Induced breeding and larval rearing of Surubı ´, Pseudoplatystoma fasciatum (Linnaeus, 1766), from the Bolivian Amazon Je¤ sus Nun ȶ ez 1,2 ,Re¤ miDugue¤ 1,3 , Navil Corcuy Arana 3 , Fabrice Duponchelle 1,2 , Jean FrancŶ ois Renno 1,4 ,Thomas Raynaud 1,3,5 , Nicolas Hubert 1 & Marc Legendre 1 1 IRD UR 175, GAMET, BP Montpellier cedex 5, France 2 UNFV-FOPCA, Calle Roma, Mira£ores, Lima, Peru¤ 3 UAGRM-CICPE, Santa Cruz de la Sierra, Bolivia 4 IIAP, Avenida Aberlardo Quin ȶ ones, Iquitos, Peru¤ 5 UMII, Creufop, Station Marine Se' te, France Correspondence: Dr J Nun ȶ ez, IRD UR 175, GAMET, BP 5095, 34196 Montpellier, cedex 5, France. E-mail : [email protected], nunez@ ird.fr Abstract Brooders of Surub|¤ ( Pseudoplatystoma fasciatum) were caught in the Ichillo River (Bolivian Amazon) and adapted to captivity conditions for 1 year in the facil- ities of the experimental aquaculture station of ‘El Prado’ (Santa Cruz de la Sierra) under natural tem- perature and photoperiod conditions. Induced repro- duction was obtained by means of Ovaprim s (Syndel, Canada) injections and arti¢cial fertilization. Sperm and ova were obtained by gentle stripping of male and female brooders. Fertilized eggs were incubated in 60 L Zug jars. A mean hatching rate of 73.7 19.0% was obtained after 24 h at 26.5 1C. For larval rearing, several protocols were tested with dif- ferent settings of photoperiod, light intensity, food type and period of distribution, and stocking density. The best survival rates were obtained with Artemia nauplii feeding in total darkness. A high level of ag- gressiveness between larvae and precocious appear- ance of jumpers was observed, but these can be controlled with appropriate rearing conditions. Keywords: Pseudoplatystoma fasciatum, larval rearing, photoperiod, induced spawning, Bolivian Amazon Introduction The ¢shing pressure keeps increasing in the Amazon region (Almeida, McGrath & Ru⁄no, 2001; Ru⁄no 2004, 2005) despite reports of over-exploitation of some of the most important species (Reinert & Win- ter 2002; Petrere, Barthem, Co¤ rdoba & Go¤ mez 2004). In this context, the development of aquaculture is a viable alternative to capture ¢sheries for providing a sustainable source of proteins for local ¢shermen communities and populations of the developing cities in the Amazon region. Currently, the freshwater mar- ketable ¢sh in this area come from inland capture (667 000 MT for 2005) and from aquaculture (246 000 MT) (FAO 2005; Ru⁄no 2005). In some in- stances, the necessity for fast-producing systems has encouraged the introduction of ‘ready-to-use’ alien species, essentially the tilapias, carps and African cat¢sh (Welcomme 1988), but at the cost of environ- mental issues pertaining to biodiversity. The culture of economically important Amazonian species must be considered in order to provide an alternative to alien species introductions in the Amazon basin. The Bolivian Amazon represents a great potential for ¢sh culture, especially in its northern and central parts, where high temperatures and good water qual- ity are available all year round. One of the limiting factors of ¢sh culture development is the lack of sus- tained ¢ngerling production for the important com- mercial species. Within the cat¢sh family Pimelodidae, three species, Pseudoplatystoma fascia- tum, P. tigrinum and P. coruscans , share very similar characteristics and constitute important commercial ¢sh resources in the Amazon and adjacent regions (Almeida 2006). Pseudoplatystoma fasciatum and Aquaculture Research, 2008, 39, 764^776 doi: 10.1111/j.1365-2109.2008.01928.x r 2008 The Authors 764 Journal Compilation r 2008 Blackwell Publishing Ltd

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Induced breeding and larval rearing of Surubı́,
Pseudoplatystoma fasciatum (Linnaeus, 1766), from
the Bolivian Amazon
Je¤ sus Nun� ez1,2, Re¤ mi Dugue¤ 1,3, Navil CorcuyArana3, Fabrice Duponchelle1,2, Jean Franc� ois
Renno1,4,Thomas Raynaud1,3,5, Nicolas Hubert1 & Marc Legendre1
1IRD UR175, GAMET, BPMontpellier cedex 5, France2UNFV-FOPCA, Calle Roma, Mira£ores, Lima, Peru¤3UAGRM-CICPE, Santa Cruz de la Sierra, Bolivia4IIAP, Avenida Aberlardo Quin� ones, Iquitos, Peru¤5UMII, Creufop, Station Marine Se' te, France
Correspondence: Dr J Nun� ez, IRD UR175, GAMET, BP 5095, 34196 Montpellier, cedex 5, France. E-mail : [email protected], nunez@
ird.fr
Abstract
Brooders of Surub|¤ (Pseudoplatystoma fasciatum) werecaught in the Ichillo River (Bolivian Amazon) andadapted to captivity conditions for1year in the facil-ities of the experimental aquaculture station of‘El Prado’ (SantaCruz de la Sierra) under natural tem-perature and photoperiod conditions. Induced repro-ductionwas obtained by means of Ovaprim
s
(Syndel,Canada) injections and arti¢cial fertilization. Spermand ova were obtained by gentle stripping of maleand female brooders. Fertilized eggs were incubatedin 60 L Zug jars. A mean hatching rate of73.7 � 19.0% was obtained after 24 h at 26.5 1C. Forlarval rearing, several protocols were tested with dif-ferent settings of photoperiod, light intensity, foodtype and period of distribution, and stocking density.The best survival rates were obtained with Artemianauplii feeding in total darkness. A high level of ag-gressiveness between larvae and precocious appear-ance of jumpers was observed, but these can becontrolled with appropriate rearing conditions.
Keywords: Pseudoplatystoma fasciatum, larvalrearing, photoperiod, induced spawning, BolivianAmazon
Introduction
The ¢shing pressure keeps increasing in theAmazonregion (Almeida, McGrath & Ru⁄no, 2001; Ru⁄no
2004, 2005) despite reports of over-exploitation ofsome of the most important species (Reinert & Win-ter 2002; Petrere, Barthem, Co¤ rdoba & Go¤ mez 2004).In this context, the development of aquaculture is aviable alternative to capture ¢sheries for providing asustainable source of proteins for local ¢shermencommunities and populations of the developing citiesin theAmazon region. Currently, the freshwater mar-ketable ¢sh in this area come from inland capture(667000 MT for 2005) and from aquaculture(246000 MT) (FAO 2005; Ru⁄no 2005). In some in-stances, the necessity for fast-producing systems hasencouraged the introduction of ‘ready-to-use’ alienspecies, essentially the tilapias, carps and Africancat¢sh (Welcomme 1988), but at the cost of environ-mental issues pertaining to biodiversity. The cultureof economically important Amazonian species mustbe considered in order to provide an alternative toalien species introductions in the Amazon basin.The Bolivian Amazon represents a great potentialfor ¢sh culture, especially in its northern and centralparts, where high temperatures and goodwater qual-ity are available all year round. One of the limitingfactors of ¢sh culture development is the lack of sus-tained ¢ngerling production for the important com-mercial species. Within the cat¢sh familyPimelodidae, three species, Pseudoplatystoma fascia-tum, P. tigrinum and P. coruscans, share very similarcharacteristics and constitute important commercial¢sh resources in the Amazon and adjacent regions(Almeida 2006). Pseudoplatystoma fasciatum and
Aquaculture Research, 2008, 39, 764^776 doi:10.1111/j.1365-2109.2008.01928.x
r 2008 TheAuthors764 Journal Compilationr 2008 Blackwell Publishing Ltd

P. tigrinum occur throughout theAmazon basin, andin the Bolivian Amazon (Loubens & Pan¢li 2000),whereas P. coruscans is distributed in the Rio SaoFrancisco and R|¤ o de la Plata Basins (Barthem &Goulding 1997; Tavares 1997). Recently, the genusPseudoplatystoma has been subdivided into eight spe-cies, three long recognized ones and ¢ve new species:P. punctifer (Castelnau), P. reticulatum Eigenmann &Eigenmann, P. orinocoense n. sp., P. metaense n. sp.and P. magdaleniatum n. (Buitrago^SuaŁ rez & Burr2007). According to the geographic distribution ofthe described species, the species studied here wouldbe P. punctifer. Nevertheless, in this paper, we willcontinue to use the former name, P. fasciatum, forthe sake of consistency with previous studies on thesame species in the region. These Pimelodidae areamong the most appreciated and expensive cat¢shspecies in the ¢sh markets of South America. Giventhe increasing demand for these species, several at-tempts of arti¢cial reproduction have been underta-ken with P. fasciatum (Kossowski & Madrid 1985;Rodriguez 1996; Padilla Pe¤ rez, AlcaŁ ntara Bocanegra& Ismin� o Orbe 2001; GervaŁ sio Leonardo, Romagosa,Borella & Batlouni 2004) and P. coruscans (Sato,Cardoso, Sallum & Godinho1997). However, all thesestudies reported low larval survival rates, and so¢ngerling mass production is not a common realityfor these species yet. Furthermore, these ¢rst experi-ments pointed out the high level of aggressivenessand cannibalism among larvae of P. fasciatum(Kossowski & Madrid 1985; Kossowski & Madrid1991; Kossowski1996).The aggressive behaviour of most cat¢sh larval
stages (Britz & Pienaar 1992; Appelbaum & Kamler2000; Giri, Sahoo, Sahu, Sahu, Mohanty, Mukhopad-hyay & Ayyappan 2002) is one of the main factors af-fecting larval mortality (Qin & Fast1996; Appelbaum& Kamler 2000). Cannibalism is frequent among cul-tured ¢sh larvae, especially in piscivorous species,which develop a wide gape and oral teeth at a preco-cious age (Baras & Jobling 2002). Among the factorsthat govern the impact of cannibalism in ¢sh, sizeheterogeneity is probably the most striking one (Qin& Fast 1996; Kestemont, Jourdan, Houbart, Melard,Paspatis, Fontaine, Cuvier, Kentouri & Baras 2003;Hseu, Huang & Chu 2007; Mandiki, Babiak, Krol,Rasolo & Kestemont 2007). Other factors that modu-late cannibalism, either directly or indirectlythrough an increase in size heterogeneity, includethe feeding environment, rearing density, watertemperature, light regime and intensity (Boeuf & LeBail 1999; Appelbaum & Kamler 2000; Kestemont
et al. 2003). The amount and quality (or type) of foodand its availability have an obvious in£uence on lar-val survival and cannibalismbecause a lackof satietyor a nutritional de¢ciency exacerbates this naturalbehaviour (Qin & Fast 1996). If the feeding periodand meal frequency can in£uence larvae survival, ithas also been reported in several species that lightphase duration can directly in£uence cannibalisticbehaviour (Appelbaum & Kamler 2000).The present study aimed at providing (1) more reli-
able bases for the hormonally induced reproductionof P. fasciatum, (2) a better characterization of thee¡ect of biotic and abiotic factors in larval intensiverearing conditions and (3) the identi¢cation of appro-priate food and feeding regimes for maximizing lar-val survival and growth. A methodology wasdeveloped for induced breeding of P. fasciatum andthree di¡erent food sources for ¢rst feeding of larvae:Artemia nauplii, decapsulated Artemia cysts andearthen pond zooplankton were compared. AsP. fasciatum is a nocturnal animal, feeding mostly dur-ing the night, we investigated in parallel the in£uenceof three important environmental factors (light regimeand intensity, stocking density and feeding period) onthe survival, growth and size heterogeneity of larvae.
Material and methods
Fish husbandry
Fifty ¢sh from 2 to12 kgwere capturedwith nets andhookand line by local ¢shermen in 2002 in the Ichiloriver and its tributaries (Fig. 1) near Puerto Villaroel(16152 0S;64146 0W). The ¢shwere transported in aer-ated tanks to the facilities of the ‘El Prado’ researchstation, about 150 km south of their capture place,tagged individually with PIT-tags and transferred to2000m2 outdoor earthen ponds for acclimatization.During the following year, Surub|¤ brooders were
reared at a low stocking density (one ¢sh per 40m2)and fed with live ¢sh (Prochilodus nigricans,Aequidenssp.) pig and beef liver and commercial pelleted feed.
Male and female selection
During the reproductive season (from November toMarch), sexual maturity of each brood¢sh waschecked every month after anaesthesia in 2-phenoxyethanol (0.4mL L�1) to minimize trauma to the ¢shand operators (Surub|¤ possess strong pectoralspines). For males, the emission of sperm following
Aquaculture Research, 2008, 39, 764^776 Induced breeding of Pseudoplatystoma fasciatum J Nun� ez et al.
r 2008 TheAuthorsJournal Compilationr 2008 Blackwell Publishing Ltd, Aquaculture Research, 39, 764^776 765

gentle abdominal pressurewas veri¢ed.Thematurityof females was determined by examination of folliclesamples taken byan intra-ovarian biopsy.Ovarian samples were placed in physiological
saline and observed under the stereomicroscope(magni¢cation � 20) to determine the proportionof atretic oocytes. Thereafter, saline solution wasremoved and Serra’s solution was added to examinethe position of the germinal vesicle. A subsample(containing at least 50 oocytes) of each biopsy waspreserved for 24 h in physiological saline with 2%formalin, and photographed under the stereomicro-scope. Oocyte diameter distributions were deter-mined from digital photographs, using the IMAGE J
(NIH) software package. For each female, themean oocyte diameter and its coe⁄cient of variationwere calculated.Only males giving sperm following a gentle
abdominal massage were injected with Ovaprims
(Syndel, Qualicum Beach, Canada).
Hormonal treatment, arti¢cial fertilizationand incubation
Females were injected intraperitoneally with Ova-prim at a total dose of 0.5mL kg�1 body weight, ad-ministered as two injections; a priming one at10% ofthe total dose and,12 h later, a resolving one at 90%of the total dose. Females were individually examinedby a gentle abdominal massage starting 6 h after thesecond injection, and then every hour until ovula-tion. When ovulated eggs were present, the femalewas stripped after drying the papilla with a paper to-wel and ova were colleted in dry plastic recipients.Males were treated with 50% of the total female dosein a single injection given at the same time as the fe-male priming injection. The sperm was collected inclean dry syringes before the ¢rst female check in or-der to fertilize ovulated eggs straight after their col-lection. Sperm fractions were stored on ice in cleandry test tubes. One sample of each collected sperm
Figure 1 Schematic map of the capture area of Pseudoplatystoma fasciatum breeding stock in the Bolivian Amazon. TheGrey circular area corresponds approximately to the ¢shing area in the R|¤ o Ichilo basin.
Induced breeding of Pseudoplatystoma fasciatum J Nun� ez et al. Aquaculture Research, 2008, 39, 764^776
r 2008 TheAuthors766 Journal Compilationr 2008 Blackwell Publishing Ltd, Aquaculture Research, 39, 764^776

fraction was tested for motility before being used forarti¢cial fertilization. The routine test consisted inobservation under a microscope (magni¢cation� 400) of sperm activation after a 1:100 dilution indistilled water.Batches of ovulated eggs (100 g) were gently mixed
together with 5mL of 0.154M NaCl and 200 mL ofmilt for 60 s, and then 30mL of distilled water wasadded with constant gentle stirring for another 60 s.Fertilized eggs were rinsed three times with 100mLof water from the incubation circuit, and then trans-ferred into 60 L Zug cylindro-conic jars in a recircu-lating and thermoregulated (26.5 � 2.0 1C) watersystem. In the rest of the manuscript, we refer to ¢shage by reference to the moment of hatching (i.e.hours or days post hatch, hph and dph respectively),which took place 24 h after fertilization. At 24 hph,embryos were collected, counted by a volumetricmethod and distributed in 60 L tanks in a £ow-through (60 L h�1) thermoregulated recirculatingwater system (mean temperature of 26.5 � 2.0 1C).Food distribution started generally at 4 dph.
E¡ects of food type on the growth and survivalof larvae
Three di¡erent food types were tested: decapsulatedcysts of brine shrimp, brine shrimp nauplii and nat-ural zooplankton. Natural zooplankton (mainlyMoina and copepods) was provided by continuouswater pumping from a plankton-enriched earthenpond in the vicinity of the experimental tanks.Waterwas ¢ltered through a 200 mm mesh in order to pre-vent the entrance of insect larvae into the experi-mental tanks. All diets were distributed in excess(controlling the presence of live food 30minafter dis-tribution) ¢ve times a day (8:00, 10:00, 12:00, 14:00and 6:00 hours). Food comparison trials were per-formed using 100 full sibling Surub|¤ larvae aged4 dph. Larvae were counted and distributed in ninecircular 60 L tanks (three replications per food type)connected to a (60 L h�1) water recirculating system(for Artemia cysts and nauplii treatments) or£ow-through pond water at 60 L h�1 (naturalzooplankton treatment). The ammonia and nitriteconcentrations were monitored during the experi-ment and they never exceeded 0.3 and 0.05mg L�1
of N^NH4 and N^NO2 respectively. The meantemperature was maintained nearly constant at27.0 � 1.2 1C during the experiment and so was dis-solved oxygen (between 8 and 9mg L�1) by continu-
ous aeration.The experiment was conducted under anatural photoperiod:12 h of light and12h of darkness(12L:12D).Three controls were performed at15,28 and35 dph to determine the survival rates; standardlengthwasmeasured onlyat 28 dphby image analysis.
E¡ects of light intensity on the growth andsurvival of larvae
In its natural environment, the Surub|¤ , P. fasciatum, isknown as a nocturnal feeding ¢sh (negative photo-taxis).We observed that larvae became photophobicvery rapidly after yolk sac absorption when theyacquired swimming capabilities. For this reason, wetested three levels of light intensity during larvalrearing (o0.01, 1 and 10 lx), corresponding to dark,shaded light, and low light respectively, under a12L:12D photoperiod regime (except for the 24 h darktreatment). No light intensity brighter than10 lx wastested in order to avoid additional stress due to exces-sive light intensity.Twelve sets of1900 larvae aged 2 dphwere distrib-
uted in twelve 60-L tanks (three treatments with fourreplicates). The average temperature in the tanks was28.5 � 1.0 1C. All tanks received the same amount offood in slight excess (Artemia nauplii), distributed sixtimes daily (3:00, 7:00, 11:00, 15:00, 19:00 and 21:00hours), with control half an hour after each distribu-tion to make sure that the number of preys availablewas in excess. At 9 dph, larvae were sampled for sur-vival and size determination using image analysis.
E¡ects of stocking density on the growth andsurvival of larvae
Three stocking densities were evaluated using(almost) logarithmic steps between densities (i.e. 10,30, 100 larvae L�1). Two light regimes (12L:12D and0L:24D) were tested for each stocking density, in or-der to evaluate the interaction between the two envir-onmental variables (six treatments, three replicationsper treatment combination). Fishwere fed ¢ve times aday (8:00, 10:00, 12:00, 14:00 and 16:00 hours) inslight excess with newly hatched Artemia nauplii.Water temperature was maintained at 26.7 � 1.0 1C.
E¡ect of photoperiod and food distributionschedule on the growth and survival of larvae
Three light regimes were evaluated: 0L:24D;12L:12Dand 24L:0D. Artemia nauplii were distributed in
Aquaculture Research, 2008, 39, 764^776 Induced breeding of Pseudoplatystoma fasciatum J Nun� ez et al.
r 2008 TheAuthorsJournal Compilationr 2008 Blackwell Publishing Ltd, Aquaculture Research, 39, 764^776 767
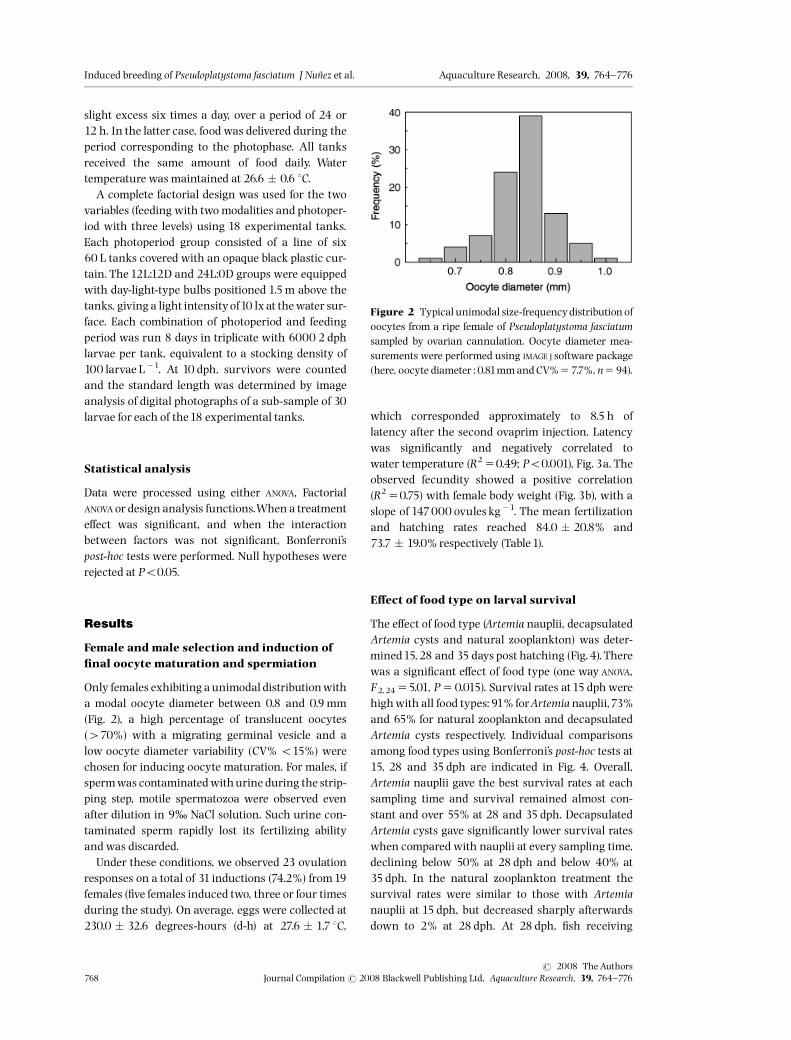
slight excess six times a day, over a period of 24 or12 h. In the latter case, food was delivered during theperiod corresponding to the photophase. All tanksreceived the same amount of food daily. Watertemperature was maintained at 26.6 � 0.6 1C.A complete factorial design was used for the two
variables (feeding with two modalities and photoper-iod with three levels) using 18 experimental tanks.Each photoperiod group consisted of a line of six60 L tanks covered with an opaque black plastic cur-tain. The12L:12D and 24L:0D groups were equippedwith day-light-type bulbs positioned 1.5m above thetanks, giving a light intensityof10 lx at thewater sur-face. Each combination of photoperiod and feedingperiod was run 8 days in triplicate with 6000 2 dphlarvae per tank, equivalent to a stocking density of100 larvae L�1. At 10 dph, survivors were countedand the standard length was determined by imageanalysis of digital photographs of a sub-sample of 30larvae for each of the18 experimental tanks.
Statistical analysis
Data were processed using either ANOVA, FactorialANOVAor design analysis functions.When a treatmente¡ect was signi¢cant, and when the interactionbetween factors was not signi¢cant, Bonferroni’spost-hoc tests were performed. Null hypotheses wererejected at Po0.05.
Results
Female and male selection and induction of¢nal oocyte maturation and spermiation
Only females exhibiting aunimodal distributionwitha modal oocyte diameter between 0.8 and 0.9mm(Fig. 2), a high percentage of translucent oocytes(470%) with a migrating germinal vesicle and alow oocyte diameter variability (CV% o15%) werechosen for inducing oocyte maturation. For males, ifspermwas contaminatedwithurine during the strip-ping step, motile spermatozoa were observed evenafter dilution in 9% NaCl solution. Such urine con-taminated sperm rapidly lost its fertilizing abilityand was discarded.Under these conditions, we observed 23 ovulation
responses on a total of 31 inductions (74.2%) from19females (¢ve females induced two, three or four timesduring the study). On average, eggs were collected at230.0 � 32.6 degrees-hours (d-h) at 27.6 � 1.7 1C,
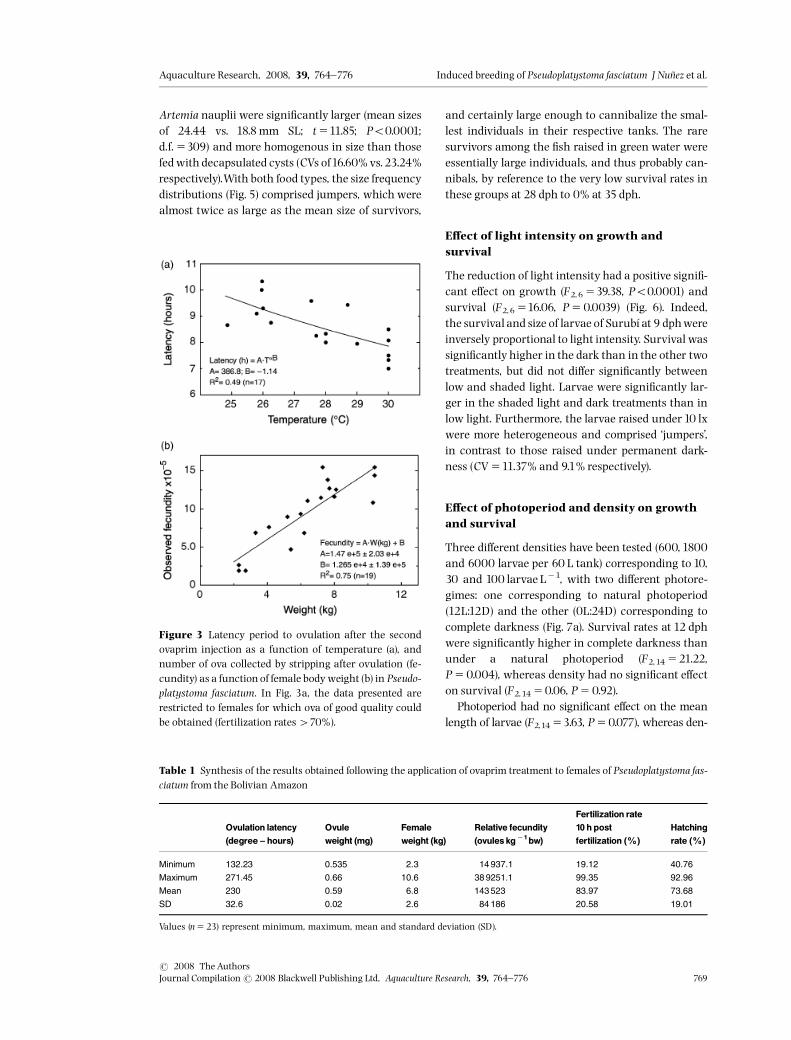
which corresponded approximately to 8.5 h oflatency after the second ovaprim injection. Latencywas signi¢cantly and negatively correlated towater temperature (R2 50.49; Po0.001), Fig.3a. Theobserved fecundity showed a positive correlation(R2 50.75) with female body weight (Fig. 3b), with aslope of 147000 ovules kg�1. The mean fertilizationand hatching rates reached 84.0 � 20.8% and73.7 � 19.0% respectively (Table1).
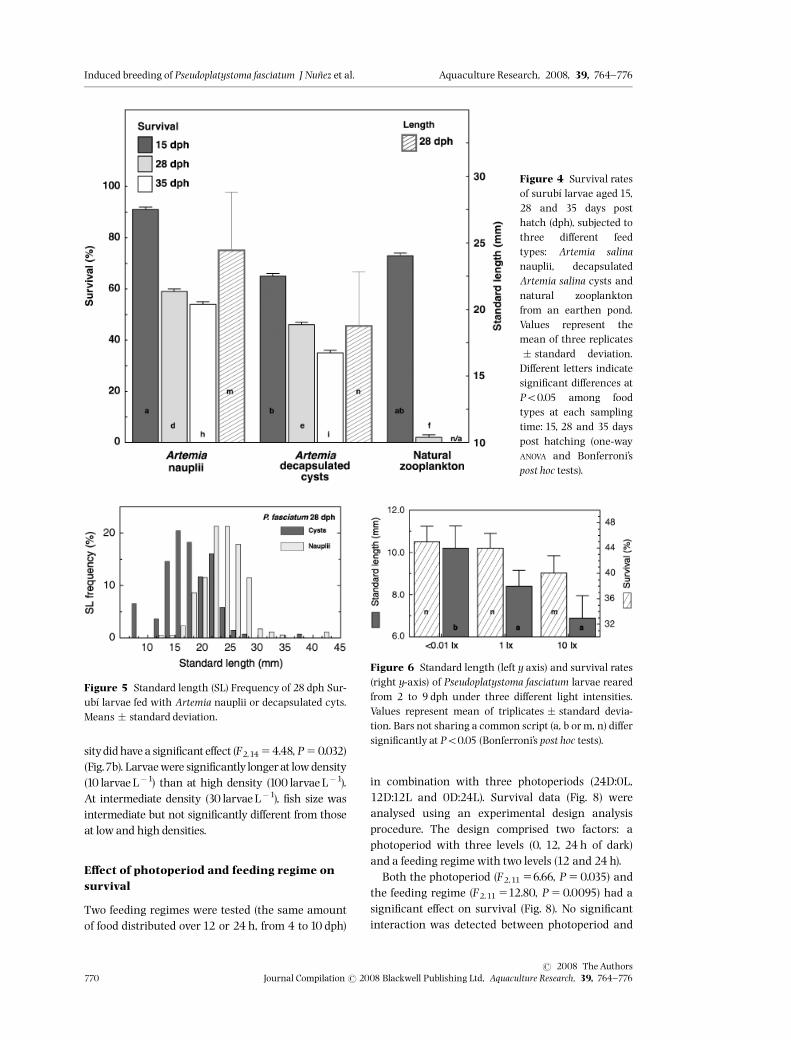
E¡ect of food type on larval survival
The e¡ect of food type (Artemia nauplii, decapsulatedArtemia cysts and natural zooplankton) was deter-mined15,28 and 35 days post hatching (Fig.4).Therewas a signi¢cant e¡ect of food type (one way ANOVA,F2,24 55.01, P50.015). Survival rates at15 dph werehighwith all food types:91% forArtemia nauplii,73%and 65% for natural zooplankton and decapsulatedArtemia cysts respectively. Individual comparisonsamong food types using Bonferroni’s post-hoc tests at15, 28 and 35 dph are indicated in Fig. 4. Overall,Artemia nauplii gave the best survival rates at eachsampling time and survival remained almost con-stant and over 55% at 28 and 35 dph. DecapsulatedArtemia cysts gave signi¢cantly lower survival rateswhen compared with nauplii at every sampling time,declining below 50% at 28 dph and below 40% at35 dph. In the natural zooplankton treatment thesurvival rates were similar to those with Artemianauplii at 15 dph, but decreased sharply afterwardsdown to 2% at 28 dph. At 28 dph, ¢sh receiving
Figure 2 Typical unimodal size-frequencydistributionofoocytes from a ripe female of Pseudoplatystoma fasciatumsampled by ovarian cannulation. Oocyte diameter mea-surements were performed using IMAGE J software package(here, oocyte diameter :0.81mmandCV%57.7%, n594).
Induced breeding of Pseudoplatystoma fasciatum J Nun� ez et al. Aquaculture Research, 2008, 39, 764^776
r 2008 TheAuthors768 Journal Compilationr 2008 Blackwell Publishing Ltd, Aquaculture Research, 39, 764^776
Artemia nauplii were signi¢cantly larger (mean sizesof 24.44 vs. 18.8mm SL; t511.85; Po0.0001;d.f.5309) and more homogenous in size than thosefedwith decapsulated cysts (CVs of16.60% vs.23.24%respectively).With both food types, the size frequencydistributions (Fig. 5) comprised jumpers, which werealmost twice as large as the mean size of survivors,
and certainly large enough to cannibalize the smal-lest individuals in their respective tanks. The raresurvivors among the ¢sh raised in green water wereessentially large individuals, and thus probably can-nibals, by reference to the very low survival rates inthese groups at 28 dph to 0% at 35 dph.
E¡ect of light intensity on growth andsurvival
The reduction of light intensity had a positive signi¢-cant e¡ect on growth (F2,6 539.38, Po0.0001) andsurvival (F2,6 516.06, P50.0039) (Fig. 6). Indeed,the survival and size of larvae of Surub|¤ at 9 dphwereinversely proportional to light intensity. Survival wassigni¢cantly higher in the dark than in the other twotreatments, but did not di¡er signi¢cantly betweenlow and shaded light. Larvae were signi¢cantly lar-ger in the shaded light and dark treatments than inlow light. Furthermore, the larvae raised under10 lxwere more heterogeneous and comprised ‘jumpers’,in contrast to those raised under permanent dark-ness (CV511.37% and 9.1% respectively).
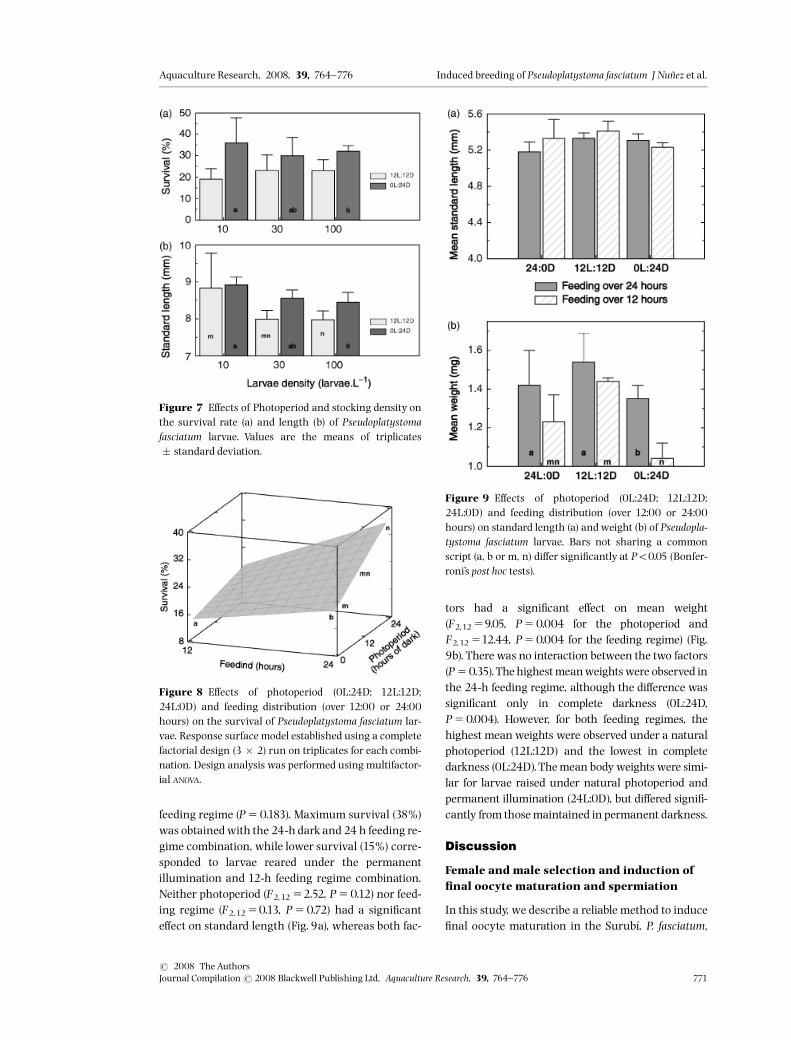
E¡ect of photoperiod and density on growthand survival
Three di¡erent densities have been tested (600,1800and 6000 larvae per 60 L tank) corresponding to 10,30 and 100 larvae L�1, with two di¡erent photore-gimes: one corresponding to natural photoperiod(12L:12D) and the other (0L:24D) corresponding tocomplete darkness (Fig. 7a). Survival rates at 12 dphwere signi¢cantly higher in complete darkness thanunder a natural photoperiod (F2,14 521.22,P50.004), whereas density had no signi¢cant e¡ecton survival (F2,14 50.06, P50.92).Photoperiod had no signi¢cant e¡ect on the mean
length of larvae (F2,14 53.63, P50.077), whereas den-
Figure 3 Latency period to ovulation after the secondovaprim injection as a function of temperature (a), andnumber of ova collected by stripping after ovulation (fe-cundity) as a function of female bodyweight (b) in Pseudo-platystoma fasciatum. In Fig. 3a, the data presented arerestricted to females for which ova of good quality couldbe obtained (fertilization rates470%).
Table 1 Synthesis of the results obtained following the application of ovaprim treatment to females of Pseudoplatystoma fas-ciatum from the Bolivian Amazon
Ovulation latency(degree ^ hours)
Ovuleweight (mg)
Femaleweight (kg)
Relative fecundity(ovules kg�1bw)
Fertilization rate10 h postfertilization (%)
Hatchingrate (%)
Minimum 132.23 0.535 2.3 14 937.1 19.12 40.76
Maximum 271.45 0.66 10.6 38 9251.1 99.35 92.96
Mean 230 0.59 6.8 143 523 83.97 73.68
SD 32.6 0.02 2.6 84 186 20.58 19.01
Values (n523) represent minimum, maximum, mean and standard deviation (SD).
Aquaculture Research, 2008, 39, 764^776 Induced breeding of Pseudoplatystoma fasciatum J Nun� ez et al.
r 2008 TheAuthorsJournal Compilationr 2008 Blackwell Publishing Ltd, Aquaculture Research, 39, 764^776 769
sitydid have a signi¢cant e¡ect (F2,14 54.48,P50.032)(Fig.7b). Larvaewere signi¢cantly longerat lowdensity(10 larvae L�1) than at high density (100 larvae L�1).At intermediate density (30 larvae L�1), ¢sh size wasintermediate but not signi¢cantly di¡erent from thoseat lowand high densities.
E¡ect of photoperiod and feeding regime onsurvival
Two feeding regimes were tested (the same amountof food distributed over 12 or 24 h, from 4 to 10 dph)
in combination with three photoperiods (24D:0L,12D:12L and 0D:24L). Survival data (Fig. 8) wereanalysed using an experimental design analysisprocedure. The design comprised two factors: aphotoperiod with three levels (0, 12, 24 h of dark)and a feeding regime with two levels (12 and 24 h).Both the photoperiod (F2,11 56.66, P50.035) and
the feeding regime (F2,11 512.80, P50.0095) had asigni¢cant e¡ect on survival (Fig. 8). No signi¢cantinteraction was detected between photoperiod and
Figure 4 Survival ratesof surub|¤ larvae aged 15,28 and 35 days posthatch (dph), subjected tothree di¡erent feedtypes: Artemia salinanauplii, decapsulatedArtemia salina cysts andnatural zooplanktonfrom an earthen pond.Values represent themean of three replicates� standard deviation.Di¡erent letters indicatesigni¢cant di¡erences atPo0.05 among foodtypes at each samplingtime: 15, 28 and 35 dayspost hatching (one-wayANOVA and Bonferroni’spost hoc tests).
Figure 5 Standard length (SL) Frequency of 28 dph Sur-ub|¤ larvae fed with Artemia nauplii or decapsulated cyts.Means � standard deviation.
Figure 6 Standard length (left y axis) and survival rates(right y-axis) of Pseudoplatystoma fasciatum larvae rearedfrom 2 to 9 dph under three di¡erent light intensities.Values represent mean of triplicates � standard devia-tion. Bars not sharing a common script (a, b or m, n) di¡ersigni¢cantly at Po0.05 (Bonferroni’s post hoc tests).
Induced breeding of Pseudoplatystoma fasciatum J Nun� ez et al. Aquaculture Research, 2008, 39, 764^776
r 2008 TheAuthors770 Journal Compilationr 2008 Blackwell Publishing Ltd, Aquaculture Research, 39, 764^776
feeding regime (P50.183). Maximum survival (38%)was obtainedwith the 24-h dark and 24 h feeding re-gime combination, while lower survival (15%) corre-sponded to larvae reared under the permanentillumination and 12-h feeding regime combination.Neither photoperiod (F2,12 52.52, P50.12) nor feed-ing regime (F2,12 50.13, P50.72) had a signi¢cante¡ect on standard length (Fig. 9a), whereas both fac-
tors had a signi¢cant e¡ect on mean weight(F2,12 59.05, P50.004 for the photoperiod andF2,12 512.44, P50.004 for the feeding regime) (Fig.9b). There was no interaction between the two factors(P50.35).The highest meanweights were observed inthe 24-h feeding regime, although the di¡erence wassigni¢cant only in complete darkness (0L:24D,P50.004). However, for both feeding regimes, thehighest mean weights were observed under a naturalphotoperiod (12L:12D) and the lowest in completedarkness (0L:24D).The mean body weights were simi-lar for larvae raised under natural photoperiod andpermanent illumination (24L:0D), but di¡ered signi¢-cantly from those maintained in permanent darkness.
Discussion
Female and male selection and induction of¢nal oocyte maturation and spermiation
In this study, we describe a reliable method to induce¢nal oocyte maturation in the Surub|¤ , P. fasciatum,
Figure 7 E¡ects of Photoperiod and stocking density onthe survival rate (a) and length (b) of Pseudoplatystomafasciatum larvae. Values are the means of triplicates� standard deviation.
Figure 8 E¡ects of photoperiod (0L:24D; 12L:12D;24L:0D) and feeding distribution (over 12:00 or 24:00hours) on the survival of Pseudoplatystoma fasciatum lar-vae. Response surface model established using a completefactorial design (3 � 2) run on triplicates for each combi-nation. Design analysis was performed using multifactor-ial ANOVA.
Figure 9 E¡ects of photoperiod (0L:24D; 12L:12D;24L:0D) and feeding distribution (over 12:00 or 24:00hours) on standard length (a) and weight (b) of Pseudopla-tystoma fasciatum larvae. Bars not sharing a commonscript (a, b or m, n) di¡er signi¢cantly at Po0.05 (Bonfer-roni’s post hoc tests).
Aquaculture Research, 2008, 39, 764^776 Induced breeding of Pseudoplatystoma fasciatum J Nun� ez et al.
r 2008 TheAuthorsJournal Compilationr 2008 Blackwell Publishing Ltd, Aquaculture Research, 39, 764^776 771

using ovaprim at the recommended dosage(0.5mL kg�1). This result further supports the ¢nd-ing that the combination of LH-RHa and an anti-dopamine factor, here domperidone, is a potentinductor of ¢nal oocyte maturation in a broad rangeof ¢sh species (Lin, van der Kraak, Liang, Peng, Li, Lu,Zhou, Chang & Peter1986; Peter, Lin & Van der Kraak1988) and particularly in cat¢sh species (De Leeuw,Goos, Richter & Eding1985; Goos, Joy, De Leeuw,VanOordt,Van Delft & Gielen 1987; Richter, Eding, Goos,De Leeuw, Scott & Van Oordt1987; Legendre, Linhart& Billard1996; Alok, Krishnan,Talwar & Garg1998).One of the advantages over carp pituitary extract(CPE), which is frequently used in ¢sh farms, is thatthe biological activity of ovaprim or other LH-RHapreparations is standardized and constant if the pro-ducts are stored properly. It is also most likely that apart of the success in inducing the ¢nal maturationoffemales emerged from the determination of objectivecriteria for female selection. These criteria werebased on the direct observation of oocytes obtainedby an intra-ovarian biopsy as already reported forthis species (GervaŁ sio Leonardo et al. 2004) and forother cultured African and Asian cat¢sh (Gilles,Dugue¤ & Slembrouck 2001; Slembrouck, Komarudin,Maskur & Legendre 2004).We observed that high fer-tilization and hatching rates were associated withhigh proportions of translucent oocytes with migrat-ing germinal vesicle and low coe⁄cients of variationof the modal diameter of postvitellogenic oocytes.Once female selection is achieved, another importantfactor is the time of latency, which has to be consid-ered very carefully because it determines high fertili-zation and hatching rates (results not shown). Werecommend using the number of d-h rather than theelapsed time, because the d-h calculation integratesall temperature variations during the maturationprocess and is thus more accurate.We observed in dif-ferent induction trials that in overripe females(4250 d-h), stripping was generally very easy, butfertilization and hatching rates were very low. Bycontrast, stripping before 180 d-h was generallyunproductive. Nevertheless, we exceptionally ob-tained ovulated eggs as early as 130 d-h for early-re-sponding females and as late as 270 d-h for late-responding females (seeTable 1). In this study, we at-tained an overall 74.2% proportion of ovulation re-sponse (n531 inductions). This methodologyenabled the production of large numbers of viablelarvae per female. The collection of sperm from malesdid not require hormonal induction, but it is recom-mended to inject males with 50% of the female total
dose in order to increase milt volume and facilitate itscollection. Attention has to be focused on the spermcol-lection procedure to avoid urine contamination, whichmayactivate spermatozoamotility. It is preferable to col-lect the sperm in separate fractions and to test each frac-tion separately before its use for arti¢cial fertilization.
E¡ect of Food type on larval survival
Artemia nauplii were shown to be adequate for feed-ing Surubi larvae as soon as they start exogenousfeeding. Larvae fed with decapsulated cysts of Arte-mia survived almost equally well, but their growthwas slower, possibly because inert cysts are less at-tractive than live prey, leading to a lower ingestion,or because they are less digestible than nauplii, espe-cially for a species that starts feeding at a relativelysmall size (ca. 4.5mmSL). Decapsulated cyts andArtemia nauplii are now widely used for a large vari-ety of cultured species. Nevertheless, other more af-fordable alternatives can be used to initiate larvaefeeding on live prey until weaning. Previous work onthis species reported good growth with natural zoo-planktonic prey during the ¢rst10 days of larval rear-ing (Padilla Pe¤ rez et al. 2001) but there was noindication as to whether survival was low or high.This study provided evidence that rearing Surubi lar-vae in green water until the age of 15 dph did notcompromise their survival. After this period, naturalzooplankton was apparently no longer su⁄cient tosustain larvae survival.This result o¡ers perspectivesfor evaluating weaning schedules straight afterplankton feeding. However, it is strongly suggestedthat larval rearing in green water be restricted tolow stocking densities; otherwise, the plankton re-sources may rapidly become limiting, thereby caus-ing growth heterogeneity and cannibalism, as wasobserved during this study. The early weaning of15 dph larvae after natural zooplankton feeding hasnot yet been tested in this species. Nevertheless, thispossibility has been tested successfully in marinespecies like Red porgy (Pagrus pagrus), with a wean-ing protocol (co-feeding) involving live prey and drydiets (AristizaŁ bal & SuaŁ rez 2006), or Sole (S. senega-lensis) (Ribeiro, Engrola & Dinis 2005).
E¡ect of light intensity on growth andsurvival
The e¡ect of light intensity has already been tested inmarine and freshwater cultured species, and there isgenerally a positive correlation of growth and survi-
Induced breeding of Pseudoplatystoma fasciatum J Nun� ez et al. Aquaculture Research, 2008, 39, 764^776
r 2008 TheAuthors772 Journal Compilationr 2008 Blackwell Publishing Ltd, Aquaculture Research, 39, 764^776

val with ambient light intensity in most ¢sh speciesstudied (Boeuf & Le Bail 1999). However, in cat¢shthe situation appears to be di¡erent because, moststudies reported better survival and sometimesgrowth under short photoperiods or even in perma-nent darkness (Sundararaj, Nath &Halberg1982; Ker-dchuen & Legendre1991; Britz & Pienaar1992; Baras,Tissier, Westerloppe, Me¤ lard & Philippart 1998; Giriet al. 2002; Almazan-Rueda, van Helmond,Verreth &Schrama 2005). This characteristic seems to be asso-ciated with their natural feeding behaviour, as adultsusually feed around sunset or during the night (Bel-benoit, Moller, Serrier & Push 1979; Sundararaj et al.1982; Rodriguez, Richardson & Lewis Jr 1990;Kerdchuen & Legendre1991; Ezenwaji1999; Hossain,Batty, Haylor & Beveridge 1999; Spotte, Petry & Zua-non 2001; Almazan-Rueda, Schrama & Verreth2004; Pohlmann, Atema & Breithaupt 2004). Cat¢shlarvae become photophobic early during the larvalperiod and exhibit a negative phototaxis (Blackshaw& Snyder 1997; Appelbaum & McGeer 1998; Hossainet al. 1999), but under arti¢cial conditions larvae ofmany species can feed either in the light or in com-plete darkness. As in some other species, in most cat-¢sh, larvae exhibit aggressive behaviour, leading tonumerous injuries and frequent cannibalism (Britz& Pienaar 1992; Appelbaum & Kamler 2000; Giriet al. 2002). Long periods of light in the African cat-¢sh, Clarias gariepinus, can induce increased plasmacortisol levels, indicating a physiological response tostressful conditions (Almazan-Rueda et al. 2005). Ac-cording to our observations, the aggressive beha-viour in P. fasciatum might be responsible for asigni¢cant part of larval mortality during the ¢rst 2or 3 weeks of life.We observed that rearing larvae inthe dark ensures less competition for space, becauselarvae are mainly distributed in the water column in-stead of being concentrated in the darker parts of thetank. Therefore, there are limited contacts betweenthem, which ¢nally improves survival rates, and tosome extent growth. As Surub|¤ larvae keep feedingnormally in complete darkness, survival rates are im-proved under these conditions. We strongly recom-mend the use of complete dark conditions for thelarval rearing of Surub|¤ .
E¡ect of photoperiod and density on growthand survival
The e¡ect of photoperiod and density on survival andgrowth in ¢sh larvae has been demonstrated in some
cultured species (Kaiser, Weyl & Hecht 1995; Alma-zan-Rueda et al. 2005). Nevertheless, in our experi-ments, with a10-fold range of variation, the e¡ect ofdensity on survival was not signi¢cant. Survival atlow densities was slightly better only in completelydark conditions, but this might be more related tothe photoperiod (day length) than to density itself.However, larval growth was signi¢cantly a¡ected bydensity: the larger larvae were observed at the lowestdensity. This situation is quite common among cul-tured species because each larva has more spaceand spends less energycompeting for foodwith othercongeners. In this study, the range of initial larvaedensity variation (10^100 larvae L�1 correspondingto 600^6000 larvaem�2) was relatively high com-pared with the initial densities reported for marinespecies (0.1^30 larvae L�1) (Hitzfelder, Holt, Fox &McKee 2006) but similar to other reported rearingdensities for fresh water ¢sh, Sander lucioperca (25^100 L�1) (Szkudlarek & Zake
’sŁ 2007) or Clarias batra-
chus (1000^5000m�2) (Sahoo, Giri & Sahu 2004),indicating that P. fasciatummay be reared under highdensity conditions but also that survival will likely beimproved using lower densities.
E¡ect of photoperiod and feeding regime onsurvival
There was a striking e¡ect of both feeding and lightregimes. In several ¢sh species (visual-based feeders),the e¡ect of long photophase generally has a positivee¡ect on growth and survival (Barlow, Pearce, Rod-gers & Clayton 1995; Ronzani Cerqueira & MacedoBrˇgger 2001; Downing & Litvak 2002; Fielder,Bardsley, Allan & Pankhurst 2002; Puvanendran &Brown 2002; Trotter, Battaglene & Pankhurst 2003;Moustakas,Watanabe & Copeland 2004; Pedro Cana-vate, Zerolo & Fernandez-Diaz 2006). As in other cat-¢sh species (Appelbaum & McGeer1998; Appelbaum& Kamler, 2000; Giri et al. 2002), survival of Surub|¤larvaewas higher under complete darkness andwitha continuous food distribution. In complete darkness,larvae activity is mainly devoted to feeding and theiraggressive behaviour is highly reduced because theyremain quiet for long periods of time after feeding.The continuous distribution of food (over 24 h) alsoprevents long periods of food deprivation, which tendto stimulate foraging behaviour and increase con-tacts between ¢sh. In some species, these higher sur-vival rates were also associated with faster growthbecause the costs of swimming activity and aggres-
Aquaculture Research, 2008, 39, 764^776 Induced breeding of Pseudoplatystoma fasciatum J Nun� ez et al.
r 2008 TheAuthorsJournal Compilationr 2008 Blackwell Publishing Ltd, Aquaculture Research, 39, 764^776 773

sive behaviour are reduced. However, this potentialgrowth gain could not be detected under the presentexperimental conditions.In conclusion, we have developed a reliable metho-
dology for ¢sh selection, and induced breeding forthis very important, yet poorly studied Amazonianspecies. The overall survival rate obtained under theexperimental conditions in this study was around40% at 35 dph, which is compatible with mass pro-duction of fry owing to the high fecundity of this spe-cies. These preliminary results should help promotthe culture of this promising species, which will bethe only way to alleviate the increasing ¢shing pres-sure it endures in most Amazonian countries and too¡er a new candidate for aquaculture of native spe-cies. Future research e¡orts should, focus amongother things, on developing weaning protocols withe⁄cient dry foods, for increasing growth rates and at-taining a sustainable number of ¢ngerlings for grow-out production of this species.
Acknowledgments
Wewould like to thank all the technical sta¡ and stu-dents of the Universidad Gabriel Rene¤ Moreno at the‘El Prado’ research station, especially Jose¤ Mamani,Jose¤ Luis Linares, Roberto Ken Atta and Ojopi Ortiz.Special thanks are due to Dr Etienne Baras for thevaluable comments, criticisms and suggestionsduring the preparation of the manuscript. This workwas funded by IRD through the UR081/UR175-UAGRM/El Prado cooperation research programme(2002^2005).
References
Almazan-Rueda P., Schrama J.W. & Verreth J.A.J. (2004)Behavioural responses under di¡erent feeding methodsand light regimes of theAfrican cat¢sh (Clarias gariepinus)juveniles. Aquaculture 231,347^359.
Almazan-Rueda P., van Helmond A.T.M., Verreth J.A.J.& Schrama J.W. (2005) Photoperiod a¡ects growth,behaviour and stress variables in Clarias gariepinus.Journal of Fish Biology 67,1029^1039.
Almeida O.T.D. (2006) A indu¤ stria pesqueira na AmazoŒ nia.Ibama/Provarzea, Manaus-AM, Brazil p.110.
Almeida O.T., Mcgrath D.G. & Ru⁄no M.L. (2001) Thecommercial ¢sheries of the lower Amazon: an economicanalysis. Fisheries Management and Ecology 8, 253^269.
Alok D., KrishnanT.,Talwar G.P. & Garg L.C. (1998) Multipleinduced spawnings of the Indian cat¢sh Heteropneustes
fossilis (Bloch) within a prolonged spawning season.Journal of theWorld Aquaculture Society 29, 252^258.
Appelbaum S. & Kamler E. (2000) Survival, growth,metabolism and behaviour of Clarias gariepinus (Burchell1822) early stages under di¡erent light conditions.Aquacultural Engineering 22, 269^287.
Appelbaum S. & Mcgeer J.C. (1998) E¡ect of diet and lightregime on growth and survival of African cat¢sh (Clariasgariepinus) larvae and early juveniles. AquacultureNutrition 4,157^164.
AristizaŁ bal E.O. & SuaŁ rez J. (2006) E⁄ciency of co-feedingred porgy (Pagrus pagrus L.) larvae with live and com-pound diet. Revista de Biolog|¤ a Marina y Oceanograf|¤ a 41,203^208.
Baras E. & Jobling M. (2002) Dynamics of intracohortcannibalism in cultured ¢sh. Aquaculture Research 33,461^479.
Baras E.,Tissier F.,Westerloppe L., Me¤ lard C. & Philippart J.-C.(1998) Feeding in darkness alleviates density-dependentgrowth of juvenile vundu cat¢sh Heterobranchus longi¢lis(Clariidae). Aquatic Living Resources11,335^340.
Barlow C.G., Pearce M.G., Rodgers L.J. & Clayton P. (1995)E¡ects of photoperiod on growth, survival and feedingperiodicity of larval and juvenile barramundi Latescalcarifer (Bloch). Aquaculture138,159^168.
Barthem R. & Goulding M. (1997) The Cat¢sh Connection.Ecology, Migration and Conservation of Amazon Predators.Columbia University Press, NewYork, NY, USA.137pp.
Belbenoit P., Moller P., Serrier J. & Push S. (1979) Ethologicalobservations on the electric organ discharge behaviourof the electric cat¢sh, Malapterurus electricus (Pisces).Behavioral Ecology and Sociobiology 4,321^330.
Blackshaw S. & Snyder S.H. (1997) Parapinopsin, a novelcat¢sh opsin localized to the parapineal organ, de¢nes anew gene family. Journal of Neuroscience17,8083^8092.
Boeuf G. & Le Bail P.-Y. (1999) Does light have an in£uenceon ¢sh growth? Aquaculture177,129^152.
Britz P. & Pienaar A. (1992) Laboratory experiments on thee¡ect of light and cover on the behaviour and growthof African cat¢sh, Clarias gariepinus (Pisces: Clariidae).Journal of Zoology 227, 43^62.
Buitrago^SuaŁ rez U.A. & Burr B.M. (2007) Taxonomy of thecat¢sh genus Pseudoplatystoma Bleeker (Siluriformes:Pimelodidae) with recognition of eight species. Zootaxa1512,1^38.
De Leeuw R., Goos H.J.T., Richter C.J.J. & Eding E.H. (1985)Pimozide-LHRHa-induced breeding of theAfrican cat¢sh,Clarias gariepinus (Burchell). Aquaculture 44, 295^302.
Downing G. & Litvak M.K. (2002) E¡ects of light intensity,spectral composition and photoperiod on developmentand hatching of haddock (Melanogrammus aegle¢nus)embryos. Aquaculture 213, 265^278.
Ezenwaji H. (1999) The abundance and trophic biologyof Clarias albopunctatus Nichols & LaMonte, 1953(Osteichthyes: Clariidae) in a tropical £oodriver basin.Hydrobiologia 392,159^168.
Induced breeding of Pseudoplatystoma fasciatum J Nun� ez et al. Aquaculture Research, 2008, 39, 764^776
r 2008 TheAuthors774 Journal Compilationr 2008 Blackwell Publishing Ltd, Aquaculture Research, 39, 764^776

FAO (2005) Inland Fisheries management in Latin America andits contribution to food security and poverty alleviation. FAO/COPESCAL/X/05/3E. PanamaŁ , Republica de PanamaŁ .
Fielder D.S., Bardsley W.J., Allan G.L. & Pankhurst P.M.(2002) E¡ect of photoperiod on growth and survivalof snapper Pagrus auratus larvae. Aquaculture 211,135^150.
GervaŁ sio LeonardoA.F., Romagosa E., BorellaM.I. & BatlouniS.R. (2004) Induced spawning of hatchery-raisedBrazilian cat¢sh, cachara Pseudoplatystoma fasciatum(Linnaeus,1766). Aquaculture 240, 451^461.
Gilles S., Dugue¤ R. & SlembrouckJ. (2001)Manuel de productiond’alevins du silure africain Heterobranchus longi¢lis, LeTechni-cien d’Agriculture Tropicale. IRD, Maisonneuve et Larose,Paris, France128pp.
Giri S.S., Sahoo S.K., Sahu B.B., Sahu A.K., Mohanty S.N.,Mukhopadhyay P.K. & Ayyappan S. (2002) Larval survivaland growth inWallago attu (Bloch and Schneider): e¡ectsof light, photoperiod and feeding regimes. Aquaculture213,151^161.
Goos H.J.T., Joy K.P., De Leeuw R., Van Oordt P.G.W.J., VanDelft A.M.L. & Gielen J.T. (1987) The e¡ect of luteinizinghormone-releasing hormone analogue (LHRHa) incombination with di¡erent drugs with anti-dopamineand anti-serotonin properties on gonadotropin releaseand ovulation in the African cat¢sh, Clarias gariepinus.Aquaculture 63,143^156.
Hitzfelder G.M., Holt G.J., Fox J.M. & Mckee D.A. (2006) Thee¡ect of rearing density on growth and survival of Cobia,Rachycentron canadum, larvae in a closed recirculatingaquaculture system. Journal of the World AquacultureSociety 37, 204^209.
Hossain M.A.R., Batty R.S., Haylor G.S. & Beveridge M.C.M.(1999) Diel rhythms of feeding activity in African cat¢sh,Clarias gariepinus (Burchell 1822). Aquaculture Research30,901^905.
Hseu J.-R., HuangW.-B. & ChuY.-T. (2007) What causes can-nibalization-associated su¡ocation in cultured brown-marbled grouper, Epinephelus fuscoguttatus (Forsskal,1775)? Aquaculture Research 38, 1056–1060.
Kaiser H., Weyl O. & Hecht T. (1995) The e¡ect ofstocking density on growth, survival and agonisticbehaviour of African cat¢sh. Aquaculture International 3,217^225.
Kerdchuen N. & Legendre M. (1991) E¡ects of frequency andperiod of feeding on growth rate and feed e⁄ciency ofan African cat¢sh, Heterobranchus longi¢lis (Teleostei,Clariidae). Aquatic Living Resources 4, 241^248.
Kestemont P., Jourdan S., Houbart M., Melard C., PaspatisM., Fontaine P., Cuvier A., Kentouri M. & Baras E. (2003)Size heterogeneity, cannibalism and competition incultured predatory ¢sh larvae: biotic and abioticin£uences. Aquaculture 227,333^356.
Kossowski C. (1996) Perspectives de l’e¤ levage des poissons-chats (Siluroidei) en Ame¤ rique du Sud. Aquatic LivingResources 9,189^195.
Kossowski C. & Madrid F. (1985) Ensayo de la reproduccio¤ ninducida en el bagre rayado cabezo¤ n Pseudoplatystomafasciatum (Linnaeus 1766) (Pisces. Siluriformes). ActaCient|¤ ¢caVenezolana 36, 284^285.
Kossowski C. & Madrid F. (1991) Observaciones de losestadios embrionario y Iarval del bagre rayado cabezo¤ nPseudoplatystoma fasciatum (Linnaeus, 1766) (Pisces,Siluriformes). BioLlania 8, 9–15.
Legendre M., Linhart O. & Billard R. (1996) Spawning andmanagement of gametes, fertilized eggs and embryos inSiluroidei. Aquatic Living Resources 9,59^80.
Lin H.R., Van der Kraak G., Liang J.-Y., Peng C., Li G.-Y.,Lu L.-Z., Zhou X.-J., Chang M.-L. & Peter R.E. (1986)The e¡ect of LHRH analogue and drugs which blockthe e¡ects of dopamine on gonadotropin secretionand ovulation in ¢sh cultured in China. In: Aquacultureof Cyprinids (ed. by R.E.M., J. Billard), pp. 139^150. INRA,Paris, France.
Loubens G. & Pan¢li J. (2000) Biologie de Pseudoplatystomafasciatum et P. tigrinum (Teleostei: Pimelodidae) dans lebassin du Mamore¤ (Amazonie Bolivienne). IchthyologicalExploration of Freshwaters11,13^34.
Mandiki S.N.M., Babiak I., Krol J., Rasolo J.F.R. & KestemontP. (2007) How initial predator-prey ratio a¡ects intra-cohort cannibalism and growth in Eurasian perch Perca£uviatilis L larvae and juveniles under controlled condi-tions. Aquaculture 268, 149–155.
Moustakas C.T., Watanabe W.O. & Copeland K.A. (2004)Combined e¡ects of photoperiod and salinity on growth,survival, and osmoregulatory ability of larval southern£ounder Paralichthys lethostigma. Aquaculture 229,159^179.
Padilla Pe¤ rez P.P., AlcaŁ ntara Bocanegra F. & Ismin� o Orbe R.(2001) Reproduccio¤ n inducida de la doncella Pseudopla-tystoma fasciatum y desarrollo embrionario - larval. FoliaAmazonica12,141^154.
Pedro Canavate J., Zerolo R. & Fernandez-Diaz C. (2006)Feeding and development of Senegal sole (Solea senegalen-sis) larvae reared in di¡erent photoperiods. Aquaculture258,368^377.
Peter R.E., Lin H.R. & Van Der Kraak G. (1988) Inducedovulation and spawning of cultured freshwater ¢sh inChina: advances in application of Gn-RH analogues anddopamine antagonists. Aquaculture 74,1^10.
Petrere M., Barthem R.B., Co¤ rdoba E.A. & Go¤ mez B.C. (2004)Review of the large cat¢sh ¢sheries in the upperAmazonand the stock depletion of pira|¤ ba (Brachyplatystoma ¢la-mentosum Lichtenstein). Reviews in Fish Biology and Fish-eries14, 403^414.
PohlmannK., AtemaJ. & BreithauptT. (2004) The importanceof the lateral line in nocturnal predation of piscivorouscat¢sh. The Journal of Experimental Biology 207,2971^2978.
PuvanendranV. & Brown J.A. (2002) Foraging, growth andsurvival of Atlantic cod larvae reared in di¡erent light in-tensities and photoperiods. Aquaculture 214,131^151.
Aquaculture Research, 2008, 39, 764^776 Induced breeding of Pseudoplatystoma fasciatum J Nun� ez et al.
r 2008 TheAuthorsJournal Compilationr 2008 Blackwell Publishing Ltd, Aquaculture Research, 39, 764^776 775

Qin J. & Fast A.W. (1996) Size and feed dependent cannibal-ismwith juvenile snakehead Channa striatus. Aquaculture144,313^320.
Reinert T.R. & Winter K.A. (2002) Sustainability of har-vested pacu¤ (Colossoma macropomum) populations in theNorthheastern Bolivian Amazon. Conservation Biology16,1344^1351.
Ribeiro L., Engrola S. & Dinis M.T. (2005) Weaning of sene-galese sole (Solea senegalensis) postlarvae to an inert dietwith a co-feeding regime. Ciuencias Marinas 31,327^337.
Richter C.J.J., Eding E.H., Goos H.J.T., De Leeuw R., Scott A.P.& Van Oordt P.G.W.J. (1987) The e¡ect of pimozide/LHRHaand17[alpha]-hydroxyprogesterone on plasma steroid le-vels and ovulation in the African cat¢sh, Clarias gariepi-nus. Aquaculture 63,157^168.
Rodriguez J.A. (1996) Avances en la reproduccio¤ n inducidade Siluridos. Dahlia1,85^88.
Rodriguez M.A., Richardson S.E. & Lewis W.M. Jr. (1990)Nocturnal behavior and aspects of the ecology of a drift-wood cat¢sh, Entomocorus gameroi (Auchenipteridae).Biotropica 22, 435^438.
Ronzani Cerqueira V. & Macedo Brˇgger A. (2001) E¡ect oflight intensity on initial survival of fat snook (Centropo-mus parallelus, Pisces: Centropomidae Larvae. BrazilianArchives of Biology andTechnology 44,343^349.
Ru⁄no M.L. (2004) A pesca e os recursos pesqueiros na Ama-zoŒ nia brasileira. Ibama/ProVaŁ rzea, Manaus, Brazil 262pp.
Ru⁄no M.L. (2005) Gesta� o do uso dos Recursos Pesqueiros naAmazoŒ nia. IBAMA, Manaus, Brazil135pp.
Sahoo S.K., Giri S.S. & Sahu A.K. (2004) E¡ect of stockingdensity on growth and survival of Clarias batrachus
(Linn.) larvae and fry during hatchery rearing. Journal ofApplied Ichthyology 20,302^305.
SatoY., Cardoso E.L., SallumW.B. & Godinho H.P. (1997) In-duc� ao experimental da desove do surubim Pseudoplatys-toma coruscans. In: Surubim (ed. by M.O.T. Miranda), pp.69^79. IBAMA, Belo Horizonte, Brazil.
Slembrouck J., Komarudin O., Maskur H. & Legendre M.(2004) Propagation of the Indonesian Cat¢sh, PangasiusDjambal. IRD EŁ ditions^DKP, Depok, Indonesia,131pp.
Spotte S., Petry P. & Zuanon J.A.S. (2001) Experimentson the feeding behavior of the hematophagous candiru,Vandellia cf. Plazaii. Environmental Biology of Fishes 60,459^464.
Sundararaj B., Nath P. & Halberg F. (1982) Circadianmeal timing in relation to lighting schedule optimizescat¢sh body weight gain. Journal of Nutrition 112,1085^1097.
SzkudlarekM. & Zake’sŁ Z. (2007) E¡ect of stocking density on
survival and growth performance of pikeperch, Sander lu-cioperca (L.), larvae under controlled conditions. Aquacul-ture International15,67^81.
Tavares M.P.O. (1997) O surubim. In: Surubim (ed. by M.O.T.Miranda), pp.9^25. IBAMA, Belo Horizonte, Brazil.
Trotter A.J., Battaglene S.C. & Pankhurst P.M. (2003) E¡ectsof photoperiod and light intensity on initial swim bladderin£ation, growth and post-in£ation viability in culturedstriped trumpeter (Latris lineata) larvae. Aquaculture224,141^158.
Welcomme R.L. (1988) International introductions of InlandAquatic Species, FAO ¢sheries technical paper T294, FAO,Rome, Italy,318pp.
Induced breeding of Pseudoplatystoma fasciatum J Nun� ez et al. Aquaculture Research, 2008, 39, 764^776
r 2008 TheAuthors776 Journal Compilationr 2008 Blackwell Publishing Ltd, Aquaculture Research, 39, 764^776
Related Documents





![Análisis multimodelo del crecimiento de Pseudoplatystoma orinocoense en … · 2019-06-17 · realizados en las especies P. metaense del río apure [2] y Pseudoplatystoma corruscans](https://static.cupdf.com/doc/110x72/5ebceca7619b9a54f554cb91/anlisis-multimodelo-del-crecimiento-de-pseudoplatystoma-orinocoense-en-2019-06-17.jpg)





