ....................................................................................................................................48
Introduction ..............................................................................................................................48
Materials and Methods .............................................................................................................50
Results ......................................................................................................................................55

x
Discussion ................................................................................................................................59
CHAPTER V: CONCLUSIONS AND FUTURE DIRECTIONS ..........................................69
REFERENCES ..............................................................................................................................75

xi
LIST OF TABLES
Table 1. List of probe sequences used in Chapter II. .................................................................... 32
Table 2. Bacterial strains used in Chapter II. ................................................................................ 33
Table 3. NIR probe sequences used in Chapter III. ...................................................................... 47
Table 4. Bacterial strains used in Chapter IV. .............................................................................. 67
Table 5. Optimization of second-generation NIR probe concentration in Chapter IV. ................ 68

xii
LIST OF FIGURES
Figure 1. Selection of the P2&3 TT-variant sequence. ................................................................. 25
Figure 2. Selection of the 2’-FL self-hybridizing sequence. ........................................................ 26
Figure 3. Multiplex activation of FAM-P2&3 and ATTO565-2’-FL-SH probes. ........................ 27
Figure 4. Multiplex imaging of skin infections with FAM-P2&3 and ATTO565-2’-FL probes. ........................................................................................................................................... 28
Figure 5. Multiplex imaging activation ratios and statistical analysis. ......................................... 29
Figure 6. Infection detection and pathogenic identification via MP imaging algorithm. ............. 30
Figure 7. Skin infection pathology................................................................................................ 31
Figure 8. Unquenched NIR probes display enhanced fluorescence. ............................................ 41
Figure 9. Fluorescence emission/excitation spectrum of Cy5.5 unquenched probe. .................... 42
Figure 9. Blood fractionation revels modest contribution to enhanced fluorescence. .................. 42
Figure 10. Fluorescence emission/excitation spectrum of Cy5.5 unquenched probe. .................. 44
Figure 11. Correlation between probe concentration and fluorescence ........................................ 45
Figure 11. Correlation between probe concentration and fluorescence. ....................................... 45
Figure 12. Self-quenching of NIR probes. .................................................................................... 46
Figure 13. Cartoon illustration of oligonucleotide probe activation/digestion by MN. ................ 61
Figure 14. Selection of the second-generation NIR probe. ........................................................... 62
Figure 14. Selection of the second-generation NIR probe. ........................................................... 62
Figure 15. Characterization of the S. aureus biofilm infection model. ......................................... 63
Figure 16. Optimization of noninvasive imaging for biofilm infection model............................. 64
Figure 17. Enhanced fluorescence with Xen29 nuclease knock-out strain. ................................. 65
Figure 18. Statistical analysis for biofilm detection with second-generation NIR probe. ............ 66

1
CHAPTER I: THE CURRENT STATE OF DIAGNOSTIC MICROBIOLOGY AND FUTURE PERSPECTIVES
Abstract
Microbial infections are presently diagnosed symptomatically or by the screening of a
sample collection (tissue, urine, blood, etc.) in clinical settings. Samples taken are often used to
run secondary tests (i.e. culture, microscopy, biochemical and/or molecular diagnostic tests) to
identify the invading pathogen. Although these diagnostic methodologies have been the standard
for infectious disease diagnostics, they continue to pose significant issues. If a patient presents
with a focal infection, then a rapid and accurate diagnosis is essential to prevent complications
such as sepsis promoting the potential for positive patient outcomes. However, current diagnostic
tests take days to weeks for a definitive identification. Therefore, physicians treat empirically with
antibiotics which have contributed to growing resistance among pathogenic bacteria. This
introductory chapter will discuss the current gold-standard methodologies being used to diagnose
pathogenic infections, and then delineate recent novel rapid diagnostic technologies that are
currently in use or under development. A handful of these novel technologies pose the potential to
definitively identify a pathogen within a few hours (the average time of a hospital visit), whereas
definitive identification using the current methodologies take at least a day and more often days to
weeks or more until a positive diagnosis is confirmed.
Introduction
The infectious disease workflow typically begins when a patient shows up to the hospital
with an infection. Based on a composite of information (including physical examination, patient
symptoms, and risk factors), physicians confirm a bacterial infection then order more specific
laboratory tests to identify the invading pathogen that is the cause of infection. The primary

2
method of pathogen identification is through cell staining followed by direct examination under a
microscope [1].The staining procedure consists of particular dyes that are incorporated into the
pathogen cell wall in which yields a distinct color or cellular morphology distinguishable from
background. Using this procedure, clinicians can recognize the pathogen through certain
characteristics including color (following the staining procedure), size and morphology. A
common staining procedure known as, the Gram stain, utilizes contrasting colors in Gram-
positive(blue) and Gram-negative (red) bacteria to aid in narrowing down the identity of the
pathogen [2]. Although the Gram stain determines whether a pathogen is Gram-negative or Gram
positive, it cannot be used to decipher the specific genus and species of the invading pathogen.
Therefore, additional testing is required to garner species-specific information. Another widely
used identification technique is the culture-based method. This method allows the bacteria in
samples taken from patients to grow over time in selective media. Additional tests are then carried
out using the cultivated bacterial sample for further identification [1, 3, 4]. Therefore, a major
limitation of the culture-based methods is that it requires a considerable amount of time (hours to
weeks) for bacterial growth which is not usually available in a clinical setting. Additionally, many
pathogenic bacteria are difficult to culture in an in vitro setting allowing for the possibility of false
negative test results. There are also immunologic-based diagnostic tests that can identify
antibodies (from the patient) to the invading pathogen and/or antigens from the pathogen [3, 5, 6].
Due to the length of time it takes for a patient to produce antibodies during the infection process, it
would be difficult to use tests based on antibody production as an early or rapid diagnostic
approach [7, 8]. Molecular-based diagnostic tests (i.e. PCR) identify a pathogen based on its
genotypic information and can also provide additional information such as genes encoding
virulence enzymes, however many of the genetic-based molecular diagnostic tests possess a limit

3
of detection for microbes with low abundance, cannot detect unknown species, or are expensive
and extremely time consuming [8-13].
A more recent type of molecular-based diagnostic test is the matrix assisted laser
desorption/ionization time of flight mass spectrometry (MALDI-TOF MS). This technology can
identify a specific species within a bacterial community and can also distinguish between both
species and genus of bacteria based on specific spectral profile peaks [14, 15]. This diagnostic test
is considered to be relatively fast since it does not require bacteria to be cultivated, can be operated
in high-throughput fashion, and generates data that is easy to interpret based on spectral profiles
[16, 17]. Conversely, the MALDI-TOF MS diagnostic possesses a detection limit for
microorganisms with low abundance. In addition, clinical samples regularly consist of a high
amount of host proteins and normal flora which could potentially confound the spectral output [16,
18]. Although the overall operating cost of a MALDI-TOF instrument is relatively low (compared
to other molecular diagnostic test), there is a high initial cost for equipment and maintenance [19,
20].
Although the methodologies examined have become the gold-standard of infectious
disease diagnostics, the underlying limitation is time. If a patient presents with a microbial
infection, he/she does not leave the hospital visit with definitive knowledge of the invading
pathogen. Therefore physicians treat empirically with broad spectrum antimicrobials until a
definitive identification is accomplished which could potentially contribute to resistance [21].
Because of this, there is currently a great need for more rapid infectious disease diagnostics so that
medical teams can approach microbial infections with more targeted and precise antimicrobial
therapies. The next section of this chapter will examine some of the new optical imaging

4
approaches that are rapid and have the potential to replace (or supplement) the gold-standard
methods that are currently in use.
Currently, real-time infection detection has depended on nuclear imaging of a potential
infection with [18F] FDG (2-fluoro-2-deoxy-D-glucose) or radiolabeled leukocytes. In this
procedure, white blood cells fighting the infection are isolated and treated with [18F] FDG and then
injected back into patients followed by positron emission tomography (PET) imaging which
enables medical professionals to identify infection foci with whole body imaging [22]. Although
this procedure is said to be highly sensitive, there is low specificity for bacterial infections that
could potentially lead to an inaccurate diagnosis [22-29]. Additional limitations of this procedure
include the expense and time-consuming efforts of isolating and radiolabeling leukocytes, the
short shelf-life due to radionuclide decay, the fact that recovering leukocytes from
immunocompromised patients is not always feasible. Furthermore, not all bacterial infections
stimulate a strong neutrophil response, such as infections by Mycobacterium tuberculosis, Listeria
monocytogenes, and Salmonella. There is also concern about radiation exposure to both the
patient and medical staff using nuclear imaging [22].
Optical imaging is a promising new technology in the medical and research fields that uses
fluorescent light to detect biological processes within the body. In the context of an infection, a
reaction (usually enzymatic) takes place at the target (infection) site that leads to an increase in
fluorescence that can be detected using an imaging apparatus. The probes used for this type of
imaging are engineered to have a fluorescent molecule (i.e. GFP) and a fluorescence quencher on
opposite ends of a target substrate. When the probe encounters the target molecule at the infection
site, the substrate is degraded separating the fluorophore and quencher creating a fluorescence
increase. This process can be detected and imaged in real time [22, 30, 31].

5
Despite the potential for optical imaging to be used in conjunction with or as a replacement
for the current gold-standard methods for infection diagnostics, this technology has not yet been
FDA-approved for clinical use. The most important reason has to do with the poor signal-to-noise
ratios of the fluorophores used on the imaging probes that are currently available. They are excited
and emit light in the visible light spectrum (400-500 nm) range. At this range of light, tissue
absorption and autofluorescence are high and light is scattered. This makes it challenging to image
a potential infection site within the tissue. In recent years, probes have been engineered with Near
Infrared (NIR) fluorophores which are excited by and emit light at longer wavelengths (700-900
nm). The longer wavelength range penetrates tissue with lower light scattering, tissue absorption,
and autofluorescence enabling real-time imaging in vivo [32, 33]. So far, there is one FDA-
approved NIR fluorophore, indocyanine green (ICG), and it has been used in cancer treatment in
the context of image-guided surgery for tumor removal [34-37]. Other problems with imaging of
infections include the low sensitivity and specificity of currently available imaging probes and the
potential for cytotoxicity. To help circumvent these shortcomings, several novel NIR-labeled
imaging probes have been designed and studied by researchers with promising results in animal
models of infection [22, 38, 39]. Our previous study (the rational for the work provided in this
dissertation) as well as other fluorescence diagnostic imaging studies will be discussed in
remainder of this chapter.
Nucleases are a class of enzymes that degrade/digest oligonucleotide substrates via
phosphodiester bond cleavage. Our group has shown that several modified synthetic
oligonucleotide sequences confer resistance to mammalian nuclease cleavage while being
susceptible to cleavage by specific bacterial nucleases [22, 40]. One of the sequences (NMTT)
characterized was sensitive to MN (enzyme and virulence factor secreted by S. aureus)

6
degradation and resistant to serum nuclease degradation [31]. The sequence was further modified
by incorporation of a Cy5.5 (5’-end) NIR fluorophore and quenching groups (3’-end) on opposite
ends of the sequence creating a FRET (fluorescence resonance energy transfer) probe system. The
probe was subsequently administered intravenously to mice infected with pyomyositis and
remained silent until it encountered MN where it was digested and activated which facilitated
identification of S. aureus infections in vivo in less than one hour.
Another NIR FRET-based probe described by Kong et al. exploits the enzymatic properties
of β-lactamase, which is encoded by a variety of bacterial species conferring resistance to β-lactam
antibiotics “i.e. amoxicillin”, for pathogen identification purposes [22, 41-44]. The β-lactamase of
Mycobacterium tuberculosis (M. tuberculosis) was selected as the target enzyme for this probe
because it has higher activity levels and the structure differs from the β-lactamases of other
bacterial species [45, 46]. Several probes were evaluated in mice with subcutaneous M.
tuberculosis infections where they accumulated in activated form at the sites of infection. The
probe with the highest signal-to-noise ratio was further assessed in mice with M. tuberculosis lung
infections. The probe was shown to become activated and accumulate at the sites of infection with
high sensitivity and was not activated or accumulate in the lungs of uninfected animals [47].
Positively charged metals such as the zinc(ii)-dipicolylamine (Zn-DPA) complex do not
interact with healthy mammalian cells, but freely associate with the negatively charged
membranes of cells in both gram-positive and gram-negative bacteria [48, 49]. The Zn-DPA
complex has been modified with various NIR moieties to enable in vivo infection imaging [48, 50-
54]. Although this technology has shown promising results in animal models of S. aureus
infections [50-52, 54], it is considered a non-specific imaging as it was found to interact with
apoptotic cell membranes as well as necrotic mammalian cells [50-52, 54-56].

7
Lectin-binding proteins, such as Concanavalin A (Con A), have high affinity interactions
with sugars within cell walls of both gram-positive and gram-negative bacteria and have also been
utilized as optical imaging tools. Tang et al. developed a Con A NIR probe (Con A-IR750
nanoprobe) by linking the protein to a nanoparticle carrier and an IR750 fluorophore. This probe
was subsequently assessed in animal models of superficial wound and transplanted catheter
infections by S. aureus where it displayed adherence to sites of infection following application and
a wash step prior to fluorescence imaging [57].
Prothrombin is a blood protein involved in the clotting during host immune defense. S.
aureus can evade host immune systems through production of the staphylocoagulase protein that
binds prothrombin and sequesters the clotting process [58-60]. This condition was targeted for an
optical imaging approach in which a prothrombin analogue was conjugated to an Alexa Fluor 680
fluorophore (AF680-Pro-T probe) by Panizzi et al. This probe was shown to accumulate at sites of
coagulase-positive infections in an endocarditis animal model but not in coagulase-negative nor
un-infected controls [61].
Nitroreductases are a family of enzymes prevalent in both gram-positive and gram-
negative bacterial species. Therefore the activity of these enzymes have been the target of
diagnostic imaging procedures [62]. This has been accomplished by the development of a self-
quenching NIR dye (CytoCy5S) in which (the dye) is activated and emits fluorescence signal upon
reduction by nitroreductase [63]. This self-quenching NIR probe has been shown to identify
pathogens in various animal infection models including Salmonella typhimurium, Escherichia
coli, and Bifidobacterium breve [22].
As the field for clinical infection diagnostics advances, that both nuclear and optical
imaging have the potential to be used and could replace the time-consuming methods currently in

8
play. Nuclear imaging could be used in a patient with a suspected infection to identify the location
of infection. Specific optical probes could then be used to image the infected region or even to
image samples taken from the region ex vivo (for definitive identification). These procedures could
be carried out within the timeframe of a hospital visit and would provide useful information for
more targeted antibiotic therapies.

9
CHAPTER II: RAPID DETECTION OF S. AUREUS AND S. PYOGENES SKIN INFECTIONS WITH NUCLEASE-ACTIVATABLE IMAGING PROBES
Abstract
There is currently an unmet need for rapid and robust diagnostic tests for common skin
infections such as impetigo and cellulitis. As these infections are usually caused by S. aureus or
S. pyogenes, they are generally treated with empiric antibiotic therapy that is effective against
both bacterial pathogens. A diagnostic that could detect and specifically identify these pathogens
at suspected skin infections could enable the use of effective narrow spectrum antibiotics,
thereby reducing the unnecessary use of toxic drugs and enabling better antibiotic stewardship.
Here we report a multiplexed molecular imaging approach that rapidly detects and identifies S.
aureus and S. pyogenes skin infections via the enzymatic properties of their nucleases. Topical
application of two fluorogenic oligonucleotide substrates (probes) that are activatable by
nucleases of the target pathogens, to skin infections in mice, enabled detection of infections via
elevated fluorescence within an hour. Additionally, the ratios of the fluorescence levels of the
spectrally distinct probe fluorophores at infection sites provided robust pathogen-specific
signatures. This multiplexed molecular imaging approach can thus rapidly detect and identify the
causative pathogen in skin infections caused by the two most common bacterial skin pathogens.
Due to the distinct antibiotic susceptibilities of these pathogens, this approach has the potential to
effectively inform therapy for common types of infections that currently lack effective
diagnostics.
Introduction
Pathogenic skin and soft tissue infections (SSTIs) currently present a curative challenge
and have been a major clinical concern due to high morbidity and mortality rates [64, 65].
Staphylococcus aureus (S. aureus) and Streptococcus pyogenes (S. pyogenes) group A

10
streptococcus (GAS) are the primary and secondary SSTI pathogens respectively, worldwide [65-
71]. To prevent chronic SSTI conditions, rapid and accurate diagnosis is essential to provide
proper antibiotic therapy. However, current diagnostic methodologies are time consuming (taking
days to weeks for definitive identification) and are also prone to false-positive results.
Additionally, isolation of the invading SSTI pathogen is restricted to current methodologies which
obscure antibiotic selection for empirical treatment [31, 72-76]. Therefore, a novel technology to
detect both S. aureus and S. pyogenes skin infections in a rapid, sensitive, and specific manner
could be extremely beneficial to the infectious disease diagnostics field.
We previously reported a first generation nuclease-activatable probe that was used to
detect and image a mouse model of a pyomyositis (thigh muscle) infection by S. aureus [31]. The
probe consisted of a Cy5.5 near infrared (NIR) fluorophore and a quencher flanked on opposite
sides of a synthetic (ssRNA) oligonucleotide sequence sensitive to micrococcal nuclease (MN)
degradation. At 48-hours post infection (PI), the probe was administered through IV injection and
remained silent until it reached the S. aureus infection site where fluorescence activation signal
was observed within 30 minutes of administration.
Based on the findings in the previous study, we postulated that the novel technology could also be
applied to a more clinically relevant class of infection such pathogenic skin infections. Herein we
report the second generation activatable probe to rapidly detect and image a S. aureus skin
infection and a first generation activatable probe to rapidly detect and image a S. pyogenes skin
infection both in mouse models of bacterial skin infections. A multiplex diagnostic approach was
used in this study where one probe (for S. aureus detection) sequence was labelled with a

11
fluorescein (green) fluorophore and the other (for S. pyogenes detection) was labelled with an
ATTO-565 (red) fluorophore. Both probes were reconstituted together in phosphate buffered
saline (PBS) and hydroxyethylcellulose (HEC) into a gel that was applied topically to either an S.
aureus or S. pyogenes skin infected mouse 24 hours PI. In less than 30 minutes of gelled-probe
application, green fluorescence activation signal was observed for S. aureus-infected animals with
minimal activation observed in the red channel, whereas red fluorescence activation signal was
observed for S. pyogenes-infected animals with no activation observed in the green channel. The
results from this chapter provide a novel optical imaging approach to rapidly diagnose the top 2
skin-invading pathogens and displays a high degree of translatability to clinical settings.
Materials and Methods
Quenched fluorescent oligonucleotide probe synthesis. All probes used in this study
were synthesized, purified (HPLC), and lyophilized by Integrated DNA Technologies (IDT).
Lyophilized probes were reconstituted in TE (10 mM Tris-HCl pH 8.0, 1 mM EDTA) at 500 µM
concentration and stored in aliquots at -80°C.
In vitro assessment of oligonucleotide probes with plate reader assays. Fluorescence
plate reader assays were carried out as previously described [40] with a Synergy™ Mx
Microplate reader. Nuclease reactions were prepared in 10 µl volumes with 1 µl of 50 µM of
each probe combined with the nuclease-containing samples indicated in the figure legends.
Micrococcal nuclease was obtained from Worthington Biochemical Corporation. Reactions were
incubated in 0.5 ml Eppendorf LoBind Tubes at 37°C for 1 hour. Following the incubation
period, 290 µl PBS without divalent cations supplemented with 10 mM EDTA 10 mM EGTA
was added to each reaction tube. 95 µl portions were then transferred in triplicate to 3 wells of 96
well plates. Fluorescence levels were measured and then analyzed using Excel.

12
In vitro kinetic evaluation of probe variants. Kinetic evaluation of probe digestion was
carried out with 2.5 µM probe concentrations in DPBS with divalent cations (Gibco), 0.000625
units/µl micrococcal nuclease (Worthington Biochemical) in 10 µl volumes in 384-well plates
using a Biotek Synergy Mx fluorescence plate reader. Plates were covered with optical film to
prevent evaporation, incubated at 37 °C and fluorescence was measured with excitation of
485/20 and emission at 528/20 every 5 minutes for an hour. Plates were shaken briefly prior to
each measurement. Background was measured with mock reactions that were prepared in the
same manner, but with no nuclease. Control reactions were used to define the fluorescence signal
of complete probe digestion. These were prepared as described above, but with 1 unit/µl
micrococcal nuclease (MN). All reactions were prepared in protein LoBind tubes (Eppendorf)
prior to transfer to the 384-well plates. For analysis, experimental values were normalized to
positive control values after background subtraction. Averages of experimental replicates with
standard deviations are shown in the results.
Genetic manipulation of S. aureus. The strains of S. aureus used in this study were
engineered to be luminescent as previously described and the nuclease-1 mutant S. aureus strain
(AH2672) was previously created by the Horswill laboratory (University of Colorado
Department of Immunology and Microbiology) [31]. Additionally, the nuclease 1 and nuclease 2
mutant S. aureus strain (AH3086) was created by Dr. Heidi Crosby from the Horswill laboratory
for this study.
Bacterial culture conditions for in vitro assays. All bacteria were cultured from frozen
stocks in the appropriate media and growth conditions (Table 2). In order to prepare cultures for
activation assays, overnight cultures were grown at 37°C with shaking at 200 r.p.m; log-phase
cultures were used for some experiments (see figure legends). Growth conditions for the

13
streptococcal strains required stationary growth at 37°C in an incubator supplemented with 5%
C02. To prepare consumed media, 5-10 ml of the cultures were centrifuged at 6,000 g for 10
minutes. The supernatants were then collected. For S. pyogenes and WT S. aureus Newman
strains, Roche© cOmplete ULTRA protease inhibitor was added to the supernatant immediately
after it was collected. For some experiments, culture supernatants were dialyzed prior to use as
input in nuclease assays (see figure legends for details).
Conditions for culture supernatant specificity screen- Cultures were grown in 5 mL
with the broth and growth conditions indicated in table 2. 1 ml of each overnight culture was
spun down at 6,000 g for 10 minutes followed by collection of the supernatant. The remainder of
the culture was then sub-cultured in fresh broth (1:50 dilution) for ~2 hours until the OD600
reached mid-log phase (~0.5). Mid-log supernatants were then collected with centrifugation
followed by use in activation assays.
Human skin lysate probe digestion assay. Human skin whole tissue lysate (Novus
Biologicals) was centrifuged at 10,000 g for 10 minutes at 4°C to remove any particulate
material. The supernatant was then aliquoted into 50µl volumes and stored at -80°C. Prior to use,
the skin lysate was dialyzed in a microdialysis cassette (Pierce) against DPBS (with divalent
cations), 1% Triton X-100 with EDTA-free protease inhibitor (Roche). 5 uM of either the FAM-
P2 & P3 TT probe or the ATTO565-2’-FL-SH probe was used for each 10 ul reaction, which
incubated for 60 minutes at 37°C. Reactions were next stopped with 290µl stop buffer (10mM
EDTA/10mM EGTA/PBS with no divalent cations) and plated in triplicate wells of a 96W
microplate. Plate was read on SynergyMx at the following settings: For the P2 & P3 TT probe-
Excitation: 485/20 nm; Emission: 528/20 nm. For the 2’-FL-SH probe- Excitation: 563/20 nm;
Emission: 592/20 nm. The source of the skin lysate used in this study was from a normal skin

14
donor who was involved in a car accident, it corresponds to a 34 year-old Asian male. Tissue
was taken (postmortem) from the upper arm portion of the body where the lysate sample was
extracted from the dermis of the skin.
S. pyogenes skin infection model. The S. pyogenes strain used in the infection model
was chosen because of its ability to cause robust localized infections with subsequent immune
responses in various models of animal infections [77-82]. Cultures were prepared for intradermal
injections in the following fashion. First, overnight cultures (~13.5 hours of growth) of HSC5
strain (Caparon lab, Washington University) were grown in Todd-Hewitt Broth with yeast
extract at 37°C to stationary phase in an incubator supplemented with 5 % C02. A subculture
(starting with OD600= 0.05) was grown for ~2 hours or until the bacteria went through two
population doublings (OD600=0.2). Next, the culture was washed once and re-suspended in cold
PBS with divalent cations to an inoculum of ~4 X 106 CFU in 50 microliters for intradermal
injections into the right flank of 6-8 week old hairless SKH1 female mice under isoflurane
anesthesia. As a control, 50 microliters of PBS with divalent cations was injected intradermally
into the left flank. Fluorescence imaging through fur and/or shaved (nicks from shaving-
compromised skin surface) is a technical challenge, therefore SKH1 mice were used in this
chapter (and Chapter IV) as this strain is ideal for in vivo imaging applications.
S. aureus skin infection model. Cultures were prepared for intradermal injections in the
following fashion. First, overnight cultures of lux+ MN-expressing WT S. aureus Newman strain
were grown in TSB supplemented with kanamycin (50 µg ml-1) at37°C with shaking at 200
r.p.m. A 1:100 diluted subculture was then grown for another 2 hours. Following 2-hour growth,
cultures were washed once then re-suspended in cold PBS with divalent cations to an inoculum
of ~1x108 CFU in 50 microliters to inject intradermally into the right flank of 6-8 week old

15
SKH1 female mice under isoflurane anesthesia. As a control, 50 microliters of PBS with divalent
cations was injected intradermally into the left flank.
Fabrication of rubber imaging patches. Small rubber rings made from FDA-grade
white rubber were attached to adhesive paper and colored black with a sharpie marker to limit
autofluorescence of the rubber material.
In vivo assessment of nuclease-activated probes. For skin infections, fluorescence
imaging was carried out using the Xenogen IVIS 200 imaging system (1-second exposure time).
For all luminescence imaging carried out (prior to and/or following fluorescence imaging), a
Xenogen IVIS 200 imaging system (1-minute exposure time) was used. Mice were first
anesthetized with 2 % isoflurane gas and then placed on the imaging platform. Fluorescence
images were acquired (with GFP or DSred filter settings) prior to and following gelled probe
topical application on the ethanol-wiped skin sites with specific time points denoted on the
figures. Living Image® 2.50.1 software was used for image acquisitions, processing, and
analysis.
2.5 nanomoles of either the ATTO565-2’-FL-SH and FAM-P2&3 TT probe was
reconstituted in 995 µl PBS with divalent cations (separately for pathogen-specific imaging or
together for multiplex imaging skin infections) and mixed with 30 mg Ashland Natrosol for the
gelling process. Once the reconstituted probe(s) began to gel, it was placed in a 1 mL syringe,
foiled and left at room temperature overnight to continue the gelling process. The next day ~ 100
µl of the gelled probe was applied on the skin lesion as well as the contralateral control site
following control images taken prior to probe application.
Imaging and processing of skin lesions (CFU, histology, and pathology) were carried out

16
at 24 hours post-infection. All animal experiments were approved by the University of Iowa
Institutional Animal Care and Use Committee (IACUC).
Target-to-background measurements. Region of interest (ROI) analysis was carried
out using the Living Image 2.50.1 software. Fluorescence intensity was measured within the ROI
for S. aureus or S. pyogenes –infected tissue and fluorescence intensity was also measured within
the ROI on the contralateral side for un-infected tissue (PBS injection) for each time point
recorded as denoted on the figures. Time “0” post probe application ROI value was subtracted
from all timepoints as a background subtraction method for each experiment.
Infection CFU counts. Following fluorescence imaging of skin infections, mice were
sacrificed, and infected tissue was carefully dissected and put into 1 ml PBS with divalent
cations in 1.5 ml microcentrifuge tubes. Tissue was then homogenized with a Biospec Products,
Inc. Tissue Tearor™. This tube was marked as 10-1 and five subsequent 10-fold dilutions were
carried out (10-1, 10-2, and so forth.) prior to plating 50 ul from each tube onto TSA or THY
plates. Plates were then incubated at 37°C overnight and CFU counts were made the following
day.
Imaging and processing of skin lesions (CFU, histology, and pathology) were carried out 24
hours post-infection.
Statistical analysis. RFU values (n=8 S. aureus-infected mice and n=6 S. pyogenes-
infected mice) from the 55-minute timepoint post probe application (background subtracted)
were analyzed with two-way ANOVA Sidak’s multiple comparisons test (infected skin
compared to healthy skin in both the green and red channels) using Graphpad Prism 7. S. aureus-
infected mice (P < 0.0035), S. pyogenes-infected mice (P < 0.0001).
Histology and pathology of skin lesions. Mice were sacrificed 24H PI via carbon

17
dioxide intoxication. The skin lesion on the right flank was carefully dissected along with control
(PBS-injected) skin on the left flank for comparison and placed in 10% neutral buffered formalin
at room temperature for 3-5 days. Next, the fixed skin tissue was gross sectioned and processed
through a series of alcohol/xylene baths. Tissue sections were made (~4-µm) on glass slides and
stained with hematoxylin and eosin (H&E) or Gram stain as previously described [83]. All slides
were examined by a veterinary pathologist (D.K.M.) for histopathologic analysis using principles
for valid histopathology [84]. Digital images were acquired with a high-resolution Olympus
DP73 camera mounted to an Olympus BX53 microscope using Olympus CellSens Software.
Results
In this study, we sought a means of specifically detecting the two most common bacterial species
that cause many types of skin infections, S. aureus and S. pyogenes, via specific detection of
their nuclease activities. A clinical assay with such a capability could inform therapy due to the
distinct antibiotic susceptibility profiles of these species. We envisioned a simple diagnostic
method in which fluorogenic substrates for S. aureus and S. pyogenes nucleases would be
topically applied to skin infections followed by fluorescence imaging to detect the presence of
the bacteria via increases in fluorescence. Topical application would have the advantage of
circumventing the variability inherent in specimen collection. To detect and distinguish the target
species, fluorogenic oligonucleotide substrates that selectively respond to nucleases of each
species, but that are also resistant to relevant endogenous human nucleases are needed.
Chemically modified oligonucleotides that are sensitive to micrococcal nuclease (MN)
and resistant to mammalian serum nucleases were previously identified. These oligos were
coupled to fluorescein amidite (FAM) (a green fluorophore) and ZEN and Iowa Black RQ
quenchers on opposite ends, yielding a “TT probe” that includes a pair of unmodified

18
deoxythymidines flanked by several 2’-O-methyl modified nucleotides [31, 40, 85, 86]. Nuclease
digestion of the oligonucleotide portion enables the quenchers to diffuse from the fluorophore,
which then exhibits fluorescence. We began here by further optimizing the TT probe. We tested
11mer oligo compositions (quenched probe format) where the pair of deoxythymidines were
placed at various positions along the sequence with all other nucleotides in the sequence being 2’-
O-methyl modified uridines. These sequences were named according to the position of the TT
(Table 1). The MN activation kinetics of the positional variants were then tested against the first-
generation TT probe (Fig. 1A). As shown in the figure, the P2&3 TT probe is the most sensitive to
MN digestion. Interestingly, it appears that MN preferentially digests sequences with the
deoxythymidine cut site closer to the 5’-end. To determine specificity of the second-generation
P2&3 TT probe, in vitro activation assays were carried out using culture supernatants of various
skin pathogens (Table 2, Figure 1B). Besides S. aureus, one species (S. caprae) appeared to
moderately activate the probe while another (S. capitis) did so to a lesser extent; however, neither
of these species is commonly associated with skin and soft tissue infections [87-89]. We also
found that the P2&3 TT probe is resistant to human skin nucleases (Fig. 1C).
We next tested FAM-labeled self-hybridizing oligonucleotides (i.e., double-stranded
substrates) with several nucleotide chemistries to identify a suitable probe for S. pyogenes.
Incubation with a S. pyogenes culture supernatant revealed a 2’-fluoro modified self-hybridizing
probe (2’-FL-SH) as sensitive to S. pyogenes nucleases (see Fig. 2A). Although a similar DNA
probe was also sensitive to S. pyogenes nucleases, the 2’-FL-SH probe was selected for further
evaluation because the 2’-fluoro modification is known to yield broader resistance to mammalian
nucleases. The FAM fluorophore (green) used for this probe in Figure 2A was exchanged for the

19
ATTO565 fluorophore (red) in order to enable subsequent multiplexing with the FAM-labeled
P2&3 TT probe.
The specificity of the 2’-FL-SH probe with respect to distinct skin pathogens was
evaluated by incubation with culture supernatants of various pathogenic bacterial species (Fig.
2B). Here, we observed that the 2’-FL-SH probe is activated by nucleases in a S. pyogenes
supernatant and to a lesser extent in S. pneumoniae supernatant, but is resistant to nucleases
present in several alternative bacterial species, including several species that can cause skin
infections [90-94]. Overall, these data suggest the 2’-FL-SH probe is quite specific for S.
pyogenes.
As for the S. aureus detection probe, a probe that would detect S. pyogenes with topical
application to skin infections must be resistant to the many endogenous nucleases found on human
skin [95]. We therefore tested 2’-FL-SH probe stability in the presence of a human skin lysate,
using S. pyogenes culture supernatant as a positive control for probe digestion (Fig. 2C). The
probe was clearly stable in the skin lysate indicating its resistance to human skin nucleases.
To assess the ability of the P2&3 TT and 2’-FL-SH probes to function in a multiplexed
format, in vitro nuclease detection assays were carried out with reaction mixtures that contained
both the FAM-labeled P2&3 TT (green) probe and the ATTO565-labeled 2’-FL-SH (red) probe
and culture supernatants of either S. aureus or S. pyogenes (Fig 3). Fluorescence in the green
channel indicated P2&3 TT probe activation in the presence of S. aureus culture supernatant, but
not in the presence of S. pyogenes culture supernatant. In contrast, fluorescence in the red channel
indicated 2’-FL-SH probe activation in the presence of S. pyogenes culture supernatant, but not in
the presence of S. aureus culture supernatant. This result demonstrates the functionality of a
multiplexed nuclease-detection format that enables detection and specific identification of S.

20
aureus and S. pyogenes. Similar results were obtained when several different S. pyogenes strains
were tested (data not shown).
Our next step was to determine if this multiplexed format could enable detection of skin
infections by either pathogen in a rapid manner by imaging a topically applied probe mixture. Skin
infections were produced by intradermal injections of bacteria in the flanks of SKH1 mice (Fig. 4
A, C). The S. aureus strain used here was engineered to produce blue luminescence by
incorporation of the lux operon of Photorhabdus luminescens, enabling confirmation of bacterial
presence and location via luminescence imaging [31, 96-98]; the skin lesions from this strain
displayed luminescence that co-localized with gross clinical pathology (Fig. 4A). The FAM-P2&3
TT and the ATTO565-2’-FL-SH probes were combined at 2.5 µM each in a gelled-matrix (see
methods) and applied to infection sites and contralateral control sites (PBS-injected) 24 hours after
bacterial inoculation. Fluorescence at the infection site was detected as early as 10 minutes after
probe application in the green channel (P2&3 TT probe activation), while fluorescence increases
were also apparent in the red channel (Fig. 4B). Intradermal skin lesions in mice from S. pyogenes
displayed similar gross pathology 24 hours post-inoculation (Fig. 4C). The same multiplexed
imaging procedure was carried out with the S. pyogenes-infected mice and yielded fluorescence at
the infection site as early as 10 minutes after probe application in the red channel (2’-FL-SH probe
activation). In contrast to the S. aureus infections, the off-target probe (P2&3 TT probe) was not
digested in the context S. pyogenes infections (Fig. 4D).
In order to analyze infection imaging data accurately, background subtracted RFU values
on infected and healthy skin sites were calculated at the 55-minute post probe application (PPA)
time point. Statistical analysis of S. aureus-infected mice revealed a significant difference in the
green channel (P2&3 probe activation) and not the red channel (2’-FL-SH probe activation) when

21
infected skin sites were compared to healthy skin sites (Fig. 5A). Analysis of the S. pyogenes-
infected mice revealed a significant difference in the red channel (2’-FL-SH probe activation) and
not the green channel (P2&3 probe activation) when infected skin was compared to healthy skin
(Fig. 5B).
Background subtracted fluorescence values at the 55-minute PPA time point for each of
the S. aureus-infected mice in this study (Fig. 5C) indicate the ability of the multiplexed probes to
detect both pathogens, but also reveal the contrasting substrate specificities of the S. aureus and S.
pyogenes nucleases. Whereas the S. pyogenes nuclease activity is selective for the 2’-FL-SH
probe, the S. aureus nuclease activates both probes, but with a preference for the P2&3 TT probe.
The infection site fluorescence levels can be used to clearly distinguish infected from
healthy skin. For instance, fluorescence values that exceed either of the thresholds for the red and
green channels shown in Figure 5C include all of the infected but none of the healthy skin sites.
Further, a simple examination of the green fluorescence level relative to the green threshold can
clearly identify S. aureus infections. However, elevated red fluorescence could indicate either
pathogen. We next considered whether the relative fluorescence levels of the two channels might
provide pathogen-specific signatures. Indeed, ratios of the background subtracted fluorescence
levels provided a robust means of identifying the pathogens (Fig. 5D). For instance, of the
infected skin sites displaying well above red or green fluorescence thresholds, those with green/red
channel fluorescence ratios of greater than 0.5 include all the S. aureus-infected sites and none of
the S. pyogenes-infected sites (Fig. 5D-left panel). Likewise, infected skin sites with red/green
channel fluorescence ratios greater than 2 include all of the S. pyogenes-infected sites and none of
the S. aureus-infected sites (Fig. 5D-right panel). The above analysis can be reduced to a simple

22
diagnostic algorithm (Fig. 6) with the outcome being a straightforward means of detecting and
identifying the bacterial pathogens that are the predominant causes of common skin infections.
Histologic sections of both skin infection types revealed similar pathologies including
inflammation, bacterial colonization, and tissue necrosis (Fig. 7). Taken together, these data
indicate the ability of a novel bacterial nuclease-detecting multiplexed approach to specifically
detect the presence of S. aureus and S. pyogenes in skin infections.
Discussion
Currently, there are no licensed optical imaging probes for clinical detection of pathogenic
infections [22]. Diagnosing bacterial infections primarily depends on nonspecific tests in addition
to infected-tissue collection and culture, which takes a considerable amount of time. In this study,
we developed two nuclease-activatable optical imaging probes to noninvasively detect S. aureus
and S. pyogenes skin infections in mice within 60 minutes using a multiplexed approach.
The second-generation FAM-P2&3 TT probe is a sequence variant of the previously
described TT Probe [31], and was selected due to its higher sensitivity to MN digestion. The
ATTO565-2’-FL-SH probe was selected due to its sensitivity to S. pyogenes culture supernatant
digestion and resistance to mammalian nucleases. The activation of the FAM-P2&3 TT probe by
S. aureus culture supernatants and infected-skin is the result of MN production as minimal probe
activation was seen in experiments where no MN was present. In contrast, the activation of the
ATTO565-2’-FL-SH probe was attributed to S. pyogenes nuclease activity in the context of culture
supernatants and to S. pyogenes or S. aureus in the context of infected-skin. Significantly, both
skin imaging probes are resistant to degradation by endogenous mouse and human nucleases,
which is likely the result of the 2’-O-methyl modifications in the P2&3 TT-variant and the 2’-
flouro modifications in the 2’-FL-SH probes as both modifications are known to confer a high

23
degree of resistance to serum nuclease digestion [31, 40, 86]. Additionally, it appears that the base
modifications to both probes enable bacterial specificity as each probe was primarily activated in
the presence of its targeted bacterial nuclease versus nucleases of other pathogens tested. The
nuclease specificity of the probes provides the basis for the identification of the bacterial species
present in infections.
The non-specific digestion of the 2’-FL-SH probe by MN indicates this enzyme exhibits
less substrate specificity than the nuclease(s) of S. pyogenes. To provide a high level of clarity
when using this multiplex imaging technology, we created a 2-step algorithm that first deciphers
between infected and uninfected (healthy) skin. The subsequent step distinguishes between S.
aureus and S. pyogenes infections. The entire process can be carried out in less than 60 minutes.
Clinically, cellulitis infections are found to be caused by S. pyogenes (GAS) and S. aureus
at an equal rate [99]. Definitive identification of the causative species cannot be made within the
length of time of an outpatient visit. Therefore, physicians treat with broad spectrum coverage
including coverage against MRSA strains. Trimethoprim/sulfamethoxazole (TMP-SMX) is used
to cover MRSA strains in combination with beta-lactam antibiotics (clindamycin if more
aggressive) to cover GAS strains. Because the multiplex imaging technology described in this
study occurs within 60 minutes, it could enable physicians to provide more targeted antibiotic
therapies. Additionally, a confirmation that S. aureus was not involved in the infection would steer
therapy selection away from drugs with more severe side effects.
The novel imaging approach described in this study has an ease of translatability into
clinical use. The probes are believed to be nontoxic, and the agent used to gel the probes for
topical application (HEC) is currently used clinically [100]. More importantly, since this approach
involves a topical gel application, the likelihood of systemic toxicity is far diminished as the

24
probes can be wiped from the skin following the imaging procedure. There is also the possibility
of differentially labeling other modified sequences to target nucleases of different bacterial
pathogens allowing the potential for identification of polymicrobial infections. In summary, the
technology in this study has the potential to address an important unmet clinical need in the
management of common infections that are currently treated with empiric antibiotic therapy.
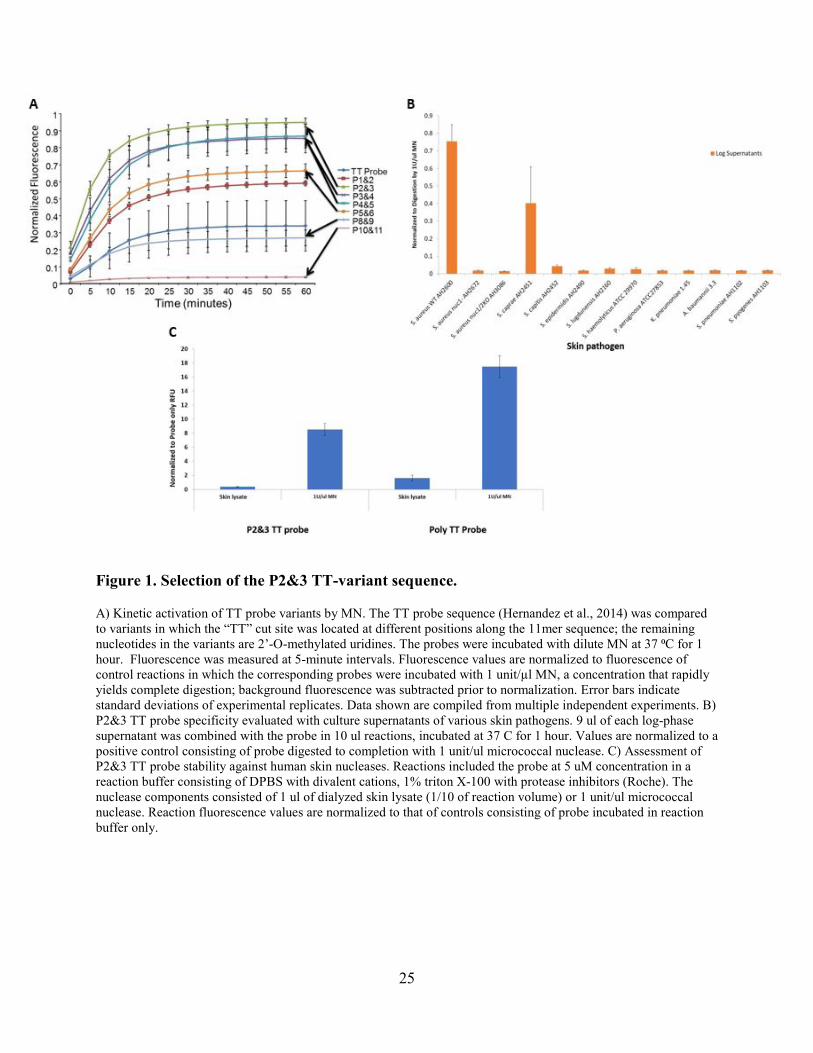
25
Figure 1. Selection of the P2&3 TT-variant sequence.
A) Kinetic activation of TT probe variants by MN. The TT probe sequence (Hernandez et al., 2014) was compared to variants in which the “TT” cut site was located at different positions along the 11mer sequence; the remaining nucleotides in the variants are 2’-O-methylated uridines. The probes were incubated with dilute MN at 37 ⁰C for 1 hour. Fluorescence was measured at 5-minute intervals. Fluorescence values are normalized to fluorescence of control reactions in which the corresponding probes were incubated with 1 unit/µl MN, a concentration that rapidly yields complete digestion; background fluorescence was subtracted prior to normalization. Error bars indicate standard deviations of experimental replicates. Data shown are compiled from multiple independent experiments. B) P2&3 TT probe specificity evaluated with culture supernatants of various skin pathogens. 9 ul of each log-phase supernatant was combined with the probe in 10 ul reactions, incubated at 37 C for 1 hour. Values are normalized to a positive control consisting of probe digested to completion with 1 unit/ul micrococcal nuclease. C) Assessment of P2&3 TT probe stability against human skin nucleases. Reactions included the probe at 5 uM concentration in a reaction buffer consisting of DPBS with divalent cations, 1% triton X-100 with protease inhibitors (Roche). The nuclease components consisted of 1 ul of dialyzed skin lysate (1/10 of reaction volume) or 1 unit/ul micrococcal nuclease. Reaction fluorescence values are normalized to that of controls consisting of probe incubated in reaction buffer only.
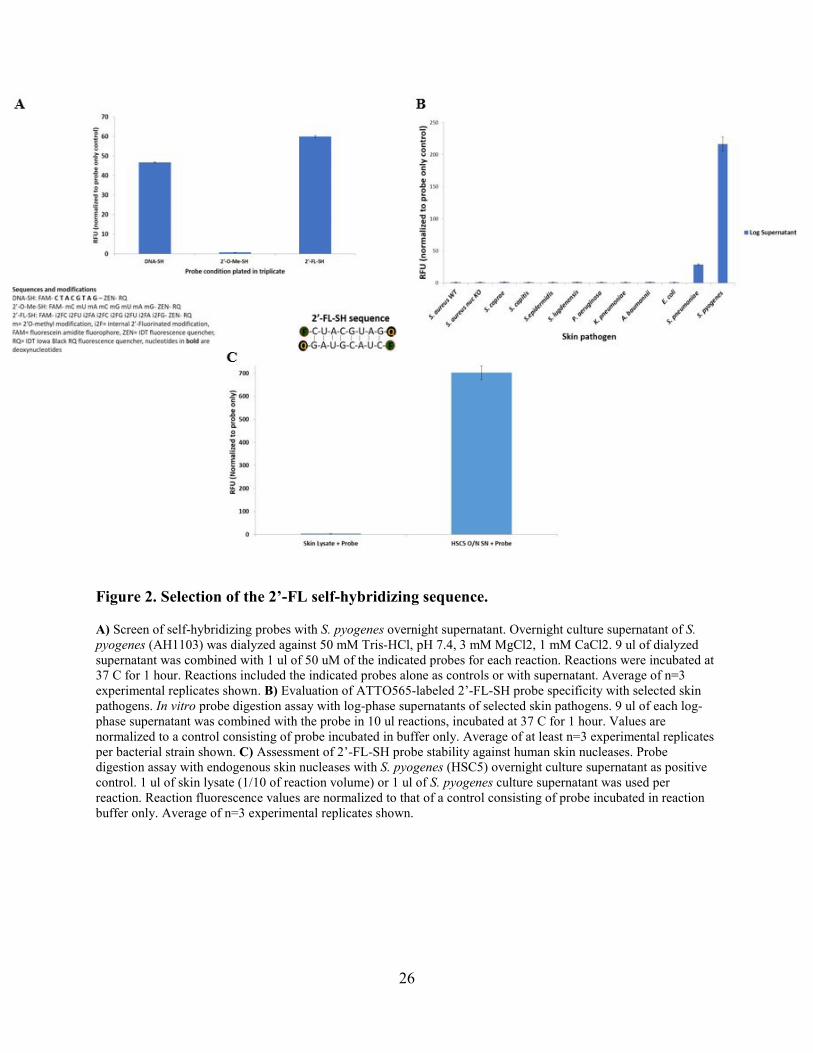
26
Figure 2. Selection of the 2’-FL self-hybridizing sequence.
A) Screen of self-hybridizing probes with S. pyogenes overnight supernatant. Overnight culture supernatant of S. pyogenes (AH1103) was dialyzed against 50 mM Tris-HCl, pH 7.4, 3 mM MgCl2, 1 mM CaCl2. 9 ul of dialyzed supernatant was combined with 1 ul of 50 uM of the indicated probes for each reaction. Reactions were incubated at 37 C for 1 hour. Reactions included the indicated probes alone as controls or with supernatant. Average of n=3 experimental replicates shown. B) Evaluation of ATTO565-labeled 2’-FL-SH probe specificity with selected skin pathogens. In vitro probe digestion assay with log-phase supernatants of selected skin pathogens. 9 ul of each log-phase supernatant was combined with the probe in 10 ul reactions, incubated at 37 C for 1 hour. Values are normalized to a control consisting of probe incubated in buffer only. Average of at least n=3 experimental replicates per bacterial strain shown. C) Assessment of 2’-FL-SH probe stability against human skin nucleases. Probe digestion assay with endogenous skin nucleases with S. pyogenes (HSC5) overnight culture supernatant as positive control. 1 ul of skin lysate (1/10 of reaction volume) or 1 ul of S. pyogenes culture supernatant was used per reaction. Reaction fluorescence values are normalized to that of a control consisting of probe incubated in reaction buffer only. Average of n=3 experimental replicates shown.
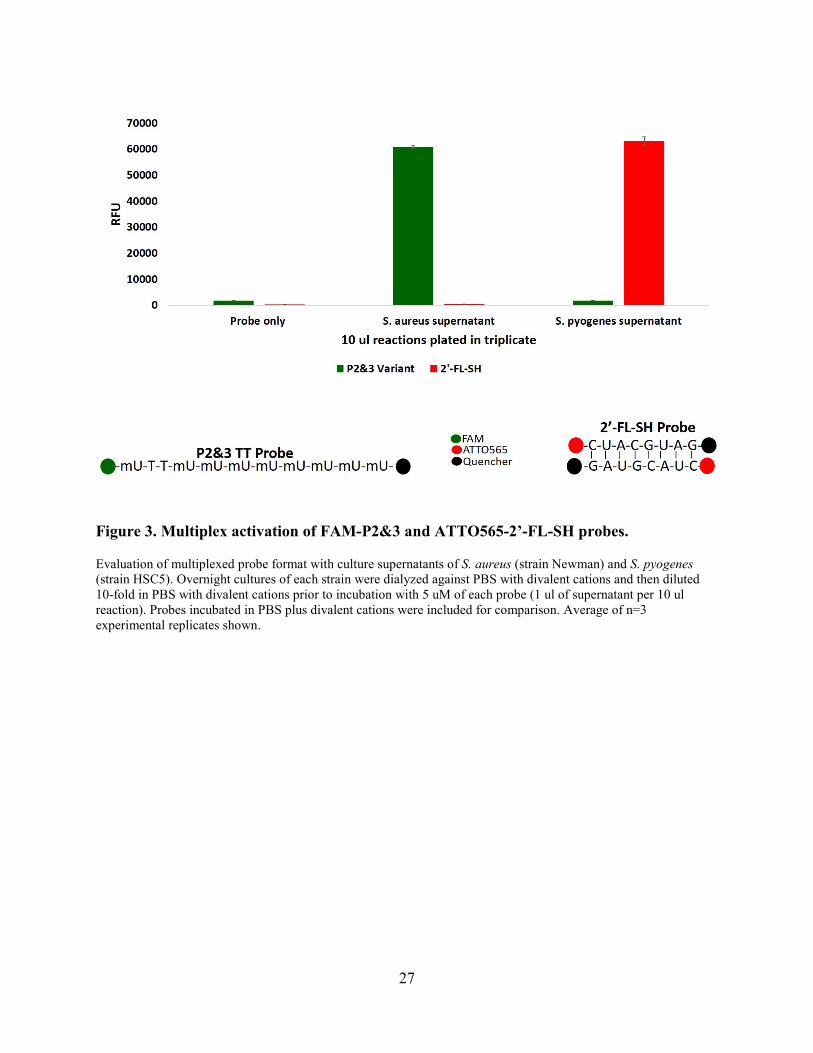
27
Figure 3. Multiplex activation of FAM-P2&3 and ATTO565-2’-FL-SH probes.
Evaluation of multiplexed probe format with culture supernatants of S. aureus (strain Newman) and S. pyogenes (strain HSC5). Overnight cultures of each strain were dialyzed against PBS with divalent cations and then diluted 10-fold in PBS with divalent cations prior to incubation with 5 uM of each probe (1 ul of supernatant per 10 ul reaction). Probes incubated in PBS plus divalent cations were included for comparison. Average of n=3 experimental replicates shown.
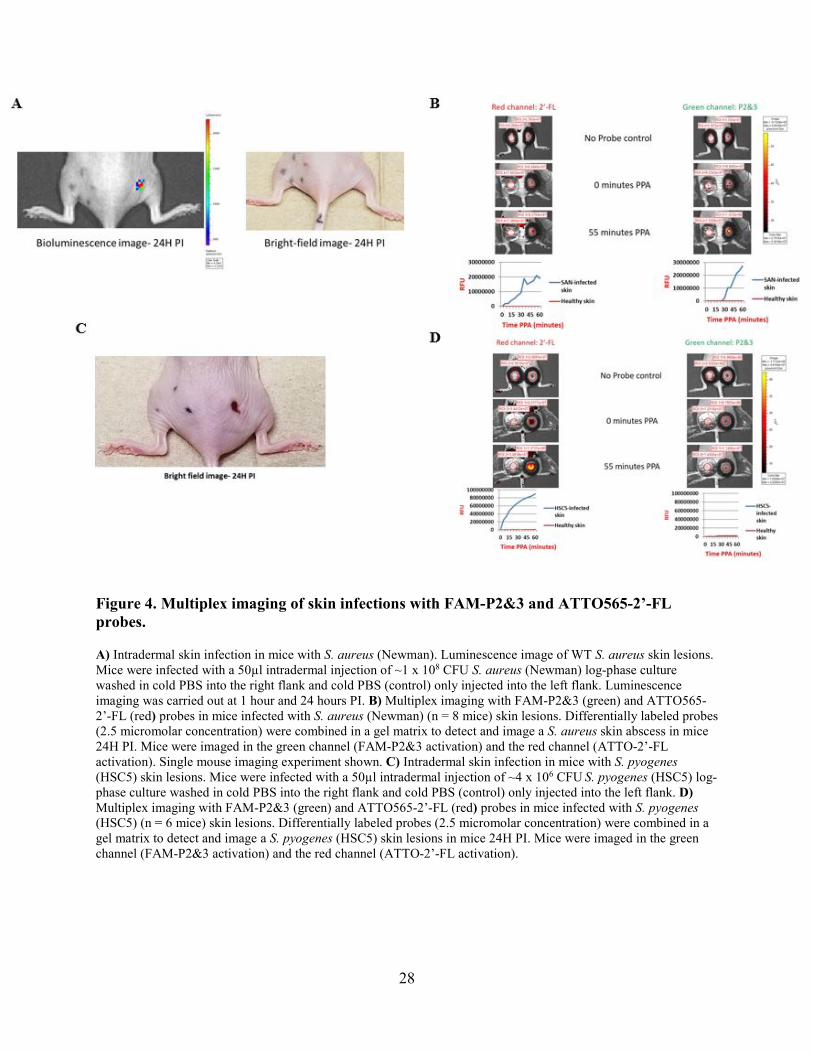
28
Figure 4. Multiplex imaging of skin infections with FAM-P2&3 and ATTO565-2’-FL probes.
A) Intradermal skin infection in mice with S. aureus (Newman). Luminescence image of WT S. aureus skin lesions. Mice were infected with a 50µl intradermal injection of ~1 x 108 CFU S. aureus (Newman) log-phase culture washed in cold PBS into the right flank and cold PBS (control) only injected into the left flank. Luminescence imaging was carried out at 1 hour and 24 hours PI. B) Multiplex imaging with FAM-P2&3 (green) and ATTO565-2’-FL (red) probes in mice infected with S. aureus (Newman) (n = 8 mice) skin lesions. Differentially labeled probes (2.5 micromolar concentration) were combined in a gel matrix to detect and image a S. aureus skin abscess in mice 24H PI. Mice were imaged in the green channel (FAM-P2&3 activation) and the red channel (ATTO-2’-FL activation). Single mouse imaging experiment shown. C) Intradermal skin infection in mice with S. pyogenes (HSC5) skin lesions. Mice were infected with a 50µl intradermal injection of ~4 x 106 CFU S. pyogenes (HSC5) log-phase culture washed in cold PBS into the right flank and cold PBS (control) only injected into the left flank. D) Multiplex imaging with FAM-P2&3 (green) and ATTO565-2’-FL (red) probes in mice infected with S. pyogenes (HSC5) (n = 6 mice) skin lesions. Differentially labeled probes (2.5 micromolar concentration) were combined in a gel matrix to detect and image a S. pyogenes (HSC5) skin lesions in mice 24H PI. Mice were imaged in the green channel (FAM-P2&3 activation) and the red channel (ATTO-2’-FL activation).
29
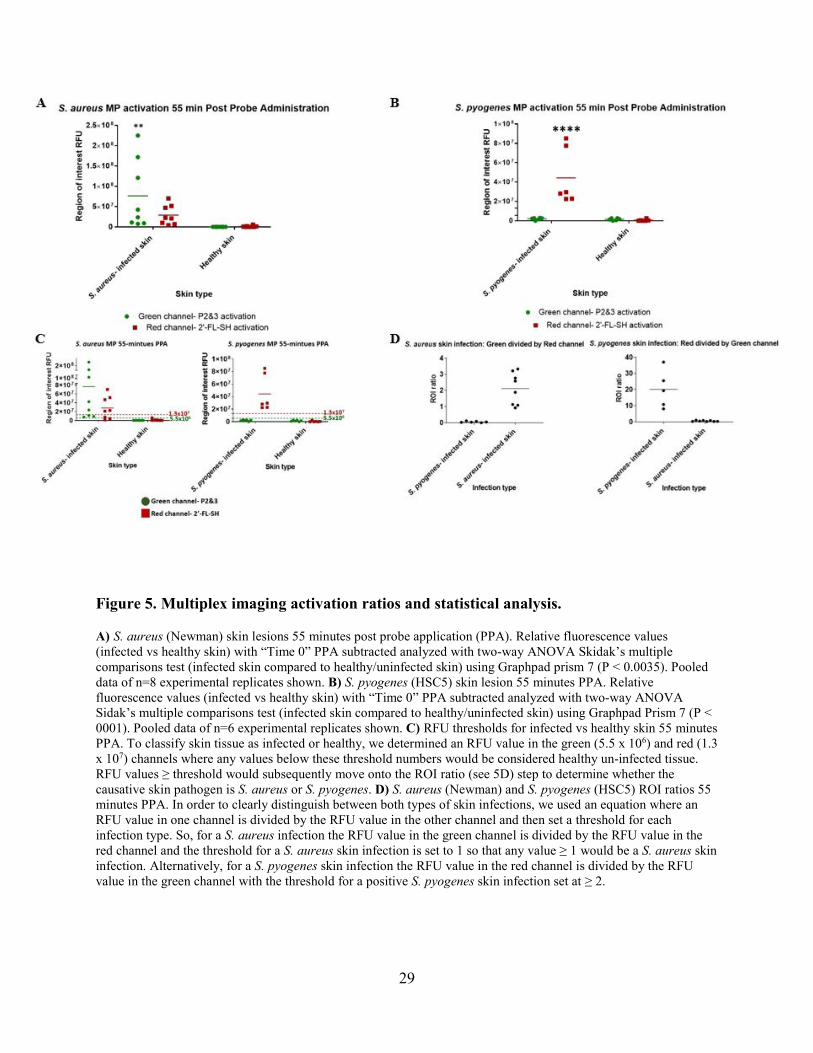
Figure 5. Multiplex imaging activation ratios and statistical analysis.
A) S. aureus (Newman) skin lesions 55 minutes post probe application (PPA). Relative fluorescence values (infected vs healthy skin) with “Time 0” PPA subtracted analyzed with two-way ANOVA Skidak’s multiple comparisons test (infected skin compared to healthy/uninfected skin) using Graphpad prism 7 (P < 0.0035). Pooled data of n=8 experimental replicates shown. B) S. pyogenes (HSC5) skin lesion 55 minutes PPA. Relative fluorescence values (infected vs healthy skin) with “Time 0” PPA subtracted analyzed with two-way ANOVA Sidak’s multiple comparisons test (infected skin compared to healthy/uninfected skin) using Graphpad Prism 7 (P < 0001). Pooled data of n=6 experimental replicates shown. C) RFU thresholds for infected vs healthy skin 55 minutes PPA. To classify skin tissue as infected or healthy, we determined an RFU value in the green (5.5 x 106) and red (1.3 x 107) channels where any values below these threshold numbers would be considered healthy un-infected tissue. RFU values ≥ threshold would subsequently move onto the ROI ratio (see 5D) step to determine whether the causative skin pathogen is S. aureus or S. pyogenes. D) S. aureus (Newman) and S. pyogenes (HSC5) ROI ratios 55 minutes PPA. In order to clearly distinguish between both types of skin infections, we used an equation where an RFU value in one channel is divided by the RFU value in the other channel and then set a threshold for each infection type. So, for a S. aureus infection the RFU value in the green channel is divided by the RFU value in the red channel and the threshold for a S. aureus skin infection is set to 1 so that any value ≥ 1 would be a S. aureus skin infection. Alternatively, for a S. pyogenes skin infection the RFU value in the red channel is divided by the RFU value in the green channel with the threshold for a positive S. pyogenes skin infection set at ≥ 2.
30
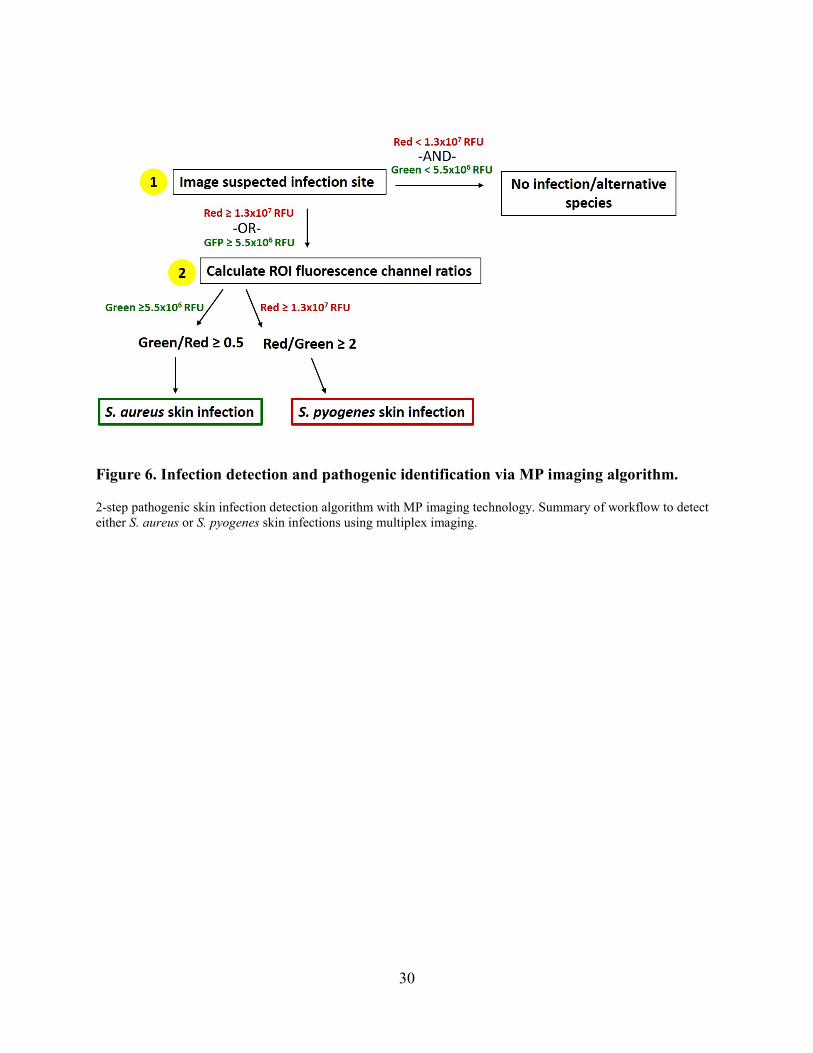
Figure 6. Infection detection and pathogenic identification via MP imaging algorithm.
2-step pathogenic skin infection detection algorithm with MP imaging technology. Summary of workflow to detect either S. aureus or S. pyogenes skin infections using multiplex imaging.
31
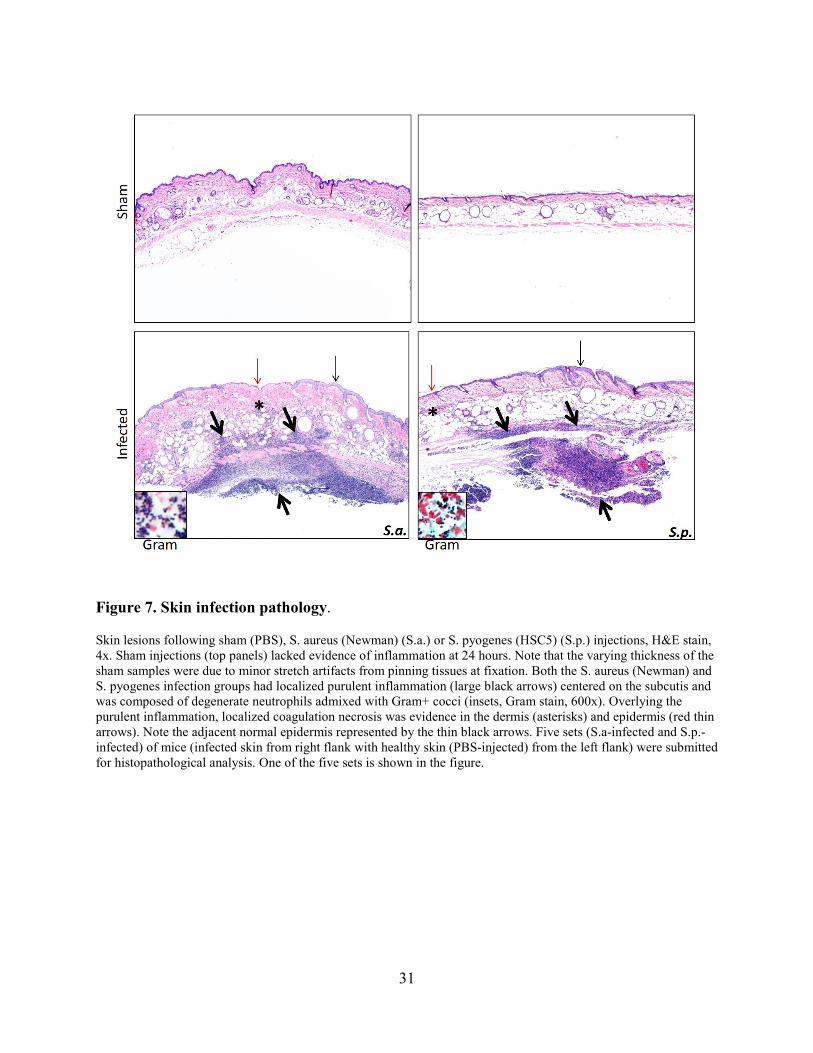
Figure 7. Skin infection pathology.
Skin lesions following sham (PBS), S. aureus (Newman) (S.a.) or S. pyogenes (HSC5) (S.p.) injections, H&E stain, 4x. Sham injections (top panels) lacked evidence of inflammation at 24 hours. Note that the varying thickness of the sham samples were due to minor stretch artifacts from pinning tissues at fixation. Both the S. aureus (Newman) and S. pyogenes infection groups had localized purulent inflammation (large black arrows) centered on the subcutis and was composed of degenerate neutrophils admixed with Gram+ cocci (insets, Gram stain, 600x). Overlying the purulent inflammation, localized coagulation necrosis was evidence in the dermis (asterisks) and epidermis (red thin arrows). Note the adjacent normal epidermis represented by the thin black arrows. Five sets (S.a-infected and S.p.-infected) of mice (infected skin from right flank with healthy skin (PBS-injected) from the left flank) were submitted for histopathological analysis. One of the five sets is shown in the figure.
32
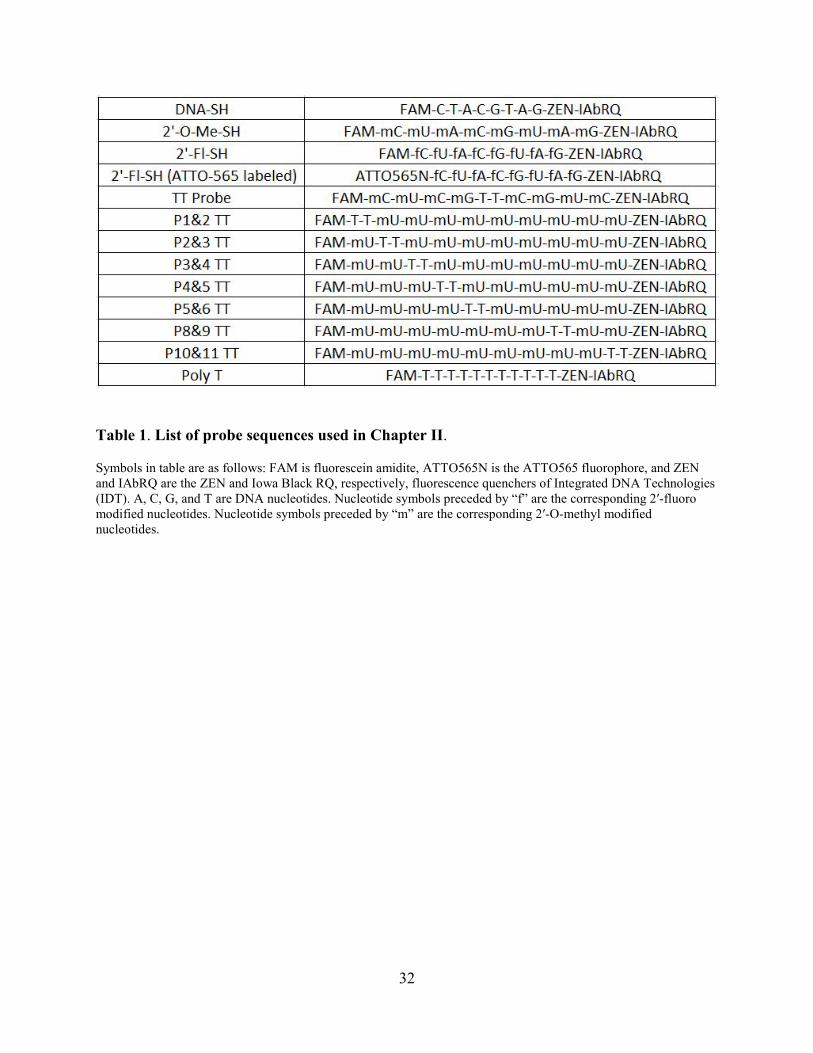
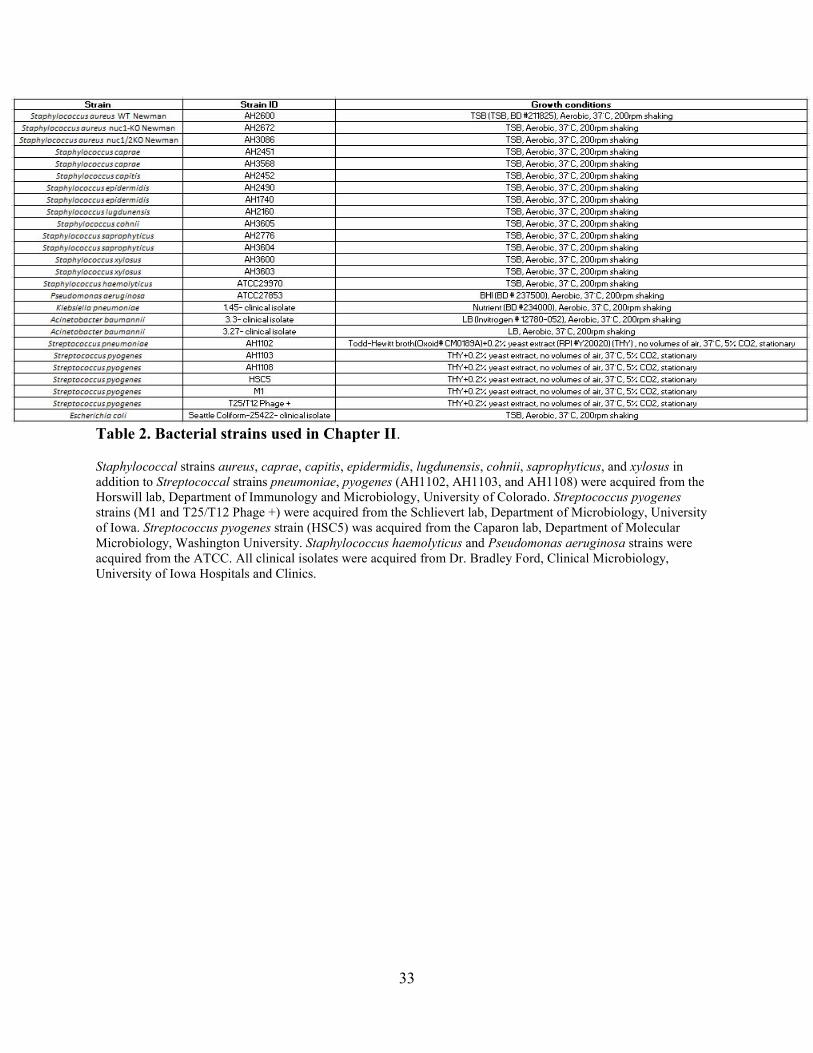
Table 1. List of probe sequences used in Chapter II.
Symbols in table are as follows: FAM is fluorescein amidite, ATTO565N is the ATTO565 fluorophore, and ZEN and IAbRQ are the ZEN and Iowa Black RQ, respectively, fluorescence quenchers of Integrated DNA Technologies (IDT). A, C, G, and T are DNA nucleotides. Nucleotide symbols preceded by “f” are the corresponding 2′-fluoro modified nucleotides. Nucleotide symbols preceded by “m” are the corresponding 2′-O-methyl modified nucleotides.
33
Table 2. Bacterial strains used in Chapter II.
Staphylococcal strains aureus, caprae, capitis, epidermidis, lugdunensis, cohnii, saprophyticus, and xylosus in addition to Streptococcal strains pneumoniae, pyogenes (AH1102, AH1103, and AH1108) were acquired from the Horswill lab, Department of Immunology and Microbiology, University of Colorado. Streptococcus pyogenes strains (M1 and T25/T12 Phage +) were acquired from the Schlievert lab, Department of Microbiology, University of Iowa. Streptococcus pyogenes strain (HSC5) was acquired from the Caparon lab, Department of Molecular Microbiology, Washington University. Staphylococcus haemolyticus and Pseudomonas aeruginosa strains were acquired from the ATCC. All clinical isolates were acquired from Dr. Bradley Ford, Clinical Microbiology, University of Iowa Hospitals and Clinics.
34
Supplementary Figure 1. Evaluation of multiplexed probe format with culture supernatants of S. aureus (strain Newman) and S. pyogenes HSC5, M1, T12, and AH1103 strains. Overnight cultures of each strain were dialyzed against PBS with divalent cations and then diluted 10-fold in PBS with divalent cations prior to incubation with 5 uM of each probe (1 ul of supernatant per 10 ul reaction). Probes incubated in PBS plus divalent cations were included for comparison. Average of n=3 experimental replicates shown.

35
CHAPTER III: NIR PROBE CHARACTERISTICS IN WHOLE BLOOD
Abstract
Near-infrared (NIR) fluorescent probes that promise to enable the accurate diagnosis of a
variety of diseases via non-invasive optical imaging are currently being developed. We
previously described a novel molecular optical imaging technology to noninvasively detect S.
aureus thigh infections in mice within 1 hour of IV probe administration. Despite the promising
data that we and others have generated with NIR fluorescent probes, the interactions of
fluorophores with whole blood have not been well characterized. We therefore sought to
compare the fluorescence levels of some commonly used unquenched NIR fluorophores (NIR
dye coupled to an oligonucleotide without quencher) in whole blood and blood components.
Here, we report a greater fluorescence intensity when NIR probes are diluted in whole blood
versus phosphate buffered saline (PBS). More modest differences in fluorescence intensity were
observed when the probes were diluted in human and mouse plasma or serum versus PBS. The
results from this study provided insights that were useful in efforts to optimize the second-
generation nuclease-activatable NIR probe used in Chapter IV.
Introduction
In recent years NIR fluorophores have gained considerable attention from researchers as
molecular fluorescence-imaging agents. This is due to the particular wavelengths, 650-900 nm, at
which these molecules are excited and emit fluorescence. Within this range of light, tissue
absorption is very low allowing deep penetration in addition to low light scattering. This range,
known as the “optical window,” enables deep tissue and whole animal imaging with a higher
signal-to-background ratio [101, 102]. Despite the promising results our group and others have

36
generated, the interaction of fluorophores in the NIR spectra with whole blood has not been well
characterized [31, 103]. In addition, NIR fluorophore specifications (i.e. extinction coefficient,
quantum yield) have only been assessed in non-biologically relevant solvents such as toluene,
MeOH, cyclohexane [104]. There are a limited number of NIR-imaging probes available for
clinical use [35, 105], therefore to help advance optical imaging with NIR labelled molecules, a
better understanding of how these dyes perform in vivo (e.g. blood) will be beneficial.
The degree of fluorescence from NIR fluorophores in blood is based, for the most part, on
the chemical composition of the fluorophore. For example, the Cy5.5 fluorophore has a
hydrophobic structure that gives it different fluorescence characteristics depending on the type of
solution in which it is being examined [106]. The objective of this study was to characterize a
panel of fluorophores that span the NIR spectra in blood samples and less viscous dilution
buffers. In addition to the Cy5.5 fluorophore, the NIR fluorophores used in this study were
Cy7.5, IRDye800 and DyLight800. These dyes possess excitations/emissions that cover the NIR
region. We hypothesized that the other NIR dyes would exhibit substantial fluorescence
differences in blood compared to solutions composed of fewer components. The data support our
hypothesis that NIR dyes display a general increased fluorescence characteristic when diluted in
whole blood compared to other biologically relevant solutions. The findings in this study provide
insight to molecular imaging probe optimization.
Materials and Methods
Unquenched oligonucleotide probe synthesis. All unquenched probes used in this study
were synthesized, purified (HPLC), and lyophilized by Integrated DNA Technologies® (IDT).
Lyophilized probes were reconstituted in TE buffer (Ambion™ Ref. AM9849 supplemented
with 10 mM TRIS-HCL pH 8.0 and 1 mM EDTA) for a 500 µM final frozen stock

37
concentration. 1 µl aliquots were then frozen at -80°C.
Note: The PBS used in all experiments for this study was Dulbecco’s phosphate-buffered saline
with (Ref. 14040-133) or without (Ref. 14190-144) physiological amounts of calcium and
magnesium from Invitrogen. For in vitro experiments, lithium-heparinized mouse blood, lithium-
heparinized plasma, serum and lithium-heparinized human whole blood were acquired from
Bioreclamation Inc.
In vitro assessment of oligonucleotide probes with microplate reader assays.
Fluorescence plate reader assays were carried out as previously described [31] with a Synergy™
Mx Microplate reader or a Li-Cor® NIR imaging machine. Probe dilution assays were carried
out in 50 µl reactions. Probes (500 pmol/µl) were reconstituted and diluted in PBS with divalent
cations, lithium-heparinized mouse blood, lithium- heparinized plasma, serum or lithium-
heparinized human whole blood. 4 µl of the specified probe was combined with 46 µl (final
probe concentration of 0.334 µM) of the specified buffer (i.e. PBS or blood) taken in triplicate
and incubated in separate 1.5 ml SealRite® Natural Microcentrifuge Tubes (Lot No. E163484O-
1739) at 37°C for 1 hour. Following the incubation period, solutions from each tube were
transferred to a Falcon® 96-Well clear bottom tissue culture treated plate (Ref. 353219) and read
on the Odyssey Classic, Odyssey Clx (Cy5.5: 700 nm channel IRDye800: 800 nm channel) or
the Synergy Mx (Cy5.5: 650 excitation 698 emission 160 Gain IRDye800: 750 excitation 700
emission 180 Gain). Data was then analyzed with Microsoft Excel.
Statistical analysis. RFU averages and standard deviation of triplicate plating conditions
(based on the experiment- see figure legends) were calculated using Microsoft Excel. Standard
deviation for all experiments was also done using Microsoft Excel, due to the age and technical

38
issues of the software in the three microplate readers it was not shown in all figures. Note:
Statistical significance was not calculated for any of the experiments in this chapter.
Results
We initially included two commonly-used NIR fluorophores in unquenched (always
“on”) probes as positive controls for in vitro MN digestion assays. All digestion reactions took
place in phosphate buffered saline (PBS) with divalent cations and mouse (and human) whole
blood. The initial observation of altered fluorescence came in the positive control group (Fig. 8).
Unquenched Cy5.5 and IRDye800 probes revealed higher RFU values when diluted in whole
blood than when diluted in PBS. To isolate the part of blood in which the NIR-probe interaction
was occurring, we next looked at Cy5.5 unquenched probe fluorescence diluted in two fractions
of whole blood: plasma and red blood cells (crude extractions) (Fig. 9A). The results revealed
only modest differences compared to whole blood. We then examined the unquenched Cy5.5 and
IRDye800 probes diluted in plasma and serum (purchased commercially) compared to PBS and
whole blood (Fig. 9B). We followed this by a closer look at unquenched Cy5.5 probe dilution in
blood fractions read in two NIR microplate reader instruments due to the fact that the Licor
instrument had greater sensitivity to NIR probe fluorescence than the SynergyMx instrument
(Fig. 9C). These results also displayed modest differences between RFUs in blood fractions
compared to whole blood. Because hemoglobin is an abundant protein in blood [107, 108], we
examined unquenched Cy5.5 IRDye800 probes at serially diluted concentrations of purified
hemoglobin to determine if this interaction was the cause of the increased fluorescence in blood
(Fig. 9D). However, only modest contributions to enhanced fluorescence were observed when
NIR probes were diluted in purified hemoglobin.

39
An emission (Fig. 10A) and excitation (Fig. 10B) spectral scan on unquenched Cy5.5
probe diluted in PBS, purified hemoglobin, and whole blood was then carried out to identify any
light spectrum changes when the probe was diluted in these buffers. Normalized data revealed
that no spectral changes occurred when the unquenched probe was diluted in the buffers tested.
Data from this experiment illustrated the fact that changes when unquenched NIR probes are
diluted in various buffers only occur in the relative fluorescence values and not spectrum of light.
Standard curves were subsequently carried out with unquenched Cy5.5 and IRDye800 probes to
discern whether probe concentration was proportional to the relative fluorescence intensity (Fig
11A). We then compared instrument sensitivity to unquenched probes using the SynergyMx and
Odyssey clx microplate readers (Fig. 11B). The data from this experiment revealed that NIR
probe concentration is correlated with fluorescence intensity.
In addition to enhanced fluorescence intensity in whole blood observation, the
unquenched NIR probes displayed much lower relative fluorescence intensities when diluted in
PBS or other hydrophilic buffers. A plausible explanation for this would be self-quenching of the
hydrophobic NIR probes in a hydrophilic environment. To this end we carried out 2-fold
standard curve dilutions in PBS and PBS + 1% sodium dodecyl sulfate (SDS) of unquenched
Cy5.5 probe with and without heat (Fig. 12A). Results from this experiment did not appear to
support the self-quenching hypothesis due to minimal difference in fluorescence values between
PBS and PBS + 1% SDS groups. We then used ethanol and methanol as dilution buffers (Fig.
12B) and observed modest differences when compared with fluorescence values in PBS. Lastly,
we examined the relative fluorescence values of unquenched NIR probes in PBS and whole
blood as a function of time over the course of 30 minutes (Fig. 12C). The relative fluorescence
value of the unquenched Cy5.5 probe diluted in blood increased to saturation levels within 10

40
minutes. In contrast, the probe diluted in PBS with divalent cations had a modest reduction in
relative fluorescence value over the course of 30 minutes. These results may suggest the
possibility that NIR probes display enhanced fluorescence in blood due to interactions with
certain blood components whereas they have a reduction in fluorescence value in more
hydrophilic dilution buffers (i.e. PBS) because of aggregating and ultimately self-quenching.
Conclusion
In conclusion, the we show that NIR fluorophores display enhanced fluorescence
characteristics when diluted in whole blood. In addition, we have provided evidence that NIR
fluorophores display diminished fluorescence characteristics when diluted in hydrophilic buffers,
most likely the result of self-quenching. The knowledge gained from this study contributed to the
development of a second-generation NIR probe used to image catheter biofilm infections in
mice. Multidisciplinary teams that can develop and assess NIR fluorophores are best suited to
maximize their potential in the context of biomedical imaging.
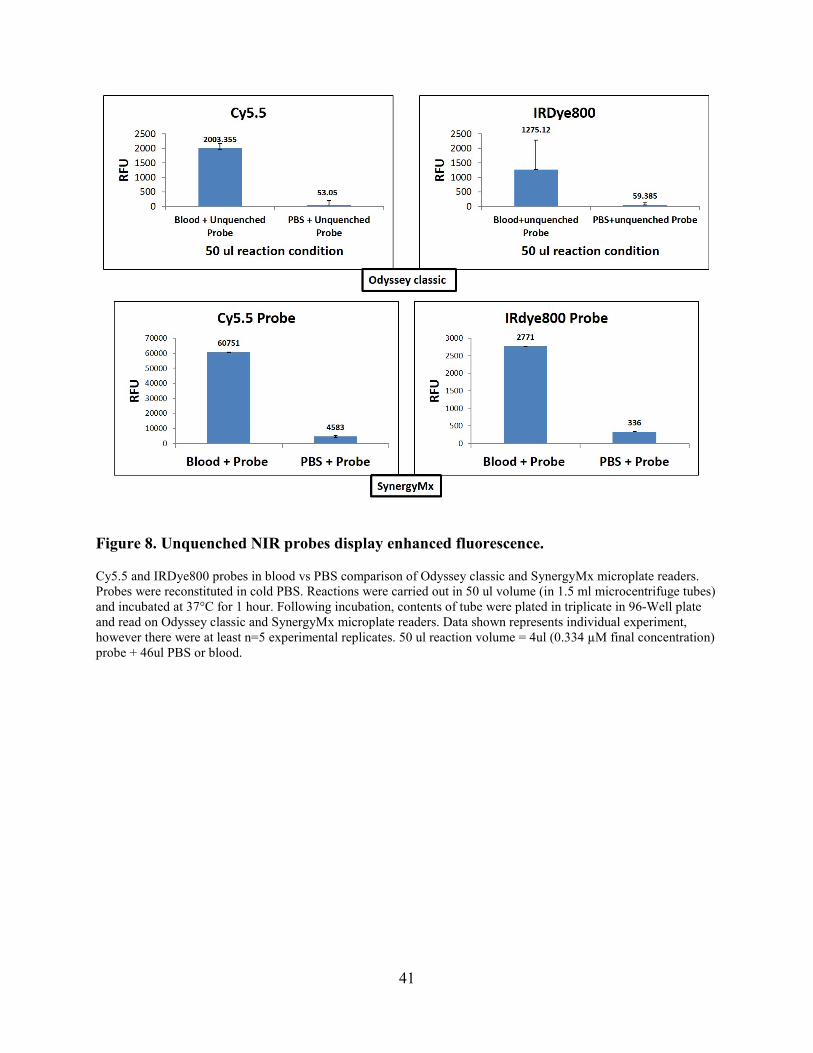
41
Figure 8. Unquenched NIR probes display enhanced fluorescence.
Cy5.5 and IRDye800 probes in blood vs PBS comparison of Odyssey classic and SynergyMx microplate readers. Probes were reconstituted in cold PBS. Reactions were carried out in 50 ul volume (in 1.5 ml microcentrifuge tubes) and incubated at 37°C for 1 hour. Following incubation, contents of tube were plated in triplicate in 96-Well plate and read on Odyssey classic and SynergyMx microplate readers. Data shown represents individual experiment, however there were at least n=5 experimental replicates. 50 ul reaction volume = 4ul (0.334 µM final concentration) probe + 46ul PBS or blood.
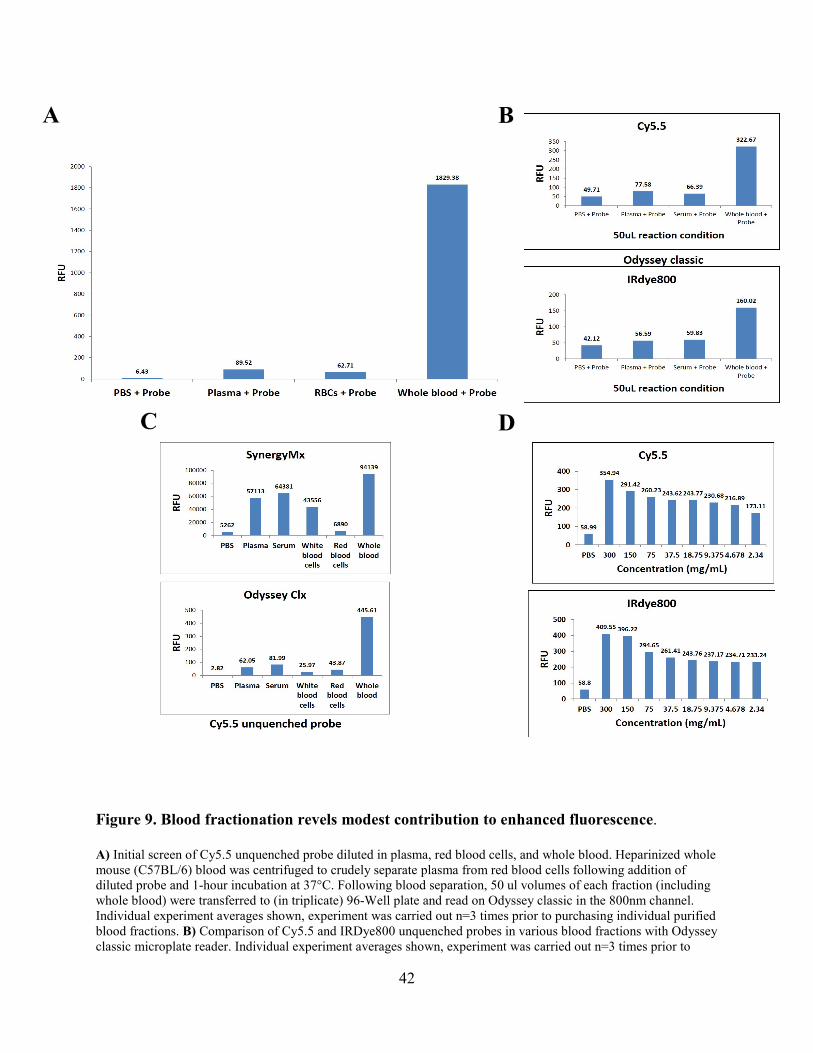
42
Figure 9. Blood fractionation revels modest contribution to enhanced fluorescence.
A) Initial screen of Cy5.5 unquenched probe diluted in plasma, red blood cells, and whole blood. Heparinized whole mouse (C57BL/6) blood was centrifuged to crudely separate plasma from red blood cells following addition of diluted probe and 1-hour incubation at 37°C. Following blood separation, 50 ul volumes of each fraction (including whole blood) were transferred to (in triplicate) 96-Well plate and read on Odyssey classic in the 800nm channel. Individual experiment averages shown, experiment was carried out n=3 times prior to purchasing individual purified blood fractions. B) Comparison of Cy5.5 and IRDye800 unquenched probes in various blood fractions with Odyssey classic microplate reader. Individual experiment averages shown, experiment was carried out n=3 times prior to
A
B
C
D

43
purchasing individual purified blood fractions. C) Comparison of Cy5.5 unquenched probe diluted in pre-separated blood fractions between SynergyMx and Odyssey Clx microplate readers. Individual experiment averages shown, experiment was carried out at least n=5 times. D) Hemoglobin screen with Cy5.5 and IRDye unquenched probes show that red blood cells along are not the cause of enhanced fluorescence. Hemoglobin (reconstituted and diluted in PBS) serially diluted with seven 2-fold dilutions starting at 300 mg/ml concentration. Same assay protocols as 9A/B. Individual experiment averages shown, experiment was carried out n=3 times. 50 ul reaction volume = 4ul (0.334 µM final concentration) probe + 46ul buffer (see figure) for all experiments performed in this figure.
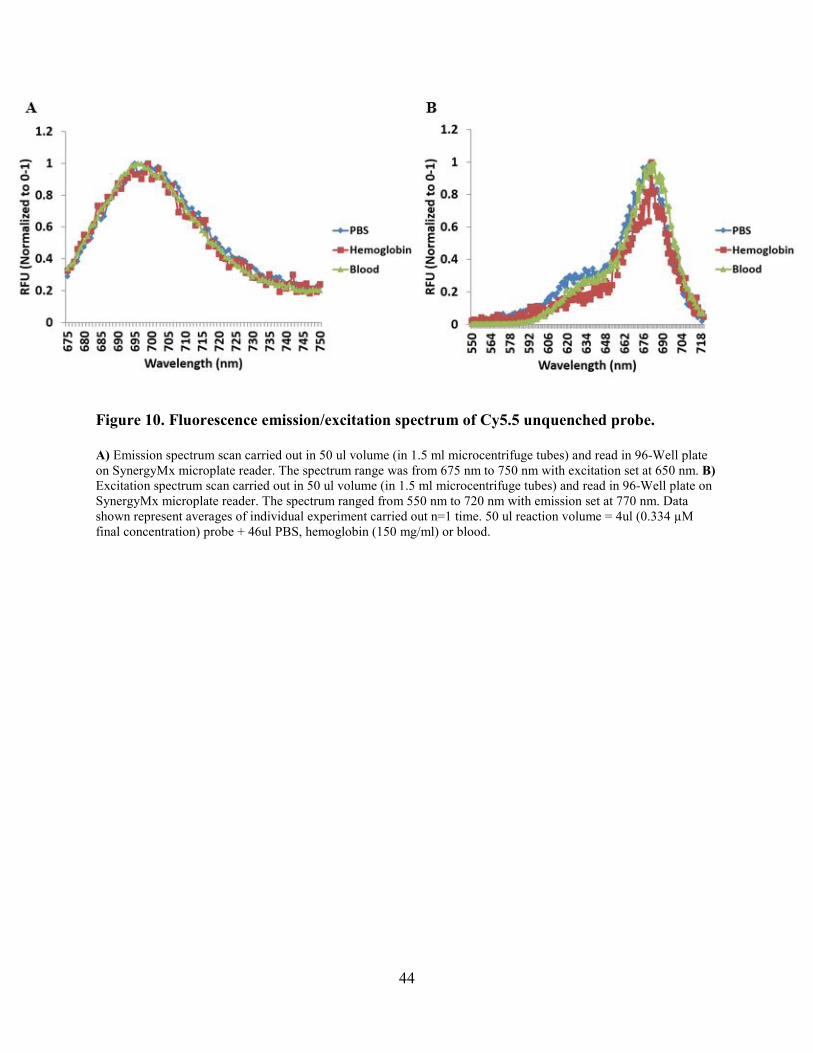
44
Figure 10. Fluorescence emission/excitation spectrum of Cy5.5 unquenched probe.
A) Emission spectrum scan carried out in 50 ul volume (in 1.5 ml microcentrifuge tubes) and read in 96-Well plate on SynergyMx microplate reader. The spectrum range was from 675 nm to 750 nm with excitation set at 650 nm. B) Excitation spectrum scan carried out in 50 ul volume (in 1.5 ml microcentrifuge tubes) and read in 96-Well plate on SynergyMx microplate reader. The spectrum ranged from 550 nm to 720 nm with emission set at 770 nm. Data shown represent averages of individual experiment carried out n=1 time. 50 ul reaction volume = 4ul (0.334 µM final concentration) probe + 46ul PBS, hemoglobin (150 mg/ml) or blood.
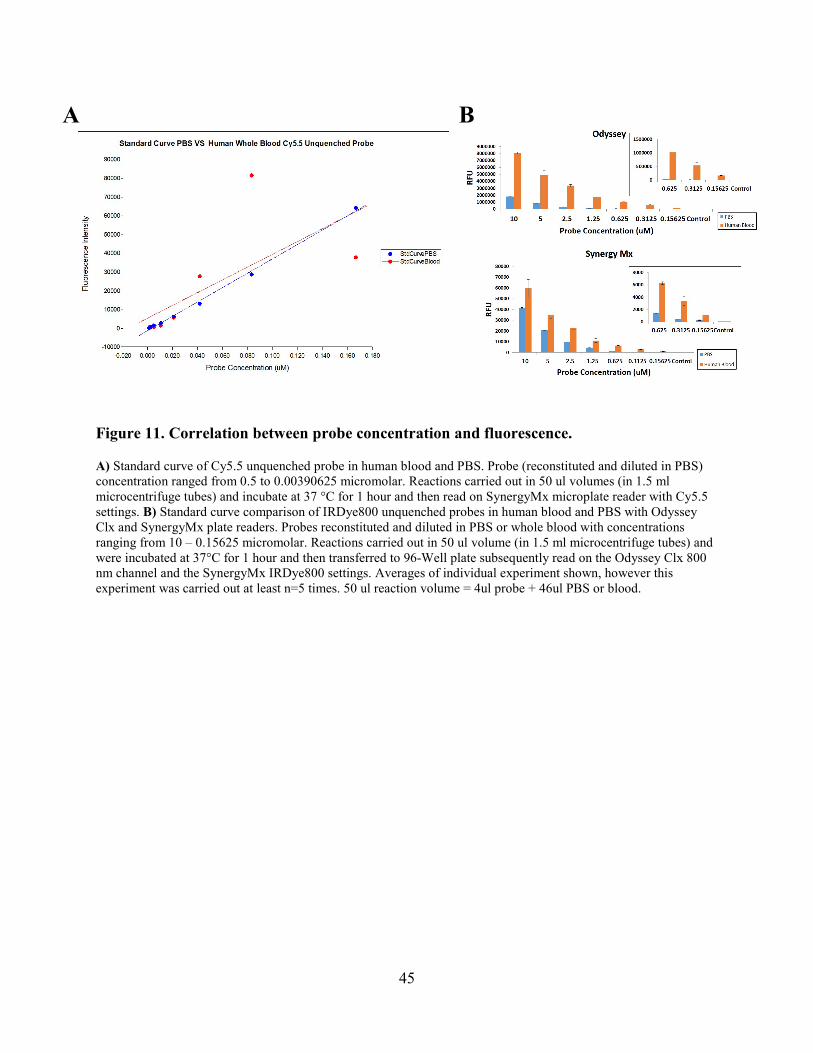
45
Figure 11. Correlation between probe concentration and fluorescence.
A) Standard curve of Cy5.5 unquenched probe in human blood and PBS. Probe (reconstituted and diluted in PBS) concentration ranged from 0.5 to 0.00390625 micromolar. Reactions carried out in 50 ul volumes (in 1.5 ml microcentrifuge tubes) and incubate at 37 °C for 1 hour and then read on SynergyMx microplate reader with Cy5.5 settings. B) Standard curve comparison of IRDye800 unquenched probes in human blood and PBS with Odyssey Clx and SynergyMx plate readers. Probes reconstituted and diluted in PBS or whole blood with concentrations ranging from 10 – 0.15625 micromolar. Reactions carried out in 50 ul volume (in 1.5 ml microcentrifuge tubes) and were incubated at 37°C for 1 hour and then transferred to 96-Well plate subsequently read on the Odyssey Clx 800 nm channel and the SynergyMx IRDye800 settings. Averages of individual experiment shown, however this experiment was carried out at least n=5 times. 50 ul reaction volume = 4ul probe + 46ul PBS or blood.
B
A
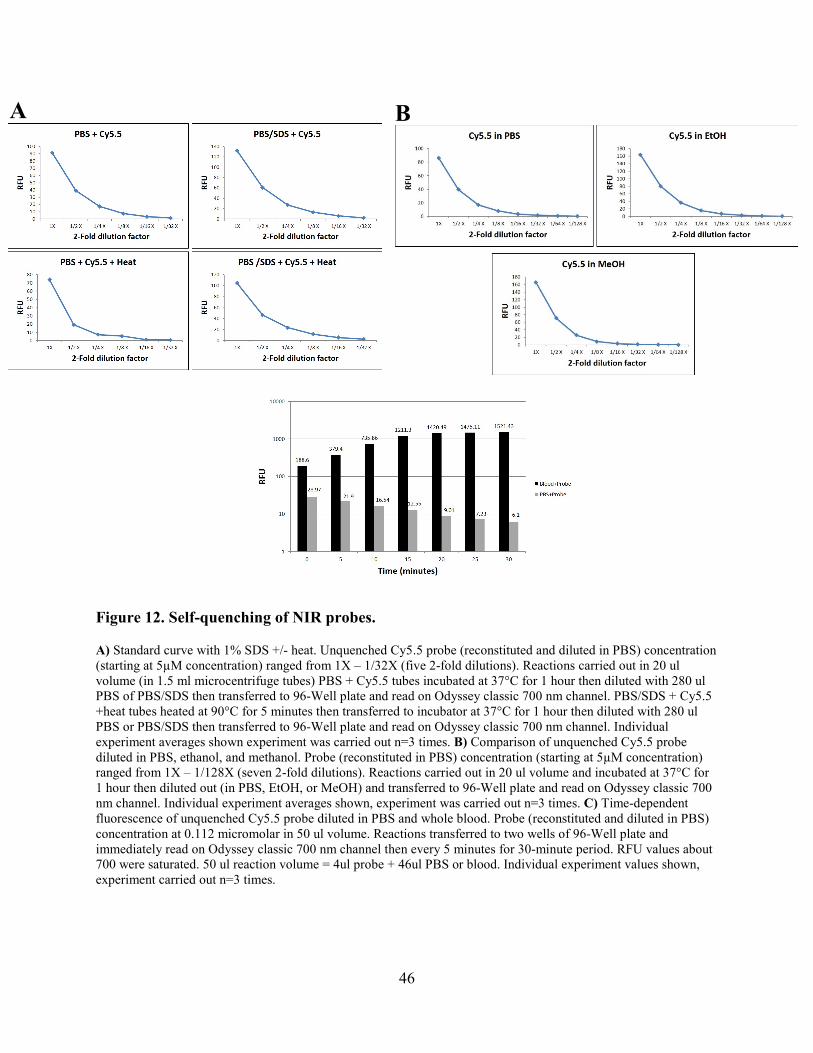
46
Figure 12. Self-quenching of NIR probes.
A) Standard curve with 1% SDS +/- heat. Unquenched Cy5.5 probe (reconstituted and diluted in PBS) concentration (starting at 5µM concentration) ranged from 1X – 1/32X (five 2-fold dilutions). Reactions carried out in 20 ul volume (in 1.5 ml microcentrifuge tubes) PBS + Cy5.5 tubes incubated at 37°C for 1 hour then diluted with 280 ul PBS of PBS/SDS then transferred to 96-Well plate and read on Odyssey classic 700 nm channel. PBS/SDS + Cy5.5 +heat tubes heated at 90°C for 5 minutes then transferred to incubator at 37°C for 1 hour then diluted with 280 ul PBS or PBS/SDS then transferred to 96-Well plate and read on Odyssey classic 700 nm channel. Individual experiment averages shown experiment was carried out n=3 times. B) Comparison of unquenched Cy5.5 probe diluted in PBS, ethanol, and methanol. Probe (reconstituted in PBS) concentration (starting at 5µM concentration) ranged from 1X – 1/128X (seven 2-fold dilutions). Reactions carried out in 20 ul volume and incubated at 37°C for 1 hour then diluted out (in PBS, EtOH, or MeOH) and transferred to 96-Well plate and read on Odyssey classic 700 nm channel. Individual experiment averages shown, experiment was carried out n=3 times. C) Time-dependent fluorescence of unquenched Cy5.5 probe diluted in PBS and whole blood. Probe (reconstituted and diluted in PBS) concentration at 0.112 micromolar in 50 ul volume. Reactions transferred to two wells of 96-Well plate and immediately read on Odyssey classic 700 nm channel then every 5 minutes for 30-minute period. RFU values about 700 were saturated. 50 ul reaction volume = 4ul probe + 46ul PBS or blood. Individual experiment values shown, experiment carried out n=3 times.
A
A
B
B

47
Table 3. NIR probe sequences used in Chapter III.
Symbols in table are as follows: Cy5.5 is the cyanine5.5 fluorophore, Cy7.5 is the cyanine7.5 fluorophore, IRdye800 is the IRDye®800 CW fluorophore, and the DyLight800 is the DyLight800 maleimide fluorophore. C and T are DNA nucleotides. Nucleotide symbols preceded by “m” are the corresponding 2′-O-methyl modified nucleotides.

48
CHAPTER IV: NONINVASIVE IMAGING OF BIOFILM INFECTIONS BY S. AUREUS WITH SECOND-GENERATION NIR PROBE
Abstract
We previously described a novel molecular imaging technology capable of rapidly
detecting focal S. aureus infections in mice. This entails intravenous administration of an
activatable near-infrared (NIR) fluorescent probe in which a fluorophore and a quencher are
attached to opposite ends of a synthetic oligonucleotide sequence. Upon encountering an S. aureus
infection, the probe is selectively cleaved by a secreted nuclease of S. aureus, yielding probe
activation (i.e., higher fluorescence). This approach enabled us to rapidly image S. aureus muscle
infections in mice. In the study described here, we sought to improve 1) the spectral properties of
the first generation NIR probe to enable deeper tissue imaging and 2) the efficiency of probe
activation to produce more rapid imaging responses. In addition, we sought to determine the
efficacy of this imaging technology with biofilm infections which present very different
microenvironments from muscle infections. We report a sequence variant that is more efficiently
digested by micrococcal nuclease (the secreted nuclease of S. aureus) and an 800-nanometer
fluorescence emitting fluorophore that exhibits robust performance in whole blood. When this
second-generation probe was tested in a mouse S. aureus catheter biofilm infection model, it
yielded elevated fluorescence at infection sites as early as 5 minutes following probe
administration. This probe also displays the ability to distinguish between S. aureus and
alternative bacterial species as well as sterile inflammation.
Introduction
Staphylococcus aureus is the leading cause of bacteremia and infective endocarditis
through intravenous catheter biofilm infections [109, 110]. There is an urgent need to detect and
diagnose infections of this type at an early stage so targeted therapy can be administered to ensure

49
complication-free survival. However currently, diagnosis often begins with a physician observing
the patient’s symptoms including swelling, pain, redness, and fever. The catheter is then removed
and cultures are grown from the catheter itself or the implant site to get a definitive diagnosis
[111]. These steps involve invasive procedures and take days or even weeks to correctly identify
the invading pathogen by which time it is often too late as the infection has spread to other areas
of the patient’s body. Therefore, a novel diagnostic technology that can definitively detect an S.
aureus catheter biofilm infection in a rapid and noninvasive manner could be extremely beneficial.
Interest in activatable molecular imaging probes has grown considerably in recent years
because they enable physicians to observe molecular processes that lead to disease in a
noninvasive manner. Moreover, these probes allow the detection of a target (or process) with high
target-to-background ratio within living systems. Due to these characteristics, imaging probes
have become highly desired in many different research areas including cancer and pharmacology
[112, 113].
Our previous work involved use of a first-generation nuclease activatable probe to rapidly
detect and image a thigh muscle infection (pyomyositis) by S. aureus in a mouse model [31]. This
was accomplished by exploiting the enzymatic activity of a prevalent nuclease secreted by S.
aureus, micrococcal nuclease (MN). The probe used for this study contained a single stranded
modified (O-methylated) RNA sequence that had a Cy5.5 near infrared (NIR) fluorophore and a
quencher on opposite ends (figure 1: activation cartoon). In the presence of S. aureus, this probe
yielded a fluorescence activation signal within 30 minutes of intravenous administration.
The findings from our previous study led us to hypothesize that this novel technology
could be applied to S. aureus catheter biofilm infections with a second-generation nuclease
activatable probe. The second-generation probe consists of an alternate single-stranded RNA

50
sequence (also O-methylated) with a DyLight800 NIR fluorophore and a quencher on opposite
ends. The DyLight800 labeling of the RNA sequence enables deeper tissue imaging than the
previous probe due to longer (~800 nanometers) wavelength emission from the fluorophore [32,
101, 104].
Here we report the rapid detection of a S. aureus catheter biofilm with the second-
generation nuclease activatable probe. The modified RNA sequence of this probe enables it to be
degraded by the MN enzyme more efficiently than the first-generation Cy5.5 probe. This allows
specific detection and imaging of wild-type S. aureus catheter biofilm infections within 30
minutes of I.V. administration at relatively low doses. The rapid and noninvasive manner in which
this probe is used allows easy translatability for infection imaging in clinical settings.
Materials and Methods
Quenched and unquenched oligonucleotide probe synthesis. All probes used in this
study were synthesized, purified (HPLC), and lyophilized by Integrated DNA Technologies®
(IDT). Lyophilized probes were reconstituted in TE buffer (Ambion™ Ref. AM9849
supplemented with 10 mM TRIS-HCL pH 8.0 and 1 mM EDTA) for a 500 µM final frozen stock
concentration. 1 µl aliquots were then frozen at -80°C.
Note: PBS used in all experiments for this study was Dulbecco’s phosphate-buffered saline
(PBS) with (Ref. 14040-133) or without (Ref. 14190-144) physiological amounts of calcium and
magnesium from Invitrogen. For in vitro experiments, lithium-heparinized mouse blood, lithium-
heparinized plasma, serum and lithium-heparinized human whole blood were acquired from
BioreclamationIVT.

51
In vitro microplate reader assays. Fluorescence microplate reader assays were carried
out as previously described [31] with a Synergy™ Mx Microplate reader or a Li-Cor® NIR
imaging machine. Nuclease digestion assays were carried out in 10 µl reactions with 1 µl of a
frozen stock of each probe (50 µM) combined with 1 µl of specified concentration of purified
micrococcal nuclease (Worthington Biochemical Corporation) or dialyzed (in PBS with divalent
cations) overnight bacterial culture supernatant into 8 µl of PBS (buffer). All 10 µl reactions
were then incubated in 0.5 ml Eppendorf® LoBind Tubes (Ref. 0030108.094) at 37°C for 1
hour. Following the incubation period, 290 µl PBS without divalent cations supplemented with
10 mM EDTA 10 mM EGTA was added to each reaction tube. 95 µl aliquots were then
transferred in triplicate to 3 wells of a Falcon® 96 Well clear bottom tissue culture treated plate
(Ref. 353219). Fluorescence levels were measured and then analyzed using Excel. Probe dilution
assays were carried out in 50 µl reactions. The probe (500 pmol/µl) was reconstituted and diluted
in PBS with divalent cations. 4 or 5 µl of the specified probe was combined with 45 or 46 µl
(final probe concentration of 0.334 µM) of the specified buffer (i.e. PBS or blood) taken in
triplicate and incubated in separate 1.5 ml SealRite® Natural Microcentrifuge Tubes (Lot No.
E163484O-1739) at 37°C for 1 hour. Following the incubation, the 50 µl reactions were
transferred to wells of a Falcon® 96-Well clear bottom tissue culture treated plate (Ref. 353219)
and fluorescence levels were measured and then analyzed using Excel.
Bacterial culture conditions. All bacteria were cultured from a frozen stock in Bacto™
Tryptic Soy Broth (TSB) (Ref. 211825) or specified media and culture conditions (Table 2). To
prepare cultures for activation assays, overnight cultures were grown at 37°C with shaking at 200
r.p.m. To prepare consumed media, 10 ml of culture was centrifuged at 17,000 G for 12 minutes.

52
The supernatant was then collected. 2-3 ml of supernatant was then dialyzed (Pierce® 96-well
Microdialysis Plate (Ref. 88262)) in PBS with divalent cations at 4°C.
Genetic manipulation of S. aureus. The strains of S. aureus used in this study were
engineered to be luminescent by way of stable insertion of the lux operon into the chromosome
(Caliper Life Sciences, PerkinElmer Corporation). The Xen29 double nuclease mutant S. aureus
strain was created for use in this study. Both procedures were performed by the Horswill
laboratory (University of Colorado, Department of Immunology and Microbiology).
Catheter-associated biofilm infection model. Cultures were prepared for catheter
inoculum in the following fashion. First, overnight cultures of WT/mutant S. aureus (Xen29) or
S. epidermidis (1457) strain (Horswill lab, University of Colorado) were grown in TSB at 37°C
with shaking at 200 r.p.m. A 1:1000 diluted subculture was then grown for another 4 hours.
Following 4-hour growth, the culture was washed twice in cold PBS with divalent cations and
then re-suspended in cold PBS to an OD600 of 1.0. BD Insyte Autoguard ™ Shielded I.V. (REF.
381444) catheters from the UIHC outgoing patient supply store were cut into 1cm segments and
first incubated in female SKH-1(Charles River Laboratories) mouse serum for 1 hour at 37°C.
Catheter segments were then transferred to 1ml of washed WT/mutant S. aureus (Xen29), S.
epidermidis (1457), or PBS with divalent cations (sham control) culture (OD600=1.0) for 1 hour
at 37°C which gave the catheters a starting inoculum ~5.0 X 106 or 0 (control) CFU.
The animals were infected by re-sheathing the catheter segment incubated in the culture
and inserting it into the alcohol-swabbed skin of the left flank (vertically) of 6-8 week old SKH-1
female mice under isoflurane anesthesia. The needle lesion from the catheter applicator was then
sealed with LiquiBand®. Catheter sites were assessed with bioluminescent imaging on the day of
and 6 days following the application. Mice were imaged 6 days post-infection. Imaging and the

53
processing of the lesions (CFU, histology, and pathology) were carried out 6-7 days post-
infection. All animal experiments were approved by the University of Iowa Institutional Animal
Care and Use Committee (IACUC).
In vivo assessment of nuclease-activated probes. A Xenogen IVIS 200 imaging system
(3-minute exposure time) was used for all luminescent imaging carried out (prior to and/or
following fluorescence imaging). For catheter-associated biofilm infections, fluorescence
imaging was carried out with a Pearl® Trilogy Small Animal Imaging System. Mice were first
anesthetized with 2 % isoflurane gas and then placed on the imaging bed. Fluorescence images
were acquired in the 800 nm channel prior to and following I.V. probe administration with
specific time points denoted in the figures in the results section. Image Studio™ 5.2 software
was used for image acquisitions, processing, and analysis.
Target-to-background measurements. Region of interest (ROI) analysis was carried
out using the Image Studio™ 5.2 software. Fluorescence intensity was measured within the ROI
for S. aureus WT, S. epidermidis, S. aureus nuclease mutant, sham–infected tissue. Fluorescence
intensity was also measured within the ROI on the contralateral side for un-infected tissue for
each time point recorded as denoted in the figures in the results section. Raw data was analyzed
with a standard concentrated ROI. Each image in the time series was zoomed to the max in the
image studio software and the standard ROI was placed over the region of the catheter (and
placed over the contralateral control site) at the site of the most concentrated and highest pixel
intensity.
Infection CFU counts. Following fluorescence imaging of catheter biofilm infections,
the mice were sacrificed and the implanted catheters were carefully dissected out of mouse
tissue. Each catheter was placed in 1 ml PBS with divalent cations in a 1.5 ml microcentrifuge

54
tube. The tube was then sonicated for 10-15 minutes with a Branson 1510 ultrasonic cleaner.
Next, five 10-fold dilutions were carried out in PBS with divalent cations in 1.5 ml
microcentrifuge tubes. The initial tube (1 ml PBS + catheter) was marked as 10-1 dilution and the
subsequent diluted tubes were marked accordingly up to 10-6. 50 ul from each dilution tube was
then plated on TSA plates and spread with a sterile inoculation loop. Plates were then incubated
at 37°C overnight and CFU counts were made on the following day.
Statistical analysis. RFU values (n=6 Xen29 WT, n=10 Xen29 mut, n=4 S. epidermidis,
n=3 sham mice) from the 90-minute timepoint post probe administration (background
subtracted) were analyzed with a student’s unpaired nonparametric T test (catheter-infected site
compared to contralateral control site) using Graphpad Prism 7. Xen29 WT-infected mice
(P=0.0005), S. epdermidis-infected mice (P= 0.6098), sham-infected mice (P=0.8067), Xen29
nuc mutant-infected mice (P=0.1357).
Histology and pathology of biofilm infections. Mice were sacrificed 6 days PI via
carbon dioxide intoxication. The infected-catheter with attached tissue margins in the left flank
was carefully dissected along with contralateral control healthy tissue site in the right flank for
comparison and placed in 10% neutral buffered formalin at room temperature for 3-5 days. Next,
the fixed catheter was gross sectioned and processed through a series of alcohol/xylene baths.
Tissue sections were made (~4-µm) on glass slides and stained with hematoxylin and eosin
(H&E) [83]. All slides were examined by a veterinary pathologist (D.K.M.) for histopathologic
analysis using principles for valid histopathology [84]. Digital images were acquired with a high-
resolution Olympus DP73 camera mounted to an Olympus BX53 microscope using Olympus
CellSens Software.

55
In vitro probe digestion with excised catheter supernatants. Mice infected with either
Xen29 WT or Xen29 double nuclease mutant S. aureus catheter biofilm infections were
sacrificed 6 days PI and catheters (including wide-margin of attached tissue) were carefully
excised using sterile surgical blades. Infected catheters were then placed in a 1.5 ml
microcentrifuge tube filled with 1 ml PBS with divalent cations, mashed with a sterile pestle, and
then sonicated for 1 hour in a Branson ultrasonic cleaner. The tube was then spun down at
17,000 G for 10 minutes and the supernatant was transferred to a clean tube. The supernatant was
subsequently used for 10 ul probe digestion assays (see in vitro microplate reader assays).
Results
The utility of the DyLight800 P2&3 imaging probe allows quenching of fluorescence
(OFF-position) when the probe is in contact with an off-target nuclease, however when it comes in
contact with the target bacterial nuclease (MN) fluorescence is unquenched (ON-position) by way
of enzymatic digestion of the oligonucleotide (Fig. 13). We found that the original NMTT probe
evaluated by Hernandez et al. was not the most efficiently digested by MN within our sequence
library. We therefore compared kinetic probe digestion by MN between the NMTT probe and TT-
probe variants where the MN TT-cut site was located at various positions along the 11mer
sequence with all other nucleotides being 2’-O-methyl modified uridines (Fig. 14A). The results
showed that the P2&3 positional variant was the most efficiently digested over the course of 60
minutes. Our next step in second-generation NIR probe selection was to compare a few different
NIR fluorophore representatives along the NIR spectrum. This was accomplished by comparing in
vitro MN digestion assays in PBS and whole blood (Fig. 14B). The fluorophore selection criteria
involved the highest activation ratio (RFU of fully activated probe/RFU of intact probe) in blood
because whole blood mimics in vivo conditions more so than PBS. Therefore, the DyLight800

56
fluorophore was selected as the NIR fluorophore to pair with the P2&3 sequence to form the
second-generation NIR probe.
Our next step was to establish a S. aureus catheter-associated biofilm infection mouse
model (Fig. 15A). This biofilm infection protocol was established in SKH1 mice with WT
Xenogen29 strain S. aureus. The Xen29 strain was engineered to constitutively express
bioluminescence [97, 114-116] which enabled infection monitoring independent of fluorescence
imaging (Fig. 15B). Histological sections were then taken for H & E stains to compare the 4
biofilm catheter infection types (Fig. 15C). The comparison shows that there is a higher amount of
leukocyte infiltration with necrotic debris extending into the surrounding tissue in the bacterial
catheter infections whereas there is only sterile inflammation observed in the sham catheter
infection. To evaluate Xen29 biofilm formation on catheter implants, catheter segments were
removed and stained with SYT09 (live) followed by ToPro3 (dead) cell stains 6 days PI. Stained
catheters were then sectioned and imaged with a fluorescence microscope (Fig. 15D). Prominent
live cell staining was observed along with dead cell staining to a lesser extent on implanted
catheters.
Due to the high sensitivity of the Licor Pearl Trilogy NIR small animal imaging to 800
nanometer fluorophores, WT S. aureus Hg001 strain catheter biofilm infections were imaged with
3 concentrations (0.25, 0.5, and 1 nanomole concentrations) of the DyLight800-P2&3 probe to
determine the optimal imaging concentration for subsequent experiments (Table 3). The
concentration of probe that enabled the greatest target-to-background ratio while conferring the
lowest off-target fluorescence from the spleen/liver was 0.5 nanomoles. Fig. 16A shows the
comparison of DyLight800-P2&3 concentrations evaluated with the best target-to-background
ratio being shown for the best of 3 separate imaging experiments per probe concentration. A full

57
biofilm imaging experiment of 0.5 nanomole probe concentration is displayed in Fig. 16B with the
highest target-to-background ROI timepoint image included. The difference in RFU values
between the biofilm infection and contralateral control sites can be distinguished in as little as 25
minutes.
The specificity of the DyLight800-P2&3 NIR probe was evaluated by imaging catheter-
associated biofilm infections that were established with 3 control groups: S. epidermidis 1457,
sham-infection, and Xen29 nuclease mutant. S. epidermidis is the second-leading cause of I.V.
catheter biofilm infections world-wide [117]. Therefore, a noninvasive imaging technology to
rapidly distinguish between a WT S. aureus and S. epidermidis biofilm infection would be
beneficial. To this end, catheter biofilm infections were established with a commonly used S.
epidermidis (1457) strain capable of biofilm formation and imaged with 0.5 nanomoles of the
DyLight800-P2&3 probe (Fig. 17A). As illustrated in the figure, there is no difference observed
between the biofilm infection site and the contralateral control site over the 2.5-hour imaging
period following I.V. probe administration. Another significant characteristic of a noninvasive
imaging technology for S. aureus catheter biofilm infections is the ability to distinguish between
an actual infection and sterile inflammation. To evaluate second-generation NIR probe activation
in the context of sterile inflammation, sham catheter infections were established in mice and
imaged with a 0.5 nanomole concentration of probe (Fig. 17B). Results indicate that WT S. aureus
biofilm infections can be distinguished from sterile inflammation using the DyLight800-P2&3
probe. Because the DyLight800-P2&3 probe is a MN-activatable probe, it was obvious to evaluate
activation of this probe in the context of a catheter biofilm infection established with a S. aureus
mutant strain that lacked MN production. The Xen29 nuclease 1 and 2 (nuclease 1/2 KO)
knockout mutant was used to establish catheter biofilm infections and then subsequently imaged

58
with 0.5 nanomoles DyLight800-P2&3 6 days PI (Fig. 17C). Results indicate that the nuclease 1/2
KO control group also had elevated signal at the infection site when compared to the contralateral
control site. We compared activation ratios (fluorescence of fully activated probe/ fluorescence of
intact probe) between all imaging experiments at the 90-minute PPA timepoint and found that a
distinction between the WT and mutant catheter biofilm infections could not be made using this
probe (Fig. 17D- left). To investigate further into this issue, we excised WT and nuclease mutant
infected catheters from mice 6 days PI. The catheters were then placed in 1 mL PBS with divalent
cations, mashed with a sterile pestle, and then sonicated for 30 minutes. Following sonication, the
tubes were spun down and supernatant was removed. The supernatant was then used in 10 uL in
vitro probe digestion assays (Fig. 17D- right). From these results, it does not appear that the
nuclease mutant is producing any alternate nucleases or proteases that are digesting the second-
generation NIR probe. We therefore believe that the issues with this control go beyond the scope
of this study.
Despite these results, statistical analysis was performed on each catheter biofilm infection
type. A student’s unpaired nonparametric T test (P < 0.05) was performed on the WT Xen29 S.
aureus group using Graphpad Prism version 7 (Fig. 18A). As illustrated in the figure, the WT
catheter biofilm can be detected from uninfected tissue site within 90 minutes of IV probe
administration. S. epidermidis (Fig. 18B), sham (Fig. 18C), and Xen29 nuclease KO (Fig. 18D)
catheter biofilm control groups did not display differences from the respective control tissue ROI
sites. The analysis shows that a S. aureus catheter biofilm infection can be detected and imaged
within 90-minute probe administration using the second-generation NIR imaging probe.
Additionally, a WT S. aureus biofilm infection can be discerned from other relevant strain
infection types as well as sterile inflammation.

59
Discussion
The imaging technologies that are currently available cannot be used to directly detect
bacterial infections. Instead they can only be used to gage downstream effects of the infection such
as immune response and tissue necrosis. Therefore, the type of bacteria involved, and the site of
potential infection cannot be determined using current technologies. The second-generation
imaging probe generated in our study displays a proof-of-concept that can be translated clinically
once properly modified.
The modified RNA P2&3 variant sequence portion of the probe was selected due to higher
sensitivity to MN digestion when compared to its predecessor. The DyLight800 NIR fluorophore
was selected to label the sequence because it displayed the highest activation ratio in whole blood
of all the 700 and 800-nm fluorophores tested in addition to longer wavelength fluorescence
emission than the Cy5.5 NIR fluorophore. Activation of the second-generation probe in culture
supernatants and infected animals can be attributed to MN (by WT S. aureus) because other
species including the nuclease mutant staph species did not digest the probe. This probe also
confers resistance to degradation by human and mouse serum nucleases, a characteristic that can
be attributed to the 2’-O-methyl modifications to nucleotides within the probe sequence. This
modification is known to confer a high degree of resistance to degradation by endogenous
nucleases.
Although we observed increased signal when imaging nuclease mutant infected mice, we
did not observe probe activation when those infected catheters were removed from mice and
processed for in vitro digestion assays with the second-generation NIR probe. The results from the
digestion assays support the idea of an enhanced permeability and retention effect (EPR) when
infected tissue gets damaged to the point that it becomes highly permeable allowing certain
molecules in circulation to accumulate [118, 119]. However, digestion by other enzymes secreted

60
by S. aureus (not previously considered) cannot be ruled out [120]. Lastly, our laboratory has
benchmark values of where we expect the activation ratio (≥ 5) of an imaging probe used in
animal studies should be in order to be a successful diagnostic candidate. As shown in Figure 17D,
the activation ratio of Xen29 WT S. aureus catheter biofilm was less than 4. Generally, probes that
display activation ratio this low are not promising diagnostic candidates due to the poor signal-to-
noise ratio. However, the second-generation NIR probe still has the potential to address an unmet
clinical need with minimal modifications that can perhaps be addressed in the biotechnology
sector.

61
Figure 13. Cartoon illustration of oligonucleotide probe activation/digestion by MN.
In the presence of an off-target nuclease (i.e. endogenous serum nuclease) probe remains intact in an “OFF” state, however when the probe is in the presence of the target bacterial nuclease (i.e. MN) the oligonucleotide sequence is cleaved whereas the fluorophore is separated from the quencher transitioning to the “ON” state.
Off On
OffOff
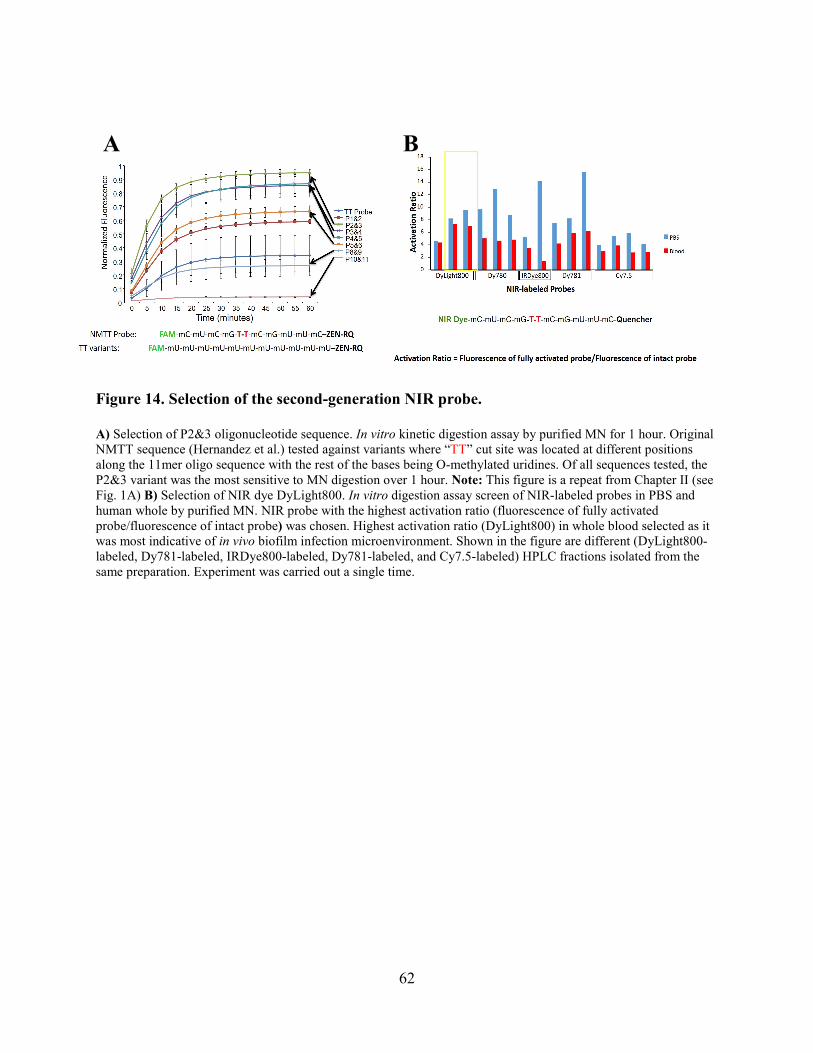
62
Figure 14. Selection of the second-generation NIR probe.
A) Selection of P2&3 oligonucleotide sequence. In vitro kinetic digestion assay by purified MN for 1 hour. Original NMTT sequence (Hernandez et al.) tested against variants where “TT” cut site was located at different positions along the 11mer oligo sequence with the rest of the bases being O-methylated uridines. Of all sequences tested, the P2&3 variant was the most sensitive to MN digestion over 1 hour. Note: This figure is a repeat from Chapter II (see Fig. 1A) B) Selection of NIR dye DyLight800. In vitro digestion assay screen of NIR-labeled probes in PBS and human whole by purified MN. NIR probe with the highest activation ratio (fluorescence of fully activated probe/fluorescence of intact probe) was chosen. Highest activation ratio (DyLight800) in whole blood selected as it was most indicative of in vivo biofilm infection microenvironment. Shown in the figure are different (DyLight800-labeled, Dy781-labeled, IRDye800-labeled, Dy781-labeled, and Cy7.5-labeled) HPLC fractions isolated from the same preparation. Experiment was carried out a single time.
A
B
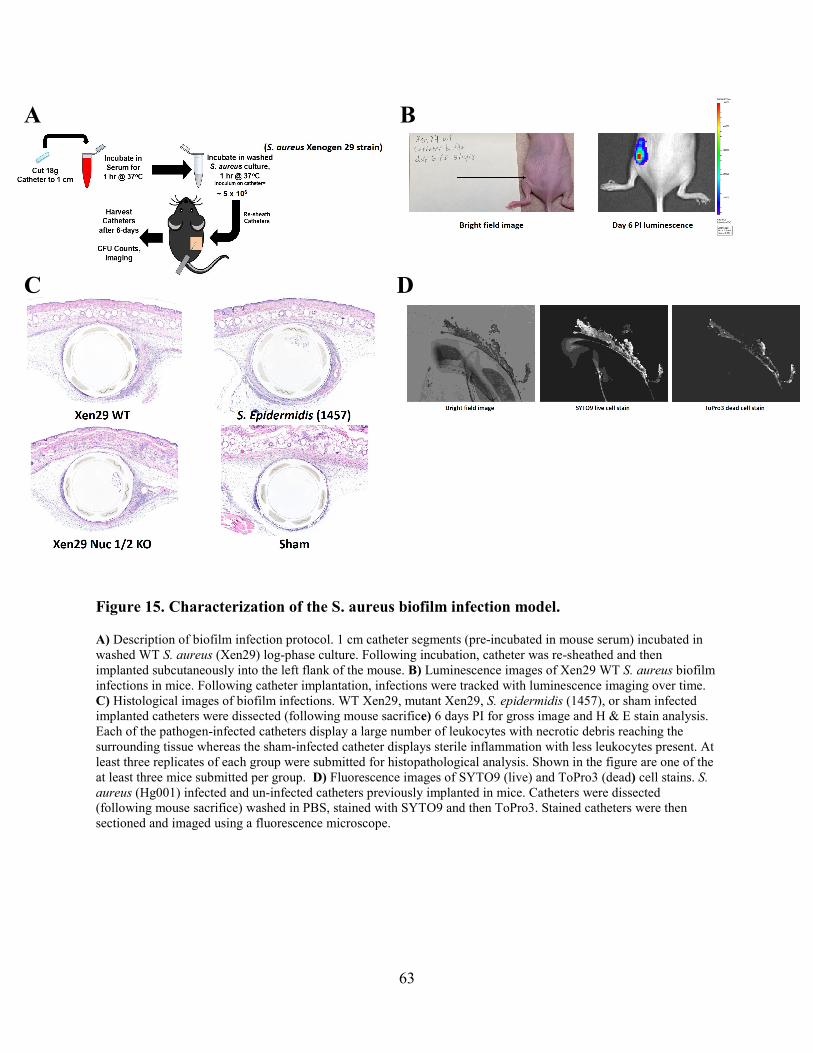
63
Figure 15. Characterization of the S. aureus biofilm infection model.
A) Description of biofilm infection protocol. 1 cm catheter segments (pre-incubated in mouse serum) incubated in washed WT S. aureus (Xen29) log-phase culture. Following incubation, catheter was re-sheathed and then implanted subcutaneously into the left flank of the mouse. B) Luminescence images of Xen29 WT S. aureus biofilm infections in mice. Following catheter implantation, infections were tracked with luminescence imaging over time. C) Histological images of biofilm infections. WT Xen29, mutant Xen29, S. epidermidis (1457), or sham infected implanted catheters were dissected (following mouse sacrifice) 6 days PI for gross image and H & E stain analysis. Each of the pathogen-infected catheters display a large number of leukocytes with necrotic debris reaching the surrounding tissue whereas the sham-infected catheter displays sterile inflammation with less leukocytes present. At least three replicates of each group were submitted for histopathological analysis. Shown in the figure are one of the at least three mice submitted per group. D) Fluorescence images of SYTO9 (live) and ToPro3 (dead) cell stains. S. aureus (Hg001) infected and un-infected catheters previously implanted in mice. Catheters were dissected (following mouse sacrifice) washed in PBS, stained with SYTO9 and then ToPro3. Stained catheters were then sectioned and imaged using a fluorescence microscope.
B
A
A
C
D
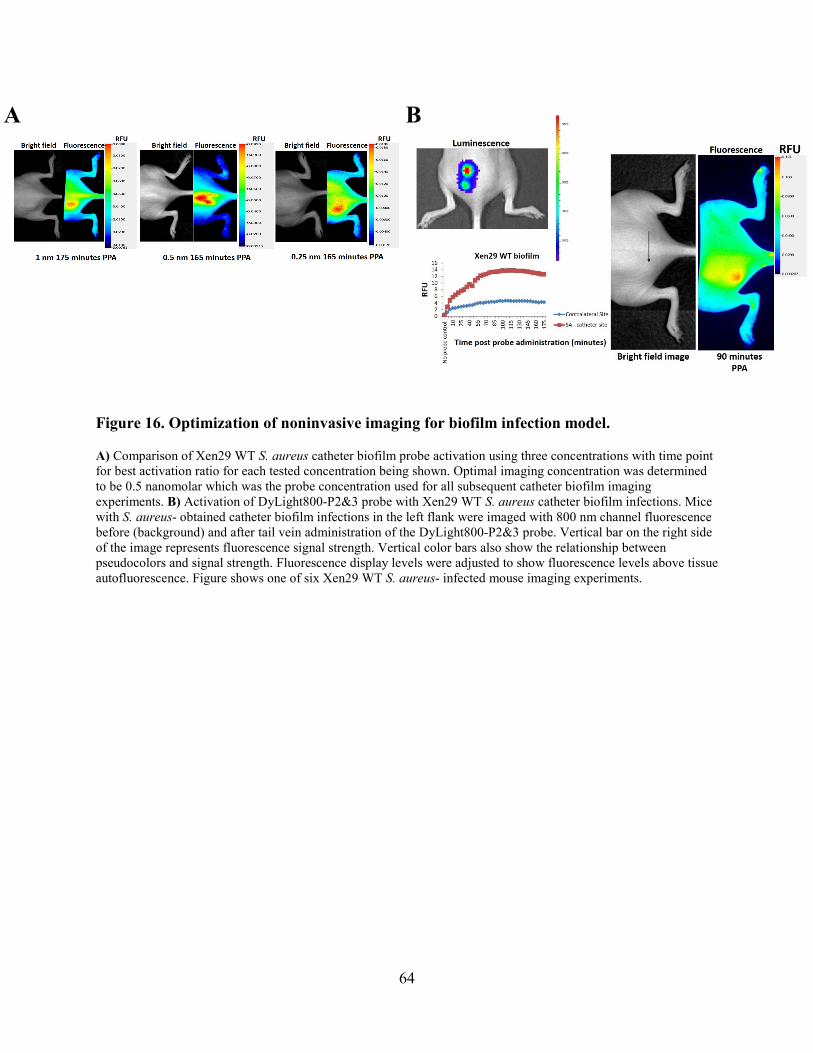
64
Figure 16. Optimization of noninvasive imaging for biofilm infection model.
A) Comparison of Xen29 WT S. aureus catheter biofilm probe activation using three concentrations with time point for best activation ratio for each tested concentration being shown. Optimal imaging concentration was determined to be 0.5 nanomolar which was the probe concentration used for all subsequent catheter biofilm imaging experiments. B) Activation of DyLight800-P2&3 probe with Xen29 WT S. aureus catheter biofilm infections. Mice with S. aureus- obtained catheter biofilm infections in the left flank were imaged with 800 nm channel fluorescence before (background) and after tail vein administration of the DyLight800-P2&3 probe. Vertical bar on the right side of the image represents fluorescence signal strength. Vertical color bars also show the relationship between pseudocolors and signal strength. Fluorescence display levels were adjusted to show fluorescence levels above tissue autofluorescence. Figure shows one of six Xen29 WT S. aureus- infected mouse imaging experiments.
A
B
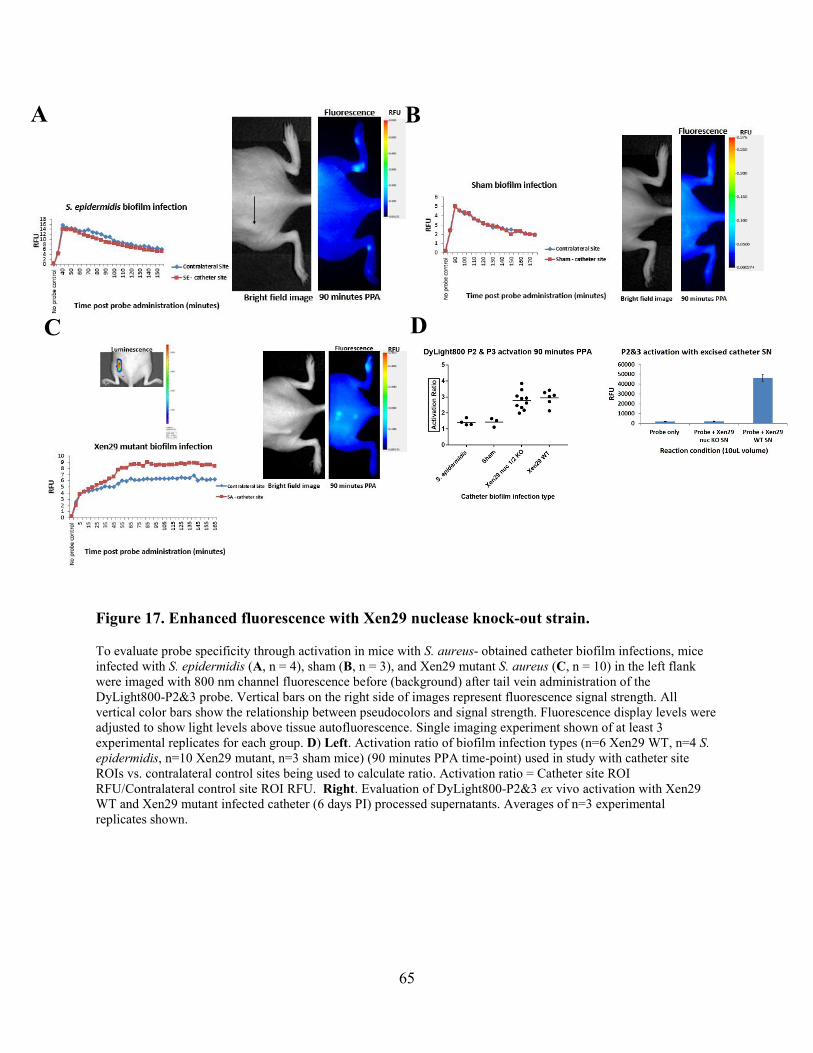
65
Figure 17. Enhanced fluorescence with Xen29 nuclease knock-out strain.
To evaluate probe specificity through activation in mice with S. aureus- obtained catheter biofilm infections, mice infected with S. epidermidis (A, n = 4), sham (B, n = 3), and Xen29 mutant S. aureus (C, n = 10) in the left flank were imaged with 800 nm channel fluorescence before (background) after tail vein administration of the DyLight800-P2&3 probe. Vertical bars on the right side of images represent fluorescence signal strength. All vertical color bars show the relationship between pseudocolors and signal strength. Fluorescence display levels were adjusted to show light levels above tissue autofluorescence. Single imaging experiment shown of at least 3 experimental replicates for each group. D) Left. Activation ratio of biofilm infection types (n=6 Xen29 WT, n=4 S. epidermidis, n=10 Xen29 mutant, n=3 sham mice) (90 minutes PPA time-point) used in study with catheter site ROIs vs. contralateral control sites being used to calculate ratio. Activation ratio = Catheter site ROI RFU/Contralateral control site ROI RFU. Right. Evaluation of DyLight800-P2&3 ex vivo activation with Xen29 WT and Xen29 mutant infected catheter (6 days PI) processed supernatants. Averages of n=3 experimental replicates shown.
A
B
D
C
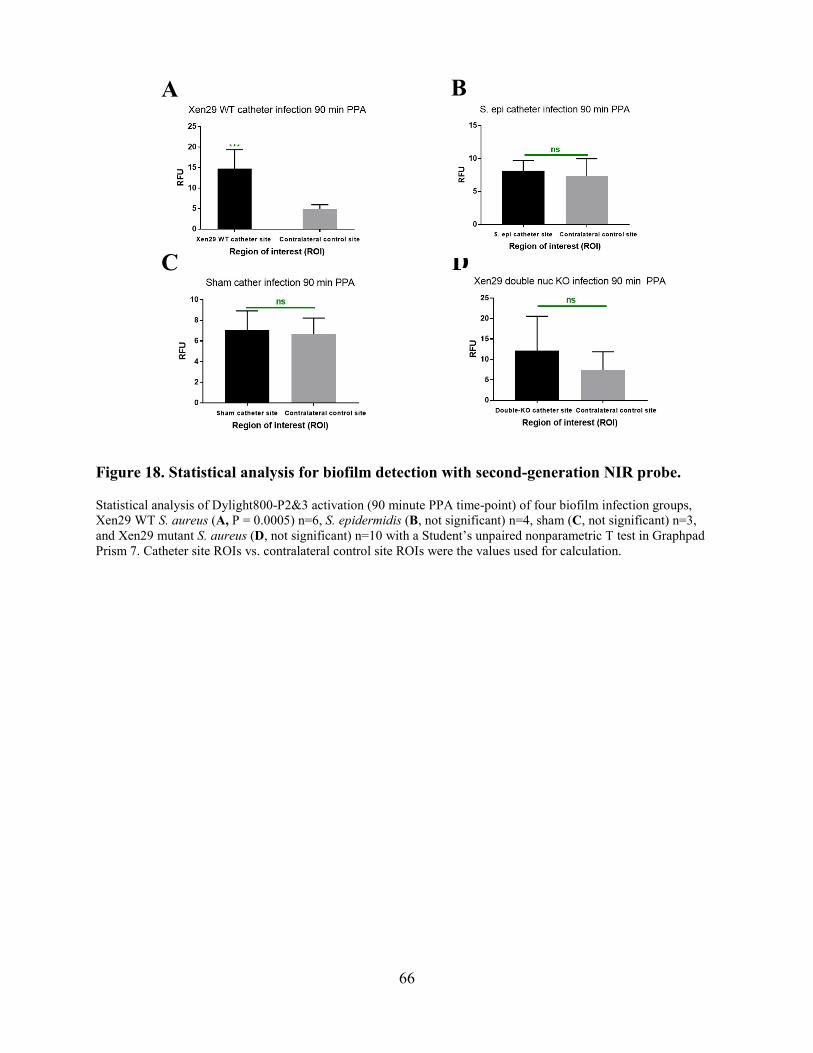
66
Figure 18. Statistical analysis for biofilm detection with second-generation NIR probe.
Statistical analysis of Dylight800-P2&3 activation (90 minute PPA time-point) of four biofilm infection groups, Xen29 WT S. aureus (A, P = 0.0005) n=6, S. epidermidis (B, not significant) n=4, sham (C, not significant) n=3, and Xen29 mutant S. aureus (D, not significant) n=10 with a Student’s unpaired nonparametric T test in Graphpad Prism 7. Catheter site ROIs vs. contralateral control site ROIs were the values used for calculation.
D
A
B
C

67
Table 4. Bacterial strains used in Chapter IV.
All bacterial strains used in this chapter were biofilm-forming strains and were acquired from the Horswill laboratory, Department of Immunology and Microbiology, University of Colorado.
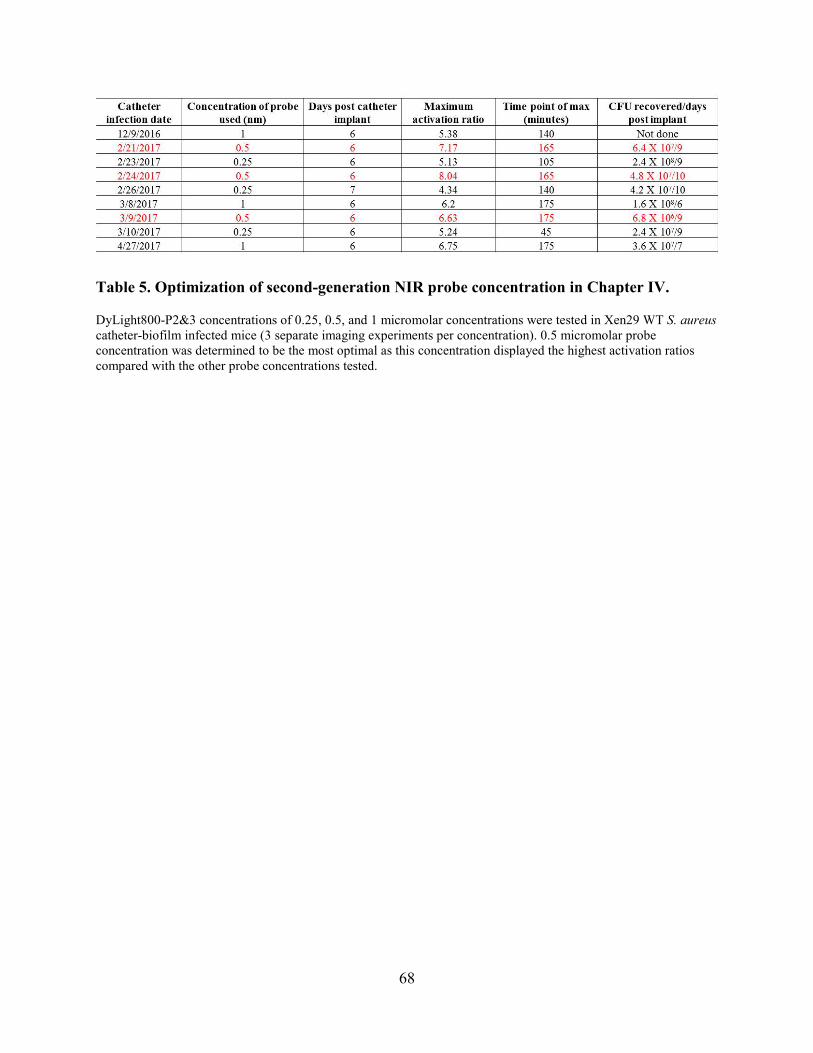
68
Table 5. Optimization of second-generation NIR probe concentration in Chapter IV.
DyLight800-P2&3 concentrations of 0.25, 0.5, and 1 micromolar concentrations were tested in Xen29 WT S. aureus catheter-biofilm infected mice (3 separate imaging experiments per concentration). 0.5 micromolar probe concentration was determined to be the most optimal as this concentration displayed the highest activation ratios compared with the other probe concentrations tested.

69
CHAPTER V: CONCLUSIONS AND FUTURE DIRECTIONS
As resistance and susceptibility profiles of bacteria continue to change, it is crucial that
diagnostic methodologies evolve as well to provide more rapid and definitive information. This
information could enable physicians to make more informed decisions about antibiotic therapy.
Optical imaging of bacterial infections is a recent methodology that offers great promise for
detecting pathogens in a shorter timeframe with greater certainty. Because there are no molecular
imaging probes approved for imaging pathogenic infections, we conducted two proof-of-concept
studies (Chapter II and IV) with high translatability to the clinic. In a previous study, our group
was the first to demonstrate noninvasive imaging of a S. aureus pyomyositis infection in mice
using a nuclease-activatable imaging probe [31]. This novel technology allowed us to detect and
image S. aureus thigh muscle infections in mice within 60 minutes of IV probe administration.
Although this imaging technology was shown to be effective in real-time detection of S. aureus
pyomyositis infections, we sought to investigate the detection of additional clinically relevant
infectious disease states commonly seen in hospitals worldwide. This thesis describes a
multiplex imaging approach that allows rapid diagnosis of skin infections by two leading SSTI
pathogens, S. aureus and S. pyogenes (Chapter II). It also characterizes NIR probes in
hydrophobic and hydrophilic environments (Chapter III) and the development of a second-
generation NIR imaging probe to noninvasively image S. aureus catheter biofilm infections in
mice (Chapter IV).
In chapter II, we describe a novel rapid optical imaging approach based on a multiplex
concept capable of detecting two different pathogens involved in skin infections. One of the
probes used in this approach (FAM-P2&3 TT-variant) was selected for its sensitivity to the MN
enzyme (Fig. 1A) secreted by S. aureus while the other probe (ATTO-56502’-FL-SH) was
selected for its sensitivity to an enzyme secreted by S. pyogenes GAS (Fig. 2A). Both probes

70
displayed resistance to digestion by the nucleases of other skin pathogens in addition to
endogenous skin nucleases (Figs. 1/2B and 1/2C) likely as a result of the modifications to the
oligonucleotide sequences in each probe. We initially tested the multiplex methodology to
validate optical imaging application using an equal concentration of both probes in the in vitro
activation reaction (Fig. 3). Following confirmation, imaging experiments were carried out with
S. aureus and S. pyogenes skin-infected mice 24H PI using the multiplexed probe combination in
a gel matrix. Statistical analysis revealed that one skin infection type could be distinguished
from the other within 55 minutes of gelled multiplex probe application using a simple 2-step
algorithm (Fig. 5). Moreover, this multiplex imaging approach has the potential to provide
physicians with rapid (less than an hour) diagnostic information that enables targeted antibiotic
therapy while avoiding the side-effects of broad-spectrum therapies.
The results of the above study show the utility of optical imaging as a rapid clinical
diagnostic tool. Given the timeframe typically associated with identifying pathogens involved in
infections using traditional culture methods, efforts to introduce novel rapid diagnostic methods
are greatly needed. The multiplex concept described in this study is the first optical imaging
methodology capable of identifying more than one pathogen, as far as we know. Another
possible clinical application of rapid diagnostic methodologies using the multiplex concept
would be through an in vitro assay where a swabbed sample (from potential infection site) could
be taken and used to digest one (or the other) probe in the multiplex matrix prior to reading the
enzymatic digestion on a plate reader.
The work in chapter II demonstrates a proof-of-concept realization for an alternative
methodology of rapidly detecting clinically relevant infections in a rapid manner with optical
imaging. Additional optimization of the ATTO-565-2’-FL-SH through sequence modification

71
could eliminate the digestion of this probe by the MN enzyme (at higher concentrations)
observed in the multiplex infection imaging studies (Fig. 5A).
Further studies to determine if the multiplex imaging concept can be applied to
infections that are polymicrobial such as Diabetic Foot Infections (DFI) would be extremely
beneficial [121-125]. Researchers could first select oligonucleotide sequences that are selectively
digested by nucleases of particular bacterial species involved in an infection and follow this by
differentially labeling sequences with fluorophores of different wavelengths/colors. The
multiplex imaging concept would then be used to detect different bacterial species involved in a
polymicrobial infection.
Polymicrobial infections present a growing healthcare problem [126]. These types of
infections are more often associated with patients that have underlying medical conditions, such
as diabetes mellitus [127]. Due to the progressive nature of these types of infections, by the time
the pathogens are identified the patient has already began to experience severe complications
such as bacteremia and toxic shock syndrome (TSS). The platform capability of the multiplex
imaging technology described in Chapter II could be applied to polymicrobial infections prior to
the spread of infection. The main challenge would be developing individual modified sequences
that target the enzymatic activity of specific nucleases. To address this issue, developing a high
throughput enzymatic screening assay could be developed to profile nuclease activity and
identify preferred substrates [128]. However, this method could present substantial challenges
especially if the target nuclease is not identified and characterized. Therefore, individually
targeting nucleases of pathogenic bacterial strains in a step-wise manner is currently the most
direct method to multiplex various strains. Culture supernatants would initially be tested with
single and double-stranded unmodified sequences to determine preferential cleavage. The

72
subsequent steps would involve base modifications to the selected sequence in order to tailor its
digestion to the targeted nuclease [31, 86, 129]. Once a modified sequence is developed for the
targeted nuclease, further characterization could then be accomplished. The same process would
be applied to other pathogenic species commonly involved in polymicrobial infections. The final
step would involve labelling each nuclease-specific modified sequence with different color
fluorophores to be incorporated into the multiplex imaging technology.
In chapter III, we compare NIR probes diluted in whole mouse/human blood to NIR
probes diluted in less aqueous solutions such as PBS. Unquenched probe conditions were
initially used as positive controls for in vitro MN digestion assays that also included quenched
versions. We found that unquenched NIR probes display enhanced fluorescence when diluted in
whole blood. Conversely, we found that these same unquenched probes displayed decreased
fluorescence when diluted in PBS. These observations are likely the results of NIR probes
interacting with proteins in blood to display higher fluorescence while self-quenching in
hydrophilic environments like PBS. Data from this study contributed to the development of the
second-generation NIR probe used for the catheter biofilm imaging described in chapter IV.
It is important to understand which interactions of NIR probes in blood are associated
with increased fluorescence as this correlates to higher background fluorescence which lowers
signal to noise ratio in fluorescence imaging studies [105]. Characterization of a wider range of
800-900 nanometer probes would be useful for further optimization of a deep tissue imaging
probe. As stated in the chapter, there should be a multidisciplinary approach to develop these
probes in the field of biomedical imaging.
In chapter IV, we describe an investigation to noninvasively image S. aureus catheter-
biofilm infections with the second-generation DyLight800-P2&3 probe. The oligonucleotide

73
sequence for this probe was selected because its sensitivity surpasses the first-generation TT
probe (Fig. 1B). The fluorophore used to label the sequence was the DyLight800 which was
selected because it has the highest activation ratio in blood when compared to the other NIR
fluorophores tested (Fig. 1C). We carried out fluorescence imaging experiments with S. aureus
(Xen29 WT), S, epidermidis (1457), S. aureus (Xen29 nuclease mutant), and sham-infected
catheter biofilm infections 6 days PI using the DyLight800-P2&3 probe. Statistical analysis
showed that the WT S. aureus biofilm infection was detected within 90 minutes of IV probe
administration (Fig. 5A) while the probe remained relatively silent in the 3 control biofilm
infections (Figs. 5BCD). Despite these results, we observed an elevated fluorescence signal at
the biofilm infection site in the Xen29 nuclease mutant control group (Fig. 4D left panel). To
determine whether the mutant strain produced nucleases/proteases that were not accounted for,
we processed biofilm infections 6 days PI and used the supernatant for in vitro digestion assays
(Fig, 4D right panel). Data from this experiment support the EPR theory that damaged tissue
becomes permeable allowing undigested probe to accumulate in those regions [118, 119, 130].
However, alternate nuclease/protease activity cannot be completely ruled out [120].
To look further into the nuclease mutant issue in the future, we plan to excise biofilm-
infected tissue with wide margins (including catheter) 6 days PI followed by direct fluorescence
imaging of the tissue compared to the WT infection. Results from this experiment could give
further insight to either the EPR effect or unaccounted for enzymes in the context of biofilm
infection environments. There is also potential to better optimize the second-generation NIR
probe to alleviate this issue at the biotech/pharma level. With proper modifications to this probe,
we believe that this technology is readily translatable to clinical practice as a noninvasive and
rapid infection diagnostic tool.

74
Collectively this thesis describes the selection and optimization of molecular imaging
probes to detect two clinically relevant classes of bacterial infections. Most optical imaging
methodologies available for use indirectly detect bacteria involved in pathogenic infections. Both
the skin and biofilm infection imaging technologies described here can directly detect bacteria in
an infection within the length of time of an average hospital visit. Additionally, the multiplex
imaging concept described in chapter II has the potential to be applied as an inexpensive in vitro
clinical infection diagnostic tool in the context of infections of the throat or other regions where
samples can be taken. Lastly, there is also potential to address definitive identification of
pathogens associated with polymicrobial infections using the multiplex imaging concept. We
believe that the work generated here contributes to both the literature on biomedical imaging as
well as clinical infection diagnostics fields and provides the groundwork for the development of
potential clinical applications in the near future.

75
REFERENCES
[1] J. Washington, Principles of Diagnosis, 4th ed., University of Texas Medical Branch at Galveston, Galveston (TX), 1996. [2] T.J. Beveridge, Use of the gram stain in microbiology, Biotechnic & histochemistry : official publication of the Biological Stain Commission 76(3) (2001) 111-8. [3] Quenching of Fluorescence, in: J. Lakowicz (Ed.), Principles of Fluorescence Spectroscopy, Springer US2006, pp. 277-330. [4] J.C. Lagier, S. Edouard, I. Pagnier, O. Mediannikov, M. Drancourt, D. Raoult, Current and past strategies for bacterial culture in clinical microbiology, Clinical microbiology reviews 28(1) (2015) 208-36. [5] O. Lazcka, F.J.D. Campo, F.X. Muñoz, Pathogen detection: A perspective of traditional methods and biosensors, Biosensors and Bioelectronics 22(7) (2007) 1205-1217. [6] J.R. Crowther, ELISA. Theory and practice, Methods in molecular biology (Clifton, N.J.) 42 (1995) 1-218. [7] P.E. Andreotti, G.V. Ludwig, A.H. Peruski, J.J. Tuite, S.S. Morse, L.F. Peruski, Jr., Immunoassay of infectious agents, BioTechniques 35(4) (2003) 850-9. [8] J. Weile, C. Knabbe, Current applications and future trends of molecular diagnostics in clinical bacteriology, Analytical and bioanalytical chemistry 394(3) (2009) 731-42. [9] W. Ludwig, Reprint of “Nucleic acid techniques in bacterial systematics and identification" [Int. J. Food Microbiol., 120 (2007) 225–236], International Journal of Food Microbiology 125(1) (2008) I-XII. [10] K.T. Konstantinidis, A. Ramette, J.M. Tiedje, The bacterial species definition in the genomic era, Philosophical transactions of the Royal Society of London. Series B, Biological sciences 361(1475) (2006) 1929-1940. [11] D. De Vos, A. Lim, Jr., J.P. Pirnay, M. Struelens, C. Vandenvelde, L. Duinslaeger, A. Vanderkelen, P. Cornelis, Direct detection and identification of Pseudomonas aeruginosa in clinical samples such as skin biopsy specimens and expectorations by multiplex PCR based on two outer membrane lipoprotein genes, oprI and oprL, Journal of clinical microbiology 35(6) (1997) 1295-1299. [12] R.H. Deurenberg, C. Vink, C. Driessen, M. Bes, N. London, J. Etienne, E.E. Stobberingh, Rapid detection of Panton–Valentine leukocidin from clinical isolates of Staphylococcus aureus strains by real-time PCR, FEMS Microbiology Letters 240(2) (2004) 225-228. [13] L.S. Burrack, D.E. Higgins, Genomic approaches to understanding bacterial virulence, Current Opinion in Microbiology 10(1) (2007) 4-9. [14] J.L. Moore, R.M. Caprioli, E.P. Skaar, Advanced mass spectrometry technologies for the study of microbial pathogenesis, Current opinion in microbiology 19 (2014) 45-51. [15] M. Welker, Proteomics for routine identification of microorganisms, Proteomics 11(15) (2011) 3143-53. [16] Y.P. Ho, P.M. Reddy, Advances in mass spectrometry for the identification of pathogens, Mass spectrometry reviews 30(6) (2011) 1203-24. [17] J.O. Lay, Jr., MALDI-TOF mass spectrometry of bacteria, Mass spectrometry reviews 20(4) (2001) 172-94. [18] P.A.C. Braga, A. Tata, V. Gonçalves dos Santos, J.R. Barreiro, N.V. Schwab, M. Veiga dos Santos, M.N. Eberlin, C.R. Ferreira, Bacterial identification: from the agar plate to the mass spectrometer, RSC Advances 3(4) (2013) 994-1008. [19] N. Singhal, M. Kumar, P.K. Kanaujia, J.S. Virdi, MALDI-TOF mass spectrometry: an emerging technology for microbial identification and diagnosis, Frontiers in microbiology 6 (2015) 791-791.

76
[20] A. Wieser, L. Schneider, J. Jung, S. Schubert, MALDI-TOF MS in microbiological diagnostics-identification of microorganisms and beyond (mini review), Applied microbiology and biotechnology 93(3) (2012) 965-74. [21] A. Mehl, B.O. Åsvold, A. Kümmel, S. Lydersen, J. Paulsen, I. Haugan, E. Solligård, J.K. Damås, S. Harthug, T.-H. Edna, Trends in antimicrobial resistance and empiric antibiotic therapy of bloodstream infections at a general hospital in Mid-Norway: a prospective observational study, BMC infectious diseases 17(1) (2017) 116-116. [22] B. Mills, M. Bradley, K. Dhaliwal, Optical imaging of bacterial infections, Clinical and Translational Imaging 4 (2016) 163-174. [23] T.C. Kwee, R.M. Kwee, A. Alavi, FDG-PET for diagnosing prosthetic joint infection: systematic review and metaanalysis, European Journal of Nuclear Medicine and Molecular Imaging 35(11) (2008) 2122-2132. [24] F. De Winter, D. Vogelaers, F. Gemmel, R.A. Dierckx, Promising role of 18-F-fluoro-D-deoxyglucose positron emission tomography in clinical infectious diseases, European journal of clinical microbiology & infectious diseases : official publication of the European Society of Clinical Microbiology 21(4) (2002) 247-57. [25] T.K. Chacko, H. Zhuang, K.Z. Nakhoda, B. Moussavian, A. Alavi, Applications of fluorodeoxyglucose positron emission tomography in the diagnosis of infection, Nucl Med Commun 24(6) (2003) 615-24. [26] K.D. Stumpe, H. Dazzi, A. Schaffner, G.K. von Schulthess, Infection imaging using whole-body FDG-PET, European journal of nuclear medicine 27(7) (2000) 822-32. [27] H. Zhuang, J.Q. Yu, A. Alavi, Applications of fluorodeoxyglucose-PET imaging in the detection of infection and inflammation and other benign disorders, Radiologic clinics of North America 43(1) (2005) 121-34. [28] H. Zhuang, P.S. Duarte, M. Pourdehnad, A. Maes, F. Van Acker, D. Shnier, J.P. Garino, R.H. Fitzgerald, A. Alavi, The promising role of 18F-FDG PET in detecting infected lower limb prosthesis implants, Journal of nuclear medicine : official publication, Society of Nuclear Medicine 42(1) (2001) 44-8. [29] A. Haroon, A. Zumla, J. Bomanji, Role of fluorine 18 fluorodeoxyglucose positron emission tomography-computed tomography in focal and generalized infectious and inflammatory disorders, Clinical infectious diseases : an official publication of the Infectious Diseases Society of America 54(9) (2012) 1333-41. [30] K. Shah, R. Weissleder, Molecular optical imaging: applications leading to the development of present day therapeutics, NeuroRx : the journal of the American Society for Experimental NeuroTherapeutics 2(2) (2005) 215-225. [31] F.J. Hernandez, L. Huang, M.E. Olson, K.M. Powers, L.I. Hernandez, D.K. Meyerholz, D.R. Thedens, M.A. Behlke, A.R. Horswill, J.O. McNamara Ii, Noninvasive imaging of Staphylococcus aureus infections with a nuclease-activated probe, Nat Med 20(3) (2014) 301-306. [32] V. Marx, Probes: seeing in the near infrared, Nat Meth 11(7) (2014) 717-720. [33] R. Weissleder, V. Ntziachristos, Shedding light onto live molecular targets, Nat Med 9(1) (2003) 123-8. [34] J.T. Alander, I. Kaartinen, A. Laakso, T. Pätilä, T. Spillmann, V.V. Tuchin, M. Venermo, P. Välisuo, A Review of Indocyanine Green Fluorescent Imaging in Surgery, International Journal of Biomedical Imaging 2012 (2012) 940585. [35] M.V. Marshall, J.C. Rasmussen, I.C. Tan, M.B. Aldrich, K.E. Adams, X. Wang, C.E. Fife, E.A. Maus, L.A. Smith, E.M. Sevick-Muraca, Near-Infrared Fluorescence Imaging in Humans with Indocyanine Green: A Review and Update, Open surgical oncology journal (Online) 2(2) (2010) 12-25. [36] S. Keereweer, P.B.A.A. Van Driel, T.J.A. Snoeks, J.D.F. Kerrebijn, R.J. Baatenburg de Jong, A.L. Vahrmeijer, H.J.C.M. Sterenborg, C.W.G.M. Löwik, Optical Image-Guided Cancer Surgery: Challenges and Limitations, Clinical Cancer Research 19(14) (2013) 3745.

77
[37] S. Gioux, H.S. Choi, J.V. Frangioni, Image-Guided Surgery using Invisible Near-Infrared Light: Fundamentals of Clinical Translation, Molecular imaging 9(5) (2010) 237-255. [38] W.M. Leevy, N. Serazin, B.D. Smith, Optical Imaging of Bacterial Infection Models, Drug discovery today. Disease models 4(3) (2007) 91-97. [39] M. van Oosten, M. Hahn, L.M.A. Crane, R.G. Pleijhuis, K.P. Francis, J.M. van Dijl, G.M. van Dam, Targeted imaging of bacterial infections: advances, hurdles and hopes, FEMS Microbiology Reviews 39(6) (2015) 892-916. [40] F.J. Hernandez, K.R. Stockdale, L. Huang, A.R. Horswill, M.A. Behlke, J.O. McNamara, 2nd, Degradation of nuclease-stabilized RNA oligonucleotides in Mycoplasma-contaminated cell culture media, Nucleic acid therapeutics 22(1) (2012) 58-68. [41] J.R. Knox, P.C. Moews, J.-M. Frere, Molecular evolution of bacterial β-lactam resistance, Chemistry & Biology 3(11) (1996) 937-947. [42] F.F. Jed, M. Shahriar, Three Decades of the Class A β-Lactamase Acyl-Enzyme, Current Protein & Peptide Science 10(5) (2009) 401-407. [43] J.C. Hermann, L. Ridder, H.-D. Holtje, A.J. Mulholland, Molecular mechanisms of antibiotic resistance: QM/MM modelling of deacylation in a class A [small beta]-lactamase, Organic & Biomolecular Chemistry 4(2) (2006) 206-210. [44] J. Davies, Inactivation of antibiotics and the dissemination of resistance genes, Science 264(5157) (1994) 375. [45] R.E. Campbell, Realization of β-lactamase as a versatile fluorogenic reporter, Trends in Biotechnology 22(5) (2004) 208-211. [46] F. Wang, C. Cassidy, J.C. Sacchettini, Crystal Structure and Activity Studies of the Mycobacterium tuberculosis β-Lactamase Reveal Its Critical Role in Resistance to β-Lactam Antibiotics, Antimicrobial Agents and Chemotherapy 50(8) (2006) 2762-2771. [47] Y. Kong, H. Yao, H. Ren, S. Subbian, S.L.G. Cirillo, J.C. Sacchettini, J. Rao, J.D. Cirillo, Imaging tuberculosis with endogenous β-lactamase reporter enzyme fluorescence in live mice, Proceedings of the National Academy of Sciences of the United States of America 107(27) (2010) 12239-12244. [48] K.M. DiVittorio, W.M. Leevy, E.J. O’Neil, J.R. Johnson, S. Vakulenko, J.D. Morris, K.D. Rosek, N. Serazin, S. Hilkert, S. Hurley, M. Marquez, B.D. Smith, Zinc(II)-Coordination Complexes as Membrane Active Fluorescent Probes and Antibiotics, Chembiochem : a European journal of chemical biology 9(2) (2008) 286-293. [49] W.M. Leevy, J.R. Johnson, C. Lakshmi, J. Morris, M. Marquez, B.D. Smith, Selective recognition of bacterial membranes by zinc(ii)-coordination complexes, Chemical Communications (15) (2006) 1595-1597. [50] A.G. White, B.D. Gray, K.Y. Pak, B.D. Smith, Deep-red fluorescent imaging probe for bacteria, Bioorganic & medicinal chemistry letters 22(8) (2012) 2833-2836. [51] W.M. Leevy, S.T. Gammon, H. Jiang, J.R. Johnson, D.J. Maxwell, E.N. Jackson, M. Marquez, D. Piwnica-Worms, B.D. Smith, Optical Imaging of Bacterial Infection in Living Mice Using a Fluorescent Near-Infrared Molecular Probe, Journal of the American Chemical Society 128(51) (2006) 16476-16477. [52] W.M. Leevy, S.T. Gammon, J.R. Johnson, A.J. Lampkins, H. Jiang, M. Marquez, D. Piwnica-Worms, M.A. Suckow, B.D. Smith, Noninvasive Optical Imaging of Staphylococcus aureus Bacterial Infection in Living Mice Using a Bis-Dipicolylamine-Zinc(II) Affinity Group Conjugated to a Near-Infrared Fluorophore, Bioconjugate Chemistry 19(3) (2008) 686-692. [53] J.R. Johnson, N. Fu, E. Arunkumar, W.M. Leevy, S.T. Gammon, D. Piwnica-Worms, B.D. Smith, Squaraine rotaxanes: superior substitutes for Cy-5 in molecular probes for near-infrared fluorescence cell imaging, Angewandte Chemie (International ed. in English) 46(29) (2007) 5528-31.

78
[54] A.G. White, N. Fu, W.M. Leevy, J.-J. Lee, M.A. Blasco, B.D. Smith, Optical Imaging of Bacterial Infection in Living Mice Using Deep-Red Fluorescent Squaraine Rotaxane Probes, Bioconjugate Chemistry 21(7) (2010) 1297-1304. [55] R.G. Hanshaw, C. Lakshmi, T.N. Lambert, J.R. Johnson, B.D. Smith, Fluorescent Detection of Apoptotic Cells by Using Zinc Coordination Complexes with a Selective Affinity for Membrane Surfaces Enriched with Phosphatidylserine, ChemBioChem 6(12) (2005) 2214-2220. [56] M.L. Thakur, K. Zhang, B. Paudyal, D. Devakumar, M.Y. Covarrubias, C. Cheng, B.D. Gray, E. Wickstrom, K.Y. Pak, Targeting Apoptosis for Optical Imaging of Infection, Molecular Imaging and Biology 14(2) (2012) 163-171. [57] E.N. Tang, A. Nair, D.W. Baker, W. Hu, J. Zhou, In Vivo Imaging of Infection Using a Bacteria-Targeting Optical Nanoprobe, Journal of biomedical nanotechnology 10(5) (2014) 856-863. [58] R. Friedrich, P. Panizzi, P. Fuentes-Prior, K. Richter, I. Verhamme, P.J. Anderson, S.-I. Kawabata, R. Huber, W. Bode, P.E. Bock, Staphylocoagulase is a prototype for the mechanism of cofactor-induced zymogen activation, Nature 425 (2003) 535. [59] P. Panizzi, R. Friedrich, P. Fuentes-Prior, K. Richter, P.E. Bock, W. Bode, Fibrinogen Substrate Recognition by Staphylocoagulase·(Pro)thrombin Complexes, The Journal of biological chemistry 281(2) (2006) 1179-1187. [60] L. Loeb, The Influence of certain Bacteria on the Coagulation of the Blood, The Journal of Medical Research 10(3) (1903) 407-419. [61] P. Panizzi, M. Nahrendorf, J.-L. Figueiredo, J. Panizzi, B. Marinelli, Y. Iwamoto, E. Keliher, A.A. Maddur, P. Waterman, H.K. Kroh, F. Leuschner, E. Aikawa, F.K. Swirski, M.J. Pittet, T.M. Hackeng, P. Fuentes-Prior, O. Schneewind, P.E. Bock, R. Weissleder, In vivo detection of Staphylococcus aureus endocarditis by targeting pathogen-specific prothrombin activation, Nature Medicine 17 (2011) 1142. [62] S. Michael, C. Michelle, L. Panos, T. Mark, In Vivo Bacterial Imaging without Engineering; A Novel Probe-Based Strategy Facilitated by Endogenous Nitroreductase Enzymes, Current Gene Therapy 15(3) (2015) 277-288. [63] M.D. Roldán, E. Pérez-Reinado, F. Castillo, C. Moreno-Vivián, Reduction of polynitroaromatic compounds: the bacterial nitroreductases, FEMS Microbiology Reviews 32(3) (2008) 474-500. [64] L.F. Malheiro, R. Magano, A. Ferreira, A. Sarmento, L. Santos, Skin and soft tissue infections in the intensive care unit: a retrospective study in a tertiary care center, Revista Brasileira de Terapia Intensiva 29(2) (2017) 195-205. [65] A.F. Cardona, S.E. Wilson, Skin and soft-tissue infections: a critical review and the role of telavancin in their treatment, Clinical infectious diseases : an official publication of the Infectious Diseases Society of America 61 Suppl 2 (2015) S69-78. [66] L.J. Eron, B.A. Lipsky, D.E. Low, D. Nathwani, A.D. Tice, G.A. Volturo, Managing skin and soft tissue infections: expert panel recommendations on key decision points, The Journal of antimicrobial chemotherapy 52 Suppl 1 (2003) i3-17. [67] D.C. Vinh, J.M. Embil, Rapidly progressive soft tissue infections, The Lancet. Infectious diseases 5(8) (2005) 501-13. [68] M. Dryden, Complicated skin and soft tissue infections caused by methicillin-resistant Staphylococcus aureus: epidemiology, risk factors, and presentation, Surgical infections 9 Suppl 1 (2008) s3-10. [69] A. Jeng, M. Beheshti, J. Li, R. Nathan, The role of beta-hemolytic streptococci in causing diffuse, nonculturable cellulitis: a prospective investigation, Medicine 89(4) (2010) 217-26. [70] G.T. Ray, J.A. Suaya, R. Baxter, Microbiology of skin and soft tissue infections in the age of community-acquired methicillin-resistant Staphylococcus aureus, Diagnostic microbiology and infectious disease 76(1) (2013) 24-30.

79
[71] D.T.J.J.G.B.A.R.M.G.A.M.C.D.B.E.P.J.A.M.P. Musgrove, Disease Control Priorities in Developing Countries (Second Edition). [72] J. Hatzenbuehler, T.J. Pulling, Diagnosis and management of osteomyelitis, American family physician 84(9) (2011) 1027-33. [73] J.P. Burnham, J.P. Kirby, M.H. Kollef, Diagnosis and management of skin and soft tissue infections in the intensive care unit: a review, Intensive Care Medicine 42(12) (2016) 1899-1911. [74] European Centre for Disease Prevention and Control (2013) Annual Epidemiological Report. Reporting on 2010 surveillance data and 2011 epidemic intelligence data. ECDC, Stockholm, (2012). [75] J.G. Crisp, S.S. Takhar, G.J. Moran, A. Krishnadasan, S.E. Dowd, S.M. Finegold, P.H. Summanen, D.A. Talan, Inability of polymerase chain reaction, pyrosequencing, and culture of infected and uninfected site skin biopsy specimens to identify the cause of cellulitis, Clinical infectious diseases : an official publication of the Infectious Diseases Society of America 61(11) (2015) 1679-87. [76] D.L. Stevens, A.L. Bisno, H.F. Chambers, E.P. Dellinger, E.J.C. Goldstein, S.L. Gorbach, J.V. Hirschmann, S.L. Kaplan, J.G. Montoya, J.C. Wade, Practice Guidelines for the Diagnosis and Management of Skin and Soft Tissue Infections: 2014 Update by the Infectious Diseases Society of America, Clinical Infectious Diseases 59(2) (2014) e10-e52. [77] N.M. Watson ME Jr., Caparon MG, Animal Models of Streptococcus pyogenes Infection, Streptococcus pyogenes : Basic Biology to Clinical Manifestations [Internet] (2016). [78] G.C. Port, E. Paluscio, M.G. Caparon, Complete Genome Sequence of emm Type 14 Streptococcus pyogenes Strain HSC5, Genome Announc 1(4) (2013) e00612-13. [79] M.E. Watson, Jr., H.V. Nielsen, S.J. Hultgren, M.G. Caparon, Murine vaginal colonization model for investigating asymptomatic mucosal carriage of Streptococcus pyogenes, Infection and immunity 81(5) (2013) 1606-17. [80] A. Brenot, K.Y. King, B. Janowiak, O. Griffith, M.G. Caparon, Contribution of glutathione peroxidase to the virulence of Streptococcus pyogenes, Infection and immunity 72(1) (2004) 408-13. [81] M.N. Neely, J.D. Pfeifer, M. Caparon, Streptococcus-zebrafish model of bacterial pathogenesis, Infection and immunity 70(7) (2002) 3904-14. [82] E. Hanski, P.A. Horwitz, M.G. Caparon, Expression of protein F, the fibronectin-binding protein of Streptococcus pyogenes JRS4, in heterologous streptococcal and enterococcal strains promotes their adherence to respiratory epithelial cells, Infection and immunity 60(12) (1992) 5119-25. [83] D.A. Stoltz, D.K. Meyerholz, A.A. Pezzulo, S. Ramachandran, M.P. Rogan, G.J. Davis, R.A. Hanfland, C. Wohlford-Lenane, C.L. Dohrn, J.A. Bartlett, G.A.t. Nelson, E.H. Chang, P.J. Taft, P.S. Ludwig, M. Estin, E.E. Hornick, J.L. Launspach, M. Samuel, T. Rokhlina, P.H. Karp, L.S. Ostedgaard, A. Uc, T.D. Starner, A.R. Horswill, K.A. Brogden, R.S. Prather, S.S. Richter, J. Shilyansky, P.B. McCray, Jr., J. Zabner, M.J. Welsh, Cystic fibrosis pigs develop lung disease and exhibit defective bacterial eradication at birth, Sci Transl Med 2(29) (2010) 29ra31. [84] D.K. Meyerholz, A.P. Beck, Principles and approaches for reproducible scoring of tissue stains in research, Laboratory investigation; a journal of technical methods and pathology 98(7) (2018) 844-855. [85] B.R. Kelemen, T.A. Klink, M.A. Behlke, S.R. Eubanks, P.A. Leland, R.T. Raines, Hypersensitive substrate for ribonucleases, Nucleic acids research 27(18) (1999) 3696-3701. [86] M.A. Behlke, Chemical modification of siRNAs for in vivo use, Oligonucleotides 18(4) (2008) 305-19. [87] C.A. Hilliard, J. El Masri, M. Goto, Staphylococcus caprae bacteraemia and native bone infection complicated by therapeutic failure and elevated MIC: a case report, JMM case reports 4(9) (2017) e005112-e005112. [88] P. Seng, M. Barbe, P.O. Pinelli, F. Gouriet, M. Drancourt, A. Minebois, N. Cellier, C. Lechiche, G. Asencio, J.P. Lavigne, A. Sotto, A. Stein, Staphylococcus caprae bone and joint infections: a re-emerging infection?, Clinical microbiology and infection : the official publication of the European Society of Clinical Microbiology and Infectious Diseases 20(12) (2014) O1052-8.

80
[89] N.E. Natsis, P.R. Cohen, Coagulase-Negative Staphylococcus Skin and Soft Tissue Infections, American Journal of Clinical Dermatology 19(5) (2018) 671-677. [90] C. Chalmers, A.H.J. Khemlani, C.R. Sohn, J.M.S. Loh, C. Tsai, T. Proft, Streptococcus pyogenes nuclease A (SpnA) mediated virulence does not exclusively depend on nuclease activity, Journal of Microbiology, Immunology and Infection (2017). [91] W.G. Yasmineh, E.D. Gray, L.W. Wannamaker, Streptococcal nucleases. III. Kinetics of action and inhibition by transfer ribonucleic acid, Biochemistry 7(1) (1968) 91-97. [92] P. Ferrieri, E.D. Gray, L.W. Wannamaker, Biochemical and immunological characterization of the extracellular nucleases of group B streptococci, The Journal of experimental medicine 151(1) (1980) 56-68. [93] A.F. Moon, M. Midon, G. Meiss, A. Pingoud, R.E. London, L.C. Pedersen, Structural insights into catalytic and substrate binding mechanisms of the strategic EndA nuclease from Streptococcus pneumoniae, Nucleic Acids Res 39(7) (2011) 2943-53. [94] B.T. Ferreira, L.C. Benchetrit, A.C.D.D. Castro, T.G.F.M. Batista, L. Barrucand, Extracellular deoxyribonucleases of streptococci: a comparison of their occurrence and levels of production Smong beta-hemolytic strains of various serological groups, Zentralblatt für Bakteriologie 277(4) (1992) 493-503. [95] J. Probst, S. Brechtel, B. Scheel, I. Hoerr, G. Jung, H.-G. Rammensee, S. Pascolo, Characterization of the ribonuclease activity on the skin surface, Genetic Vaccines and Therapy 4(1) (2006) 4. [96] C.H. Contag, P.R. Contag, J.I. Mullins, S.D. Spilman, D.K. Stevenson, D.A. Benaron, Photonic detection of bacterial pathogens in living hosts, Molecular microbiology 18(4) (1995) 593-603. [97] Y.Q. Xiong, J. Willard, J.L. Kadurugamuwa, J. Yu, K.P. Francis, A.S. Bayer, Real-time in vivo bioluminescent imaging for evaluating the efficacy of antibiotics in a rat Staphylococcus aureus endocarditis model, Antimicrob Agents Chemother 49(1) (2005) 380-7. [98] K.P. Francis, D. Joh, C. Bellinger-Kawahara, M.J. Hawkinson, T.F. Purchio, P.R. Contag, Monitoring bioluminescent Staphylococcus aureus infections in living mice using a novel luxABCDE construct, Infection and immunity 68(6) (2000) 3594-600. [99] D.L. Stevens, A.E. Bryant, Impetigo, Erysipelas and Cellulitis, in: J.J. Ferretti, D.L. Stevens, V.A. Fischetti (Eds.), Streptococcus pyogenes : Basic Biology to Clinical Manifestations, University of Oklahoma Health Sciences Center
(c) The University of Oklahoma Health Sciences Center., Oklahoma City (OK), 2016. [100] N.K. Brogden, M. Milewski, P. Ghosh, L. Hardi, L.J. Crofford, A.L. Stinchcomb, Diclofenac delays micropore closure following microneedle treatment in human subjects, Journal of controlled release : official journal of the Controlled Release Society 163(2) (2012) 220-229. [101] V.J. Pansare, S. Hejazi, W.J. Faenza, R.K. Prud’homme, Review of Long-Wavelength Optical and NIR Imaging Materials: Contrast Agents, Fluorophores, and Multifunctional Nano Carriers, Chemistry of Materials 24(5) (2012) 812-827. [102] K. Umezawa, A. Matsui, Y. Nakamura, D. Citterio, K. Suzuki, Bright, Color-Tunable Fluorescent Dyes in the Vis/NIR Region: Establishment of New “Tailor-Made” Multicolor Fluorophores Based on Borondipyrromethene, Chemistry – A European Journal 15(5) (2009) 1096-1106. [103] X. Zhang, S. Bloch, W. Akers, S. Achilefu, Near-infrared molecular probes for in vivo imaging, Curr Protoc Cytom Chapter 12 (2012) Unit12.27. [104] K. Umezawa, D. Citterio, K. Suzuki, New trends in near-infrared fluorophores for bioimaging, Anal Sci 30(3) (2014) 327-49. [105] J.V. Frangioni, In vivo near-infrared fluorescence imaging, Current Opinion in Chemical Biology 7(5) (2003) 626-634.

81
[106] L.C. Zanetti-Domingues, C.J. Tynan, D.J. Rolfe, D.T. Clarke, M. Martin-Fernandez, Hydrophobic Fluorescent Probes Introduce Artifacts into Single Molecule Tracking Experiments Due to Non-Specific Binding, PLOS ONE 8(9) (2013) e74200. [107] S. Zhang, G. Li, J. Wang, D. Wang, Y. Han, H. Cao, L. Lin, Nondestructive Measurement of Hemoglobin in Blood Bags Based on Multi-Pathlength VIS-NIR Spectroscopy, Scientific Reports 8(1) (2018) 2204. [108] G. Kumar, J.M. Schmitt, Optimum wavelengths for measurement of blood hemoglobin content and tissue hydration by NIR spectrophotometry, SPIE1996. [109] R. Gahlot, C. Nigam, V. Kumar, G. Yadav, S. Anupurba, Catheter-related bloodstream infections, International Journal of Critical Illness and Injury Science 4(2) (2014) 162-167. [110] S. Hogan, M. Zapotoczna, N.T. Stevens, H. Humphreys, J.P. O'Gara, E. O'Neill, Eradication of Staphylococcus aureus Catheter-Related Biofilm Infections Using ML:8 and Citrox, Antimicrobial Agents and Chemotherapy 60(10) (2016) 5968-5975. [111] K.M. Bullard, D.L. Dunn, Diagnosis and treatment of bacteremia and intravascular catheter infections, The American Journal of Surgery 172(6, Supplement 1) (1996) 13s-19s. [112] J.M. Specht, D.A. Mankoff, Advances in molecular imaging for breast cancer detection and characterization, Breast Cancer Res 14(2) (2012) 206. [113] C.J. Anderson, J.W. Bulte, K. Chen, X. Chen, B.A. Khaw, M. Shokeen, K.L. Wooley, H.F. VanBrocklin, Design of targeted cardiovascular molecular imaging probes, Journal of nuclear medicine : official publication, Society of Nuclear Medicine 51 Suppl 1 (2010) 3s-17s. [114] J.L. Kadurugamuwa, L.V. Sin, J. Yu, K.P. Francis, R. Kimura, T. Purchio, P.R. Contag, Rapid direct method for monitoring antibiotics in a mouse model of bacterial biofilm infection, Antimicrobial agents and chemotherapy 47(10) (2003) 3130-3137. [115] J.L. Kadurugamuwa, L. Sin, E. Albert, J. Yu, K. Francis, M. DeBoer, M. Rubin, C. Bellinger-Kawahara, T.R. Parr, Jr., P.R. Contag, Direct continuous method for monitoring biofilm infection in a mouse model, Infection and immunity 71(2) (2003) 882-890. [116] J.L. Kadurugamuwa, L.V. Sin, J. Yu, K.P. Francis, T.F. Purchio, P.R. Contag, Noninvasive optical imaging method to evaluate postantibiotic effects on biofilm infection in vivo, Antimicrobial agents and chemotherapy 48(6) (2004) 2283-2287. [117] H. Shah, W. Bosch, K.M. Thompson, W.C. Hellinger, Intravascular Catheter-Related Bloodstream Infection, The Neurohospitalist 3(3) (2013) 144-151. [118] E.A. Azzopardi, E.L. Ferguson, D.W. Thomas, The enhanced permeability retention effect: a new paradigm for drug targeting in infection, Journal of Antimicrobial Chemotherapy 68(2) (2013) 257-274. [119] F. Romero Pastrana, J.M. Thompson, M. Heuker, H. Hoekstra, C.A. Dillen, R.V. Ortines, A.G. Ashbaugh, J.E. Pickett, M.D. Linssen, N.M. Bernthal, K.P. Francis, G. Buist, M. van Oosten, G.M. van Dam, D.L.J. Thorek, L.S. Miller, J.M. van Dijl, Noninvasive optical and nuclear imaging of Staphylococcus-specific infection with a human monoclonal antibody-based probe, Virulence 9(1) (2018) 262-272. [120] P. Tiet, K.C. Clark, J.O. McNamara, J.M. Berlin, Colorimetric Detection of Staphylococcus aureus Contaminated Solutions without Purification, Bioconjugate Chemistry 28(1) (2017) 183-193. [121] A.B. Taha, Relationship and susceptibility profile of Staphylococcus aureus infection diabetic foot ulcers with Staphylococcus aureus nasal carriage, The Foot 23(1) (2013) 11-16. [122] S. Saeed, J. Zafar, B. Khan, A. Akhtar, S. Qurieshi, S. Fatima, N. Ahmad, J. Irfanullah, Utility of 99mTc-labelled antimicrobial peptide ubiquicidin (29-41) in the diagnosis of diabetic foot infection, European Journal of Nuclear Medicine and Molecular Imaging 40(5) (2013) 737-743. [123] B.A. Lipsky, A.R. Berendt, P.B. Cornia, J.C. Pile, E.J.G. Peters, D.G. Armstrong, H.G. Deery, J.M. Embil, W.S. Joseph, A.W. Karchmer, M.S. Pinzur, E. Senneville, 2012 Infectious Diseases Society of America Clinical Practice Guideline for the Diagnosis and Treatment of Diabetic Foot Infections, Clinical Infectious Diseases 54(12) (2012) e132-e173.

82
[124] V.K. Sharma, P.B. Khadka, A. Joshi, R. Sharma, Common pathogens isolated in diabetic foot infection in Bir Hospital, Kathmandu University medical journal (KUMJ) 4(3) (2006) 295-301. [125] B.A. Lipsky, A.R. Berendt, H.G. Deery, J.M. Embil, W.S. Joseph, A.W. Karchmer, J.L. LeFrock, D.P. Lew, J.T. Mader, C. Norden, J.S. Tan, Diagnosis and Treatment of Diabetic Foot Infections, Clinical Infectious Diseases 39(7) (2004) 885-910. [126] K.A. Brogden, J.M. Guthmiller, C.E. Taylor, Human polymicrobial infections, Lancet (London, England) 365(9455) (2005) 253-5. [127] S.S. Wong, Y.S. Lin, L. Mathew, L. Rajagopal, D. Sepkowitz, Increase in group G streptococcal infections in a community hospital, New York, USA, Emerg Infect Dis 15(6) (2009) 991-993. [128] J.-P. Goddard, J.-L. Reymond, Enzyme assays for high-throughput screening, Current opinion in biotechnology 15 (2004) 314-22. [129] K.W. Thiel, P.H. Giangrande, Therapeutic applications of DNA and RNA aptamers, Oligonucleotides 19(3) (2009) 209-22. [130] H. Maeda, Vascular permeability in cancer and infection as related to macromolecular drug delivery, with emphasis on the EPR effect for tumor-selective drug targeting, Proceedings of the Japan Academy. Series B, Physical and biological sciences 88(3) (2012) 53-71.
Related Documents











