Plant Physiol. (1984) 74, 856-861 0032-0889/84/74/0856/06/$0 1.00/0 Purification and Properties of Spinach Leaf Debranching Enzyme' Received for publication July 1, 1983 and in revised form Novermber 18, 1983 ISABELLA LUDWIG, PAUL ZIEGLER, AND ERWIN BECK* Lehrstuhl Pflanzenphysiologie, Universitat Bayreuth, D-8580 Bayreuth, West Germany ABSTRACT Starch debranching enzyme was purified from intact spinach (Spinacia okracea L. cv Vital) chloroplasts and from a spinach leaf extract using affinity chromatography on Sepharose 6B-bound cycloheptaamylose (Schardinger,B-dextrin). The enzyme from both sources was homogene- ous upon sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Spinach leaf debranching enzyme appears to consist of a single polypep- tide chain, since the molecular weight of the native protein (110,000 daltons) was not changed by treatment with sodium dodecyl sulfate. Only one spinach leaf debranching enzyme band could be detected after elec- trophoresis of a leaf extract on amylopectin-containing polyacrylamide gel, the retardation factor of which coincided with that of the single band seen with the chloroplast enzyme. The purified enzyme exhibited strong pullulanase activity, the specific activity being 69 units per milligram protein with pullulan and 22 units per milligram protein with amylopectin. Cycloheptnamylose is a potent competitive inhibitor of spinach leaf debranching enzyme. The pH optimum of the enzyme was found to be 5.5. The purified enzyme is rather unstable at both 200 and 0°C. Part of the activity lost under storage or at a suboptimal pH could immediately be restored by the addition of thiols. The reactivatable protein, being of the same molecular weight as the native enzyme, exhibited a somewhat altered electrophoretic mobility resulting in one or two minor bands on a zymogram. In contrast to the enzymes of assimilatory starch formation, which are located exclusively in the chloroplast, the enzymes of starch degradation have been found in the cytosol as well as in the chloroplasts (1 1, 14). Their activities in the former compart- ment usually exceed those in the plastids by far (13). The chloroplasts of at least spinach have been shown to contain all three types of amylases (endo-, exoamylase, and the debranching enzyme [1]) as well as phosphorylase (17). It has been demon- strated that amylolysis can act as pacemaker for the final phos- phorolytic degradation of assimilatory starch (1). Whereas the phosphorylases have been purified to homogeneity (14, 16), pure chloroplastic amylases could not be obtained by the classical separation techniques. Affinity chromatography with inhibitors (4) or substrate ana- logs ( 15, 19) has been successfully applied to the purification of amylases from various non-leaf sources. These findings suggest that leaf amylases, too, could be isolated according to such techniques. The present communication describes the purifica- tion of the starch debranching enzyme from spinach leaves and isolated chloroplasts and reports some of the properties of this enzyme. 'Supported by the Deutsche Forschungsgemeinschaft Grant Be 473/ 10 and 13/2. MATERIALS AND METHODS Preparation of Leaf Extract. Deribbed spinach leaves (Spina- cia oleracea L. cv Vital; 820 g) were cut into small pieces and homogenized in 1,500 ml 50 mm sodium acetate buffer (pH 6.0) with a Multimix. The homogenate was then centrifuged at 16,300g for 30 min at 0°C. Solid (NH4)2 S04 was added to the supematant and the precipitate which formed between 40 and 50% saturation was used for further purification of the enzyme. The precipitated protein was dissolved in 20 to 30 ml of acetate buffer (as described above) and dialyzed for 12 h against bidis- tilled H20. Prior to affinity chromatography, the solution was freed from precipitated protein by another centrifugation step (45,000g for 30 min). Preparation of Chloroplast Extract. Intact chloroplasts were isolated from spinach leaves using the Mes and Hepes buffers 'A' and 'B' as described by Jensen and Bassham (7). The whole procedure was performed at 0°C. Deribbed spinach leaves (250 g) were homogenized in buffer A for 30 s with an Ultraturrax. The homogenate was filtered through eight layers of gauze and centrifuged for 2 min at 2,600g. The precipitate was suspended in 120 ml buffer B and cells and cell debris were removed by centrifugation at 30g (twice). From the supernatant, the chloro- plasts were harvested at 750g (1 min) and washed once with the same buffer. The intact chloroplasts were then disrupted by suspension in 40 ml bidistilled H20. The membranes were subsequently removed by centrifugation at 45,000g (30 min). Preparation of Sepharose-Bound Cycloheptaamylose. The gel for affinity chromatography was prepared according to the method of Vretblad (18). However, j3-cyclodextrin (cycloheptaa- mylose, Sigma) was used instead of a-cyclodextrin. Epoxy-acti- vated Sepharose 6B was obtained from Sigma. Prior to applica- tion of the sample, the gel was equilibrated with 50 mm sodium acetate buffer, pH 6.0. The column size was 1.5 x 4.3 cm. Affinity Chromatography of Spinach Debranching Enzyme. The column was placed in a cold room and loaded with leaf or chloroplast extract equivalent to about 500 mg of protein and washed with 50 mM sodium acetate buffer (pH 6.0) until the effluent was free of material absorbing at 280 nm. Subsequently, the same buffer containing 1 mg (leaf extract enzyme) or 10 mg cycloheptaamylose (chloroplast enzyme) per ml was applied to release the bound protein. Fractions of 2.25 ml were collected at a flow rate of the elution buffer of 17 ml h-'. The debranching enzyme-containing fractions were combined and cycloheptaa- mylose was removed by filtration over a Sephadex G-25 medium column (2.5 x 20 cm). Assay of Enzyme Activity. Debranching activity was assayed in a stop time assay by measurement of the produced reducing groups (as maltose units) with dinitrosalicylic acid reagent (2). Pullulan was routinely used as substrate. The reaction mixture for glucan hydrolysis contained in a volume of 1 ml: sodium citrate buffer (60 mm, pH 6.0), pullulan (3 mg), and enzyme (0.02-0.1 ml). After 30 min at 370C, the reaction was stopped 856 www.plantphysiol.org on July 25, 2018 - Published by Downloaded from Copyright © 1984 American Society of Plant Biologists. All rights reserved.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Plant Physiol. (1984) 74, 856-8610032-0889/84/74/0856/06/$0 1.00/0
Purification and Properties of Spinach Leaf DebranchingEnzyme'
Received for publication July 1, 1983 and in revised form Novermber 18, 1983
ISABELLA LUDWIG, PAUL ZIEGLER, AND ERWIN BECK*Lehrstuhl Pflanzenphysiologie, Universitat Bayreuth, D-8580 Bayreuth, West Germany
ABSTRACT
Starch debranching enzyme was purified from intact spinach (Spinaciaokracea L. cv Vital) chloroplasts and from a spinach leaf extract usingaffinity chromatography on Sepharose 6B-bound cycloheptaamylose(Schardinger,B-dextrin). The enzyme from both sources was homogene-ous upon sodium dodecyl sulfate-polyacrylamide gel electrophoresis.Spinach leaf debranching enzyme appears to consist of a single polypep-tide chain, since the molecular weight of the native protein (110,000daltons) was not changed by treatment with sodium dodecyl sulfate. Onlyone spinach leaf debranching enzyme band could be detected after elec-trophoresis of a leaf extract on amylopectin-containing polyacrylamidegel, the retardation factor of which coincided with that of the single bandseen with the chloroplast enzyme. The purified enzyme exhibited strongpullulanase activity, the specific activity being 69 units per milligramprotein with pullulan and 22 units per milligram protein with amylopectin.Cycloheptnamylose is a potent competitive inhibitor of spinach leafdebranching enzyme. The pH optimum of the enzyme was found to be5.5. The purified enzyme is rather unstable at both 200 and 0°C. Part ofthe activity lost under storage or at a suboptimal pH could immediatelybe restored by the addition of thiols. The reactivatable protein, being ofthe same molecular weight as the native enzyme, exhibited a somewhataltered electrophoretic mobility resulting in one or two minor bands on azymogram.
In contrast to the enzymes of assimilatory starch formation,which are located exclusively in the chloroplast, the enzymes ofstarch degradation have been found in the cytosol as well as inthe chloroplasts (1 1, 14). Their activities in the former compart-ment usually exceed those in the plastids by far (13). Thechloroplasts of at least spinach have been shown to contain allthree types of amylases (endo-, exoamylase, and the debranchingenzyme [1]) as well as phosphorylase (17). It has been demon-strated that amylolysis can act as pacemaker for the final phos-phorolytic degradation of assimilatory starch (1). Whereas thephosphorylases have been purified to homogeneity (14, 16), purechloroplastic amylases could not be obtained by the classicalseparation techniques.
Affinity chromatography with inhibitors (4) or substrate ana-logs ( 15, 19) has been successfully applied to the purification ofamylases from various non-leaf sources. These findings suggestthat leaf amylases, too, could be isolated according to suchtechniques. The present communication describes the purifica-tion of the starch debranching enzyme from spinach leaves andisolated chloroplasts and reports some of the properties of thisenzyme.
'Supported by the Deutsche Forschungsgemeinschaft Grant Be 473/10 and 13/2.
MATERIALS AND METHODS
Preparation of Leaf Extract. Deribbed spinach leaves (Spina-cia oleracea L. cv Vital; 820 g) were cut into small pieces andhomogenized in 1,500 ml 50 mm sodium acetate buffer (pH 6.0)with a Multimix. The homogenate was then centrifuged at16,300g for 30 min at 0°C. Solid (NH4)2 S04 was added to thesupematant and the precipitate which formed between 40 and50% saturation was used for further purification of the enzyme.The precipitated protein was dissolved in 20 to 30 ml of acetatebuffer (as described above) and dialyzed for 12 h against bidis-tilled H20. Prior to affinity chromatography, the solution wasfreed from precipitated protein by another centrifugation step(45,000g for 30 min).
Preparation of Chloroplast Extract. Intact chloroplasts wereisolated from spinach leaves using the Mes and Hepes buffers'A' and 'B' as described by Jensen and Bassham (7). The wholeprocedure was performed at 0°C. Deribbed spinach leaves (250g) were homogenized in buffer A for 30 s with an Ultraturrax.The homogenate was filtered through eight layers of gauze andcentrifuged for 2 min at 2,600g. The precipitate was suspendedin 120 ml buffer B and cells and cell debris were removed bycentrifugation at 30g (twice). From the supernatant, the chloro-plasts were harvested at 750g (1 min) and washed once with thesame buffer. The intact chloroplasts were then disrupted bysuspension in 40 ml bidistilled H20. The membranes weresubsequently removed by centrifugation at 45,000g (30 min).
Preparation of Sepharose-Bound Cycloheptaamylose. The gelfor affinity chromatography was prepared according to themethod of Vretblad (18). However, j3-cyclodextrin (cycloheptaa-mylose, Sigma) was used instead of a-cyclodextrin. Epoxy-acti-vated Sepharose 6B was obtained from Sigma. Prior to applica-tion of the sample, the gel was equilibrated with 50 mm sodiumacetate buffer, pH 6.0. The column size was 1.5 x 4.3 cm.
Affinity Chromatography of Spinach Debranching Enzyme.The column was placed in a cold room and loaded with leaf orchloroplast extract equivalent to about 500 mg of protein andwashed with 50 mM sodium acetate buffer (pH 6.0) until theeffluent was free of material absorbing at 280 nm. Subsequently,the same buffer containing 1 mg (leaf extract enzyme) or 10 mgcycloheptaamylose (chloroplast enzyme) per ml was applied torelease the bound protein. Fractions of 2.25 ml were collected ata flow rate of the elution buffer of 17 ml h-'. The debranchingenzyme-containing fractions were combined and cycloheptaa-mylose was removed by filtration over a Sephadex G-25 mediumcolumn (2.5 x 20 cm).Assay of Enzyme Activity. Debranching activity was assayed
in a stop time assay by measurement of the produced reducinggroups (as maltose units) with dinitrosalicylic acid reagent (2).Pullulan was routinely used as substrate. The reaction mixturefor glucan hydrolysis contained in a volume of 1 ml: sodiumcitrate buffer (60 mm, pH 6.0), pullulan (3 mg), and enzyme(0.02-0.1 ml). After 30 min at 370C, the reaction was stopped
856 www.plantphysiol.orgon July 25, 2018 - Published by Downloaded from Copyright © 1984 American Society of Plant Biologists. All rights reserved.
SPINACH LEAF DEBRANCHING ENZYME
.2 40- * O(-4)Amylase- enzyme-activity.0activity,
30-
20-
10-
0
0 10 20 30 40
Fractions
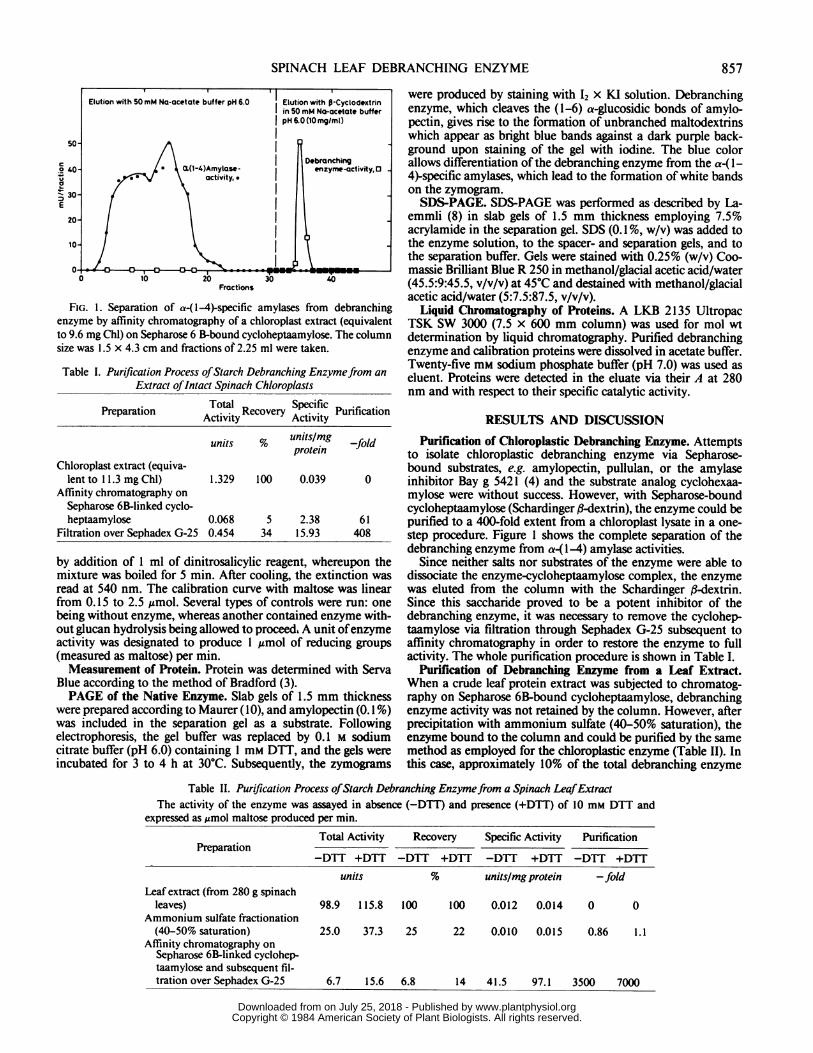
FIG. 1. Separation of a-(1-4)-specific amylases from debranchingenzyme by affinity chromatography of a chloroplast extract (equivalentto 9.6 mg Chl) on Sepharose 6 B-bound cycloheptaamylose. The columnsize was 1.5 x 4.3 cm and fractions of 2.25 ml were taken.
Table I. Purification Process ofStarch Debranching Enzymefrom an
Extract ofIntact Spinach ChloroplastsTotal SpecificPreparation Toty Recovery Acivic PurificationActivity Activity
units % units/mg -foldproteinChloroplast extract (equiva-
lent to 11.3 mg Chl) 1.329 100 0.039 0Affinity chromatography on
Sepharose 6B-linked cyclo-heptaamylose 0.068 5 2.38 61
Filtration over Sephadex G-25 0.454 34 15.93 408
by addition of 1 ml of dinitrosalicylic reagent, whereupon themixture was boiled for 5 min. After cooling, the extinction wasread at 540 nm. The calibration curve with maltose was linearfrom 0.15 to 2.5 gmol. Several types of controls were run: onebeing without enzyme, whereas another contained enzyme with-out glucan hydrolysis being allowed to proceed, A unit ofenzymeactivity was designated to produce 1 tmol of reducing groups(measured as maltose) per min.Measurement of Protein. Protein was determined with Serva
Blue according to the method of Bradford (3).PAGE of the Native Enzyme. Slab gels of 1.5 mm thickness
were prepared according to Maurer (10), and amylopectin (0.1 %)was included in the separation gel as a substrate. Followingelectrophoresis, the gel buffer was replaced by 0.1 M sodiumcitrate buffer (pH 6.0) containing 1 mm DTT, and the gels wereincubated for 3 to 4 h at 30C. Subsequently, the zymograms
were produced by staining with I2 X KI solution. Debranchingenzyme, which cleaves the (1-6) a-glucosidic bonds of amylo-pectin, gives rise to the formation of unbranched maltodextrinswhich appear as bright blue bands against a dark purple back-ground upon staining of the gel with iodine. The blue colorallows differentiation of the debranching enzyme from the a-( 1-
4)-specific amylases, which lead to the formation of white bandson the zymogram.SDS-PAGE. SDS-PAGE was performed as described by La-
emmli (8) in slab gels of 1.5 mm thickness employing 7.5%acrylamide in the separation gel. SDS (0.1%, w/v) was added tothe enzyme solution, to the spacer- and separation gels, and tothe separation buffer. Gels were stained with 0.25% (w/v) Coo-massie Brilliant Blue R 250 in methanol/glacial acetic acid/water(45.5:9:45.5, v/v/v) at 45C and destained with methanol/glacialacetic acid/water (5:7.5:87.5, v/v/v).
Liquid Chromatography of Proteins. A LKB 2135 UltropacTSK SW 3000 (7.5 x 600 mm column) was used for mol wtdetermination by liquid chromatography. Purified debranchingenzyme and calibration proteins were dissolved in acetate buffer.Twenty-five mm sodium phosphate buffer (pH 7.0) was used aseluent. Proteins were detected in the eluate via their A at 280nm and with respect to their specific catalytic activity.
RESULTS AND DISCUSSIONPurification of Chloroplastic Debranching Enzyme. Attempts
to isolate chloroplastic debranching enzyme via Sepharose-bound substrates, e.g. amylopectin, pullulan, or the amylaseinhibitor Bay g 5421 (4) and the substrate analog cyclohexaa-mylose were without success. However, with Sepharose-boundcycloheptaamylose (Schardinger l-dextrin), the enzyme could bepurified to a 400-fold extent from a chloroplast lysate in a one-step procedure. Figure 1 shows the complete separation of thedebranching enzyme from a(1-4) amylase activities.
Since neither salts nor substrates of the enzyme were able todissociate the enzyme-cycloheptaamylose complex, the enzymewas eluted from the column with the Schardinger ,8-dextrin.Since this saccharide proved to be a potent inhibitor of thedebranching enzyme, it was necessary to remove the cyclohep-taamylose via filtration through Sephadex G-25 subsequent toaffinity chromatography in order to restore the enzyme to fullactivity. The whole purification procedure is shown in Table I.
Purification of Debranching Enzyme from a Leaf Extract.When a crude leaf protein extract was subjected to chromatog-raphy on Sepharose 6B-bound cycloheptaamylose, debranchingenzyme activity was not retained by the column. However, afterprecipitation with ammonium sulfate (40-50% saturation), theenzyme bound to the column and could be purified by the samemethod as employed for the chloroplastic enzyme (Table II). Inthis case, approximately 10% of the total debranching enzyme
Table II. Purification Process ofStarch Debranching Enzymefrom a Spinach LeafExtractThe activity of the enzyme was assayed in absence (-DTT) and presence (+DTT) of 10 mM DTT and
expressed as Mmol maltose produced per min.
Total Activity Recovery Specific Activity PurificationPreparation
-DTT +DTT -DTT +DTT -DTT +DTT -DTT +DTTunits % units/mg protein - fold
Leaf extract (from 280 g spinachleaves) 98.9 115.8 100 100 0.012 0.014 0 0
Ammonium sulfate fractionation(40-50% saturation) 25.0 37.3 25 22 0.010 0.015 0.86 1.1
Affinity chromatography onSepharose 6B-linked cyclohep-taamylose and subsequent fil-tration over Sephadex G-25 6.7 15.6 6.8 14 41.5 97.1 3500 7000
857
www.plantphysiol.orgon July 25, 2018 - Published by Downloaded from Copyright © 1984 American Society of Plant Biologists. All rights reserved.
Plant Physiol. Vol. 74, 1984
cl
1 2 3 4 5f
6
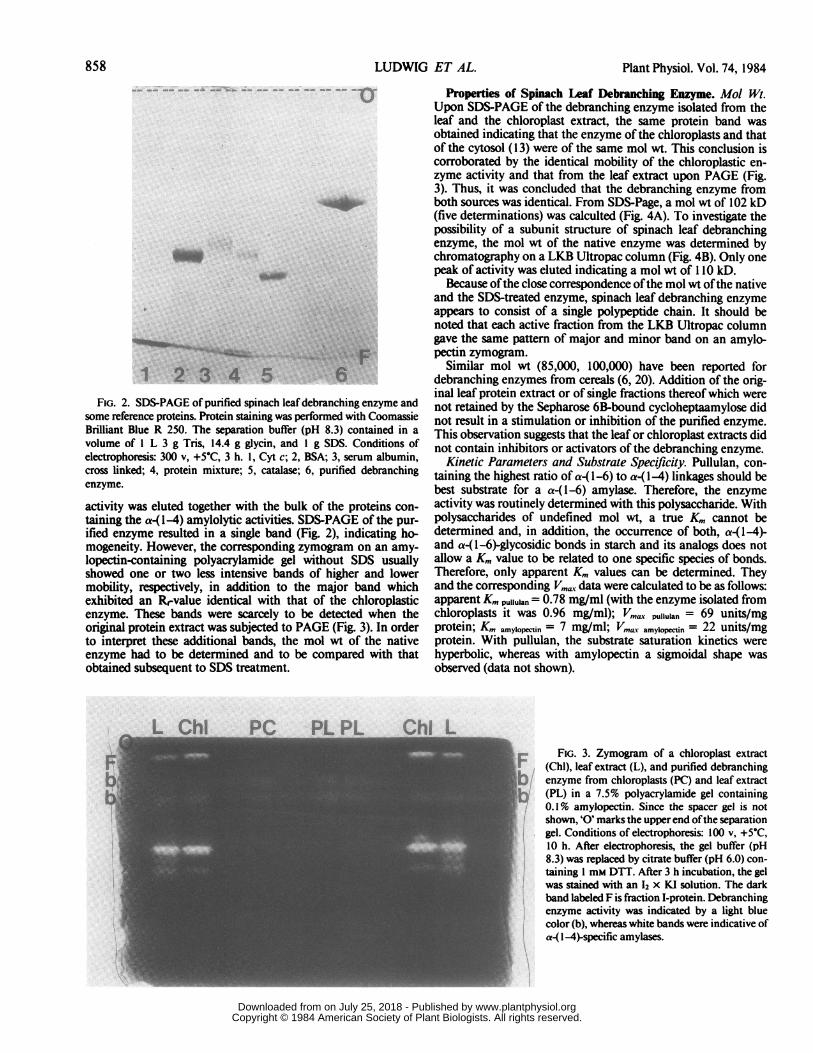
FIG. 2. SDS-PAGE of purified spinach leaf debranching enzyme andsome reference proteins. Protein staining was performed with CoomassieBrilliant Blue R 250. The separation buffer (pH 8.3) contained in a
volume of I L 3 g Tris, 14.4 g glycin, and I g SDS. Conditions ofelectrophoresis: 300 v, +5°C, 3 h. 1, Cyt c; 2, BSA; 3, serum albumin,cross linked; 4, protein mixture; 5, catalase; 6, purified debranchingenzyme.
activity was eluted together with the bulk of the proteins con-taining the a-1-4) amylolytic activities. SDS-PAGE of the pur-ified enzyme resulted in a single band (Fig. 2), indicating ho-mogeneity. However, the corresponding zymogram on an amy-lopectin-containing polyacrylamide gel without SDS usuallyshowed one or two less intensive bands of higher and lowermobility, respectively, in addition to the major band whichexhibited an R-value identical with that of the chloroplasticenzyme. These bands were scarcely to be detected when theoriginal protein extract was subjected to PAGE (Fig. 3). In orderto interpret these additional bands, the mol wt of the nativeenzyme had to be determined and to be compared with thatobtained subsequent to SDS treatment.
Properties of Spinach Leaf Debranching Enzyme. Mol Wt.Upon SDS-PAGE of the debranching enzyme isolated from theleaf and the chloroplast extract, the same protein band wasobtained indicating that the enzyme of the chloroplasts and thatof the cytosol (13) were of the same mol wt. This conclusion iscorroborated by the identical mobility of the chloroplastic en-zyme activity and that from the leaf extract upon PAGE (Fig.3). Thus, it was concluded that the debranching enzyme fromboth sources was identical. From SDS-Page, a mol wt of 102 kD(five determinations) was calculted (Fig. 4A). To investigate thepossibility of a subunit structure of spinach leaf debranchingenzyme, the mol wt of the native enzyme was determined bychromatography on a LKB Ultropac column (Fig. 4B). Only onepeak of activity was eluted indicating a mol wt of 1 10 kD.
Because ofthe close correspondence ofthe mol wt ofthe nativeand the SDS-treated enzyme, spinach leaf debranching enzymeappears to consist of a single polypeptide chain. It should benoted that each active fraction from the LKB Ultropac columngave the same pattern of major and minor band on an amylo-pectin zymogram.
Similar mol wt (85,000, 100,000) have been reported fordebranching enzymes from cereals (6, 20). Addition of the orig-inal leaf protein extract or of single fractions thereof which werenot retained by the Sepharose 6B-bound cycloheptaamylose didnot result in a stimulation or inhibition of the purified enzyme.This observation suggests that the leaf or chloroplast extracts didnot contain inhibitors or activators of the debranching enzyme.
Kinetic Parameters and Substrate Specificity. Pullulan, con-taining the highest ratio of a-1-6) to a-(14) linkages should bebest substrate for a a-(1-6) amylase. Therefore, the enzymeactivity was routinely determined with this polysaccharide. Withpolysaccharides of undefined mol wt, a true Km cannot bedetermined and, in addition, the occurrence of both, a-(14)-and a( 1-6)-glycosidic bonds in starch and its analogs does notallow a Km value to be related to one specific species of bonds.Therefore, only apparent Km values can be determined. Theyand the corresponding Vmax data were calculated to be as follows:apparent Km pullulan = 0.78 mg/ml (with the enzyme isolated fromchloroplasts it was 0.96 mg/ml); Vma. p.1 =1an 69 units/mgprotein; Km amylopecin = 7 mg/ml; Vmcv amylopecin = 22 units/mgprotein. With pullulan, the substrate saturation kinetics werehyperbolic, whereas with amylopectin a sigmoidal shape wasobserved (data not shown).
LFIG. 3. Zymogram of a chloroplast extract
F (Chl), leaf extract (L), and purified debranchingb enzyme from chloroplasts (PC) and leaf extractb (PL) in a 7.5% polyacrylamide gel containing
0.1% amylopectin. Since the spacer gel is notshown, 'O' marks the upper end ofthe separationgel. Conditions of electrophoresis: 100 v, +5C,10 h. After electrophoresis, the gel buffer (pH8.3) was replaced by citrate buffer (pH 6.0) con-taining I mm DTT. After 3 h incubation, the gelwas stained with an I2 x KI solution. The darkband labeled F is fraction I-protein. Debranchingenzyme activity was indicated by a light bluecolor (b), whereas white bands were indicative ofa-l-4)-specific amylases.
858 LUDWIG ET AL.
www.plantphysiol.orgon July 25, 2018 - Published by Downloaded from Copyright © 1984 American Society of Plant Biologists. All rights reserved.

SPINACH LEAF DEBRANCHING ENZYME
_________----as ncning ~ BA1nzyme BSA--
E Catclase (Subunit)cm Ovalbumir\
4.5- carbonianhydrase J cro e
4 91~~~~~~~~
4 0L2 0.40.6 0.8 20 40 50
RF Ftctio
FIG. 4. Determination of the mol wt of spinach leaf debranchingenzyme. A, R, values of debranching enzyme and calibration proteinsupon SDS-PAGE. Experimental details as in Figure 2. A mol wt of 102kD was calculated for the SDS-treated debranching enzyme. B, Liquidchromatography of purified debranching enzyme together with calibra-tion proteins on a LKB 2135 Ultropac TSK 3000 column. Debranchingenzyme activity was measured without (A) and in the presence (A) of 10mM DTT. The mol wt of the enzyme was calculated to be I 10 kD.
Table III. Substrate Specificity ofPurified Spinach LeafDebranchingEnzyme
The concentration of the substrates was 5 mg/ml. Degradation was
allowed to proceed for 30 min at 37°C and pH 6.0.
Substrate Relative Activity
Pullulan 100Soluble starch 37Amylopectin 17.5Amylose 0Glycogen type II (oyster) 0Glycogen type IX (bovine liver) 0Glycogen type X (rabbit liver) 0
Table IV. Activity ofPurified Spinach LeafDebranching Enzyme asInfluenced by Thiols
Thiols in the Specific Activity StimulationIncubation Mixture
units/mg protein -foldControl 20.2 0+ 10 mMDTT 84.8 4.2+ 20 mm DTT 95.5 4.7+ 10 mM GSH 67.4 3.3+ 20 mm GSH 66.5 3.3+ 10 mM mercaptoethanol 63.5 3.1+ 20 mM mercaptoethanol 71.9 3.6
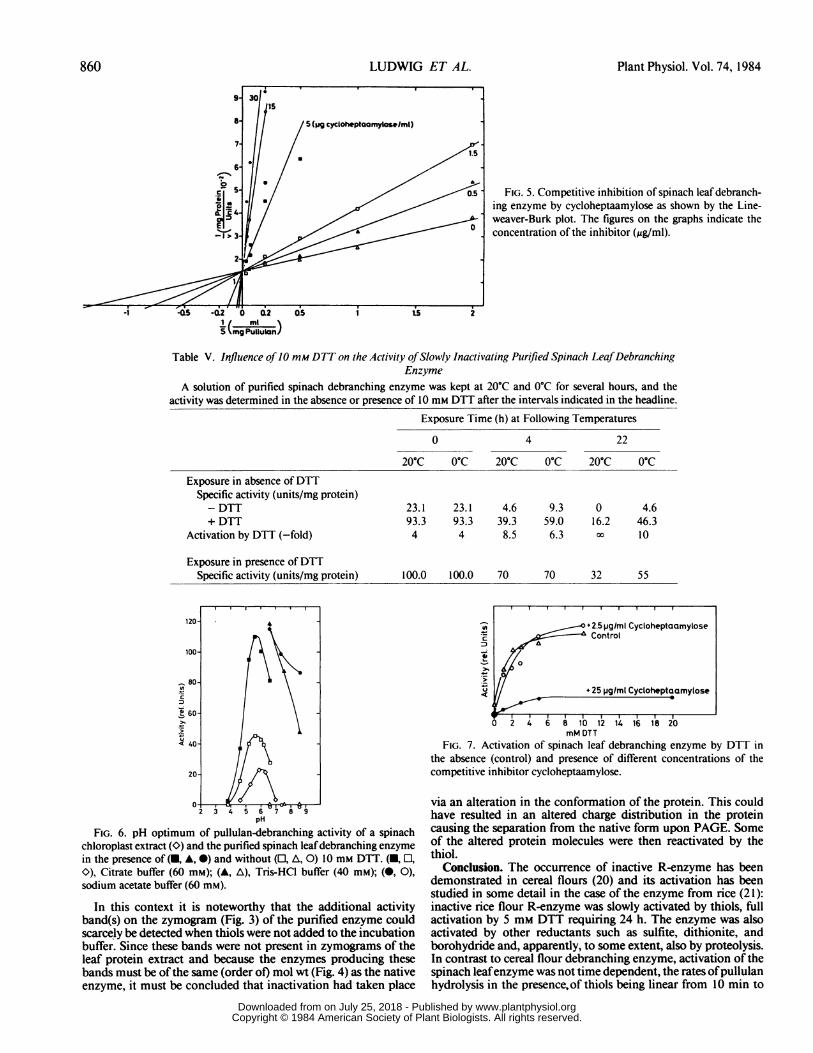
With various homoglucans as starch analogs, the spinach leafdebranching enzyme exhibited specificity as shown in Table III.Km and V values proved pullulan to be the most specificsubstrate. This has been observed also for rice seed R-enzyme(6), and an earlier report on a partially purified debranchingenzyme from spinach leaves (1 1) mentioned that only the so-called a-limit dextrins, which actually are branched oligosaccha-rides of a degree of polymerization of 6 to 7, were more readilyattacked than was pullulan. Spinach debranching enzyme isstrongly inhibited by cycloheptaamylose, whereas cyclohexoa-mylase was inhibitory only at concentrations of two orders ofmagnitude greater than that of the Schardinger (8-dextrin. Inhi-bition by cycloheptaamylose was competitive (Fig. 5). The K1was determined to be 0.0009 mg/ml. Competitive inhibition byboth cycloamyloses has also been reported for pullulanases fromvarious microorganisms (9).pH Optimum. With a protein extract from chloroplasts, pul-
lulan was optimally degraded at pH 6.0 (Fig. 6). However, withthe purified debranching enzyme, this optimum shifted to pH 5to 5.5. The difference of both optima could be explained by theadditional action of a-( 1-4) amylases in the chloroplast extracton the liberated maltotriose, which exhibit optimal reaction atpH 6.0. The purified enzyme from oat flour exhibited maximalactivity at pH 5.6 (20).
Stability of the Enzyme. Diluted solutions (0.15 Ag proteinml-') of the purified enzyme proved to be rather unstable. Atroom temperature, 80% of the activity was lost after 4 h and no
activity was found after 22 h. At O°C, the losses of activity were
lower but still significant: 60% after 4 h and 80% after 22 h.When kept frozen, the enzyme lost its activity within 1 week.
Stimulation of Spinach Debranching Enzyme by Thiols. Con-tradictory effects of thiols on debranching enzyme activity havebeen reported in the literature: a 4-to 5-fold activation was foundwith the enzyme from germinating rice seeds (21), whereas on
the other hand, Okita and Preiss (11) did not find a significanteffect of such compounds on chloroplastic amylases. Therefore,the effect of thiols was studied in more detail.
Addition of various thiols clearly increased the activity ofpurified spinach R-enzyme several-fold, the effect of a dithiolbeing greater than that of monothiols (Table IV).
This effect was different from that of thiols on the reducingpower of carbohydrates (5) which, under the conditions appliedin this work, resulted in an increase of extinction by a factor of2. To differentiate between the actual and the apparent activationof the enzyme, calibration curves with and without thiols were
established.The stimulatory effect of thiols showed saturation kinetics
(Fig. 7) similar to those reported for R-enzyme from rice flour(21). Since half-saturation of the thiol-specific effect did notdepend on the absence or presence or on the concentration ofthe inhibitor cycloheptaamylose, the stimulatory effect of DTTin the experiment shown in Figure 7 cannot be explained as a
reversal of the competitive inhibition exerted by the substrateanalog. In turn, this interpretation suggests that thiols do notinteract with the active site of spinach leaf debranching enzyme.
When the activating effect of DTT on the debranching enzyme
activity of several chloroplast extracts was compared (e.g. on a
chlorophyll basis), it became apparent that the degree of stimu-lation was inversely proportional to the initial activity. Similarly,the effect of 10 mm DTT, employed in the purification of theenzyme (Table II) was small with respect to the total leaf extract,but significant with respect to the purified enzyme. These obser-vations could be explained by the assumption that thiols are
capable of activating inactive enzyme, irrespective of the reasonsfor inactivity. For instance, the addition of DTT did not shiftthe pH optimum of debranching enzyme, but considerably ex-
tended the basis of the activity peak towards the alkaline region(Fig. 6). Thus, the stimulatory effect was small at the optimumpH, but dramatically increased above pH 6. Furthermore, whenthe purified enzyme was stored at 20°C or 0°C and a more or
less rapid inactivation was observed, part of the lost activitycould be restored by the addition of DTT to the assay mixture(Table V). Again, the activating effect was inversely proportionalto the activity of the enzyme. It should be emphasized that inthis case the effect ofthiols cannot be explained as a preservationofthe activity, but must be understood as a conversion ofalreadyinactive protein into an active form. Consequently, DTT alsoproved to be a potent stabilizing reagent for the enzyme activity(Table V). The concept of a reactivating effect of thiols couldalso explain the results shown in Figure 7. The activation factorof 9 found with the control indicated a considerable proportionof inactive enzyme in the preparation which was reactivated byDTT to such an extent that it could at least partly overcome theinhibition due to cycloheptaamylose.
859
www.plantphysiol.orgon July 25, 2018 - Published by Downloaded from Copyright © 1984 American Society of Plant Biologists. All rights reserved.
Plant Physiol. Vol. 74, 1984
/.FIG. 5. Competitive inhibition of spinach leaf debranch-
ing enzyme by cycloheptaamylose as shown by the Line-
weaver-Burk plot. The figures on the graphs indicate the
-1 1t3- ]concentration of the inhibitor (,ig/ml).
-a5 -0.2 0 02 05 1 21 mlS (mgPulluon)
Table V. Influence of 10 mM DTT on the Activity ofSlowly Inactivating Purified Spinach LeafDebranchingEnzyme
A solution of purified spinach debranching enzyme was kept at 20°C and OC for several hours, and theactivity was determined in the absence or presence of 10 mM DTT after the intervals indicated in the headline.
Exposure Time (h) at Following Temperatures
0 4 22
20°C OOC 200C 0°C 200C 0°C
Exposure in absence of DTTSpecific activity (units/mg protein)- DTT+ DTT
Activation by DTT (-fold)
Exposure in presence ofDTTSpecific activity (units/mg protein)
pH
FIG. 6. pH optimum of pullulan-debranching activity of a spinachchloroplast extract (O) and the purified spinach leafdebranching enzymein the presence of (l, A, 0) and without (0, A, 0) 10 mM DTT. (U, 0,
C), Citrate buffer (60 mM); (A, A), Tris-HCI buffer (40 mM); (-, 0),sodium acetate buffer (60 mM).
In this context it is noteworthy that the additional activityband(s) on the zymogram (Fig. 3) of the purified enzyme couldscarcely be detected when thiols were not added to the incubationbuffer. Since these bands were not present in zymograms of theleaf protein extract and because the enzymes producing thesebands must be ofthe same (order of) mol wt (Fig. 4) as the nativeenzyme, it must be concluded that inactivation had taken place
1 23.1 4.6 9.3 0 4.6.3 93.3 39.3 59.0 16.2 46.3
4 8.5 6.3 00 10
.0 100.0 70 70 32 55
o_ *2.5 pg/ml CycloheptaamyloseC: < Control
-0
.25 pg/mI Cycloheptoamylose
2 4 6 8 10 12 14 16 18 20mM DTT
FIG. 7. Activation of spinach leaf debranching enzyme by DTT inthe absence (control) and presence of different concentrations of thecompetitive inhibitor cycloheptaamylose.
via an alteration in the conformation of the protein. This couldhave resulted in an altered charge distribution in the proteincausing the separation from the native form upon PAGE. Someof the altered protein molecules were then reactivated by thethiol.
Conclusion. The occurrence of inactive R-enzyme has beendemonstrated in cereal flours (20) and its activation has beenstudied in some detail in the case of the enzyme from rice (21):inactive rice flour R-enzyme was slowly activated by thiols, fullactivation by 5 mM DTT requiring 24 h. The enzyme was alsoactivated by other reductants such as sulfite, dithionite, andborohydride and, apparently, to some extent, also by proteolysis.In contrast to cereal flour debranching enzyme, activation of thespinach leafenzyme was not time dependent, the rates ofpullulanhydrolysis in the presence,of thiols being linear from 10 min to
860 LUDWIG ET AL.
www.plantphysiol.orgon July 25, 2018 - Published by Downloaded from Copyright © 1984 American Society of Plant Biologists. All rights reserved.

SPINACH LEAF DEBRANCHING ENZYME
more than 5 h (data not shown). In addition, treatment of achloroplast extract with proteases (papain, trypsin) did not sig-nificantly increase the debranching enzyme activity and, after 3h incubation rather resulted in decrease of pullulanase activity.From these results, it is obvious that the effect of thiols onspinach leaf debranching enzyme is quite different from that oncereal R-enzymes. Therefore, it appears to be unlikely that thiolsare directly involved in the achievement of the diumal oscilla-tions exhibited by chloroplastic debranching enzyme (1, 12).
Acknowledgment-The authors would like to thank M. Hofmann for skillfultechnical assistance.
LITERATURE CITED
1. BECK E, P PONGRATZ, I REUTER 1981 The amylolytic system of isolatedspinach chloroplasts and its role in the breakdown of assimilatory starch. InG Akoyunoglou, ed, Proceedings of the Fifth International Congress onPhotosynthesis, Vol 4. Balaban International Science Services, Philadelphia,pp 529-538
2. BERNFELD P 1955 Amylases, a and i. Methods Enzymol 1: 149-1583. BRADFORD MM 1976 A rapid and sensitive method for quantitation of
microgram quantities of protein. Anal Biochem 72: 248-2544. BURRILL PH, PM BRANNON, N KRETCHMER 1981 A single-step purification of
rat pancreatic and salivary amylase by affinity chromatography. Anal Bio-chem 117: 402-405
5. CLAYTON JW, WOS MEREDITH 1966 The effect of thiols on the dinitrosalicylicacid test for reducing sugars. J Inst Brew 72: 537-540
6. IWAKI K, H FuwA 1981 Purification and some properties of debranchingenzyme ofgerminating rice endosperm. Agric Biol Chem 45: 2683-2688
7. JENSEN RG, JA BASSHAM 1966 Photosynthesis by isolated chloroplasts. Proc
Natl Acad Sci USA 56: 1095-1 1018. LAEMMLI UK 1970 Cleavage of structural proteins during the assembly of the
head of bacteriophage T4. Nature 227: 680-6859. MARSHALL JJ 1973 Inhibition of pullulanase by Schardinger dextrins. FEBS
Lett 37: 269-27310. MAURER HR 1971 Disc-Electrophoresis. Walter de Gruyter, Berlin, pp 22-4411. OKITA TW, J PREISS 1980 Starch degradation in spinach leaves. Isolation and
characterization of the amylases and R-enzyme of spinach leaves. PlantPhysiol 66:870-876
12. PONGRATZ P, E BECK 1978 Diurnal oscillation ofamylolytic activity in spinachchloroplasts. Plant Physiol 62: 687-689
13. PREISS J 1982 Regulation of the biosynthesis and degradation of starch. AnnuRev Plant Physiol 33: 431-454
14. PREISS J, TW OKITA, E GREENBERG 1980 Characterization of the spinach leafphosphorylases. Plant Physiol 66:864-869
15. SILVANOVICH MP, RD HILL 1976 Affinity chromatography ofcereal a-amylase.Anal Biochem 73: 430-433
16. STEUP M 1981 Purification of chloroplast al,4-glucan phosphorylase fromspinach leaves by chromatography on Sepharose-bound starch. BiochimBiophys Acta 659: 123-131
17. STEUP M, C SCHAECHTELE, E LATZKO 1980 Separation and partial characteri-zation of chloroplast and non-chloroplast a-glucan phosphorylases fromspinach leaves. Z Pflanzenphysiologie 96: 365-374
18. VRETBLAD P 1974 Immobilization of ligands for biospecific affinity chroma-tography via their hydroxyl groups. The cyclohexaamylose-fl-amylase system.FEBS Lett 47: 86-89
19. WESELAKE RJ, RD HILL 1982 Cycloheptaamylose as an affinity ligand ofcerealalpha amylase. Characteristics and possible mechanism of the interaction.Carbohydr Res 108: 153-161
20. YAMADA J 1981 Purification of oat debranching enzyme and occurrence ofinactive debranching enzyme in cereals. Agric Biol Chem 45: 1013-1015
21. YAMADA J 1981 Inactive debranching-enzyme in rice seeds, and its activation.Carbohydr Res 90: 153-157
861
www.plantphysiol.orgon July 25, 2018 - Published by Downloaded from Copyright © 1984 American Society of Plant Biologists. All rights reserved.
Related Documents











