Protein Function and Evolution

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Protein Function and Evolution

Role of Globins in O2 Transport and Storage
Red Blood Cells (Erythrocytes)
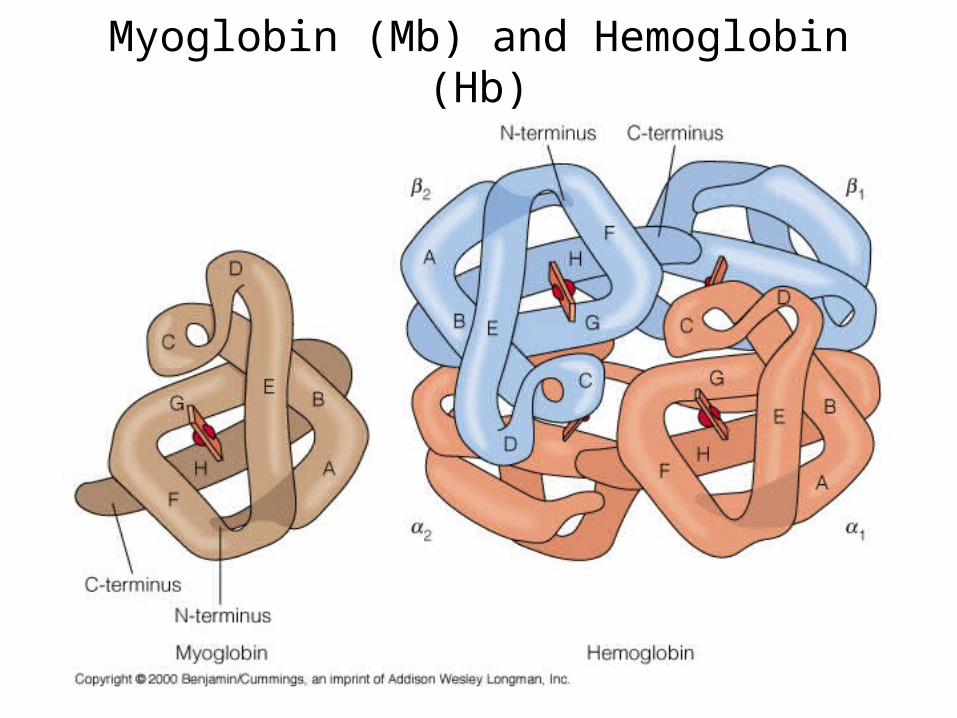
Myoglobin (Mb) and Hemoglobin (Hb)

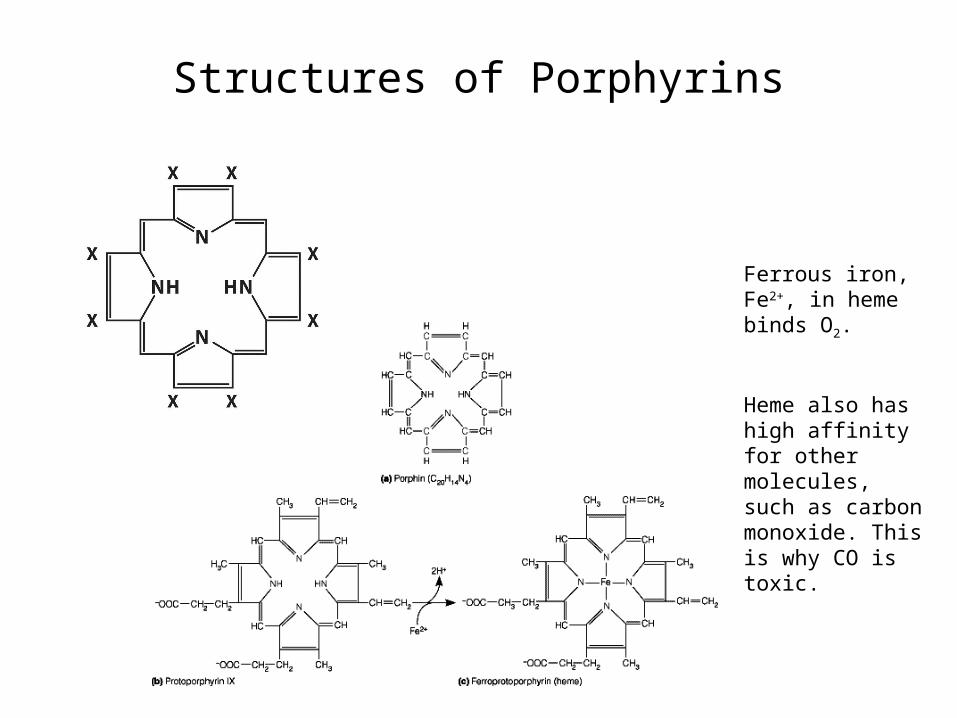
Structures of Porphyrins
Ferrous iron, Fe2+, in heme binds O2.
Heme also has high affinity for other molecules, such as carbon monoxide. This is why CO is toxic.

The Heme Prosthetic Group

Geometry of Iron in Oxyhemoglobin

Iron in Oxyhemoglobin
Heme bound to protein as prosthetic group (tightly bound co-factor) protects heme iron from oxidation (from Fe2+ to Fe3+ oxidation state), lowers heme’s extremely high affinity for CO, and allows for regulation of O2 binding affinity in hemoglobin.
His93 (F8) = proximal HisHis64 (E7) = distal His

Binding of Oxygen and Carbon Monoxide to Heme Iron


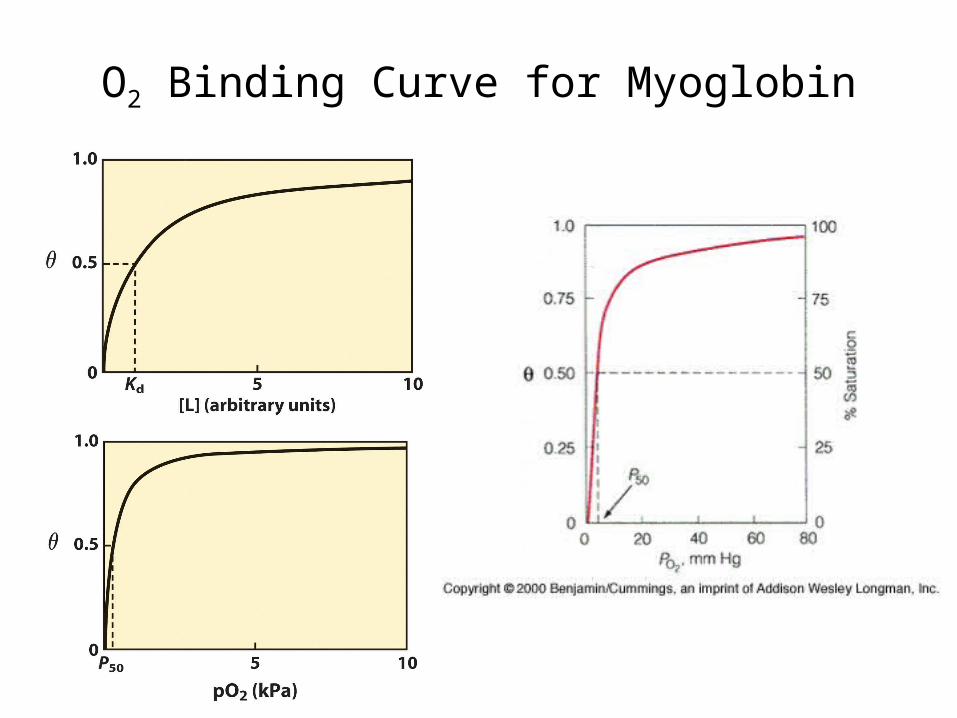
O2 Binding Curve for Myoglobin

Equations for Myoglobin Binding O2
Mb + O2 <-> MbO2
K (association constant) = [MbO2]/([Mb][O2])
(theta; fractional occupancy) = sites occupied/total available sites
= [MbO2]/([Mb]+[MbO2]) = K[Mb][O2]/([Mb]+ K[Mb][O2])
= K[O2]/(1+ K[O2]) = [O2]/(1/K + [O2])
1/K = Kd (dissociation constant) = P50 (O2 partial pressure for half-maximal saturation), so
= PO2/(P50 + PO2)

Dynamics of 02 Release by Myoglobin
Kd = 1/K = ([Mb][O2])/[MbO2] = koff/kon
von (on rate) = kon[O2][Mb]
voff (off rate) = koff[MbO2]

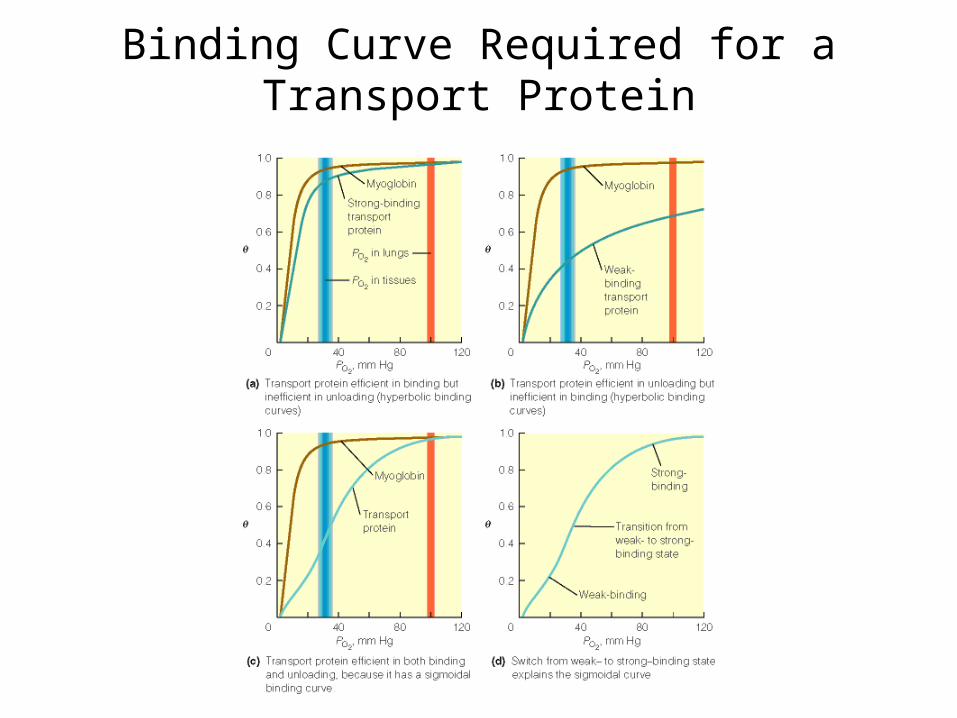
Binding Curve Required for a Transport Protein

Cooperative Binding and Allostery
• T (“tense”) conformational state: low-affinity ligand-binding state of protein.
• R (“relaxed”) conformational state: high-affinity binding binding state of protein.
• Homotropic allosteric interaction: effector and ligand regulated by the effector are the same molecule (e.g., O2 binding affects subsequent O2 binding).
• Heterotropic allosteric interaction: effector and ligand are different molecules (e.g., H+ or BPG binding affects O2 binding).
• Positive allosteric interaction: effector binding increases affinity for ligand.
• Negative allosteric interaction: effector binding decreases affinity for ligand.
allostery = “other site”

Evaluating CooperativityFractional O2 occupancy:
= PO2n/(P50
n + PO2n)
Rearrange and take logarithms for Hill plot:
log(/(1-)) = nlogP02 - nlogP50(assumes n = # of O2 binding sites & all O2 bind simultaneously)
In fact, experimentally determined “n” value is Hill coefficient (nH).nH n except for hypothetical, wholly cooperative process where all ligand molecules would bind simultaneously.nH < n for all real systems.
Hill coefficient very useful in describing cooperativity, since:nH = 1: non-cooperative processnH > 1: positive cooperativitynH < 1: negative cooperativity
Archibald Hill (1910)

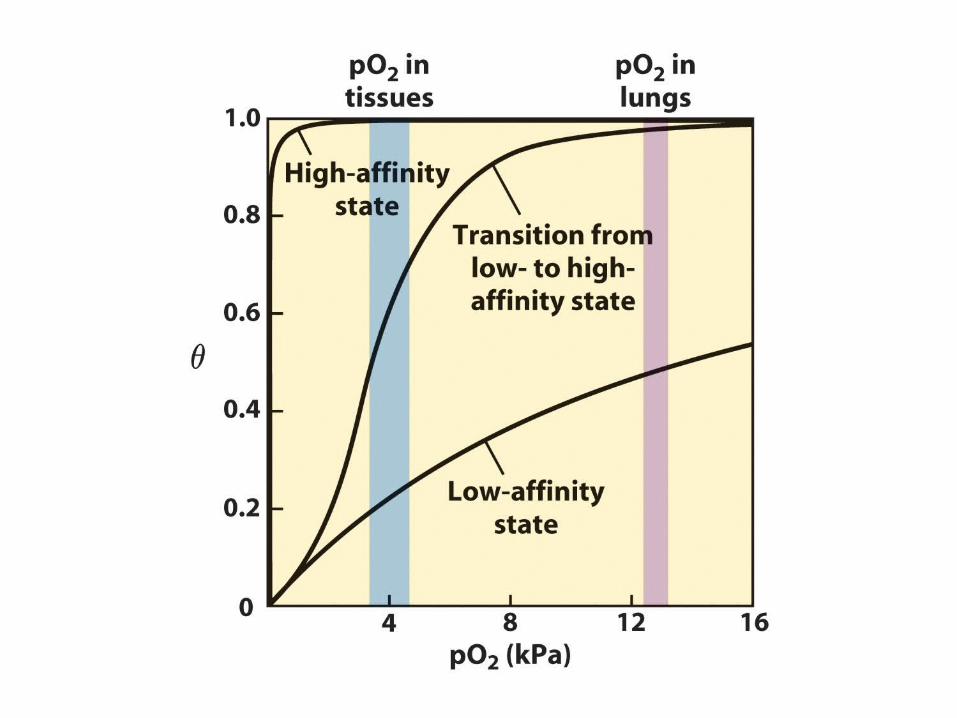
Hill Plots for O2 Binding for Mb and Hb
For Mb:nH = 1This indicates non-cooperative process.
For Hb:nH (max slope) = 3.0-3.5This indicates positive cooperativity (since nH > 1) with at least 4 binding sites for O2 (since n always > nH).
•Intercepts with broken black line at the 0 value for log(/(1-)) indicate P50 and so O2 binding affinity (lower P50 = higher affinity)•Hb high-affinity 02-binding logP50 = upper asymptote intercept•Hb low-affinity 02-binding logP50 = lower asymptote intercept

Scatchard PlotsBound ligand/free ligand vs. bound ligand
X-axis intercept indicates maximum amount of ligand bound (Bmax) or total number of ligand binding sites (n), e.g., 1 for Mb, 4 for Hb.
Slope = -K = -1/Kd (or -1/P50)
Shape of curve gives indication of whether there is cooperativity (positive or negative) or not.
Note: Scatchard plots and other data plotting methods in biochemistry are used a great deal for visual/graphical representation even today. Biochemical parameters used to be determined by manual plotting but now computers are used, since regression analysis on a computer is much more accurate for determining n, Kd, etc.

Subunit Interactions in deoxyHb(T State)
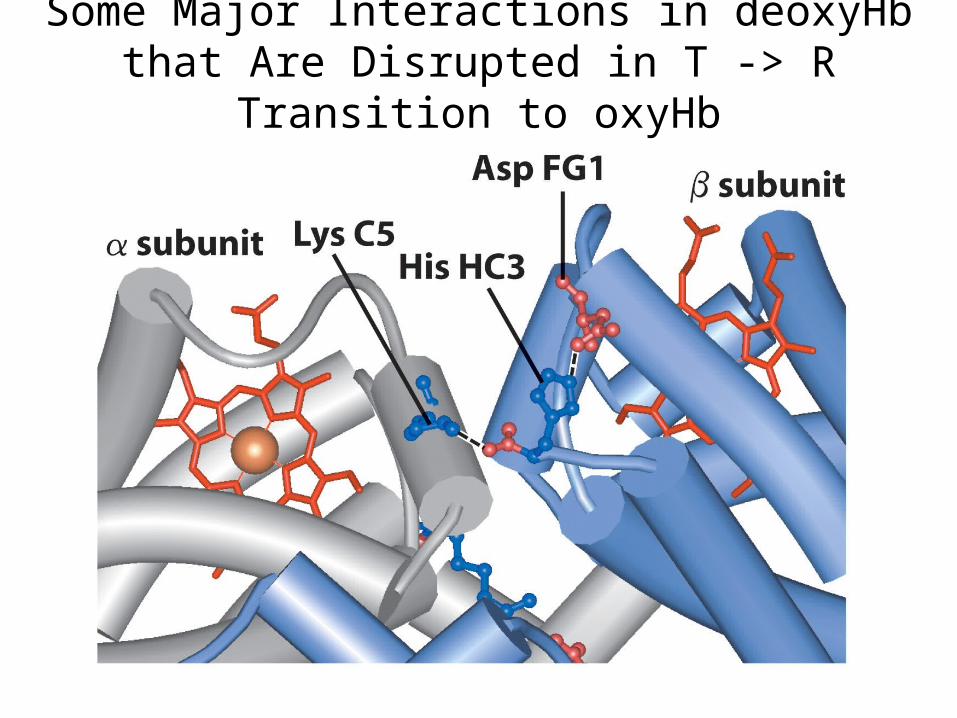
Some Major Interactions in deoxyHb that Are Disrupted in T -> R Transition to
oxyHb

Some Major Interactions in deoxyHb that Are Disrupted in T -> R Transition to
oxyHb

Changes at - and - Interface in T -> R Transition in Hb

Change in Hb 4o Structure with O2 Binding
Some major changes:
•Rotation of 11 relative to 22.•Change in size of central cavity.•Shift of C-termini and FG corners of chains relative to C helices of chains •C-termini of chains interact with C helices of chains in T (deoxy) state. These interactions are broken in transition to R (oxy) state.


Mechanism of T -> R Transition: Iron Pulled into Heme Plane when O2
Binds
His F8 (proximal His) also dragged along in T -> R transition, pulling F helix and shifting subunits relative to one another, increasing O2 affinity of binding sites on other subunits.
Perutz Model (1970)

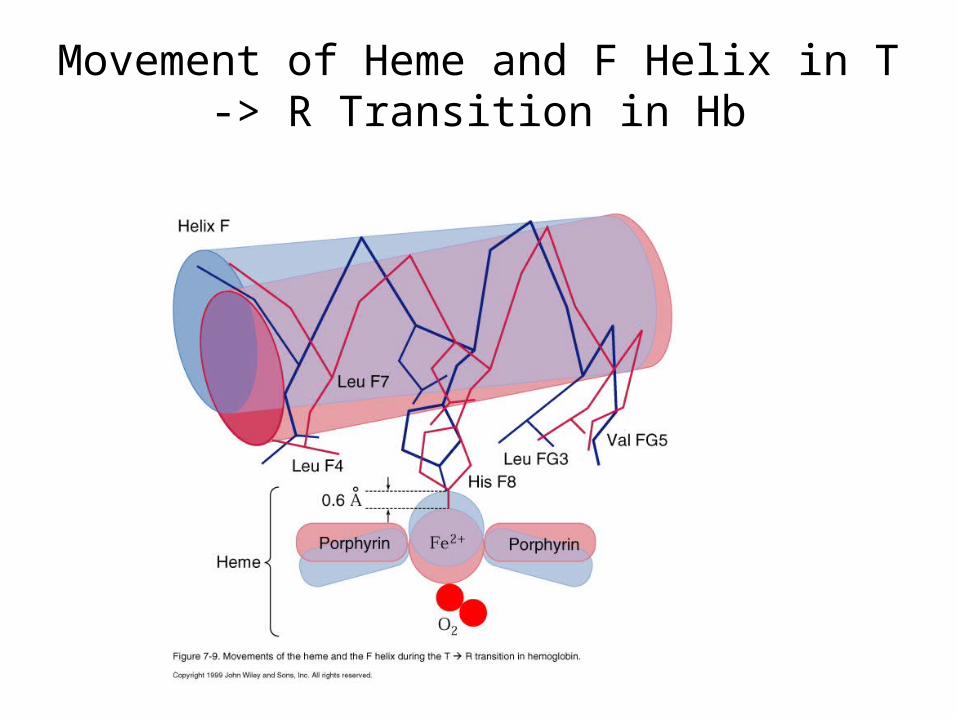
Movement of Heme and F Helix in T -> R Transition in Hb

Effect of Replacing Proximal His in Hb with Gly and Adding Imidazole
Replacement:No cooperativity
Normal Hb: Cooperativity

Negative Allosteric Effectors of O2 Binding in Hb: Stabilizers of T State of
Hb• H+ (“The Bohr effect,” Christian Bohr, 1904)• 2,3-bisphosphoglycerate (BPG)• Carbon dioxide (transported in blood as bicarbonate and
carbamates): – Bicarbonate formation: CO2 + H20 <-> HCO3
- + H+
– Carbamate formation:Hb-NH3
+ + HCO3- <=> Hb-NH-COO- + H+ + H2O
– CO2 lowers O2 binding affinity through H+ released (contributing to Bohr effect) and formation of carbamate at N-termini of Hb subunits, stabilizing T state interactions between and chains.
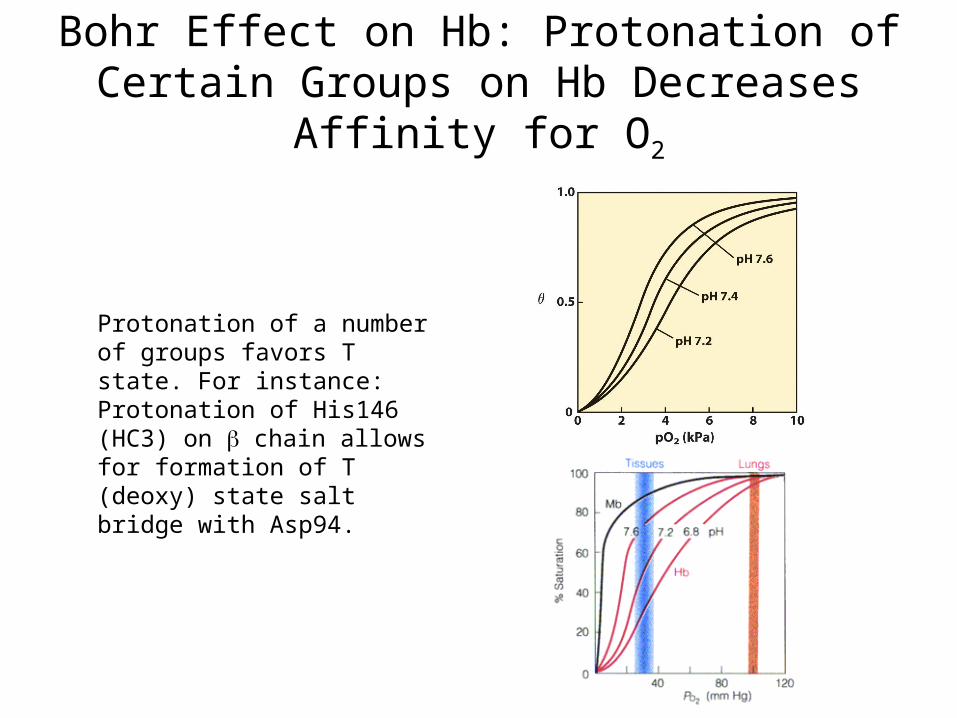
Bohr Effect on Hb: Protonation of Certain Groups on Hb Decreases Affinity for O2
Protonation of a number of groups favors T state. For instance:Protonation of His146 (HC3) on chain allows for formation of T (deoxy) state salt bridge with Asp94.
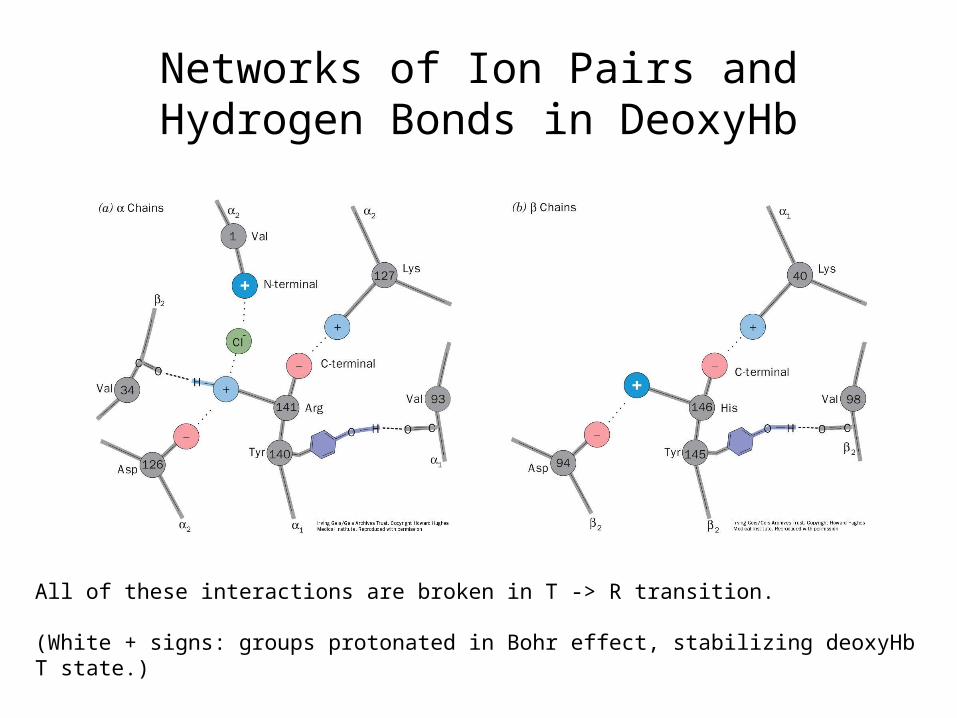
Networks of Ion Pairs and Hydrogen Bonds in DeoxyHb
All of these interactions are broken in T -> R transition.
(White + signs: groups protonated in Bohr effect, stabilizing deoxyHbT state.)

2,3-Bisphosphoglycerate (BPG)
In mammals
In birds

Binding of BPG to DeoxyHb: Stabilization of T State of Hb

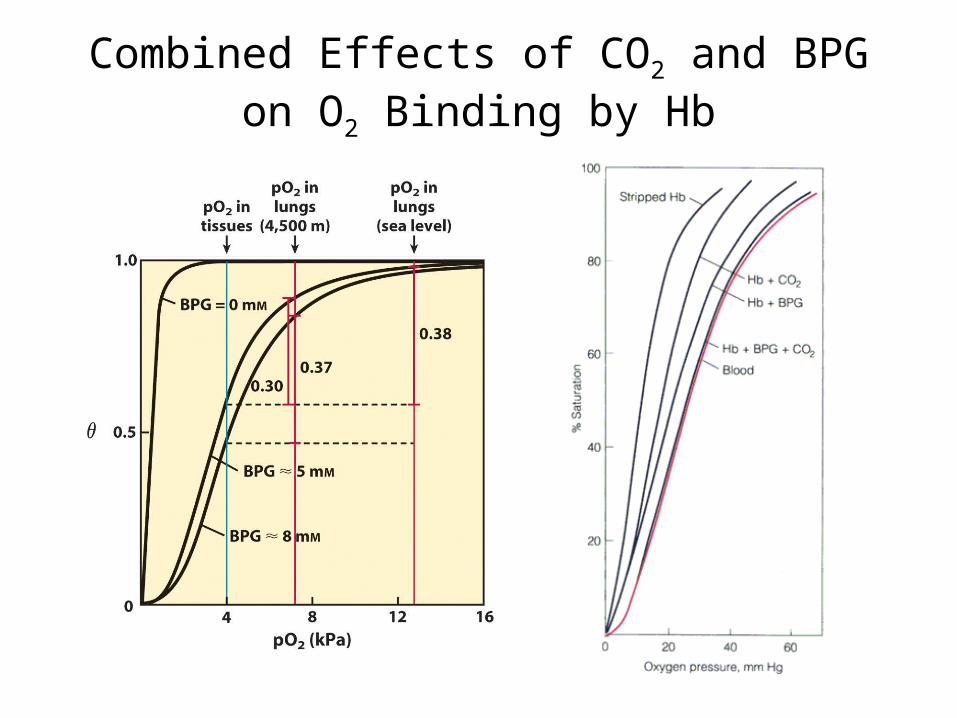
Combined Effects of CO2 and BPG on O2 Binding by Hb

Role of Globins in O2 Transport and Storage
CO2, H+ and BPG decrease Hb’s affinity for O2 and so favor release of O2 in tissues.
OxyHb carries O2 in arteries.
CO2 carried in veins as bicarbonate. Also, deoxyHb carries CO2 as carbamates.
[CO2] (and [H+]) high in tissues as a result of respiration.
Release of CO2 in lungs (or gills in fish). Oxygenation of Hb in lungs.

Two Models of Allostery
Koshland, Nemethy, Filmer (KNF) Model (1966): Sequential or Induced Fit Model•Ligand binding at one site causes protein conformational change (induced fit), shifting binding affinity in adjacent subunits only, so complete T -> R transition is a sequential process.•Can account for both positive and negative cooperativity.
Monod, Wyman, Changeux (MWC) Model (1965): Concerted or Symmetry Model•Equilibrium between T and R states.•Transition is a concerted process, affecting all subunits simultaneously in the same way.•In absence of ligand, equilibrium favors T state.•Ligand binding shifts equilibrium toward R state.•Only models positive cooperativity.
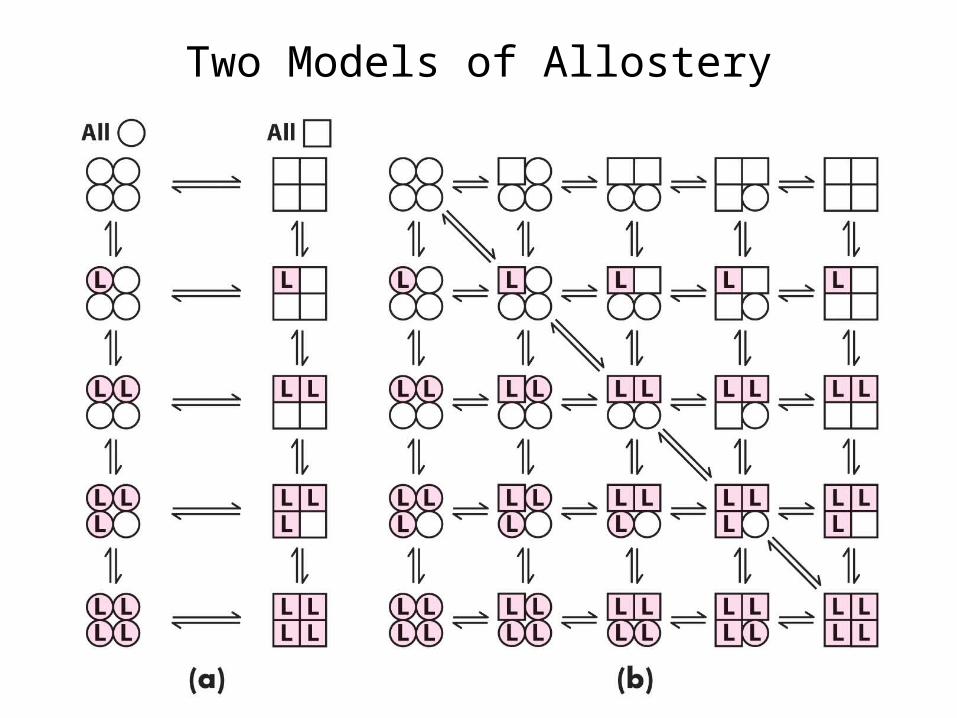
Two Models of Allostery

Two Models of Allostery
Koshland, Nemethy, Filmer (KNF) Model (1966): Sequential or Induced Fit Model
Monod, Wyman, Changeux (MWC) Model (1965): Concerted or Symmetry Model

Recent Model for Cooperative Transition of Hb
If both and each contain at least one O2 bound, T -> R transition occurs.

Protein Evolution and Diversity
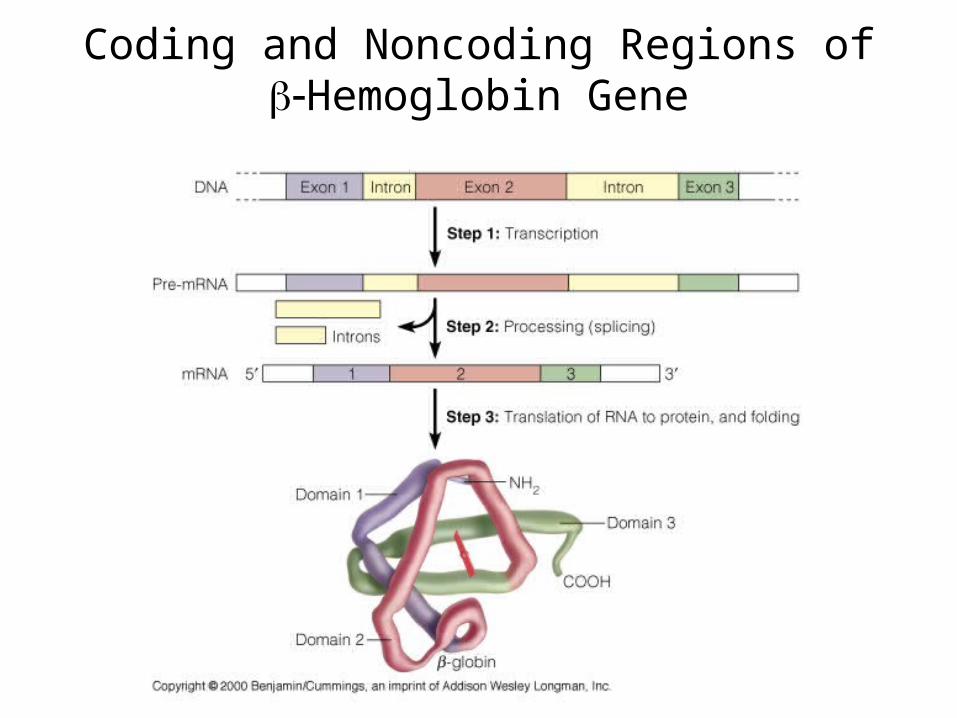
Coding and Noncoding Regions of Hemoglobin Gene

Some Mutagenic Agents

Types of Mutations

Comparison of Sequences of Mb and the and Chains of Hb

Evolutionary Conservation of the Globin Folding Pattern

Evolution of the Globin Genes

Expression of Human Globin Genes at Different Stages of Development
Fetal Hb (22) has low affinity for BPG, which facilitates transfer of O2 to fetus, since in the presence of BPG, fetal 22 Hb has higher affinity for O2 than does adult 22 Hb.

Some Missense Mutations in Human Hemoglobins
In addition to missense mutations in human hemoglobins, there are other hemoglobin diseases called thalassemias in which or chains are not produced at all or produced in insufficient quantities.
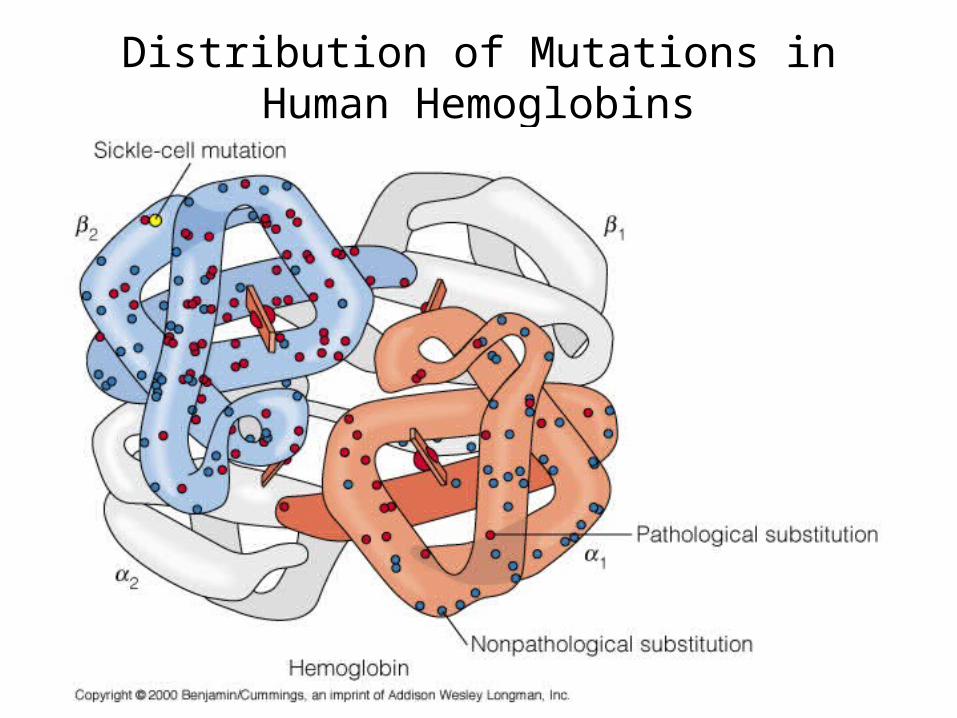
Distribution of Mutations in Human Hemoglobins
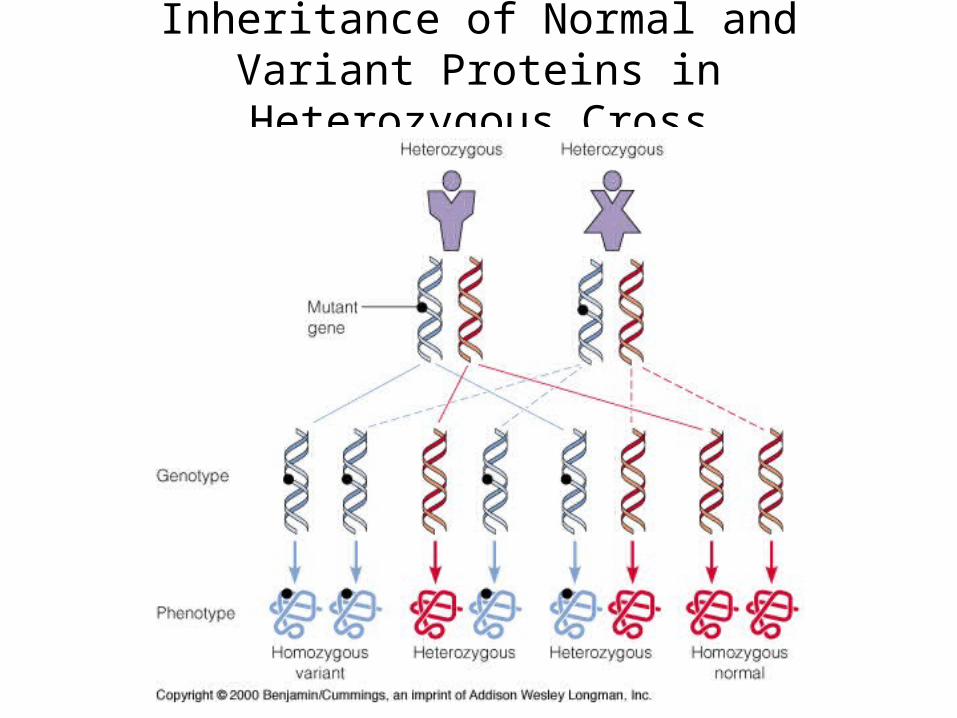
Inheritance of Normal and Variant Proteins in Heterozygous Cross

Sickle-Cell Anemia
Red blood cells become abnormally elongated and sickle-shaped. Sickled cells block capillaries and die prematurely.
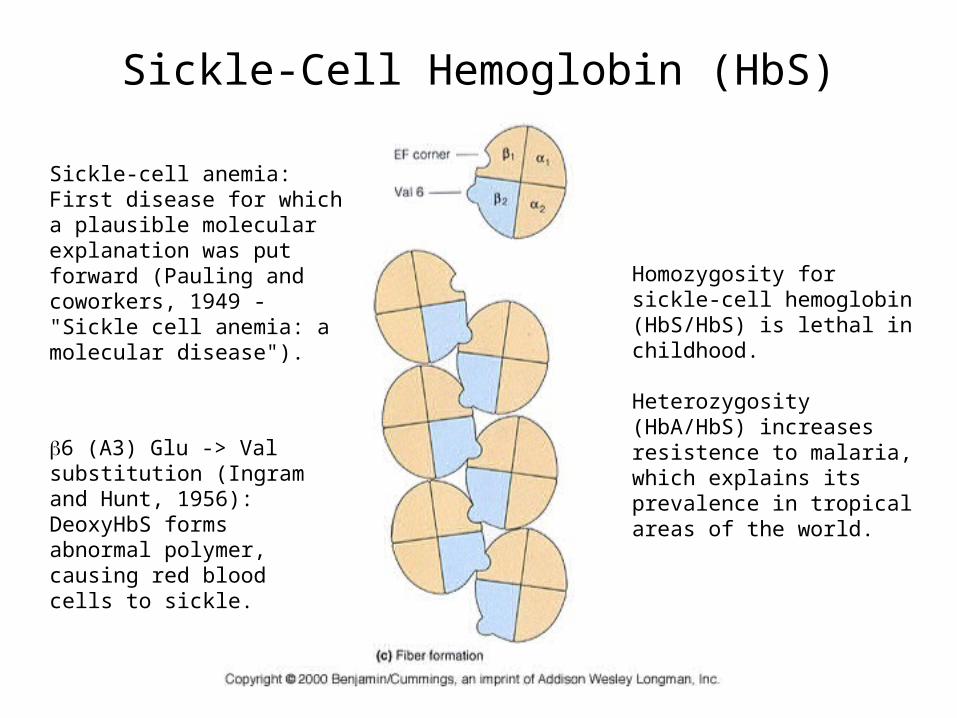
Sickle-Cell Hemoglobin (HbS)
Sickle-cell anemia:First disease for which a plausible molecular explanation was put forward (Pauling and coworkers, 1949 - "Sickle cell anemia: a molecular disease").
Homozygosity for sickle-cell hemoglobin (HbS/HbS) is lethal in childhood.
Heterozygosity (HbA/HbS) increases resistence to malaria, which explains its prevalence in tropical areas of the world.
6 (A3) Glu -> Val substitution (Ingram and Hunt, 1956):DeoxyHbS forms abnormal polymer, causing red blood cells to sickle.
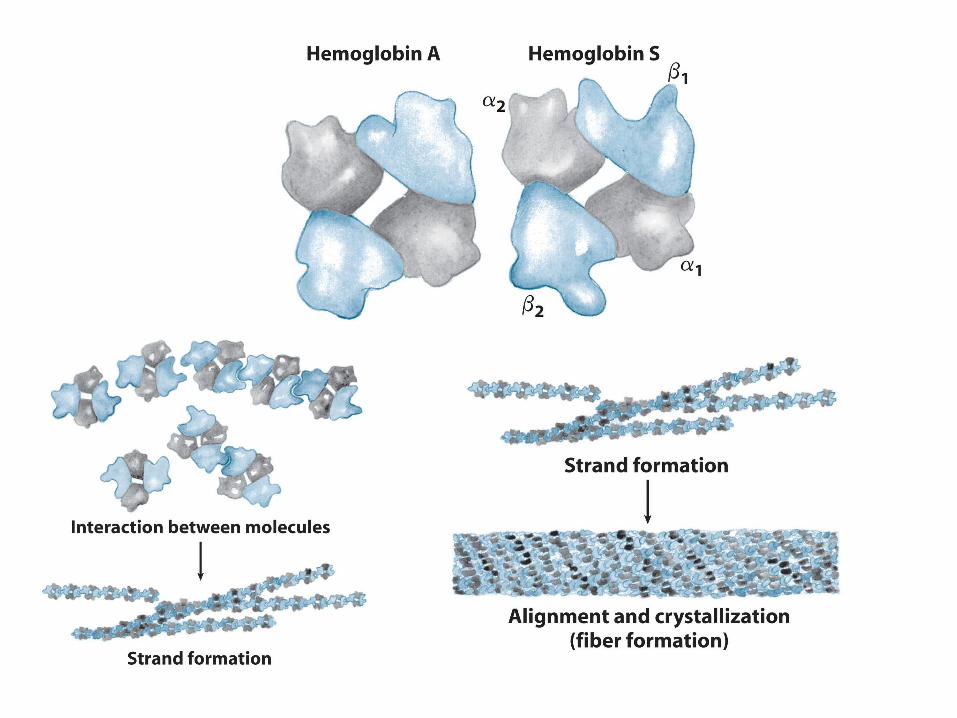

Structure of Sickle-Cell Hemoglobin (HbS) FIbers

Immunoglobulins (Antibodies):Diversity in Structure and Binding
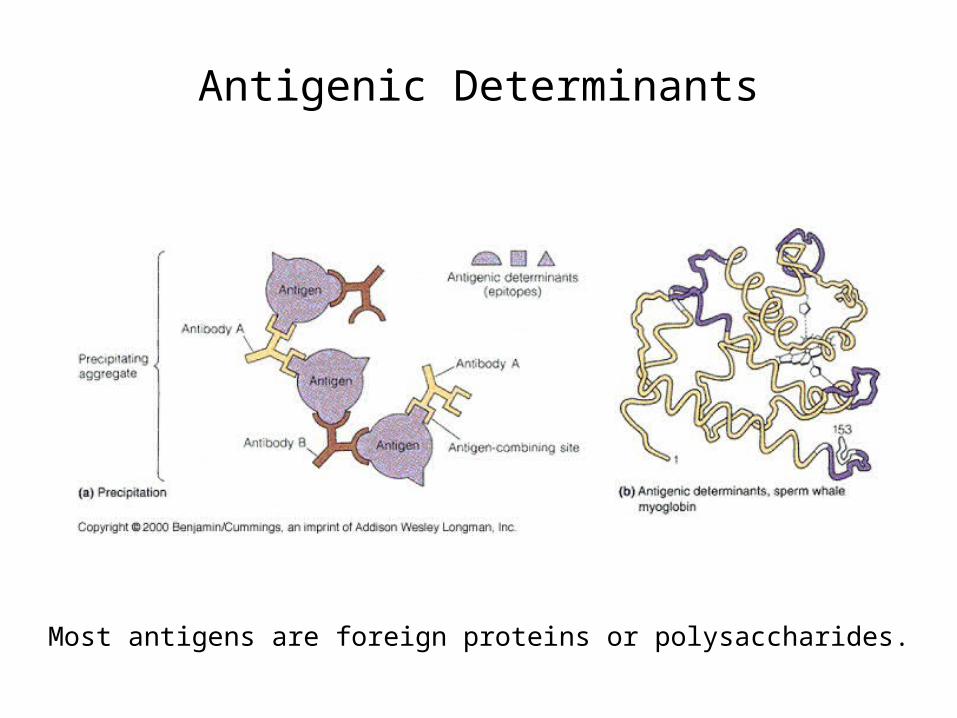
Antigenic Determinants
Most antigens are foreign proteins or polysaccharides.

Interactions of Antigen with Antibody
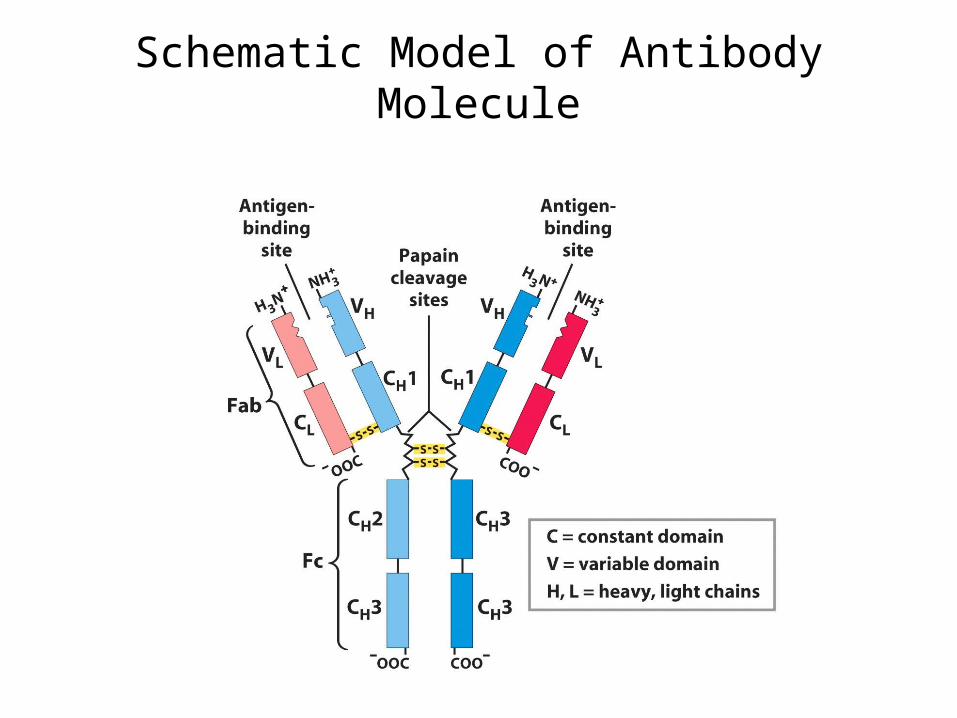
Schematic Model of Antibody Molecule
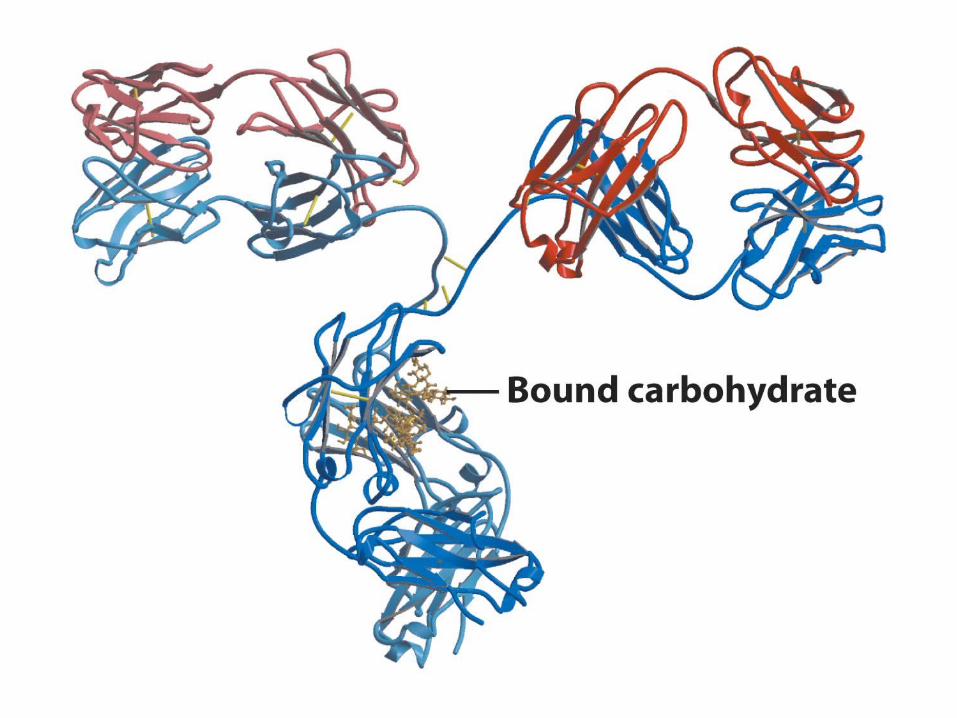


Model of X-Ray Structure of IgG
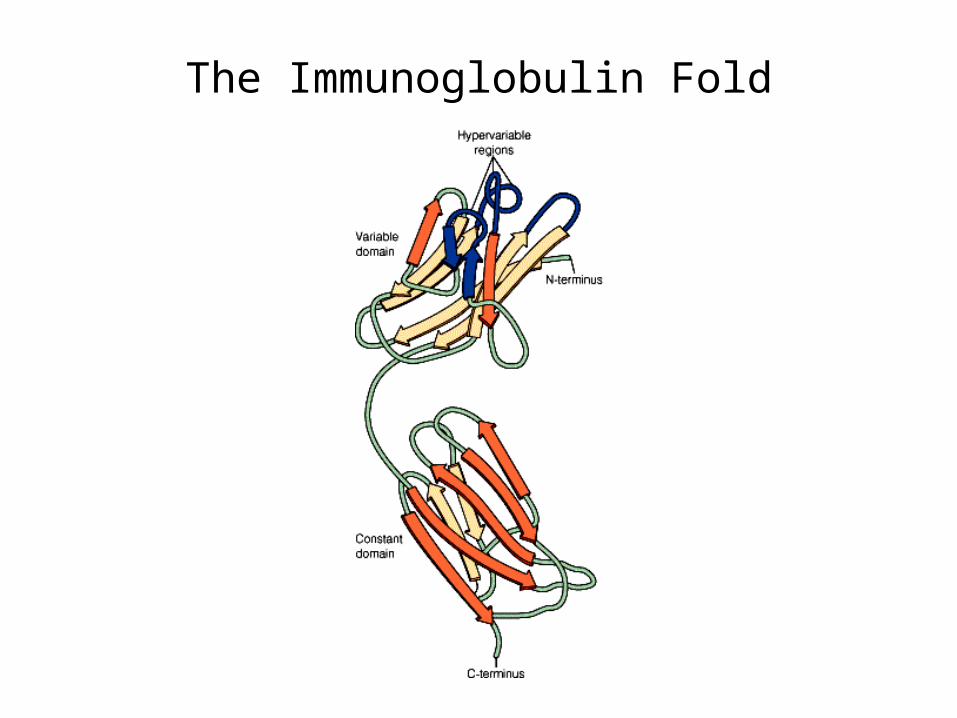
The Immunoglobulin Fold


Generation of Antibody DiversityVDJ Recombination

Generation of Antibody DiversitySomatic hypermutation (point mutations)
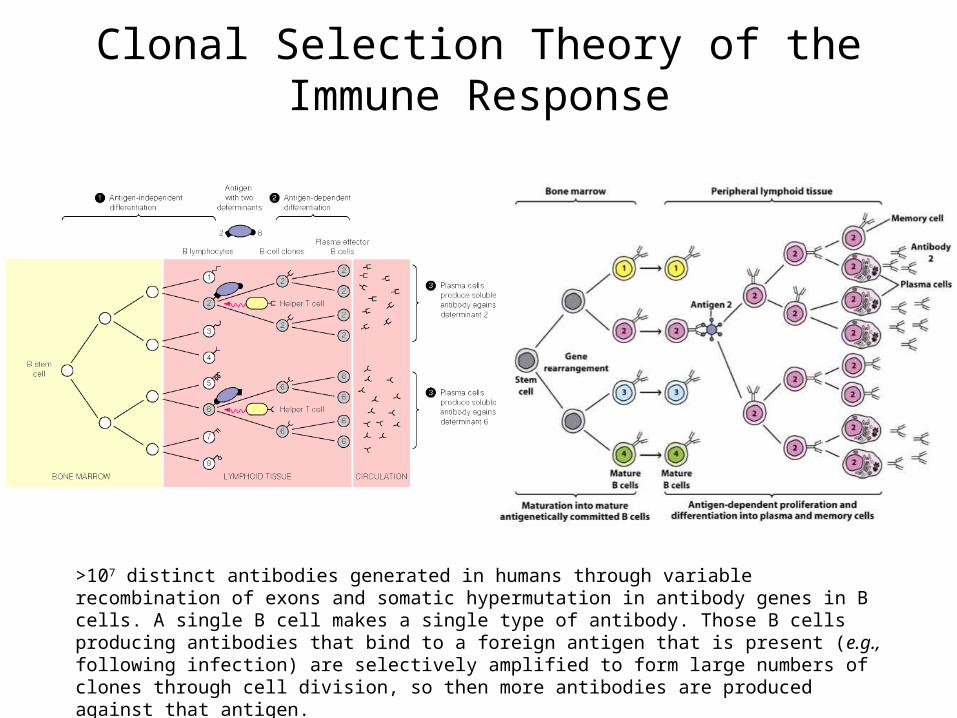
Clonal Selection Theory of the Immune Response
>107 distinct antibodies generated in humans through variable recombination of exons and somatic hypermutation in antibody genes in B cells. A single B cell makes a single type of antibody. Those B cells producing antibodies that bind to a foreign antigen that is present (e.g., following infection) are selectively amplified to form large numbers of clones through cell division, so then more antibodies are produced against that antigen.

Two Developmental Pathways for Stimulated B Lymphocytes
Antibody attached to B cell membrane = B-cell receptor
Soluble antibodies released from effector B cells (plasma cells).
Most abundant circulating antibodies: immunglobulin G (IgG).

Humoral and Cellular Immune Responses
Humoral: secreted antibodies (mainly IgG)
Cellular: B-cell receptor on B cells and T-cell receptor on killer T cells.


Human Immunodeficiency Virus
HIV binds to a specific cell-surface protein (CD4) on helper T cells, enters these cells and kills them, leading to immunodeficiency.

Technical Applications of Antibodies

Preparation of Polyclonal Antibodies
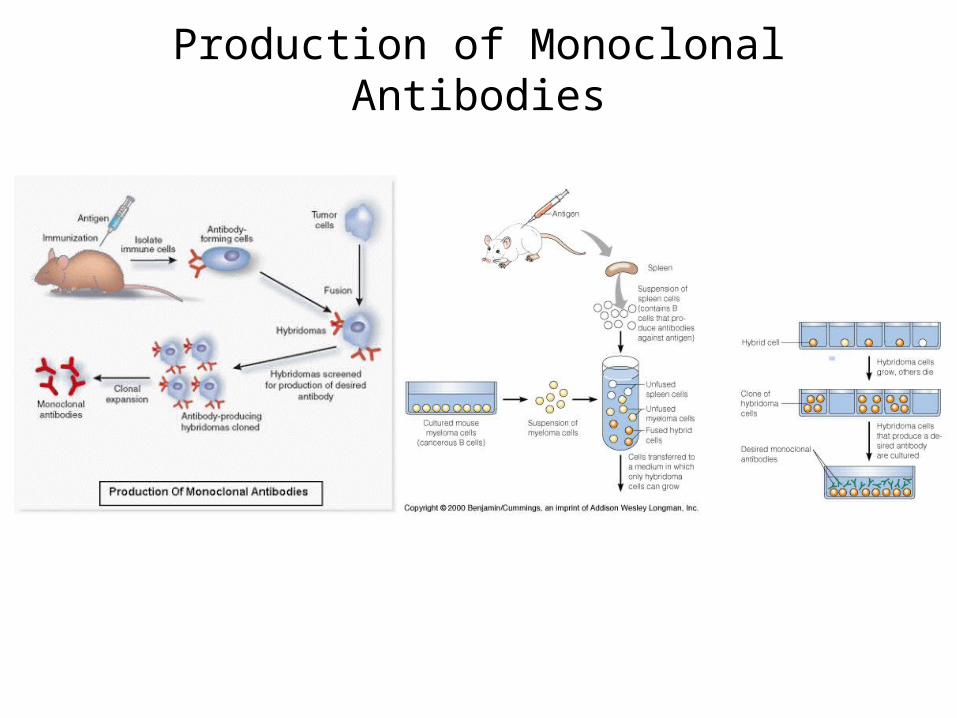
Production of Monoclonal Antibodies

Enzyme-Linked Immunosorbent Assay (ELISA)
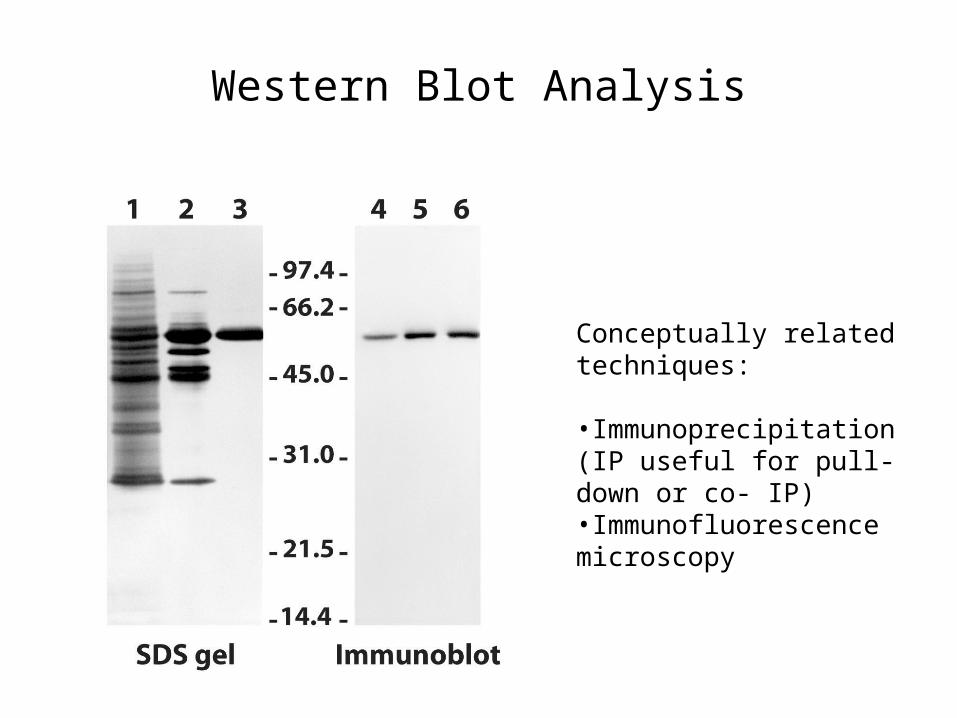
Western Blot Analysis
Conceptually related techniques:
•Immunoprecipitation (IP useful for pull-down or co- IP)•Immunofluorescence microscopy
Related Documents











