RAPD-PCR typing of Acinetobacter isolates from activated sludge systems designed to remove phosphorus microbiologically E. Carr 1 , H. Eason 1 , S. Feng 1 , A. Hoogenraad 2 , R. Croome 2 , J. Soddell 1 , K. Lindrea 1 and R. Seviour 1 1 Biotechnology Research Centre, La Trobe University, Bendigo, Victoria, and 2 Department of Environmental Management and Ecology, La Trobe University, Wodonga, Victoria, Australia 514/8/00: received 18 August 2000 and accepted 31 October 2000 E. CARR, H. EASON, S. FENG, A. HOOGENRAAD, R. CROOME, J. SODDELL, K. LINDREA AND R. SEVIOUR. 2001. Aims: This study investigated whether there were differences in RAPD fingerprints between already described genomic species of Acinetobacter and those from activated sludge systems. Whether plant-specific populations of acinetobacters exist was also examined. Methods and Results: Fifty-two isolates of Acinetobacter from four biological phosphorus removal (EBPR) systems of different configurations, and the known genomic species, were characterized using RAPD-PCR, and fragments separated on agarose gels. Patterns were analysed using Gel Pro software and data analysed numerically. RAPD-PCR produced patterns suggesting that many environmental isolates differ from known genomic species. In two cases, strains from individual plants clustered closely enough together to imply that there may be plant-specific populations of acinetobacters. Conclusions: The data suggest that current understanding of the taxonomic status of Acinetobacter may need modifying to accommodate non-clinical isolates, as many of the clusters emerging after numerical analysis of RAPD-PCR fragments from activated sludge isolates were quite separate from the clusters containing the already described genomic species. Some evidence was also obtained from the clusters generated to support a view that particular populations of Acinetobacter may occur in individual activated sludge plants. Significance and Impact of the Study: These data suggest that the current understanding of the systematics of Acinetobacter, based as it is almost exclusively on clinical isolates, may need drastic revision to accommodate environmental strains. They also suggest that a re-examination of the importance and role of Acinetobacter in the activated sludge process may be appropriate. INTRODUCTION Members of the genus Acinetobacter are ubiquitous organ- isms of increasing interest and importance to clinical microbiologists, as some are serious nosocomial pathogens and show increasing resistance to many commonly pre- scribed antibiotics (Bergogne-Berezin and Towner 1996). This genus has a very confused taxonomic history but is now placed in the c-Proteobacteria on 16S rDNA data; it is considered to be phylogenetically coherent (Rainey et al. 1994; Ibrahim et al. 1997) and can be delineated unequivo- cally using the transformation assay of Juni (1972). The species delineation of Acinetobacter still poses prob- lems, even after Bouvet and Grimont (1986), Bouvet and Jeanjean (1989) and Tjernberg and Ursing (1989) recognized 19 genomic species using DNA:DNA hybridization. Only seven of these species have been validly named. Tjernberg and Ursing (1989) showed that their DNA group 14 corresponded to DNA group 13 of Bouvet and Jeanjean (1989), while their DNA group 15 was novel and quite different to the Bouvet Correspondence to: Dr R. Seviour, Biotechnology Research Centre, La Trobe University, Bendigo, Victoria 3552, Australia (e-mail: [email protected]). ª 2001 The Society for Applied Microbiology Journal of Applied Microbiology 2001, 90, 309–319

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

RAPD-PCR typing of Acinetobacter isolates from activatedsludge systems designed to remove phosphorusmicrobiologically
E. Carr1, H. Eason1, S. Feng1, A. Hoogenraad2, R. Croome2, J. Soddell1,K. Lindrea1 and R. Seviour1
1Biotechnology Research Centre, La Trobe University, Bendigo, Victoria, and 2Department of Environmental
Management and Ecology, La Trobe University, Wodonga, Victoria, Australia
514/8/00: received 18 August 2000 and accepted 31 October 2000
E. CARR, H. EASON, S. FENG, A . HOOGENRAAD, R. CROOME, J . SODDELL, K . L INDREA
AND R. SEVIOUR. 2001.
Aims: This study investigated whether there were differences in RAPD ®ngerprints between
already described genomic species of Acinetobacter and those from activated sludge systems.
Whether plant-speci®c populations of acinetobacters exist was also examined.
Methods and Results: Fifty-two isolates of Acinetobacter from four biological phosphorus
removal (EBPR) systems of different con®gurations, and the known genomic species, were
characterized using RAPD-PCR, and fragments separated on agarose gels. Patterns were
analysed using Gel Pro software and data analysed numerically. RAPD-PCR produced patterns
suggesting that many environmental isolates differ from known genomic species. In two cases,
strains from individual plants clustered closely enough together to imply that there may be
plant-speci®c populations of acinetobacters.
Conclusions: The data suggest that current understanding of the taxonomic status of
Acinetobacter may need modifying to accommodate non-clinical isolates, as many of the clusters
emerging after numerical analysis of RAPD-PCR fragments from activated sludge isolates were
quite separate from the clusters containing the already described genomic species. Some
evidence was also obtained from the clusters generated to support a view that particular
populations of Acinetobacter may occur in individual activated sludge plants.
Signi®cance and Impact of the Study: These data suggest that the current understanding
of the systematics of Acinetobacter, based as it is almost exclusively on clinical isolates, may need
drastic revision to accommodate environmental strains. They also suggest that a re-examination
of the importance and role of Acinetobacter in the activated sludge process may be appropriate.
INTRODUCTION
Members of the genus Acinetobacter are ubiquitous organ-
isms of increasing interest and importance to clinical
microbiologists, as some are serious nosocomial pathogens
and show increasing resistance to many commonly pre-
scribed antibiotics (Bergogne-Berezin and Towner 1996).
This genus has a very confused taxonomic history but is now
placed in the c-Proteobacteria on 16S rDNA data; it is
considered to be phylogenetically coherent (Rainey et al.1994; Ibrahim et al. 1997) and can be delineated unequivo-
cally using the transformation assay of Juni (1972).
The species delineation of Acinetobacter still poses prob-
lems, even after Bouvet and Grimont (1986), Bouvet and
Jeanjean (1989) and Tjernberg and Ursing (1989) recognized
19 genomic species using DNA:DNA hybridization. Only
seven of these species have been validly named. Tjernberg and
Ursing (1989) showed that their DNA group 14 corresponded
to DNA group 13 of Bouvet and Jeanjean (1989), while their
DNA group 15 was novel and quite different to the BouvetCorrespondence to: Dr R. Seviour, Biotechnology Research Centre, La Trobe
University, Bendigo, Victoria 3552, Australia
(e-mail: [email protected]).
ã 2001 The Society for Applied Microbiology
Journal of Applied Microbiology 2001, 90, 309±319

and Jeanjean (1989) group 15. Tjernberg and Ursing (1989)
also proposed that their DNA group 13 was closely related to
the Bouvet and Grimont groups 1, 2 and 3, and they and
colleagues have proposed that all four should be amalgamated
into a single species, A. calcoaceticus. Tjernberg and Ursing
(1989) also suggested their DNA groups 10 and 11 were most
closely related to each other, as were groups 4 and 6 and
Bouvet and Jeanjean (1989) groups 15 and 16. However, such
interspeci®c relationships have not always been con®rmed
when other characterization methods, both phenotypic and
molecular, have been used (e.g. Nowak and Kur 1995;
Bergogne-Berezin and Towner 1996; Vila et al. 1996; Dijk-
shoorn et al. 1998; Jawad et al. 1998; Koeleman et al. 1998).
Certainly, 16S rDNA sequence comparisons (Rainey et al.1994) gave different interspecies relationships to some of
these (e.g. Rudant et al. 1999), although Yamamoto et al.(1999) obtained a better, if not complete, agreement with the
DNA groupings using gyr B DNA sequence comparisons. As
a consequence of the inconsistencies arising from the many
different typing methods used for Acinetobacter, reliable
methods for unequivocally identifying many members of this
genus are still not available. Furthermore, it is now clear that
more clinical genomic species than those currently recognized
from the collective work of Bouvet and Grimont (1986),
Bouvet and Jeanjean (1989) and Tjernberg and Ursing (1989)
exist (Dijkshoorn et al. 1998; Yamamoto et al. 1999).
Acinetobacters are readily isolated from activated sludge
systems removing phosphorus (Beacham et al. 1990; Mino
et al. 1998). There is some evidence that genomic species 5,
7 and 8/9 are more commonly seen in these environmental
samples (Wiedmann-Al-Ahmad et al. 1994; Knight et al.1995) than DNA groups 2 and 3 and Tjernberg and Ursing
(1989) group 13, which predominate in clinical species.
Identi®cation methods developed for the clinical isolates are
unlikely always to be applicable to environmental isolates
(Soddell et al. 1993). It is also likely that many novel isolates
of Acinetobacter await description from habitats other than
clinical specimens (Vaneechoutte et al. 1999), so it is
probable that we still understand little about the ecology
and true biodiversity of this genus (Dijkshoorn et al. 1998).
The importance of Acinetobacter in biological phosphate
removal in activated sludge systems, once widely accepted
(e.g. Jenkins and Tandoi 1991), has been questioned from
studies using 16S rRNA sequences as determinants of
biodiversity (Wagner et al. 1994; Bond et al. 1995, 1999;
Kampfer et al. 1996; Christensson et al. 1998).
The current study attempted to understand better the
biodiversity and ecology of Acinetobacter in four enhanced
biological phosphate removal (EBPR) activated sludge
systems, by clarifying their taxonomic status in relation to
the currently recognized DNA groups. The characterization
method chosen was RAPD-PCR, which has been success-
fully applied to epidemiological studies with bacteria, and in
resolving interspeci®c relationships in other genera (Pacheco
et al. 1996; Moschetti et al. 1998; Vickery et al. 1998). The
antibiotic resistance patterns of individual isolates was also
determined to see if these patterns differed in populations
from the different plants.
MATERIALS AND METHODS
Cultures of Acinetobacter used in study
Four different EBPR plants were sampled at regular (usually
fortnightly) intervals from April to December in 1995.
These plants were at Bendigo, Ballarat and Wodonga in
Victoria and Albury in NSW, Australia. Their plant
operational parameters are given in Table 1, and all were
removing phosphate during the study period. The methods
for isolating Acinetobacter spp. from biomass samples within
24 h of taking them were those described by Knight et al.(1995). All the Gram-negative, oxidase-negative coccobacilli
obtained were then screened using the Juni (1972) transfor-
mation assay for members of the genus Acinetobacter. The 52
isolates used in this study were selected randomly from the
total number obtained to re¯ect different sampling times.
All were stored at )80°C. Those from Albury, Bendigo,
Ballarat and Wodonga are pre®xed with A (11 strains), B (16
strains), C (12 strains) and D (13 strains), respectively. Also
included were the 19 recognized genomic species, kindly
supplied by Prof. Bouvet, Institut Pasteur, Paris (referred
to as BG strains) and Prof. Tjernberg, Lund University,
Sweden (TU strains). Their sources and culture collection
numbers are given in Table 2. All the environmental isolates
were screened for their ability to accumulate polyphosphate
as detailed by Beacham et al. (1992).
Phenotypic identi®cation of Acinetobacterisolates to species level
The Biolog GN system was used in attempts to characterize
and identify the isolates con®rmed as Acinetobacter spp.
Table 1 Operation parameters of the four activated sludge plants
used in study
Wastewater treatment plants
Albury Ballarat Wodonga Bendigo
P mg g)1 (MLSS) 24á34 38á11 14á03 31á26
P mg l)1 117á75 87á00 133á25 126á58
Typical MLSS mg l)1 4800 2300 9500 4100
MLVSS/MLSS 0á77 0á80 0á82 0á80
Sludge age (days) 16 (est) 3±5 18 25
P = Phosphorus; MLSS = Mixed Liquor Suspended Solids; MLVSS
= Mixed Liquor Volatile Suspended Solids.
310 E. CARR ET AL .
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 309±319

These isolates were also identi®ed using the phenotypic
identi®cation schemes described by Soddell et al. (1993).
Numerical analysis of the Biolog data was carried out
and dendrograms drawn based on SSM coef®cients and the
UPGMA algorithm using the NTSYS-pc software (Exeter
Software, New York, USA).
DNA extraction for RAPD-PCRof Acinetobacter isolates
All Acinetobacter isolates were grown in nutrient broth
(Oxoid) at 30°C for 48 h with shaking, and cells were
harvested by centrifugation. Pellets were resuspended in
TE buffer (10 mmol l)1 Tris; 1 mmol l)1 EDTA, pH 8),
and cell lysis occurred with addition of 30 ll 10% v/v
sodium dodecyl sulphate (SDS) plus 3 ll proteinase K
(20 mg ml)1, Sigma). Many of these isolates produced
substantial capsular material which seemed to interfere with
cell lysis and DNA extraction. Therefore, 100 ll 5 mol l)1
NaCl, and 80 ll 1% w/v cetyl trimethyl ammonium
bromide (CTAB) in 0á7 mol l)1 NaCl, were added, and the
mixture incubated at 65°C for 10 min before extraction with
phenol:chloroform:isoamyl alcohol (25:24:1). After centrif-
ugation, the aqueous phase was again extracted with
chloroform:isoamyl alcohol (24:1). Genomic DNA was then
precipitated with ice-cold ethanol; the DNA was collected
by centrifugation, washed in cold 70% (v/v) ethanol and air-
dried before being resuspended in TE buffer. Finally, 3 ll
RNAse (1 mg ml)1 Sigma) were added and the mixture
incubated at 37°C for 30 min. The DNA was again
precipitated with ethanol as above and the pellet air dried
and redissolved in TE buffer overnight. Purity of DNA was
checked by the A260:A280 nm ratio which, if greater than 1á8,
was taken to indicate that purity was acceptable. If below
1á8, the DNA solution was re-extracted with chloroform:iso-
amyl alcohol (24:1).
Genomic ®ngerprinting of Acinetobacterusing RAPD-PCR
Seven primers, all obtained from Bresatec (Adelaide,
Australia), were used in this study. Four were discarded as
when screened, the reproducibility of the band patterns
obtained with the known genomic species was poor between
replicate runs (data not presented). Primers 5¢GCTTGTGAAC 3¢ and 5¢ CGGCAGCGCC 3¢ gave
reproducible patterns but often, only a small numbers of
bands was generated with many of the strains (data not
presented). The one eventually chosen for this study was the
decamer 5¢ GCTTGTGAAC 3¢ because it gave the highest
number of bands with the known genomic species in
screening trials. The importance of standardizing genomic
®ngerprinting protocols has been convincingly demonstrated
by Koeleman et al. (1998), so the following method was
Table 2 Known genomic species of
Acinetobacter used in this studyAbbreviation used in study Species name Origin/Culture collection numbers
BG1 Acinetobacter calcoaceticus ATCC 23055T/CIP 81á08T
BG2 Acinetobacter baumannii ATCC 19606T/CIP 70á34T
BG3 Acinetobacter sp. 3 ATCC 19004/CIP 70á29
BG4 Acinetoabcter haemolyticus ATCC 17906T/CIP 64á3T
BG5 Acinetobacter junii ATCC 17908T/CIP 64á5T
BG6 Acinetobacter sp. 6 ATCC 17979/CIP A165
BG7 Acinetobacter johnsonii ATCC17909T/CIP 64á6T
BG8 Acinetobacter lwof®i NCTC 5866T/CIP 64á10T
BG9 Acinetobacter sp. 9 ATCC 9957/CIP 70á31
BG10 Acinetobacter sp. 10 ATCC 17924/CIP 70á12
BG11 Acinetobacter sp. 11 ATCC 11171/CIP 63á46
BG12 Acinetobacter radioresistens SEIP 12á81
BJ13 Acinetobacter sp. 13 (BJ) ATCC 17905/CIP 64á2BJ14 Acinetobacter sp. 14 (BJ) K.Irino 105/85
BJ15 Acinetobacter sp. 15 (BJ) M.M. Adam Ac606 180 : 40 va
BJ16 Acinetobacter sp. 16 (BJ) ATCC 17988/CIP 70á18
BJ17 Acinetobacter sp. 17 (BJ) SEIP Ac87á314
TU13 Acinetobacter sp. 13 (TU) ATCC 17903
TU14 Acinetobacter sp. 14 (TU) ATCC 151a1
TU15 Acinetobacter sp. 15 (TU) ATCC 71
ATCC = American Type Culture Collection, Maryland, USA; CIP = Collection de l'Institut
Pasteur, Paris; SEIP = Collection du Service des Enterobacteries, Institut Pasteur, Paris.
ACINETOBACTER FROM ACTIVATED SLUDGE 311
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 309±319

rigorously adhered to for all the analyses in this study. In
most cases, DNA that was extracted at least twice from each
strain almost always gave identical RAPD patterns, even
when analyses were carried out up to ®ve months apart. Any
occasional differences noticed in the patterns between runs
of the same strain were limited to presence or absence of one
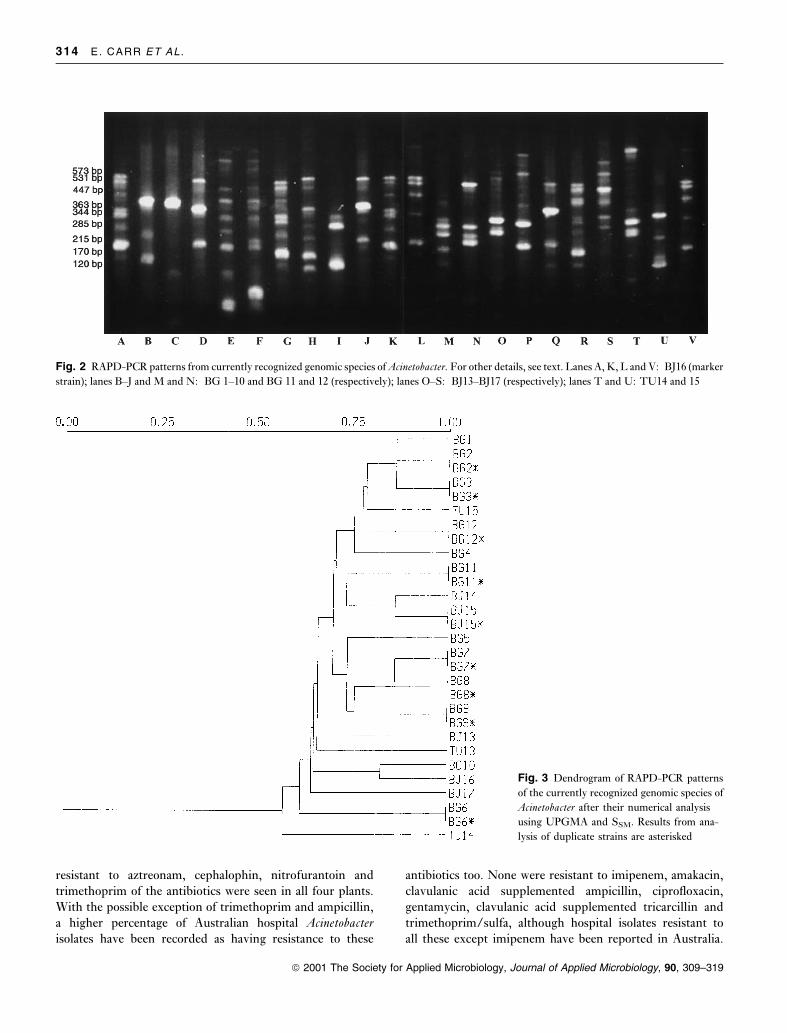
or two faint bands (e.g. as in lanes K and L in Fig. 2, which
are the poorest replicate patterns obtained in this study).
Some of the data obtained for replicate runs after numerical
analysis of the patterns for several strains are presented in
the dendrograms given in Figs 3 and 5.
PCR reaction mixes contained 2á5 ll of a 10´ reaction
buffer (200 mmol l)1 (NH4)2SO4; 750 mmol l)1 Tris HCl
buffer pH 9; 0á1% v/v Tween (80), 2á5 ll 2 mmol l)1
deoxynucleotide triphosphate mixture (Pharmacia), 3á5 ll
25 mmol l)1 MgCl2, 0á9 ll 20 lmol l)1 primer (Bresatec),
2 units Taq polymerase (Integrated Sciences, Melbourne)
and 100 ng DNA template from each Acinetobacter isolate.
DNA ampli®cation was achieved with a Hybaid Omnigene
thermocycler (Integrated Sciences, Melbourne, Australia)
and the following protocol. After initial denaturation at 94°C
for 3 min, followed by 34 cycles at 92°C for 30 s, 40°C for
60 s and 72°C for 3 min, a ®nal single cycle at 92°C for 30 s,
40°C for 60 s and 72°C for 10 min was performed.
Ampli®cation products were run on 1% agarose gels with
1´ TAE buffer at 90 V for 1á5 h, and bands visualized with
ethidium bromide. Gels were viewed and photographed
with a Polaroid/FotoDyne (FotoDyne, Hartland, Wiscon-
sin, USA) camera illuminator using Polaroid 665 ®lm.
Negatives of gel images were scanned into Adobe Photoshop
(Macintosh version 4á01), and the gel patterns analysed
using Gel Pro Analyser (Media Cybernetics, Silver Spring,
Maryland, USA), which could compensate for slightly
slanted gel runs. One Acinetobacter strain, BG 16, was run
on all gels as an internal marker, and all bands from other
isolates normalized against those of this isolate using GelPro
Analyser (Media Cybernetics, Silver Spring, Maryland,
USA). Numerical analyses and dendrogram constructions
were carried out as described earlier.
Antibiotic resistance patterns for activatedsludge Acinetobacter isolates
Antibiotic resistance for selected isolates was determined
using the Vitek AMS system (bioMerieux Vitek, Melbourne,
Australia) and the GNS-406 susceptibility card containing
the following antibiotics: amikacin, amoxicillin/clavulanic
acid, ampicillin, aztreonam, cefotaxime, ceftazidime, ceph-
alothin, cipro¯oxacin, gentamycin, imipenem, nitrofuran-
toin, nor¯oxacin, tricarcillin/clavulanic acid, tobramycin,
trimethaprim/sulfamethoxazole and trimethoprim. Resist-
ance or susceptibility was based on MIC values determined
for each antibiotic by Vitek (Vitek Technical Bulletin).
RESULTS
Phenotypic properties of the Acinetobacterisolates
All 52 strains used in this study were con®rmed as belonging
to the genus Acinetobacter with the Juni (1972) assay.
However, most could not be identi®ed to species level with
either the Biolog GN or the phenotypic characterization
schemes used earlier by Soddell et al. (1993). Of those
which were (<30%), most emerged as genomic species 7
(A. johnsonii), 11 and 15. Many which identi®ed as genomic
species BG15 with Biolog GN appeared as A. johnsonii when
the other schemes were used. Numerical analyses using the
95 Biolog characters for a selection of these strains and the
known genomic species produced a dendrogram (Fig. 1)
where few of the isolates from the four plants cluster closely
with the recognized genomic species of Bouvet and Grimont
(1986), Bouvet and Jeanjean (1989) or Tjernberg and Ursing
(1989). Isolates from individual plants were usually scattered
widely throughout the dendrogram, although some strains
from a single plant (e.g. C2, C5, C6 and C7 from Ballarat,
and D2, D5, D6, D7, D12 and D13 from Wodonga), and
occasionally different plants (e.g. A6, A15 and B9, and B17,
D4 and D11), were very closely related to each other on
their substrate utilization patterns. Less than 50% of these
isolates (23/52) could store polyphosphate when examined
in pure culture with the methylene blue and Neisser stains,
and these did not belong to any particular genomic species
where identi®cation was possible.
PCR ®ngerprinting of known genomic speciesof Acinetobacter
The patterns obtained with the primer 5¢ GCTTGTGAAC
3¢ after RAPD-PCR of the known genomic species are
shown in Fig. 2. Most yielded a distinctive RAPD ®nger-
print with the primer used, and the relationships between
them following numerical analysis of these patterns are
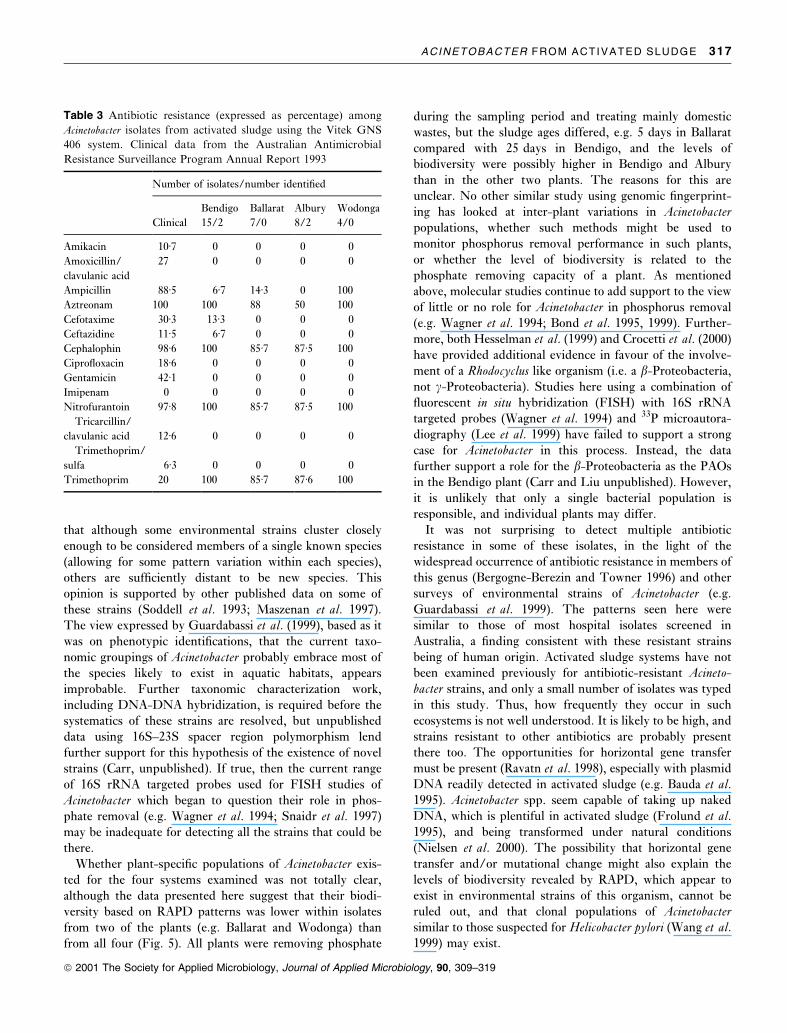
shown in the dendrogram (Fig. 3). Brie¯y, all strains cluster
at an SSM of 0á55. Several distinct clusters emerged and
among these, isolates BG1, BG2 and BG3, BG7, BG8 and
BG9, and BJ14 and BJ15, clustered most closely together.
Only one strain of each of these known genomic species was
examined here.
RAPD-PCR of activated sludge Acinetobacterisolates
Examples of the patterns obtained from the Acinetobacterisolates from the four different plants are given in Fig. 4 and
the dendrogram constructed after numerical analysis of
these, and including the known genomic species, is presen-
312 E. CARR ET AL .
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 309±319

ted in Fig. 5. They show that generally, the environmental
strains produced patterns quite different to those of the
known genomic species and thus, did not cluster closely
with them. There were a few exceptions (see below).
In general, isolates from each particular EBPR plant
tended to cluster together, as shown with those from Ballarat
(C isolates) which form a single group except for C8 and
C12, the latter being most closely related to some Bendigo
isolates. The Albury (A) strains, like the Bendigo (B)
isolates, fall into one quite tight cluster or are distributed
throughout the dendrogram. Strains from Wodonga (D) fall
into three main clusters that are scattered throughout the
dendrogram, while some isolates e.g. D4, D11 and D13
appear to be identical to each other, even though obtained
from the plant at different times of the year. Only in a very
few cases do these activated sludge isolates form clusters at
high SSM values with the known genomic species. In some
cases, these groupings support those obtained after analysis
of the Biolog characterization data (Fig. 1), as in the case of
most of the isolates from Ballarat (C) and Wodonga (D
isolates), which clustered together in both.
Antibiotic resistance patterns of activatedsludge isolates of Acinetobacter
The antibiotic resistance pattern results determined by the
Vitek system are given in Table 3. Not all the isolates could
be tested with this system because many, although not
identi®ed as A. johnsonii with Biolog, failed to grow at 37°C,
the temperature at which this system operates. With the
possible exception of ampicillin and ceftazidine, very similar
resistance patterns were obtained for isolates from the four
plants, although the percentages of isolates resistant to each
of the individual antibiotics did vary. Thus, only isolates
Fig. 1 Dendrogram of Acinetobacter spp. after
numerical analysis with UPGMA and SSM of
data obtained from characterization of strains
using Biolog
ACINETOBACTER FROM ACTIVATED SLUDGE 313
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 309±319
resistant to aztreonam, cephalophin, nitrofurantoin and
trimethoprim of the antibiotics were seen in all four plants.
With the possible exception of trimethoprim and ampicillin,
a higher percentage of Australian hospital Acinetobacterisolates have been recorded as having resistance to these
antibiotics too. None were resistant to imipenem, amakacin,
clavulanic acid supplemented ampicillin, cipro¯oxacin,
gentamycin, clavulanic acid supplemented tricarcillin and
trimethoprim/sulfa, although hospital isolates resistant to
all these except imipenem have been reported in Australia.
Fig. 2 RAPD-PCR patterns from currently recognized genomic species of Acinetobacter. For other details, see text. Lanes A, K, L and V: BJ16 (marker
strain); lanes B±J and M and N: BG 1±10 and BG 11 and 12 (respectively); lanes O±S: BJ13±BJ17 (respectively); lanes T and U: TU14 and 15
Fig. 3 Dendrogram of RAPD-PCR patterns
of the currently recognized genomic species of
Acinetobacter after their numerical analysis
using UPGMA and SSM. Results from ana-
lysis of duplicate strains are asterisked
314 E. CARR ET AL .
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 309±319

None of the Acinetobacter isolates screened here were
identi®ed as members of the A. calcoaceticus±A. baumanniicomplex, i.e., those normally associated with clinical spec-
imens.
DISCUSSION
This study has investigated the biodiversity of Acinetobacterstrains from four activated sludge plants of different
con®gurations, but all of which were removing phosphorus.
It has determined, on the RAPD-PCR criteria used here,
that strains different to the described genomic species from
clinical habitats exist in activated sludge, an ecosystem long
known to contain large numbers of culturable Acinetobacterstrains (e.g. Knight et al. 1995). This supports earlier work
based on phenotypic (Soddell et al. 1993) and soluble
protein PAGE (Maszenan et al. 1997) characterization. Even
isolates obtained from clinical specimens have often fallen
outside the currently accepted taxonomic framework for this
genus (e.g. Gerner-Smidt and Frederiksen 1993).
Many different methods have been applied in attempts
to sort out the confused interspecies relationships within
Acinetobacter, but none give results which agree completely
with the relationships suggested from the DNA-DNA
hybridization data (Bouvet and Grimont 1986; Bouvet and
Jeanjean 1989; Tjernberg and Ursing 1989). The character-
ization method selected for the isolates in this study was
RAPD-PCR, which has been used successfully elsewhere to
resolve intrageneric relationships in bacteria, including
acinetobacters from activated sludge (Wiedmann-Al-Ahmad
et al. 1994). In some, if not all (e.g. Tynkkynen et al. 1999)
comparative studies, RAPD-PCR has proved to be as
discriminatory in achieving this with both prokaryotes and
eukaryotes as several of the other genomic ®ngerprinting
techniques available (Koeleman et al. 1998; Arenal et al.1999).
Results obtained here showed that PCR-RAPD could
differentiate between all the currently recognized genomic
species of Acinetobacter. Although BG8 and BG9 could not
be separated either phenotypically by Bouvet and Grimont
(1986) or using other criteria (e.g. Tjernberg and Ursing
1989; Vaneechoutte et al. 1995), they were distinguished
readily here on their RAPD patterns (Fig. 2). An ability to
separate BG8 and BG9 was reported by Weidmann-
Al-Ahmad et al. (1994) although in their RAPD study,
BG7 and BG9 gave the same RAPD patterns, as they did
also in the study by Nowak and Kur (1995) with REP-PCR.
Numerical analysis of the RAPD patterns from BG1, BG2,
BG3 and TU13 (Fig. 3) supports the close relationship
between BG1, BG2 and BG3 (but not TU13) proposed
earlier from other methods of characterization (Gerner-
Smidt 1992; Gerner-Smidt and Tjernberg 1993). Genomic
species 10 and 11, also considered very similar (Tjernberg
and Ursing 1989; Bernards et al. 1995; Ibrahim et al. 1997;
Maszenan et al. 1997; Vaneechoutte et al. 1999), gave easily
recognizable different RAPD patterns (Fig. 2). Other
genomic ®ngerprinting techniques (Wiedmann-Al-Ahmad
et al. 1994; Nowak and Kur 1995; Janssen and Dijkshoorn
1996) could also separate them.
The reasons for these variations in interspecies relation-
ships with different typing methods are not clear, but the
patterns obtained here were highly reproducible. It is
recognized that only a single primer for PCR-RAPD was
used and only one strain of each genomic species was
analysed in this study. Clearly, more strains of each of these
known genomic species should be analysed but as detailed
Fig. 4 RAPD-PCR patterns of representative strains of Acinetobacter obtained from activated sludge systems. Lanes A, F, K, L, P and U: BJ16
(marker strain); lanes B, C, D, E and G: C2, C4, C6, C8 and C10; lanes H, I and J: D4, D8 and D10; lanes M and N: A29 and A34; lanes O, Q, R, S
and T:B6, B10, B11, B17 and B19
ACINETOBACTER FROM ACTIVATED SLUDGE 315
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 309±319

above, the literature suggests that disagreements like those
noted here are not uncommon within Acinetobacter charac-
terization studies.
Even allowing for the fact that only single isolates of each
of the known genomic species of Acinetobacter were used
here, it is thought that the main ®nding of this study still
holds. With a few exceptions (e.g. D10 and BJ15), the
activated sludge strains analysed here did not cluster closely
with them. Instead, these usually clustered quite separately,
as seen with cluster BG5, BG7, BG8, BG9, BJ13 and BJ14,
and cluster BG1, BG2, BG3, BG4, and BG12. (Fig. 5). Of
the 30% of strains which were identi®ed phenotypically
here, most emerged as BG7, BG11 and BJ15, and the former
is thought to dominate such communities of Acinetobacter(Kampfer et al. 1992; Knight et al. 1995; Guardabassi et al.1999). However, none of the activated sludge isolates
analysed here clustered closely with the BG7 type strain
(Fig. 5), so, with a few exceptions, it was not possible to
identify con®dently any of the unknown strains by compar-
ing their RAPD patterns with those of the known genomic
species. This technique can distinguish between different
strains of a single species with some bacteria (e.g. Soto et al.1999), although whether that is the case with Acinetobacterspp. is not known. The dendrogram (Fig. 5) would suggest
Fig. 5 Dendrogram of RAPD-PCR patterns
of all Acinetobacter isolates examined by
RAPD-PCR after their numerical analysis
using UPGMA and SSM. Results from ana-
lysis of duplicate strains are asterisked
316 E. CARR ET AL .
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 309±319
that although some environmental strains cluster closely
enough to be considered members of a single known species
(allowing for some pattern variation within each species),
others are suf®ciently distant to be new species. This
opinion is supported by other published data on some of
these strains (Soddell et al. 1993; Maszenan et al. 1997).
The view expressed by Guardabassi et al. (1999), based as it
was on phenotypic identi®cations, that the current taxo-
nomic groupings of Acinetobacter probably embrace most of
the species likely to exist in aquatic habitats, appears
improbable. Further taxonomic characterization work,
including DNA-DNA hybridization, is required before the
systematics of these strains are resolved, but unpublished
data using 16S±23S spacer region polymorphism lend
further support for this hypothesis of the existence of novel
strains (Carr, unpublished). If true, then the current range
of 16S rRNA targeted probes used for FISH studies of
Acinetobacter which began to question their role in phos-
phate removal (e.g. Wagner et al. 1994; Snaidr et al. 1997)
may be inadequate for detecting all the strains that could be
there.
Whether plant-speci®c populations of Acinetobacter exis-
ted for the four systems examined was not totally clear,
although the data presented here suggest that their biodi-
versity based on RAPD patterns was lower within isolates
from two of the plants (e.g. Ballarat and Wodonga) than
from all four (Fig. 5). All plants were removing phosphate
during the sampling period and treating mainly domestic
wastes, but the sludge ages differed, e.g. 5 days in Ballarat
compared with 25 days in Bendigo, and the levels of
biodiversity were possibly higher in Bendigo and Albury
than in the other two plants. The reasons for this are
unclear. No other similar study using genomic ®ngerprint-
ing has looked at inter-plant variations in Acinetobacterpopulations, whether such methods might be used to
monitor phosphorus removal performance in such plants,
or whether the level of biodiversity is related to the
phosphate removing capacity of a plant. As mentioned
above, molecular studies continue to add support to the view
of little or no role for Acinetobacter in phosphorus removal
(e.g. Wagner et al. 1994; Bond et al. 1995, 1999). Further-
more, both Hesselman et al. (1999) and Crocetti et al. (2000)
have provided additional evidence in favour of the involve-
ment of a Rhodocyclus like organism (i.e. a b-Proteobacteria,
not c-Proteobacteria). Studies here using a combination of
¯uorescent in situ hybridization (FISH) with 16S rRNA
targeted probes (Wagner et al. 1994) and 33P microautora-
diography (Lee et al. 1999) have failed to support a strong
case for Acinetobacter in this process. Instead, the data
further support a role for the b-Proteobacteria as the PAOs
in the Bendigo plant (Carr and Liu unpublished). However,
it is unlikely that only a single bacterial population is
responsible, and individual plants may differ.
It was not surprising to detect multiple antibiotic
resistance in some of these isolates, in the light of the
widespread occurrence of antibiotic resistance in members of
this genus (Bergogne-Berezin and Towner 1996) and other
surveys of environmental strains of Acinetobacter (e.g.
Guardabassi et al. 1999). The patterns seen here were
similar to those of most hospital isolates screened in
Australia, a ®nding consistent with these resistant strains
being of human origin. Activated sludge systems have not
been examined previously for antibiotic-resistant Acineto-bacter strains, and only a small number of isolates was typed
in this study. Thus, how frequently they occur in such
ecosystems is not well understood. It is likely to be high, and
strains resistant to other antibiotics are probably present
there too. The opportunities for horizontal gene transfer
must be present (Ravatn et al. 1998), especially with plasmid
DNA readily detected in activated sludge (e.g. Bauda et al.1995). Acinetobacter spp. seem capable of taking up naked
DNA, which is plentiful in activated sludge (Frolund et al.1995), and being transformed under natural conditions
(Nielsen et al. 2000). The possibility that horizontal gene
transfer and/or mutational change might also explain the
levels of biodiversity revealed by RAPD, which appear to
exist in environmental strains of this organism, cannot be
ruled out, and that clonal populations of Acinetobactersimilar to those suspected for Helicobacter pylori (Wang et al.1999) may exist.
Table 3 Antibiotic resistance (expressed as percentage) among
Acinetobacter isolates from activated sludge using the Vitek GNS
406 system. Clinical data from the Australian Antimicrobial
Resistance Surveillance Program Annual Report 1993
Number of isolates/number identi®ed
Bendigo Ballarat Albury Wodonga
Clinical 15/2 7/0 8/2 4/0
Amikacin 10á7 0 0 0 0
Amoxicillin/ 27 0 0 0 0
clavulanic acid
Ampicillin 88á5 6á7 14á3 0 100
Aztreonam 100 100 88 50 100
Cefotaxime 30á3 13á3 0 0 0
Ceftazidine 11á5 6á7 0 0 0
Cephalophin 98á6 100 85á7 87á5 100
Cipro¯oxacin 18á6 0 0 0 0
Gentamicin 42á1 0 0 0 0
Imipenam 0 0 0 0 0
Nitrofurantoin 97á8 100 85á7 87á5 100
Tricarcillin/
clavulanic acid 12á6 0 0 0 0
Trimethoprim/
sulfa 6á3 0 0 0 0
Trimethoprim 20 100 85á7 87á6 100
ACINETOBACTER FROM ACTIVATED SLUDGE 317
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 309±319

REFERENCES
Arenal, F., Platas, G., Martin, J., Salazar, O. and Pelaez, F. (1999)
Evaluation of different PCR-based DNA ®ngerprinting techniques
for assessing the genetic variability of isolates of the fungus
Epicoccum nigrum. Journal of Applied Microbiology 87, 898±906.
Bauda, P., Lalhement, C. and Manem, J. (1995) Plasmid content
evaluation of activated sludge. Water Research 29, 371±374.
Beacham, A.M., Seviour, R.J. and Lindrea, K.C. (1992) Polyphosphate
accumulating abilities of Acinetobacter isolates obtained from a
biological nutrient removal pilot plant. Water Research 26, 121±122.
Beacham, A.M., Seviour, R.J., Lindrea, K.C. and Livingston, I. (1990)
Genospecies diversity of Acinetobacter isolates obtained from a
biological nutrient removal pilot plant of a modi®ed UCT
con®guration. Water Research 24, 23±29.
Bergogne-Berezin, E. and Towner, K.J. (1996) Acinetobacter spp. as
nosocomial pathogens: microbiological, clinical and epidemiological
features. Clinical Microbiology Reviews 9, 148±165.
Bernards, A.T., Dijkshoorn, L., Van Der Toorn, J., Bochner, B.R. and
Boven, C.P.A. (1995) Phenotypic characterization of Acinetobacter
strains of 13 DNA-DNA hybridization groups by means of the
Biolog system. Journal of Medical Microbiology 42, 113±119.
Bond, P., Hugenholtz, P., Keller, J. and Blackall, L. (1995) Bacterial
community structures of phosphate-removing and non-phosphate-
removing activated sludges from sequencing batch reactors. Applied
and Environmental Microbiology 61, 1910±1916.
Bond, P.L., Erhart, R., Wagner, M., Keller, J. and Blackall, L.L. (1999)
Identi®cation of some of the major groups of bacteria in ef®cient and
nonef®cient biological phosphorus removal activated sludge systems.
Applied and Environmental Microbiology 65, 4077±4084.
Bouvet, P.J.M. and Grimont, P.A.D. (1986) Taxonomy of the genus
Acinetobacter with the recognition of Acinetobacter baumannii sp.
nov., Acinetobacter haemolyticus sp. nov., Acinetobacter johnsonnii sp.
nov. & Acinetobacter junii sp. nov. & emended description of
Acinetobacter calcoaceticus and Acinetobacter lwof®i. International
Journal of Systematic Bacteriology 36, 228±240.
Bouvet, P.J.M. and Jeanjean, S. (1989) Delineation of a new proteolytic
species in the genus Acinetobacter. Research Microbiology 140,
291±299.
Christensson, M., Blackall, L.L. and Welander, T. (1998) Metabolic
transformations and characterization of the sludge community in an
enhanced biological phosphorus removal system. Applied Microbiol-
ogy and Biotechnology 49, 222±234.
Crocetti, G.R., Hugenholtz, P., Bond, P.L. et al. (2000) Identi®cation
of polyphosphate-accumulating organisms and design of 16S rRNA-
directed probes for their detection and quantitation. Applied and
Environmental Microbiology 66, 1175±1182.
Dijkshoorn, L., Van Harsselaar, B., Tjernberg, I., Bouvet, P.J.M. and
Vaneechoutte, M. (1998) Evaluation of ampli®ed ribosomal DNA
restriction analysis for identi®cation of Acinetobacter genomic
species. Systematic and Applied Microbiology 21, 33±39.
Frolund, B., Griebe, T. and Nielsen, P.H. (1995) Enzymatic activity in
the activated-sludge ¯oc matrix. Applied Microbiology and Biotech-
nology. 43, 755±761.
Gerner-Smidt, P. (1992) Ribotyping of the Acinetobacter calcoaceticus±
Acinetobacter baumannii complex. Journal of Clinical Microbiology 30,
2680±2685.
Gerner-Smidt, P. and Frederiksen, W. (1993) Acinetobacter in Den-
mark: I. Taxonomy, antiobiotic susceptibility and pathogenicity of
112 clinical strains. APMIS 101, 815±825.
Gerner-Smidt, P. and Tjernberg, I. (1993) Acinetobacter in Denmark:
II. Molecular studies of the Acinetobacter calcoaceticus±Acinetobacter
baumannii complex. APMIS 101, 826±832.
Guardabassi, L., Dalsgaard, A. and Olsen, J.E. (1999) Phenotypic
characterization and antibiotic resistance of Acinetobacter spp.
isolated from aquatic sources. Journal of Applied Microbiology 87,
659±667.
Hesselmann, R.P.X., Werlen, C., Hahn, D., Van Der Meer, J.R. and
Zehnder, A.J.B. (1999) Enrichment, phylogenetic analysis and
detection of a bacterium that performs enhanced biological phos-
phate removal in activated sludge. Systematic and Applied Micro-
biology 22, 454±465.
Ibrahim, A., Gerner-Smidt, P. and Liesack, W. (1997) Phylogenetic
relationships of genus Acinetobacter. FEMS Microbiology Letters 124,
349±354.
Janssen, P. and Dijkshoorn, L. (1996) High resolution DNA ®nger-
printing of Acinetobacter outbreak strains. FEMS Microbiology
Letters 142, 194±194.
Jawad, A., Snelling, A.M., Heritage, J. and Hawkey, P.M. (1998)
Comparison of ARDRA and recA-RFLP analysis for genomic
species identi®cation of Acinetobacter spp. FEMS Microbiology
Letters 165, 357±362.
Jenkins, D. and Tandoi, V. (1991) The applied microbiology of
enhanced biological phosphate removalÐaccomplishments and
needs. Water Research 25, 1471±1478.
Juni, E. (1972) Interspecies transformation of Acinetobacter: genetic
evidence for a ubiquitous genus. Journal of Bacteriology 112, 917±931.
Kampfer, P., Bark, K., Busse, H.J., Auling, G. and Dott, W. (1992)
Numerical and chemotaxonomy of polyphosphate accumulating
Acinetobacter strains with high polyphosphate:AMP phosphotrans-
ferase (PPAT) activity. Systematic and Applied Microbiology 15,
409±419.
Kampfer, P., Erhart, R., Beimfohr, J., Wagner, M. and Amann, R.
(1996) Characterization of bacterial communities from activated
sludge: culture-dependant numerical identi®cation versus in situ
identi®cation using group- and genus-speci®c rRNA-targeted
oligonucleotide probes. Microbial Ecology 32, 101±121.
Knight, G.C., Seviour, E.M., Seviour, R.J. et al. (1995) Development
of the microbial community of a full-scale biological nutrient
removal activated sludge plant during start-up. Water Research 29,
2085±2093.
Koeleman, J.G.M., Stoof, J., Biesmans, D.J., Savelkoul, P.H.M. and
Vandenbroucke-Grauls, C.M.J.E. (1998) Comparison of ampli®ed
ribosomal DNA restriction analysis, random ampli®ed polymorphic
DNA analysis and ampli®ed fragment length polymorphism ®nger-
printing for identi®cation of Acinetobacter genomic species and
typing of Acinetobacter baumannii. Journal of Clinical Microbiology 36,
2522±2529.
Lee, N., Nielsen, P.H., Andreasen, K.H. et al. (1999) Combination of
¯uorescent in situ hybridization and microautoradiographyÐa new
tool for structure-function analyses in microbial ecology. Applied and
Environmental Microbiology 65, 1289±1297.
Maszenan, A.M., Seviour, R.J., McDougall, B.M. and Soddell, J.A.
(1997) Diversity of isolates of Acinetobacter from activated sludge
318 E. CARR ET AL .
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 309±319

systems based on their whole cell protein patterns. Journal of
Industrial Microbiology and Biotechnology 18, 267±271.
Mino, T., Van Loosdrecht, M.C.M. and Heijnen, J.J. (1998)
Microbiology and biochemistry of the enhanced biological phosphate
removal process. Water Research 32, 3193±3207.
Moschetti, G., Blaiotta, G., Aponte, M. et al. (1998) Random ampli®ed
polymorphic DNA and ampli®ed ribosomal DNA spacer poly-
morphism: powerful methods to differentiate Streptococcus thermo-
philus strains. Journal of Applied Microbiology 85, 25±36.
Nielsen, K.M., Smalla, K. and van Elsas, J.D. (2000) Natural transfor-
mation of Acinetobacter sp. strain BD413 with cell lysates of
Acinetobacter sp., Pseudomonas ¯uorescens and Burkholderia cepacia in
soil microcosms. Applied and Environmental Microbiology 66, 206±212.
Nowak, A. and Kur, J. (1995) Genomic species typing of Acineto-
bacters by polymerase chain reaction ampli®cation of the recA gene.
FEMS Microbiology Letters 130, 327±332.
Pacheco, A.B.F., Guth, B.E.C., de Almeida, D.F. and Ferreira, L.C.S.
(1996) Characterization of enterotoxigenic Escherichia coli by random
ampli®cation of polymorphic DNA. Research Microbiology 147,
175±182.
Rainey, F.A., Lang, E. and Stackebrandt, E. (1994) The phylogenetic
structure of the twenty-one DNA groups of the genus Acinetobacter
as revealed by 16S ribosomal DNA sequence analysis. International
Journal of Systematic Bacteriology 47, 837±841.
Ravatn, R., Zehnder, A.J.B. and Van der Meer, J.R. (1998) Low-
frequency horizontal transfer of an element containing the chloro-
catechol degradation genes from Pseudomonas sp. strain B13 to
Pseudomonas putida F1 and to indigenous bacteria in laboratory-scale
activated-sludge microcosms. Applied and Environmental Micro-
biology 64, 2126±2132.
Rudant, E., Bouvet, P., Courvalin, P. and Lambert, T. (1999)
Phylogenetic analysis of proteolytic Acinetobacter strains based on
the sequence of genes encoding aminoglycoside 6¢-N- acetyltrans-
ferases. Systematic and Applied Microbiology 22, 59±67.
Snaidr, J., Amann, R., Huber, I., Ludwig, W. and Schleifer, K. (1997)
Phylogenetic analysis and in situ identi®cation of bacteria in
activated sludge. Applied and Environmental Microbiology 63, 2884±
2896.
Soddell, J.A., Beacham, A.M. and Seviour, R.J. (1993) Phenotypic
identi®cation of non-clinical isolates of Acinetobacter species. Journal
of Applied Bacteriology 74, 210±214.
Soto, S.M., Guerra, B., Gonzalez-Hevia, M.A. and Mendoza, M.C.
(1999) Potential of three-way randomly ampli®ed polymorphic DNA
analysis as a typing method for twelve Salmonella serotypes. Applied
and Environmental Microbiology 65, 4830±4836.
Tjernberg, I. and Ursing, J. (1989) Clinical strains of Acinetobacter
classi®ed by DNA-DNA hybridisation. APMIS 97, 596±605.
Tynkkynen, S., Satokari, R., Saarela, M., Mattila-Sandholm, T. and
Saxelin, M. (1999) Comparison of ribotyping, randomly ampli®ed
polymorphic DNA analysis and pulsed-®eld gel electrophoresis in
typing of Lactobacillus rhamnosus and L. casei strains. Applied and
Environmental Microbiology 65, 3908±3914.
Vaneechoutte, M., Dijkshoorn, L., Tjernberg, I. et al. (1995) Identi-
®cation of Acinetobacter genomic species by ampli®ed ribosomal
DNA restriction analysis. Journal of Clinical Microbiology 33, 11±15.
Vaneechoutte, M., Tjernberg, I., Baldi, F. et al. (1999) Oil-degrading
Acinetobacter strain RAG-1 and strains described as Acinetobacter
venetianus sp. nov. belong to the same genomic species. Research
Microbiology 150, 69±73.
Vickery, M., Smith, A., DePaola, A., Jones, D., Steffan, R. and Bej, A.
(1998) Optimization of the arbitraily-primed polymerase chain
reaction (AP-PCR) from intra-species differentiation of Vibrio
vulni®cus. Journal of Microbiological Methods 33, 181±189.
Vila, J., Marcos, M.A. and Jimenez De Anta, M.T. (1996) A
comparative study of different PCR-based DNA ®ngerprinting
techniques for typing of the Acinetobacter calcoaceticus±A. baumannii
complex. Journal of Medical Microbiology 44, 482±489.
Wagner, M., Erhart, R., Manz, W. et al. (1994) Development of an
rRNA-targeted oligonucleotide probe speci®c for the genus Acine-
tobacter and its application for in situ monitoring in activated sludge.
Applied and Environmental Microbiology 60, 792±800.
Wang, G., Humayun, M.Z. and Taylor, D.E. (1999) Mutation as an
origin of genetic variability in Helicobacter pylori. Trends in
Microbiology 7, 488±493.
Wiedmann-Al-Ahmad, M., Tichy, H.V. and Schon, G. (1994)
Characterization of Acinetobacter type strains and isolates obtained
from wastewater treatment plants by PCR ®ngerprinting. Applied
and Environmental Microbiology 60, 4066±4071.
Yamamoto, S., Bouvet, P.J.M. and Harayama, S. (1999) Phylogenetic
structures of the genus Acinetobacter based on gyrB sequences:
comparison with the grouping with DNA-DNA hybridization.
International Journal of Systematic Bacteriology 49, 87±95.
ACINETOBACTER FROM ACTIVATED SLUDGE 319
ã 2001 The Society for Applied Microbiology, Journal of Applied Microbiology, 90, 309±319
Related Documents











