psbA Mutation (valine 219 to isoleucine) in Poa annua resistant to metribuzin and diuron Lemma W Mengistu, 1² George W Mueller-Warrant, 2 * Aaron Liston 3 and Reed E Barker 2 1 Crop and Soil Science Department, Oregon State University, Corvallis, OR 97330 USA 2 USDA-ARS, National Forage Seed Production Research Center, 3450 SW Campus Way, Corvallis, OR 97331-7102 USA 3 Botany and Plant Pathology Department, Oregon State University, Corvallis, OR 97330 USA Abstract: The herbicide-binding region of the chloroplast psbA gene from a total of 20 biotypes of Poa annua L resistant and susceptible to metribuzin and diuron was selectively amplified using PCR. Sequence analysis of the fragment from six herbicide-resistant biotypes of P annua exhibited a substitution from valine to isoleucine at position 219 of the D1 protein encoded by the psbA gene. This is the same mutation as reported for Chlamydomonas and Synechococcus through site-directed mutagenesis and in cell cultures of Chenopodium rubrum L. To our knowledge this is the first report of a higher plant exhibiting resistance in the field to photosystem II inhibitors due to a psbA mutation other than at position 264. The existence of additional biotypes of P annua resistant to diuron or metribuzin but lacking mutation in the herbicide-binding region indicates that resistance to these herbicides can also be attained by other mechanisms. # 2000 Society of Chemical Industry Keywords: herbicide resistance; grass-seed production; field selection; diuron; metribuzin 1 INTRODUCTION A range of herbicides, including s-triazines, substituted ureas, and phenolic derivatives, block the photosyn- thetic electron transport chain on the reducing side of photosystem II (PS II). 1,2 PS II herbicides act by displacing the secondary plastoquinone Q B from its binding site in PS II. 3 The protein target of the PS II herbicides has been identified as the D1 polypeptide subunit of PS II, also called the herbicide or Q B binding protein. 4 The D1 polypeptide is encoded by the chloroplast psbA gene, 5 and is highly conserved in plants, algae and cyanobacteria. 6 In higher plants the chloroplast psbA gene is located in the large single copy region close to the left junction of the inverted repeat. 7 The first report of herbicide resistance to a photo- synthesis inhibitor involved a triazine herbicide, 8 and since then biotypes of more than 60 species have been documented to possess triazine resistance. 9 The 1995/ 96 International Survey of Herbicide Resistant Weeds revealed 183 herbicide-resistant weed biotypes (124 different species) in 42 countries, and 32% of these were triazine-resistant. 10 Resistance to triazine herbi- cides has been attributed either to a rapid metabolic detoxification of the herbicide 11 or to a modified site of action that prevents herbicide binding within the chloroplast. 12 Susceptible as well as resistant biotypes of some weed species can rapidly metabolize atrazine via hydroxylation reactions in the plant’s metab- olism. 12 Triazine-resistant weeds, however, degrade atrazine at a much lower rate than maize (Zea mays L). 13 Triazine resistance is generally target-site based, resulting from a modification at the herbicide binding site, the D1 protein of PS II. 14 The substituted urea herbicides, although chemically distinct from triazines, are also potent PS II inhibitors 14 and bind at overlapping but not identical sites with the triazines. 15 Biotypes highly resistant to triazine herbicides as a result of a modified D1 protein fail to show resistance to substituted urea herbicides 14 because mutation Ser 264 to Gly providing resistance to triazine herbicides does not affect binding of substituted urea herbi- cides. 2,15 Triazine-resistant plants, however, have been reported to show resistance to other PS II inhibitors such as triazinones, uracils and pyridazi- nones. 16,17 The psbA gene from several organisms has been sequenced, and, quite frequently, resistance by photo- synthetic organisms to PS II herbicides, like triazi- nones, triazines, and DCMU (diuron), results from mutations in the psbA gene, leading to an amino acid exchange in the Q B binding protein. In higher plants a (Received 21 October 1999; accepted 1 November 1999) * Correspondence to: George W Mueller-Warrant, USDA-ARS, National Forage Seed Production Research Center, 3450 SW Campus Way, Corvallis, OR 97331-7102 USA E-mail: [email protected] ² Current address: Department of Plant Sciences, North Dakota State University, Fargo, ND 58105 USA Contract/grant sponsor: Oregon Seed Council # 2000 Society of Chemical Industry. Pest Manag Sci 1526–498X/2000/$17.50 209 Pest Management Science Pest Manag Sci 56:209–217 (2000)

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Pest Management Science Pest Manag Sci 56:209±217 (2000)
psbA Mutation (valine 219 to isoleucine) in Poaannua resistant to metribuzin and diuronLemma W Mengistu,1† George W Mueller-Warrant,2* Aaron Liston3 andReed E Barker21Crop and Soil Science Department, Oregon State University, Corvallis, OR 97330 USA2USDA-ARS, National Forage Seed Production Research Center, 3450 SW Campus Way, Corvallis, OR 97331-7102 USA3Botany and Plant Pathology Department, Oregon State University, Corvallis, OR 97330 USA
(Rec
* CoCorvE-ma† CurCont
# 2
Abstract: The herbicide-binding region of the chloroplast psbA gene from a total of 20 biotypes of Poa
annua L resistant and susceptible to metribuzin and diuron was selectively ampli®ed using PCR.
Sequence analysis of the fragment from six herbicide-resistant biotypes of P annua exhibited a
substitution from valine to isoleucine at position 219 of the D1 protein encoded by the psbA gene. This
is the same mutation as reported for Chlamydomonas and Synechococcus through site-directed
mutagenesis and in cell cultures of Chenopodium rubrum L. To our knowledge this is the ®rst report of
a higher plant exhibiting resistance in the ®eld to photosystem II inhibitors due to a psbA mutation
other than at position 264. The existence of additional biotypes of P annua resistant to diuron or
metribuzin but lacking mutation in the herbicide-binding region indicates that resistance to these
herbicides can also be attained by other mechanisms.
# 2000 Society of Chemical Industry
Keywords: herbicide resistance; grass-seed production; ®eld selection; diuron; metribuzin
1 INTRODUCTIONA range of herbicides, including s-triazines, substituted
ureas, and phenolic derivatives, block the photosyn-
thetic electron transport chain on the reducing side of
photosystem II (PS II).1,2 PS II herbicides act by
displacing the secondary plastoquinone QB from its
binding site in PS II.3 The protein target of the PS II
herbicides has been identi®ed as the D1 polypeptide
subunit of PS II, also called the herbicide or QB
binding protein.4 The D1 polypeptide is encoded by
the chloroplast psbA gene,5 and is highly conserved in
plants, algae and cyanobacteria.6 In higher plants the
chloroplast psbA gene is located in the large single copy
region close to the left junction of the inverted repeat.7
The ®rst report of herbicide resistance to a photo-
synthesis inhibitor involved a triazine herbicide,8 and
since then biotypes of more than 60 species have been
documented to possess triazine resistance.9 The 1995/
96 International Survey of Herbicide Resistant Weeds
revealed 183 herbicide-resistant weed biotypes (124
different species) in 42 countries, and 32% of these
were triazine-resistant.10 Resistance to triazine herbi-
cides has been attributed either to a rapid metabolic
detoxi®cation of the herbicide11 or to a modi®ed site of
action that prevents herbicide binding within the
chloroplast.12 Susceptible as well as resistant biotypes
eived 21 October 1999; accepted 1 November 1999)
rrespondence to: George W Mueller-Warrant, USDA-ARS, National Fallis, OR 97331-7102 USAil: [email protected] address: Department of Plant Sciences, North Dakota State Univract/grant sponsor: Oregon Seed Council
000 Society of Chemical Industry. Pest Manag Sci 1526±498X/2
of some weed species can rapidly metabolize atrazine
via hydroxylation reactions in the plant's metab-
olism.12 Triazine-resistant weeds, however, degrade
atrazine at a much lower rate than maize (Zea maysL).13
Triazine resistance is generally target-site based,
resulting from a modi®cation at the herbicide binding
site, the D1 protein of PS II.14 The substituted urea
herbicides, although chemically distinct from triazines,
are also potent PS II inhibitors14 and bind at
overlapping but not identical sites with the triazines.15
Biotypes highly resistant to triazine herbicides as a
result of a modi®ed D1 protein fail to show resistance
to substituted urea herbicides14 because mutation
Ser264 to Gly providing resistance to triazine herbicides
does not affect binding of substituted urea herbi-
cides.2,15 Triazine-resistant plants, however, have
been reported to show resistance to other PS II
inhibitors such as triazinones, uracils and pyridazi-
nones.16,17
The psbA gene from several organisms has been
sequenced, and, quite frequently, resistance by photo-
synthetic organisms to PS II herbicides, like triazi-
nones, triazines, and DCMU (diuron), results from
mutations in the psbA gene, leading to an amino acid
exchange in the QB binding protein. In higher plants a
orage Seed Production Research Center, 3450 SW Campus Way,
ersity, Fargo, ND 58105 USA
000/$17.50 209

LW Mengistu et al
mutation affecting amino acid 264 (serine) leads to
resistance against triazine herbicides, and in algae,
mutations in any of positions 215, 219, 255, 256 or
275 are known to confer resistance to several classes of
herbicides.18
Double and triple mutants of Chenopodium rubrum L
cell cultures were characterized where a valine to
isoleucine change at position 219 was decisive for
metribuzin resistance, although there were some other
changes between amino acid residues 209 and 291.
For example, metribuzin-resistant cell cultures pos-
sessed psbA mutations in most cases at 219 and 251, or
at 219, 251 and 256, or at 219, 229 and 270, or at 219,
220 and 270, or at 219, 251 and 272. All these changes
resulted in resistance to metribuzin. None of the
mutant cell cultures possessed the classical ®eld
mutation at residue 264.18 Most of the mutations in
the D1 protein leading to herbicide resistance in
cyanobacteria and green algae were identi®ed by
randomly induced mutagenesis or by site-directed
gene technology methods for the psbA gene.19
A detailed model of the herbicide-binding region in
PS II has been developed.20 Based on this model, 17
amino acids of the psbA gene are in contact with the QB
binding site. These include: Phe211, Met214, His215,
Leu218, Val219, Thr237, Ile248, Ala251, His252, Phe255,
Gly256, Ala263, Ser264, Phe265, Asn266, Ser268 and
Leu275. The eight site mutations including Val219 that
have been reported to cause resistance to herbicides all
involve residues that have contact with QB. The
relative importance of each residue in herbicide
binding has been further characterized.20 For exam-
ple, mutation Ser264 had greatest impact on binding of
most PS-II-inhibiting herbicides. Val219 and Leu275
were characterized as being located peripherally to the
QB binding site, as compared to the other amino acids
found nearer the center. Mutations to the more bulky
amino acids Ile219 or Phe275 will only touch those
inhibitors extending into this peripheral space. As a
result, the Val219 to Ile mutant showed R/S I50 ratios of
20, 2, 2, and 3 for metribuzin, atrazine, s-triazine and
s-thiazole, respectively.20
High levels of resistance to atrazine have been
correlated with a single amino acid substitution (Ser to
Gly) in higher plants at position 264 in the D1
protein.21±23 A change of Ser to Ala at the same
position in this protein is correlated with lower levels of
resistance to atrazine and diuron in algal mutants24
and a cyanobacterial transformant.25 In higher plants
spontaneous herbicide resistance has been found in
the ®eld in various weed plants.21,26,27 In all reported
cases involving the psbA gene, a Ser264 to Gly change is
responsible for the acquired resistance.28 This muta-
tion is reported for dicotyledenous species such as
Amaranthus hybridus L,21,29Solanum nigrum L,23 Che-nopodium album L,30,31 and Senecio vulgaris L.28 The
only monocotyledenous species with a psbA gene
mutation at residue 264 giving resistance to atrazine
is Poa annua L.32 This mutation in higher plants is
accompanied by a slowing down of electron transfer
210
between the primary QA and secondary QB quinones
of PS II reaction centers,33±35 leading to an increased
susceptibility to photoinhibition in the resistant
biotypes.36 The triazine-resistant mutants display a
marked decrease in growth rate at normal light
intensities37 and a reduction in competitive ®tness.38
However, mutations of the D1 at positions 219 and
275 have been demonstrated to have no role in
electron transfer system of PS II.6 A study conducted
on the molecular basis of resistance to atrazine and
cross-resistance to diuron used uniparental mutants of
Chlamydomonas reinhardi Dang resistant to diuron
(strain Dr2) and to atrazine (strain Ar207) that
possessed Val219 to Ile and Phe255 to Tyr, respec-
tively.6 Strain Dr2 showed a two-fold resistance to
atrazine and a 15-fold resistance to diuron, while strain
Ar207 had a 15-fold resistance to atrazine and a 0.5-
fold resistance (two-fold increased susceptibility) to
diuron. Electron transport for these two mutants was
normal, as opposed to Ser264 to Ala or Ser264 to Gly
mutations that alter the electron transport system.6 An
extended study of the mutant strain Dr-18 of Creinhardi 39 revealed that the mutant required 10-fold
higher concentrations of DCMU and CMU to show
the same degree of inhibition as the wild type. This
mutant was later sequenced, and determined to
consist of a Val219 to Ile exchange in the psbA gene.6
The resistance to PS II inhibitors due to alteration of
the psbA gene is maternally inherited. However,
hybrids with resistance from paternal plastids were
found at a frequency of 0.2 to 2%.40
Poa annua is one of the worst weed problems
confronting grass-seed growers in the Willamette
Valley of western Oregon. Its presence in harvested
seeds complicates seed cleaning and reduces crop
value if seed cannot be cleaned to `Poa-free' status.41
Over 20 populations of P annua in grass ®elds of
Oregon have been shown to be ethofumesate-resistant,
triazine-resistant and urea herbicide-resistant.10 Sev-
eral of the P annua populations have shown multiple
resistance to atrazine, diuron, terbacil and ethofume-
sate,41±43 with greenhouse experiments indicating a
10-fold increase in resistance to both atrazine and
diuron.42 Managing herbicide-resistant weeds requires
a good understanding of the genetics of resistance.
Understanding the mechanisms of resistance involved
in these populations of P annua for various herbicides
is vital. Wise management decisions will require full
understanding of the mechanisms of resistance, the
gene or genes involved, inheritance of resistance and
gene ¯ow, as well as the details of the biology of the
species. This study therefore was designed to ful®l the
objective of understanding the mechanism of resis-
tance involved in populations of P annua resistant to
diuron and metribuzin in western Oregon grass-seed
crops.
2 MATERIALS AND METHODSSeeds for these experiments were obtained from
Pest Manag Sci 56:209±217 (2000)

psbA mutation in Poa annua resistant to metribuzin and diuron
greenhouse-grown P annua plants collected in 1994
and 1995 from grass-seed ®elds of western Oregon for
studies of genetic diversity.44 Seeds of several addi-
tional herbicide-resistant types and one susceptible
type were obtained from Dr Carol Mallory-Smith
(Dept Crop & Soil Sci, Oregon State Univ, Corvallis,
OR).
2.1 Greenhouse experimentsIn the greenhouse experiments, we tested the tolerance
to ®eld rates of diuron and metribuzin using 225
accessions of P annua. Seeds from single plants of the
225 accessions were planted in rows using ¯ats in the
greenhouse. Every ¯at consisted of ten rows, and each
row was a single accession of P annua. Eighty of 225
accessions were tested for their reactions both to
metribuzin and diuron, while the other 145 accessions
were only tested for diuron resistance because quan-
tities of seed were limited.
Every accession's seedlings were counted after
emergence and recorded before herbicide treatment.
On 30 May 1997, seedlings in the two-leaf stage were
treated with diuron at 1.1kg AI haÿ1 for all 225
accessions and metribuzin at 0.37kg AI haÿ1 for 80
accessions. Two ¯ats from the metribuzin treatments
were divided into halves, and one half was covered
with a plastic sheet while spraying the herbicide, to
serve as an untreated control. Other untreated controls
from earlier plantings were also grown in the green-
house for comparisons. The treated seedlings were
kept in the greenhouse at 12h supplemental light,
21°C day, 15°C night and watered daily. Seedlings
were counted again at two and three weeks after
herbicide treatment, and the progeny classes and
accession classes were determined based on their
reaction to the herbicides as (R) resistant, (I) inter-
mediate and (S) susceptible. Progeny were scored as
resistant if uninjured by herbicide, intermediate if
injured but alive, and susceptible if killed. Accessions
were scored as resistant if any progeny were resistant,
susceptible if all progeny were killed, and otherwise
intermediate. After identifying the response of P annuaaccessions to diuron and metribuzin, the mechanism
of resistance was studied through DNA sequence
analysis of the psbA gene.
2.2 DNA extractionWe extracted total DNA using a modi®ed procedure of
Figure 1. A polyacrylamide gel electrophoresisindicating the amplified 933 base pair fragmentDNA of the herbicide-binding region of the psbAgene from Poa annua accessions. The two primersused to amplify this DNA fragment were 5'GGATGGTTTGGTGTTTTG 3' and 5'TAGAGGGAAGTTGTGAGC 3'. The PCR reactionconditions are explained in the text. Numbers (1–7)are P annua samples and M is the marker.
Pest Manag Sci 56:209±217 (2000)
a rapid one-step extraction of DNA (ROSE) buffer
[Tris HCl (pH 8; 10mM), EDTA (pH 8; 312mM),
lauryl sarkocyl (10g litreÿ1), PVPP (1% per 50ml ®nal
volume) and b-mercaptoethanol (2g litreÿ1)] as ex-
plained by Steiner et al. 45 The following steps were
added to clean DNA samples further and to quantify
the DNA yield. The supernatant was placed in clean
tubes, two-thirds volume (330ml) cold isopropanol
was added, each sample was then well mixed and held
at ÿ20°C for 10min. The samples were then
centrifuged for 10min at 12000rev minÿ1 and pellets
were carefully saved in the tube while the supernatant
was decanted. The pellets were washed with etha-
nol�water (80�20 by volume, 300ml) by spinning for
5min, then air-dried. The dried pellets were resus-
pended in Tris base�EDTA pH 8(TE; 400ml).
RNAse (0.5mgmlÿ1, 8ml; ®nal concentration
10mgmlÿ1) was added to the samples and incubated
for 30min at 37°C. The DNA was precipitated by
rocking the tubes after adding ammonium acetate
(7.5 M, 200ml) absolute ethanol (1.2 ml), and then
held for 10min at ÿ20°C. Samples were ®nally spun
for 10min, decanted, washed, and air-dried. The
pellets were then resuspended in TE (80ml) and kept
in storage at ÿ20°C until needed for ampli®cation.
DNA was quanti®ed for every sample using a
¯uorometer, and the DNA samples were diluted to
®nal concentrations of 10ngmlÿ1.
2.3 PCR conditions and DNA amplificationTwo primers were designed and synthesized based on
homologous regions of sequences available from
GenBank for the chloroplast psbA gene of Arabidopsisthaliana (GI:515373) and Nicotiana plumbaginifolia(GI:11754). These primers should amplify a 933 base
pair fragment of the chloroplast psbA gene including
the herbicide-binding region. The primer sequence 5'GGATGGTTTGGTGTTTTG 3' corresponded to
bases 91±108 of the psbA 5' region. The second primer
5' TAGAGGGAAGTTGTGAGC 3' corresponded to
bases 1006±1023 of the 3' region of the DNA
sequence. Both primers were used to PCR-amplify
the target region from total DNA (Fig 1). The same
two primers were also used for sequencing the
fragment in separate reactions, and results were
aligned during analysis. Another primer was synthe-
sized based on the speci®c sequence we obtained from
our P annua, and this primer was only used for
211

LW Mengistu et al
sequencing the fragment. The sequence of the third
primer was (5' CTCCTGTTGCAGCTGCTACT
3'), which starts just before the region of amino acid
residue 150 and corresponds to bases 446±465 of the
psb A sequence. This primer was designed to give a
very clear sequence of the herbicide-binding region.
PCR was performed in a 100ml reaction volume with
90ml of reaction mix [1� Stoffel buffer, 3.75mM
MgCl2, 50mM of each dNTP, 1.0pM of each primer,
and 1 unit Taq Stoffel fragment] aliquotted into a 96-
well assay plate and then overlaid with three drops of
mineral oil. Finally, 10ml of 10ngmlÿ1 diluted DNA
sample was added to the reaction mixture under the
oil. The thermal cycling program started with dena-
turation for 7min at 94°C followed by 43 cycles of 30-
s denaturation at 47C, a 1°C per 3-s ramp, and
extension at 72°C for 2min. A ®nal extension at 72°Cfor 5min concluded the DNA ampli®cation. After
ampli®cation the samples were kept at 4°C for a short
period (<24h) or at ÿ20°C for longer periods until
electrophoresis.
2.4 Electrophoresis, DNA extraction from gels andpurificationThe PCR product was run in both TBE (Tris
base�boric acid�EDTA) agarose gel electrophoresis
and polyacrylamide gel electrophoresis. Polyacryla-
mide gels were made from Tris-HCl (pH 8.8),
acrylamide (7.5% ®nal), ammonium persulfate, and
TEMED (N,N,N ',N '-tetramethylethylenediamine),
and resulted in the approximately 933 base pair single
fragment seen in Fig 1, along with several very faint
bands of lower molecular mass. DNA was extracted
from the 933 base pair fragment on the agarose gel,
and puri®ed for sequencing by following the QIAquick
Gel Extraction protocol (QIAGEN Inc, Valencia,
California 1997) and Kit (buffers, tubes, ®lters). The
puri®ed product for each sample was submitted to the
Central Services Laboratory at Oregon State Uni-
versity. Multiple sequences were obtained for most
accessions. Analyses of the sequences were made ®rst
by editing multiple sequences, and once a consensus
sequence was obtained homology tests were made with
known psbA sequences. Translation was made using
Expasy, an Internet translation tool [http://www.
expasy.ch/tools/dna.html].
3 RESULTS3.1 Poa annua accessions reaction to metribuzinand diuron in greenhouseA large number of P annua seedlings were susceptible
to diuron and metribuzin, with treatments causing 46
to 94% and 75 to 96% lethality, respectively (Table 1).
However, a large percentage of the accessions con-
tained at least one individual (out of approximately 50
tested) highly resistant to diuron. Some individual
accessions were relatively uniform in their response to
diuron, while others possessed resistant, intermediate,
and susceptible types, suggesting that many of these
212
populations were still segregating for resistance to
diuron. In general, these accessions were more
sensitive to metribuzin than to diuron. However, three
accessions from the Bowers farm near Harrisburg,
Oregon, possessed individuals highly resistant to
metribuzin, and several accessions at two other sites
were intermediate in their response to metribuzin.
Only one accession (#145) out of 80 tested was
resistant to both diuron and metribuzin. Its reaction
may have been maternally inherited because all the
progeny from the single plant parent showed uniform
reaction to the herbicides. Since all 225 of the
accessions were tested for diuron while only 80 were
also tested for metribuzin, we presume some of the
diuron-resistant accessions were also resistant to
metribuzin. The uniformity of their response to diuron
across the rows of progenies was similar to that of
accession #145, indicating that these other accessions
were similar to accession #145.
3.2 Sequence analysisInitial attempts to sequence the 933 base pair fragment
of the psbA gene from our accessions used either one or
the other of the two 18-mer primers from PCR
ampli®cation as their sequencing template. The
DNA sequencer used (ABI Prism2 Model 377) gave
no results in the template region. Nucleotide identi-
®cation became ambiguous somewhere between 300
and 600 bases after the primer in any individual run,
sometimes making it dif®cult to align the 5' and 3'sequences and be con®dent of the results in the
herbicide-binding region (amino acid residues 211 to
275, bases 541 to 735 from the 5' end or bases 199 to
393 from the 3' end of our fragment). In order to
obtain clearer results in this critical region, we
generated a new (nested) primer for use as a
sequencing template based on the sequence already
obtained for bases 446 to 465 of the psbA gene (bases
356 to 375 from the 5' end of our fragment). This third
primer usually gave good readings all the way out to
the 3' end of the fragment (psbA codon 1023).
The only detectable difference in the DNA se-
quences of the 20 accessions occurred at codon 655,
which changed from wild-type guanine to mutant
adenine. This mutation was present in samples
extracted from three separate plants grown out from
seed produced by accession #145. The mutation was
detectable in DNA sequence runs using each of the
three primer templates for four separate accessions.
The mutation was detected using the nested primer in
two other accessions. Wild-type in the herbicide-
binding region was con®rmed in nine accessions with
the nested primer, two accessions with the 3' primer,
and three accessions with the 5' primer. For the 15
accessions sequenced with the nested primer, the high
quality of the data strongly implies that there were no
psbA differences among these accessions from codon
469 to 1023, except for the mutation at 655. The seven
accessions sequenced using the 5' PCR primer were
reliably sequenced (and showed no differences) from
Pest Manag Sci 56:209±217 (2000)
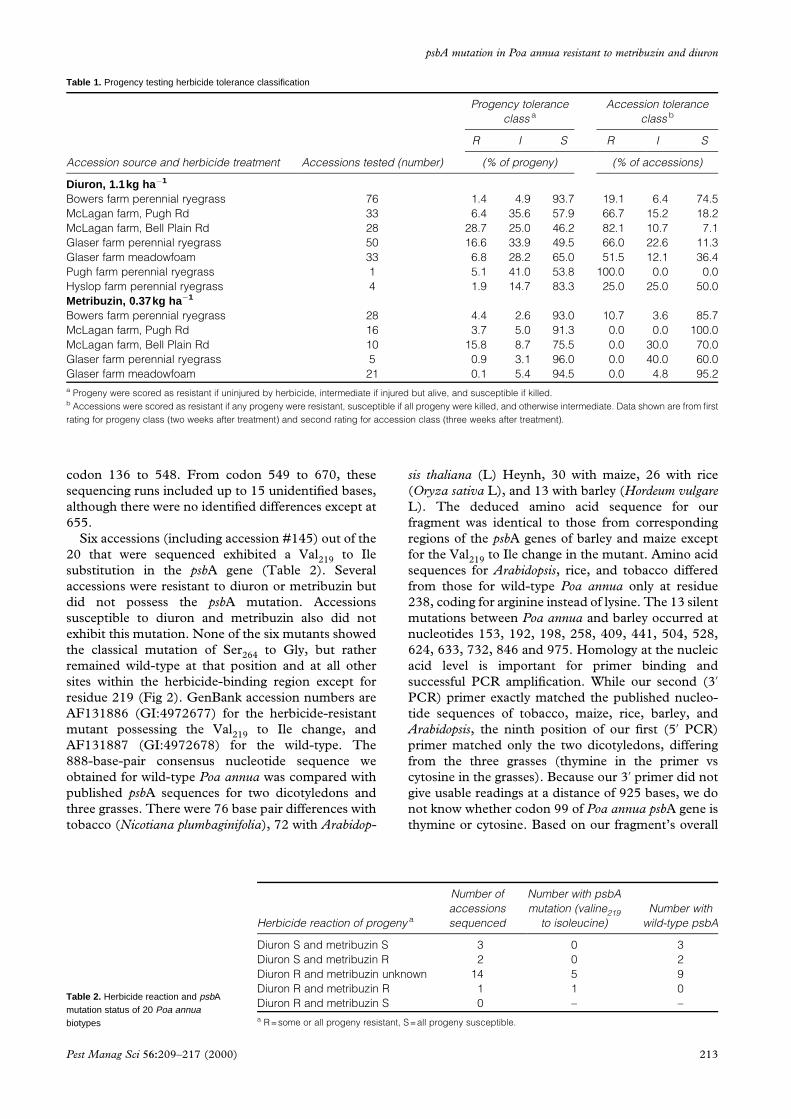
Table 1. Progency testing herbicide tolerance classification
Accession source and herbicide treatment Accessions tested (number)
Progency tolerance
class a
Accession tolerance
class b
R I S R I S
(% of progeny) (% of accessions)
Diuron, 1.1kg ha ÿ1
Bowers farm perennial ryegrass 76 1.4 4.9 93.7 19.1 6.4 74.5
McLagan farm, Pugh Rd 33 6.4 35.6 57.9 66.7 15.2 18.2
McLagan farm, Bell Plain Rd 28 28.7 25.0 46.2 82.1 10.7 7.1
Glaser farm perennial ryegrass 50 16.6 33.9 49.5 66.0 22.6 11.3
Glaser farm meadowfoam 33 6.8 28.2 65.0 51.5 12.1 36.4
Pugh farm perennial ryegrass 1 5.1 41.0 53.8 100.0 0.0 0.0
Hyslop farm perennial ryegrass 4 1.9 14.7 83.3 25.0 25.0 50.0
Metribuzin, 0.37kg ha ÿ1
Bowers farm perennial ryegrass 28 4.4 2.6 93.0 10.7 3.6 85.7
McLagan farm, Pugh Rd 16 3.7 5.0 91.3 0.0 0.0 100.0
McLagan farm, Bell Plain Rd 10 15.8 8.7 75.5 0.0 30.0 70.0
Glaser farm perennial ryegrass 5 0.9 3.1 96.0 0.0 40.0 60.0
Glaser farm meadowfoam 21 0.1 5.4 94.5 0.0 4.8 95.2
a Progeny were scored as resistant if uninjured by herbicide, intermediate if injured but alive, and susceptible if killed.b Accessions were scored as resistant if any progeny were resistant, susceptible if all progeny were killed, and otherwise intermediate. Data shown are from ®rst
rating for progeny class (two weeks after treatment) and second rating for accession class (three weeks after treatment).
psbA mutation in Poa annua resistant to metribuzin and diuron
codon 136 to 548. From codon 549 to 670, these
sequencing runs included up to 15 unidenti®ed bases,
although there were no identi®ed differences except at
655.
Six accessions (including accession #145) out of the
20 that were sequenced exhibited a Val219 to Ile
substitution in the psbA gene (Table 2). Several
accessions were resistant to diuron or metribuzin but
did not possess the psbA mutation. Accessions
susceptible to diuron and metribuzin also did not
exhibit this mutation. None of the six mutants showed
the classical mutation of Ser264 to Gly, but rather
remained wild-type at that position and at all other
sites within the herbicide-binding region except for
residue 219 (Fig 2). GenBank accession numbers are
AF131886 (GI:4972677) for the herbicide-resistant
mutant possessing the Val219 to Ile change, and
AF131887 (GI:4972678) for the wild-type. The
888-base-pair consensus nucleotide sequence we
obtained for wild-type Poa annua was compared with
published psbA sequences for two dicotyledons and
three grasses. There were 76 base pair differences with
tobacco (Nicotiana plumbaginifolia), 72 with Arabidop-
Table 2. Herbicide reaction and psbAmutation status of 20 Poa annuabiotypes
Herbicide reaction of progeny
Diuron S and metribuzin S
Diuron S and metribuzin R
Diuron R and metribuzin unkn
Diuron R and metribuzin R
Diuron R and metribuzin S
a R=some or all progeny resistant, S
Pest Manag Sci 56:209±217 (2000)
sis thaliana (L) Heynh, 30 with maize, 26 with rice
(Oryza sativa L), and 13 with barley (Hordeum vulgareL). The deduced amino acid sequence for our
fragment was identical to those from corresponding
regions of the psbA genes of barley and maize except
for the Val219 to Ile change in the mutant. Amino acid
sequences for Arabidopsis, rice, and tobacco differed
from those for wild-type Poa annua only at residue
238, coding for arginine instead of lysine. The 13 silent
mutations between Poa annua and barley occurred at
nucleotides 153, 192, 198, 258, 409, 441, 504, 528,
624, 633, 732, 846 and 975. Homology at the nucleic
acid level is important for primer binding and
successful PCR ampli®cation. While our second (3'PCR) primer exactly matched the published nucleo-
tide sequences of tobacco, maize, rice, barley, and
Arabidopsis, the ninth position of our ®rst (5' PCR)
primer matched only the two dicotyledons, differing
from the three grasses (thymine in the primer vs
cytosine in the grasses). Because our 3' primer did not
give usable readings at a distance of 925 bases, we do
not know whether codon 99 of Poa annua psbA gene is
thymine or cytosine. Based on our fragment's overall
a
Number of
accessions
sequenced
Number with psbA
mutation (valine219
to isoleucine)
Number with
wild-type psbA
3 0 3
2 0 2
own 14 5 9
1 1 0
0 ± ±
=all progeny susceptible.
213

Figure 2. Consensus nucleotide sequence for psbA gene fragments of 14 wild-type Poa annua covering the herbicide-binding region of the D1 protein (middlelines), nucleotide sequence for the barley psbA gene where it differs from Poa annua (upper lines), and deduced amino acids (lower lines). The metribuzin- anddiuron-resistant mutant psbA sequence possessed a change of Val219 to Ile at the amino acid residue and nucleotide position (in the box) as a result of a changefrom GTA to ATA. Numbers at the right refer to the residue number of amino acids from the entire psbA gene. The underlined bases correspond to the third primer(5' CTCCTGTTGCAGCTGCTACT 3') we used for sequencing close to the herbicide binding region. We present here only the region that resulted in consistentsequences for all accessions, covering from amino acid residues 46 to 341 of the entire chloroplast psbA gene. Single letter amino acid code: A: alanine, C:cysteine, D: aspartate, E: glutamate, F: phenylalanine, G: glycine, H: histidine, I: isoleucine, K: lysine, L: leucine, M: methionine, N: asparagine, P: proline, Q:glutamine, R: arginine, S: serine, T: threonine, V: valine, Y: tyrosine, W: tryptophan.
LW Mengistu et al
closer homology to the grasses, however, and particu-
larly to barley, it seems probable that codon 99 in the
Poa annua psbA gene is cytosine, and that we had a
mismatch at the ninth position of our 5' PCR primer.
The six accessions that showed the Val219 to Ile
mutation came from two ®eld sites located 10 km
apart: two accessions from Glaser perennial ryegrass
and four from McLagan Bell Plain Rd perennial
214
ryegrass. These sites were well known for herbicide-
resistant P annua. Using randomly ampli®ed poly-
morphic DNA (RAPDs), we found that herbicide
selection pressure at these ®elds has reduced the
among population genetic variation in P annua.44 The
six accessions possessing the Val219 to Ile mutation
showed ®ve different RAPD banding patterns, with
the two accessions from Glaser perennial ryegrass
Pest Manag Sci 56:209±217 (2000)

psbA mutation in Poa annua resistant to metribuzin and diuron
having the same banding pattern.44 Interestingly, one
of the RAPD banding patterns was shared by a psbA
mutant at McLagan Bell Plain Rd perennial ryegrass
and psbA wild-types at Glaser perennial ryegrass and
McLagan Pugh Rd perennial ryegrass. RAPD banding
patterns of the six psbA mutants occurred in an
average of 14 individuals over the entire collection,
®ve time more frequently than those of the average
accession.44
4 DISCUSSIONFailure to ®nd psbA mutants of higher plants in the
®eld other than Ser264 to Gly has long puzzled
researchers. Our sequence analysis on herbicide-
resistant P annua accessions revealed that there are
biotypes of higher plants with site mutations other
than residue 264, and these biotypes resist PS II
inhibitors at ®eld rates. In addition to Poa annua's
short life cycle and signi®cant amount of outcrossing,
herbicide rates in western Oregon grass-seed crops
may have been nearly ideal for selection of this psbA
mutation. Grass-seed growers have applied 1 to
1.5kghaÿ1 atrazine, 2.2kghaÿ1 simazine and 1.1 to
2.2kg haÿ1 diuron for approximately 30 years. These
rates are limited by the tolerance of crops such as
perennial ryegrass to these herbicides. We do not yet
know whether the Val219 to Ile mutant accessions also
possessed additional resistance from the nuclear-
controlled mechanism of resistance apparently present
in other accessions. Selection in the ®eld may have
played a role in combining multiple resistant mechan-
isms into single individuals.
The number of mutant types within the accessions
we sequenced is far higher than expected by mutation
rates alone of about 10ÿ5 per generation. For example,
out of 20 P annua accessions we sequenced for psbA,
three were susceptible to metribuzin and diuron, 14
were resistant to diuron with reaction to metribuzin
untested, two were resistant to metribuzin but
susceptible to diuron, and one was resistant to both
metribuzin and diuron. Of the 14 diuron-resistant,
metribuzin-untested biotypes, ®ve exhibited a Val219
to Ile substitution in their psbA gene. Sequences from
the remaining nine diuron-resistant biotypes and the
two metribuzin-resistant biotypes were wild-type in
their psbA gene. The metribuzin-and diuron-resistant
biotype possessed the mutation at the position we have
indicated. It is likely that more psbA mutants would
have been found had we sequenced all diuron-resistant
accessions within our population. We suspect that the
rate and pattern of herbicide use by western Oregon
grass-seed growers must have provided suitable selec-
tion pressure to select these mutants. Higher rates
might have killed the mutants, while lower rates might
have allowed the susceptible types to survive.
The existence of both maternally inherited, single-
gene, site-mutation herbicide resistance and appar-
ently nuclear controlled resistance for the same
herbicides within populations of P annua in western
Pest Manag Sci 56:209±217 (2000)
Oregon grass-seed crops will complicate the problem
of addressing herbicide resistance. Gressel et al46
emphasized that `the tendency to cut dose rate is
increasing resistance due to multiple cumulative
events (polygenic, ampli®cation, or sequential muta-
tions within a gene).' The dilemma here is whether
crop rotation or rate of herbicide use should target the
major gene or minor genes with cumulative effects,
since both types exist in the same populations.
The mutation of the herbicide-binding D1 protein
at residue 264 has been characterized to cause
inhibition of the electron transport system, leading
the mutant to be less ®t than susceptible biotypes,
especially without herbicide use. However, the Val219
to Ile mutation we report now has been characterized
as causing resistance to herbicides but having no effect
on electron transfer. Because the mutant may be
equally ®t as the wild-type, use of no herbicide or low
selection pressure of herbicide may fail to reduce the
frequency of this mutant in the populations. Another
aspect of the mutation of Val219 to Ile is its capability of
resisting more than one kind of herbicide, as it is
reported to result in resistance to metribuzin, atrazine,
diuron and ioxynil in various organisms. Unlike the
Ser264 to Gly change in other weed species that exhibit
negative cross-resistance to diuron, the absence of this
in Val219 to Ile mutants is a problem to growers, as it
eliminates several alternative herbicides.
The degree of resistance by site mutation of Val219
to Ile has been reported to range from 10- to 20-fold
which is close to the level of resistance found via
detoxi®cation of herbicides by enzyme action through
polygenic effects. This would be an advantage if high
doses of herbicides could be used to delay herbicide
resistance due to minor genes with cumulative effects
and mutants of the kind we found. However, grass-
seed growers of western Oregon usually grow per-
ennial ryegrass or tall fescue that would be susceptible
to herbicides themselves should rates be increased
substantially above those currently used. One option
may be to rotate to crops like orchardgrass that better
tolerate those herbicides or tolerate other herbicides
such as pronamide, and use higher doses of herbicides
to eliminate resistant weeds from their ®elds. Alter-
native crops like meadowfoam (Limnanthes species) or
clovers (Trifolium species) are another option, allowing
use of herbicides with different modes of action. The
alternative is that continued increases in populations
of P annua biotypes with multiple herbicide resistance
in western Oregon grass-seed crops may affect seed
producers and companies, leading to shifts in produc-
tion, increased seed cleaning costs, and decreased
pro®tability.
We believe that more populations of P annua must
be studied to determine the level and type of resistance
in the speci®c ®elds. Monitoring is needed to avoid
contamination of non-infested ®elds. The procedure
we presented here could be further modi®ed and
simpli®ed for the ef®cient screening of a large number
of samples for speci®c mutations, such as Val to Ile.
219215

LW Mengistu et al
ACKNOWLEDGEMENTSThe authors thank the Oregon Seed Council for partial
funding of this project. Contribution of the Agricul-
tural Research Service, USDA, in cooperation with the
Agricultural Experimental Station, Oregon State Uni-
versity Tech. Paper No. 11504 of the latter.
REFERENCES1 Trebst A, Inhibitors in electron ¯ow: Tools for the functional and
structural localization of carriers and energy conservation sites.
Methods Enzymol 69:675±715 (1980).
2 Arntzen CJ, P®ster K and Stienback KE, The mechanism of
chloroplast triazine resistance: Alterations in the site herbicide
action, in Herbicide Resistance in Plants, ed by LeBaron HM and
Gressel J, John Wiley & Sons, New York. pp 185±214 (1982).
3 Velthuys BR, Electron-dependent competition between plasto-
quinone and inhibitors for binding to photosystem II. FEBS
Lett 126:277±281 (1981).
4 P®ster K, Steinback KE, Gardner G and Arntzen CJ, Photo-
af®nity labelling of a herbicide receptor protein in chloroplast
membranes. Proc Natl Acad Sci USA 78:981±985 (1981).
5 Zurawski G, Bohnet H, Whitfeld P and Bottomley W, Nucleotide
sequence of the gene for the Mr 32000 thylakoid membrane
protein from Spinacia oleracea and Nicotinia debneyi predicts a
totally conserved primary translocation protein of Mr 38 950.
Proc Natl Acad Sci USA 79:7699±7703 (1982).
6 Erickson JM, Rahire M and Rochaix JD, Herbicide resistance
and cross-resistance: Changes at three distinct sites in the
herbicide binding protein. Science (Washington) 228:204±207
(1985).
7 Driesel AJ, Speirs J and Bohnert HJ, Spinach chloroplast mRNA
for a 32000 Dalton polypeptide. Size and localization on the
physical map of the chloroplast DNA. Biochem Biophys Acta
610:287±310 (1980).
8 Ryan GF, Resistance of common groundsel to simazine and
atrazine. Weed Sci 18:614±616 (1970).
9 LeBaron HM, Distribution and seriousness of herbicide-resistant
weed infestations worldwide, in Herbicide Resistance in Weeds
and Crops, ed by Caseley JC, Cussans GW and Atkin RK,
Butterworth-Heinemann, Oxford. pp 27±43 (1991).
10 Heap IM, The occurrence of herbicide-resistant weeds world-
wide. Pestic Sci 51:235±243 (1997).
11 Shimabukuro RH, Atrazine metabolism in resistant corn and
sorghum. Plant Physiol 43:1925±1930 (1968).
12 Ali A and Machado VS, A comparative analysis of leaf
chlorophyll ¯uorescence, Hill reaction activity and 14C-
atrazine tracer studies to explain differential triazine suscept-
ibility in wild turnip rape (Brassica campestris) biotypes. Can J
Plant Sci 644:707±713 (1984).
13 Gressel J, Herbicide tolerance and resistance: Alteration of site of
activity, in Weed Physiology, Vol II Herbicide Physiology, ed by
Duke SO, CRC Press, Boca Raton, Florida. pp 159±189
(1985).
14 Gronwald JW, Resistance to photosystem II inhibiting herbi-
cides, in Herbicide Resistance in Plants: Biology and Biochemistry,
ed by Powles SB and Holtum JAM, Lewis Publishers, Boca
Raton, Florida. pp 27±60 (1994).
15 Trebst A, The molecular basis of resistance to photosystem II
herbicides, in Herbicide Resistance in Weeds and Crops, ed by
Caseley JC, Cussans GW and Atkin RK, Butterworth-
Heinemann, Oxford. pp 145±164 (1991).
16 Oettmeier W, Masson K, Fedtke C, Konze J and Schmidt RR,
Effect of photosystem II inhibitors on chloroplasts from species
either susceptible or resistant towards s-triazine herbicides.
Pestic Biochem Physiol 18:357±367 (1982).
17 Fuerst EP, Arntzen CJ, P®ster K and Penner D, Herbicide cross-
resistance in triazine-resistant biotypes of four species. Weed
Sci 34:344±353 (1986).
18 Schwenger-Erger C, Thiemann J, Barz W, Johanningmeier U
216
and Naber D, Metribuzin resistance in photoautotrophic
Chenopodium rubrum cell cultures. Characterization of double
and triple mutations in the psbA gene. FEBS Lett 329:43±46
(1993).
19 Kless H, Shamir MO, Malkin S, McIntosh L and Edelman M,
The D-E region of the D1 protein is involved in multiple
quinone and herbicide interactions in photosystem II. Bio-
chemistry 33:10501±10507 (1994).
20 Tietjen KG, Kluth JF, Andree R, Haug M, Lindig M, MuÈller
KH, Wroblowsky HJ and Trebst A, The herbicide binding
niche of photosystem II ± A model. Pestic Sci 31:65±72 (1991).
21 Hirschberg J and McIntosh L, Molecular basis of herbicide
resistance in Amaranthus hybridus. Science (Washington).
222:1346±1349 (1983).
22 Hirschberg J, Bleecker A, Kyle DJ and McIntosh L, The
molecular basis of triazine herbicide resistance in higher plant
chloroplasts. Z Naturforsch 39:412±420 (1984).
23 GolouÈbinoff P, Edelman M and Hallick RB, Chloroplast coded
atrazine-resistance in Solanum nigrum: psbA loci from suscep-
tible and resistant biotypes are isogenic except for a single
codon change. Nucleic Acids Res 12:9489±9496 (1984).
24 Erickson JM, Rahire M, Bennoun MP, Delepelaire P, Dines B
and Rochaix JD, Herbicide resistance in C reinhardii results
from a mutation in the chloroplast gene for the 32 kiloDalton
protein in photosystem II. Proc Natl Acad Sci USA 81:3617±
3621 (1984).
25 Golden SS and Heselkorn R, Mutation to herbicide resistance
maps within the psbA gene of Anacystis nidulans R2. Science
(Washington) 229:1104±1107 (1985).
26 Van Oorschot JLP, Chloroplastic resistance of weeds to triazines
in Europe, in Herbicide Resistance in Weeds and Crops, ed by
Caseley JC, Cussans GW and Atkin RK, Butterworth-
Heinemann, Oxford. pp 87±101 (1991).
27 Rubin B, Herbicide resistance in weeds and crops, progress and
prospects, in Herbicide Resistance in Weeds and Crops, Vol II, ed
by Caseley JC, Cussans GW and Atkin RK, Butterworth-
Heinemann, Oxford. pp 387±414 (1991).
28 Trebst A, The molecular basis of plant resistance to photosystem
II herbicides, in Molecular Genetics and Evolution of Pesticide
Resistance, ed by Brown TM, ACS symposium series 645,
Washington, DC. pp 44±51 (1996).
29 McNally S, Bettini P, Sevignac M, Darmency H, Gasquez J and
Dron M, A rapid method to test for chloroplast DNA
involvement in atrazine resistance. Plant Physiol 83:248±250
(1987).
30 Gasquez J, Mouemar AAL and Darmency H, Triazine herbicide
resistance in Chenopodium album L. Occurrence and charac-
teristics of an intermediate biotype. Pestic Sci 16:390±395
(1985).
31 Bettini P, Sevignac M, McNally S, Darmency H, Gasquez J and
Dron M, Molecular mechanisms involved in the appearance of
atrazine resistance in Chenopodium album. J Cell Biochem 10:44
(1986).
32 Barros MDC and Dyer TA, Atrazine resistance in grass Poa
annua is due to a single base change in the chloroplast gene for
the D1 protein of photosystem II. Theor Appl Genet 75:610±
616 (1988).
33 Arntzen CJ, Ditto CL and Brewer PE, Chloroplast membrane in
triazine-resistant Amaranthus retro¯exus biotypes. Proc Natl
Acad Sci USA 76:278±282 (1979).
34 Bowes J, Crofts AR and Arntzen CJ, Redox reactions on the
reducing side of photosystem II in chloroplasts with altered
herbicide-binding properties. Arch Biochem Biophys 200:303±
308 (1980).
35 Ort OR, Ahrens WH, Martin B and Stoller EW, Comparison of
photosynthetic performance in triazine-resistant and suscep-
tible biotypes of Amaranthus hybridus. Plant Physiol 72:925±930
(1983).
36 Sundby C, Chow WS and Anderson JM, Effects on photosystem
II function, photoinhibition, and plant performance of the
spontaneous mutation of serine 264 in photosystem II reaction
Pest Manag Sci 56:209±217 (2000)

psbA mutation in Poa annua resistant to metribuzin and diuron
center D1 protein in triazine-resistant Brassica napus L. Plant
Physiol 103:105±113 (1993).
37 Hart JJ and Stemler A, Similar photosynthetic performance in
low light-grown isonuclear triazine-resistant and susceptible
Brassica napus L. Plant Physiol 94:1295±1300 (1990).
38 Holt JS and Thill DC, Growth and productivity of resistant
plants, in Herbicide Resistance in Plants: Biology and Biochem-
istry, ed by Powles SB and Holtum JAM, Lewis Publishers,
Boca Raton, Florida. pp 293±316 (1994).
39 Lien S, McBride JC, McBride AC, Togasaki RK and Pietro AS,
A comparative study of photosystem II speci®c inhibitors: The
differential action on a DCMU resistant mutant strain of C
reinhardi, in Photosynthetic Organelles, Plant & Cell Physiol
(Special Issue) 243±256 (1977).
40 Darmency H, Genetics of herbicide resistance in weeds and
crops, in Herbicide Resistance in Plants: Biology and Biochemistry,
ed by Powles SB and Holtum JAM, Lewis Publishers, Boca
Raton, Florida. pp 263±297 (1994).
41 Mueller-Warrant GW, Treatment of diuron-resistant annual
bluegrass in a perennial ryegrass seed crop, in 1996 Seed
Production Research at Oregon State University, USDA-ARS
Pest Manag Sci 56:209±217 (2000)
Cooperating, ed by Young III, WC, Department Crop and Soil
Science Ext/CrS 110. pp 29±32 (1997).
42 Heap IM, Multiple herbicide resistance in annual bluegrass (Poa
annua). WSSA Abstracts 42:56 (1995).
43 Gamroth DM, Brewster BD and Mallory-Smith CA, Herbicide
banding in grass-seed crops, in 1996 Seed Production Research at
Oregon State University, USDA-ARS Cooperating, ed by Young
III, WC, Department Crop and Soil Science Ext/CrS 110. p 33
(1997).
44 Mengistu LW, Genetic diversity and herbicide resistance in
annual bluegrass (Poa annua L). PhD Thesis, Oregon State
University, Corvallis, Oregon pp 1±122 (1999). Dissertation
abstract no. AAT 9842465.
45 Steiner JJ, Poklemba CJ, Fjellstrom RG, and Elliott LF, A rapid
one-step DNA extraction technique for PCR and RAPD
analysis. Nucleic Acids Res 23:2569±2570 (1995).
46 Gressel J, Gardner SN and Mangel M, Prevention versus
remediation in resistance management, in Molecular Genetics
and Evolution of Pesticide Resistance, ed by Brown TM, ACS
symposium series 645, Washington, DC. pp 169±186 (1996).
217
Related Documents











