Protein kinase CK2 in postsynaptic densities: phosphorylation of PSD-95/SAP90 and NMDA receptor regulation q Dagoberto Soto a,1 , Floria Pancetti b,1 , Juan Jose ´ Marengo c , Mauricio Sandoval a , Rodrigo Sandoval a , Fernando Orrego a , Ursula Wyneken a, * a Neuroscience Laboratory, Faculty of Medicine, Universidad de Los Andes, Santiago-6782468, Chile b Laboratory of Neurosciences, Department of Biology, Faculty of Chemistry and Biology, University of Santiago, Chile c ICBM, Faculty of Medicine, Universidad de Chile, and Instituto de Neurocirugı ´a Asenjo, Chile Received 19 July 2004 Abstract Protein kinase CK2 (CK2) is highly expressed in rat forebrain where its function is not well understood. Subcellular distribution studies showed that the catalytic subunit of CK2 (CK2a) was enriched in postsynaptic densities (PSDs) by 68%. We studied the putative role of CK2 activity on N-methyl-D-aspartate receptor (NMDAR) function using isolated, patch-clamped PSDs in the pres- ence of 2 mM extracellular Mg 2+ . The usual activation by phosphorylation of the NMDARs in the presence of ATP was inhibited by the selective CK2 inhibitor 5,6-dichloro-1-b-ribofuranosyl benzimidazole (DRB). This inhibition was voltage-dependent, i.e., 100% at positive membrane potentials, while at negative potentials, inhibition was incomplete. Endogenous CK2 substrates were characterized by their ability to use GTP as a phosphoryl donor and susceptibility to inhibition by DRB. Immunoprecipitation assays and 2D gels indicated that PSD-95/SAP90, the NMDAR scaffolding protein, was a CK2 substrate, while the NR2A/B and NR1 NMDAR subunits were not. These results suggest that postsynaptic NMDAR regulation by CK2 is mediated by indirect mechanisms possibly involving PSD-95/SAP90. Ó 2004 Elsevier Inc. All rights reserved. Keywords: NMDA receptor; Protein kinase CK2; Postsynaptic density; PSD-95/SAP90; Excitatory neurotransmission; Rat forebrain Protein kinase CK2 (CK2; formerly called casein ki- nase 2) is a ubiquitous serine-threonine kinase [1–3], that is highly concentrated in the brain [4], where it phospho- rylates at least 78 different proteins, including several ones related to nucleic acid and protein synthesis, tran- scription factors, cytoskeletal and other structural pro- teins, as well as a large number of elements involved in signal transduction [5]. In cells, the majority of CK2 is found as a heterotetramer consisting of two catalytic subunits (CK2a or CK2a 0 ) and two regulatory subunits (CK2b) [2,6]. CK2 is activated in the hippocampus during long- term potentiation (LTP) [7] and following the applica- tion of brain-derived neurotrophic factor (BDNF) [8], and is able to phosphorylate proteins related to plastic- ity, such as transcription factors of the HMG class, GAP-43 (B-50) and DARPP-32 [9–12]. In non-neuronal cells under proliferative activity, CK2 is concentrated in the nucleus [4,13], where it plays funda- mental roles in signalling, gene expression, and other nu- clear processes [14]. In the brain, it has been shown to be enriched in microsomal and soluble, postmicrosomal fractions where it phosphorylates and is found associated 0006-291X/$ - see front matter Ó 2004 Elsevier Inc. All rights reserved. doi:10.1016/j.bbrc.2004.07.158 q Abbreviations: AMPA, a-amino-3 hydroxy-5 methylisoxazole-4- propionate; BDNF, brain-derived neurotrophic factor; DRB, 5,6- dichloro-1-b-ribofuranosyl benzimidazole; ( ) o , extracellular; NMDA, N-methyl-D-aspartate; PSD, postsynaptic density. * Corresponding author. Fax: +5622141752. E-mail address: [email protected] (U. Wyneken). 1 Both authors contributed equally to this work. www.elsevier.com/locate/ybbrc Biochemical and Biophysical Research Communications 322 (2004) 542–550 BBRC

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

www.elsevier.com/locate/ybbrc
Biochemical and Biophysical Research Communications 322 (2004) 542–550
BBRC
Protein kinase CK2 in postsynaptic densities: phosphorylationof PSD-95/SAP90 and NMDA receptor regulationq
Dagoberto Sotoa,1, Floria Pancettib,1, Juan Jose Marengoc, Mauricio Sandovala,Rodrigo Sandovala, Fernando Orregoa, Ursula Wynekena,*
a Neuroscience Laboratory, Faculty of Medicine, Universidad de Los Andes, Santiago-6782468, Chileb Laboratory of Neurosciences, Department of Biology, Faculty of Chemistry and Biology, University of Santiago, Chile
c ICBM, Faculty of Medicine, Universidad de Chile, and Instituto de Neurocirugıa Asenjo, Chile
Received 19 July 2004
Abstract
Protein kinase CK2 (CK2) is highly expressed in rat forebrain where its function is not well understood. Subcellular distribution
studies showed that the catalytic subunit of CK2 (CK2a) was enriched in postsynaptic densities (PSDs) by 68%. We studied the
putative role of CK2 activity on N-methyl-DD-aspartate receptor (NMDAR) function using isolated, patch-clamped PSDs in the pres-
ence of 2 mM extracellular Mg2+. The usual activation by phosphorylation of the NMDARs in the presence of ATP was inhibited
by the selective CK2 inhibitor 5,6-dichloro-1-b-ribofuranosyl benzimidazole (DRB). This inhibition was voltage-dependent, i.e.,
100% at positive membrane potentials, while at negative potentials, inhibition was incomplete. Endogenous CK2 substrates were
characterized by their ability to use GTP as a phosphoryl donor and susceptibility to inhibition by DRB. Immunoprecipitation
assays and 2D gels indicated that PSD-95/SAP90, the NMDAR scaffolding protein, was a CK2 substrate, while the NR2A/B
and NR1 NMDAR subunits were not. These results suggest that postsynaptic NMDAR regulation by CK2 is mediated by indirect
mechanisms possibly involving PSD-95/SAP90.
� 2004 Elsevier Inc. All rights reserved.
Keywords: NMDA receptor; Protein kinase CK2; Postsynaptic density; PSD-95/SAP90; Excitatory neurotransmission; Rat forebrain
Protein kinase CK2 (CK2; formerly called casein ki-
nase 2) is a ubiquitous serine-threonine kinase [1–3], that
is highly concentrated in the brain [4], where it phospho-rylates at least 78 different proteins, including several
ones related to nucleic acid and protein synthesis, tran-
scription factors, cytoskeletal and other structural pro-
teins, as well as a large number of elements involved
in signal transduction [5]. In cells, the majority of CK2
0006-291X/$ - see front matter � 2004 Elsevier Inc. All rights reserved.
doi:10.1016/j.bbrc.2004.07.158
q Abbreviations: AMPA, a-amino-3 hydroxy-5 methylisoxazole-4-
propionate; BDNF, brain-derived neurotrophic factor; DRB, 5,6-
dichloro-1-b-ribofuranosyl benzimidazole; ( )o, extracellular; NMDA,
N-methyl-DD-aspartate; PSD, postsynaptic density.* Corresponding author. Fax: +5622141752.
E-mail address: [email protected] (U. Wyneken).1 Both authors contributed equally to this work.
is found as a heterotetramer consisting of two catalytic
subunits (CK2a or CK2a 0) and two regulatory subunits
(CK2b) [2,6].CK2 is activated in the hippocampus during long-
term potentiation (LTP) [7] and following the applica-
tion of brain-derived neurotrophic factor (BDNF) [8],
and is able to phosphorylate proteins related to plastic-
ity, such as transcription factors of the HMG class,
GAP-43 (B-50) and DARPP-32 [9–12].
In non-neuronal cells under proliferative activity, CK2
is concentrated in the nucleus [4,13], where it plays funda-mental roles in signalling, gene expression, and other nu-
clear processes [14]. In the brain, it has been shown to be
enriched in microsomal and soluble, postmicrosomal
fractions where it phosphorylates and is found associated

D. Soto et al. / Biochemical and Biophysical Research Communications 322 (2004) 542–550 543
to tubulin [15,16]. CK2 activity has been also found in the
crude synaptosomal fraction (P2), where, following fur-
ther fractionation, it was higher in synaptosomes when
compared to light membranes, myelin, and mitochondria
[11]. However, its presence in postsynaptic densities
(PSDs) was not studied by these authors.In a previous work, in which NMDA receptor
(NMDAR) function was studied in acutely dissociated
rat dentate granule cells [17], it was found that inhibitors
of CK2, such as DRB or chrysin, were able to reversibly
reduce NMDAR activity, while spermine, a CK2 activa-
tor, had opposite effects. Synaptic transmission medi-
ated by NMDA but not by AMPA receptors was
reduced by inhibitors of CK2. In that study, however,patch-clamp recordings were done on extrasynaptic
NMDA receptors, and in the absence of extracellular
Mg2+ (Mg2+)o, that induces voltage-dependent block
of the receptors [18]; and the study of synaptic transmis-
sion was done in low (0.3 mM) (Mg2+)o and without
voltage clamping. In many aspects, synaptic receptors
behave differently to extrasynaptic ones [19,20]. Synaptic
NMDARs are anchored to the postsynaptic density(PSD) by means of its scaffolding protein PSD-95/
SAP90 that couples receptor activation to specific down-
stream signalling pathways [21]. Type I PSDs, that are
associated with glutamatergic excitatory synapses, are
electron-dense assemblies of proteins that contain recep-
tors as well as numerous associated proteins, like scaf-
fold and cytoskeletal proteins, that organize the signal
transduction cascades [22]. It is also known that themolecular architecture of PSDs defines the physiological
substrates of kinases in them that in turn may form part
of the several intracellular pathways activated by synap-
tic stimulation [23].
The present work was done to better define the role of
CK2 in the regulation of exclusively synaptic NMDA
receptors from rat forebrain. The subcellular distribu-
tion of CK2, with special reference to its presence inPSDs, was studied, as well as its capacity to phosphory-
late selectively some PSD proteins. We have found that
PSD95/SAP90, an essential protein for the assembly of
the NMDAR complex, is a CK2 substrate.
For the study of NMDAR function, the relatively no-
vel preparation of isolated, patch-clamped PSDs were
used [23–25], and the activity of the NMDARs was stud-
ied in the presence of physiological, millimolar (Mg2+)o[26,27]. This gives evidence that CK2 modulates a Mg2+-
independent component of synaptic NMDAR function.
Materials and methods
Materials. NMDA was purchased from RBI; DRB from Calbio-
chem, [c-32P]GTP from Amersham; protease inhibitor mixture from
Boehringer–Mannheim; antibodies against the a1 subunit of CK2 from
Calbiochem; anti-PSD-95 from Transduction Laboratories; anti-
PKCa/b from Calbiochem; anti-b-tubulin (III) from Sigma; NMDA
receptor subunit NR1 from Pharmingen; and NMDA receptor subunit
NR2A/B from Chemicon. Appropriate horseradish peroxidase-conju-
gated secondary antibodies were used in each case (Calbiochem). All
other chemicals were obtained from Sigma.
Animals. Sprague–Dawley rats (10 weeks old) were maintained and
sacrificed following the recommendations of the government agency
Fondecyt, and the experimental protocols were approved by the Ethics
Committee of Universidad de Los Andes.
Subcellular fractionation. For PSD isolation, the method of Carlin
et al. [28] was slightly modified. Cortices and hippocampi of 10 male
Sprague–Dawley rats were minced and then homogenized, using a
Teflon-glass homogenizer in 5 ml/g wet weight of homogenization
buffer [0.32 M sucrose, 5 mM Hepes, and 0.5 mM EGTA, pH 7.4,
containing a protease inhibitor mixture (Boehringer–Mannheim)].
The homogenate was centrifuged at 1000g for 10 min and the
supernatant (S1) was collected. Pellets were washed once in homog-
enization buffer (5 ml/g wet weight) and cell debris and nuclei (P1)
were removed by a 5 min centrifugation at 500g. Both supernatants
(S1) were centrifuged for 20 min at 12,000g, resulting in a S2
supernatant and a P2 pellet (crude synaptosomal fraction). P2 was
further fractionated by centrifugation in a discontinuous sucrose
gradient (0.85/1/1.2 M sucrose). The gradient loading buffer con-
tained 0.32 M sucrose, 0.5 mM EGTA, 1 mM dithiothreitol (DTT),
and 5 mM Tris, pH 8.1. The following fractions were collected:
myelin (0.32/0.85 M interphase), light membranes (0.85/1 M inter-
phase), synaptosomes (1/1.2 M sucrose interphase), and mitochondria
(pellet). The synaptosomal fraction was lysed for 30 min in 5 volumes
of a hyposmotic solution (0.5 mM EGTA, 5 mM Tris–Cl, pH 8.1)
and centrifuged at 33,000g for 30 min. The resulting pellet was
resuspended for a second centrifugation in the same (0.85/1/1.2 M)
sucrose step gradient to yield synaptic junctional complexes that
equilibrated at the 1/1.2 M sucrose interphase. This fraction was
suspended in 0.32 M sucrose, 5 mM Tris–HCl, pH 8.1, and delipi-
dated by an equal volume of 0.32 M sucrose, 1% Triton X-100,
0.025 mM CaCl2, 2 mM DTT, and 10 mM Tris, pH 8.1 (60 ml/10 g
wet tissue). The suspension was kept on ice for 15 min and centri-
fuged for 30 min at 33,000g. The crystal clear pellet, that contained
the PSDs, was resuspended in 2 ml/g wet tissue of 120 mM KCl,
50 mM Hepes, pH 7.4, and pelleted by a 10 min centrifugation at
100,000g. All steps were performed at 0–4 �C. Protein content was
determined by the bicinchoninic acid method [29].
Western blotting. Twenty micrograms of protein of each subcellular
fraction was dissolved at 1 mg/ml in gel loading buffer, separated by
sodium dodecyl sulfate–polyacrylamide electrophoresis (SDS–PAGE)
on 5–20% gels under fully reducing conditions, and transferred onto
nitrocellulose membranes. Membranes were incubated overnight with
primary antibodies followed by incubation with horseradish peroxi-
dase-conjugated secondary antibody. Immunoreactivity was visualized
using the ECL detection system (Amersham Biosciences).
PSD phosphorylation and autoradiography. PSDs (1 lg/ll) were
incubated for 10 min at 30 �C in a final volume of 50 ll of the buffer
used for electrophysiological recordings (140 mM NaCl, 1 mM DTT,
100 lM EDTA, and 10 mM Hepes–Na, pH 7.4) containing 2 mM
GTP supplemented with 20 lCi [c-32P]GTP, 2 mM Mg2+, 2 mM NaF,
and protease inhibitor mixture (Boehringer–Mannheim). When indi-
cated, the reaction mixture contained 6, 12, 50 or 100 lM DRB dis-
solved in DMSO (1:416 in the reaction mixture); the control contained
1:416 DMSO alone. Reaction was stopped by addition of ice-cold
trichloroacetic acid (final concentration 10% v/v) followed by two
washes with acetone at �20 �C. Precipitated proteins were dried and
dissolved in loading buffer and proteins were separated by SDS–
PAGE. Gels were dried and autoradiographed on Kodak X-Omat MR
films at �70 �C.2D gel electrophoresis. The procedure was carried out using
immobilized pH gradients and standard procedures following
Amersham Biosciences recommendations. PSD samples (200 lg) were

Fig. 1. Subcellular distribution of CK2a subunit and of PSD-95 in rat
cortex and hippocampus. Fractions correspond to those obtained
following Carlin et al. [28]. Equal amounts of protein of each fraction
were loaded on a 5–20% SDS–polyacrylamide gel, and immunodetec-
tion of PSD-95 and of CK2a was performed. P1 and S1 represent the
pellet and supernatant, respectively, following the centrifugation at
1000g, while P2 and S2 represent the pellet and supernatant obtained
following the centrifugation of S1 at 12,000g. Myelin, light mem-
branes, synaptosomes, and mitochondria represent the fractions
obtained when the resuspended P2 was loaded on a discontinuous
(0.32/0.85/1/1.2 M) sucrose gradient. The CK2a and PSD-95 immun-
odetections were done in at least three different subcellular fraction-
ation procedures.
544 D. Soto et al. / Biochemical and Biophysical Research Communications 322 (2004) 542–550
solubilized in 250 ll lysis buffer for 1 h (7 M urea, 2 M thiourea, 2%
CHAPS, 20 mM Tris, pH 7.5, 1% DTT, and 0.5% IPG buffer 3–10)
and then centrifuged at 40,000g for 1 h. Two hundred forty micro-
liters of the supernatant was used for the rehydration and loading of
the proteins to IPG strips (Immobiline 13 cm DryStrip 3–10 NL,
Amersham) at 50 V for 12 h. The voltage was increased to 500 V for
1 h, 1000 V for 1 h, and 8000 V for 8 h. Immediately after being fo-
cused, IPG strips were wrapped in plastic foil and stored at �80 �C.Prior to SDS–PAGE, IPG strips were equilibrated in 6 M urea, 2%
SDS, 1% DTT, 75 mM Tris, pH 8.8, and 30% glycerol for 10 min,
followed by equilibration in 6 M urea, 2% SDS, 2.5% iodoacetamide,
75 mM Tris, pH 8.8, and 30% glycerol for 10 min. The second
dimension separation was run overnight in 1.5 mm 11% Duracryl gels
(Genomic Solutions) using a Hoefer SE600 Ruby electrophoresis
unit at 12 mA/gel at 15 �C. After electrophoresis, gels were trans-
ferred to nitrocellulose membrane for Western blot analysis and
autoradiography.
Electrophysiological recordings. Giant liposomes containing PDSs
were prepared as described earlier [24,25]. Briefly, a mixture of 100–
150 lg PSD proteins was added to 20 mg of azolectin (LL-a-lecithin,type II-S, Sigma) vesicles. The mixture was subjected to a partial
dehydration/rehydration cycle at 4 �C in a medium containing 5%
ethylene glycol. The giant liposomes obtained, into which PSDs had
been incorporated, were placed in a 35 mm Petri dish and covered with
1 ml of recording buffer (140 mM NaCl, 1 mM DTT, 100 lM EDTA,
and 10 mM Hepes–Na, pH 7.4). Giga-seals (10–20 GX) were obtainedwith pipettes of 5–10 MX resistance for the open electrode. After
sealing, withdrawal of the pipette from the liposome surface resulted in
an inside-out configuration. Reconstituted glutamate receptors were
activated by injection of 1 ll of 50 lM NMDA and 5 lM glycine into
the recording pipette, which contained about 5 ll of recording solutionand 2 mMMg2+, through a fine plastic catheter positioned within
3 mm of its tip. The holding potential was applied to the interior of the
patch pipette, and the bath electrode was maintained at ground. Patch
clamp recordings were obtained using an EPC-9 patch amplifier
(HEKA Electronics) at a gain of 50 mV/pA. Current records were
stored using a video recorder and a filter setting of 10 kHz. For
analysis, recordings were filtered at 2 kHz and digitized at 4 kHz. Data
analysis, like mean current values and current amplitude histograms of
1-min current traces, was performed using the Microcal Origin soft-
ware. All measurements were performed at room temperature.
For inhibition experiments, 12 or 24 ll of 0.5 mM DRB in 1:10
DMSO was added to the ‘‘intracellular’’ recording solution (i.e., to the
petri dish) to obtain final concentrations of 6 or 12 lM and maximally
1:416 of DMSO.
Immunoprecipitation. Immunoprecipitations were performed in
phosphorylated and solubilized PSDs. Briefly, 200 lg of PSD proteins
was solubilized during 2 h in solubilization buffer (50 mM Tris–HCl,
pH 7.5, 150 mM NaCl, 10 mM EDTA, 2 mM EGTA, 0.1% SDS, 1%
Triton X-100, 1% CHAPS, 0.5% NP-40, 0.1% BSA, and 50 mM NaF
plus proteases inhibitors), under constant agitation at 4 �C. The
remaining particulate material was discarded by centrifugation (5 min
at 9500g) and 1 lg of the corresponding primary antibody was added
to the supernatant to interact overnight at 4 �C. For controls, normal
IgG was used. Subsequently 150 ll protein G–Sepharose (pre-washed
with solubilization buffer) was added and incubated for 1 h at 4 �Cunder agitation. The samples were centrifuged for 5 min at 10,000g and
the supernatant was discarded. The immunoprecipitates were washed
three times with buffer containing 50 mM Tris–HCl, pH 7.5; 150 mM
NaCl; 0.1% SDS; and 1% Triton X-100, 1% CHAPS; and 0.5% NP-40
plus protease inhibitors and were resuspended in 50 ll of electropho-resis loading buffer.
Statistical analysis of densitometric quantification of the Western
blots was performed (two-tailed, one sample Student�s t test) using
InStat3 obtained from GraphPad (San Diego, CA, USA). For statis-
tical comparisons of fractional changes ranging from 0 to 1 the arcsine
transformation was applied.
Results
Subcellular distribution of CK2a subunit
CK2a subunit was present in all the subcellular
fractions studied in a similar relative quantity whencompared with homogenate, with the exception of mito-
chondria (Fig. 1). However, it was enriched in PSDs by
68% ± 9% (mean ± SEM, p < 0.01), in relation to the ini-
tial homogenate. A very low concentration was found in
myelin and it was not detectable in mitochondria. CK2awas also present in the microsomal fraction (P3) at a sim-
ilar concentration relative to the homogenate, and was
enriched by 42% in the postmicrosomal (S3) fraction(not shown). Although CK2a is enriched in PSDs, as
these structures represent only about 0.03% of the protein
present in the original homogenate (Table 1), the total
amount of the enzyme bound to PSDs is comparatively
low.
As a control for the subcellular fractionation method,
the distribution of PSD-95/SAP90, the scaffolding pro-
tein present in excitatory PSDs, is also shown (Fig. 1,lower panel). As it was expected, PSD-95/SAP90 was en-
riched by 18.8 ± 2.3-fold (mean ± SEM, n = 4) in PSDs
when compared with homogenates. This protein was
also present in all those fractions where synaptosomes
are known to be present [30,31]: in the P1 ‘‘nuclear’’
fraction, that also contains large synaptosomes; in the
crude ‘‘synaptosomal’’ P2 fraction and purified synapto-
somal subfractions. PSD-95/SAP90 was also detected inthe light membrane fraction, where its presence has been
associated with intracellular trafficking on the cytoplas-
mic surface of endomembranes to and from postsynap-
tic sites [32,33]. It was absent from myelin and

Table 1
Protein distribution among subcellular fractions
Percent of homogenate protein
Primary fractions
P1 55.4
P2 17.7
P3 4.4
S3 23.3
Percent of P2 protein
P2 subfractions
Myelin 25.8
Light membranes 16.2
Synaptosomes 37.7
Mitochondria 10.5
PSDs 0.16
Protein concentration was measured in each subcellular fraction and
the mean protein content of the fractions calculated in at least three
independent experiments.
D. Soto et al. / Biochemical and Biophysical Research Communications 322 (2004) 542–550 545
mitochondria, and also from S2, that contains frag-
mented endoplasmic reticulum and other cell subfrac-
tions, including the soluble cytoplasm. This pattern of
distribution was much more restricted than that of
CK2a, due to the well-known ubiquity of this protein,whereas PSD-95/SAP90 is a specific marker of PSDs.
As CK2a subunit was present in PSDs, we further
wanted to test whether it was able to regulate synaptic
NMDAR currents and Mg2+ blockade.
Effect of endogenous protein kinase CK2 on the activity of
NMDA receptors present in PSDs
As previously described [23–25], addition of 10 lMNMDA plus 1 lM glycine to isolated patch-clamped
PSDs induced, in a previously electrically silent seal,
complex currents due to the asynchronous openings
and closures of the NMDA receptor channels present
in this structure (Fig. 2A-a). As 2 mM Mg2+ was present
at the ‘‘extracellular’’ face of the PSD (i.e., in the record-
ing pipette), the typical voltage-dependent block was ob-served at negative ‘‘intracellular’’ potentials, while at
positive membrane potentials the currents were much
larger (Fig. 2A-a). This may be clearly seen also in the
one-minute current amplitude histograms on the right
hand of the figure. In the same experiment, the subse-
quent addition of 2 mM ATP-Mg2+ to the bath led to
the usual activation of several endogenous protein ki-
nases present in the PSD. An increase in the currents re-corded both at positive and negative membrane
potentials was obtained (Fig. 2A-b). When 6 lMDRB, a selective inhibitor of CK2 [34], was added to
the seal followed by a 10 min incubation, a marked inhi-
bition of the ATP-enhanced current was seen at positive
potentials which returned to control, pre-phosphoryla-
tion values (no significant difference with control,
n = 3, p > 0.5). On the contrary, a small inhibitory effect
was observed on the ATP-enhanced currents seen at
negative potentials, which remained increased by
74 ± 21% (SD, n = 3, p < 0.05, when compared to con-
trols) (Fig. 2A-c).
In another set of experiments, in which NMDARs
present in the PSD were also activated by 10 lMNMDA and 1 lM glycine (Fig. 2B-a), addition of
Mg2+-ATP plus 12 lM DRB induced an increase in
the currents seen at negative, but not at positive, poten-
tials (Fig. 2B-b). When the same seal was perfused ‘‘in-
tracellularly’’ with the bath fluid supplemented with
Mg2+-ATP but in the absence of DRB, it was found
that the inhibitory effect of DRB on positive currents
disappeared, and that ATP led to the usual enhance-ment of NMDAR currents both at positive and nega-
tive potentials (Fig. 2B-c). Mean conductances in the
membrane patch (±SD) within each minute, for each
consecutive minute of recording, were calculated for
each of the experiments done at ±50, ±70, and
±90 mV. It should be pointed out that no single channel
measurements are possible in this system, because the
NMDAR channel clusters in PSDs flicker rapidly andopen asynchronously. For this reason, the mean one-
minute conductance of a seal is a measure of activity
that integrates the charge movement in one-minute of
recording. Mean patch conductances were plotted in
Fig. 2C for each situation. After the addition of ATP-
Mg2+ plus 6 lM DRB, conductances at positive mem-
brane potentials remained unchanged when compared
to controls, while at negative potentials an increase of71 ± 5% (mean ± SEM) was observed. When DRB
was washed out, positive currents, following a 10 min
lag, increased by 145 ± 11.8% whereas at negative mem-
brane potentials, an additional increase of only 32.4%
was observed. No significant differences were found
with 6 or 12 lM DRB. The time lag between DRB
washout and conductance increase was about 10 min.
This indicated that DRB had no direct effect on theNMDA receptor channel that should be immediate fol-
lowing DRB washout.
CK2 substrates in PSDs
To look for endogenous CK2 substrates in isolated
PSDs, these were incubated with 2 mM Mg2+ and
[c-32P]GTP, a phosphoryl donor that CK2, but not anumber of other protein kinases, can use almost as effi-
ciently as ATP [35]. The phosphorylation of the different
PSD proteins by an endogenous CK2 activity was par-
tially inhibited by 6 and 12 lM DRB, thus suggesting
that they correspond to synaptic proteins whose func-
tion may be modified by this kinase (Fig. 3). In order
to visualize the high molecular weight CK2 substrates,
short exposure times were used (Fig. 3A) whereas longerexposure times were used for the low molecular weight
substrates (Fig. 3B). The quantification of inhibition
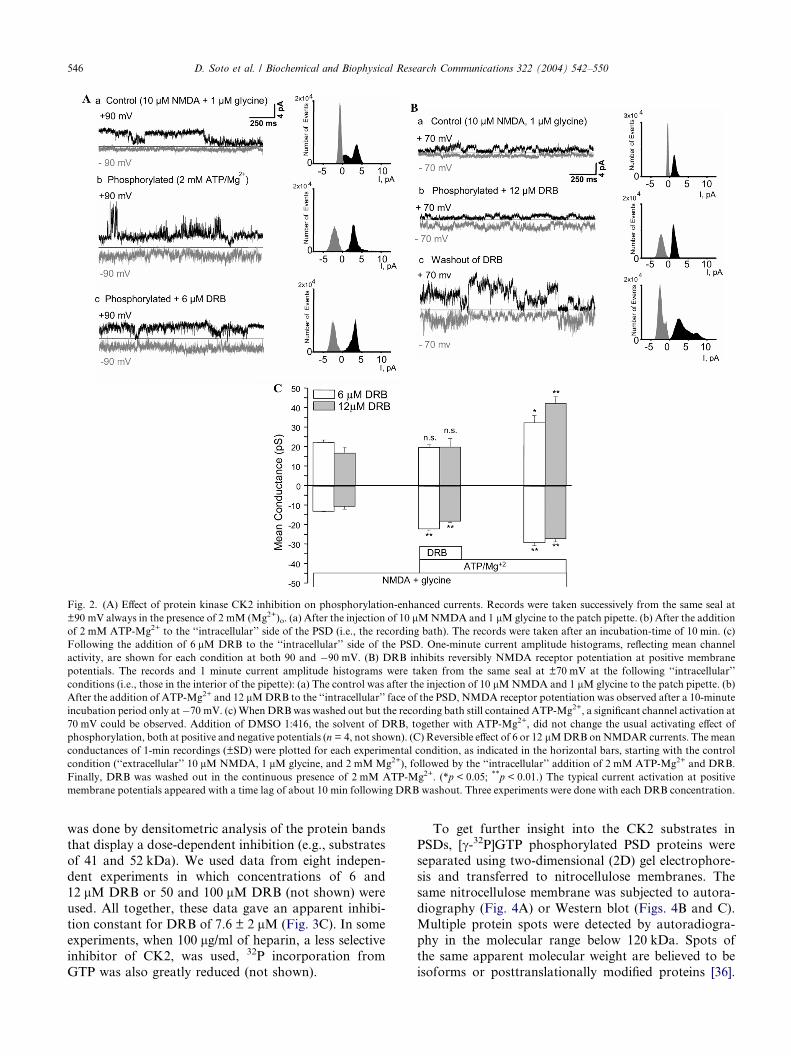
Fig. 2. (A) Effect of protein kinase CK2 inhibition on phosphorylation-enhanced currents. Records were taken successively from the same seal at
±90 mV always in the presence of 2 mM (Mg2+)o. (a) After the injection of 10 lMNMDA and 1 lMglycine to the patch pipette. (b) After the addition
of 2 mM ATP-Mg2+ to the ‘‘intracellular’’ side of the PSD (i.e., the recording bath). The records were taken after an incubation-time of 10 min. (c)
Following the addition of 6 lM DRB to the ‘‘intracellular’’ side of the PSD. One-minute current amplitude histograms, reflecting mean channel
activity, are shown for each condition at both 90 and �90 mV. (B) DRB inhibits reversibly NMDA receptor potentiation at positive membrane
potentials. The records and 1 minute current amplitude histograms were taken from the same seal at ±70 mV at the following ‘‘intracellular’’
conditions (i.e., those in the interior of the pipette): (a) The control was after the injection of 10 lMNMDA and 1 lM glycine to the patch pipette. (b)
After the addition of ATP-Mg2+ and 12 lMDRB to the ‘‘intracellular’’ face of the PSD, NMDA receptor potentiation was observed after a 10-minute
incubation period only at�70 mV. (c)WhenDRBwas washed out but the recording bath still contained ATP-Mg2+, a significant channel activation at
70 mV could be observed. Addition of DMSO 1:416, the solvent of DRB, together with ATP-Mg2+, did not change the usual activating effect of
phosphorylation, both at positive and negative potentials (n = 4, not shown). (C) Reversible effect of 6 or 12 lMDRB onNMDAR currents. The mean
conductances of 1-min recordings (±SD) were plotted for each experimental condition, as indicated in the horizontal bars, starting with the control
condition (‘‘extracellular’’ 10 lM NMDA, 1 lM glycine, and 2 mMMg2+), followed by the ‘‘intracellular’’ addition of 2 mM ATP-Mg2+ and DRB.
Finally, DRB was washed out in the continuous presence of 2 mM ATP-Mg2+. (*p < 0.05; **p < 0.01.) The typical current activation at positive
membrane potentials appeared with a time lag of about 10 min following DRB washout. Three experiments were done with each DRB concentration.
546 D. Soto et al. / Biochemical and Biophysical Research Communications 322 (2004) 542–550
was done by densitometric analysis of the protein bands
that display a dose-dependent inhibition (e.g., substrates
of 41 and 52 kDa). We used data from eight indepen-dent experiments in which concentrations of 6 and
12 lM DRB or 50 and 100 lM DRB (not shown) were
used. All together, these data gave an apparent inhibi-
tion constant for DRB of 7.6 ± 2 lM (Fig. 3C). In some
experiments, when 100 lg/ml of heparin, a less selective
inhibitor of CK2, was used, 32P incorporation from
GTP was also greatly reduced (not shown).
To get further insight into the CK2 substrates in
PSDs, [c-32P]GTP phosphorylated PSD proteins were
separated using two-dimensional (2D) gel electrophore-sis and transferred to nitrocellulose membranes. The
same nitrocellulose membrane was subjected to autora-
diography (Fig. 4A) or Western blot (Figs. 4B and C).
Multiple protein spots were detected by autoradiogra-
phy in the molecular range below 120 kDa. Spots of
the same apparent molecular weight are believed to be
isoforms or posttranslationally modified proteins [36].
D. Soto et al. / Biochemical and Biophysical Research Communications 322 (2004) 542–550 547
PSD-95/SAP90, an intracellular protein that anchors the
NMDAR to the PSD, was immunodetected as an elon-
gated spot that could be superimposed on an autoradio-
graphic spot. As a positive control of a known CK2
substrate, tubulin was immunodetected. In a different
set of experiments, PSD proteins were phosphorylatedin the presence or absence of 12 lM DRB and resolved
by 2D electrophoresis (n = 2). It could be seen that the
spot corresponding to PSD-95/SAP90 was DRB sensi-
tive (Figs. 4D and E). In these experiments, a mean inhi-
bition of 38 ± 6% was observed using well-resolved
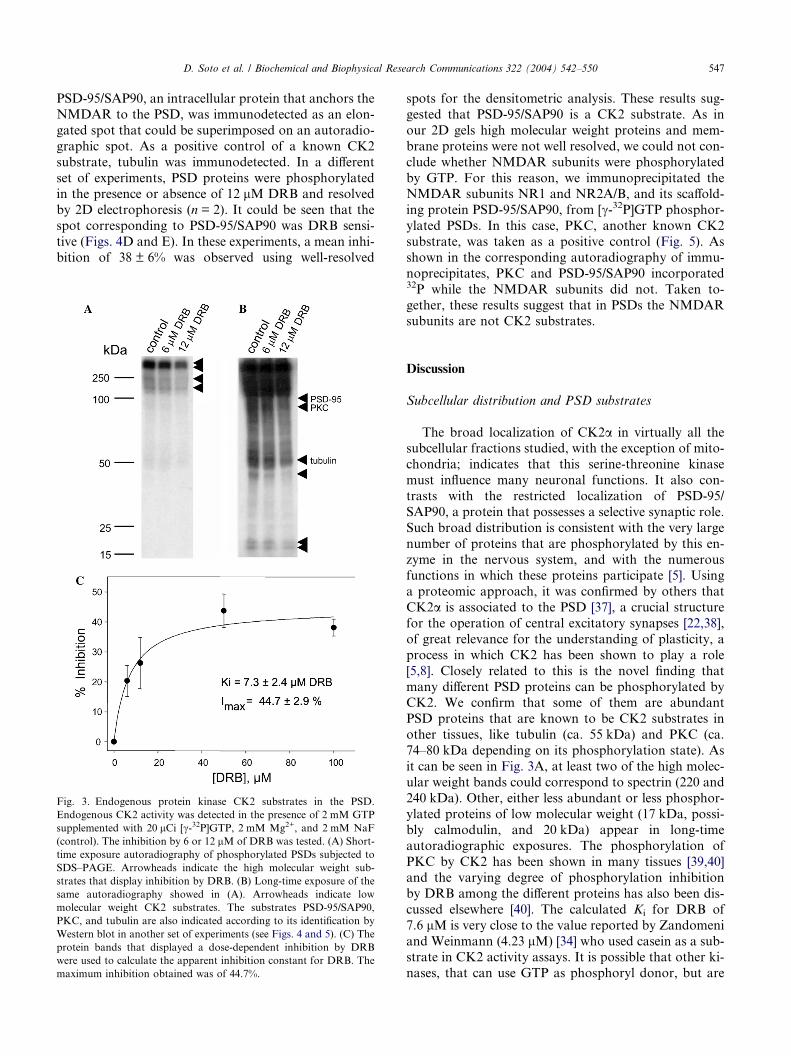
Fig. 3. Endogenous protein kinase CK2 substrates in the PSD.
Endogenous CK2 activity was detected in the presence of 2 mM GTP
supplemented with 20 lCi [c-32P]GTP, 2 mM Mg2+, and 2 mM NaF
(control). The inhibition by 6 or 12 lM of DRB was tested. (A) Short-
time exposure autoradiography of phosphorylated PSDs subjected to
SDS–PAGE. Arrowheads indicate the high molecular weight sub-
strates that display inhibition by DRB. (B) Long-time exposure of the
same autoradiography showed in (A). Arrowheads indicate low
molecular weight CK2 substrates. The substrates PSD-95/SAP90,
PKC, and tubulin are also indicated according to its identification by
Western blot in another set of experiments (see Figs. 4 and 5). (C) The
protein bands that displayed a dose-dependent inhibition by DRB
were used to calculate the apparent inhibition constant for DRB. The
maximum inhibition obtained was of 44.7%.
spots for the densitometric analysis. These results sug-
gested that PSD-95/SAP90 is a CK2 substrate. As in
our 2D gels high molecular weight proteins and mem-
brane proteins were not well resolved, we could not con-
clude whether NMDAR subunits were phosphorylated
by GTP. For this reason, we immunoprecipitated theNMDAR subunits NR1 and NR2A/B, and its scaffold-
ing protein PSD-95/SAP90, from [c-32P]GTP phosphor-
ylated PSDs. In this case, PKC, another known CK2
substrate, was taken as a positive control (Fig. 5). As
shown in the corresponding autoradiography of immu-
noprecipitates, PKC and PSD-95/SAP90 incorporated32P while the NMDAR subunits did not. Taken to-
gether, these results suggest that in PSDs the NMDARsubunits are not CK2 substrates.
Discussion
Subcellular distribution and PSD substrates
The broad localization of CK2a in virtually all thesubcellular fractions studied, with the exception of mito-
chondria; indicates that this serine-threonine kinase
must influence many neuronal functions. It also con-
trasts with the restricted localization of PSD-95/
SAP90, a protein that possesses a selective synaptic role.
Such broad distribution is consistent with the very large
number of proteins that are phosphorylated by this en-
zyme in the nervous system, and with the numerousfunctions in which these proteins participate [5]. Using
a proteomic approach, it was confirmed by others that
CK2a is associated to the PSD [37], a crucial structure
for the operation of central excitatory synapses [22,38],
of great relevance for the understanding of plasticity, a
process in which CK2 has been shown to play a role
[5,8]. Closely related to this is the novel finding that
many different PSD proteins can be phosphorylated byCK2. We confirm that some of them are abundant
PSD proteins that are known to be CK2 substrates in
other tissues, like tubulin (ca. 55 kDa) and PKC (ca.
74–80 kDa depending on its phosphorylation state). As
it can be seen in Fig. 3A, at least two of the high molec-
ular weight bands could correspond to spectrin (220 and
240 kDa). Other, either less abundant or less phosphor-
ylated proteins of low molecular weight (17 kDa, possi-bly calmodulin, and 20 kDa) appear in long-time
autoradiographic exposures. The phosphorylation of
PKC by CK2 has been shown in many tissues [39,40]
and the varying degree of phosphorylation inhibition
by DRB among the different proteins has also been dis-
cussed elsewhere [40]. The calculated Ki for DRB of
7.6 lM is very close to the value reported by Zandomeni
and Weinmann (4.23 lM) [34] who used casein as a sub-strate in CK2 activity assays. It is possible that other ki-
nases, that can use GTP as phosphoryl donor, but are
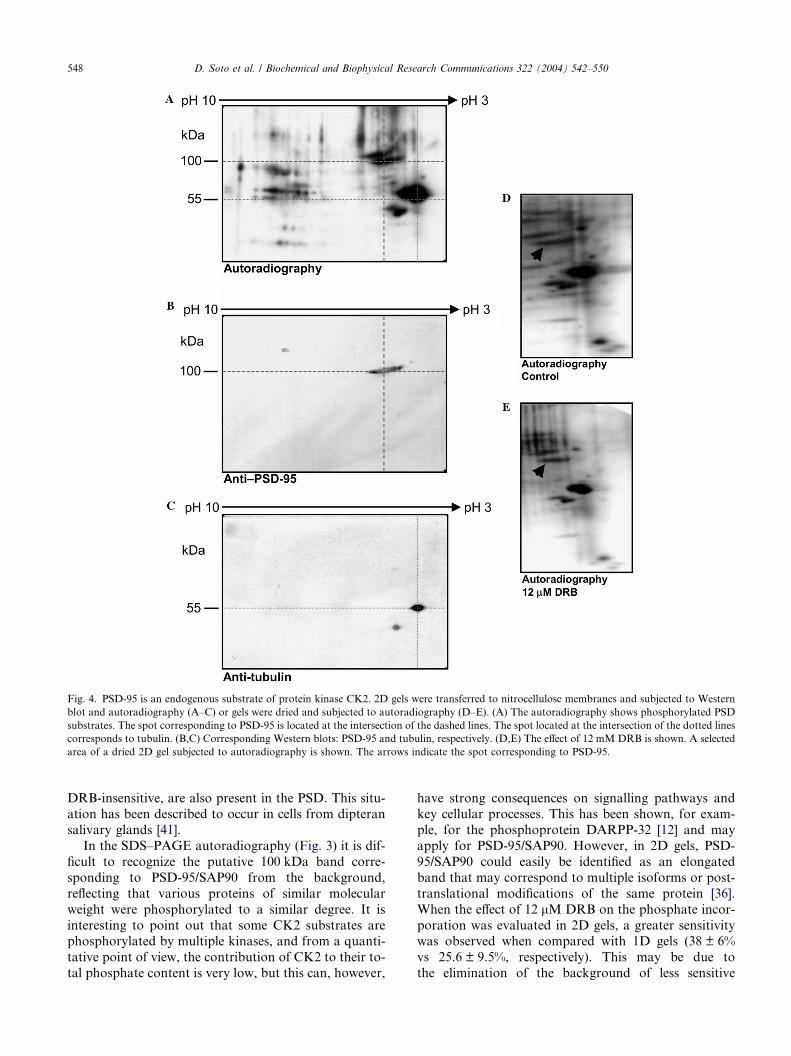
Fig. 4. PSD-95 is an endogenous substrate of protein kinase CK2. 2D gels were transferred to nitrocellulose membranes and subjected to Western
blot and autoradiography (A–C) or gels were dried and subjected to autoradiography (D–E). (A) The autoradiography shows phosphorylated PSD
substrates. The spot corresponding to PSD-95 is located at the intersection of the dashed lines. The spot located at the intersection of the dotted lines
corresponds to tubulin. (B,C) Corresponding Western blots: PSD-95 and tubulin, respectively. (D,E) The effect of 12 mM DRB is shown. A selected
area of a dried 2D gel subjected to autoradiography is shown. The arrows indicate the spot corresponding to PSD-95.
548 D. Soto et al. / Biochemical and Biophysical Research Communications 322 (2004) 542–550
DRB-insensitive, are also present in the PSD. This situ-ation has been described to occur in cells from dipteran
salivary glands [41].
In the SDS–PAGE autoradiography (Fig. 3) it is dif-
ficult to recognize the putative 100 kDa band corre-
sponding to PSD-95/SAP90 from the background,
reflecting that various proteins of similar molecular
weight were phosphorylated to a similar degree. It is
interesting to point out that some CK2 substrates arephosphorylated by multiple kinases, and from a quanti-
tative point of view, the contribution of CK2 to their to-
tal phosphate content is very low, but this can, however,
have strong consequences on signalling pathways andkey cellular processes. This has been shown, for exam-
ple, for the phosphoprotein DARPP-32 [12] and may
apply for PSD-95/SAP90. However, in 2D gels, PSD-
95/SAP90 could easily be identified as an elongated
band that may correspond to multiple isoforms or post-
translational modifications of the same protein [36].
When the effect of 12 lM DRB on the phosphate incor-
poration was evaluated in 2D gels, a greater sensitivitywas observed when compared with 1D gels (38 ± 6%
vs 25.6 ± 9.5%, respectively). This may be due to
the elimination of the background of less sensitive

Fig. 5. Immunoprecipitation of PSD proteins after phosphorylation.
PSD proteins were phosphorylated with GTP supplemented with
[c-32P]GTP. The left panel corresponds to the Western blot of the
indicated proteins: NMDAR subunits NR1 and NR2 A/B; PKC and
PSD-95. The immunodetection was performed in the PSD (positive
control) and in the immunoprecipitates (IPP). As control for the
immunoprecipitation a normal IgG of the corresponding type was
used (mouse or rabbit). The right panel shows the corresponding
autoradiographies.
D. Soto et al. / Biochemical and Biophysical Research Communications 322 (2004) 542–550 549
substrates present in the same molecular weight band,
because of in 2D gels each protein is much better re-
solved. The immunoprecipitation experiments also con-firmed that PSD-95/SAP90, but not the NMDAR
subunits NR1 and NR2A/B, was a CK2 substrate.
Effect of CK2 on NMDA receptors
The results obtained, in which inhibition of CK2
activity by DRB markedly depressed the activity of the
receptors at positive membrane potentials, i.e., whenthey were not blocked by (Mg2+)o, support the findings
of Lieberman and Mody [17] in patch-clamped extrasy-
naptic receptors that were obtained in the absence of
(Mg2+)o. However, in the present study, inhibition of
CK2 had less effect at negative membrane potentials in
the presence of (Mg2+)o, where voltage-dependent block
of the receptor channels is present [18]. This shows that,
under physiological conditions, in which (Mg2+)o ispresent at about 0.9 mM [26,27], CK2 will only enhance
the activity of NMDA receptors when the cells are depo-
larized. In the synaptic receptors studied here, such
CK2-dependent activation should be quite large when
the dendritic spines, where PSDs are predominantly lo-
cated, become depolarized by the activation of other
glutamatergic receptors and thus, may affect specifically
plastic phenomena that are induced in a LTP-like man-ner. In contrast, the NMDAR activation by phosphory-
lation at negative membrane potentials should be caused
by a different set of kinases that may include src kinase,
PKC or others [42,43]. The molecular mechanism of
activation of NMDARs by CK2 is not yet clear. It has
been reported that the interaction of PSD-95/SAP90
with NMDARs modulates their function [44–47]. It re-
mains to be figured out whether the interaction of the
NMDAR subunits NR2A/B with PSD-95/SAP90 is reg-
ulated by CK2. This, among other mechanisms, could
be due to the possibility that phosphorylated PSD-95/
SAP90 could bring an inhibitory protein close to theNMDAR in a phosphorylation-dependent manner. This
may be the case for calmodulin or protein phosphatases
[48–50]. In any case, our results show that CK2 activates
synaptic NMDARs through indirect mechanisms that
may involve components of the NMDAR complex like
PSD-95/SAP90.
Acknowledgments
We thank Professor Dr. Jorge Allende for helpful
comments, and Fermelo for providing us with Amer-
sham equipment. This work was supported by Fondecyt
Project 1020257 and by Universidad de los Andes
Projects.
References
[1] L.A. Pinna, Casein kinase 2: an ‘‘eminence grise’’ in cellular
regulation?, Biochim. Biophys. Acta 1054 (1990) 267–284.
[2] J.E. Allende, C.C. Allende, Protein kinase C2: an enzyme with
multiple substrates and a puzzling regulation, FASEB J. 9 (1995)
313–323.
[3] H. Meisner, M.P. Czech, Casein kinase II (vertebrates), in: G.
Hardie, S. Hanks (Eds.), The Protein Kinase Facts Book. Protein-
Serine Kinases, Academic Press, London, 1995, pp. 240–242.
[4] W. Krek, G. Maridor, E.A. Nigg, Casein kinase II is a
predominantly nuclear enzyme, J. Cell Biol. 116 (1992) 43–55.
[5] P.R. Blanquet, Identification of two persistently activated neuro-
trophin-regulated pathways in rat hippocampus, Neuroscience 95
(2000) 705–719.
[6] D.W. Litchfield, Protein kinase CK2: structure, regulation and
role in cellular decisions of life and death, Biochem. J. 369 (2003)
1–15.
[7] C. Charriaut-Marlangue, S. Otani, C. Creuzet, Y. Ben-Ari, J.
Loeb, Rapid activation of hippocampal casein kinase II during
long-term potentiation, Proc. Natl. Acad. Sci. USA 88 (1991)
10232–10236.
[8] P.R. Blanquet, Neurotrophin-induced activation of casein kinase
2 in rat hippocampal slices, Neuroscience 86 (1998) 739–749.
[9] B.A. Reikhardt, O.G. Kulikova, G.Y. Borisova, I.Y. Aleksandr-
ova, N.S. Sapronov, Status of the protein kinase CK2-HMG14
system in age-related amnesia in rats, Neurosci. Behav. Physiol. 33
(2003) 799–804.
[10] M. Pisano, M.A. Hegazi, E.M. Reimann, L.A. Dokas, Phosphor-
ylation of protein B-50 (GAP-43) from adult rat brain cortex by
casein kinase 2, Biochim. Biophys. Res. Commun. 155 (1988)
1207–1212.
[11] J.A. Girault, H.C. Hemmings, S.H. Zorn, E.L. Gustafson, P.
Greengard, Characterization in mammalian brain of a DARPP-32
serine kinase identical to casein kinase II, J. Neurochem. 55 (1990)
1772–1783.
[12] P. Svenningsson, A. Nishi, G. Fisone, J.A. Girault, A.C. Nairn,
P. Greengard, DARPP-32: an integrator of neurotransmission,
Annu. Rev. Pharmacol. Toxicol. 44 (2004) 269–296.

550 D. Soto et al. / Biochemical and Biophysical Research Communications 322 (2004) 542–550
[13] O.G. Issinger, Casein kinases: pleiotropic mediators of cellular
regulation, Pharmacol. Ther. 59 (1993) 1–30.
[14] L.A. Pinna, Protein kinase CK2: a challenge to canons, J. Cell Sci.
115 (2002) 3873–3878.
[15] L. Serrano, J. Dıaz-Nido, F. Wandosell, J. Avila, Tubulin
phosphorylation by casein kinase II is similar to that found in
vivo, J. Cell Biol. 105 (1987) 1731–1739.
[16] M.E. Martin, A. Alcazar, M. Salinas, Subcellular and regional
distribution of casein kinase II and initiation factor 2 activities
during rat brain development, Int. J. Dev. Neurosci. 8 (1990) 47–
54.
[17] D.N. Lieberman, I. Mody, Casein kinase II regulates NMDA
channel function in hippocampal neurons, Nat. Neurosci. 2 (1999)
125–132.
[18] L. Nowak, P. Bregestovski, P. Ascher, A. Herbet, A. Prochiantz,
Magnesium gates glutamate-activated channels in mouse central
neurons, Nature (Lond.) 307 (1984) 462–465.
[19] G.E. Hardingham, Y. Fukunaga, H. Bading, Extrasynaptic
NMDARs oppose synaptic NMDARs by triggering CREB
shut-off and cell death pathways, Nat. Neurosci. 5 (2002) 405–
414.
[20] P. Vanhoutte, H. Bading, Opposing roles of synaptic and
extrasynaptic NMDA receptors in neuronal calcium signalling
and BDNF gene regulation, Curr. Opin. Neurobiol. 13 (2003)
366–371.
[21] M. Sheng, C. Sala, PDZ domains and the organization of
supramolecular complexes, Annu. Rev. Neurosci. 24 (2001) 1–29.
[22] M.B. Kennedy, Signal-processing machines at the postsynaptic
density, Science 290 (2000) 750–754.
[23] U. Wyneken, J.J. Marengo, F. Orrego, Electrophysiology and
plasticity in isolated postsynaptic densities, Brain Res. Rev. (2004)
(in press).
[24] G. Riquelme, U. Wyneken, S. Villanueva, F. Orrego, Recordings
of glutamate receptor channels in isolated postsynaptic densities,
Neuroreport 4 (1993) 1163–1166.
[25] U. Wyneken, G. Riquelme, S. Villanueva, F. Orrego, Effect of
glutamate receptor phosphorylation by endogenous protein
kinases on electrical activity of isolated postsynaptic densities
of rat cortex and hippocampus, Neurosci. Lett. 224 (1997) 131–
135.
[26] M.W. Bradbury, C.R. Kleeman, H. Bagdoyan, A. Berberian, The
calcium and magnesium content of skeletal muscle, brain and
cerebrospinal fluid as determined by atomic absorption flame
photometry, J. Lab. Clin. Med. 71 (1968) 884–892.
[27] J.G. Chutkow, Metabolism of magnesium in central nervous
system, Neurology 24 (1974) 780–787.
[28] R. Carlin, D.J. Grab, R.S. Cohen, P. Siekevitz, Isolation and
characterization of postsynaptic densities from various brain
regions: enrichment of different types of postsynaptic densities, J.
Cell Biol. 86 (1980) 831–843.
[29] P. Smith, R. Krohn, G. Hermanson, A. Mallia, F. Gartner, M.
Provenzano, E. Fujimoto, N. Goeke, B. Olson, D. Klenk,
Measurement of protein using bicinchoninic acid, Anal. Bio-
chem. 150 (1985) 76–85 [and erratum in Anal. Biochem. 163
(1985) 279.
[30] E. De Robertis, A. Pellegrino de Iraldi, G. Rodrıguez de Lores
Arnaiz, L. Salganicoff, Cholinergic and non-cholinergic nerve
endings in rat brain, I. Isolation and subcellular distribution of
acetylcholine and acetylcholinesterase, J. Neurochem. 9 (1962) 23–
35.
[31] V.P. Whittaker, The application of subcellular fractionation
techniques to the study of brain function, Prog. Biophys. Mol.
Biol. 15 (1965) 39–96.
[32] H. Wu, J.E. Nash, P. Zamorano, C.C. Garner, Interaction of
SAP97 with minus-end-directed actin motor myosin VI. Implica-
tions for AMPA receptor trafficking, J. Biol. Chem. 34 (2002)
30928–30934.
[33] D.M Chetkovich, R.C. Bunn, S.-H. Kuo, Y. Kawasaki, M.
Kohwi, D.S. Bredt, Postsynaptic targeting of alternative postsyn-
aptic density-95 isoforms by distinct mechanisms, J. Neurosci. 22
(2002) 6415–6425.
[34] R. Zandomeni, R. Weinmann, Inhibitory effect of 5,6-dichloro-1-
b-DD-ribofuranosylbenzimidazole on a protein kinase, J. Biol.
Chem. 259 (1984) 14804–14811.
[35] G.M. Hathaway, J.A. Traugh, Cyclic nucleotide independent
protein kinases from rabbit reticulocytes. Purification of casein
kinases, J. Biol. Chem. 254 (1979) 762–768.
[36] K.W. Li, M.P. Hornshaw, R.C. Van Der Schors, R. Watson, S.
Tate, B. Casetta, C.R. Jimenez, Y. Gouwenberg, E.D. Gundel-
finger, K.H. Smalla, A.B. Smit, Proteomics analysis of rat brain
postsynaptic density. Implications of the diverse protein func-
tional groups for the integration of synatic physiology, J. Biol.
Chem. 279 (2004) 987–1002.
[37] Y. Yoshimura, Y. Yamauchi, T. Shinkawa, M. Taoka, H. Donai,
N. Takahashi, T. Isobe, T. Yamauchi, Molecular constituents of
the postsynaptic density fraction revealed by proteomic analysis
using multidimensional liquid chromatography–tandem mass
spectrometry, J. Neurochem. 88 (2004) 759–768.
[38] E.B. Ziff, Enlightening the postsynaptic density, Neuron 19 (1997)
1163–1174.
[39] C. Borner, I. Filipuzzi, M. Wartmann, U. Eppenberger, D.
Fabbro, Biosynthesis and posttranslational modifications of
protein kinase C in human breast cancer cells, J. Biol. Chem.
264 (1989) 13902–13909.
[40] D. Boehning, C. Moon, S. Sharma, J. Hurt, L.D. Hester, G.V.
Ronnett, D. Shugar, S.H. Snyder, Carbon monoxide neurotrans-
mission activated by CK2 phosphorylation of Heme Oxygenase-2,
Neuron 40 (2003) 129–137.
[41] M. Holst, E. Egyhazi, Differential kinase systems are involved in
the rapidly turning over phosphorylation of prominent nuclear
proteins, Biochim. Biophys. Acta 931 (1987) 224–233.
[42] M.W. Salter, L.V. Kalia, Src kinases: a hub for NMDA receptor
regulation, Nat. Rev. Neurosci. 5 (2004) 317–328.
[43] W.Y. Lu, Z.G. Xiong, S. Lei, B.A. Orser, E. Dudek, M.D.
Browning, J.F. MacDonald, G-protein-coupled receptors act via
protein kinase C and Src to regulate NMDA receptors, Nat.
Neurosci. 2 (1999) 331–338.
[44] T. Tezuka, H. Umemori, T. Akiyama, S. Nakanishi, T. Yamam-
oto, PSD-95 promotes fyn-mediated tyrosine phosphorylation of
the N-methyl-DD-aspartate receptor subunit NR2A, Proc. Natl.
Acad. Sci. USA 96 (1999) 435–440.
[45] Y. Yamada, Y. Chochi, K. Takamiya, K. Sobue, M. Inui, Modu-
lation of the channel activity of the e2/f1-subtype N-methyl-DD-
aspartate receptor by PSD-95, J. Biol. Chem. 274 (1999) 6647–6652.
[46] Y. Yamada, T. Iwamoto, Y. Watanabe, K. Sobue, M. Inui, PSD-
95 eliminates src-induced potentiation of NR1/NR2A-subtype
NMDA receptor channels and reduces high-affinity zinc inhibi-
tion, J. Neurochem. 81 (2002) 758–764.
[47] T. Iwamoto, Y. Yamada, K. Hori, Y. Watanabe, K. Sobue, M.
Inui, Differential modulation of NR1-NR2A and NR1-NR2B
subtypes of NMDA receptor by PDZ domain containing proteins,
J. Neurochem. 89 (2004) 100–108.
[48] K.A. Pelkey, R. Askalan, S. Paul, L.V. Kalia, T.H. Nguyen,
G.M. Pitcher, M.W. Salter, P.J. Lombroso, Tyrosine phosphatase
STEP is a tonic brake on induction of long-term potentiation,
Neuron 34 (2002) 127–138.
[49] A. Jouvenceau, J.M. Billard, U. Haditsch, I.M. Mansuy, P.
Dutar, Different phosphatase-dependent mechanisms mediate
long-term depression and depotentiation of long-term potentia-
tion in mouse hippocampal CA1 area, Eur. J. Neurosci. 18 (2003)
1279–1285.
[50] B.K. Rycroft, A.J. Gibb, Regulation of single NMDA receptor
channel activity by a-actinin and calmodulin in rat hippocampal
granule cells, J. Physiol. 557 (Part 3) (2004) 795–808.
Related Documents











