Nephrol Dial Transplant (2008) 1 of 12 doi: 10.1093/ndt/gfn570 Original Article Rab 23 is expressed in the glomerulus and plays a role in the development of focal segmental glomerulosclerosis Tzu-Hao Huang 1 , Hao-Ai Shui 1 , Shuk-Man Ka 2 , Bor-Luen Tang 3 , Tai-Kuang Chao 2 , Jin-Shuen Chen 4 , Yuh-Feng Lin 4 and Ann Chen 2 1 Graduate Institute of Medical Sciences, 2 Department of Pathology, Tri-Service General Hospital, National Defense Medical Center, Taipei, Taiwan, ROC, 3 Department of Biochemistry, Yong Loo Lin School of Medicine, National University of Singapore, 8 Medical Drive, Singapore 117597 and 4 Division of Nephrology, Department of Internal Medicine, Tri-Service General Hospital, National Defense Medical Center, Taipei, Taiwan, ROC Abstract Background. Rab23, a member of the Rab family of small GTPase, has a function in antagonizing sonic hedge- hog signal transduction. Both Rab-family and hedgehog- related proteins are involved in sclerosis and fibrosis in certain pathological states, but their roles in focal segmen- tal glomerulosclerosis (FSGS) remain unclear. Methods. The FSGS model was established in Balb/c mice by a single injection of adriamycin. Serum, urine and mice kidneys were collected on Days 0, 7, 15 and 20. Western blot analysis was performed to detect the levels of Rab23 in the samples. Immunohistochemistry was used to examine the expressional profiles of Rab23 in kidneys. The expres- sions of transcripts of Rab23, extracellular matrix (ECM) proteins, and various hedgehog signalling pathway genes in kidneys or mesangial cells were evaluated by real-time RT- PCR. The effect of Rab23 on ECM protein expressions was evaluated by the knockdown or overexpression of Rab23 in mesangial cells. Results. Our results show that elevations of Rab23 were observed in the urine, but not in the serum, of the FSGS mice. Rab23 and hedgehog signalling pathway genes were constitutively expressed in normal kidneys and were sig- nificantly up-regulated in the kidneys of FSGS mice. The basal expression of Rab23 was identified in glomeruli, and mesangial cells displayed obvious elevation of Rab23 in the FSGS state. The knockdown or overexpression of Rab23 af- fected the collagen expression in cultured mesangial cells. Conclusions. An autocrine loop of hedgehog signalling could be activated in mesangial cells in the FSGS state, and Rab23 may be elevated to suppress hedgehog signalling and/or influence collagen synthesis. Importantly, Rab23 Correspondence and offprint requests to: Ann Chen, Department of Pathology, Tri-Service General Hospital, National Defense Medical Center, No. 325, Sec. 2, Cheng-Gung Road, Taipei, Taiwan, ROC. Tel: +886-2-8792-7008; Fax: +886-2-8792-7009; E-mail: doc31717@ ndmctsgh.edu.tw and Dr. Hao-Ai Shui, Graduate Institute of Medical Sci- ences, National Defense Medical Center, 161, Min-Chuan East Road, 6th Section, Taipei, Taiwan, ROC. E-mail: [email protected] could serve as a biomarker that indicates the severity of FSGS. Keywords: biomarker; focal segmental glomerulosclerosis; hedgehog signal transduction; mesangial cells; Rab23 Introduction Focal segmental glomerulosclerosis (FSGS) is a form of chronic nephropathy characterized by scattered sclerosis of glomeruli in which only a segment of the capillary is af- fected [1,2]. The molecules involved in FSGS pathogenesis are being gradually uncovered. These include proteins that cause oxidative stress, podocyte damage, hyperfiltration and sclerosis, which lead to marked proteinuria and scat- tered glomerular sclerosis seen in the disease [2,3]. Occa- sionally, some proteins that were originally not considered to play any role in FSGS were demonstrated to be involved in the pathogenesis of the disease. For example, osteopon- tin, a matrixcellular protein with a chemoattractive property for monocytes, has been shown to play a role in epithelial hyperplasia lesion in FSGS [4]. Recently, in our proteomic profiling of FSGS, we have identified Rab23 in the urine of an FSGS mouse model (unpublished data). However, the changes in expressional profiles of Rab23 and its associated genes in FSGS remain unknown. Rab23 is a member of the large Rab family of small GT- Pase, which has about 60 members that play varying roles in the intracellular traffic of proteins and membrane vesicles [5–7]. The expression and function of Rab23 in kidneys are unclear. In contrast to Rab23, many other members of the Rab family have been identified in the kidney. Rabs 4, 5, 7, 11, 13, 18, 20, 25, 34 and 38 are located in tubular cells [8–11], while Rab 3A is located in podocytes [12]. Rab11 and Rab20 co-localize with vacuolar H + -ATPase in tubu- lar and intercalated cells and play roles in regulating pro- ton secretion by transferring the H + -ATPase to the plasma C The Author [2008]. Published by Oxford University Press on behalf of ERA-EDTA. All rights reserved. For Permissions, please e-mail: [email protected] NDT Advance Access published October 16, 2008 by guest on July 12, 2015 http://ndt.oxfordjournals.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Nephrol Dial Transplant (2008) 1 of 12doi: 10.1093/ndt/gfn570
Original Article
Rab 23 is expressed in the glomerulus and plays a rolein the development of focal segmental glomerulosclerosis
Tzu-Hao Huang1, Hao-Ai Shui1, Shuk-Man Ka2, Bor-Luen Tang3, Tai-Kuang Chao2, Jin-Shuen Chen4,Yuh-Feng Lin4 and Ann Chen2
1Graduate Institute of Medical Sciences, 2Department of Pathology, Tri-Service General Hospital, National Defense Medical Center,Taipei, Taiwan, ROC, 3Department of Biochemistry, Yong Loo Lin School of Medicine, National University of Singapore, 8 MedicalDrive, Singapore 117597 and 4Division of Nephrology, Department of Internal Medicine, Tri-Service General Hospital, NationalDefense Medical Center, Taipei, Taiwan, ROC
AbstractBackground. Rab23, a member of the Rab family ofsmall GTPase, has a function in antagonizing sonic hedge-hog signal transduction. Both Rab-family and hedgehog-related proteins are involved in sclerosis and fibrosis incertain pathological states, but their roles in focal segmen-tal glomerulosclerosis (FSGS) remain unclear.Methods. The FSGS model was established in Balb/c miceby a single injection of adriamycin. Serum, urine and micekidneys were collected on Days 0, 7, 15 and 20. Westernblot analysis was performed to detect the levels of Rab23 inthe samples. Immunohistochemistry was used to examinethe expressional profiles of Rab23 in kidneys. The expres-sions of transcripts of Rab23, extracellular matrix (ECM)proteins, and various hedgehog signalling pathway genes inkidneys or mesangial cells were evaluated by real-time RT-PCR. The effect of Rab23 on ECM protein expressions wasevaluated by the knockdown or overexpression of Rab23 inmesangial cells.Results. Our results show that elevations of Rab23 wereobserved in the urine, but not in the serum, of the FSGSmice. Rab23 and hedgehog signalling pathway genes wereconstitutively expressed in normal kidneys and were sig-nificantly up-regulated in the kidneys of FSGS mice. Thebasal expression of Rab23 was identified in glomeruli, andmesangial cells displayed obvious elevation of Rab23 in theFSGS state. The knockdown or overexpression of Rab23 af-fected the collagen expression in cultured mesangial cells.Conclusions. An autocrine loop of hedgehog signallingcould be activated in mesangial cells in the FSGS state, andRab23 may be elevated to suppress hedgehog signallingand/or influence collagen synthesis. Importantly, Rab23
Correspondence and offprint requests to: Ann Chen, Department ofPathology, Tri-Service General Hospital, National Defense MedicalCenter, No. 325, Sec. 2, Cheng-Gung Road, Taipei, Taiwan, ROC.Tel: +886-2-8792-7008; Fax: +886-2-8792-7009; E-mail: [email protected] and Dr. Hao-Ai Shui, Graduate Institute of Medical Sci-ences, National Defense Medical Center, 161, Min-Chuan East Road, 6thSection, Taipei, Taiwan, ROC. E-mail: [email protected]
could serve as a biomarker that indicates the severity ofFSGS.
Keywords: biomarker; focal segmentalglomerulosclerosis; hedgehog signal transduction;mesangial cells; Rab23
Introduction
Focal segmental glomerulosclerosis (FSGS) is a form ofchronic nephropathy characterized by scattered sclerosis ofglomeruli in which only a segment of the capillary is af-fected [1,2]. The molecules involved in FSGS pathogenesisare being gradually uncovered. These include proteins thatcause oxidative stress, podocyte damage, hyperfiltrationand sclerosis, which lead to marked proteinuria and scat-tered glomerular sclerosis seen in the disease [2,3]. Occa-sionally, some proteins that were originally not consideredto play any role in FSGS were demonstrated to be involvedin the pathogenesis of the disease. For example, osteopon-tin, a matrixcellular protein with a chemoattractive propertyfor monocytes, has been shown to play a role in epithelialhyperplasia lesion in FSGS [4]. Recently, in our proteomicprofiling of FSGS, we have identified Rab23 in the urineof an FSGS mouse model (unpublished data). However, thechanges in expressional profiles of Rab23 and its associatedgenes in FSGS remain unknown.
Rab23 is a member of the large Rab family of small GT-Pase, which has about 60 members that play varying roles inthe intracellular traffic of proteins and membrane vesicles[5–7]. The expression and function of Rab23 in kidneys areunclear. In contrast to Rab23, many other members of theRab family have been identified in the kidney. Rabs 4, 5,7, 11, 13, 18, 20, 25, 34 and 38 are located in tubular cells[8–11], while Rab 3A is located in podocytes [12]. Rab11and Rab20 co-localize with vacuolar H+-ATPase in tubu-lar and intercalated cells and play roles in regulating pro-ton secretion by transferring the H+-ATPase to the plasma
C© The Author [2008]. Published by Oxford University Press on behalf of ERA-EDTA. All rights reserved.For Permissions, please e-mail: [email protected]
NDT Advance Access published October 16, 2008 by guest on July 12, 2015
http://ndt.oxfordjournals.org/D
ownloaded from

2 T.-H. Huang et al.
membrane [9]. Rab11 also mediates the vasopressin-induced transport of aquaporin-2 water channels to the cellmembrane, and therefore plays a role in water re-absorptionin collecting duct cells [8]. In particular, Rab3A and Rab38are two known factors associated with sclerosis and protein-uria in FSGS. Rab3A is expressed in podocytes and changesin its levels correlate with FSGS in mice and proteinuria inpatients [12]. A Rab38 mutation causes proteinuria in fawn-hooded hypertensive rats and is associated with a familialform of FSGS in human [13,14].
Unlike the general trafficking function of other Rabfamily members, Rab23 plays a unique role in regulatingthe signal transduction of sonic hedgehog, a secreted mor-phogen that controls the development of multiple organsduring embryogenesis [5,15–19]. Hedgehog signalling isinitiated by binding of the ligand sonic hedgehog (Shh),desert hedgehog (Dhh) or Indian hedgehog (Ihh) to thereceptor patched (PTCH, including PTCH1 and PTCH2).This disinhibits the activity of smoothened (SMO), whichthen activates glioma-associated oncogene homologue(Gli) transcription factors (including Gli1, Gli2 and Gli3)to turn on the expressions of their target genes [20]. Rab23appears to antagonize hedgehog signalling, and mutation ofRab23 gene causes a severe defect in the developing neuraltube in mice [15,18] and abnormalities of multiple organsin patients of two congenital diseases—Carpenter and Gor-lin syndromes [21,22]. In addition, since hedgehog genes,such as shh, possess oncogenic properties [20], Rab23 hasbeen implicated to be involved in carcinogenesis. Indeed,the abnormal expression of Rab23 has been observed inhepatocellular as well as thyroid carcinoma [23,24]; theoverexpression of Rab23 has been reported in atrophic gas-tritis, a pre-stage of gastric cancer [25]. However, althoughhedgehog-related genes also play roles in fibrosis and/orsclerosis in brain, lung and liver [26–28], the expressionalprofiles of Rab23 under both of the pathological conditionsare still unclear. Similarly, although hedgehog signaltransduction participates in kidney development [29,30],the expression of Rab23 in the adult kidney is unknown,especially in the kidney that shows glomerular sclerosis.
Although Rab23 was considered to play a major rolein embryogenesis, Rab23 is also found to be highly ex-pressed in adult rodent brain and expressed in low levels inmultiple tissues [5,19]. Rab23 may, therefore, have a post-natal physiological function beyond its role in embryonicdevelopment. We demonstrated in the present study thatRab23 is elevated in the urine of FSGS mice. Using spe-cific antibodies [19] against Rab23, the expression levelsof the protein in normal and FSGS-associated mice wereinvestigated, in conjunction with other hedgehog signallingpathway genes. Our results suggest that Rab23 could playa role in the development of FSGS and could be a markerof FSGS pathology.
Subjects and methods
FSGS animal model and sample collections
The experiments were performed using 8-week-oldBALB/c mice with the approval of the Institutional An-
imal Care and Use Committee of the National DefenseMedical Center, Taiwan, and were consistent with the NIHGuide for the Care and Use of Laboratory Animals. Themice were injected intravenously with a single dose of adri-amycin (AD) (0.1 mg/10 g body weight). Days 0, 7, 15 and20, which cover different severities of FSGS in the mousemodel [3,4,31,32], were chosen for collection of blood andurine samples and for sacrifice of the animals for harvestingof kidneys. Blood was collected through the retro-orbitalvenous plexus [4,31]. To prevent the degradation of uri-nary protein and contamination by cellular proteins due tocell lysis, long-period collection of urine was avoided; in-stead, spot urine was collected at the same time of day onDays 0, 7, 15 and 20. To avoid contamination by food, wa-ter and faeces that can happen in using metabolic cages,mouse urine was obtained by gentle bladder massage andcollected on a Parafilm film [3]. Serum creatinine, bloodurea nitrogen (BUN) and urine protein concentration werethen measured.
Measurement of serum creatinine, BUN and urine proteinconcentration
Samples of serum or urine were cleared by centrifugationand stored in liquid nitrogen until analysis. A modifiedBradford method that can minimize interference from ureain urine was used to measure urine protein concentration[33]; Serum creatinine was measured using a picric acidmethod and BUN was measured using a urease assay aspreviously described [34].
Histopathology and immunohistochemistry
Renal tissues were fixed in 10% buffered formalin and em-bedded in paraffin for routine histopathology. Sections ofthe formalin-fixed renal tissues were immersed in xylene toremove paraffin, rehydrated in graded ethanol, stained withhaematoxylin and eosin, and examined under a light micro-scope (Olympus, Tokyo, Japan). Semi-quantitative analysisof sclerosis was performed with an optical microscope ac-cording to our previously published criterion [3,4,31,32].
For single immunostaining of Rab23, OCT-embeddedtissues were cut into 5-µm-thick sections. After immer-sion in acetone for 5 min and air drying, the sections werethen incubated with Tris-buffered saline, pH 7.4, contain-ing 0.05% Tween 20 (TBST) and 2% bovine serum albumin(BSA) at room temperature for 30 min for the blocking step.TBST was used for all of the following washing steps. Thesections were incubated with a rabbit anti-mouse Rab23antibody (1:200 dilution) at 4◦C overnight, washed withTBST, and incubated with a horseradish peroxidase (HRP)-conjugated goat anti-rabbit antibody (1:200) (JacksonImmunoResearch, West Grove, PA, USA) at room temper-ature for 1 h. Bound antibodies were visualized with 3,3′-diaminobenzidine (DAKO, Carpinteria, CA, USA), and theslides were lightly counterstained with haematoxylin. Semi-quantitative analysis of immunohistochemistry (IHC) sec-tions was performed by optical microscope as previouslydescribed [4,31,35]. Briefly, fifty glomeruli were exam-ined on each slide, and the Rab23 intensity of glomeruli,
by guest on July 12, 2015http://ndt.oxfordjournals.org/
Dow
nloaded from

Rab23, glomerular cells and FSGS 3
podocytes or mesangial cells was assigned a score from0 to 3. The total intensity score for counted glomeruli,podocytes or mesangial cells was calculated according tothe following equation for each specimen: total intensityscore = (% intensity negative × 0) + (% intensity trace in-tensity × 0.5) + (% 1 + intensity × 1) + (% 2 + intensity ×2) + (% 3 + intensity × 3). The values ranged from 0 to amaximum of 300.
For double immunostaining for Rab23/nephrin, the pro-cedure is similar to single immunostaining except that pri-mary antibodies used were rabbit anti-Rab23 (1:150)/goatanti-nephrin (1:20) (Santa Cruz Biotechnology, Santa Cruz,CA, USA), and the secondary antibodies used were fluores-cein isothiocyanate (FITC)-conjugated donkey anti-rabbitIgG (1:500) (Jackson ImmunoResearch, West Grove, PA,USA)/Alexa Fluor 594-conjugated donkey anti-goat IgG(1:500) (Invitrogen, CA, USA). Semi-quantification of to-tal Rab23 fluorescence intensity was performed using thesame equation described above [4,31,35]. For evaluationof the Rab23 expression in podocytes, the area of overlapof the two fluorophores was estimated and presented as apercentage of the total area of glomeruli.
Proteomics analysis
The urine collected by bladder massage was desalted andconcentrated by ultrafiltration as previously described [3].Urinary proteins (400 µg) from the control or FSGSmice were loaded onto an IPG strip (Immobiline DryS-trip 3-10, GE Healthcare, NJ, USA) for simultaneous re-hydration. Isoelectric focusing and SDS–PAGE were per-formed according to the protocols previously described[3,36]. The spots were digested with trypsin and sub-jected to direct mass spectrometry protein identification.The mass spectrometer used for protein analysis was aBruker Biflex IV MALDI-TOF MS (Bruker Daltonics,Bremen, Germany). Peptide mass fingerprint (PMF) wasobtained by averaging signals generated from at least500 laser shots. The mass spectra were processed usingFlexanalysisTM and BiotoolsTM software (Bruker Dalton-ics, Bremen, Germany) and the data subjected to a searchagainst the UniProt database (http://www.pir.uniprot.org)by the MS-Fit database searching engine (http://prospector.ucsf.edu/ucsfhtml4.0/msfit.htm). For each PMF search, themass tolerance was set to 100 ppm and one missed trypticcleavage was allowed.
Cell culture and adriamycin treatment
Mouse mesangial cells CRL-1927 were obtained from theAmerican Type Culture Collection (Rockville, MD, USA)and routinely maintained in a 3:1 mixture of Dulbecco’smodified Eagle’s medium and Ham’s F-12 medium supple-mented with 5% fetal bovine serum and 14 mM HEPESas previously described [37]. Cultured mesangial cells(6 × 105 cells/plate) were seeded onto 10-cm diameter plas-tic dishes before the experiments. After cell attachment,mesangial cells were treated with adriamycin (0–0.2 µg/ml)for 0, 12 or 24 h according to our previous protocol [37].Then, the cells were harvested for immunocytochemistry,real-time RT-PCR and western blot analysis.
Immunocytochemistry
Harvested CRL-1927 cells were attached onto slide glassesby Cytospin (Cytospin 3, Shandon), and then fixed in 2%paraformaldehyde for 15 min. After blocking with 2%BSA in Tris-buffered saline, cells were treated with ananti-Rab23 antibody (1:200) for 1 h and then incubatedwith HRP-conjugated goat anti-rabbit IgG (1:200) (JacksonImmunoResearch, West Grove, PA, USA) for 2 h. Boundantibodies were visualized with 3,3′-diaminobenzidine(DAKO).
Western blot analysis
Urine samples (15 µl for each), serum samples (15 µl foreach) or kidney proteins (50 µg) from adriamycin-treatedmice (Days 0, 7, 15 and 20) or proteins (50 µg) from cul-tured mesangial cells were subjected to 10% SDS–PAGE.Proteins were then transferred to a Hybond PVDF mem-brane (GE Healthcare). The membrane was blocked in a20 ml blocking buffer (Tris-buffered saline, pH 8.0, con-taining 0.05% Tween 20 and 5% skimmed milk) at roomtemperature for 1 h, incubated with a rabbit anti-Rab23 anti-body (1:1000) at room temperature for 1 h, and then washedwith TBST three times, followed by incubation with the goatanti-rabbit secondary antibody for 1 h at room temperature.The proteins on the membrane were visualized with an ECLdetection kit, and protein intensity was quantified by a den-sitometer. For tissue or cell samples, Rab23 intensity wasnormalized by the intensity of the internal control GAPDH.
Real-time reverse transcription-polymerase chain reaction(real-time RT-PCR)
Kidney tissues or cultured mesangial cells were sub-jected to total RNA extraction using Trizol reagent (LifeTechnologies, Rockville, MD, USA) according to themanufacturer’s instruction. For first-strand cDNA syn-thesis, 3 µg of total RNA was used in a single-roundRT reaction, together with 200 ng of random hexamerprimer, 0.5 mM dNTPs, 1× first-strand buffer, 5 mMDTT, 10 units of superscript III reverse transcriptase (In-vitrogen) in a total volume of 20 µl. PCR was per-formed using 1 µl of the RT reaction mixture, 0.1 µMof specific primers (Table 1), 1× PCR buffer, 50 µMdNTPs and one unit of KlenTag DNA polymerase in atotal volume of 25 µl.
Real-time PCR was carried out by the iCycler real-time PCR instrument (Bio-Rad, Hercules, CA, USA), using10 µl of the RT reaction mixture, 0.07 µM gene-specificprimers and 12.5 µl ABsoluteTM QPCR SYBR Green Flu-orescein Mix (AB gene, Epsom, UK), in a total volume of25 µl. The gene-specific primers are listed in Table 1. Thethermal cycler conditions were as follows: real-time PCRwas initiated by enzyme activation at 95◦C for 15 min, fol-lowed by 50 cycles of 94◦C for 15 s, 55◦C for 35 s and72◦C for 60 s and a final extension at 72◦C for 10 min.Levels of test genes were normalized to GAPDH mRNAlevels by a comparative threshold cycle (2−[delta] Ct) methodthat converts differences of the cycle numbers to Test
by guest on July 12, 2015http://ndt.oxfordjournals.org/
Dow
nloaded from

4 T.-H. Huang et al.
Table 1. Primers and gene information for amplification in PCR
Gene Name Accession No. Position of primer in cDNA Primer Sequence Product (bp)
Shh NM_009170 662–681 F: 5′-TCTGTGATGAACCAGTGGCC-3′ 242903–884 R: 5′-GCCACGGAGTTCTCTGCTTT-3′
Ihh NM_010544 1209–1228 F: 5′-CAGGTCATCGAGACTCAGGA-3′ 3321540–1521 R: 5′-GGAAACAGTCGCAGGGGCCA-3′
Dhh NM_007857 600–619 F: 5′-CCATCGCGGTGATGAACATG-3′ 238837–818 R: 5′-TTATCAGCTTTGACCGATAC-3′
PTCH1 NM_008957 886–907 F: 5′-AACAAAAATTCAACCAAACCTC-3′ 2441129–1109 R: 5′-TGTCTTCATTCCAGTTGATGT-3′
PTCH2 NM_008958 684–703 F: 5′-TGCCTCTCTGGAGGGCTTCC-3′ 208891–872 R: 5′-CAGTTCCTCCTGCCAGTGCA-3′
SMO NM_176996 1497–1516 F: 5′-GGCTGGAGTAGTCTGGTTCG-3′ 1911687–1668 R: 5′-AGATGCCACTCACGGAGTCT-3′
Rab23 NM_008999 891–914 F: 5′-GAAGGAGGACCTCAACGTGAGTGA-3′ 1521042–1020 R: 5′-CCAAGTGACTTCTGACCGATGCA-3′
Gli1 NM_010296 796–818 F: 5′-ATCACCTGTTGGGGATGCTGGAT-3′ 3181113–1091 R: 5′-GGCGTGAATAGGACTTCCGACAG-3′
Gli2 NM_001081125 1818–1840 F: 5′-GTTCCAAGGCCTACTCTCGCCTG-3′ 3052122–2100 R: 5′-CTTGAGCAGTGGAGCACGGACAT-3′
Gli3 NM_008130 1839–1861 F: 5′-AGCAAGCAGGAGCCTGAAGTCAT-3′ 2712109–2087 R: 5′-GTCTTGAGTAGGCTTTTGTGCAA-3′
GAPDH BC110311 582–601 F: 5′-ACCACAGTCCATGCCATCAC-3′ 4521033–1014 R: 5′-TCCACCACCCTGTTGCTGTA-3′
Fibronectin NM_010233 6928–6947 F: 5′-GCGACTCTGACTGGCCTTAC-3′ 1667093–7074 R: 5′-CCGTGTAAGGGTCAAAGCAT-3′
COL1A1 NM_007742 4048–4067 F: 5′-GAGAGCATGACCGATGGATT-3′ 1784225–4206 R: 5′-CCTTCTTGAGGTTGCCAGTC-3′
COL1A2 NM_007743 3947–3966 F: 5′-CCGTGCTTCTCAGAACATCA-3′ 1934139–4120 R: 5′-CTTGCCCCATTCATTTGTCT-3′
COL4A1 NM_009931 5029–5048 F: 5′-TCGCCACCATAGAGAGAAGC-3′ 1325160–5141 R: 5′-GCAGAGGCGAGCATCATAGT-3′
COL4A2 NM_009932 3731–3949 F: 5′-ACCAAAGGCTTCCCTGGAT-3′ 1083838–3817 R: 5′-CTGGAAGTGTGTTGGCCTCT-3′
The gene name, accession number of each gene in the NCBI database, position of primer sequence within each cDNA, primer sequence, and productsize are shown. The forward and reverse primers were designed by the website-based program primer 3 (http://frodo.wi.mit.edu/primer3/input.htm) andare located at different exons to prevent amplification of genomic DNA.
gene/GAPDH ratios. Fold changes of mRNA were calcu-lated by a (2−[delta] [delta] Ct) method.
Overexpression and knockdown of Rab23
CRL-1927 mesangial cells were seeded onto 6-cm diam-eter plastic dishes. The overexpression of Rab23 was per-formed as previously reported [19]. Briefly, the pCIneoconstruct carrying full-length Rab23 was transfectedinto cells using Lipofectamine 2000TM (Invitrogen), andcells were harvested 48 h after the transfection. Knock-down of Rab23 was performed by transfection with adouble-stranded small interfering RNA (siRNA) that wasmade by hybridization of a sense (5′-GAUGCAUGA-AUCAUCCAGCAGAUCGAU-3′) RNA oligo and an anti-sense (5′-CGAUCUGCUGGAUGAUUCAAUGCATC-3′)RNA oligo.
Statistics
All data are presented as mean ± SEM. Statistical analysiswas performed with one-way ANOVA by using the SPSSstatistics analysis software program. A Newman–Keuls testwas carried out when the ANOVA comparisons gave a sig-nificant result. Differences were considered significant atP < 0.05.
Results
Pathophysiological manifestations exhibited by mice in theFSGS model
Consistent with our previous reports [3,4,31,32], severeproteinuria and deteriorated renal clearance function wereexhibited by the adriamycin-treated mice. Compared tobasal levels (0.59 ± 0.082 mg/ml), the urine protein levelswere significantly elevated on Day 15 (16.84 ± 3.13 mg/ml) (F = 13.81, P < 0.01) (Figure 1A). Serum creatinineand BUN, two general parameters for evaluating renal func-tion, were also higher in adriamycin-treated mice comparedto control mice. Serum creatinine showed significant eleva-tions on Day 15 (2.02 ± 0.17 mg/dl versus the basal levelsof 0.48 ± 0.03 mg/dl; F = 25.17, P < 0.01) (Figure 1B),and BUN also showed significant increases on Day 15(71.42 ± 8.09 mg/dl versus the basal levels of 21.85 ±1.99 mg/dl; F = 38.79, P < 0.01) (Figure 1C).
Glomerular sclerosis in the FSGS model
Histopathological examination was performed on kidneysections obtained on Days 0, 7, 15 and 20. As shown inFigure 2A, expansion of the extracellular matrix (ECM) anddeposition of hyaline mass in the glomeruli were obvious
by guest on July 12, 2015http://ndt.oxfordjournals.org/
Dow
nloaded from

Rab23, glomerular cells and FSGS 5
Fig. 1. Pathophysiological manifestations of FSGS mice. Three typicalparameters for evaluation of renal function, (A) urine protein, (B) serumcreatinine and (C) BUN, were measured at the three time points afteradriamycin treatment. n = 6 for each day, ∗∗P < 0.01 compared to Day 0.The dashed line shows the mean values of normal mice.
in FSGS mice in comparison to the control mice. Sclerosisscores were significantly higher in FSGS mice than in con-trol mice on Day 7 (13.17 ± 2.06 compared to basal levelsof 0.0 ± 0.0; F = 74.48, P < 0.01) and continued to ele-vate on Day 15 (63.83 ± 6.20 compared to the basal levels;F = 74.48, P < 0.01) and Day 20 (76.83 ± 5.75 comparedto the basal levels; F = 74.48, P < 0.01) (Figure 2B). Theextent of sclerosis correlated well with the severity of theclinical manifestations as shown in Figure 1.
Identification of Rab23 in urine from FSGS mice byproteomic analysis
We have previously reported serial changes of urinary pro-teome in FSGS mice [3] based on more abundant proteins.However, after the concentration of the urine proteins, we
Fig. 2. Glomerular histopathology in FSGS mice. (A) Kidney tissuehistopathological sections on Days 0, 7, 15 or 20, showing a gradualincrease of sclerosis in the glomeruli (haematoxylin and eosin staining).Original magnification, ×400. (B) A semi-quantitative plot showing thesclerosis scores for the glomeruli in the tissue sections. The sclerosisscores were calculated as described in the Subjects and methods section.n = 6 for each day, ∗P < 0.05 or ∗∗P < 0.01 compared to Day 0.
have now identified a new spot corresponding to Rab23 in2D gels of urine from FSGS mice (white arrow in Figure 3A,right panel) that was absent (undetectable) in gels of con-trol mice (left panel). The peptide mass fingerprint of theidentified protein matches 10 peptides that cover 46.4%Rab23 amino acid sequence (Figure 3B). The expressionof Rab23 during FSGS has never been identified by ouror other labs’ proteomics work. In contrast, most of otherspots shown in Figure 3 have been published in our recentpaper [3].
Serial increase of Rab23 levels in mouse urine duringFSGS
In order to confirm the identification of Rab23, we usedthe specific anti-mouse Rab23 antibody [19] to performwestern blot analysis of the concentrated urine proteins.As shown in Figure 4, urine Rab23 protein can be de-tected by western blot analysis in FSGS mice on Days 7, 15and 20, but undetectable on Day 0 when FSGS was absent(F = 26.18, P < 0.01). The Rab23 levels increased with theprogression and severity of FSGS in mice (Figure 4 versusFigure 2; Figure 4 versus Figure 1). In addition, blood isunlikely to be the source of urine Rab23 protein as Rab23was not detectable in mouse serum at any stage of FSGS(Figure 4A).
by guest on July 12, 2015http://ndt.oxfordjournals.org/
Dow
nloaded from

6 T.-H. Huang et al.
Fig. 3. Detection of Rab23 in FSGS mouse urine by proteomic analysis. (A) Magnified image of the areas containing Rab23 in the 2D gels of urine ofcontrol and FSGS mice. The position of Rab23 is indicated by a white arrow. The molecular mass and pI ranges are indicated, respectively, at the rightand at the top of the images. (B) Peptide mass fingerprint of the identified Rab23 protein. The mass spectrum and identified Rab23 amino acid sequenceare shown. The sequences that match the peptide masses derived from the digested protein are indicated (U: unmodified, O: oxidized methionine, C:alkylated cysteine, P: phosphorylated amino acids). There are 10 matched peptides, and the sequence coverage of Rab23 is 46.4%.
Increases of Rab23 expression in mouse kidneys duringFSGS
In order to delineate the source of urinary Rab23, wechecked both mRNA and protein expressions levels ofRab23 in kidneys of the FSGS mice. As shown inFigure 5A, renal Rab23 mRNA levels on Day 7 (0.12 ±0.019), Day 15 (0.18 ± 0.028) and Day 20 (0.27 ± 0.059)were significantly higher than those on Day 0 (0.013 ±0.0025) (F = 9.85, P < 0.01), again correlating with theincrease in severity of FSGS. Similarly, renal Rab23 pro-tein levels on Day 7 (0.61 ± 0.078), Day 15 (1.02 ± 0.11)and Day 20 (0.93 ± 0.090) were also significantly higher
than those on Day 0 (0.24 ± 0.028) (F = 19.18, P < 0.01),indicating that kidney tissues could be a major source ofurinary Rab23 protein in FSGS mice.
Localizing Rab23 expression in mouse kidneys
In order to determine the tissue/cell location of Rab23 ex-pression in kidneys, we performed single and double im-munostaining to respectively evaluate Rab23 expressionbased on counting colour and fluorescence intensity scores.
Figure 6A shows the semi-quantitative data of single im-munohistochemical stain. Total intensity scores of Rab23
by guest on July 12, 2015http://ndt.oxfordjournals.org/
Dow
nloaded from

Rab23, glomerular cells and FSGS 7
Fig. 4. Western blot analysis of Rab23 in urine and serum of FSGS mice.(A) Representative blots showing Rab23 bands in mouse urine (the topblot) and serum (the bottom blot) at Days 0, 7, 15 and 20. (B) Statisticaldata based on densitometric quantification for Rab23 concentration. n = 6,∗∗P < 0.01 between the two indicated groups.
in glomeruli were elevated on Days 7, 15 and 20 (139.50 ±19.93, 202.33 ± 18.20 and 208.50 ± 23.91, respectively),significantly higher than those on Day 0 (61.83 ± 7.86) (F =14.12, P < 0.01). Similarly, the expression levels of mesan-gial Rab23 in FSGS mice on Days 7, 15 and 20 (145.33 ±17.83, 189.33 ± 21.63, and 181.83 ± 25.07, respectively)were significantly higher than those of control mice on Day0 (67.33 ± 4.72) (F = 8.65, P < 0.01) (Figure 6A), correlat-ing well with the increases of Rab23 transcripts (real-timeRT-PCR) and protein (western blot analysis) shown above(Figure 5). The expression of Rab23 was also noticed inthe podocytes (black arrows in the tissue sections,Figure 6A), but these did not show a significant eleva-tion in FSGS mice on Day 7, 15 or 20 in comparison to thecontrol mice (39.83 ± 3.95) (F = 1.53, P > 0.05).
Figure 6B shows the semi-quantitative data of dou-ble immunostaining of Rab23/nephrin. Consistent withFigure 6A, counting of the fluorescence intensity of to-tal glomerular Rab23 showed that total Rab23 expressionelevated along the course of FSGS disease progression(F = 6.62, P < 0.05). However, the major areas of Rab23elevation do not coincide with nephrin staining. Indeed,based on calculating the overlapping fluorescent areas ofRab23/nephrin, a mild but not significant (F = 0.514,P > 0.05) elevation of Rab23 was observed in the nephrin-positive area.
Constitutive expression of Rab23 and hedgehog signallingpathway genes in mesangial cells
Since Rab23 elevation can only be observed in mesangialcells in FSGS mice, we wondered whether it is merely adrug response to adriamycin stimulation.
Fig. 5. Real-time RT-PCR and western blot analyses of Rab23 in kidneysat different FSGS stages. (A) Statistical data of real-time PCR showing thetime course of change for Rab23/GAPDH mRNA ratios after adriamycintreatment. (B) Western blots showing the changes in Rab23/GAPDH pro-tein ratios. The upper panels show representative gel images, and thelower panels display the statistical data based on densitometric quantifi-cation showing the Rab23/GAPDH ratios. n = 6, ∗P < 0.05 or ∗∗P < 0.01compared to Day 0.
In order to confirm the Rab23 expression in glomerulimesangial cells, we analysed Rab23 expression in a mesan-gial cell line. In addition, since Rab23 is known to play a rolein regulating hedgehog signal transduction, we, therefore,also checked the expressions of hedgehog signalling path-way genes in the mesangial cell line. As shown in Figure 7,constitutive expression of Rab23 can be observed in themouse mesangial cell line CRL-1927 (immunocytochem-istry in Figure 7A, 0 h bar in Figure 7B, and 0 h lane inFigure 7C) and basal constitutive expression of Shh, Ihh,Dhh, PTCH1, PTCH2, SMO, Gli1, Gli2 and Gli3 can alsobe detected in these cells (0 hr bars in Figure 7D).
As shown in Figure 7B and C, the expressions of bothmRNA and protein of Rab23 did not change signifi-cantly after adriamycin (0–0.2 µg/ml) treatment for 24 h(F = 1.315 and 0.029, respectively, P > 0.05). We can,therefore, rule out an immediate and direct drug-elicitedeffect by adriamycin on Rab23 expression in the cells. Sim-ilarly, in Figure 7D, the expressions of hedgehog signalling
by guest on July 12, 2015http://ndt.oxfordjournals.org/
Dow
nloaded from

8 T.-H. Huang et al.
Fig. 6. Expression of Rab23 in kidney tissues at different FSGS stages. (A) Representative IHC-stained sections are displayed in the upper panels,showing the basal expression of Rab23 on Day 0 in both podocytes (black arrows) and mesangial cells (white arrow heads), and elevated expressionsof Rab23 in the mesangium on Days 7, 15 and 20. The lower panels show the scores of colorimetric intensity of Rab23 in glomeruli (left), as well asthe scores of podocytes and mesangial cells (right). n = 6, ∗∗P < 0.01 compared to Day 0. (B) Double immnuofluorescent staining of Rab23/nephrin atdifferent FSGS stages. The upper panels show the representative single-fluorescence images of nephrin and Rab23, and the merged images. The lowerpanels show the scores of total fluorescent intensity of Rab23 in glomeruli (left), and the area of overlapping fluorescence (right).
by guest on July 12, 2015http://ndt.oxfordjournals.org/
Dow
nloaded from

Rab23, glomerular cells and FSGS 9
Fig. 7. Expressions of Rab23 and hedgehog signalling pathway genes in cultured mesangial cells. (A) Immunocytochemical staining of Rab23 incultured mesangial cells. The cells stained with the second antibody are only shown as the negative control. (B) Statistical data of real-time PCRshowing the insignificant change of Rab23 expression after adriamycin (0.2 µg/ml) stimulation. (C) Western blot analysis showing insignificantchanges in Rab23 expression after adriamycin (0.2 µg/ml) treatment. Typical gels representing Rab23 and GAPDH proteins are shown in the upperpanels, and statistical data based on densitometric quantification are shown in the lower panels. P > 0.05 compared to 0 h. (D) Statistical data ofreal-time PCR showing insignificant changes in the expression of hedgehog signalling pathway genes (Shh, Ihh, Dhh, PTCH1, PTCH2, SMO, Gli1,Gli2, Gli3) after adriamycin (0.2 µg/ml) treatment.
pathway genes (Shh, Ihh, Dhh, PTCH1, PTCH2, SMO,Gli1, Gli2 and Gli3) were not significantly altered by adri-amycin (0–0.2 µg/ml) treatment of the mesangial cell line(F = 0.011, 1.54, 1.79. 1.43, 0.32, 0.97, 0.58, 2.30, and0.22, respectively, P > 0.05).
Expressional profiles of hedgehog signalling pathwaygenes in kidneys of FSGS mice
The expression of Rab23 and hedgehog signalling-relatedgenes in adult kidneys has never been studied before.
We, therefore, profiled the expression of these hedgehogsignalling-related genes in the kidneys of both control andFSGS mice. As shown in Figure 8, the expressions of allgenes of the hedgehog signalling pathway can be detectedin kidneys of the normal control mice on Day 0, indicat-ing that these genes are constitutively expressed in normaladult mice kidneys. This finding is interesting as the devel-opmentally regulated hedgehog signalling genes were notexpected to be expressed in high levels in adult kidneys.
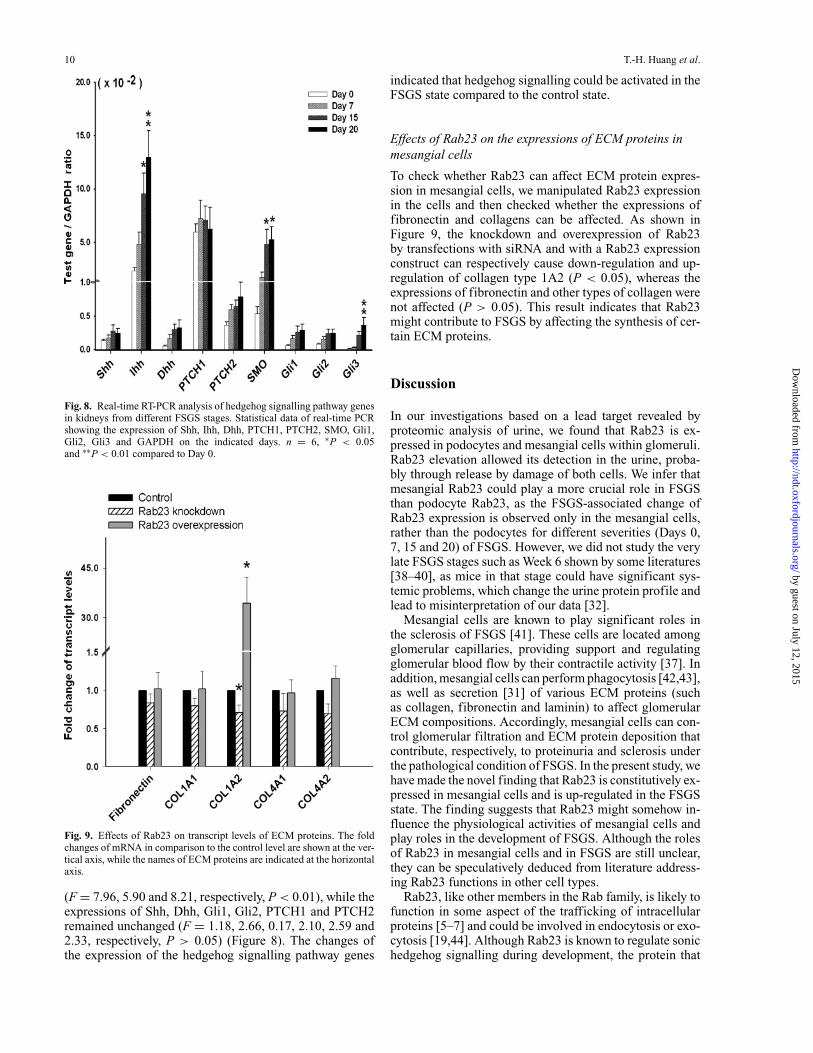
Upon induction of FSGS, the expressions of Ihh, SMOand Gli3 were significantly enhanced by Day 15 and Day 20
by guest on July 12, 2015http://ndt.oxfordjournals.org/
Dow
nloaded from
10 T.-H. Huang et al.
Fig. 8. Real-time RT-PCR analysis of hedgehog signalling pathway genesin kidneys from different FSGS stages. Statistical data of real-time PCRshowing the expression of Shh, Ihh, Dhh, PTCH1, PTCH2, SMO, Gli1,Gli2, Gli3 and GAPDH on the indicated days. n = 6, ∗P < 0.05and ∗∗P < 0.01 compared to Day 0.
Fig. 9. Effects of Rab23 on transcript levels of ECM proteins. The foldchanges of mRNA in comparison to the control level are shown at the ver-tical axis, while the names of ECM proteins are indicated at the horizontalaxis.
(F = 7.96, 5.90 and 8.21, respectively, P < 0.01), while theexpressions of Shh, Dhh, Gli1, Gli2, PTCH1 and PTCH2remained unchanged (F = 1.18, 2.66, 0.17, 2.10, 2.59 and2.33, respectively, P > 0.05) (Figure 8). The changes ofthe expression of the hedgehog signalling pathway genes
indicated that hedgehog signalling could be activated in theFSGS state compared to the control state.
Effects of Rab23 on the expressions of ECM proteins inmesangial cells
To check whether Rab23 can affect ECM protein expres-sion in mesangial cells, we manipulated Rab23 expressionin the cells and then checked whether the expressions offibronectin and collagens can be affected. As shown inFigure 9, the knockdown and overexpression of Rab23by transfections with siRNA and with a Rab23 expressionconstruct can respectively cause down-regulation and up-regulation of collagen type 1A2 (P < 0.05), whereas theexpressions of fibronectin and other types of collagen werenot affected (P > 0.05). This result indicates that Rab23might contribute to FSGS by affecting the synthesis of cer-tain ECM proteins.
Discussion
In our investigations based on a lead target revealed byproteomic analysis of urine, we found that Rab23 is ex-pressed in podocytes and mesangial cells within glomeruli.Rab23 elevation allowed its detection in the urine, proba-bly through release by damage of both cells. We infer thatmesangial Rab23 could play a more crucial role in FSGSthan podocyte Rab23, as the FSGS-associated change ofRab23 expression is observed only in the mesangial cells,rather than the podocytes for different severities (Days 0,7, 15 and 20) of FSGS. However, we did not study the verylate FSGS stages such as Week 6 shown by some literatures[38–40], as mice in that stage could have significant sys-temic problems, which change the urine protein profile andlead to misinterpretation of our data [32].
Mesangial cells are known to play significant roles inthe sclerosis of FSGS [41]. These cells are located amongglomerular capillaries, providing support and regulatingglomerular blood flow by their contractile activity [37]. Inaddition, mesangial cells can perform phagocytosis [42,43],as well as secretion [31] of various ECM proteins (suchas collagen, fibronectin and laminin) to affect glomerularECM compositions. Accordingly, mesangial cells can con-trol glomerular filtration and ECM protein deposition thatcontribute, respectively, to proteinuria and sclerosis underthe pathological condition of FSGS. In the present study, wehave made the novel finding that Rab23 is constitutively ex-pressed in mesangial cells and is up-regulated in the FSGSstate. The finding suggests that Rab23 might somehow in-fluence the physiological activities of mesangial cells andplay roles in the development of FSGS. Although the rolesof Rab23 in mesangial cells and in FSGS are still unclear,they can be speculatively deduced from literature address-ing Rab23 functions in other cell types.
Rab23, like other members in the Rab family, is likely tofunction in some aspect of the trafficking of intracellularproteins [5–7] and could be involved in endocytosis or exo-cytosis [19,44]. Although Rab23 is known to regulate sonichedgehog signalling during development, the protein that
by guest on July 12, 2015http://ndt.oxfordjournals.org/
Dow
nloaded from

Rab23, glomerular cells and FSGS 11
is transported by Rab23 in the hedgehog signalling path-way remains unidentified [5]. Likewise, although Rab23is expressed in high abundance in neurons of adult mousebrains, transport pathways and cargoes that are regulated byRab23 in neurons are also unknown [5,19]. However, us-ing HeLa cells’ phagocytosis of mutant S. Typhimuriumas a model system, Rab23 has been demonstrated tobe involved in the phagocytic membrane traffic [44].Since Rab23 is constitutively expressed in mesangial cells(Figures 6A and 7A–C), Rab23 could control the trafficof cargo proteins whose physiological functions are es-sential to mesangial cells. However, since Rab23 expres-sion is up-regulated in mesangial cells in the FSGS state(Figures 5 and 6), some of the proteins that are transportedby Rab23 in mesangial cells might also play a role in thedevelopment, or disease progress, of FSGS. Mesangial cellshave been reported to be capable of phagocytosing many ex-tracellular proteins, including immune complexes, growthfactors and ECM proteins [42,43]. Since phagocytosis ofECM proteins can influence the development of sclerosis inFSGS, future investigations should address whether Rab23is involved in the phagocytosis function of mesangial cells.
Rab23 expression can apparently influence cell prolifera-tion. Recently, some reports have shown that the overexpres-sion of Rab23 can be observed in certain types of cancers[23–25]. Knockdown of Rab23 expression can actually in-hibit the growth of hepatocarcinoma cells [23], implicatingthat Rab23 has a proliferation-promoting effect, at least inhepatomas. However, since Rab23 can block hedgehog sig-nal transduction [5,20] that is growth promoting in certaincancer cells [45–47], Rab23 might also be anti-proliferativefor certain cells. Excessive proliferation of mesangial cellsis a common phenomenon observed in various types ofnephropathy [48,49], which disrupts the delicate structureof glomeruli and affects their filtration function. Althoughproliferation of mesangial cells in FSGS is not as signifi-cant as other forms of acute nephropathies [48,49], it canoccasionally be observed in the tissue sections of FSGSkidneys (data not shown). The proliferative state of mesan-gial cells might affect severity of sclerosis as proliferationcan increase the number of the cells that are able to phago-cytise and secrete ECM proteins, thus affecting the ECMdeposition. Future studies would address whether Rab23and hedgehog signal transduction can influence the prolif-eration rate of mesangial cells.
The hedgehog signal transduction is initiated by the bind-ing of Shh, Dhh or Ihh to the receptor PTCH, a repressorof SMO. This results in the disinhibition of SMO, which inturn activates the Gli transcription factors and downstreamtarget genes [20]. Secreted hedgehogs play both autocrineand paracrine roles in embryogenesis and carcinogenesis[45–47] and also play pathogenic roles in fibrosis and/orsclerosis in certain organs [26–28]. Since both the ligands(Shh, Dhh and Ihh) and receptors (PTCH1 and PTCH2)are expressed in mesangial cells (Figure 7D), an autocrinerole of the mesangial-derived hedgehog is possible. In ad-dition, since Rab23 is a negative regulator of hedgehogsignalling [5,15–19], the constitutive expression of Rab23in mesangial cells (Figure 7A–C) could play a negativecounterbalancing role in modulating hedgehog signallingin these cells. Since Rab23 and some of the hedgehog sig-
nalling pathway genes were up-regulated in FSGS kidneys(Figures 5, 6 and 8), the predicted autocrine action of hedge-hog in mesangial cells might be activated in the FSGS stateand may play a role in the development and progression ofthe disease.
The exact mechanisms underlying the up-regulation ofRab23 and hedgehog signalling pathway genes in FSGS arestill unclear. The elevated proteins could play either a pro-tective or a pathogenic role in FSGS development. What isclear is that the up-regulation of the genes is not caused byadriamycin directly, as the phenomenon was not observedin adriamycin-treated mesangial cell line (Figure 7). SinceRab23 levels in both kidney tissue and urine correlated withthe severity of FSGS (Figures 4–6 versus Figures 1 and 2),Rab23 is a potential biomarker of chronic nephropathies.Since the knockdown or overexpression of Rab23 can af-fect the collagen expression in cultured mesangial cells(Figure 9), Rab23 may contribute to FSGS by influencingECM protein deposition.
In conclusion, using various approaches, we have demon-strated that Rab23 and hedgehog signalling pathway geneswere constitutively expressed in podocytes and mesangialcells and were up-regulated in mesangial cells in the FSGSstate. Based on the data, we suggested that an autocrineloop of the hedgehog signalling pathway could be activatedin mesangial cells during FSGS in mice, and Rab23 couldplay a feedback role in suppressing the activated hedgehogsignalling transduction. Importantly, Rab23’s elevation inFSGS could serve as a biomarker for chronic nephropathies.
Acknowledgements. This work was supported by grants NSC 97-2320-B-016-010-MY3, NSC 96-2320-B-016-011-MY3 and NSC 96-2320-B-016-012-MY2 from the National Science Council, Taiwan, ROC. B.L.T.is supported by a grant (R-183-000-102-112) from the Ministry of Educa-tion, Singapore, and the academic staff research fund, National Universityof Singapore. T.H.H. is supported by a grant (EZRPF-360221-09511N023)from Chang-Gung Institute of Technology, Taiwan, ROC. H.A.S. is sup-ported by a grant (DOH97-TD-I-111-TM022) from Department of Health,Executive Yuan, Taiwan, ROC.
Conflict of interest statement. None declared.
References
1. Benchimol C. Focal segmental glomerulosclerosis: pathogenesis andtreatment. Curr Opin Pediatr 2003; 15: 171–180
2. Fogo AB. Animal models of FSGS: lessons for pathogenesis andtreatment. Semin Nephrol 2003; 23: 161–171
3. Shui HA, Huang TH, Ka SM et al. Urinary proteome and potentialbiomarkers associated with serial pathogenesis steps of focal segmen-tal glomerulosclerosis. Nephrol Dial Transplant 2008; 23: 176–185
4. Shui HA, Ka SM, Yang SM et al. Osteopontin as an injury markerexpressing in epithelial hyperplasia lesions helpful in prognosis offocal segmental glomerulosclerosis. Transl Res 2007; 150: 216–222
5. Wang Y, Ng EL, Tang BL. Rab23: What exactly does it traffic? Traffic2006; 7: 746–750
6. Pfeffer SR. Structural clues to Rab GTPase functional diversity. J BiolChem 2005; 280: 15485–15488
7. Stenmark H, Olkkonen VM. The Rab GTPase family. Genome Biol2001; 2: reviews 3007
8. Barile M, Pisitkun T, Yu MJ et al. Large scale protein identifica-tion in intracellular aquaporin-2 vesicles from renal inner medullarycollecting duct. Mol Cell Proteomics 2005; 4: 1095–1106
by guest on July 12, 2015http://ndt.oxfordjournals.org/
Dow
nloaded from

12 T.-H. Huang et al.
9. Curtis LM, Gluck S. Distribution of Rab GTPases in mouse kidneyand comparison with vacuolar H+-ATPase. Nephron Physiol 2005;100: 31–42
10. Speight P, Silverman M. Diacylglycerol-activated Hmunc13 serves asan effector of the GTPase Rab34. Traffic 2005; 6: 858–865
11. Tajika Y, Matsuzaki T, Suzuki T et al. Differential regulation ofAQP2 trafficking in endosomes by microtubules and actin filaments.Histochem Cell Biol 2005; 124: 1–12
12. Rastaldi MP, Armelloni S, Berra S et al. Glomerular podocytes pos-sess the synaptic vesicle molecule Rab3A and its specific effectorrabphilin-3a. Am J Pathol 2003; 163: 889–899
13. Osanai K, Takahashi K, Nakamura K et al. Expression and character-ization of Rab38, a new member of the Rab small G protein family.Biol Chem 2005; 386: 143–153
14. Rangel-Filho A, Sharma M, Datta YH et al. RF-2 gene modulatesproteinuria and albuminuria independently of changes in glomerularpermeability in the fawn-hooded hypertensive rat. J Am Soc Nephrol2005; 16: 852–856
15. Eggenschwiler JT, Bulgakov OV, Qin J et al. Mouse Rab23 regulateshedgehog signaling from smoothened to Gli proteins. Dev Biol 2006;290: 1–12
16. Evans TM, Simpson F, Parton RG et al. Characterization of Rab23,a negative regulator of sonic hedgehog signaling. Methods Enzymol2005; 403: 759–777
17. Evans TM, Ferguson C, Wainwright BJ et al. Rab23, a negative reg-ulator of hedgehog signaling, localizes to the plasma membrane andthe endocytic pathway. Traffic 2003; 4: 869–884
18. Eggenschwiler JT, Espinoza E, Anderson KV. Rab23 is an essentialnegative regulator of the mouse Sonic hedgehog signalling pathway.Nature 2001; 412: 194–198
19. Guo A, Wang T, Ng EL et al. Open brain gene product Rab23: expres-sion pattern in the adult mouse brain and functional characterization.J Neurosci Res 2006; 83: 1118–1127
20. Ruiz i Altaba A. Gli proteins and Hedgehog signaling: developmentand cancer. Trends Genet 1999; 15: 418–425
21. Oostra RJ, Maas M. Bifid ribs and unusual vertebral anomalies diag-nosed in an anatomical specimen. Gorlin syndrome? Am J Med GenetA 2006; 140: 2135–2138
22. Jenkins D, Seelow D, Jehee FS et al. Rab23 mutations in Carpentersyndrome imply an unexpected role for hedgehog signaling in cranial-suture development and obesity. Am J Hum Genet 2007; 80: 1162–1170
23. Liu YJ, Wang Q, Li W et al. Rab23 is a potential biological target fortreating hepatocellular carcinoma. World J Gastroenterol 2007; 13:1010–1017
24. Denning KM, Smyth PC, Cahill SF et al. A molecular expressionsignature distinguishing follicular lesions in thyroid carcinoma us-ing preamplification RT-PCR in archival samples. Mod Pathol 2007;20:1095–1102
25. Kim KR, Oh SY, Park UC et al. Gene expression profiling usingoligonucleotide microarray in atrophic gastritis and intestinal meta-plasia. Korean J Gastroenterol 2007; 49: 209–224
26. Mastronardi FG, daCruz LA, Wang H et al. The amount of sonichedgehog in multiple sclerosis white matter is decreased and cleavageto the signaling peptide is deficient. Mult Scler 2003; 9: 362–371
27. Sicklick JK, Li YX, Choi SS et al. Role for hedgehog signaling inhepatic stellate cell activation and viability. Lab Invest 2005; 85:1368–1380
28. Stewart GA, Hoyne GF, Ahmad SA et al. Expression of the devel-opmental Sonic hedgehog (Shh) signalling pathway is up-regulatedin chronic lung fibrosis and the Shh receptor patched 1 is present incirculating T lymphocytes. J Pathol 2003; 199: 488–495
29. Yu J, Carroll TJ, McMahon AP. Sonic hedgehog regulates pro-liferation and differentiation of mesenchymal cells in the mousemetanephric kidney. Development 2002; 129: 5301–5312
30. Zhang L, Hua F, Yuan GH et al. Sonic hedgehog signaling is critical forcytodifferentiation and cusp formation in developing mouse molars.J Mol Histol 2007
31. Shui HA, Ka SM, Lin JC et al. Fibronectin in blood invokes thedevelopment of focal segmental glomerulosclerosis in mouse model.Nephrol Dial Transplant 2006; 21: 1794–1802
32. Chen A, Sheu LF, Ho YS et al. Experimental focal segmental glomeru-losclerosis in mice. Nephron 1998; 78: 440–452
33. Ramagli LS. Quantifying protein in 2-D PAGE solubilization buffer.In: Link, AJ, ed. 2-D Proteome analysis protocols Humana Press,Totowa, NJ: 1999; 99–103
34. Montinaro V, Hevey K, Aventaggiato L et al. Extrarenal cytokinesmodulate the glomerular response to IgA immune complexes. KidneyInt 1992; 42: 341–353
35. Ka SM, Rifai A, Chen JH et al. Glomerular crescent-related biomark-ers in a murine model of chronic graft versus host disease. NephrolDial Transplant 2006; 21: 288–298
36. Shui HA, Ho ST, Wang JJ et al. Proteomic analysis of spinal proteinexpression in rats exposed to repeated intrathecal morphine injection.Proteomics 2007; 7: 796–803
37. Jin JS, Yao CW, Chin TY et al. Adriamycin impairs the contractionof mesangial cells through the inhibition of protein kinase C andintracellular calcium. Am J Physiol Renal Physiol 2004; 287: F188–F194
38. Vielhauer V, Berning E, Eis V et al. CCR1 blockade reduces inter-stitial inflammation and fibrosis in mice with glomerulosclerosis andnephrotic syndrome. Kidney Int 2004; 66: 2264–2278
39. Wang Y, Wang YP, Tay YC et al. Progressive adriamycin nephropathyin mice: sequence of histologic and immunohistochemical events.Kidney Int 2000; 58: 1797–1804
40. Wang Y, Wang YP, Tay YC et al. Role of CD8(+) cells in the progres-sion of murine adriamycin nephropathy. Kidney Int 2001; 59: 941–949
41. Cornacchia F, Fornoni A, Plati AR et al. Glomerulosclerosis is trans-mitted by bone marrow-derived mesangial cell progenitors. J ClinInvest 2001; 108: 1649–1656
42. Mancilla-Jimenez R, Bellon B, Kuhn J et al. Phagocytosis of heat-aggregated immunoglobulins by mesangial cells: an immunoperoxi-dase and acid phosphatase study. Lab Invest 1982; 46: 243–253
43. Cosio FG, Sedmak DD, Nahman NS Jr. Cellular receptors for matrixproteins in normal human kidney and human mesangial cells. KidneyInt 1990; 38: 886–895
44. Smith AC, Heo WD, Braun V et al. A network of Rab GTPases con-trols phagosome maturation and is modulated by Salmonella entericaserovar Typhimurium. J Cell Biol 2007; 176: 263–268
45. Yuan Z, Goetz JA, Singh S et al. Frequent requirement of hedgehogsignaling in non-small cell lung carcinoma. Oncogene 2007; 26: 1046–1055
46. Zhang J, Lipinski R, Shaw A et al. Lack of demonstrable autocrinehedgehog signaling in human prostate cancer cell lines. J Urol 2007;177: 1179–1185
47. Zhu G, Zhau HE, He H et al. Sonic and desert hedgehog signaling inhuman fetal prostate development. Prostate 2007; 67: 674–684
48. Baker AJ, Mooney A, Hughes J et al. Mesangial cell apoptosis: themajor mechanism for resolution of glomerular hypercellularity in ex-perimental mesangial proliferative nephritis. J Clin Invest 1994; 94:2105–2116
49. Shui HA, Ka SM, Wu WM et al. LPS-evoked IL-18 expression inmesangial cells plays a role in accelerating lupus nephritis. Rheuma-tology (Oxford) 2007; 46: 1277–1284
Received for publication: 6.1.08Accepted in revised form: 18.9.08
by guest on July 12, 2015http://ndt.oxfordjournals.org/
Dow
nloaded from
Related Documents











