Quantitative Evaluation of HIV-1 Coreceptor Use in the GHOST(3) Cell Assay Dalma Vo ¨dro ¨s,* , † , ‡ ,1 Charlotte Tscherning-Casper,* Leonor Navea,§ Dominique Schols, ¶ Erik De Clercq, ¶ and Eva Maria Fenyo ¨* , † *Microbiology and Tumorbiology Center, Karolinska Institute, Stockholm, Sweden; †Department of Medical Microbiology, Dermatology and Infection, Lund University, Lund, Sweden; ‡National Center for Epidemiology, Budapest, Hungary; §National AIDS Research Laboratory, Havana, Cuba; and ¶ Rega Institute for Medical Research, K.U. Leuven, Leuven, Belgium Received February 27, 2001; returned to author for revision July 15, 2001; accepted August 16, 2001 The utility of the GHOST(3) cell assay has been evaluated for testing coreceptor use of primary human immunodeficiency virus type 1 (HIV-1) isolates. GHOST(3) cells were derived from the human osteosarcoma cell line, HOS, and have been engineered to stably express CD4 and one or another of the chemokine receptors CCR3, CCR5, CXCR4, Bonzo, or the orphan receptor BOB. The indicator cell line carries the HIV-2 long terminal repeat-driven green fluorescence protein (GFP) gene, which becomes activated upon infection with HIV or simian immunodeficiency virus. Viral entry is followed by Tat activation of transcription and GFP becomes expressed. Infected cells can be detected 2 or 3 days after infection by simple fluorescence microscopic observation. This simplicity is the main advantage of the GHOST(3) cell system and makes it particularly suitable for screening of a large number of isolates. In addition, the efficiency of coreceptor use can be accurately quantitated with flow cytometric analysis. Here, we evaluated the coreceptor use of 59 primary HIV-1 isolates of different subtypes. © 2001 Elsevier Science Key Words: GHOST(3); GFP; coreceptor; quantitative assay; HIV; SIV. INTRODUCTION Human and simian immunodeficiency viruses (HIV and SIV) enter target cells using CD4, the primary cell surface receptor, and a secondary receptor (coreceptor) from the G-protein-coupled seven-transmembrane che- mokine receptor family (Chen et al., 1997; Deng et al., 1996, 1997; Dragic et al., 1996; Edinger et al., 1997, 1998a,b; Farzan et al., 1997; Feng et al., 1996; Hoffman et al., 1998; Marcon et al., 1997). Biological features of HIV-1 correlate with the coreceptor used. CCR5 using (R5) viruses are isolated during the early, asymptomatic phase of infection, while CXCR4 using (X4) or dual-tropic R5X4 viruses can often be detected at the late phase of infection concomitant with development of AIDS (Bjo ¨rn- dal et al., 1997; Scarlatti et al., 1997). Based on their phenotype in cultures of peripheral blood mononuclear cells (PBMC), R5 viruses have previously been classified as slow/low or non-syncytium-inducing (NSI), whereas X4 viruses were designated rapid/high or syncytium- inducing (SI) (Fenyo ¨ et al., 1988; Tersmette et al., 1988, 1989). Viruses from the latter group were able to infect established cell lines of T-lymphoid or monocytoid origin. While a clear relationship between HIV-1 biological phe- notype and severity of infection could be established, the pattern of SIV coreceptor use in relation to pathogenesis remained unclear (Chen et al., 1997; Edinger et al., 1997, 1999; Rudensey et al., 1998). For determination of HIV and SIV coreceptor use, dif- ferent kinds of indicator cell lines have been utilized. These cell lines contain reporter genes such as the gene for chloramphenicol acetyltransferase (CAT) (Merzouki et al., 1995), b-galactosidase (b-gal) (Kimpton and Emer- man, 1992), luciferase (luc) (Schwartz et al., 1990), or alkaline phosphatase (Means et al., 1997), under the control of HIV-1 or HIV-2 long terminal repeat (LTR). Upon infection the viral Tat protein increases transcription from the HIV LTR promoter, leading to high-level expression of the reporter gene (Dorsky et al., 1996). Detection of the reporter gene products, however, requires processing of the cells by lysis, by fixation, or by addition of a substrate. Here we describe an indicator cell system in which the readout is a simple microscopic observation, without cumbersome processing of cells. The GHOST cells (de- rived from the human osteosarcoma cell line, HOS) con- tain the gene of the green fluorescence protein (GFP) driven by the HIV-2 ROD LTR. The cells have been engi- neered to stably express CD4, the primary receptor used by HIV and SIV, and one of several coreceptors (Cecilia et al., 1998; Mo ¨rner et al., 1999; KewalRamani, unpub- lished data). Upon infection, viral entry is followed by Tat activation of transcription and GFP becomes expressed. Since these are early steps in the virus replication cycle, infected cells can be detected already 2 or 3 days after 1 To whom correspondence and reprint requests should be addressed at Department of Medical Microbiology, Dermatology, and Infection, Divi- sion of Virology, Lund University, So ¨lvegatan 23; S – 223 62 Lund, Sweden. Fax: 146-46-176-033. E-mail: [email protected]. Virology 291, 1–11 (2001) doi:10.1006/viro.2001.1163, available online at http://www.idealibrary.com on 0042-6822/01 $35.00 © 2001 Elsevier Science All rights reserved. 1

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Virology 291, 1–11 (2001)doi:10.1006/viro.2001.1163, available online at http://www.idealibrary.com on
Quantitative Evaluation of HIV-1 Coreceptor Use in the GHOST(3) Cell Assay
Dalma Vodros,*,†,‡,1 Charlotte Tscherning-Casper,* Leonor Navea,§ Dominique Schols,¶
Erik De Clercq,¶ and Eva Maria Fenyo*,†
*Microbiology and Tumorbiology Center, Karolinska Institute, Stockholm, Sweden; †Department of Medical Microbiology, Dermatology andInfection, Lund University, Lund, Sweden; ‡National Center for Epidemiology, Budapest, Hungary; §National AIDS
Research Laboratory, Havana, Cuba; and ¶Rega Institute for Medical Research, K.U. Leuven, Leuven, Belgium
Received February 27, 2001; returned to author for revision July 15, 2001; accepted August 16, 2001
The utility of the GHOST(3) cell assay has been evaluated for testing coreceptor use of primary human immunodeficiencyvirus type 1 (HIV-1) isolates. GHOST(3) cells were derived from the human osteosarcoma cell line, HOS, and have beenengineered to stably express CD4 and one or another of the chemokine receptors CCR3, CCR5, CXCR4, Bonzo, or the orphanreceptor BOB. The indicator cell line carries the HIV-2 long terminal repeat-driven green fluorescence protein (GFP) gene,which becomes activated upon infection with HIV or simian immunodeficiency virus. Viral entry is followed by Tat activationof transcription and GFP becomes expressed. Infected cells can be detected 2 or 3 days after infection by simplefluorescence microscopic observation. This simplicity is the main advantage of the GHOST(3) cell system and makes itparticularly suitable for screening of a large number of isolates. In addition, the efficiency of coreceptor use can be accuratelyquantitated with flow cytometric analysis. Here, we evaluated the coreceptor use of 59 primary HIV-1 isolates of different
subtypes. © 2001 Elsevier ScienceKey Words: GHOST(3); GFP; coreceptor; quantitative assay; HIV; SIV.
INTRODUCTION
Human and simian immunodeficiency viruses (HIVand SIV) enter target cells using CD4, the primary cellsurface receptor, and a secondary receptor (coreceptor)from the G-protein-coupled seven-transmembrane che-mokine receptor family (Chen et al., 1997; Deng et al.,1996, 1997; Dragic et al., 1996; Edinger et al., 1997,1998a,b; Farzan et al., 1997; Feng et al., 1996; Hoffman etal., 1998; Marcon et al., 1997). Biological features of HIV-1correlate with the coreceptor used. CCR5 using (R5)viruses are isolated during the early, asymptomaticphase of infection, while CXCR4 using (X4) or dual-tropicR5X4 viruses can often be detected at the late phase ofinfection concomitant with development of AIDS (Bjorn-dal et al., 1997; Scarlatti et al., 1997). Based on theirphenotype in cultures of peripheral blood mononuclearcells (PBMC), R5 viruses have previously been classifiedas slow/low or non-syncytium-inducing (NSI), whereasX4 viruses were designated rapid/high or syncytium-inducing (SI) (Fenyo et al., 1988; Tersmette et al., 1988,1989). Viruses from the latter group were able to infectestablished cell lines of T-lymphoid or monocytoid origin.While a clear relationship between HIV-1 biological phe-notype and severity of infection could be established, the
1 To whom correspondence and reprint requests should be addressedat Department of Medical Microbiology, Dermatology, and Infection, Divi-
sion of Virology, Lund University, Solvegatan 23; S – 223 62 Lund, Sweden.Fax: 146-46-176-033. E-mail: [email protected].1
pattern of SIV coreceptor use in relation to pathogenesisremained unclear (Chen et al., 1997; Edinger et al., 1997,1999; Rudensey et al., 1998).
For determination of HIV and SIV coreceptor use, dif-ferent kinds of indicator cell lines have been utilized.These cell lines contain reporter genes such as the genefor chloramphenicol acetyltransferase (CAT) (Merzouki etal., 1995), b-galactosidase (b-gal) (Kimpton and Emer-man, 1992), luciferase (luc) (Schwartz et al., 1990), oralkaline phosphatase (Means et al., 1997), under thecontrol of HIV-1 or HIV-2 long terminal repeat (LTR). Uponinfection the viral Tat protein increases transcription fromthe HIV LTR promoter, leading to high-level expression ofthe reporter gene (Dorsky et al., 1996). Detection of thereporter gene products, however, requires processing ofthe cells by lysis, by fixation, or by addition of a substrate.Here we describe an indicator cell system in which thereadout is a simple microscopic observation, withoutcumbersome processing of cells. The GHOST cells (de-rived from the human osteosarcoma cell line, HOS) con-tain the gene of the green fluorescence protein (GFP)driven by the HIV-2ROD LTR. The cells have been engi-neered to stably express CD4, the primary receptor usedby HIV and SIV, and one of several coreceptors (Ceciliaet al., 1998; Morner et al., 1999; KewalRamani, unpub-lished data). Upon infection, viral entry is followed by Tatactivation of transcription and GFP becomes expressed.
Since these are early steps in the virus replication cycle,infected cells can be detected already 2 or 3 days after0042-6822/01 $35.00© 2001 Elsevier ScienceAll rights reserved.

ing-Ca
2 VODROS ET AL.
infection with the help of a fluorescence microscope.This simplicity is the main advantage of the GHOST cellsystem. In the present work, we used the GHOST cellsystem and obtained quantitative data by instrumentalmeasurements using a flow cytometer (FACS). By mea-suring fluorescence intensity and the proportion of in-fected cells, the efficiency of viruses in using a certaintype of coreceptor could be determined. In addition,productive infection was evaluated by measuring theHIV-1 p24 antigen content of culture supernatants byenzyme-linked immunosorbent assays (ELISA).
RESULTS
Evaluation of data obtained by flow cytometry
GHOST(3) cell lines expressing CD4 and one of eachcoreceptor CCR3, CCR5, CXCR4, BOB, or Bonzo were in-fected with 59 different HIV-1 isolates (Table 1). Infectedcells were analyzed on a flow cytometer 3 days after infec-tion. In order to find a quantitative way of evaluating theefficiency of infection we took into consideration both thepercentage of fluorescence-positive cells (%) and the meanfluorescence intensity (FI) obtained for each virus–corecep-tor combination. These two parameters were multiplied foreach sample and the fold difference from uninfected controlcells was calculated as
T
Features of
Origin Subtype U87 assaya
Cameroon A CCR5CCR5
A/Ja CCR5CCR5
A/G (IBNG) CCR5CCR5
B CXCR4D CCR5F CCR5
U/F CCR5, CCR3, CXCR4CCR5, CXCR4
Cuba B CXCR4CCR5, CXCR4CCR3, CXCR4
Tanzania D CXCR4India C CCR5
Sweden C Not tested25 B CCR2b, CCR5, CCR3, CXCR
a Tested on U87.CD4–CCR1, -CCR2b, -CCR3, -CCR5, and -CXCR4 ceb Tested on GHOST(3) cells expressing CD4 and one of the CCR3, Cc A/J, A subtype by env and J by the protease sequences (Tschern
sequencing; gag subtype F.
RTCN 5 ~% 3 FI!virus/~% 3 FI!control,
where RTCN (ratio to cell negative) gives a quantitativemeasure of the efficiency of infection. Controls weremock-infected cultures from the corresponding corecep-tor-expressing cell lines. The FI cut-off level was set to0.1% (0.08–0.13%) fluorescence-positive cells for the un-infected cultures. Thus the mean (% 3 FI/FIcutoff) valuesfor the negative controls ranged from 0.14, for the paren-tal and BOB- and Bonzo-expressing cell lines, to 0.17, forthe CCR5-expressing cell line. The standard deviationdid not exceed 0.04 for any of the cell lines (data basedon 13 independent experiments). The close similarity ofthese values in the different negative control culturesallowed us to compare the efficiencies with which thedifferent receptors are used by viruses.
We then made the assumption that RTCN valuesabove 10 are positive, those below 5 are negative, andthose between 5 and 10 are indeterminate. To seewhether this assumption holds, we examined the rela-tionship of the calculated RTCN values to the percentagefluorescence-positive cells and the fluorescence inten-sity (fold increase above the negative control) for eachdata point obtained on GHOST(3).CCR5 cells (Fig. 1). Thedata points enclosed in the area formed by 0.5% positivecells and FI/FIneg 5 2 included the mock-infected cul-tures and viruses that were negative on U87.CD4-CCR5
ses Tested
ptor use
Number of isolatesGHOST(3) assay
CCR5 23CCR5, Bonzo 2CCR5 5CCR5, Bonzo 1CCR5 9CCR5, Bonzo 2CXCR4 2CCR5 1CCR5 2CCR5, CCR3, CXCR4 2CCR5, CXCR4 1
CXCR4 2CCR5, CXCR4 1CCR3, CXCR4 2
CXCR4 1CCR5 1
CCR5 1CCR5, CCR3, CXCR4, BOB, Bonzo 1
herning-Casper et al., 2000a).XCR4, BOB, or Bonzo receptors.
sper et al., 2000b). U/F, env subtype unclassified both by HMA and
ABLE 1
the Viru
Corece
4
lls (TscCR5, C
cells. RTCN values for this on the GHOST(3).CCR5 cellswere less than 5. The indeterminate data points (5 ,

, 10 ,p of th
3HIV-1 CORECEPTOR USE IN THE GHOST(3) CELL ASSAY
RTCN , 10) within or immediately outside this areaeither were classified as X4 viruses on U87.CD4 cells(points C, E, F, G in Fig. 1) (Bjorndal et al., 1997; Tschern-ing-Casper et al., 2000a) or had R5 phenotype (points A,B, and D) but were used in very small amounts (55 ID50,infectious dose-50, on PBMC) in the present experimentsinvolving GHOST(3) cells. The viruses depicted in datapoints H and I had low but positive RTCN values (10.9and 11.4, respectively). One of these viruses had the R5phenotype (data point I) but was tested in small amounts(ID50 5 5); the other was of X4 phenotype (data point H)and infected GHOST(3).CCR5 cells due to the endoge-nous CXCR4 expression in these cells (see Discussion).
All other data points in Fig. 1 indicate that RTCN valuesrepresent an adequate measurement of the efficiency ofcoreceptor use and, in the following, will be applied tocharacterize the efficiency of virus–coreceptor interac-tions.
Comparison of flow cytometry with microscopicobservation and HIV-1 p24 antigen production
Infected GHOST cells were observed in a fluores-cence microscope 3 days after infection. The same dayFACS analysis was carried out and RTCN values were
FIG. 1. Correlation between the proportion of fluorescence-positive(FI/FIneg). Individual points represent RTCN values: }, RTCN . 100; ■
GHOST(3).CCR5 cells are included in this figure. The inset is a blow-u
calculated. Viral antigen production was determined inculture supernatants by ELISA 6 days after infection.
Thus, for each culture, data were obtained by threedifferent methods.
Results obtained by microscopic observation and flowcytometric analysis showed strict correlation on bothCCR5- and CXCR4-expressing cells (Figs. 2A and 2B).Microscopic observation is thus sufficient and can beused alone for screening of a large amount of material.
For the comparison of RTCN values and HIV-1 p24antigen production, viruses were divided into severalgroups: R5, X4 or R5X4, and Cuban isolates. This wasnecessary because of the known endogenous expres-sion of CXCR4 on the GHOST(3) cells. The data obtainedby the different methods were validated within eachgroup.
In general, infection of the GHOST(3).CCR5 cells withR5 viruses showed that once RTCN was positive, p24antigen could also be detected in culture supernatants(Fig. 3A). However, large quantitative variations in anti-gen production were present. In fact, in two cases (iso-late 4164C and 1986B from Cameroon) culture superna-tants remained antigen negative. Discordant RTCN andantigen values are explained by extensive cell death atthe time of sampling for antigen production (day 6 postin-fection). In contrast, CXCR4-expressing cells infected
nd the increase of fluorescence intensity above the negative controlRTCN , 100; ‚, 5 , RTCN , 10; E, RTCN , 5. All tests done on thee critical part of the diagram.
cells a
with R5 viruses remained negative by both parameters(Fig. 3B). Conversely, infection with X4 viruses clearly

4 VODROS ET AL.
induced GFP and yielded antigen-producingGHOST(3).CXCR4 cultures. However, three of four X4viruses gave low but definitely positive RTCN onGHOST(3) cells engineered to express CCR5. These cul-tures nevertheless remained antigen production nega-tive. As expected, infections with R5X4 dual-tropic vi-ruses led to strong fluorescence induction and highlevels of antigen production in both cell types.
Only one Cuban HIV-1 isolate was classified as R5X4by using the U87.CD4 cell system (Table 1). It was there-fore surprising that these viruses induced GFP in CCR5-as well as CXCR4-expressing GHOST(3) cells. However,antigen production was highly variable in CCR5-express-ing cells. In fact the only virus that yielded large amountsof p24 antigen was the one classified as R5X4 dual-tropicon U87.CD4 cells. These results suggested that the Cu-ban viruses either used the endogenously expressed
FIG. 2. Comparison of observed (microscope) and measured (flowcytometry) fluorescence following infection with HIV-1 isolates. (A)GHOST(3).CCR5 cells; (B) GHOST(3).CXCR4 cells. Microscopic evalua-tion of fluorescence was validated according to the amount of fluores-cence-positive cells: 1, 1–5% fluorescent cells; 11, 5–10% fluorescentcells; 111, more than 10% fluorescent cells in the well. Flow cytomet-ric analysis: both the fluorescence intensity and the number of fluores-cence-positive cells (percentage gated) were considered and the folddifference to uninfected cultures was calculated (RTCN). Mean RTCNand standard deviation (SD) of the RTCN are shown for each valuedetermined by microscopic observation. Results of repeated experi-ments are included in this figure.
CXCR4 very efficiently or used yet another undefinedreceptor present on the GHOST(3) cells. To distinguish
between these two possibilities, inhibition experimentswere carried out with the specific CXCR4 antagonistAMD3100.
Inhibition of CXCR4-using viruses with the CXCR4antagonist AMD3100
To be able to dissect the contribution of CXCR4 use onthe evaluation of receptor use of CXCR4-using viruses,infection of GHOST(3) cells was performed in the pres-ence or absence of the specific CXCR4 antagonist,AMD3100 (Table 2). In all cases, whether the viruseswere multi-, dual-, or mono-tropic (R3R5X4, R5X4 or X4,respectively), infection of the parental cells could beinhibited. Similarly, infection of CCR3-expressing cellscould also be inhibited, except with the Cuban 95Cu132virus and the Cameroonian 001A and 001E isolatesknown to use CCR3. As expected, infection of CCR5-expressing cells by R5X4 viruses could not be inhibited,whereas CXCR4 use was inhibited in all cases. Thisallows us to conclude that the receptor used by HIV-1 toenter the parental cells (or CCR3- or CCR5-expressingcells by viruses not using these receptors) is solelyCXCR4. The results in the GHOST(3) cell system agreewith those obtained on U87.CD4 cells (Table 1).
The influence of endogenous CXCR4 expression onthe detection of coreceptor use
First, we examined infection of the parental GHOST(3)cells (Fig. 4A). None of the R5 viruses gave a positiveRTCN value on the parental cells (mean RTCN is 1.6),whereas in all other groups, consisting of CXCR4-usingviruses, at least some isolates induced GFP in the pa-rental cells. Notably, dual- or multi-tropic R5X4 or R3R5X4Cameroonian isolates and Cuban viruses were the mostefficient in this respect. These cultures, however, pro-duced much less viral antigen than the correspondingGHOST(3).CXCR4 cultures (data not shown). Similar re-sults were obtained on CXCR4- or CCR3-expressingcells, in that R5 viruses were negative (mean RTCN is 0.6and 1.3, respectively). However, 2 of 30 isolates in thisgroup induced GFP in BOB-expressing cells, althoughcultures remained antigen production negative, whileboth GFP induction and antigen production character-ized infection of the GHOST(3).CCR5 cells when infectedwith R5 viruses.
Such a clear distinction could not be found withCXCR4-using viruses. As expected, all of them efficientlyinfected CXCR4-expressing cells (Fig. 4D), but could alsoinduce GFP to a variable extent in other GHOST(3) sub-lines. In this respect, monotropic X4 viruses appeared tobe the most specific, whereas the dual-tropic R5X4 orR3X4 and the multi-tropic R3R5X4 HIV-1 isolates gavepositive results on the GHOST(3) cells expressing CCR3
or BOB. The RTCN values obtained with these viruses onGHOST(3).CCR3 cells were as high as with virus 25,
5HIV-1 CORECEPTOR USE IN THE GHOST(3) CELL ASSAY
FIG. 3. Comparison of flow cytometric measurement and viral antigen production. GHOST(3) cells expressing CCR5 (A) or CXCR4 (B) coreceptorwere infected with HIV-1 isolates. Antigen production represented as OD490 values of the infected culture supernatants and GFP induction expressedas RTCN are shown. Viruses were tested one to three times, the data of one representative experiment are shown here. The R5 3344 and the
multitropic 25 control viruses were included in each experiment along with an uninfected culture. Results obtained with these control viruses areshown as the average of all experiments.
6 VODROS ET AL.
which is known to use CCR3 (Bjorndal et al., 1997).However, productive infection was more pronounced inthose cases when CCR3 usage could be demonstratedon U87.CD4–CCR3 cells.
The effect of virus dose
Parental cells and CXCR4- or CCR5-expressing cellswere repeatedly infected with the multitropic 25 virus. Ahigher dose of infectious virus (measured as ID50 onPBMC) gave higher RTCN values in all three cultures.With a lower virus dose the parental cells became neg-ative.
To further examine the dilution effect we carried outexperiments with serial virus dilutions (Fig. 5). RTCNvalues strictly correlated with the amount of infectingvirus, indicating that GFP induction measures viral entry.Correlation was also seen between infectious virus doseand the amount of HIV-1 p24 antigen production 6 daysafter infection. With all three viruses and all coreceptorstested (CCR5, CXCR4, and Bonzo) 50 ID50 (measured onPBMC) appears to be sufficient for quantitative evalua-tion of coreceptor use in the GHOST(3) cell system. Ifinfection is performed with a lower virus dose, the mostefficiently used coreceptor(s) can still be identified butadditional coreceptor use may be overlooked.
Expression of CD4 and coreceptors on the GHOST(3)cells
An interesting observation was that RTCN values onthe parental cells (or on CCR3- or CCR5-expressing cells
TABLE 2
Inhibition of CXCR4 Receptor on the GHOST(3) Cells
Virusisolate Phenotypea
AMD3100CXCR4
antagonist
RTCN on GHOST(3) cells
Parental CCR5 CXCR4 CCR3
001A R3R5X4 2 ,10 144 164 36.31 ,10 119 ,10 23.8
001E R3R5X4 2 ,10 280 358 53.71 ,10 313 ,10 36.2
001B R5X4 2 ,10 146 122 12.71 — 182 ,10 ,10
909B X4 2 25.5 12.9 292 21.41 ,10 ,10 ,10 ,10
1388C X4 2 21.8 ,10 90 11.21 11.9 — ,10 ,10
1388D X4 2 23.0 30.5 149 19.31 ,10 ,10 ,10 ,10
95Cu110 X4 2 31.2 28.4 401 45.31 ,10 ,10 ,10 11.1
95Cu132 R3X4 2 42.6 66.6 828 99.11 ,10 ,10 ,10 62.9
a Coreceptor use on U87.CD4 cells.
when infected by viruses not using these receptors)were in all cases lower than on the CXCR4- (or CCR5-)
transfected cells. This indicated more efficient infectionof the cells engineered to express CXCR4 compared tocells expressing “background” levels of CXCR4. Thisprompted us to analyze receptor expression in the dif-ferent GHOST(3) sublines.
The GHOST(3).CCR5 and -CCR3 cell lines were dou-ble-stained for CD4 and CCR5 or CCR3, respectively, andall GHOST(3) cell lines were double-stained for CXCR4and CD4. As judged from the parallel dislocation of cellpopulations after staining with the specific monoclonalantibodies, CD4 and CXCR4 expression was present inall cells of each GHOST(3) subline (Fig. 6). The disloca-tion of cells stained for CD4 was more pronounced (foldincrease of median values was 11.87–17.01) than thosestained for CXCR4 (fold increase of median values was1.73–2.87), suggesting higher levels of CD4 expressionthan of CXCR4 expression. The levels of CCR5 and CCR3expression on the CCR5- and CCR3-transfected cells,respectively, were comparable to those of CXCR4 ex-pression (fold increase of median values in populationsstained with the specific monoclonal antibodies was 2.29and 1.83, respectively). It is surprising that the GHOST(3)cell sublines, whether transfected with CXCR4 or not,express comparable levels of CXCR4 when visualized bystaining with monoclonal antibody, yet the efficiency ofinfection by CXCR4-using viruses is lower when the en-dogenously expressed CXCR4 receptor is utilized forentry. If a part of the receptors detected by the antibodyis not functional as coreceptors for HIV-1, such a resultwould be expected. It is therefore mandatory to usestandard virus controls in each experiment, these willgive a better estimate of coreceptor function.
DISCUSSION
In the present work we describe a simple and sensi-tive assay for HIV-1 coreceptor use that gives quantita-tive results in 3 days. We used the GHOST(3) indicatorcells, which express CD4 and different coreceptors forHIV and SIV and carry the gene for GFP driven by theHIV-2 LTR. GFP becomes activated upon infection andthe read-out is optimally performed 3 days postinfection.Qualitative data can be obtained by simple microscopicobservation. With the help of flow cytometric analysis theefficiency of coreceptor use can be quantitated. Here weshow that by taking into consideration both the percent-age of fluorescent cells and their fluorescence intensityand calculating the fold difference from the uninfectedcontrol, each virus–receptor combination can be quanti-tatively determined. With the use of 59 HIV-1 isolates and6 sublines of the GHOST(3) cells (the parental cells and5 cell lines transfected with different coreceptors) theassay has been validated. The same validation applies toSIV and the accompanying paper (Vodros et al., 2001)
describes the coreceptor use of 20 sequentially obtainedSIVsm isolates.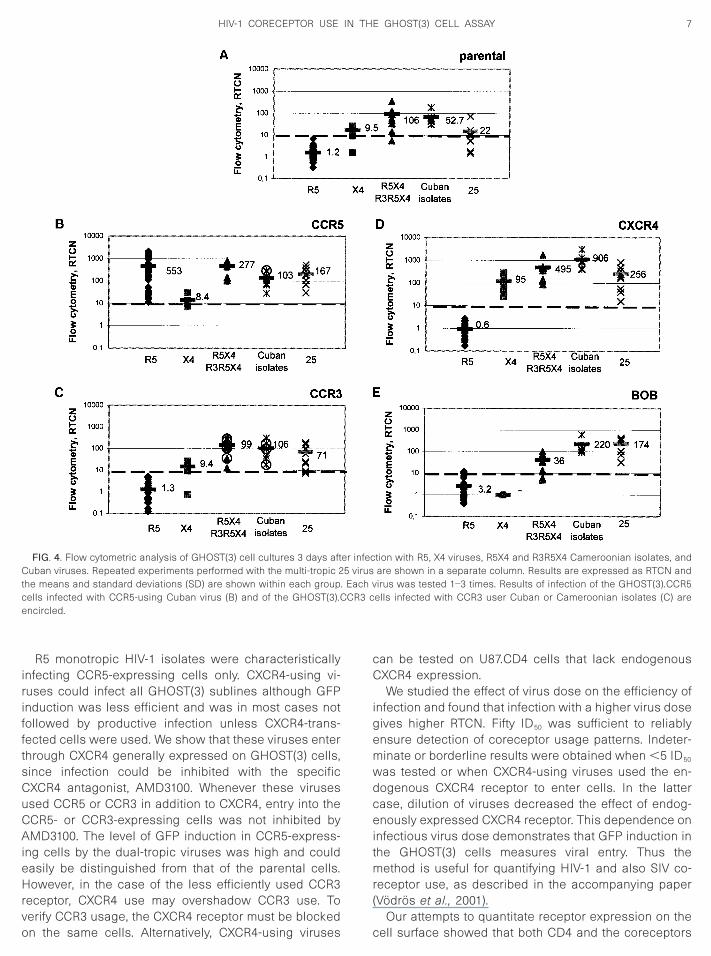
CR3 c
7HIV-1 CORECEPTOR USE IN THE GHOST(3) CELL ASSAY
R5 monotropic HIV-1 isolates were characteristicallyinfecting CCR5-expressing cells only. CXCR4-using vi-ruses could infect all GHOST(3) sublines although GFPinduction was less efficient and was in most cases notfollowed by productive infection unless CXCR4-trans-fected cells were used. We show that these viruses enterthrough CXCR4 generally expressed on GHOST(3) cells,since infection could be inhibited with the specificCXCR4 antagonist, AMD3100. Whenever these virusesused CCR5 or CCR3 in addition to CXCR4, entry into theCCR5- or CCR3-expressing cells was not inhibited byAMD3100. The level of GFP induction in CCR5-express-ing cells by the dual-tropic viruses was high and couldeasily be distinguished from that of the parental cells.However, in the case of the less efficiently used CCR3receptor, CXCR4 use may overshadow CCR3 use. To
FIG. 4. Flow cytometric analysis of GHOST(3) cell cultures 3 days aftCuban viruses. Repeated experiments performed with the multi-tropic 2the means and standard deviations (SD) are shown within each groupcells infected with CCR5-using Cuban virus (B) and of the GHOST(3).Cencircled.
verify CCR3 usage, the CXCR4 receptor must be blockedon the same cells. Alternatively, CXCR4-using viruses
can be tested on U87.CD4 cells that lack endogenousCXCR4 expression.
We studied the effect of virus dose on the efficiency ofinfection and found that infection with a higher virus dosegives higher RTCN. Fifty ID50 was sufficient to reliablyensure detection of coreceptor usage patterns. Indeter-minate or borderline results were obtained when ,5 ID50
was tested or when CXCR4-using viruses used the en-dogenous CXCR4 receptor to enter cells. In the lattercase, dilution of viruses decreased the effect of endog-enously expressed CXCR4 receptor. This dependence oninfectious virus dose demonstrates that GFP induction inthe GHOST(3) cells measures viral entry. Thus themethod is useful for quantifying HIV-1 and also SIV co-receptor use, as described in the accompanying paper(Vodros et al., 2001).
tion with R5, X4 viruses, R5X4 and R3R5X4 Cameroonian isolates, andare shown in a separate column. Results are expressed as RTCN andirus was tested 1–3 times. Results of infection of the GHOST(3).CCR5
ells infected with CCR3 user Cuban or Cameroonian isolates (C) are
er infec5 virus
. Each v
Our attempts to quantitate receptor expression on thecell surface showed that both CD4 and the coreceptors

ction o
8 VODROS ET AL.
were expressed on all cells of a given population. Addi-tion of the specific monoclonal antibodies resulted in ashift of the entire cell population. We expressed the
FIG. 5. Effect of virus dilution on the infection of the GHOST(3) cellsisolates were tested in different dilutions on the GHOST(3) parentalCalculated RTCN values are shown as columns while antigen produinfectious virus, titrated on PBMC, are also shown.
FIG. 6. Level of receptor expression on the GHOST(3) cells. The GHOand -CCR3 cells were stained for CCR5 and CCR3, respectively, and the
samples without specific antibody added, dotted lines show distribution of cellsthe fold difference in median values of stained cells compared to the negativmagnitude of the shift as the difference in median valuesof stained/unstained populations, as suggested by Leeet al. (1999). In this way we found a large difference
CR5- and Bonzo-using 2236B and R3R5X4 multitropic 001E and 001Ae (par) and cells expressing CCR5, CXCR4, or Bonzo, as indicated.
f the infected cultures is represented by the lines. ID50 values of the
ell lines were stained for CD4 and CXCR4. In addition, GHOST(3).CCR5pulations were analyzed by flow cytometry. Continuous lines represent
. The Ccell lin
ST(3) ccell po
with specific antibody added. The numbers in each histogram indicatee control samples without specific antibody added.

9HIV-1 CORECEPTOR USE IN THE GHOST(3) CELL ASSAY
between CD4 and coreceptor expression in general, inthat CD4 showed a higher level of expression than anyother coreceptors. Edinger et al. (1998a) made a similarobservation when comparing CD4 and CCR5 expressionon 293T and GHOST cells. In our experiments not onlyCCR5, but also CXCR4 and CCR3 showed similar lowlevels of expression when cells were tested with thedifferent specific monoclonal antibodies. The most sur-prising finding was that CXCR4 expression, as detectedby the antibody, appeared to differ very little betweenparental cells and CXCR4-transfected cells, while virusinfection was more efficient on the CXCR4-transfectedcells than on parental cells. This indicates that the anti-body binding assay does not give information on recep-tor function and the most important controls in the assayare standardized virus controls.
MATERIALS AND METHODS
Viruses
Viruses of different origins and subtypes were used tocalibrate the GHOST cell system. HIV-1 isolates fromCameroonian pregnant women, mainly subtype A, wereobtained within the framework of a European Networkfor “In Utero Transmission of HIV” (Tscherning-Casper etal., 2000a). HIV-1 subtype B isolates from Cuba and theTZ98010 and IN97003 isolates from Tanzania and India,respectively, were obtained within the framework of theUNAIDS Network for HIV Characterization. HIV-1 strain25, known to use all the coreceptors tested, was in-cluded in all experiments as “positive control” virus(Bjorndal et al., 1997). Virus stocks were produced inhuman PBMC and cell-free supernatants were used toinfect GHOST(3) cells. The characteristics of the virusesincluded in this study are shown in detail in Table 1.
Cell lines
The human osteosarcoma cell line, GHOST(3), wasengineered to stably express CD4 and one or another ofthe chemokine receptors CCR3, CCR5, CXCR4, Bonzo, orthe orphan receptor BOB (Cecilia et al., 1998; Matloubianet al., 2000; Morner et al., 1999; KewalRamani, unpub-lished data). The parental cell line has been engineeredto express CD4 but none of the coreceptors. The cellswere stably transfected with the GFP gene driven by theHIV-2ROD LTR. In the case of infection when the virusenters the cells by using CD4 and the appropriate core-ceptor, the viral Tat protein becomes expressed andtransactivates the GFP gene by the LTR. GFP expressionin infected cells is easily detected in a fluorescencemicroscope and by flow cytometry. The GHOST(3) celllines were maintained in Dulbecco’s modified Eagle’smedium (Life Technologies, Palsley, Scotland) containing
7.5% fetal calf serum (FCS, Life Technologies, Karlsruhe,Germany) and antibiotics and incubated in a humidifiedatmosphere with 5% CO2 at 37°C. Cultures were splittwice a week by use of 5 mM EDTA.
Infection of the GHOST cells
One day before infection 24-well plates were preparedwith 2–3 3 104 cells/well in 1 ml medium. Before infec-tion, medium was replaced with 200 ml fresh mediumand virus was added to duplicate wells in a volume of300 ml/well. In parallel experiments, using PBMC, wedetermined the ID50 for each virus (Weber et al., 1996).Two hours after infection, medium was added to 1 ml/well. After an overnight incubation, cells were washedwith PBS (Life Technologies, Palsley, Scotland), 1 mlmedium was added to each well, and the plates werefurther incubated. Three days after infection cultureswere observed in a fluorescence microscope and cellsfrom one of the parallel wells were prepared for flowcytometry by adding paraformaldehyde to a final concen-tration of 2% for at least 2 h. Remaining wells were split1:5–1:10 and cultures were maintained for another 3 daysfor viral antigen detection by ELISA.
In one series of experiments the specific CXCR4 an-tagonist AMD3100 was used (Schols et al., 1997).AMD3100 was added to the cells prior to infection in 200ml medium at a concentration of 1 mg/ml and virus wasadded 5 min later. Parallel wells without inhibitor wereincluded in each experiment and treated in the sameway.
Flow cytometric analysis
We selected the GHOST cell population on the sidescatter–forward scatter diagram and measured the fluo-rescence intensity of 1–1.5 3 104 cells (FACScan, BectonDickinson, San Jose, CA). The percentage of fluores-cence-positive cells and their fluorescence intensitywere determined and further used to calculate a charac-teristic number to evaluate the efficiency of infection asdetailed under Results.
Viral antigen detection
To detect the antigen produced by the infected cells,an in-house HIV-1 p24 antigen ELISA was used. Themethod has been described previously (Sundqvist et al.,1989). In brief, 100-ml aliquots of cell-free supernatantscontaining 0.5% Triton 100 were added to 96-well micro-titer plates previously coated with rabbit anti-Gag sera.Antigen was allowed to bind during an overnight incu-bation at 4°C. Plates were then washed and a secondaryantibody (anti-HIV-1 p24 mouse monoclonal IgG) conju-gated with horseradish peroxidase was added. Follow-ing incubation for 2 h at 37°C, the plates were washedand the substrate, o-phenylenediamine-dichloride acti-vated with H2O2, was added. The reaction was stopped
with 2.5 M H2SO4 and the optical density of the productwas measured spectrophotometrically at 490 nm (OD490).
10 VODROS ET AL.
ID50 titration
Virus isolates were titrated on PBMC in an ID50 assayas previously described (Weber et al., 1996). Briefly, 6fivefold dilutions were made, starting from 1:5. Seventy-five microliters of each virus dilution was added to fiveparallel wells in 96-well round-bottom microtiter plates(Nunc, Roskilde, Denmark). Thereafter 1 3 105 phytohe-magglutinin (Sigma) stimulated PBMC from two healthydonors in a 150-ml volume were added to each well andthe plates were incubated for 6 days. At days 1 and 3 theplates were washed by centrifugation and change of 200ml medium. At day 6, supernatants from each well wereanalyzed for the presence of viral antigen by ELISA. TheID50 was defined as the reciprocal of the virus dilutionresulting in 50% positive wells (Reed–Muench calcula-tion).
Receptor staining
For staining, 1 3 106 GHOST(3) cells were washedwith staining buffer (SB: PBS, containing 2% FCS) andantibodies were added. For CXCR4 labeling, 20 ml undi-luted antibody (12G5; Endres et al., 1996) was added tothe samples. For CCR5 and CCR3 labeling, the antibod-ies 2D7 and 7B11 (Heath et al., 1997), respectively, werefirst diluted in SB to a final concentration of 10 ng/ml, and25 ml of diluted antibody was added to each sample. Asnegative control 10 ml of triple-color reagent (Dako,Glostrup, Denmark) was used. Cells were then incubatedon ice for 30 min. After two washings with SB, 50 ml of1:20 diluted anti-mouse-IgG FITC conjugate was addedto the samples. Double-staining for CD4 was performedat this step. For this we used 10 ml of the CD4/RPE-Cy5-conjugated antibody (Dako; Catalog No. C7069) accord-ing to the manufacturer’s recommendation. Followingincubation on ice for 30 min in the dark, the cells werewashed twice with SB and fixed by addition of 300 ml of4% paraformaldehyde. Analysis was then performed on aFACScan using Cellquest software.
ACKNOWLEDGMENTS
We thank Dr. Dan R. Littman and his colleagues at the HowardHughes Medical Institute, Skirball Institute for BioMolecular Medicine,New York University, New York, for kindly providing the GHOST(3)indicator cell lines. Monoclonal antibodies to CCR5 and CCR3 wereprovided by Leukosite Inc., and the NIBSC Centralized Facility for AIDSReagents. Antibodies to CXCR4 (12G4) were provided by Dr. J. Hoxiethrough the AIDS Reagent Project. We thank Kerstin Andreasson andRobert Fredriksson for their technical assistance. Support for the workperformed herein was received from the Swedish Medical ResearchCouncil, the Swedish International Development Cooperation Agency,and the Biomed 2 Programme of the European Commission.
REFERENCES
Bjorndal, Å., Deng, H., Jansson, M., Fiore, J. R., Colognesi, C., Karlsson,
A., Albert, J., Scarlatti, G., Littman, D. R., and Fenyo, E. M. (1997).Co-receptor usage of primary human immunodeficiency virus type 1isolates varies according to biological phenotype. J. Virol. 71(10),7478–7487.
Cecilia, D., KewalRamani, V. N., O’Leary, J., Volsky, B., Nyambi, P., Burda,S., Xu, S., Littman, D. R., and Zolla-Pazner, S. (1998). Neutralizationprofiles of primary human immunodeficiency virus type 1 isolates inthe context of coreceptor usage. J. Virol. 72(9), 6988–6996.
Chen, Z. W., Zhou, P., Ho, D. D., Landau, N. R., and Marx, P. (1997).Genetically divergent strains of simian immunodeficiency virus useCCR5 as a coreceptor for entry. J. Virol. 71(4), 2705–2714.
Deng, H., Unutmaz, D., KewalRamani, V. N., and Littman, D. R. (1997).Expression cloning of new receptors used by simian and humanimmunodeficiency viruses. Nature 388(6639), 296–300.
Deng, H. K., Liu, R., Ellmeier, W., Choe, S., Unutmaz, D., Burkhart, M.,Dimarzio, P., Marmon, S., Sutton, R. E., Hill, C. M., Davis, C. B., Peiper,S. C., Schall, T. J., Littman, D. R., and Landau, N. R. (1996). Identifi-cation of a major co-receptor for primary isolates of HIV-1. Nature381(6584), 661–666.
Dorsky, D. I., Wells, M., and Harrington, R. D. (1996). Detection of HIV-1infection with a green fluorescent protein reporter system. J. Acquir.Immune Defic. Syndr. Hum. Retrovirol. 13(4), 308–313.
Dragic, T., Litwin, V., Allaway, G. P., Martin, S. R., Huang, Y. X., Na-gashima, K. A., Cayanan, C., Maddon, P. J., Koup, R. A., Moore, J. P.,and Paxton, W. A. (1996). HIV-1 entry into CD4(1) cells is mediated bythe chemokine receptor CC-CKR-5. Nature 381(6584), 667–673.
Edinger, A. L., Amedee, A., Miller, K., Doranz, B. J., Enders, M., Sharron,M., Samson, M., Lu, Z.-H., Clements, J. E., Murphey-Corb, M., Peiper,S. C., Parmentier, M., Broder, C. C., and Doms, R. W. (1997). Differ-ential utilization of CCR5 by macrophage and T cell tropic simainimmunodeficiency virus strains. Proc. Natl. Acad. Sci. USA 94, 4005–4010.
Edinger, A. L., Clements, J. E., and Doms, R. W. (1999). Chemokine andorphan receptors in HIV-2 and SIV tropism and pathogenesis. Virol-ogy 260, 211–221.
Edinger, A. L., Hoffman, T. L., Sharron, M., Lee, B., O’Dowd, B., andDoms, R. W. (1998a). Use of GPR1, GPR15, and STRL33 as corecep-tors by diverse human immunodeficiency virus type 1 and simianimmunodeficiency virus envelope proteins. Virology 249, 367–378.
Edinger, A. L., Hoffman, T. L., Sharron, M., Lee, B., Yi, Y., Choe, W.,Kolson, D. L., Mitrovic, B., Zhou, Y., Faulds, D., Collman, R. G.,Hesselgesser, J., Horuk, R., and Doms, R. W. (1998b). An orphanseven-transmembrane domain receptor expressed widely in thebrain functions as a coreceptor for human immunodeficiency virustype 1 and simian immunodeficiency virus. J. Virol. 72(10), 7934–7940.
Endres, M. J., Clapham, P. R., Marsh, M., Ahuja, M., Turner, J. D.,McKnight, A., Thomas, J. F., Stoebenau-Haggarty, B., Choe, S., Vance,P. J., Wells, T. N., Power, C. A., Sutterwala, S. S., Doms, R. W., Landau,N. R., and Hoxie, J. A. (1996). CD4-independent infection by HIV-2 ismediated by fusin/CXCR4. Cell 87(4), 745–756.
Farzan, M., Choe, H., Martin, K., Marcon, L., Hofmann, W., Karlsson, G.,Sun, Y., Barrett, P., Marchand, N., Sullivan, N., Gerard, N., Gerard, C.,and Sodroski, J. (1997). Two orphan seven-transmembrane segmentreceptors which are expressed in CD4-positive cells support simianimmunodeficiency virus infection. J. Exp. Med. 86(3), 405–411.
Feng, Y., Broder, C. C., Kennedy, P. E., and Berger, E. A. (1996). Hiv-1entry cofactor—Functional cDNA cloning of a seven-transmem-brane, G protein-coupled receptor. Science 272(5263), 872–877.
Fenyo, E. M., Morfeldt-Månson, L., Chiodi, F., Lind, A., von Gegerfelt, A.,Albert, J., Olausson, E., and Åsjo, B. (1988). Distinct replicative andcytopathic characteristics of human immunodeficiency virus isolates.J. Virol. 62(11), 4414–4419.
Heath, H., Qin, S., Rao, P., Wu, L., LaRosa, G., Kassam, N., Ponath, P. D.,and Mackay, C. R. (1997). Chemokine receptor usage by humaneosinophils. The importance of CCR3 demonstrated using an antag-onistic monoclonal antibody. J. Clin. Invest. 99(2), 178–184.
Hoffman, T. L., Stephens, E. B., Narayan, O., and Doms, R. W. (1998). HIV
type I envelope determinants for use of the CCR2b, CCR3, STRL33,and APJ coreceptors. Proc. Natl. Acad. Sci. USA 95(19), 11360–11365.
11HIV-1 CORECEPTOR USE IN THE GHOST(3) CELL ASSAY
Kimpton, J., and Emerman, M. (1992). Detection of replication-compe-tent and pseudotyped human immunodeficiency virus with a sensi-tive cell line on the basis of activation of an integrated beta-galac-tosidase gene. J. Virol. 66(4), 2232–2239.
Lee, B., Sharron, M., Montaner, L. J., Weissman, D., and Doms, R. W.(1999). Quantification of CD4, CCR5, and CXCR4 levels on lympho-cyte subsets, dendritic cells, and differentially conditioned mono-cyte-derived macrophages. Proc. Natl. Acad. Sci. USA 96(9), 5215–5220.
Marcon, L., Choe, H., Martin, K. A., Farzan, M., Ponath, P. D., Wu, L. J.,Newman, W., Gerard, N., Gerard, C., and Sodroski, J. (1997). Utiliza-tion of C-C chemokine receptor 5 by the envelope glycoproteins of apathogenic simian immunodeficiency virus, SIVmac239. J. Virol.71(3), 2522–2527.
Matloubian, M., David, A., Engel, S., Ryan, J. E., and Cyster, J. G. (2000).A transmembrane CXC chemokine is a ligand for HIV-coreceptorBonzo. Nat. Immunol. 1(4), 298–304.
Means, R. E., Greenough, T., and Desrosiers, R. C. (1997). Neutralizationsensitivity of cell culture-passaged simian immunodeficiency virus.J. Virol. 71(10), 7895–7902.
Merzouki, A., Patel, P., Cassol, S., Ennaji, M., Tailor, P., Turcotte, F. R.,O’Shaughnessy, M., and Arella, M. (1995). HIV-1 gp120/160 express-ing cells upregulate HIV-1 LTR directed gene expression in a cell linetransfected with HIV-1 LTR-reporter gene constructs. Cell. Mol. Biol.41(3), 445–452.
Morner, A., Bjorndal, Å., Albert, J., KewalRamani, V., Littman, D. R., Inoue,R., Thorstensson, R., Fenyo, E. M., and Bjorling, E. (1999). Primaryhuman immunodeficiency virus type 2 (HIV-2) isolates, like HIV-1isolates, frequently use CCR5 but show promiscuity in coreceptorusage. J. Virol. 73(3), 2343–2349.
Rudensey, L. M., Kimata, J. T., Long, E. M., Chackerian, B., and Over-baugh, J. (1998). Changes in the extracellular envelope glycoproteinof variants that evolve during the course of simian immunodeficiencyvirus SIVMne infection affect neutralizing antibody recognition, syn-cytium formation, and macrophage tropism but not replication, cyto-pathicity, or CCR-5 coreceptor recognition. J. Virol. 72(1), 209–217.
Scarlatti, G., Tresoldi, E., Bjorndal, Å., Fredriksson, R., Colognesi, C.,Deng, H., Malnati, M. S., Plebani, A., Siccardi, A. G., Littman, D. R.,and Lusso, P. (1997). In vivo evolution of HIV-1 coreceptor usage andsensitivity to chemokine-mediated suppression. Nat. Med. 3, 1259–
1265.Schols, D., Struyf, S., Van Damme, J., Este, J. A., Henson, G., and De
Clercq, E. (1997). Inhibition of T-tropic HIV strains by selective an-tagonization of the chemokine receptor CXCR4. J. Exp. Med. 186(8),1383–1388.
Schwartz, O., Virelizier, J. L., Montagnier, L., and Hazan, U. (1990). Amicrotransfection method using the luciferase-encoding reportergene for the assay of human immunodeficiency virus LTR promoteractivity. Gene 88(2), 197–205.
Sundqvist, V. A., Albert, J., Ohlsson, E., Hinkula, J., Fenyo, E. M., andWahren, B. (1989). Human immunodeficiency virus type 1 p24 pro-duction and antigenic variation in tissue culture of isolates withvarious growth characteristics. J. Med. Virol. 29, 170–175.
Tersmette, M., De Goede, R. E. Y., Al, B. J., Winkel, I., Gruters, B.,Cuypers, H. T., Huisman, H. G., and Miedema, F. (1988). Differentialsyncytium-inducing capacity of human immunodeficiency virus iso-lates: Frequent detection of syncytium-inducing isolates in patientswith acquired immunodeficiency syndrome (AIDS) and AIDS-relatedcomplex. J. Virol. 62(6), 2026–2032.
Tersmette, M., Lange, J. M. A., De Goede, R. E. Y., De Wolf, F., Eeftink-Schattenkerk, J. K. M., Coutinho, R. A., Huisman, J. G., Goudsmit, J.,and Miedema, F. (1989). Association between biological properties ofhuman immunodeficiency virus variants and risk for AIDS and AIDSmortality. Lancet 1, 983–985.
Tscherning-Casper, C., Vodros, D., Menu, E., Aperia, K., Fredriksson, R.,Dolcini, G., Chaouat, G., Barre-Sinoussi, F., Albert, J., and Fenyo, E. M.(2000a). Coreceptor usage of HIV-1 isolates representing differentgenetic subtypes obtained from pregnant Cameroonian women. Eu-ropean Network for in Utero Transmission of HIV-1. J. Acquir. ImmuneDefic. Syndr. 24(1), 1–9.
Tscherning-Casper, C., Dolcini, G., Mauclere, P., Fenyo, E.M., Barre-Sinoussi, F., Albert, J., and Menu, E. (2000b). Evidence of the exis-tence of a new circulating recombinant form of HIV type 1 subtype A/Jin Cameroon. The European Network on the Study of in UteroTransmission of HIV-1. AIDS Res. Hum. Retroviruses 16(13), 1313–1318.
Vodros, D., Thorstensson, R., Biberfeld, G., Schols, D., De Clercq, E.,and Fenyo, E. M. (2001). Coreceptor usage of sequential isolatesfrom cynomolgus monkeys experimentally infected with simian im-munodeficiency virus (SIVsm). Virology 290, 12–21.
Weber, J., Fenyo, E. M., Nara, P., Beddows, S., Kaleebu, P., Bjorndal, Å.,and the WHO Network for HIV Isolation and Characterization. (1996).
Neutralisation serotypes of HIV-1 field isolates are not predicted bygenetic subtype. J. Virol. 70(11), 7827–7832.
Related Documents











