Quantitative Genetic Perspectives on Loss of Diversity in Elite Maize Breeding Germplasm Jode W. Edwards USDA ARS CICG [email protected]

Quantitative Genetic Perspectives on Loss of Diversity in Elite Maize Breeding Germplasm Jode W. Edwards USDA ARS CICG [email protected].
Dec 19, 2015
Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Quantitative Genetic Perspectives on Loss of Diversity
in Elite Maize Breeding Germplasm
Jode W. Edwards
USDA ARS [email protected]

Outline
• Diversity
• Population genetics of maize
• Quantitative genetic processes– Bottlenecks– Selection
• Implications

What is diversity?
• D = 1 – pi2
– pi = allele frequency
• At Hardy-Weinberg equilibrium D is an estimator of heterozygosity, H
• With population subdivision, heterozygosity is related to Fst:
– Ht = (1-1/2N)tH0 = (1-Fst)H0
Sources: Nei, M, 1973, PNAS , 70:3321-3323; Wright, S., 1943, Genetics, 28:114-138

Diversity in Maize Inbreds and LandracesTenaillon, Sawkins, Long, Gaut, Doebley, and Gaut, 2001
• Estimated SNP diversity by sequencing– 7 known genes, – 6 cDNA clones– 8 RFLP clones– All chromosome 1
• Germplasm– 16 exotic landraces (1 inbred per landrace)– 9 U.S. inbreds (B73, Mo24W, Mo17, W153R, Ky21,
NC258, Oh43, Tx601, T8)• Inbreds contained 77% as much diversity as the
landraces (DI/DL)Source: Tenaillon et al., 2001, PNAS, 98:9161-9166

• ‘the U.S. inbred sample retains a high proportion of diversity, which is difficult to explain given that U.S. elite germplasm has a narrow origin based largely on two open-pollinated varieties, Reid yellow dent and Lancaster (14)’ – [“(14)” is Major Goodman’s paper in Heredity]
Tenaillon et al. Conclusion

Is 77% Hard to Explain?
• 1 - Fst = 1 - 0.77
• For Fst of 0.23, N=2.2
If inbreds were:• Sampled randomly,
E[1-Fst] = 0.89
• Subpopulation with Fst = 0.87, E[1-Fst] = 0.77
0
0.2
0.4
0.6
0.8
1
0 25 50 75 100
Inbred lines
He
tero
zyg
os
ity

How should we measure diversity?
Heterozygosity (formally)?Number of alleles?Number of polymorphic loci?Number of rare alleles?JE thoughts:
Diversity is important, but we don’t know how to measure it (or what it is)Something else may be more important

Sustainable Selection Response
• Plant breeders’ main goal is selection – Short term: Maximum response– Long term: Sustainable response
• In order to address sustainability of selection response, we need to understand phenotype– Population genetics of maize– Quantitative genetics of population bottlenecks– Quantitative genetics of selection with finite
size

Maize Population Genetics: BSSS
• Started with maize land races (O.P.) and develop ‘first cycle’ inbreds
• 16 lines intermated to form BSSSC0– Expected diversity = 87.5% of ancestor
• B14, B37 emerge from Cycle zero– Expected diversity = .875 x .5 = 43.75%
• B73, B84 emerge from C5, C7

Corn Belt Maize Land Races
• Outcrossing, monoecious populations
• Large Ne (?)
• Mass selected for visual characteristics (low h2?)• Corn belt dents existed 100+ generations, longer
for other groups• Corn belt dents (Labate et al., 2003)
– Accessions: Fst = 0.15
– Varieties: Fst = 0.04
• “Almost” one large randomly mated populationSource: Labate, J.A. et al., 2003, Crop Science, 43:80-91

Maize Land Races
• Hardy-Weinberg equilibrium
• Linkage equilibrium
• Mutation-selection equilibrium

Haldane (1937) Principle
• Mutation frequencies determined by equilibrium– New mutations are constantly added to the population– Mutations removed by selection (and drift)– Mutation rates estimated to be 0.4 – 1.0 per diploid
individual per generation
• At equilibrium– Individuals carry many mutations– Reduction in fitness due to mutations = “genetic
mutation load” (Muller)
Source: Haldane, J.B.S., 1937, The American Naturalist, 71:337-359; Crow, J.F., 1993, Oxford Surveys in Evolutionary Biology, 9:3-42

Does Mutation Load Apply to Maize?
• Inbreeding depression– Severe in first cycle inbreds– Less in germplasm with inbreeding history
(purging of recessives)– If many loci carry mutations, complete purging
takes many generations
• Observation of major “lethal” mutations
• Empirical work in maize is needed!

Significance of Haldane Principle
• Mutation load provides a model of quantitative genetic variation more realistic than ‘infinitessimal theory’
• Provides a basis for understanding quantitative genetic variation, and thus,
• Basis for predicting effects of bottlenecks and artificial selection

Bottlenecks
• Population is formed from small number of individuals– Change allele frequencies– Hardy-Weinberg and linkage disequilibria
• Under additive model– ‘within subpoplation variance’, Vw = (1-Fst) 2
A
– ‘among subpopulation variance’, Vb =2Fst 2A
• Non-additive model: effects of bottlenecks are complex
Source: Wang, J., et al., 1998, Genetics, 150:435-447
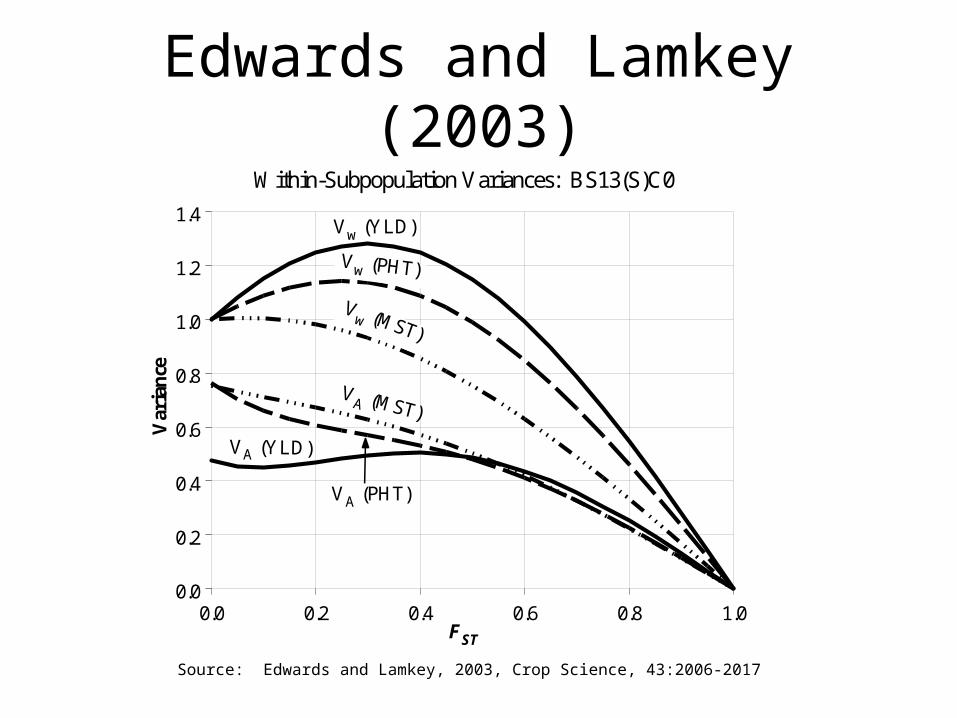
Edwards and Lamkey (2003)
0.0 0.2 0.4 0.6 0.8 1.0FST
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
Variance
Within-Subpopulation Variances: BS13(S)C0
Vw (YLD)
Vw (PHT)
Vw (MST)
VA (YLD)
VA (MST)
VA (PHT)
Source: Edwards and Lamkey, 2003, Crop Science, 43:2006-2017

Garcia, Lopez-Fanjul, and Garcia-Dorado, 1994D. melanogaster, Full-sib lines
0
40
80
120
160
0 0.25 0.5 0.75
Inbreeding Coefficient
Var
ian
ce
Source: Garcia, N., et al., 1994, Evolution, 48:1277-1285

Gene Effect Sizes Wang, Caballero, Keightley, and Hill, 1998
0
10
20
30
40
50
60
70
80
0-50 50-100 100-200
200-300
300-400
400-500
>500 lethals
Effect size (Ne x s)
Per
cen
t o
f ad
dit
ive
vari
ance
Source: Wang, J., et al., 1998, Genetics, 150:435-447

Gene Effects and Bottlenecks
• Genes of all sizes important in the base
• After a bottleneck: large recessives become much more important (and hence large increase in dominance)
• Explanation: Nonlinear relationship between frequency and variance: small increase in frequency = large increase in variance

Limits to Selection ResponseRobertson, 1960
• Max response = 2 Ne times initial response• Half-life occurs at 1.4 Ne generations• Total response is maximized at 50%
intensity (greater with linkage)• Based on ‘infinitessimal’ theory
– Many genes of ‘infinitely’ small effect– Can we understand ‘side effects’ of selection
under more realistic conditions?Source: Robertson, A., 1960, Proc. Roy. Soc. London, Ser. B, 153:235-249

Selection Effects
• Loss of heterozygosity (diversity)
• Linkage disequilibrium– Bulmer– Hill-Robertson
• Epistasis

Linkage and Selection
• Bulmer effect– Correlation between alleles induced by
selection– Causes excess of coupling phase linkages
and reduced genetic variance
• Hill-Robertson effect– Effect of repulsion phase linkages– Unfavorable alleles become fixed because of
selection for favorable alleles linked in repulsion phase
Sources: Bulmer, M.G., 1971, American Naturalist, 105:201-211; Hill, W.G. and Robertson, A., 1968, Theor. Appl. Genet., 38:226-231

Zhang and Hill, 2005
• Simulated selection in cage populations derived from ‘equilibrium natural populations’ of D. melanogaster
• Conditions– Genetic model: mutation-selection balance under
joint pleiotropic and stabilizing selection– 40% intensity– Recombine 40 individuals
– VG0 = 0.5 VE
– 3 chromosomes of varying lengthSource: Zhang, X.S., and Hill, W.G., 2005, Genetics, 169:411-425

Selection and Linkage
Zhang and Hill, 2005
Source: Zhang, X.S., and Hill, W.G., 2005, Genetics, 169:411-425

Gene Numbers and EffectsZhang and Hill, 2005
• Distribution of gene effects– 90% of genes have a<0.1p and account for
27% of genetic variance
– 10% of genes have a>0.1p and account for the rest of the genetic variance
• Estimated that 103 – 104 loci are polymorphic in a cage population
Source: Zhang, X.S., and Hill, W.G., 2005, Genetics, 169:411-425

Evidence of Linkage in Maize
• Degree of dominance, d, can be estimated as a ratio, D
2/A2, in F2-derived
populations• Linkage disequilibrium causes
a bias called ‘associative overdominance’
• Random mating breaks up linkage and reduces bias
AA Aa -> d=1
aa
Aa -> d=2
Aa -> d=0

Maize NCIII Experiments
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
0 3 6 9 12 15
Generations random mated
Av
era
ge
de
gre
e o
f d
om
ina
ce
GardnerLonnquist
Lonnquist, J.H., 1980, Anal. Acad. Nac. Cs. Ex. Fis. Nat., 32:195-201; Gardner, C. O., Personal communication to E.T. Bingham

Epistasis
• Favorable epistatic interactions are increased by selection
• Lamkey, Schnicker, and Melchinger, 1995– Began with BSSS lines B73 (cycle 5) and B84 (cycle
7)
– Formed the F1, F2, BC1 (to both parents) and intermated F2
– Testcrossed all generations onto Mo17– With additive model (no epistasis) there is a linear
relationship among generations
Source: Lamkey, K.R., et al., 1995, Crop Science, 35:1272-1281

Epistasis in B73 and B84Lamkey, Schnicker, and Melchinger, 1995
Source: Lamkey, K.R., et al., 1995, Crop Science, 35:1272-1281

How did we get here?
• Bottleneck followed by 5 and 7 cycles of selection
• During selection– Linkage disequilibrium increases– Epistatic combinations become more important– Selection may be dominated by genes of large effect
• Slow increase in frequency of many small favorable alleles is not a good model– For positive effects, i.e., response– For negative effects

Sustainable Response is a Function of More than Diversity
• Loss of alleles (diversity)
• Increase in linkage disequilibrium (reduced variance)
• Increased dependence on specific epistatic combinations
• Shift in size of genes that contribute to genetic variance (small to big)

Implications for Elite x Exotic Crosses
• Genetic variance within a single population is due mostly to genes of large effect
• Linkage disequilibrium within the cross may reduce genetic variance
• Any new alleles from the exotic parent are preferentially lost if:– Linked to negative alleles at physiologically selected
loci, e.g., photoperiod– There are favorable epistatic interactions among elite
alleles

What can be done?
• Map major genes (especially photoperiod) and use markers to break linkages
• Recycle lines from different crosses• Enhance or improve land races directly to
maintain more variation and reduce disequilibrium– If major genes were identified, could speed up with
markers– Preserve more variation due to genes of small effect
• Random mate individual crosses

Basic Research Questions
• How differentiated are maize land races from each other and from elite lines?– At neutral loci– At selected loci
• Can we identify major genes that– Differentiate elite lines from ancestral varieties– Corn belt dent from tropical races
• Genetic architecture– Can we estimate mutation load parameters?– Can we distinguish purging of recessive load from
selection for physiological effects

We can succeed doing what we are already doing
However, can we be more successful?
Related Documents











