PURIFICATION AND CHARACTERIZATION OF POLYPHENOL OXIDASE FROM POTATO: I. PURIFICATION AND PROPERTIES YONG K. CHO' and HYE K. AHN Department of Biochemistry Changwon National University Changwon, Kyungnam 641-773 S. Korea Received for Publication August 5, 1998 Accepted for Publication December 16, 1998 ABSTRACT Polyphenol oxidase (Isozyme I) from potato was extracted and purified with ammonium sulfate, cation-exchange (Bio-Rad Bio-Scale S2) and Sephadex G-I 00 column chromatography. The enzyme was purified 11.8 fold resulting in a spectfic activity of 250.3 unitdmg. Optimum pH of the enzyme was 6.6. Optimum temperature of the enzyme was 40C and its half-life was 0.8 min at 70C. The K, for catechol, pyrogallol, 4-methyl catechol, caffeic acid and L-DOPA were 4. I I mM, 0.61 mM, 0.78 mM, 0.50 mM and 32 mM, respectively. However, monophenols such as tyrosine,p-cresol and I-naphtol did not show any activity. Data for V,JKm which represents catalytic efficiency show that 4-methyl catechol has the highest value. The molecular weight of the active enzyme was 86,000 Da, composed of two identical subunits. The number of Cu2' ions bound wasfound to be 2 per enzyme molecule. INTRODUCTION Enzymatic browning is the result of polyphenol oxidase (PPO: monophenol, dihydroxy-L-phenylalanine: oxygen oxidoreductase; EC 1.14.18.1) catalyzed oxidation of mono- and diphenols to o-quinones as shown in Scheme I. Polyphenol oxidase is a mixed function oxidase that catalyzes both the ortho hydroxylation of monophenols to o-diphenols (cresolase activity) and the further oxidation o-diphenols to o-quinones (catecholase activity). The o-quinones 'Corresponding author: [email protected] Journal of Food Biochemistry 23 (1999) 577-592. All Rights Reserved. "Copyright 2000 by Food & Nutrition Press, Inc., Trumbull. Connecticut. 577

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

PURIFICATION AND CHARACTERIZATION OF POLYPHENOL OXIDASE FROM POTATO: I. PURIFICATION
AND PROPERTIES
YONG K. CHO' and HYE K. AHN
Department of Biochemistry Changwon National University Changwon, Kyungnam 641-773
S. Korea
Received for Publication August 5, 1998 Accepted for Publication December 16, 1998
ABSTRACT
Polyphenol oxidase (Isozyme I) from potato was extracted and purified with ammonium sulfate, cation-exchange (Bio-Rad Bio-Scale S2) and Sephadex G-I 00 column chromatography. The enzyme was purified 11.8 fold resulting in a spectfic activity of 250.3 unitdmg. Optimum pH of the enzyme was 6.6. Optimum temperature of the enzyme was 40C and its half-life was 0.8 min at 70C. The K, for catechol, pyrogallol, 4-methyl catechol, caffeic acid and L-DOPA were 4. I I mM, 0.61 mM, 0.78 mM, 0.50 mM and 32 mM, respectively. However, monophenols such as tyrosine, p-cresol and I-naphtol did not show any activity. Data for V,JKm which represents catalytic efficiency show that 4-methyl catechol has the highest value. The molecular weight of the active enzyme was 86,000 Da, composed of two identical subunits. The number of Cu2' ions bound was found to be 2 per enzyme molecule.
INTRODUCTION
Enzymatic browning is the result of polyphenol oxidase (PPO: monophenol, dihydroxy-L-phenylalanine: oxygen oxidoreductase; EC 1.14.18.1) catalyzed oxidation of mono- and diphenols to o-quinones as shown in Scheme I. Polyphenol oxidase is a mixed function oxidase that catalyzes both the ortho hydroxylation of monophenols to o-diphenols (cresolase activity) and the further oxidation o-diphenols to o-quinones (catecholase activity). The o-quinones
'Corresponding author: [email protected]
Journal of Food Biochemistry 23 (1999) 577-592. All Rights Reserved. "Copyright 2000 by Food & Nutrition Press, Inc., Trumbull. Connecticut. 577

578 Y.K. CHO and H.K. AHN
Scheme I. Pathway of Polyphenoloxidase-Catalyzed Reaction
are highly reactive compounds and can polymerize spontaneously to form brown pigment (melanin), or react with amino acids and proteins (Joslyn and Pointing 1951; Mayer and Hare1 1979; Vamos-Vigyazo 1981).
Enzymatic browning of raw agro-materials is a major problem in the food industry and one of the main causes of quality loss during postharvest handling and processing (Mathew and Parpia 1971). It results in unpleasant appearance and the concomitant development of off flavors. To prevent enzymatic browning reaction, many techniques have been tried: use of reducing agents (Ingraham 1956; Scharf and Dawson 1958; Golan-Goldhirsh et al. 1984), chelating agents (Gardner et aZ. 199 I), complexing agents (McEvily et al. 1992), acidulants (Aylward and Haisman 1969) as well as enzyme inhibitors (Saruno et al. 1979; Otwell et aZ. 199 1 ; Iyenga et al. 1991) and enzyme treatment (Finkle and Nelson 1963; Kelly and Finkle 1969).
There have been many studies on the purification of polyphenol oxidases from microorganisms (Bull and Carter 1973; Gutterigde and Robb 1975), fruits (Stelzig et al. 1977; Kahn 1977; Flurkey and Jen 1978; Das et al. 1997) and plants (Patil and Zucker 1965; Gregory and Bendall 1966; Vaughan et al. 1975; Oda el al. 1989). However, there are few studies on the catalytic mechanism of polyphenol oxidase. In order to understand catalytic mechanism of polyphenol oxidase from potato as a standard model, purification of the enzyme from potato was attempted. In this study, we report purification and some properties of polyphenol oxidase (Isozyme I) from potato.
MATERIALS AND METHODS
Materials
Catechol, 4-methyl catechol, pyrogallol, caffeic acid, L-3,4-dihydroxy phenylalanine (L-DOPA), tyrosine, 1-naphtol and p-cresol were purchased from Sigma Chemical Co. (USA). Sephadex G-100 and G-150 were from Pharmacia Chemicals (Sweden). Bio-Scale S2 column was from Bio-Rad (USA). SDS- polyacrylamide electrophoresis and gel filtration molecular weight markers were from Boehringer Mannheim (Germany). All other chemicals and buffers were

I. PURIFICATION AND PROPERTIES 579
obtained from commercially available sources and were of the highest quality available.
Enzyme Extraction
Freshly harvested potatoes from Kimhae, Kyungnam were washed and hand peeled. A 500 g sample was cut with 5 mm width and length. Then, it was homogenized with 500 mL of cold 50 mM Mops buffer (pH 6.6) with 20 mM EDTA, 10 mM cysteine and 0.35 M NaCl for 2 min in Waring blender (Waring Product Co, USA). The homogenate was kept at 4C for 1 h and filtered. The filrate was centrifuged at 10,000 g for 30 rnin at 4C twice. The supernatant was collected and used as a crude enzyme extract. The crude enzyme was brought to 30% ammonium sulfate saturation with solid ammonium sulfate with continuous stirring at 4C. After 6 h, the solution was centrifuged at 13,000 g for 1 h at 4C. The supernatant was brought to 80% ammonium sulfate saturation and, after 6 h, Centrifuged at 13,000 g for 1 h. The precipitate was dissolved in a minimal volume of 50 mM Mops buffer (PH 6.6) containing 20 mM EDTA. The salt was removed by dialysis (dialysis membrane: SpectraPor M W cut 6000-8000) in the same buffer for 12 h with 6 changes of the buffer at 4C. The dialyzed enzyme was stored at - 80C before further purification.
Chromatography on Bio-Scale S2
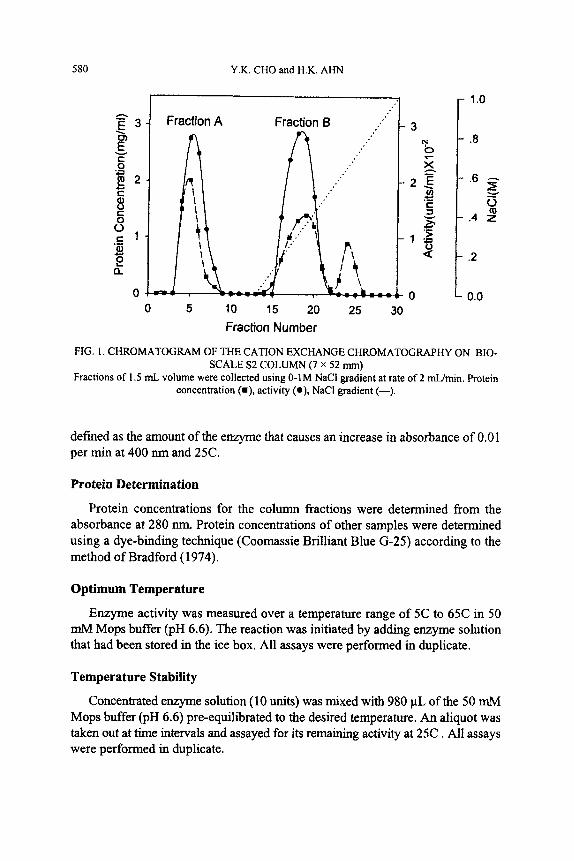
The dialyzed enzyme was injected into a Bio-Scale S2 cation exchange column which was equilibrated with 50 mM Mops buffer (pH 6.6) of the Bio-Rad Automated Protein Purification System 8000T (Bio-Rad, USA). The protein was eluted using 0-1 M NaCl gradient at the rate of 2 mL/min. Fractions of 1.5 mL volume were collected.
Chromatography on Sephadex G-100
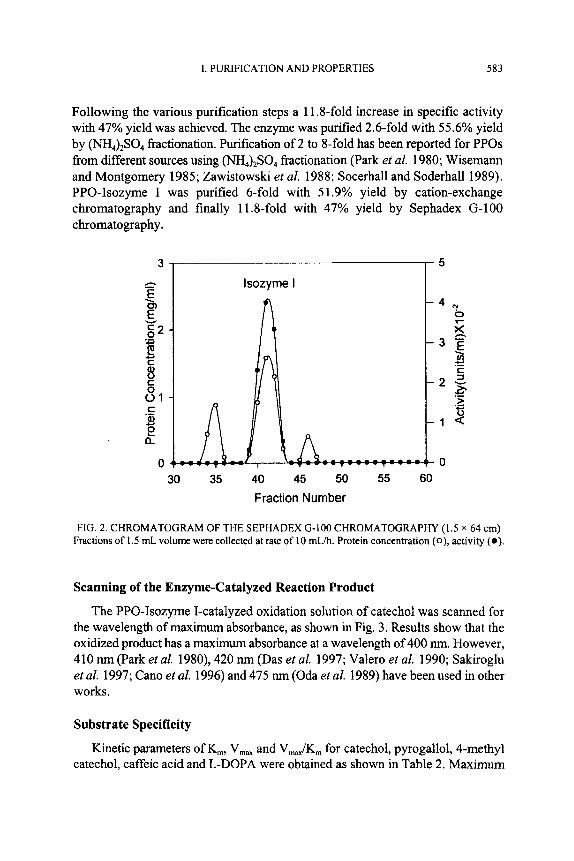
The enzyme solution from “Fraction B” in Fig. 1 was concentrated and chromatographed on a column (1.5 x 62 cm) of Sephadex G-100 equilibrated with 50 mM Mops buffer (pH 6.6). The active fractions were pooled to provide a purified enzyme preparation.
Enzyme Assay
Polyphenol oxidase activity was determined by measuring the increase in absorbance at 400 nm with a UV-Vis spectrophotometer (Varian Cary 3C, USA). The sample cuvette (1 d) contained 1 unit of the enzyme and variable concentrations of substrates. The reaction was initiated by adding enzyme solution and carried out at various temperatures and pH values as below. Enzyme activity was calculated from the initial reaction portion. One unit of enzyme activity was
580 Y.K. CHO and H.K. AHN
0 5 10 15 20 25 30
Fraction Number
FIG. 1. CHROMATOGRAM OF THE CATION EXCHANGE CHROMATOGRAPHY ON BIO- SCALE S2 COLUMN (7 x 52 mm)
Fractions of 1.5 mL volume were collected using 0-1 M NaCl gradient at rate of 2 mUmin. Protein concentration (m), activity (0). NaCl gradient (-).
defined as the amount of the enzyme that causes an increase in absorbance of 0.01 per min at 400 nm and 25C.
Protein Determination
Protein concentrations for the column fractions were determined from the absorbance at 280 nm. Protein concentrations of other samples were determined using a dye-binding technique (Coomassie Brilliant Blue G-25) according to the method of Bradford (1974).
Optimum Temperature
Enzyme activity was measured over a temperature range of 5C to 65C in 50 mM Mops buffer (pH 6.6). The reaction was initiated by adding enzyme solution that had been stored in the ice box. All assays were performed in duplicate.
Temperature Stability
Concentrated enzyme solution (10 units) was mixed with 980 pL of the 50 mM Mops buffer (pH 6.6) pre-equilibrated to the desired temperature. An aliquot was taken out at time intervals and assayed for its remaining activity at 25C . All assays were performed in duplicate.

I. PURIFICATION AND PROPERTIES 581
Optimum pH
All buffers were titrated to the desired pH with KOH. In all cases, overlaps were obtained when buffers were changed so that correction could be made for spurious buffer effects. The pH of the reaction mixture was measured with a Radiometer PHM82 pH meter with a combined microelectrode before and after sufficient data were collected for determination of initial velocities. Negligible pH changes were observed before and after reaction.
Enzyme activity was determined with 20 mM catechol over a pH range of 4.0- 8.8 in 50 mM sodium acetate buffer (PH 4.0-5.5), 50 mM Mes buffer (pH 5.5-6.5), 50 mM Mops buffer (PH 6.5-7.5) and 50 mM Tris buffer (pH 7.5-8.8) at 25C. The pH of the reaction mixtures were checked before and after the reaction. All assays were performed in duplicate.
Enzyme Kinetics
Michaelis-Menten constant (K,,,), maximum velocity (V,,,=) and V,,/K,,, were determined with catechol, pyrogallol, 4-methylcatechol, caffeic acid 3,4-dihydroxy phenylalanine, tyrosine,p-cresol and l-naphtol using 50 mM Mops buffer (pH 6.6). Kinetic parameters were determined from the double reciprocal plot of Lineweaver-Burk (1934). All assays were performed in duplicate.
Electrophoresis
Polyacrylamide gel electrophoresis (PAGE) was conducted by the method of Laemmli (1970) using a mini-gel apparatus, Mini-Protean (Bio-Rad, USA). The gel composition was 12% polyacrylamide and 4% acrylamide for running and stacking gels, respectively. The electrode buffer (PH 8.3) consisted of 25 mM Tris-HC1 and 0.192 M glycine. The sample solution was prepared in a sample buffer (6.3 mM Tris-HC1 and 8 x bromophenol blue) and electrophoresis was run at lOOV in the stacking and 200V in the running gel. Sodium dodecyl sulfate (SDS)-PAGE was performed with the same conditions used for PAGE except for the addition of SDS. The electrode buffer (pH 8.3) contained 25 mM Tris-HC1 10% (VN) glycerol, 2% SDS, 1.54% (WN) dithiothreitol and 8 x 1 0-4% bromophenol blue. Marker proteins used were lysozyme (MW 14,400), soy bean trypsin inhibitor (MW 21,500), carbonic anhydrase (MN 3 1 ,OW), ovalbumin (MW 45,000), bovine serum albumin (MW 66,200) and phosphorylase (MW 97,500).
Molecular Weight Determination
The molecular weight of the native enzyme was estimated using a Sephadex G- 150 (1.5 x 100 cm) column equilibrated with 50 mM Mops buffer (PH 6.6). Marker proteins were carbonic anhydrase (MW 3 1,000), ovalbumin (MW 45,000), bovine serum albumin (MW 66,200), rabbit phosphorylase (MW 97,500), and p-

582 Y.K. CHO and H.K. AHN
galactosidase (MW 116,000), The molecular weight was obtained from the relationship between log(MW) and elution volume.
Determination of Copper Content
SpectrAA-200). All assays were performed in duplicate. Copper was assayed with an atomic absorption spectrophotometer (Varian
Data Processing
program of Cleland (1979). Data for initial velocity studies were fitted to Eq. 1. All data were fitted to the appropriate rate equations using the FORTRAN
v = VA/[K+A] (1)
Here, V is the maximum velocity and A is the concentration of the reactant.
RESULTS AND DISCUSSION
Extraction of Polyphenol Oxidase
Extraction of PPO was carried out in 50 mM Mops buffer (pH 6.6) containing 10 mM cysteine, 20 mM EDTA and 0.35% NaCl. Precipitates formed with (MQ2S04 between 0- lo%, 10-20%, 20-30%, 30-40%, 40-50%, 50-60%, 60-70%, 70-80% and 80-90% were tested to find the proper saturation point. The precipitate obtained from 30-80% (NH,),SO, saturation contained most of the activity (data not shown) and this saturation range was used all the extraction processes. It is well documented that oxidation of phenolics by PPO produces quinones which would inhibit PPO (Walker 1964). Therefore, cysteine was used to reduce quinones to phenolic substrates during extraction.
Purification of Polyphenol Oxidase
Desalted enzyme obtained from dialysis in 50 mM Mops buffer (pH 6.6) was injected into a Bio-Scale S2 column incorporated in a Bio-Rad Automated Protein Purification System. The elution profile is shown in Fig. 1. Two enzyme fractions appeared, i.e. Fraction A and Fraction B. In this study, Fraction B which eluted at 0.3 M NaCl was further fractionated. It was pooled, concentrated and chromatographed in a Sephadex (3-100 column (1.5 x 64 cm) pre-equilibrated with same buffer. The chromatogram shown in Fig. 2 indicates that the enzyme protein was clearly separated. The pooled enzyme from the peak was used for all experiments.
The results of PPO-Isozyme I purification are summarized in Table 1.
1. PURIFICATION AND PROPERTIES 583
Following the various purification steps a 11.8-fold increase in specific activity with 47% yield was achieved. The enzyme was purified 2.6-fold with 55.6% yield by (NH4),S04 fractionation. Purification of 2 to 8-fold has been reported for PPOs from different sources using (NH,),SO, fractionation (Park et al. 1980; Wisemann and Montgomery 1985; Zawistowski et aI. 1988; Socerhall and Soderhall 1989). PPO-Isozyme I was purified 6-fold with 5 1.9% yield by cation-exchange chromatography and finally 11.8-fold with 47% yield by Sephadex G-100 chromatography.
g - 3 E
3 .-
E
- 1 Q
C 3 - 2 - > 'ij 0
30 35 40 45 50 55 60 Fraction Number
FIG. 2. CHROMATOGRAM OF THE SEPHADEX (3-100 CHROMATOGRAPHY (1.5 x 64 cm) Fractions of 1.5 mL volume were collected at rate of 10 mLh. Protein concentration ( O ) , activity (a).
Scanning of the Enzyme-Catalyzed Reaction Product
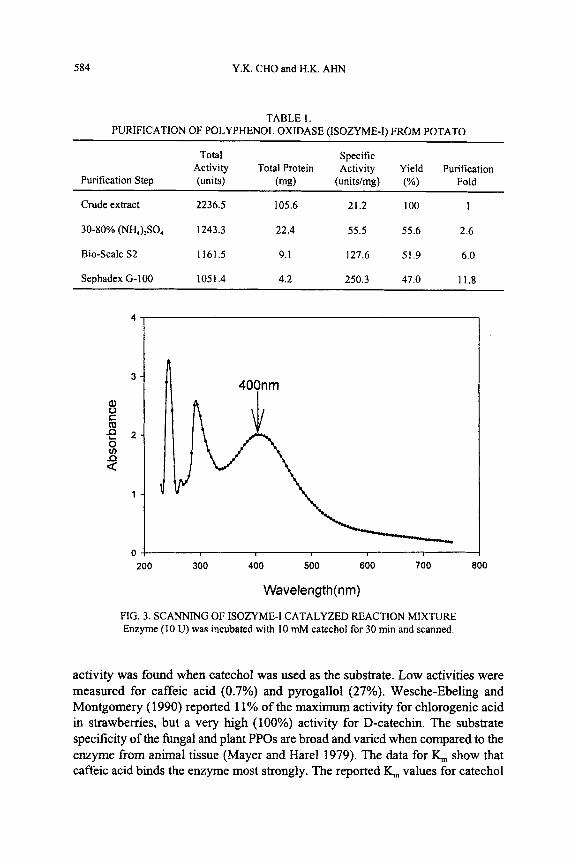
The PPO-Isozyme I-catalyzed oxidation solution of catechol was scanned for the wavelength of maximum absorbance, as shown in Fig. 3. Results show that the oxidized product has a maximum absorbance at a wavelength of 400 nm. However, 410 nm (Park et al. 1980), 420 nm (Das et al. 1997; Valero et al. 1990; Sakiroglu et al. 1997; Can0 et al. 1996) and 475 nm (Oda et al. 1989) have been used in other works.
Substrate Specificity
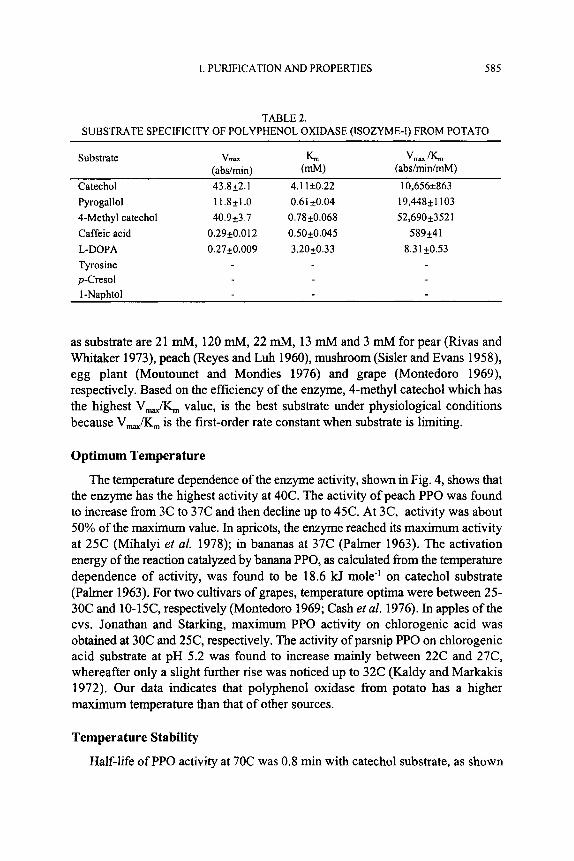
Kinetic parameters of K,,,, V,, and V,,/K, for catechol, pyrogallol, 4-methyl catechol, caffeic acid and L-DOPA were obtained as shown in Table 2. Maximum
584 Y.K. CHO and H.K. AHN
TABLE 1. PURIFICATION OF POLYPHENOL OXIDASE (ISOZYME-I) FROM POTATO
Total Specific Activity Total Protein Activity Yield Purification
Purification Step (units) (mg) (unitdmg) (%) Fold ~~ ~~
Crude extract 2236.5 105.6 21.2 100 1
30-80% (NHJZSO, 1243.3 22.4 55.5 55.6 2.6
Bio-Scale S2 1161.5 9.1 127.6 51.9 6.0
Sephadex G-100 1051.4 4.2 250.3 47.0 11.8
200 300 400 500 600 700 800
Wavelength( nm)
FIG. 3. SCANNING OF ISOZYME-I CATALYZED REACTION MIXTURE Enzyme (10 U) was incubated with 10 mM catechol for 30 min and scanned.
activity was found when catechol was used as the substrate. Low activities were measured for caffeic acid (0.7%) and pyrogallol (27%). Wesche-Ebeling and Montgomery (1990) reported 11% of the maximum activity for chlorogenic acid in strawberries, but a very high (100%) activity for D-catechin. The substrate specificity of the fungal and plant PPOs are broad and varied when compared to the enzyme from animal tissue (Mayer and Hare1 1979). The data for K, show that caffeic acid binds the enzyme most strongly. The reported K,,, values for catechol
I. PURIFICATION AND PROPERTIES 585
TABLE 2. SUBSTRATE SPECIFICITY OF POLYPHENOL OXIDASE (ISOZYME-I) FROM POTATO
Substrate V W L vm, /K, I abs/min) (abs/min/mM)
Catechol 43.8k2.1 4.1 1i0.22 10,656i863 Pyrogallol 1 1.8+1 .O 0.61 t0.04 19,448*1103 4-Methyl catechol 40.9t3.7 0.78*0.068 52,690k3521 Caffeic acid 0.29+0.012 0.50+0.045 589k41 L-DOPA 0.27k0.009 3.20k0.33 8.3 1 k0.53 Tyrosine p-Cresol I-Naphtol
as substrate are 21 mM, 120 mM, 22 mM, 13 mM and 3 mM for pear (Rivas and Whitaker 1973), peach (Reyes and Luh 1960), mushroom (Sisler and Evans 1958), egg plant (Moutounet and Mondies 1976) and grape (Montedoro 1969), respectively. Based on the efficiency of the enzyme, 4-methyl catechol which has the highest V,,& value, is the best substrate under physiological conditions because V,,iTS, is the first-order rate constant when substrate is limiting.
Optimum Temperature
The temperature dependence of the enzyme activity, shown in Fig. 4, shows that the enzyme has the highest activity at 40C. The activity of peach PPO was found to increase from 3C to 37C and then decline up to 45C. At 3C, activity was about 50% of the maximum value. In apricots, the enzyme reached its maximum activity at 25C (Mihalyi et al. 1978); in bananas at 37C (Palmer 1963). The activation energy of the reaction catalyzed by banana PPO, as calculated from the temperature dependence of activity, was found to be 18.6 kJ mole-' on catechol substrate (Palmer 1963). For two cultivars of grapes, temperature optima were between 25- 30C and 10-15C, respectively (Montedoro 1969; Cash et al. 1976). In apples of the cvs. Jonathan and Starking, maximum PPO activity on chlorogenic acid was obtained at 30C and 25C, respectively. The activity of parsnip PPO on chlorogenic acid substrate at pH 5.2 was found to increase mainly between 22C and 27C, whereafter only a slight further rise was noticed up to 32C (Kaldy and Markakis 1972). Our data indicates that polyphenol oxidase from potato has a higher maximum temperature than that of other sources.
Temperature Stability
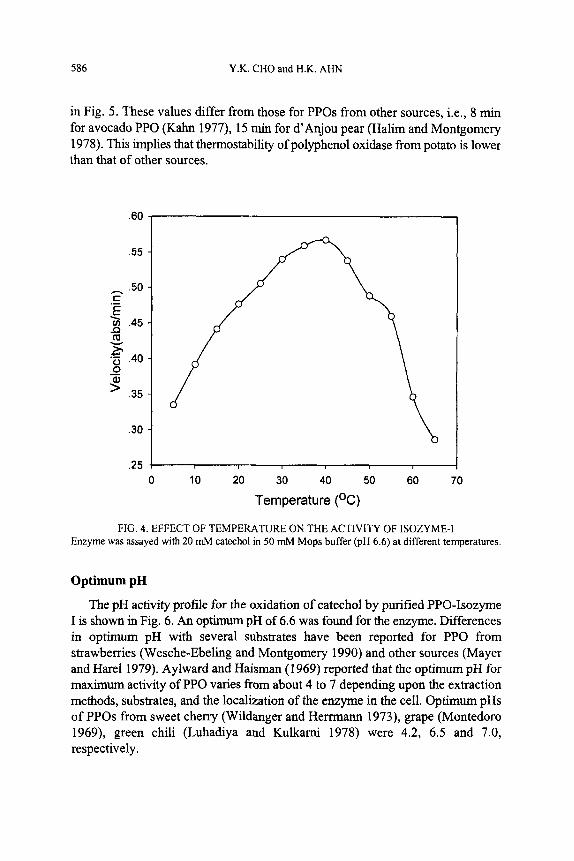
Half-life of PPO activity at 70C was 0.8 min with catechol substrate, as shown
586 Y.K. CHO and H.K. AHN
in Fig. 5. These values differ from those for PPOs from other sources, i.e., 8 min for avocado PPO (Kahn 1977), 15 min for d’Anjou pear (Halim and Montgomery 1978). This implies that thermostability of polyphenol oxidase from potato is lower than that of other sources.
.60
.25 I
0 10 20 30 40 50 60 70
Temperature (OC)
FIG. 4. EFFECT OF TEMPERATURE ON THE ACTIVITY OF ISOZYME-I Enzyme was assayed with 20 mM catechol in 50 mM Mops buffer (pH 6.6) at different temperatures.
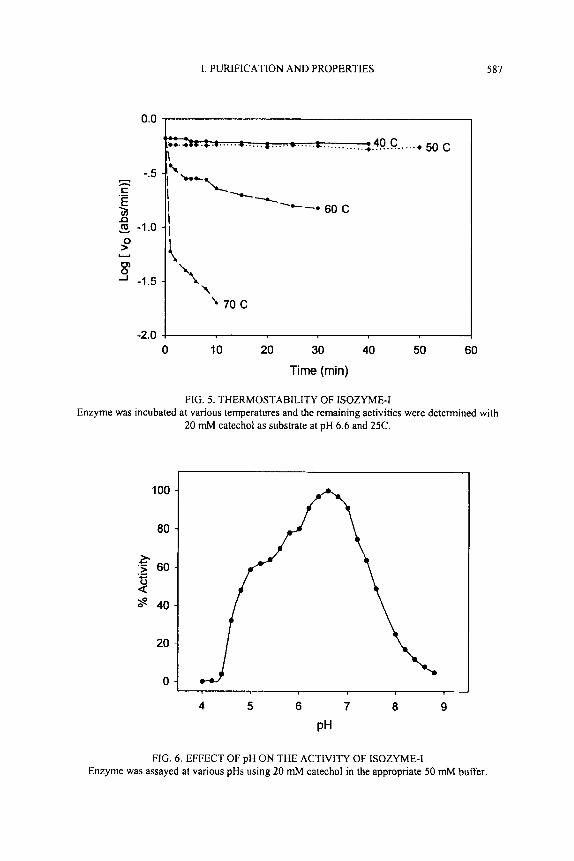
Optimum pH
The pH activity profile for the oxidation of catechol by purified PPO-Isozyme I is shown in Fig. 6. An optimum pH of 6.6 was found for the enzyme. Differences in optimum pH with several substrates have been reported for PPO from strawberries (Wesche-Ebeling and Montgomery 1990) and other sources (Mayer and Hare1 1979). Aylward and Haisman (1969) reported that the optimum pH for maximum activity of PPO varies from about 4 to 7 depending upon the extraction methods, substrates, and the localization of the enzyme in the cell. Optimum pHs of PPOs from sweet cherry (Wildanger and Henmann 1973), grape (Montedoro 1969), green chili (Luhadiya and Kulkami 1978) were 4.2, 6.5 and 7.0, respectively.
1. PURIFICATION AND PROPERTIES 587
0.0
-5 - h C .- E
5 -1.0 2 0 > 0) 0
u
-1.5
-2.0 0 10 20 30 40 50 60
Time (min)
FIG. 5. THERMOSTABILITY OF ISOZYME-I Enzyme was incubated at various temperatures and the remaining activities were determined with
20 mM catechol as substrate at pH 6.6 and 25C.
100
80
3 ‘5 60
&? 40
.- - 2
20
0
4 5 6 7 8 9
PH
FIG. 6. EFFECT OF pH ON THE ACTIVITY OF ISOZYME-I Enzyme was assayed at various pHs using 20 mM catechol in the appropriate 50 mM buffer.
588 Y.K. CHO and H.K. AHN
Molecular Weight
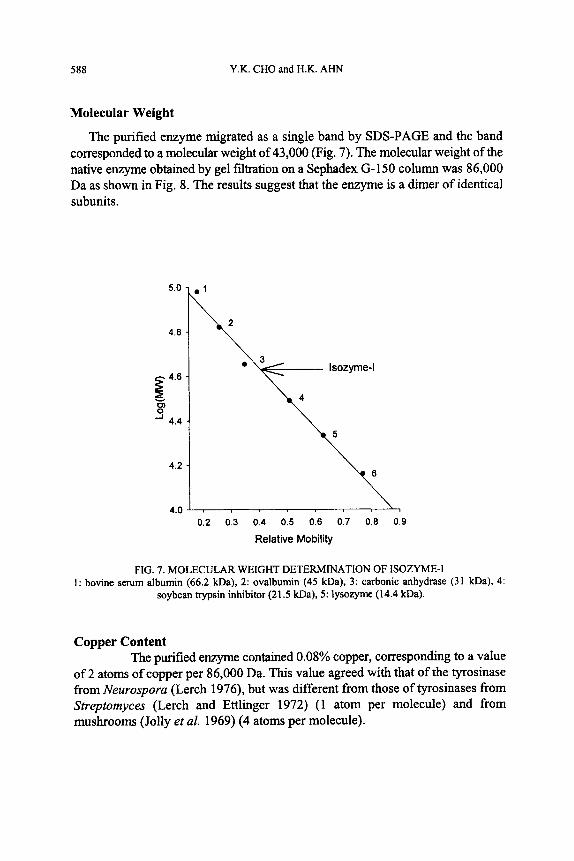
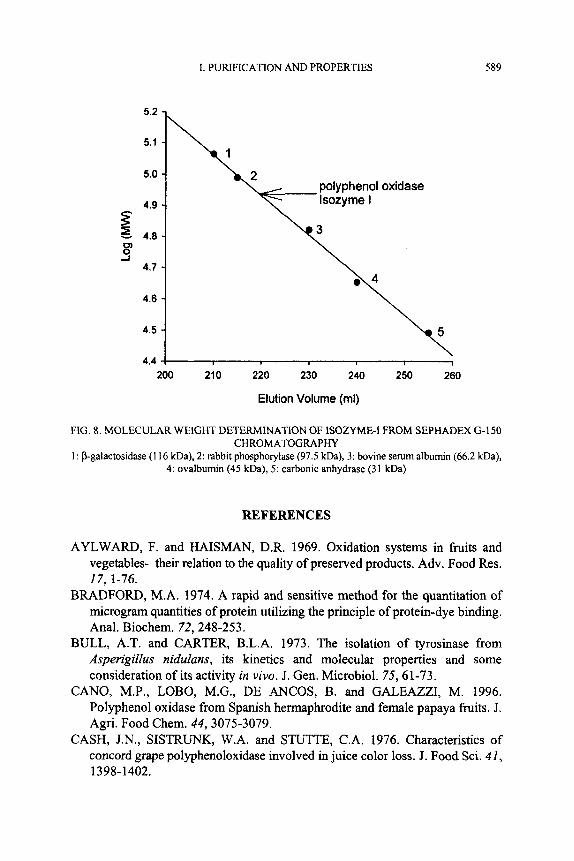
The purified enzyme migrated as a single band by SDS-PAGE and the band corresponded to a molecular weight of 43,000 (Fig. 7). The molecular weight of the native enzyme obtained by gel filtration on a Sephadex G-150 column was 86,000 Da as shown in Fig. 8. The results suggest that the enzyme is a dimer of identical subunits.
4.8 5'0 n 4.0
0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9
Relative Mobility
FIG. 7. MOLECULAR WEIGHT DETERMINATION OF ISOZYME-I 1: bovine serum albumin (66.2 kDa), 2: ovalbumin (45 kDa), 3: carbonic anhydrase (31 kDa), 4:
soybean trypsin inhibitor (21.5 kDa), 5: lysozyme (14.4 kDa).
Copper Content The purified enzyme contained 0.08% copper, corresponding to a value
of 2 atoms of copper per 86,000 Da. This value agreed with that of the tyrosinase from Neurosporu (Lerch 1976), but was different from those of tyrosinases from Streptomyces (Lerch and Ettlinger 1972) (1 atom per molecule) and from mushrooms (Jolly et ul. 1969) (4 atoms per molecule).
I. PURIFICATION AND PROPERTIES 589
200 210 220 230 240 250 260
Elution Volume (ml)
FIG. 8. MOLECULAR WElGHT DETERMlNATlON OF ISOZYME-I FROM SEPHADEX G-150 CHROMATOGRAPHY
1 : P-galactosidase (1 16 kDa), 2: rabbit phosphorylase (97.5 kDa), 3: bovine serum albumin (66.2 kDa), 4: ovalbumin (45 kDa), 5: carbonic anhydrase (31 kDa)
REFERENCES
AYLWARD, F. and HAISMAN, D.R. 1969. Oxidation systems in fruits and vegetables- their relation to the quality of preserved products. Adv. Food Res.
BRADFORD, M.A. 1974. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72,248-253.
BULL, A.T. and CARTER, B.L.A. 1973. The isolation of tyrosinase from Asperigillus nidulans, its kinetics and molecular properties and some consideration of its activity in vivo. J. Gen. Microbiol. 75,61-73.
CANO, M.P., LOBO, M.G., DE ANCOS, B. and GALEAZZI, M. 1996. Polyphenol oxidase from Spanish hermaphrodite and female papaya fruits. J. Agri. Food Chem. 44,3075-3079.
CASH, J.N., SISTRUNK, W.A. and STU?TE, C.A. 1976. Characteristics of concord grape polyphenoloxidase involved in juice color loss. J. Food Sci. 4 I ,
17, 1-76.
1398- 1402.

590 Y.K. CHO and H.K. AHN
CLELAND, W.W. 1979. Statistical analysis of kinetic data. Methods in Enzymol.
DAS, J.R., BHAT, S.G. and GOWDA, L.R. 1997. Purification and characterization of a polyphenol oxidase from the kew cultivar of Indian pineapple fruit. J. Agri. Food Chem. 45,203 1-2035.
FINKLE, B.J. and NELSON, R.F. 1963. Enzyme reactions with phenolic compounds: effect of o-methyl transferase on a natural substrate of fruit polyphenol oxidase. Nature 197, 902-904.
FLURKEY, W.H. and E N , J.J. 1978. Peroxidase and polyphenol oxidase activities in developing peaches. J. Food Sci. 43, 1826- 183 1.
GARDNER, J., MANOHAR, S. and BORISENOK, W.S. 1991. Method and composition for preserving fresh peeled fruits and vegetables. U.S. Patent 4 988, 523.
GOLAN-GOLDHIRSH, A. and WHITAKER, J.R. 1984. Effect of ascorbic axid, sodium bisulfite and thiol compounds on mushroom polyphenol oxidase. J. Agric. Food Chem. 32, 1003-1009.
GOLAN-GOLDHIRSH, A., WHITAKER, J.R. and KAHN, V. 1984. Relation between structure of polyphenol oxidase and prevention of browning. Adv. Exp. Med. Biol. 177,437-456.
GREGORY, R.P.F. and BENDALL, D.S. 1966. The purification and some properties of the polyphenol oxidase from tea (Camellia sinensis L), Biochem.
GUTTERIDGE, S. and ROBB, D. 1975. The catecholase activity of Neurospora tyrosinase. Eur. J. Biochem. 54, 107-1 16.
HALIM, D.H. and MONTGOMERY, M.W. 1978. Polyphenol oxidase of d'Anjou pears. J. Food Sci. 43, 603-608.
INGRAHAM, L.L.J. 1956. Effect of ascorbic acid on polyphenol oxidase. J. Am. Chem. SOC. 78,5095-5099.
IYENGA, R., BOHMONT, C.W. and MCEVILY, A.J. 1991.4-hexylresorcinol and prevention of shrimp melanosis: residual analyses. J. Food Comp. Anal. 4, 148- 153.
JOLLY, R.L., NELSON, R.M. and ROBB, D.A. 1969. Characterization of polyphenol oxidase from mushroom. J. Biol. Chem. 244,3251-3256.
JOSLYN, M.A. and PONTING, J.D. 195 1. Enzyme-catalyzed oxidative browning of fruit products. Adv. Food Res. 3, 1-39.
KAHN, V. 1977. Some biochemical properties of polyphenol oxidase from two avocado varieties differing in their browning rates. J. Food Sci. 42, 38-43.
KALDY, M.S. and MARKAKIS, P. 1972. The browning of parsnip. Can. Inst. Food Sci. Technol. J. 5, 37-42.
KELLY, S.H. and FINKLE, B.J. 1969. Action of a ring-cleaving oxygenase in preventing oxidative darkening of apple juice. J. Sci. Food Agric. 20,629-633.
63, 103-139.
J. 101,569-574.

I. PURIFICATION AND PROPERTIES 591
LAMMELI, V.K. 1970. Cleavage of structural proteins during the assembly of the
LERCH, L. 1976. Characterization of tyrosinase from Neurosporu. FEBS Lett. 69,
LERCH, L. and ETTLINGER, L. 1972. Purification and characterization of tyrosinase from Streptomyces. Eur. J. Biochem. 31,429-433.
LINEWEAVER, H. and BURK, D. 1934. The determination of enzyme dissociation constant. J. Am. Chem. SOC. 56, 658-661.
LUHADIYA, A.P. and KULKARNI, P.R. 1978. Polyphenol oxidase of Capsicum frutescens var. grossa sendt. J. Food Sci. Technol. 15,214.
MATHEW, A.G. and PARPIA, H.A.B. 1971. Food browning as a polyphenol reaction. Adv. Food Res. 19,75-145.
MAYER, A.M. and HAREL, E. 1979. Polyphenol oxidase in plants. Phytochemistry 18, 193-215.
MCEVILY, A.J., IYENGAR, R. and OTWELL, W.S. 1992. Inhibition of enzymatic browning in foods and beverages. Crit. Rev. Food. Sci. Nutr. 32,
MIHALYI, K., VAMOS-VIGYAZO, L., KISS-KUTZ, N. and BABOS- SZEBENYI, E. 1978. The activities of polyphenol oxidase and peroxidase in fruits and vegetables as to pH and temperature. Acta Aliment. Acad. Sci. Hung.
MONTEDORO, M. 1969. Ricerche sulla polifenolossidasi delle uve: 11. Estrazione, purificazione caratterizzazione dell’enzima. Ind. Agrar. 7,259-264.
MOUTOUNET, M. and MONDIES, H. 1976. La polyphenoloxidase de la pruine d’Ente. Ann. Technol. Agric. 25, 343-348.
ODA, Y., KATO, H., ISODA, Y., TAKAHASHI, N., YAMAMOTO, T., TAKADA, Y. and KUDO, S. 1989. Purification and properties of phenoloxidase from spinach leaves. Agri. Biol. Chem. 53,2053-2061.
OTWELL, W.S., IYENGAR, R. and MCEVILY, A.J. 1991. Inhibition of shrimp melanosis by 4-hexylresorcinol. J. Aquatic Food Prod. Technol. I, 53-57.
PALMER, J.K. 1963. Banana polyphenoloxidase. Plant Physiol. 38, 508-5 13. PARK, Y.K., SATO, H.H., ALMEIDA, T.D. and MOLETTI, R.H. 1980.
Polyphenol oxidase of Mango. J. Food Sci. 45, 1619-1624. PATIL, S.S. and ZUCKER, M. 1965. Potato phenolases: purification and
properties. J. Biol. Chem. 240,3938-3943. REYES, P. and LUH, B.S. 1960. Characteristics of browning enzymes in Fay
Elberta freestone peaches. Food Technol. 14,570-575. RIVAS, N.J. and WHITAKER, J.R. 1973. Purification and some properties of two
polyphenol oxidases from Bartlett pear. Plant Physiol. 52, 501-507. SAKIROGLU, H., KUFREVIOGLU, O.I., KOCACALISKAN, I., OKTAY, M.
and ONGANER, Y. 1997. Purification and characterization of dog-rose polyphenol oxidase. J. Agri. Food Chem. 44,2982-2987.
head bacteriophage T4. Nature 227,680-682.
157- 16 1.
253-273.
7, 57-61.

592 Y.K. CHO and H.K. AHN
SARUNO, R., KATO, F. and IKENO, T. 1979. Kojic acid, a tyrosinase inhibitor from Aspergillus albus. Agric. Biol. Chem. 43, 1337-1340.
SCHARF, W. and DAWSON, C.R. 1958. The effect of ascorbic acid on the inactivation of tyrosinase. J. Am. Chem. SOC. 80,4627-463 1.
SISLER, E.C. and EVANS, H.J. 1958. A comparison of chlorogenic acid and catechol as substrates for the polyphenol oxidase from tobacco and mushroom. Plant Physiol. 33,255-260.
SOCERKALL, I. and SODERHALL, K. 1989. Purification of prophenol oxidase from Dacus caroia cell cultures. Phytochem. 28, 1805- 1809.
STELZIG, D.A., AKHTAR, S. and RIBEIRO, S. 1972. Catechol oxidase of red delicious apple peel. Phytochem. 11,535-539.
VALERO, E., VARON, R. and GARCIA-CARMONA, F. 1990. Inhibition of grape polyphenol oxidase by serveral natural aliphatic alcohols. J. Agri. Food Chem. 38,1097- 1 10 1.
VAMOS-VIGYAZO, L. 198 1. Polyphenol oxidase and peroxidase in fruits and vegetables. Crit. Rev. Food Sci. Nutr. 15,49-127.
VAUGHAN, P.F.T., EASON, R., PATON, J.Y. and RITCHIE, G.A. 1975. Molecular weight and amino acid composition of purified spinach beet phenolase. Phytochem. 14, 2383-2386.
WALKER, J.R.L. 1964. Studies on the enzymic browning of apples. 11. Properties of apple polyphenoloxidase. Aust. J. Biol. Sci. 17,360-365.
WESCHE-EBELING, P. and MONTGOMERY, M.W. 1990. Strawberry polyphenol oxidase: purification and characterization. J. Food Sci. 55, 131 5- 1319.
WILDANGER, W. and HERRMANN, K. 1973. Die phenolischen Inhaltsstoffe des Obstes. 11. Die Flavonole des Obstes. Lebensm. Unters. Forsch. 251, 103-107.
WISEMANN, K.W. and MONTGOMERY, M.W. 1985. Purification of d’Anjou pear (Pyrus communis L.) polyphenol oxidase. Plant physiol. 78,256-260.
ZAWISTOWSKI, J., BILLIADERIS, C.G. and MURRAY, E.D. 1988. Purification and characterization of Jerusalem artichoke (Helianfhus tuberosus L. ) polyphenol oxidase. J. Food Biochemistry 12, 1-7.
Related Documents







![Paper Class Xiii[Leader(Xii Xiii)]](https://static.cupdf.com/doc/110x72/577cc5851a28aba7119ca7e3/paper-class-xiiileaderxii-xiii.jpg)



