Pergamon C’omp. Biochem. PI~wol. Vol. I I IA, No. 3. pp. 351-359. 1995 Elsevier Science Ltd Printed in Great Britam 0300-%29(%)00043-7 0300-9629195 $9.50 + 0.00 Protein nutrition and defence mechanisms in rainbow trout Oncorhynchus mykiss Viswanath Kiron, Takeshi Watanabe, Hideo Fukuda, Nobuaki Okamoto and Toshio Takeuchi Department of Aquatic Biosciences, Tokyo University of Fisheries, Konan 4, Minato, Tokyo 108, Japan The modulation of non-specific and specific immune mechanism by dietary protein was examined in rainbow trout. The lysozyme activity was reduced in protein-deficient fish. The variation of C-reactive protein level in fish at different nutritional statuses merits further examination. Antibody production was maintained, even in protein-deficient fish, indicating that defence systems operate preferentially to ward off infection. Pathogenic challenge trials reveal the importance of an optimum level of dietary protein, from the point of health; resistance to viruses varied among the different strains of trout. Key words: Antibody production; C-reactive protein; Health; Immune responses: Infection; Lysozyme; Nutrition; Protein; Rainbow trout. Comp. Biochem. Physiol. 11 IA, 351-359, 1995. Introduction The concept that nutritional deficiencies or excesses can mar the delicate and complicated mechanisms operating in the maintenance of the well-being of an organism, has been firmly established over the years. Nutritional factors impair immune responses and alter the suscepti- bility to infection and diseases (Chandra and Newberne, 1977; Gross and Newberne, 1980; Gershwin et al., 1985; Chandra, 1993). This is true for both man (Chandra, 1992) and animals (Burkholder and Swecker, 1990). In fact, several of the clues on immunomodulation by nutrients in man have been drawn from laboratory animals (Gershwin and Merchant, 1981). The piscine immune system employs both non-specific and specific mechanisms in response to infectious agents, as in the case for other vertebrates, including man. Research during the last two decades, has revealed several aspects of the immune functions operating in fish (Corbel, 1975; Ellis, 1981; Correspondence to: V. Kiron, Dept. of Aquatic Biosciences, Tokyo University of Fisheries, Konan 4, Minato, Tokyo 108, Japan. Tel. 3 5463 0555; Fax 3 5463 0553. Received 27 June 1994; revised 19 December 1994; accepted 20 December 1994. Ingram, 1980; Rijkers, 1982; Zeeman, 1986). However, it was during the eighties that investi- gators started looking into the role of nutrients as immunomodulators in fish. The reviews of La11 (1988) Landolt (1989) and Blazer (1991) provide an insight into the research. Among the nutrients, protein has been widely studied in clinical and laboratory evaluations of nutritional-immunological inter- actions (Gershwin et al., 1985). Except for the investigations of Thomas and Woo (1990) and Kiron et al. (1993) hardly any attempt has been made to work out the diseasedefence mechan- isms in fish linked to its protein nutritional status. The purpose of this study was to evaluate the changes in both non-specific and specific im- mune responses in rainbow trout Oncorhynchus mykiss maintained at different levels of protein nutrition. Pathogenic-challenge tests were also carried out for further evidence. Materials and Methods The experiments were performed on rainbow trout Oncorhynchus mykiss obtained either 351

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Pergamon C’omp. Biochem. PI~wol. Vol. I I IA, No. 3. pp. 351-359. 1995
Elsevier Science Ltd Printed in Great Britam
0300-%29(%)00043-7 0300-9629195 $9.50 + 0.00
Protein nutrition and defence mechanisms in rainbow trout Oncorhynchus mykiss
Viswanath Kiron, Takeshi Watanabe, Hideo Fukuda, Nobuaki Okamoto and Toshio Takeuchi Department of Aquatic Biosciences, Tokyo University of Fisheries, Konan 4, Minato, Tokyo 108, Japan
The modulation of non-specific and specific immune mechanism by dietary protein was examined in rainbow trout. The lysozyme activity was reduced in protein-deficient fish. The variation of C-reactive protein level in fish at different nutritional statuses merits further examination. Antibody production was maintained, even in protein-deficient fish, indicating that defence systems operate preferentially to ward off infection. Pathogenic challenge trials reveal the importance of an optimum level of dietary protein, from the point of health; resistance to viruses varied among the different strains of trout.
Key words: Antibody production; C-reactive protein; Health; Immune responses: Infection; Lysozyme; Nutrition; Protein; Rainbow trout.
Comp. Biochem. Physiol. 11 IA, 351-359, 1995.
Introduction
The concept that nutritional deficiencies or excesses can mar the delicate and complicated mechanisms operating in the maintenance of the well-being of an organism, has been firmly established over the years. Nutritional factors impair immune responses and alter the suscepti- bility to infection and diseases (Chandra and Newberne, 1977; Gross and Newberne, 1980; Gershwin et al., 1985; Chandra, 1993). This is true for both man (Chandra, 1992) and animals (Burkholder and Swecker, 1990). In fact, several of the clues on immunomodulation by nutrients in man have been drawn from laboratory animals (Gershwin and Merchant, 1981).
The piscine immune system employs both non-specific and specific mechanisms in response to infectious agents, as in the case for other vertebrates, including man. Research during the last two decades, has revealed several aspects of the immune functions operating in fish (Corbel, 1975; Ellis, 1981;
Correspondence to: V. Kiron, Dept. of Aquatic Biosciences, Tokyo University of Fisheries, Konan 4, Minato, Tokyo 108, Japan. Tel. 3 5463 0555; Fax 3 5463 0553.
Received 27 June 1994; revised 19 December 1994; accepted 20 December 1994.
Ingram, 1980; Rijkers, 1982; Zeeman, 1986). However, it was during the eighties that investi- gators started looking into the role of nutrients as immunomodulators in fish. The reviews of La11 (1988) Landolt (1989) and Blazer (1991) provide an insight into the research.
Among the nutrients, protein has been widely studied in clinical and laboratory evaluations of nutritional-immunological inter- actions (Gershwin et al., 1985). Except for the investigations of Thomas and Woo (1990) and Kiron et al. (1993) hardly any attempt has been made to work out the diseasedefence mechan- isms in fish linked to its protein nutritional status.
The purpose of this study was to evaluate the changes in both non-specific and specific im- mune responses in rainbow trout Oncorhynchus mykiss maintained at different levels of protein nutrition. Pathogenic-challenge tests were also carried out for further evidence.
Materials and Methods
The experiments were performed on rainbow trout Oncorhynchus mykiss obtained either
351

352 V. Kiron et al.
from the Okutama branch of the Tokyo Metropolitan Fisheries Experimental Station, or from the Fuji Trout Hatchery of the Shizuoka Prefectural Fisheries Experimental Station. The two different stocks of the trout were called the Okutama population and Fuji population, respectively. The former was used for the majority of the experiments, unless otherwise mentioned. Eyed eggs were brought to the rearing unit of our laboratory, hatched out and fed preferentially on commercial micro- diets. The fish were maintained initially at the ambient winter temperatures of 10 + 5°C and thereafter in the primary rearing system kept at about 15 f 2°C. The flow rate of dechlorinated tap water into the stock units as well as experimental units was regulated in the range 600-800 ml/min. The stock fish were fed com- mercial trout rations, twice every day. The fish were selected at certain size stages for the vari- ous experiments, depending on the purpose. The initial selection and pre-experimental feeding regimes were carefully planned as the nutri- tional background of organisms had a direct impact on the physiology. The experimental diets were prepared from purified ingredients which were mixed and pelleted mechanically (manually pelleted in the case of small-size feed), vacuum freeze-dried for about 16 hr and down-sized (only for experiments using small fish) to suit the fish. The casein-based purified diets were made isocaloric to the possible extent. Since the experiment was intended to look into
the effects related to protein nutrition status, and as variations in energy content of the diet influence the immunological functions (Gershwin er al., 1985), any manipulation of the protein component had to be compensated through dietary carbohydrates and lipids. The details on test feeding are given separately under the following headings.
Non -speciJic defence mechanisms
The non-specific defence mechanisms investi- gated in relation to the amount of protein in diet were the lysozyme levels and C-reactive protein (CRP) in the serum of rainbow trout.
Fish produced from the eyed eggs were weaned on commercial trout rations up to 34.3 + 2.5 g, when they were selected for the experiments to assess changes in lysozyme and CRP. The fish were maintained at temperatures of 15-l 7°C and gradually introduced to the experimental diets (Table 1). The purified casein diets IP, , IP, and IP, were formulated to contain 10, 35 and 50% protein, respectively. The fish were fed to satiation at 09.00 and at 17.00 hr. The diets were offered to 10 fish in duplicate lots for a period of 24 weeks, whereafter the assessments were made.
Lysozyme. Lysozyme present in the serum was assayed based upon the lysis of lysozyme- sensitive gram positive bacterium Micro- coccus lysodeikticus (lyophilized; Sigma Chemical Company). The basic lysoplate technique employed was the one developed by
Table I. Composition of &he diets: !. In the experiment to determine lysozyme activity and CRP response; 2. In the pathogenic challenge experiment*
Ingredient a/100 R
Diet groups (percent protein)
IP, (10) IP, (35) IP, (50)
Casein IO 35 50 Amino acid mixturet 1.3 1.3 1.3 cc-Starch IO 10 IO Dextrin 63 36 I5 Oils 8.7 4 4 Vitamin mixture$ 2 2 2 Mineral mixture7 5 5 5 Cellulose 0 6.7 12.7
Digestible energy (kcal/lOO g) 319.0 318.3 327.0
*The diet composition in the pathogenic challenge experiment was the same as shown above but for the change in diet codes as IIIP,, IIIP, & IIIP, for the 10, 35 and 50% protein, respectively.
tL-Cystine, 0.9 g; L-arginine; 0.4 g. ISoybean oil and Pollack liver oil at the ratio 3 : 2. gvitamin mixture: 1.5g vitamin blend; 0.1 g DL-CL tocopherol (50% purity); 0.4g choline
chloride. The vitamin blend included the respective amount of the following vitamins per 100 g of diet: B, 9 mg; B, 15 mg; B, 6 mg; Bi, 0.015 mg; C 750 mg; niacin &I mg; calcium pantothenate 15 mg; inositol 300 mg; biotin 0.9 mg; folic acid 2.25 mg; PABA 7.5 mg; K, 7.5 mg; A 6000 IU; D, 6000 IU.
TMineral mixture: IOOg of the mixture contained the following compounds. NaCl 1 g; MgSO, .7H,O 15 g;
.6H,O 0.1 g; KIO, 0.3 g; cellulose 43.5 g.

Protein nutrition and immune responses in trout 353
Ossermann and Lawlor (1966) and modified incorporating suggestions of later workers. In the procedure adopted, 40 mg of M. lysodeikti- cus was suspended in 100 ml of 0.04 M NaH, PO,/Na, HPO,-buffer (pH 6.2) containing 1% (w/v) agarose gel. The agar was dissolved by heating, and 15 ml of this was poured into a 90 mm-diameter Petri dish kept on a perfectly flat surface. The dishes were not moved until the gel had hardened, as uneven gelling would cause variations in lysis zone formation. Three equidistant wells of 4 mm-diameter were punched in the gel in each plate. The test serum samples or the standard rainbow trout samples or the hen egg white lysozyme (HEWL; Sigma Chemical Company) were introduced (10 p 1 each) into the wells, and the sealed plates were incubated for 72 hr at 25°C. The area cleared by lysis was measured three times using a digital area meter (Tamaya Planix 5000) and the mean value corresponded to the area of clearance of one particular sample. Each sample was tested in triplicate. HEWL was used to quantify the rainbow trout standard lysozyme, since the former could not be used as standard as it has already been established that fish lysozyme had a much higher rate of diffusion in agar, resulting in overestimation of activity, even up to 1700 times as high in the case of rainbow trout (Grinde et al., 1988a; Ochiai, 1993). The internal standard was chosen after sampling 50 healthy rainbow trout maintained on commercial rations, belonging to the same parentage as the experimental fish. This was prompted by the findings on genetic variation in the lysozyme activity of rainbow trout by Rered et al. (1993). The HEWL was prepared as dilutions in phosphate buffer and the final volume added to the plates contained 1 06, 1 O5 and 1 O4 units/ml. The factorial increment in lysis area was calculated for the dilutions. The area cleared by lysis of the bacterium was measured for the standard and samples and the lysozyme activity, y, was calculated, using the equation y = 104.1 ()(lOE.~ - logumg2.8
3 where y was the
lysozyme activity (units/ml); x the measured area of lysis (mm*); a the lysis area for the least dilution of the HEWL; and 2.8 the factorial increase in lysis area for HEWL. The lysozyme activities of the experimental groups are expressed relative to the activity of the rainbow trout lysozyme standards assumed as loo%, a combination of which was repetitively tested along with the samples. The values in each group were the mean + SD of lo-12 fish.
C-reactive protein. The CRP in serum was quantitatively determined with test packs in Abbott Vision System (Abbott Laboratories, 1989). The system employed a turbidimetric immunoassay for C-reactive protein, where the turbidity formed by antigen-antibody com- plexes in the presence of polyethylene glycol (PEG) was measured bichromatically with a 340/600 nm filter pair. The end point is pro- portional to the CRP concentration. The values were determined for 10-l 2 samples per treatment.
Specific defence mechanism
The effect of protein nutrition on humoral immune response constituted the study on specific defence mechanism. Antibody pro- duction against Aeromonas salmonicida was quantified in this experiment.
Stock fish maintained on commercial rations were trained on a purified diet at first and subsequently chosen for the experiments. The initial size was 127 _+ 12 g. Six lots of 15 fish each were fed three experimental diets (Table 2) in duplicate, a low protein (10%) group UP,, an adequate protein (35%) group IIP,, and an excess protein (65%) group IIP,. During the rearing period on the experimental diets, they were offered food twice a day to satiation. The rearing facilities were as described earlier. After 12 weeks, one lot from each treatment was injected with formalin-killed Aeromonas salmonicida in adjuvant, and the other, which served as the control group, was injected with Freund’s adjuvant alone.
Table 2. Composition of the diets in the experiment on antibody production
Ingredient
g/l00 g
Casein Amino acid mixture* a-Starch Dextrin Oil* Vitamin mixture* Mineral mixture* Cellulose
Digestible energy (kcal/lOO g)
*Refer to footnote of Table I.
Diet groups (percent protein)
HP, (10) HP, (35) IIP, (65)
IO 35 65 1.3 1.3 I.3
15.7 10 9 55 39 II 5 4 2 2 2 5 5 5 0 2.7 13.7
330. I 334.7 349.7

354 V. Kiron et al.
Antibody production. Aeromonas salmonicida cultured in trypto-soy broth was harvested by centrifugation (48OOg), and the collected bac- teria were washed three times in phosphate buffer saline (PBS; pH 7.2). They were then suspended in 0.3% formaldehyde solution and left overnight on a shaker at 25°C. After the removal of formaldehyde the next day, a 10% (w/v) suspension of the killed bacteria was reconstituted with PBS. The bacterial inviability was confirmed simultaneously. The bacterial suspension in PBS was emulsified in an equal volume of Freund’s complete adjuvant to be employed as inoculum. The fish received 0.5 ml of this as an intraperitoneal injection. The con- trol fish were injected with an identical volume of adjuvant alone. Six weeks after immuniz- ation, blood samples were collected from both groups to serve as the antisera and control sera. Sera collected from some fish before immuniz- ation were used for reference purposes.
The antibody production in the fish against Aeromonas salmonicida was detected by enzyme- linked immunosorbent assay (ELISA) with suit- able modifications to the technique described by Kodama et al. (1985). A. salmonicida were collected according to the methods detailed earlier, and later a 10% suspension (w/v) was constituted in PBS. The bacterial lysate was prepared by ultrasonification (200 W, 20 set) employing a sonicator (Ohtake). The resulting solution was centrifuged (2OOOg, 15 min) and the supernatant was removed to be used as antigen. The antigen was diluted with coating buffer (Na,CO,/NaHCO, , pH 9.6) to give a resultant protein concentration (Lowry et al., 1951) of 10 pg/ml. One hundred microlitres of the antigen were introduced to each well of a 96-well microplate, sealed and kept overnight in a 25°C incubator. The next day, the wells were drained and washed gently with PBS-Tween three times; this step was repeated at the end of each of the succeeding steps. After washing, 200 ~1 of the blocking solution (2% bovine serum albumin in 50 mM Tris hydrochloride, pH 8.0/0.9% sodium chloride) was added. The antibody containing serum (or dilutions) was added, 100 ~1 per well in triplicates. After the lapse of the incubation period and washings, 100 ,ul of diluted ( x 100) biotin-labelled anti- rainbow trout immunoglobulin rabbit IgG was introduced. The specificity of the rabbit IgG was confirmed by Western-blot technique, prior to application. An hour later, this was replaced by an equal volume of avidin conjugated peroxidase of identical dilution, In the next replacement step, 100~1 of OPD solution (containing 0.1 M citric acid, 0.2 M Na, HPO,, orthophenylene diamine and H,Oz) was added to each well. The last stage of incubation was for
only 30 min; in the previous steps, it ranged from 1 to 2 hr at 25-30°C. The reaction was halted at this stage on addition of 50 ,~l 2 M sulphuric acid. The color developed in the wells was read immediately at 492 nm using a microplate reader (Tosoh MPR A4). The values were expressed in relation to those of control sera.
Disease resistance capacity
The infectivity trial conducted on rainbow trout weaned on some of the experimental diets described in the earlier sections was intended as a complement to the defence mechanism studies.
The eyed eggs (Fuji population) of fish were hatched out and reared up to 0.40 ) 0.03 g when they were chosen for the experiment. The exper- imental diets (Table 1) were to contain 10% (IIIP,), 35% (IIIP,) and 50% (HIP,) protein. The dry diets, made to suit the experimental fish size, were offered for nearly 5 weeks to 20 fish each in duplicate tanks per treatment. The sizes of the fry prior to infection were 0.56 and 0.54 g for group IIIP,, 0.69 and 0.69 for group IIIPZ and 0.67 and 0.68 for group IIIP,.
The fish were exposed to Infectious Hematopoietic Necrosis Virus which had been isolated from the Tokyo Fisheries Experimental Station. A hyperosmotic infiltration technique was adopted using the virus at 105~“TCID,,/ml. The exposure timings were 2 min in 5.4% NaCl solution and 6 min in virus solution. The fish were maintained for 3 weeks in the rearing system (13-15°C) of the pathology unit, taking extreme caution during the progress of the disease. Mortality was monitored frequently, and dead fish were removed every day at 08.00 hr. The results are expressed as mean cumulative mortality for the duplicate tanks during the 3 weeks.
Statistical analysis
Various responses under the different dietary treatment were processed with Duncan’s Mul- tiple range tests. The results of the ANOVA were considered significant at P < 0.05.
Results
Non -spec$c defence mechanisms
Lysozyme. The lysozyme activity detected in the low protein diet group (IP,) was signifi- cantly lower (P < 0.01) than the other groups (IP, and IP,) which had a sufficient amount of the nutrient. Comparing the latter two groups, an increment in protein level did not bring about any significant variation in the enzyme activity (Fig. 1). The mean (n = 10) relative
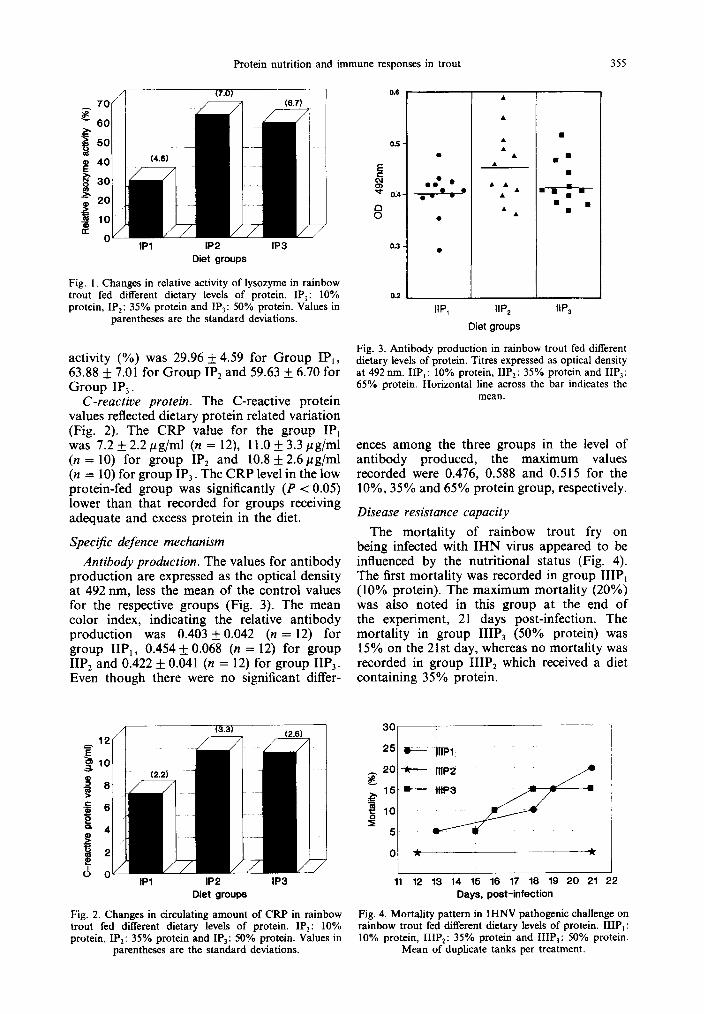
Protein nutrition and immune responses in trout 355
”
IPI IP2 IP3 Diet groups
Fig. I. Changes in relative activity of lysozyme in rainbow trout fed different dietary levels of protein. IP,: 10% protein, IP,: 35% protein and IP,: 50% protein. Values in
parentheses are the standard deviations.
activity (%) was 29.96 + 4.59 for Group IP,, 63.88 + 7.01 for Group IP, and 59.63 + 6.70 for Group IP,.
C-reactive protein. The C-reactive protein values reflected dietary protein related variation (Fig. 2). The CRP value for the group IP, was 7.2 + 2.2 pgg/ml (n = 12) 11.0 f 3.3 pgg/ml (n = 10) for group IP, and 10.8 f 2.6pgg/ml (n = 10) for group IP, . The CRP level in the low protein-fed group was significantly (P < 0.05) lower than that recorded for groups receiving adequate and excess protein in the diet.
Specific defence mechanism
Antibody production. The values for antibody production are expressed as the optical density at 492 nm, less the mean of the control values for the respective groups (Fig. 3). The mean color index, indicating the relative antibody production was 0.403 t_ 0.042 (n = 12) for group IIP,, 0.454 * 0.068 (n = 12) for group IIP, and 0.422 + 0.041 (n = 12) for group IIP3. Even though there were no significant differ-
=‘ ‘2
5 10
I 8 i8
tj: J 0
IPI IP2 IP3 Diet groups
25 );-_ ~FP’,
;$Zzt/+
0 1 *
II 12 13 14 15 18 17 18 19 20 21 : Days, post-infection
1 2. 2
Fig. 2. Changes in circulating amount of CRP in rainbow Fig. 4. Mortality pattern in 1HNV pathogenic challenge on trout fed different dietary levels of protein. IP,: 10% rainbow trout fed different dietary levels of protein. IIIP,: protein, IP,: 35% protein and IP,: 50% protein. Values in 10% protein, IIIP,: 35% protein and IIIP,: 50% protein.
parentheses are the standard deviations. Mean of duplicate tanks per treatment.
0.6
0.6
.
..* l
0’ -
.
.
IIP,
* I
.
. * * *
L m=
.
AA. n = . I
*
. *
.
. .
IIP, IIP,
Diet groups
Fig. 3. Antibody production in rainbow trout fed different dietary levels of protein. Titres expressed as optical density at 492 nm. IIP,: 10% protein, IIP,: 35% protein and IIP,: 65% protein. Horizontal line across the bar indicates the
mean.
ences among the three groups in the level of antibody produced, the maximum values recorded were 0.476, 0.588 and 0.515 for the lo%, 35% and 65% protein group, respectively.
Disease resistance capacity
The mortality of rainbow trout fry on being infected with IHN virus appeared to be influenced by the nutritional status (Fig. 4). The first mortality was recorded in group IIIP, (10% protein). The maximum mortality (20%) was also noted in this group at the end of the experiment, 21 days post-infection. The mortality in group IIIP, (50% protein) was 15% on the 21st day, whereas no mortality was recorded in group IIIP, which received a diet containing 35% protein.

Discussion the course of a disease (Ingram, 1980). CRP analogs have been demonstrated in the sera of
Many aspects of lysozyme have been described in vertebrates as well as invertebrates
fish such as plaice Pleuronectes platessa, cod
(Jolles and Jollts, 1984). Lysozyme activity has Gadus morhua, tilapia Tilapia mossambica and
been characterised in several fish species rainbow trout Oncorhynchus mykiss (Bald0 and
(Fletcher and White, 1976; Grinde et al., 1988b; Fletcher, 1973; Murai et al., 1990; Pepys et al., 1982; Ramos and Smith, 1978).
Lie et al., 1989) and is thought to play a role in disease defence (Lindsay, 1986); it is particularly
Commenting on the appearance of CRP in
associated with leucocyte-rich tissues and sites normal plaice sera, Ingram (1980) has remarked that this might have resulted from stress due to
where the risk of bacterial invasion is high. handling, capture, bleeding or removal from Lysozyme appears in the blood as secretions from the lysozomes of neutrophils and macro-
their natural environment or from previous exposure to infectious organisms. Nevertheless,
phages (Goldstein et af., 1975; Murray and in the present experiment on normal rainbow Fletcher, 1976). The action of lysozyme on the trout fed different diets containing 10, 35 and Gram-negative bacteria which are pathogenic to 50% protein, it was noted that the CRP levels fish, is effected after complement and various were significantly (P -C 0.05) lower in the low enzymes have acted on the outer bacterial wall, protein diet, but almost similar at the other exposing the inner murin layer (Neeman et al., two levels. It appears that rainbow trout, a cold 1974; Hjelmeland et al., 1983). water species like plaice, maintains a certain
As the lysozyme is of immunological signifi- level (in this case, usually betwen 8 and cance, modulations in its activity could be 18 pg/ml) of CRP in their serum. It was surpris- employed as indirect measures for disease resist- ing that we observed a diet-based variation in ance. Reed et al. (1993) has suggested that it the level of CRP. Murai et al. (1990) recorded may be possible to influence lysozyme activity of rainbow trout through genetic selection pro-
a three-fold increase in the CRP-level of exper- imentally infected rainbow trout, compared
grammes. An analysis of our results opens up the possibility of manipulating lysozyme activity
with control animals. Although, currently we have not authenticated our results with induced
through dietary modifications. Low levels of inflammation/injury tests, experiments in pro- protein in the diet reduced the level of activity gress may add new information. The extent to compared with optimal levels of protein, How- which the synthesis of acute phase proteins can ever, the activity in fish receiving the largest be influenced by nutrition in man is not clear amount of protein was not significantly different from the 35% protein fed group. Since dietary
(Fleck, 1989). Cell mediated immunity was severely depressed in patients with the lowest
insufficiency of the nutrient was easily reflected concentration of albumin and the highest con- in lysozyme activity, it is an indication of the centration of CRP (Nagendran et al., 1985a.b). importance of proper nutrition in maintaining Nehaus et al. (1963) and Cruickshank et al. the health of fish. Investigators in human nutri- (1989) have, however, reported that the acute tion have asserted that secretory substances, such as lysozyme, which are important in host
phase protein response may be attenuated by
defence functions are lowered during protein malnutrition. In a recent work, Jennings et al. (1992) asserted that protein deficiency can alter
malnutrition (McMurray et al., 1977; Watson the pattern and magnitude of acute phase et al., 1978). It has been validated that infections responses (albumin and c(? macroglobulin) in (Studnicka et al., 1986) and stressors (Mock and circulating protein concentrations to an extent Peters, 1990) cause a suppression of lysozyme that is dependent on the severity of protein activity in fish. It would, therefore, be inter- deficiency. They have suggested that high energy esting to examine the synergistic effect of low protein diets could reduce the availability of malnutrition and infections on the priming of amino acids, and their flux from peripheral lysozyme activity. tissues to central organs like the liver where
Acute phase proteins, including C-reactive acute phase proteins including CRP are syn- protein (CRP) appear in the serum of man and other mammals in response to infection, inflam-
thesized. A similar reduction in synthesis might have caused the low level of circulating CRP in
mation and tissue damage (Gewurz et al., 1982). the low protein fed fish in our study. Murai eb al. CRP is reported to be involved in phagocytic (1990) has attributed the presence of serum activity, opsonisation, mutagenesis, interaction CRP in normal trouts as a natural defence with the complement, antibacterial lysis and mechanism, since specific immune responses are particularly in binding to cell surfaces in relatively slow. Since the antibody responses are agglutination (Cook, 1977). Therefore, CRP is inhibited at low temperatures (Avtalion et al., a useful aid for the detection of inflammatory 1973), CRP-precipitins have been suggested by conditions, secondary infections and following Ingram (1980) as a valuable adjunct to humoral
356 V. Kiron et al.

Protein nutrition and immune responses in trout 351
defences. He has concluded that it would be beneficial for lower vertebrates to possess serum components with a wide specificity for cell walls and surface structures of invading pathogens.
The response patterns of both CRP and lysozyme activity were identical in our exper- iments on rainbow trout. An early work by Fletcher et al. (1977) mentions a parallel seasonal rise in serum CRP and lysozyme of lumpsucker, Cylopterus lumpus.
The antibody titres produced in response to A. salmonicida seem to fall within identical ranges (Fig. 3). Even though the mean observations were similar, greater titres were recorded for the fish receiving 35% protein. However, statisti- cally they were not different from each other. Such observations were made in an earlier exper- iment (Kiron et al., 1993). In the cited study, the group of fish not receiving any protein in the diet, maintained antibody titres comparable with those receiving different levels of protein. It was stated that the antibody production was above the consideration of dietary protein status. However, in another experiment on rain- bow trout, Thomas and Woo (1990) investigated the dietary modulation of humoral immune response when infected with Cryptobia salmosit- ica, a pathogenic hemoflagellate. The antibody titres were low in the fish fed on 19% protein diet; but strikingly there was not much variation in the titres for the 29% and 38% protein group. In assessments of functional competence of antibody-mediated immunity in children, quantified based on total immunoglobulin pre- sent in serum, several workers (Keet and Thorn, 1969; McFarlane et al., 1970; Watson and Freesemann, 1970; Purtilo et al., 1976) have concluded that levels in protein-energy malnour- ished subjects were similar to or above those of the control. The almost identical response of the three protein groups (IIIP, , IIIP, and IIIP,) in the present study on rainbow trout could there- fore be interpreted on similar lines. Earlier, Cohen and Hansen (1962) had described that, even though there was a reduction in albumin production, the y-globulin synthesis remained unaffected in protein-malnourished human subjects. An increased rate of immunoglobulin synthesis was noted in these groups when pathogenically challenged.
Notwithstanding, Gershwin et al. (1985) concluded that dietary protein indeed has a potent role in the development of humoral immune responses. Though they ascribe to the views of the above authors, they remark that humoral immunity cannot be singled out as a phenomenon. Instead, it is a multitude of inter- actions, and protein deficiency may influence the immune response in various ways, depending on the antigen, its dosage and the number
ap 10
36 60
0 20 40 60 80 100 % Mortality
Fig. 5. Cumulative results of 1HNV pathogenic challenge on rainbow trout in three different trials. Data in Trials I and 2 from Kiron ef al. (1993); Trial 3-present experiment. Trials 1 and 2 contained a protein-deprived group. Other protein groups in the two trials were 20, 35 and 50%. Note that no mortality was recorded for the 35% protein
group in Trials 1 and 3.
of challenges. They suggest that, as with many other nutrients, the major influence may involve alterations in sub-populations of regu- latory T-cells. Our study specifically observed humoral responses, but it could be that cellular immunity is also concomitantly influenced by protein nutrition, as mentioned earlier. In addition, Watson and Haffer (1980) have shown that the immature immune system is more susceptible to protein deficiency than is the immune system of older mice. We cannot dis- count such probabilities from our experiment. As the fish had already been fed earlier during its life with balanced commercial rations, a nutritional insult in later life might not be severe enough to cause imbalances in vital mechanisms functioning in disease prevention. Nonetheless, this hypothesis has not been verified in this study.
The experiment on disease resistance against IHN virus indicated that malnourished fish are more prone to infections compared with those receiving an optimum level (35%) of protein in diet. Mortality was also recorded at the higher level (50%) of protein, whereas no fish died in the 35% protein group. These results, the third in a series of infectivity trials, were similar to our early observations (Kiron et al., 1993; Fig. 5) The pattern of mortality in those experiments (Trial 1, Trial 2) resembled that of the exper- iment reported here (Trial 3), but the big difference in mortality rates had alarmed us since all experimental conditions were the same, except for the fish size and population (Trial 1, Fuji population: mean weight at infection ranged from 2.35 g for the protein-deprived group to 5.70 g for other groups; Trial 2, Okutama population: mean weight at infection, 0.36 g for the protein-deprived group and

358 V. Kiron et al.
ranged from 0.42 to 0.48 g for the other three groups; Trial 3, Fuji population: 0.54-0.56 g for the low-protein group and 0.67-0.69 g for the two other groups). Earlier, we concluded that such variations in mortality might have been due to the size difference of the fish, but the result of the current experiment performed on the Fuji population of fish, which were very small in size compared with those in Trial 1, exhibited mortality patterns as before. It could therefore be deduced that resistance to the virus varied among different populations of fish. Nevertheless, since mortality patterns were identical in Trial 1 and Trial 3, irrespective of size, it could be validated that diets did affect the resistance capacity to infective agents. A low-protein diet invariably resulted in greater mortality, but higher levels (around 50% pro- tein) also seemed to depress disease resistance when compared with the intermediate ranges (20-35% protein). Hardy et al. (1979) had also noted pathogenic invasion at high levels of dietary protein in chinook salmon. It appears, therefore, that an optimum level of protein is required in the diets to ward off pathogens. Extensive studies have to prove this and until such time, it cannot be firmly established that high levels of protein may be harmful, particu- larly based on our observations in Trial 2, where the results might have been influenced by the general weakness of the trout population,
In conclusion, it appears that protein nutrients are important in non-specific defence mechanisms, as indicated by lysozyme activity and the C-reactive protein response. Specific defence in terms of humoral antibody levels does not seem to depend on protein quantity in the diet. Disease resistance capability is achieved by a certain level of dietary protein. Dietary protein has a prominent role in the health of fish. While these results have partially clarified the role of protein nutrition in immunocompe- tence of fish, future studies could consider the precise interactions.
References
Abbot Laboratories (1989) Vision System Operators’ Manual, pp. 7.17-1.79. Abbott Laboratories, Abbott Park, Illinois.
Avtalion R. R., Wojdani A., Malik Z., Shahrabani R. and Duczyminer M. (1973) Influence of environmental temperature on the immune response in fish. Curr. Top. Microbial. Immunol. 61, l-35.
Baldo B. A. and Fletcher T. C. (1973) C-reactive protein- like precipitins in plaice. Nature 246, 145-146.
Blazer V. S. (1991) Piscine macrophage function and nutritional influences: A review. J. Aquat. Anim. Hlth 3, 77-86.
Burkholder W. J. and Swecker W. S. Jr (1990) Nutritional influences on immunity. Semin. Vef. Med. Surg. 5, 154-166.
Chandra R. K. (1992) Nutrition and Immunology. Experience of an old traveller and recent experiences. In Nutrition and Immunology (Edited by Chandra R. K.), pp. 9-44. ARTS Biomedical, Newfoundland.
Chandra R. K. (1993) Nutrition and the immune system. Proc. Nurr. Sot. 52, 77-84.
Chandra R. K. and Newberne P. M. (1977) Nutrition Immunity and Infection: Mechanisms of Interactions. Plenum Press, New York.
Cohen S. and Hansen J. D. L. (1962) Metabolism of albumin and globulin in kwashiorkor. Clin. Sci. 23, 351-359.
Cook G. D, (1977) The value of C-reactive protein determinations. Lab-Lore. 7, 465-467.
Corbel M. J. (1975) The immune response in fish: A review. J. Fish Biol. 7, 539-563.
Cruickshank A. M., Hansell D. T., Bunns H. J. G. and Shenkin A. (1989) Effect of nutritional status on acute phase protein response to elective surgery. Br. J. Surg. 76, 1655168.
Ellis A. E. (1981) Non-specific defense mechanisms in fish and their role in disease processes. Deul. Biol. Stand. 49, 331-352.
Fleck A. (1989) Clinical and nutritional aspects of changes in acute-phase proteins during inflammation. Proc. Nutr. sot. 48, 341-354.
Fletcher T. C. and White A. (1976) The lysozyme of the plaice Pleuronectes platessa L. Comp. Biochem. Physiol. 55B, 207-2 10.
Fletcher T. C., White A. and Baldo B. A. (1977) C-reactive protein-like precipitin and lysozyme in the lumpsucker Cyclopterus lumpus L. during the breeding season. Comp. Biochem. Physiol. 57, 353-357.
Gershwin M. E., Beach R. S. and Hurley L. S. (1985) Nutrition and Immunily, pp. 1288183. Academic Press, London.
Gershwin M. E. and Merchant B. (1981) Immunologic Defects in Laborator.v Animals, Vol. 2. Plenum Press, New York.
Gewurz H.. Mold C., Siegel J. and Fiedel B. (1982) C-reactive protein and the acute phase response. Adv. Int. Med. 27, 345-372.
Goldstein I. M., Hoffstein S. T. and Weissmann G. (1975) Mechanisms of lyzosomal enzyme release from human polymorphonuclear leucocytes. J. Cell Biol. 66, 647-652.
Grinde B., Jolles J. and Jollts P. (1988a) Purification and characterization of two lysozymes from rainbow trout (Salmo gairdneri ). Eur. J. Biochem. 173, 269-273.
Grinde B., Lie 0.. Poppe T. and Salte R. (1988b) Species and individual variation in lysozyme activity in fish of interest in aquaculture. Aquaculture 68, 299-304.
Gross R. L. and Newberne P. M. (1980) Role of nutrition in immunologic function. Physiol. Reu. 60, 188-302.
Hardy R. W., Halver J. E. and Brannon E. L. (1979) Effect of dietary protein level on the pyridoxine requirement and disease resistance of chinook salmon. In Finfish Nunition and Fishfeed Technology (Edited by Hal& J. E. and Tiews K.), Vol. 1, pp. 2533260. Heenemann, Berlin.
Hjelmeland K., Christie M. and Raa J. (1983) Skin mucus protease from rainbow trout. Salmo gairdneri Richardson, and its biological significance. J. Fish Biol. 23, 13-22.
Ingram G. A. (1980) Substances involved in the natural resistance of fish to infection-A review. J. Fish Biol. 16, 23-60.
Jennings G., Bourgeois C. and Elia M. (1992) The magni- tude of the acute phase protein response is attenuated by protein deficiency in rats. J. Nutr. 122, 132551331.
Jolles P. and Jolles J. (1984) What’s new in lysozyme research? Always a model system, today as yesterday. Molec. Cell Biochem. 63, 165-189.
Keet M. P. and Thorn H. (1969) Serum immunoglobulins in kwashiorkor. Arch. Dis. Child. 44, 600-603.

Protein nutrition and immune responses in trout 359
Kiron V.. Fukuda H., Takeuchi T. and Watanabe T. (1993) Dietary protein related humoral immune response and disease resistance of rainbow trout Oncorhynchus mykiss. In Fish Nutrition and Practice (Edited by Kaushik S. J. and Luquet P.), pp. 119-126. INRA. Paris.
Kodama, H., Honda A., Moustafa M., Mikami T. and Izawa H. (1985) Detection of antibody in rainbow trout against Aeromonas salmonicido by enzyme linked immunosorbent assay. Fish Pathol. 20, 237-242.
Lall S. P. (1988) Disease control through nutrition. In Proc. Aquaculture International Congress and E.rposition, pp. 607-610. Vancouver.
Landolt M. L. (1989) The relationship between diet and immune response of fish. Aquaculture 79, 193-206.
Lie 0.. Evensen 0.. Anita S. and Froysadai E. (1989) Study on lysozyme activity in some fish species. Dis. Aquar. Org. 6. I-5.
Lindsay G. J. H. (1986) The significance of chitinolytic enzymes and lysozyme in rainbow trout (Salmo gairdneri ) defense. Aquaculture 51, 1699173.
Lowry 0. H.: Rosebrough N. J.. Farr A. L. and Randall R. J. (1951) Protein measurement with the Folin ohenol reagent. J. ‘hiol. Chem. 193, 265-275.
McFarlane H., Reddy S.. Adcock K. J.. Adeshina H.. Cooke A. R. and Akene J. (1970) Immunity. transferrin and survival in kwashiorkor. Br. Med. J. 4, 2688270.
McMurray D. N., Rey H.. Casazza L. J. and Watson R. R. (1977) Effect of moderate malnutrition on concentrations of immunoglobulins and enzymes in tears and saliva of young Colombian children. Am. J. clin. Nutr. 30, 1944 -1948.
Murai T.. Kodama H., Naiki M.. Mikami T. and Izawa H. (1990) Isolation and characterization of rainbow trout C-reactive protein. Decl. camp. Immunol. 14, 49-58.
Murray C. K. and Fletcher T. C. (1976) The immunohisto- chemical localization of lysozyme in plaice (Pleuronecres phtessa L.) tissues. J. Fish Biol. 8, 329-334.
Mock A. and Peters G. (1990) Lysozyme activity in rain- bow trout, Oncorhynchus mykiss (Walbaum), stressed by handling, transport and water pollution. J. Fish Bioi. 37, 873-885.
Nagendran V.. Myers M. A. and Fleck A. (1985a) Depression of cell mediated immunity in cancer patients may not solely be due to malnutrition. In Proc. XIII International Congress N/ Nutrition, p. 78. Brighton, Abstract S16-0102.
Nagendran V., Myers M. A. and Fleck A. (1985b) Further evidence that serum albumin concentration is not an appropriate index of nutritional status in the presence of disease. In Proc. XIII Internalional Congress of Nulrifion. p. 95. Brighton. Abstract CO4-0044.
Neeman N.. Lahav M. and Ginsburg I. (1974) The effect of
leukocyte hydrolases on bacteria. II. The synergistic action of lysozyme and extracts of PMN. macrophages, lymphocytes and platelets in bacteriolysis. Proc. Sot. esp. hiol: Meb. 146, Ii377I 145.
Nehaus 0. W.. Baleeno H. and Miiauskas A. 11963) Biochemical significance of serum glycoproteins. II.‘EtIect of dietary protein on changes in rat serum following injury. Exp. Molec. Path. 2, 183-187.
Ochiai M. (1993) The relationship between Iclhpophonus disease in rainbow trout and serum lysozyme activity. MS. Thesis, Tokyo University of Fisheries. Tokyo tin Japanese).
Ossermann E. F. and Lawlor D. P. (1966) Serum and urinary lysozyme (Muramidase) in monocytic and monomyelocytic leukemia. J. exp. Med. 124. 921-951.
Pepys M. B., De Beer F. C.. Milstein C. P.. March J. F.. Feinstein A.. Butress N., Clamp J. R., Taylor J., Bruton C. and Fletcher T. C. (1982) C-reactive protein and serum amyloid P component in the plaice (Pleuronecre.7 p/ute.w L.). a marine teleost, are homologous with their human counterparts. Biochim. biophys. Acta 704. 123 133.
Purtilo D. T.. Rices R. S.. Evans R. and Neatie R. C. ( 1976) Humoral imminity of parasitized, malnourished children: Am. J. Trap. Med. Hyg. 25, 229-232.
Ramos F. and Smith A. C. (1978) The c-reactive protein (CRP) test for the detection of early disease in tishes. Aquaculture 14, 261 -266.
Rijkers G. T. (1982) Non-lymphoid defense mechanisms in fish. Der:l. camp. Immunol. 6, l-13.
Reed K. H.. Larsen H. J. S.. Linder R. D. and Refstie T. (1993) Genetic variation in lysozyme activity in rainbow trout (Oncorh~wchus mykiss). Aquaculrure 109, 231 244.
Studnicka M.. Siwicki A. and Ryka B. (1986) Lysozymc level in carp (Cyprinus carpio L.). Bamidgeh 38, 22 25.
Thomas,P. T. and Woo P. T. K. (1990) Dietary modulation of humoral immune response and anaemia in rainbow trout, Oncorh~nchus mykiss (Walbaum). infected with Cryptobia salmosirica Katz. 1951. J. Fi.~h Di.v. 13, 435-446.
Watson C. E. and Freesemann D. (1970) Immunoglobulins in protein-calorie malnutrition. Arch. Dis. Child 45. 282-284.
Watson R. R. and Haffer K. (1980) Modifications of cell-mediated immune responses by moderate dietary protein stress in immunologically immature and mature BALBic mice. Mech. Agng Der. 12, 269 273.
Watson R. R., Reyes M. A. and McMurray D. N. (1978) Influence of malnutrition on the concentration of IgG. lysozyme. amylase and aminopeptidase in children’s tears. Proc. Sot. esp. biol. Med. 157, 215-219.
Zeeman M. (1986) Modulation of the immune response in fish. Pet. Immunol. Immunoparhol. 12, 235-241.
Related Documents











