Abstract Hippocampal 5-HT 1A receptors have been shown to be suppressed by glucocorticoids in a variety of animal studies, however the molecular mechanism and the functional meaning of this effect are still not well un- derstood. The present study was designed to investigate the im- pact of repeated administration of corticosterone (10 mg/kg s.c. twice daily for 7 days) on the functional conse- quences of 5-HT 1A receptor stimulation measured electro- physiologically in hippocampal slices. Additionally, the effects of corticosterone on 5-HT 1A receptor binding and on receptor mRNA levels in the hippocampus were stud- ied. Prolonged, but not acute treatment with cortico- sterone attenuated (±)-8-hydroxy-2-di-N-propylamino)tetra- lin hydrobromide (8-OH-DPAT)-induced inhibition of pop- ulation spikes, and 8-OH-DPAT-induced hyperpolariza- tion in rat CA1 hippocampal neurons. Chronic, but not acute treatment with corticosterone also decreased 5-HT 1A receptor binding in the CA1 region (in the ventral part only) and the dentate gyrus. A single dose of cortico- sterone increased [ 3 H]8-OHDPAT binding in the dentate gyrus and in the CA3 and CA4 hippocampal regions. Only acute, but not prolonged treatment with cortico- sterone decreased the level of 5-HT 1A receptor mRNA in the CA1 region and dentate gyrus of the hippocampus. 5-HT turnover in the hippocampus was not influenced by chronic corticosterone. It is concluded that a chronically elevated level of cor- ticosterone can induce functional desensitization of 5-HT 1A receptors in the CA1 area of the hippocampus, although this effect is not always followed consequently by de- creases in 5-HT 1A receptor synthesis in this or other areas of the hippocampus. Keywords Corticosterone · Electrophysiology · Hippocampus · 5-HT 1A receptor · Hyperpolarization · In situ hybridization · 8-OH-DPAT binding · Population spike Introduction Glucocorticoids, the final products of hypothalamo-pitu- itary-adrenal (HPA) axis activation, affect mood and be- haviour (McEwen 1987; Wolkowitz et al. 1997; Lopez et al. 1998). Although the effects induced by glucocorticoids are critical mainly with respect to the ability of individu- als to adapt metabolically and behaviourally to stressful events (de Kloet et al. 1990), it has also been hypothe- sized that sustained alterations in the HPA axis activity that induce elevated glucocorticoid levels may underlie the pathophysiology of psychiatric disorders such as de- pression, precipitation of psychoses and increased vulner- ability to addictive substances (Modell et al. 1997, 1998; Murphy 1997; Piazza and Le Moal 1997; Wolkowitz et al. 1997; Lopez et al. 1998). Corticosterone, a main adrenocorticosteroid hormone in the rat, readily enters the central nervous system due to its lipophilic structure. There it binds to two types of in- tracellular receptors: the high-affinity mineralocorticoid receptors (MR) and the lower-affinity glucocorticoid re- ceptors (GR) (de Kloet et al. 1990). This difference in affinity implies that low levels of corticosterone will oc- cupy primarily MR, whereas GR will be activated mainly after stress, application of high doses of corticosterone or at the circadian peak (Reul et al. 1987; de Kloet et al. 1990; Spencer et al. 1990). In the rat hippocampus both MR and GR receptors are present. MR appear to be highly and evenly distributed in all hippocampal CA1–4 cell fields and the dentate gyrus (Hansson et al. 2000), whereas GR Anna Czyrak · Marzena Maćkowiak · Agnieszka Chocyk · Katarzyna Fijał · Krzysztof Tokarski · Maria Bijak · Krzysztof Wędzony Prolonged corticosterone treatment alters the responsiveness of 5-HT 1A receptors to 8-OH-DPAT in rat CA1 hippocampal neurons Naunyn-Schmiedeberg’s Arch Pharmacol (2002) 366 : 357–367 DOI 10.1007/s00210-002-0586-2 Received: 27 February 2002 / Accepted: 29 April 2002 / Published online: 16 July 2002 ORIGINAL ARTICLE A. Czyrak (✉) · M. Maćkowiak · A. Chocyk · K. Fijał · K. Wędzony Laboratory of Pharmacology and Brain Biostructure, Department of Pharmacology, Institute of Pharmacology, Polish Academy of Sciences, Smętna Street 12, 31-343 Kraków, Poland e-mail: [email protected], Tel.: +48-12-6623253, Fax: +48-12-6374500 K. Tokarski · M. Bijak Department of Physiology, Institute of Pharmacology, Polish Academy of Sciences, Smętna Street 12, 31-343 Kraków, Poland © Springer-Verlag 2002

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Abstract Hippocampal 5-HT1A receptors have beenshown to be suppressed by glucocorticoids in a variety ofanimal studies, however the molecular mechanism andthe functional meaning of this effect are still not well un-derstood.
The present study was designed to investigate the im-pact of repeated administration of corticosterone (10mg/kg s.c. twice daily for 7 days) on the functional conse-quences of 5-HT1A receptor stimulation measured electro-physiologically in hippocampal slices. Additionally, theeffects of corticosterone on 5-HT1A receptor binding andon receptor mRNA levels in the hippocampus were stud-ied. Prolonged, but not acute treatment with cortico-sterone attenuated (±)-8-hydroxy-2-di-N-propylamino)tetra-lin hydrobromide (8-OH-DPAT)-induced inhibition of pop-ulation spikes, and 8-OH-DPAT-induced hyperpolariza-tion in rat CA1 hippocampal neurons. Chronic, but notacute treatment with corticosterone also decreased 5-HT1Areceptor binding in the CA1 region (in the ventral partonly) and the dentate gyrus. A single dose of cortico-sterone increased [3H]8-OHDPAT binding in the dentategyrus and in the CA3 and CA4 hippocampal regions.Only acute, but not prolonged treatment with cortico-sterone decreased the level of 5-HT1A receptor mRNA inthe CA1 region and dentate gyrus of the hippocampus. 5-HT turnover in the hippocampus was not influenced bychronic corticosterone.
It is concluded that a chronically elevated level of cor-ticosterone can induce functional desensitization of 5-HT1A
receptors in the CA1 area of the hippocampus, althoughthis effect is not always followed consequently by de-creases in 5-HT1A receptor synthesis in this or other areasof the hippocampus.
Keywords Corticosterone · Electrophysiology · Hippocampus · 5-HT1A receptor · Hyperpolarization · In situ hybridization · 8-OH-DPAT binding · Populationspike
Introduction
Glucocorticoids, the final products of hypothalamo-pitu-itary-adrenal (HPA) axis activation, affect mood and be-haviour (McEwen 1987; Wolkowitz et al. 1997; Lopez etal. 1998). Although the effects induced by glucocorticoidsare critical mainly with respect to the ability of individu-als to adapt metabolically and behaviourally to stressfulevents (de Kloet et al. 1990), it has also been hypothe-sized that sustained alterations in the HPA axis activitythat induce elevated glucocorticoid levels may underliethe pathophysiology of psychiatric disorders such as de-pression, precipitation of psychoses and increased vulner-ability to addictive substances (Modell et al. 1997, 1998;Murphy 1997; Piazza and Le Moal 1997; Wolkowitz et al.1997; Lopez et al. 1998).
Corticosterone, a main adrenocorticosteroid hormonein the rat, readily enters the central nervous system due toits lipophilic structure. There it binds to two types of in-tracellular receptors: the high-affinity mineralocorticoidreceptors (MR) and the lower-affinity glucocorticoid re-ceptors (GR) (de Kloet et al. 1990). This difference inaffinity implies that low levels of corticosterone will oc-cupy primarily MR, whereas GR will be activated mainlyafter stress, application of high doses of corticosterone orat the circadian peak (Reul et al. 1987; de Kloet et al.1990; Spencer et al. 1990). In the rat hippocampus bothMR and GR receptors are present. MR appear to be highlyand evenly distributed in all hippocampal CA1–4 cell fieldsand the dentate gyrus (Hansson et al. 2000), whereas GR
Anna Czyrak · Marzena Maćkowiak · Agnieszka Chocyk ·Katarzyna Fijał · Krzysztof Tokarski · Maria Bijak ·Krzysztof Wędzony
Prolonged corticosterone treatment alters the responsiveness of 5-HT1A receptors to 8-OH-DPAT in rat CA1 hippocampal neurons
Naunyn-Schmiedeberg’s Arch Pharmacol (2002) 366 :357–367DOI 10.1007/s00210-002-0586-2
Received: 27 February 2002 / Accepted: 29 April 2002 / Published online: 16 July 2002
ORIGINAL ARTICLE
A. Czyrak () · M. Maćkowiak · A. Chocyk · K. Fijał ·K. WędzonyLaboratory of Pharmacology and Brain Biostructure, Department of Pharmacology, Institute of Pharmacology, Polish Academy of Sciences, Smętna Street 12, 31-343 Kraków, Polande-mail: [email protected], Tel.: +48-12-6623253, Fax: +48-12-6374500
K. Tokarski · M. BijakDepartment of Physiology, Institute of Pharmacology, Polish Academy of Sciences, Smętna Street 12, 31-343 Kraków, Poland
© Springer-Verlag 2002

show marked density in the pyramidal neurons of CA1and CA2 cell fields and the granular neurons of the den-tate gyrus, and much lower density in the CA3 and CA4cell fields (de Kloet et al. 1990; Kawata et al. 1998; Hans-son et al. 2000). There is strong evidence for co-expres-sion of MR and GR in the majority of pyramidal andgranular neurons of the hippocampal formation (VanEekelen and de Kloet 1992; Hansson et al. 2000).
As already mentioned, adrenal steroids regulate di-verse processes and systems in the brain. In the hip-pocampus, changes in glucocorticoid levels influenceboth the expression and function of 5-HT1A receptors.This subtype of 5-HT receptor is expressed intensively inthe hippocampus and other limbic areas (Chalmers andWatson 1991; Pompeiano et al. 1992) and seems to be im-plicated in the pathogenesis of depression, anxiety andschizophrenia (Lesch 1991; Burnet et al. 1996; Meijer etal. 1997b; Lopez et al. 1998), i.e. pathological processesin which glucocorticoids are also implicated.
The 5-HT1A receptor belongs to the family of G pro-tein-coupled receptors, and in the hippocampus it islinked to an inwardly rectifying K+ channel (Andrade etal. 1986; Muma and Beck 1999). Activation of the 5-HT1Areceptor induces membrane hyperpolarization (Beck et al.1992, 1996; Bijak et al. 2001). In recent years, many bio-chemical and electrophysiological studies have examinedthe effect of adrenalectomy (ADX) on 5-HT1A receptormRNA expression and ligand binding, as well as the func-tional responses to 5-HT1A receptor stimulation in the hip-pocampus. ADX is followed by a rapid increase in thesynthesis of 5-HT1A receptor mRNA (total and de novo),and by an increase in 5-HT1A binding, both effects beingprevented by low concentrations of corticosterone that ac-tivate MR preferentially (Chalmers et al. 1993, 1994;Kuroda et al. 1994; Tejani-Butt and Labow 1994; Zhongand Ciaranello 1995). As shown in electrophysiologicalstudies, ADX enhances responses to 5-HT1A receptorstimulation in CA3, but not CA1 or dentate gyrus neuronsof the hippocampus (Joels and de Kloet 1992, 1994; Becket al. 1997; Karten et al. 2001; Mueller and Beck 2000).All these studies show that the 5-HT1A receptors (at leastin the hippocampus) are under control of glucocorticoids.They also suggest that in psychiatric pathologies withcharacteristic elevation of adrenocorticoid levels, thechanges induced by these hormones in 5-HT1A receptorsmay be one of the factors precipitating clinical symptoms.To examine this hypothesis studies concerning the effectsof chronic hypercorticism on 5-HT1A receptors in intact(not ADX) animals are necessary and such studies arescarce. Two recent studies have shown that in hippocam-pal slices from rats with chronically elevated level of glu-cocorticoids the responsiveness of 5-HT1A receptors ischanged (Karten et al. 1999; Bijak et al. 2001) but not as-sociated with any changes in 5-HT1A receptor mRNA, asshown with the single-cell antisense amplification tech-nique (Karten et al. 1999).
Our study was designed to characterize further themechanism whereby a prolonged increase of cortico-steroid levels alters the function, mRNA expression and
binding capacities of hippocampal 5-HT1A receptors. Inthe present study we administered corticosterone to intactanimals for 7 days (10 m/kg twice daily). This dose yieldedblood levels comparable to those obtained by Karten et al.(1999) in their electrophysiological study showing de-creased responsiveness of 5-HT1A receptors. Similar lev-els are also induced by different stressors that modulate of5-HT1A receptor expression in the hippocampus (Raghu-pathi and McGonigle 1997). Since electrophysiologicalstudies have evaluated the effect of corticosterone on 5-HT1A receptor responsiveness elicited by the endogenousagonist 5-HT (Karten et al. 1999), we decided to confirmand extend those results and to examine the effect of re-peated treatment with corticosterone on the reactivity ofCA1 neurons to (±)-8-hydroxy-2-di-N-propylamino)tetra-lin hydrobromide (8-OH-DPAT), a selective agonist at 5-HT1A receptors. The choice of 8-OH-DPAT facilitatedalso comparison between corticosterone modulation offunctional responses on one hand and of 5HT1A receptorbinding ([3H]8-OH-DPAT was used as the ligand) on theother. The effect of corticosterone on CA1 neuronal sensi-tivity to 8-OH-DPAT was evaluated by measuring the am-plitude of population spikes observed in field potentialrecordings, and by intracellular recording. These two pro-cedures allowed us to assess the effects of corticosteroneon neuronal sensitivity and also to gain insight into thecellular mechanisms underlying observed effects. Tomake comparisons between alterations in the 5-HT1A re-ceptor function and changes in the expression of receptorspotentially responsible for the observed functional alter-ations, the level of 5-HT1A receptor mRNA and the bind-ing of [3H]8-OH-DPAT to those receptors in CA1 subfieldand other areas of hippocampus were measured. Addi-tionally, the effect of corticosterone on the level of 5-HTand its metabolite 5-hydroxyindolacetic acid (5-HIAA)was also assessed.
Materials and methods
Animals and drug treatments
Male Wistar rats (250–300 g) were used in all the experiments.The animals were housed in groups of six under controlled condi-tions (12 h light/dark cycle, 22±2°C). The rats were allowed atleast 1 week to acclimatise to the animal room, and had free accessto a standard laboratory diet and tap water. Corticosterone-21-ac-etate (Sigma, St. Louis, Mo., USA) was prepared as a suspensionin 1% Tween 80 and was administered in a dose of 10 mg/kg s.c.acutely (single dose) in rats given the vehicle for the preceding 7 days, or repeatedly (twice daily for 7 days). The rats were sacri-ficed 2 h (acute treatment), or 2 or 24 h (repeated treatment) afterthe last injection of corticosterone or vehicle. The experimentalprotocols were approved by the Committee for Laboratory AnimalWelfare and Ethics of Institute of Pharmacology Polish Academyof Sciences in Kraków and met the criteria of the European Coun-cil for Laboratory Animal Guide for the care and use of laboratoryanimals (86/609/EEC).
Measurement of serum corticosterone
The animals were decapitated, blood samples collected from thetrunk and serum corticosterone levels therein determined by ra-
358

dioimmunoassay (Przegalinski et al. 1992) using rabbit antiserumraised against corticosterone-21-thyroglobulin (Sigma) as antigen.The antiserum showed a very low cross-reactivity with other majorsteroids. The assay sensitivity was 10 pg/tube.
Electrophysiological studies
Hippocampal slice preparation and recording
Rats were killed by decapitation. The brains were removed quicklyand placed in ice-cold artificial cerebrospinal fluid (ACSF) consis-ting of (in mM): NaCl 124, KCl 2, CaCl2 2.5, MgSO4 1.3, KH2PO41.25, NaHCO3 26 and glucose 10. After dissection, the hippocam-pus was cut into 350-µm-thick transverse slices using a tissueslicer (FHC Brunswick, USA). The slices were left to recover inACSF (equilibrated to pH 7.4 with 95% O2/5% CO2). A singleslice was transferred to a recording chamber, submerged betweentwo nylon nets and superfused continuously at 1.5 ml/min withACSF saturated with 95% O2/5% CO2 at 32±0.5°C, pH 7.4 (Bijaket al. 1996, 2001).
Extracellular recordings
For extracellular recording of population spikes, a glass microelec-trode filled with 2 M NaCl (resistance 2–4 MΩ) was positioned inthe stratum pyramidale of the CA1 area. A bipolar twisted-wireelectrode was placed in the stratum radiatum to stimulate theSchaffer collateral-commissural fibre pathway. The stimuli weresquare-wave pulses (0.1 ms, 0.1 Hz). Stimulus strength was ad-justed to evoke population spikes with a half-maximum amplitude.
Intracellular recordings
For intracellular recording from CA1 pyramidal cells, microelec-trodes (1.5-mm borosilicate glass, pulled on a Flaming-Brownpuller, Sutter Instruments, Novato, Calif., USA) were filled with 3 M KCl (resistance 40–100 MΩ). The recorded signals were am-plified (Axoprobe, Axon Instruments, Foster City, Calif., USA),bandpass-filtered (1 Hz–10 kHz) and stored on a PC hard disk af-ter AD conversion at 5–10 kHz (CED1401 interface and SIGAVGdata acquisition software; Cambridge Electronic, UK).
Only cells with resting membrane potentials more negativethan –60 mV, an input resistance exceeding 50 MΩ and over-shooting action potentials were selected for the study. Before drugapplication, the membrane potential was set to –65 mV by injec-tion of DC current. Input resistance was determined by measuringthe voltage response to the hyperpolarizing current injection (450 ms, 0.1 nA) at the end of the current pulse.
After stabilization of the baseline response for at least 20 min(defined as no more than 10% variation in the median amplitude ofthe population spike or stable membrane potential), the slice wassuperfused with 1 µM 8-OH-DPAT for 10 min. The drug was thenwashed with a standard solution for 20 min. The antagonist (S)-N-tert-butyl-3-[4-(2-methoxyphenyl)piperazin-1-yl]-2-phenylpropan-amide dihydrochloride [(S)WAY 100135, 5 µM], when used, wasapplied for 40 min before the application of agonist. In extracellu-lar experiments only one application per slice was made. Quantita-tive results are expressed as the mean (±SEM) percentage changefrom baseline (pre-drug) values (population spike) or the mean(±SEM) absolute value of membrane properties. Each individualcell or each slice in the extracellular recording was treated as an in-dividual, independent sample.
5-HT1A receptors
Preparation of brain sections
The rats were decapitated 2 or 24 h after the last vehicle or corti-costerone injection, the brains removed, cut rapidly into coronal
blocks containing the hippocampus, frozen by submerging inisopentane cooled in dry ice and stored at –70°C until sectioning.Coronal sections (10 µm) were cut on a cryostat (Leica LC 3000),thaw-mounted on gelatinized glass microscope slides and stored at–20 °C.
5-HT1A receptor autoradiography
At the beginning, the sections were preincubated (30 min, roomtemperature) in 50 mM TRIS-HCl buffer (pH 7.4) containing (inmM) 180 NaCl, 5 KCl, 2,5 CaCl2, and 1,2 MgCl2. A binding assaywas carried out (Wedzony et al. 1997) by incubating the sectionsfor 60 min (room temperature) with 1.5 nM [3H]8-OH-DPAT(Amersham, UK, specific activity 137 mCi/mmol) in the same 50 mMTRIS-HCl buffer supplemented with ascorbic acid (10 mg/dl) andpargyline (0.2 mg/dl). Non-specific binding was determined inadjacent sections under the same conditions except for the pres-ence of 5 µM 5-HT. After the incubation sections were rinsed,dried and apposed to a Hyperfilm-3H (Amersham) along with a tri-tium standard (3H-microscale, Amersham) in an X-ray cassette for3 weeks at 4 °C. The films were then developed with D-19 Kodakdeveloper. Autoradiographs were quantified by a computerizeddensitometry (image analysis system Image ProPlus, Media Cy-bernetics, USA). The measured light transmittance values wereconverted to a tissue equivalent radioactivity (with reference tostandards) and thus to binding values (in femtomole/milligram tis-sue, with reference to the specific activity) of the radiolabelled lig-and. Values of non-specific binding were determined directly fromfilm images of the sections incubated in the presence of the dis-placer 5-HT (5 µM). Those values were averaged and subtractedfrom the total binding to obtain the specific binding. The cresyl vi-olet staining of tissue sections in conjunction with an atlas (Paxi-nos and Watson 1998) was used to verify anatomical regions.
In situ hybridization histochemistry
Prior to hybridization, the tissue sections were fixed in 0.01 Mphosphate-buffered saline (pH 7.0) containing 4% paraformalde-hyde (10 min, 0 °C). The slides were then washed and acetylatedwith a mixture of 0.1 M triethanolamine (pH 8) and 0.25% aceticanhydride for 10 min and subsequently dehydrated in gradedethanol, defatted in chloroform, rinsed in graded ethanol and air-dried. For the in situ hybridization assay, a mixture of two 30-merdeoxyoligonucleotide probes (Genset, USA), corresponding toamino acid sequences 175–184 and 262–271 of the rat 5-HT1A re-ceptor, was used (Liao et al. 1993). An homology search (FastGENBANK) of the selected sequences showed no significant ho-mology with any known protein. The probes were labelled with[α-35S]dATP (NEN, Belgium) with terminal transferase(Boehringer-Mannheim, Germany), purified choromatographically(Micro Bio-Spin chromatography columns, BioRad, USA) andstored (3×105 dpm/µl) at –20°C until further use. The sectionswere hybridized with the [35S]dATP-labelled probe (100 µl/slide),diluted to final activity of about 2.45×105 dpm/ml in hybridizationbuffer [50% deionized formamide, 4×saline sodium citrate (SSC),25 mM sodium phosphate, 1 mM sodium pyrophosphate, 1×Den-hardt’s solution, 200 µg/ml salmon sperm DNA, 100 µg/mlpolyadenylic acid, 10% dextran sulphate, 250 µg/ml yeast tRNA]for 20 h at 42°C in a humidified incubator (Wedzony et al. 2000).After incubation, the sections were rinsed in 1×SSC (room tem-perature), washed 3 times in 1×SSC at 55°C for 15 min, then in1×SSC at room temperature for 30 min and rinsed finally in0.1×SSC and in sterile water. After washing, the slides were dehy-drated with graded ethanol, dried and exposed to an X-ray film(Hyperfilm-βmax, Amersham) for 20 days at 4 °C and processedwith D-19 Kodak developer. The specificity of the hybridizationsignal was confirmed by visualizing silver grains over individualcell bodies. To this end the slices were subsequently immersed ina nuclear photographic emulsion (Kodak NBT 3 emulsion) and leftfor 12 weeks. After processing with D-19 Kodak developer theywere counterstained with cresyl violet. The autoradiographs were
359
analysed using an image analysis system (SPOT-32 camera andImage ProPlus). For quantification, the mean optical density of aspecific hybridization signal was measured in different regions ofthe hippocampus.
Measurement of 5-HT and 5-HIAA concentrations
The rats were sacrificed by decapitation 2 h after single injection ofcorticosterone or, in the chronically treated group, 24 h after thelast dose of corticosterone, the brains removed quickly and the hip-pocampus dissected out. The tissue samples were stored at –70°C until assay. On the day of assay the tissue was homogenizedin 0.1 M HClO4 containing 0.26 mM ascorbic acid, centrifuged(14,000 g) and filtered through 0.2-µm cellulose membranes. Todetermine the 5-HT and 5-HIAA levels, 10-µl sample aliquots wereinjected onto a HPLC column (ODS, Phase II, particle size 3 µm,3.2/100 mm; BAS). A standard HPLC system (BAS 400, Bioana-lytical Systems, USA) with electrochemical detection (BAS-LC-4Bdetector with an oxidation potential of 0.65 V against the Ag/AgClreference electrode and a glassy-carbon working electrode TL5)was used. The mobile phase (flow rate 0.8 ml/min) consisted of0.06 M citrate-phosphate buffer (pH 3.5), 0.27 mM EDTA, 0.32 mMsodium octylsulphonate and 2.4% methanol. 5-HT and 5-HIAAwere quantified by a peak area comparison with standards, run onthe test day using BAS Inject software. Brain areas were verifiedhistologically in slices transferred into a solution of a phosphatebuffer containing 4% paraformaldehyde.
Drugs, data presentation and statistics
The following drugs and substances were used: corticosterone 21-acetate (Sigma, St. Louis, USA), 8-OH-DPAT (RBI, Natick,Mass., USA), (S)WAY 100135 (Tocris, Bristol, UK). HPLCreagents were purchased from Merck (Darmstadt, Germany). Allthe other reagents were purchased from Sigma.
Data are presented as means±SEM and were analysed by one-way ANOVA. When this revealed significant main effects or in-
teractions, post hoc comparisons were carried out using the New-man Keuls test. P<0.05 was accepted as significant.
Results
Serum corticosterone levels
Blood corticosterone levels in the different groups were:8.55±1.45 µg/dl in vehicle group; 36.50±3.10 µg/dl inblood of rats sacrificed 2 h after a single dose of cortico-sterone; 59.15±5.45 and 20.47±0.50 µg/dl in rats sacrificed2 or 24 h respectively after the last dose in the chronicallytreated group (all corticosterone treatments P<0.01 vs. ve-hicle group).
The action of corticosterone on 8-OH-DPAT-inducedelectrophysiological effects
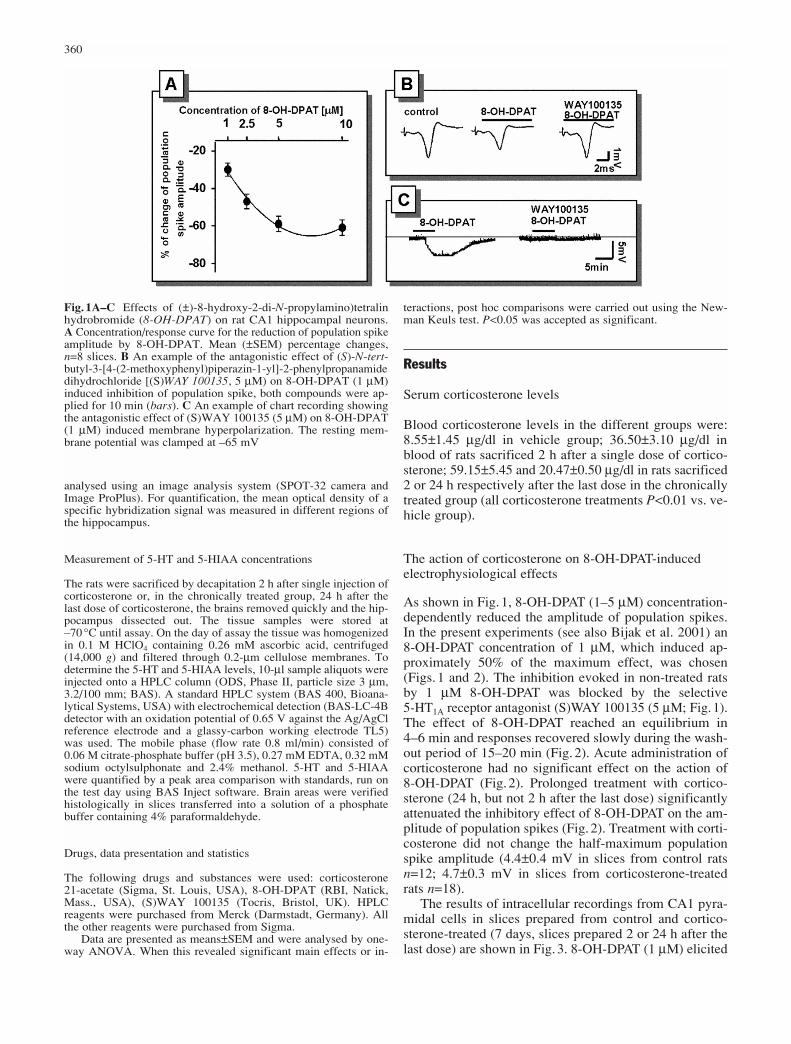
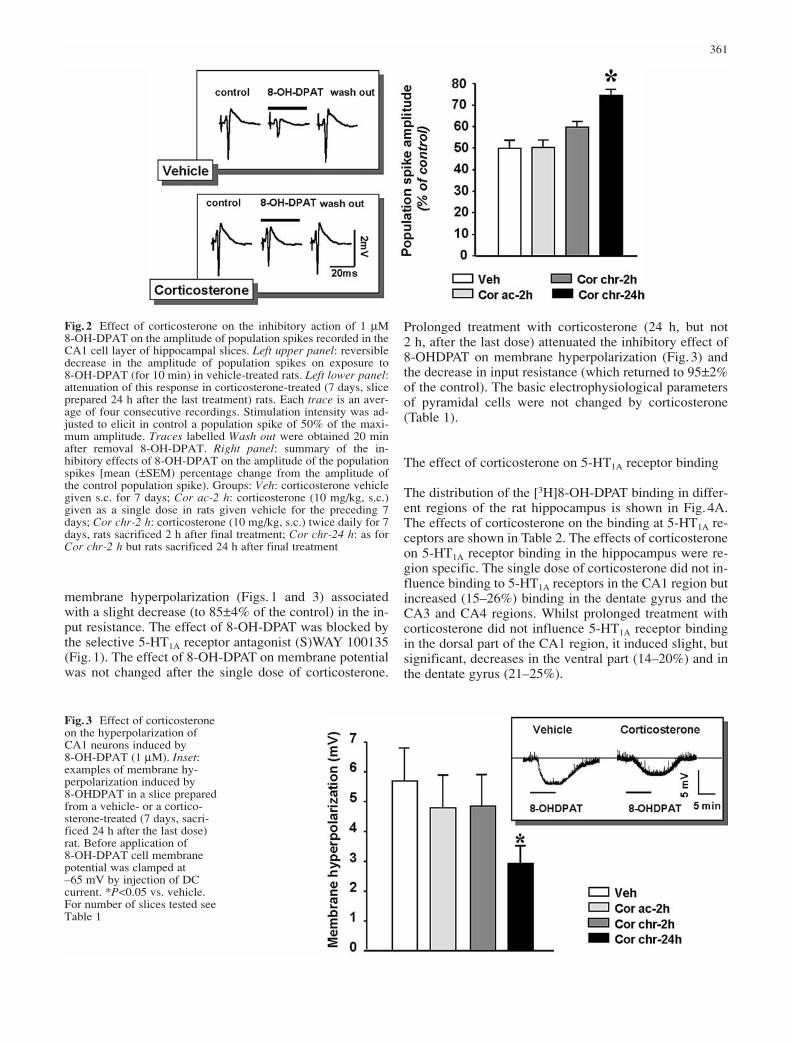
As shown in Fig.1, 8-OH-DPAT (1–5 µM) concentration-dependently reduced the amplitude of population spikes.In the present experiments (see also Bijak et al. 2001) an8-OH-DPAT concentration of 1 µM, which induced ap-proximately 50% of the maximum effect, was chosen(Figs. 1 and 2). The inhibition evoked in non-treated ratsby 1 µM 8-OH-DPAT was blocked by the selective 5-HT1A receptor antagonist (S)WAY 100135 (5 µM; Fig.1).The effect of 8-OH-DPAT reached an equilibrium in 4–6 min and responses recovered slowly during the wash-out period of 15–20 min (Fig.2). Acute administration ofcorticosterone had no significant effect on the action of 8-OH-DPAT (Fig.2). Prolonged treatment with cortico-sterone (24 h, but not 2 h after the last dose) significantlyattenuated the inhibitory effect of 8-OH-DPAT on the am-plitude of population spikes (Fig.2). Treatment with corti-costerone did not change the half-maximum populationspike amplitude (4.4±0.4 mV in slices from control ratsn=12; 4.7±0.3 mV in slices from corticosterone-treatedrats n=18).
The results of intracellular recordings from CA1 pyra-midal cells in slices prepared from control and cortico-sterone-treated (7 days, slices prepared 2 or 24 h after thelast dose) are shown in Fig.3. 8-OH-DPAT (1 µM) elicited
360
Fig.1A–C Effects of (±)-8-hydroxy-2-di-N-propylamino)tetralinhydrobromide (8-OH-DPAT) on rat CA1 hippocampal neurons.A Concentration/response curve for the reduction of population spikeamplitude by 8-OH-DPAT. Mean (±SEM) percentage changes,n=8 slices. B An example of the antagonistic effect of (S)-N-tert-butyl-3-[4-(2-methoxyphenyl)piperazin-1-yl]-2-phenylpropanamidedihydrochloride [(S)WAY 100135, 5 µM) on 8-OH-DPAT (1 µM)induced inhibition of population spike, both compounds were ap-plied for 10 min (bars). C An example of chart recording showingthe antagonistic effect of (S)WAY 100135 (5 µM) on 8-OH-DPAT(1 µM) induced membrane hyperpolarization. The resting mem-brane potential was clamped at –65 mV
membrane hyperpolarization (Figs. 1 and 3) associatedwith a slight decrease (to 85±4% of the control) in the in-put resistance. The effect of 8-OH-DPAT was blocked bythe selective 5-HT1A receptor antagonist (S)WAY 100135(Fig.1). The effect of 8-OH-DPAT on membrane potentialwas not changed after the single dose of corticosterone.
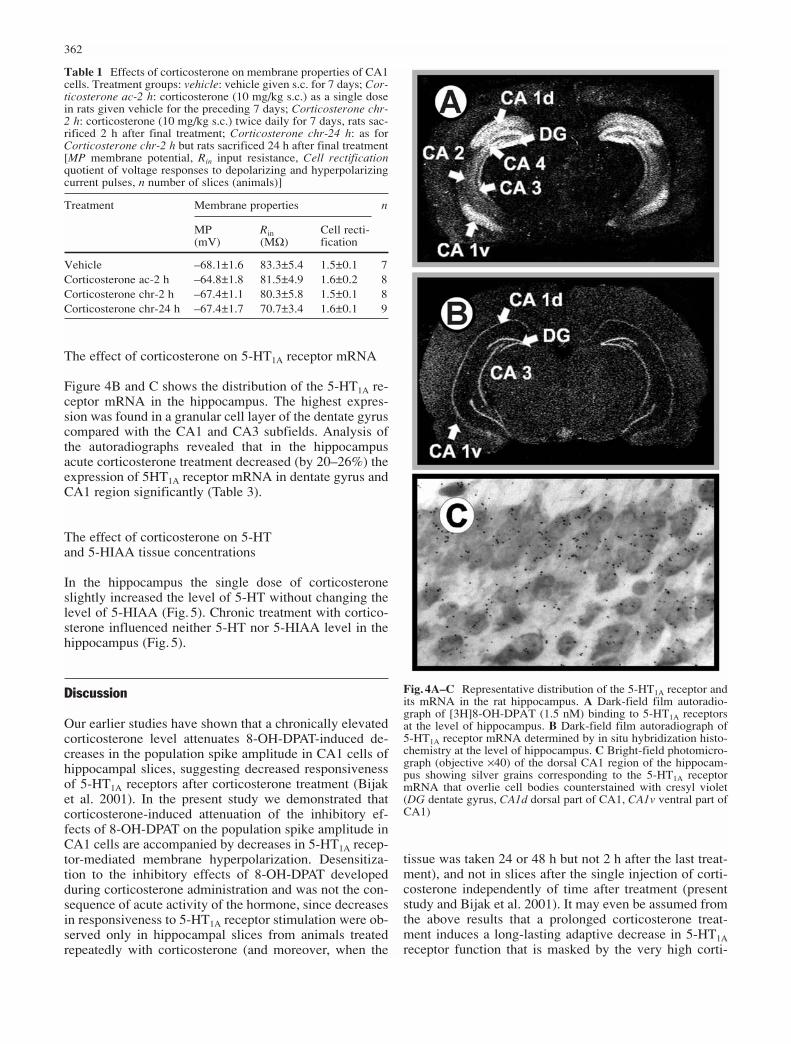
Prolonged treatment with corticosterone (24 h, but not 2 h, after the last dose) attenuated the inhibitory effect of8-OHDPAT on membrane hyperpolarization (Fig.3) andthe decrease in input resistance (which returned to 95±2%of the control). The basic electrophysiological parametersof pyramidal cells were not changed by corticosterone(Table 1).
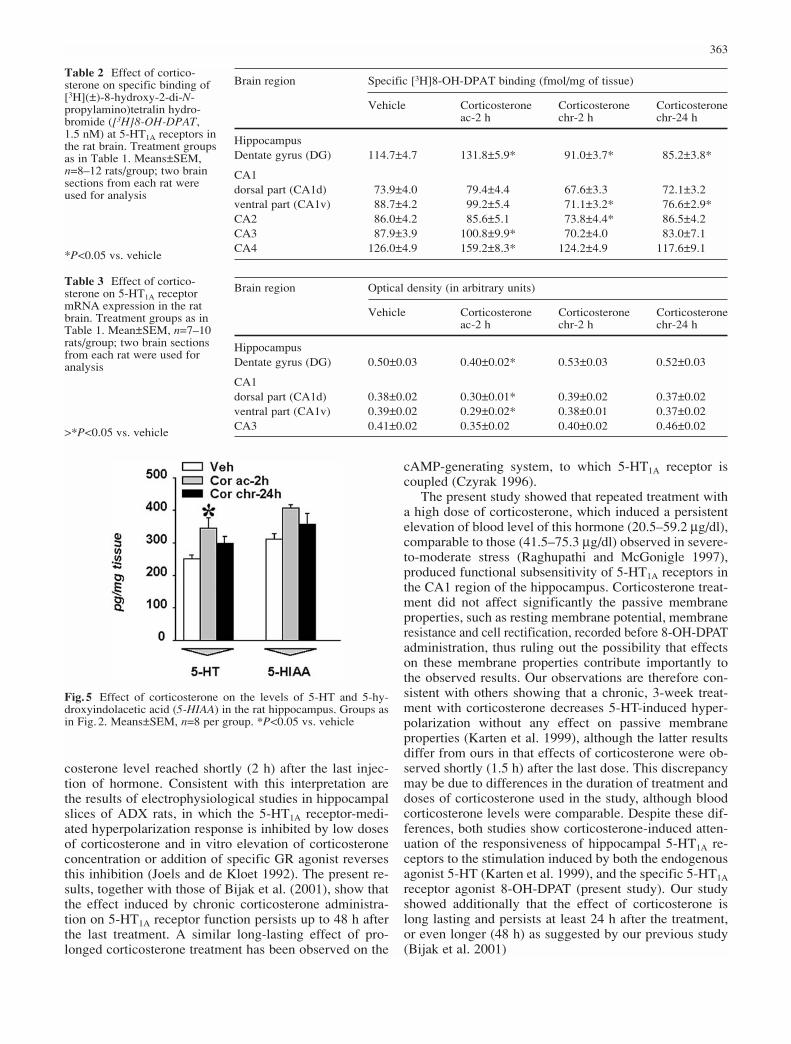
The effect of corticosterone on 5-HT1A receptor binding
The distribution of the [3H]8-OH-DPAT binding in differ-ent regions of the rat hippocampus is shown in Fig. 4A.The effects of corticosterone on the binding at 5-HT1A re-ceptors are shown in Table 2. The effects of corticosteroneon 5-HT1A receptor binding in the hippocampus were re-gion specific. The single dose of corticosterone did not in-fluence binding to 5-HT1A receptors in the CA1 region butincreased (15–26%) binding in the dentate gyrus and theCA3 and CA4 regions. Whilst prolonged treatment withcorticosterone did not influence 5-HT1A receptor bindingin the dorsal part of the CA1 region, it induced slight, butsignificant, decreases in the ventral part (14–20%) and inthe dentate gyrus (21–25%).
361
Fig.2 Effect of corticosterone on the inhibitory action of 1 µM 8-OH-DPAT on the amplitude of population spikes recorded in theCA1 cell layer of hippocampal slices. Left upper panel: reversibledecrease in the amplitude of population spikes on exposure to 8-OH-DPAT (for 10 min) in vehicle-treated rats. Left lower panel:attenuation of this response in corticosterone-treated (7 days, sliceprepared 24 h after the last treatment) rats. Each trace is an aver-age of four consecutive recordings. Stimulation intensity was ad-justed to elicit in control a population spike of 50% of the maxi-mum amplitude. Traces labelled Wash out were obtained 20 minafter removal 8-OH-DPAT. Right panel: summary of the in-hibitory effects of 8-OH-DPAT on the amplitude of the populationspikes [mean (±SEM) percentage change from the amplitude ofthe control population spike). Groups: Veh: corticosterone vehiclegiven s.c. for 7 days; Cor ac-2 h: corticosterone (10 mg/kg, s.c.)given as a single dose in rats given vehicle for the preceding 7days; Cor chr-2 h: corticosterone (10 mg/kg, s.c.) twice daily for 7days, rats sacrificed 2 h after final treatment; Cor chr-24 h: as forCor chr-2 h but rats sacrificed 24 h after final treatment
Fig.3 Effect of corticosteroneon the hyperpolarization ofCA1 neurons induced by 8-OH-DPAT (1 µM). Inset: examples of membrane hy-perpolarization induced by 8-OHDPAT in a slice preparedfrom a vehicle- or a cortico-sterone-treated (7 days, sacri-ficed 24 h after the last dose)rat. Before application of 8-OH-DPAT cell membranepotential was clamped at –65 mV by injection of DCcurrent. *P<0.05 vs. vehicle.For number of slices tested seeTable 1
The effect of corticosterone on 5-HT1A receptor mRNA
Figure 4B and C shows the distribution of the 5-HT1A re-ceptor mRNA in the hippocampus. The highest expres-sion was found in a granular cell layer of the dentate gyruscompared with the CA1 and CA3 subfields. Analysis ofthe autoradiographs revealed that in the hippocampusacute corticosterone treatment decreased (by 20–26%) theexpression of 5HT1A receptor mRNA in dentate gyrus andCA1 region significantly (Table 3).
The effect of corticosterone on 5-HT and 5-HIAA tissue concentrations
In the hippocampus the single dose of corticosteroneslightly increased the level of 5-HT without changing thelevel of 5-HIAA (Fig.5). Chronic treatment with cortico-sterone influenced neither 5-HT nor 5-HIAA level in thehippocampus (Fig.5).
Discussion
Our earlier studies have shown that a chronically elevatedcorticosterone level attenuates 8-OH-DPAT-induced de-creases in the population spike amplitude in CA1 cells ofhippocampal slices, suggesting decreased responsivenessof 5-HT1A receptors after corticosterone treatment (Bijaket al. 2001). In the present study we demonstrated thatcorticosterone-induced attenuation of the inhibitory ef-fects of 8-OH-DPAT on the population spike amplitude inCA1 cells are accompanied by decreases in 5-HT1A recep-tor-mediated membrane hyperpolarization. Desensitiza-tion to the inhibitory effects of 8-OH-DPAT developedduring corticosterone administration and was not the con-sequence of acute activity of the hormone, since decreasesin responsiveness to 5-HT1A receptor stimulation were ob-served only in hippocampal slices from animals treatedrepeatedly with corticosterone (and moreover, when the
tissue was taken 24 or 48 h but not 2 h after the last treat-ment), and not in slices after the single injection of corti-costerone independently of time after treatment (presentstudy and Bijak et al. 2001). It may even be assumed fromthe above results that a prolonged corticosterone treat-ment induces a long-lasting adaptive decrease in 5-HT1Areceptor function that is masked by the very high corti-
362
Table 1 Effects of corticosterone on membrane properties of CA1cells. Treatment groups: vehicle: vehicle given s.c. for 7 days; Cor-ticosterone ac-2 h: corticosterone (10 mg/kg s.c.) as a single dosein rats given vehicle for the preceding 7 days; Corticosterone chr-2 h: corticosterone (10 mg/kg s.c.) twice daily for 7 days, rats sac-rificed 2 h after final treatment; Corticosterone chr-24 h: as forCorticosterone chr-2 h but rats sacrificed 24 h after final treatment[MP membrane potential, Rin input resistance, Cell rectificationquotient of voltage responses to depolarizing and hyperpolarizingcurrent pulses, n number of slices (animals)]
Treatment Membrane properties n
MP Rin Cell recti-(mV) (MΩ) fication
Vehicle –68.1±1.6 83.3±5.4 1.5±0.1 7Corticosterone ac-2 h –64.8±1.8 81.5±4.9 1.6±0.2 8Corticosterone chr-2 h –67.4±1.1 80.3±5.8 1.5±0.1 8Corticosterone chr-24 h –67.4±1.7 70.7±3.4 1.6±0.1 9
Fig.4A–C Representative distribution of the 5-HT1A receptor andits mRNA in the rat hippocampus. A Dark-field film autoradio-graph of [3H]8-OH-DPAT (1.5 nM) binding to 5-HT1A receptorsat the level of hippocampus. B Dark-field film autoradiograph of5-HT1A receptor mRNA determined by in situ hybridization histo-chemistry at the level of hippocampus. C Bright-field photomicro-graph (objective ×40) of the dorsal CA1 region of the hippocam-pus showing silver grains corresponding to the 5-HT1A receptormRNA that overlie cell bodies counterstained with cresyl violet(DG dentate gyrus, CA1d dorsal part of CA1, CA1v ventral part ofCA1)
costerone level reached shortly (2 h) after the last injec-tion of hormone. Consistent with this interpretation arethe results of electrophysiological studies in hippocampalslices of ADX rats, in which the 5-HT1A receptor-medi-ated hyperpolarization response is inhibited by low dosesof corticosterone and in vitro elevation of corticosteroneconcentration or addition of specific GR agonist reversesthis inhibition (Joels and de Kloet 1992). The present re-sults, together with those of Bijak et al. (2001), show thatthe effect induced by chronic corticosterone administra-tion on 5-HT1A receptor function persists up to 48 h afterthe last treatment. A similar long-lasting effect of pro-longed corticosterone treatment has been observed on the
cAMP-generating system, to which 5-HT1A receptor iscoupled (Czyrak 1996).
The present study showed that repeated treatment witha high dose of corticosterone, which induced a persistentelevation of blood level of this hormone (20.5–59.2 µg/dl),comparable to those (41.5–75.3 µg/dl) observed in severe-to-moderate stress (Raghupathi and McGonigle 1997),produced functional subsensitivity of 5-HT1A receptors inthe CA1 region of the hippocampus. Corticosterone treat-ment did not affect significantly the passive membraneproperties, such as resting membrane potential, membraneresistance and cell rectification, recorded before 8-OH-DPATadministration, thus ruling out the possibility that effectson these membrane properties contribute importantly tothe observed results. Our observations are therefore con-sistent with others showing that a chronic, 3-week treat-ment with corticosterone decreases 5-HT-induced hyper-polarization without any effect on passive membraneproperties (Karten et al. 1999), although the latter resultsdiffer from ours in that effects of corticosterone were ob-served shortly (1.5 h) after the last dose. This discrepancymay be due to differences in the duration of treatment anddoses of corticosterone used in the study, although bloodcorticosterone levels were comparable. Despite these dif-ferences, both studies show corticosterone-induced atten-uation of the responsiveness of hippocampal 5-HT1A re-ceptors to the stimulation induced by both the endogenousagonist 5-HT (Karten et al. 1999), and the specific 5-HT1Areceptor agonist 8-OH-DPAT (present study). Our studyshowed additionally that the effect of corticosterone islong lasting and persists at least 24 h after the treatment,or even longer (48 h) as suggested by our previous study(Bijak et al. 2001)
363
Table 2 Effect of cortico-sterone on specific binding of[3H](±)-8-hydroxy-2-di-N-propylamino)tetralin hydro-bromide ([3H]8-OH-DPAT, 1.5 nM) at 5-HT1A receptors inthe rat brain. Treatment groupsas in Table 1. Means±SEM,n=8–12 rats/group; two brainsections from each rat wereused for analysis
*P<0.05 vs. vehicle
Brain region Specific [3H]8-OH-DPAT binding (fmol/mg of tissue)
Vehicle Corticosterone Corticosterone Corticosterone ac-2 h chr-2 h chr-24 h
HippocampusDentate gyrus (DG) 114.7±4.7 131.8±5.9* 91.0±3.7* 85.2±3.8*
CA1dorsal part (CA1d) 73.9±4.0 79.4±4.4 67.6±3.3 72.1±3.2ventral part (CA1v) 88.7±4.2 99.2±5.4 71.1±3.2* 76.6±2.9*CA2 86.0±4.2 85.6±5.1 73.8±4.4* 86.5±4.2 CA3 87.9±3.9 100.8±9.9* 70.2±4.0 83.0±7.1CA4 126.0±4.9 159.2±8.3* 124.2±4.9 117.6±9.1
Table 3 Effect of cortico-sterone on 5-HT1A receptormRNA expression in the ratbrain. Treatment groups as inTable 1. Mean±SEM, n=7–10rats/group; two brain sectionsfrom each rat were used foranalysis
>*P<0.05 vs. vehicle
Brain region Optical density (in arbitrary units)
Vehicle Corticosterone Corticosterone Corticosterone ac-2 h chr-2 h chr-24 h
HippocampusDentate gyrus (DG) 0.50±0.03 0.40±0.02* 0.53±0.03 0.52±0.03
CA1dorsal part (CA1d) 0.38±0.02 0.30±0.01* 0.39±0.02 0.37±0.02ventral part (CA1v) 0.39±0.02 0.29±0.02* 0.38±0.01 0.37±0.02CA3 0.41±0.02 0.35±0.02 0.40±0.02 0.46±0.02
Fig.5 Effect of corticosterone on the levels of 5-HT and 5-hy-droxyindolacetic acid (5-HIAA) in the rat hippocampus. Groups asin Fig.2. Means±SEM, n=8 per group. *P<0.05 vs. vehicle

Corticosterone may influence other cell membranecharacteristics, not measured in our study. Chronic treat-ment of ADX rats with corticosterone decreases the am-plitude and half-decay time of slow after-hyperpolariza-tions, which, theoretically, should increase cell excitabil-ity and increase the synaptic output from the area CA1 ofthe hippocampus (Mueller and Beck 2000). Interestingly,consistent with corticosterone-induced increased ex-citability of CA1 cells are results showing that chronictreatment with corticosterone increases the excitatory ef-fect of 5-HT4 receptor activation in the CA1 area of the rathippocampus (Zahorodna et al. 2000; Bijak et al. 2001).Other studies, however, have shown a reduction of 5-HT4receptor-induced effects in hippocampal slices from ADXrats treated with a high dose of corticosterone (Birnstieland Beck 1995).
In the next part of our study we addressed the questionof whether corticosterone treatment that changes 5-HT1Areceptor function in the hippocampus also modulates theexpression of these receptors. Specifically, we measured5-HT1A receptor binding and mRNA level in differentparts of the hippocampal subfields after corticosteronetreatment. A single injection of corticosterone indeed de-creased 5-HT1A receptor mRNA in the CA1 region anddentate gyrus of hippocampus, although both those changesdisappeared with prolonged administration of cortico-sterone. The binding studies conducted on the same ani-mals in which mRNA levels were measured showed cor-ticosterone-induced down-regulation of the 5-HT1A recep-tor binding after prolonged, but not after single, adminis-tration; although only in the ventral part (not in the dorsal)of the CA1 region and in the dentate gyrus. These resultssuggest that acute changes in plasma corticosterone areaccompanied very rapidly by decreases in 5-HT1A recep-tor mRNA, whereas alterations in the receptor protein lev-els occur much more slowly. This is not surprising in thelight of the 5-day half-life of the 5-HT1A receptor protein(Pinto and Battaglia 1994). Our results showing decreased5-HT1A receptor mRNA expression in the CA1 regiononly after acute corticosterone elevation are consistentwith other data (Meijer et al. 1997b; Karten et al. 1999;Lopez et al. 1999). In the dentate gyrus the decreases havebeen reported after both acute and prolonged hormone el-evation in some (Meijer et al. 1997b; Karten et al. 1999,Lopez et al. 1999), but not all (Holmes et al. 1995) stud-ies. Regarding 5-HT1A receptor binding, the general con-sensus is that the chronically elevated corticosteronedown-regulates 5-HT1A receptor binding in the dentategyrus (Chaouloff 1995; Takao et al. 1997; Lopez et al.1998), again in line with our results. Data concerning theCA1 region are inconsistent, with both down-regulation(Chaouloff 1995; Lopez et al. 1998) or no effect (Fernan-des et al. 1997) being reported. Our results showing thatthe effect of corticosterone on 5-HT1A receptor bindingdiffers between the dorsal and ventral parts of CA1 sug-gest that corticosterone regulates 5-HT1A receptor expres-sion in a cell-specific manner. They may also help to ex-plain apparent inconsistencies in the reported effects ofglucocorticoids in the CA1 region, since most of the cited
studies did not differentiate between dorsal and ventralparts of this region.
There is still discussion about the mechanism by whichcorticosterone regulates 5-HT1A receptor expression; i.e.whether it affects the 5-HT1A receptor gene directly orrather other elements of the receptor cellular cycle. Ex-periments using MR- and GR-selective ligands (rat) andgene knockout approaches (mouse) suggest that primarilyMR, but in cooperation with GR, are involved in the neg-ative regulation of 5-HT1A gene expression (Joels and deKloet 1992; Meijer and de Kloet 1995; Meijer et al.1997a; Karten et al. 1999). The precise mechanism haslong remained unclear, but Ou et al. (2001), who firstidentified nMRE/GRE (negative mineralocorticoid/gluco-corticoid response element) in 5-HT1A receptor promoter,have suggested recently that the heterodimerization ofMR and GR mediates direct corticosteroid-induced trans-repression of the 5-HT1A receptor promoter. The discov-ery that heterodimerization of MR and GR is necessaryfor efficient attenuation of 5-HT1A receptor transcriptioncorresponds well with our results showing that glucocorti-coids induce the most pronounced and consistent changesin 5-HT1A receptor expression in those regions of hippo-campus (dentate gyrus, CA1) in which both MR and GRare expressed strongly (Hansson et al. 2000).
Our results show that in the hippocampus, cortico-sterone can regulate both the functional responsiveness ofthe 5-HT1A receptor and its gene transcription and transla-tion to the receptor protein. When we tried to correlate ob-served functional changes with the alterations in expres-sion of 5-HT1A receptors it appeared that the site of func-tional changes was different in comparison with time-course and site of changes in receptor expression. Suchresults suggest that functional and receptor changes mayoccur independently; however, they do not totally excludethe possibility that corticosterone-induced desensitizationof hippocampal 5-HT1A receptor function results from (atleast partly) corticosterone-induced changes in 5-HT1A re-ceptor expression. In line with this conclusion, the studiesof Karten et al. (1999, 2001) showed also no correlationbetween the effects of corticosterone on the 5-HT1A re-ceptor expression and on the electrophysiological re-sponses induced by 5-HT1A receptor stimulation in bothidentified CA1 and dentate gyrus neurons. In fact fre-quently observed changes in the receptor binding are ap-parently not accompanied by a functional change and viceversa, as shown in the present and other studies (Bijak etal. 1996; Mongeau et al. 1997; Karten et al. 1999, 2001).
Changes in cellular processes distal to receptor proteinthus seem to be the major contributor to the altered re-sponsiveness of these receptors. The 5-HT1A receptor iscoupled to a number of different effector systems, ofwhich the opening of inwardly rectifying K+ channels andinhibition of adenylyl cyclase are most frequently de-scribed as the functional correlates of 5-HT1A receptor ac-tivation (Andrade et al. 1986; De Vivo and Maayani1986). The 5-HT1A receptor is coupled to these effectorsystems through specific G proteins, and there is convinc-ing evidence that glucocorticoids regulate levels of G pro-
364

tein α-subunits (Saito et al. 1989; Okuhara et al. 1997).Moreover, also the regulators of G protein signalling(RGS) proteins seem to be under glucocorticoid control(Ni et al. 1999). Glucocorticoids alter G protein-activated,inwardly rectifying potassium channel protein levels inthe hippocampus (Muma and Beck 1999), which is notsurprising in the light of data showing that the gene en-coding this channel contains a potential binding site(GRE, glucocorticoid response element) for the GR re-ceptor (Schoots et al. 1997). Further, chronic treatmentwith corticosterone decrease PKA (protein kinase A) ac-tivity (Dwivedi and Pandey 2000). All described aboveeffects may be relevant in corticosterone-induced func-tional attenuation of 5-HT1A receptor responsiveness.Changes in 5-HT metabolism may also, at least theoreti-cally, alter 5-HT1A receptor function, however, in ourstudy, chronic corticosterone treatment did not influencelevels of 5-HT and 5-HIAA in the hippocampus, thus ex-cluding this possibility.
Chronic elevation of plasma corticosteroid levels oc-curs in some psychiatric disorders, most commonly in af-fective disorders such as depression (Modell et al. 1997;Murphy 1997). Since hyperactivity of the HPA system ap-parently exists before manifestation of clinical symptomsof depression, it may be assumed that this hyperactivityforms one of the risk factors for precipitating this disease(Holsboer and Barden 1996; Modell et al. 1998). Themechanisms by which the hyperactivity of HPA systemtriggers clinical symptoms are still unknown. One hypoth-esis is that hypercorticism may alter the brain 5-HT sys-tem, which is important in the aetiology of depression(Meltzer 1989; Mongeau et al. 1997). Indeed, Lopez et al.(1998) have shown that suicide victims with a history ofdepression have decreases in hippocampal 5-HT1A recep-tors, which resemble those observed in chronically stressedor glucocorticoid-treated animals. Moreover, depressedpatients exhibit an increased basal cortisol secretion andsignificantly decreased ACTH and cortisol responses tothe 5-HT1A receptor agonist ipsapirone (Lesch 1991). Incontrast, chronic administration of antidepressant drugsinduces functional hypersensitivity of postsynaptic 5-HT1Areceptors, at least as detected electrophysiologically inthe hippocampus (Bijak et al. 1996, 2001; Beck et al.1997; Haddjeri et al. 1998). However, most receptor stud-ies have shown no change in the [3H]8-OH-DPAT bindingin the rat hippocampus following prolonged treatmentwith antidepressants (Watanabe et al. 1993; Hayakawa etal. 1994; Bijak et al. 1996). Comparing the effects of glu-cocorticoids and antidepressant drugs on hippocampal 5-HT1A receptor function, and considering changes in 5-HT1A receptors in depressed patients, it may be con-cluded that both functional desensitization and a decreasein the expression of 5-HT1A receptors induced by gluco-corticoids may account considerably for their “pro-de-pressive” activity.
Summing up, a prolonged rise in the corticosteronelevel in intact animals down-regulates both the functionalresponsiveness of the 5-HT1A receptor and its expressionin rat hippocampus, suggesting that these 5-HT1A recep-
tors could be controlled tonically by corticosteroids. Theability of corticosterone to influence the 5-HT1A receptorsseem to be important in the context of clinical data/hy-potheses as it points to significance of the HPA systemand 5-HT1A receptors in the hippocampus for the neurobi-ology of affective and mood disorders (Lopez et al. 1998;Modell et al. 1998; Karten et al. 1999). Further studiesinto the corticosterone/5-HT interaction in brain areasother than the hippocampus as well as post-mortem hu-man brain studies should facilitate better understanding ofthe role of elevated glucocorticoid level in certain psychi-atric diseases.
Acknowledgements This publication is dedicated to the memoryof Dr. Maria Bijak, who collaborated with Anna Czyrak on variouspsychoneuropharmacological research projects. The authors wishto thank Ms. Elżbieta Smolak for the English linguistic correctionof manuscript.
References
Andrade R, Malenka RC, Nicoll RA (1986) A G protein couplesserotonin and GABAB receptors to the same channels in hip-pocampus. Science 234:1261–1265
Beck SG, Choi KC, List TJ (1992) Comparison of 5-hydroxytrypt-amine1A-mediated hyperpolarization in CA1 and CA3 hip-pocampal pyramidal cells. J Pharmacol Exp Ther 263:350–359
Beck SG, Choi KC, List TJ, Okuhara DY, Birnsteil S (1996) Cor-ticosterone alters 5-HT1A receptor-mediated hyperpolarizationin area CA1 hippocampal pyramidal neurons. Neuropsycho-pharmacology 14:27–33
Beck SG, Birnstiel S, Choi KC, Pouliot WA (1997) Fluoxetine se-lectively alters 5-hydroxytryptamine1A and gamma-aminobu-tyric acidB receptor-mediated hyperpolarization in area CA1,but not area CA3, hippocampal pyramidal cells. J PharmacolExp Ther 281:115–122
Bijak M, Tokarski K, Czyrak A, Mackowiak M, Wedzony K(1996) Imipramine increases the 5-HT1A receptor-mediated in-hibition of hippocampal neurons without changing the 5-HT1Areceptor binding. Eur J Pharmacol 305:79–85
Bijak M, Zahorodna A, Tokarski K (2001) Opposite effects of an-tidepressants and corticosterone on the sensitivity of hip-pocampal CA1 neurons to 5-HT1A and 5-HT4 receptor activa-tion. Naunyn Schmiedeberg’s Arch Pharmacol 363:491–498
Birnstiel S, Beck SG (1995) Modulation of the 5-hydroxytrypt-amine4 receptor-mediated response by short-term and long-term administration of corticosterone in rat CA1 hippocampalpyramidal neurons. J Pharmacol Exp Ther 273:1132–1138
Burnet PW, Eastwood SL, Harrison PJ (1996) 5-HT1A and 5-HT2Areceptor mRNAs and binding site densities are differentially al-tered in schizophrenia. Neuropsychopharmacology 15:442–455
Chalmers DT, Watson SJ (1991) Comparative anatomical distrib-ution of 5-HT1A receptor mRNA and 5-HT1A binding in ratbrain–a combined in situ hybridisation/in vitro receptor autora-diographic study. Brain Res 561:51–60
Chalmers DT, Kwak SP, Mansour A, Akil H, Watson SJ (1993)Corticosteroids regulate brain hippocampal 5-HT1A receptormRNA expression. J Neurosci 13:914–923
Chalmers DT, Lopez JF, Vazquez DM, Akil H, Watson SJ (1994)Regulation of hippocampal 5-HT1A receptor gene expressionby dexamethasone. Neuropsychopharmacology 10:215–222
Chaouloff F (1995) Regulation of 5-HT receptors by cortico-steroids: where do we stand? Fundam Clin Pharmacol 9:219–233
Czyrak A (1996) Modulation of the forskolin-induced cyclic AMPaccumulation by corticosterone. Pol J Pharmacol 48:595–599
de Kloet ER, Reul JM, Sutanto W (1990) Corticosteroids and thebrain. J Steroid Biochem Mol Biol 37:387–394
365

De Vivo M, Maayani S (1986) Characterization of the 5-hydroxy-tryptamine1A receptor-mediated inhibition of forskolin-stimu-lated adenylate cyclase activity in guinea pig and rat hip-pocampal membranes. J Pharmacol Exp Ther 238:248–253
Dwivedi Y, Pandey GN (2000) Adrenal glucocorticoids modulate[3H]cyclic AMP binding to protein kinase A (PKA), cyclicAMP-dependent PKA activity, and protein levels of selectiveregulatory and catalytic subunit isoforms of PKA in rat brain. JPharmacol Exp Ther 294:103–116
Fernandes C, McKittrick CR, File SE, McEwen BS (1997) De-creased 5-HT1A and increased 5-HT2A receptor binding afterchronic corticosterone associated with a behavioural indicationof depression but not anxiety. Psychoneuroendocrinology 22:477–491
Haddjeri N, Blier P, de Montigny C (1998) Long-term antidepres-sant treatments result in a tonic activation of forebrain 5-HT1Areceptors. J Neurosci 18:10150–10156
Hansson AC, Cintra A, Belluardo N, Sommer W, Bhatnagar M,Bader M, Ganten D, Fuxe K (2000) Gluco- and mineralocorti-coid receptor-mediated regulation of neurotrophic factor geneexpression in the dorsal hippocampus and the neocortex of therat. Eur J Neurosci 12:2918–2934
Hayakawa H, Shimizu M, Nishida A, Motohashi N, Yamawaki S(1994) Increase in serotonin 1A receptors in the dentate gyrusas revealed by autoradiographic analysis following repeatedelectroconvulsive shock but not imipramine treatment. Neu-ropsychobiology 30:53–56
Holmes MC, Yau JL, French KL, Seckl JR (1995) The effect ofadrenalectomy on 5-hydroxytryptamine and corticosteroid re-ceptor subtype messenger RNA expression in rat hippocampus.Neuroscience 64:327–337
Holsboer F, Barden N (1996) Antidepressants and hypothalamic-pituitary-adrenocortical regulation. Endocrinol Rev 17:187–205
Joels M, de Kloet ER (1992) Coordinative mineralocorticoid andglucocorticoid receptor-mediated control of responses to sero-tonin in rat hippocampus. Neuroendocrinology 55:344–350
Joels M, de Kloet ER (1994) Mineralocorticoid and glucocorticoidreceptors in the brain. Implications for ion permeability andtransmitter systems. Prog Neurobiol 43:1–36
Karten YJ, Nair SM, van Essen L, Sibug R, Joels M (1999) Long-term exposure to high corticosterone levels attenuates serotoninresponses in rat hippocampal CA1 neurons. Proc Natl Acad SciUSA 96:13456–13461
Karten YJ, Stienstra CM, Joels M (2001) Corticosteroid effects onserotonin responses in granule cells of the rat dentate gyrus. J Neuroendocrinol 13:233–238
Kawata M, Yuri K, Ozawa H, Nishi M, Ito T, Hu Z, Lu H, YoshidaM (1998) Steroid hormones and their receptors in the brain. J Steroid Biochem Mol Biol 65:273–280
Kuroda Y, Watanabe Y, Albeck DS, Hastings NB, McEwen BS(1994) Effects of adrenalectomy and type I or type II glucocor-ticoid receptor activation on 5-HT1A and 5-HT2 receptor bind-ing and 5-HT transporter mRNA expression in rat brain. BrainRes 648:157–161
Lesch KP (1991) 5-HT1A receptor responsivity in anxiety disordersand depression. Prog Neuropsychopharmacol Biol Psychiatry15:723–733
Liao B, Miesak B, Azmitia EC (1993) Loss of 5-HT1A receptormRNA in the dentate gyrus of the long-term adrenalectomizedrats and rapid reversal by dexamethasone. Mol Brain Res 19:328–332
Lopez JF, Chalmers DT, Little KY, Watson SJ (1998) A.E. Ben-nett Research Award. Regulation of serotonin1A, glucocorti-coid, and mineralocorticoid receptor in rat and human hip-pocampus: implications for the neurobiology of depression.Biol Psychiatry 43:547–573
Lopez JF, Liberzon I, Vazquez DM, Young EA, Watson SJ (1999)Serotonin 1A receptor messenger RNA regulation in the hip-pocampus after acute stress. Biol Psychiatry 45:934–937
McEwen BS (1987) Glucocorticoid-biogenic amine interactions inrelation to mood and behavior. Biochem Pharmacol 36:1755–1763
Meijer OC, de Kloet ER (1995) A role for the mineralocorticoidreceptor in a rapid and transient suppression of hippocampal 5-HT1A receptor mRNA by corticosterone. J Neuroendocrinol7:653–657
Meijer OC, Cole TJ, Schmid W, Schutz G, Joels M, de Kloet ER(1997a) Regulation of hippocampal 5-HT1A receptor mRNAand binding in transgenic mice with a targeted disruption of theglucocorticoid receptor. Mol Brain Res 46:290–296
Meijer OC, Oosten RV van, de Kloet ER (1997b) Elevated basaltrough levels of corticosterone suppress hippocampal 5-hy-droxytryptamine(1A) receptor expression in adrenally intactrats: implication for the pathogenesis of depression. Neuro-science 80:419–426
Meltzer H (1989) Serotonergic dysfunction in depression. Br JPsychiatry 155 (Suppl 8):25–31
Modell S, Yassouridis A, Huber J, Holsboer F (1997) Cortico-steroid receptor function is decreased in depressed patients.Neuroendocrinology 65:216–222
Modell S, Lauer CJ, Schreiber W, Huber J, Krieg JC, Holsboer F(1998) Hormonal response pattern in the combined DEX-CRHtest is stable over time in subjects at high familial risk for af-fective disorders. Neuropsychopharmacology 18:253–262
Mongeau R, Blier P, de Montigny C (1997) The serotonergic andnoradrenergic systems of the hippocampus: their interactionsand the effects of antidepressant treatments. Brain Res Rev23:145–195
Mueller NK, Beck SG (2000) Corticosteroids alter the 5-HT(1A)receptor-mediated response in CA1 hippocampal pyramidalcells. Neuropsychopharmacology 23:419–427
Muma NA, Beck SG (1999) Corticosteroids alter G protein in-wardly rectifying potassium channels protein levels in hippo-campal subfields. Brain Res 839:331–335
Murphy BE (1997) Antiglucocorticoid therapies in major depres-sion: a review. Psychoneuroendocrinology 22 (Suppl 1):S125–S132
Ni YG, Gold SJ, Iredale PA, Terwilliger RZ, Duman RS, NestlerEJ (1999) Region-specific regulation of RGS4 (Regulator ofG-protein-signaling protein type 4) in brain by stress and glu-cocorticoids: in vivo and in vitro studies. J Neurosci 19:3674–3680
Okuhara DY, Beck SG, Muma NA (1997) Corticosterone alters Gprotein alpha-subunit levels in the rat hippocampus. Brain Res745:144–151
Ou XM, Storring JM, Kushwaha N, Albert PR (2001) Hetero-dimerization of mineralocorticoid and glucocorticoid receptorsat a novel negative response element of the 5-HT1A receptorgene. J Biol Chem 276:14299–14307
Paxinos G, Watson C (1998) The rat brain in stereotaxic coordi-nates 4th edn. Academic Press, San Diego
Piazza PV, Le Moal M (1997) Glucocorticoids as a biological sub-strate of reward: physiological and pathophysiological implica-tions. Brain Res Rev 25:359–372
Pinto W, Battaglia G (1994) Comparative recovery kinetics of 5-hydroxytryptamine 1A, 1B, and 2A receptor subtypes in ratcortex after receptor inactivation: evidence for differences inreceptor production and degradation. Mol Pharmacol 46:1111–1119
Pompeiano M, Palacios JM, Mengod G (1992) Distribution andcellular localization of mRNA coding for 5-HT1A receptor inthe rat brain: correlation with receptor binding. J Neurosci 12:440–453
Przegalinski E, Budziszewska B, Grochmal A (1992) Effect ofadenosine analogues on plasma corticosterone concentration inrats. Acta Endocrinol (Copenh) 127:471–475
Raghupathi RK, McGonigle P (1997) Differential effects of threeacute stressors on the serotonin 5-HT1A receptor system in ratbrain. Neuroendocrinology 65:246–258
366

Reul JM, Bosch FR van den, de Kloet ER (1987) Differential re-sponse of type I and type II corticosteroid receptors to changesin plasma steroid level and circadian rhythmicity. Neuroendo-crinology 45:407–412
Saito N, Guitart X, Hayward M, Tallman JF, Duman RS, NestlerEJ (1989) Corticosterone differentially regulates the expressionof Gs alpha and Gi alpha messenger RNA and protein in ratcerebral cortex. Proc Natl Acad Sci USA 86:3906–3910
Schoots O, Voskoglou T, Van Tol HH (1997) Genomic organiza-tion and promoter analysis of the human G-protein-coupled K+
channel Kir3.1 (KCNJ3/HGIRK1). Genomics 39:279–288Spencer RL, Young EA, Choo PH, McEwen BS (1990) Adrenal
steroid type I and type II receptor binding: estimates of in vivoreceptor number, occupancy, and activation with varying levelof steroid. Brain Res 514:37–48
Takao K, Nagatani T, Kitamura Y, Yamawaki S (1997) Effects ofcorticosterone on 5-HT1A and 5-HT2 receptor binding and onthe receptor-mediated behavioral responses of rats. Eur J Phar-macol 333:123–128
Tejani-Butt SM, Labow DM (1994) Time course of the effects ofadrenalectomy and corticosterone replacement on 5-HT1A re-ceptors and 5-HT uptake sites in the hippocampus and dorsalraphe nucleus of the rat brain: an autoradiographic analysis.Psychopharmacology 113:481–486
Van Eekelen JA, de Kloet ER (1992) Co-localization of brain cor-ticosteroid receptors in the rat hippocampus. Prog HistochemCytochem 26:250–258
Watanabe Y, Sakai RR, McEwen BS, Mendelson S (1993) Stressand antidepressant effects on hippocampal and cortical 5-HT1Aand 5-HT2 receptors and transport sites for serotonin. Brain Res615:87–94
Wedzony K, Mackowiak M, Czyrak A, Fijal K, Michalska B(1997) Single doses of MK-801, a non-competitive antagonistof NMDA receptors, increase the number of 5-HT1A serotoninreceptors in the rat brain. Brain Res 756:84–91
Wedzony K, Mackowiak M, Zajaczkowski W, Fijal K, Chocyk A,Czyrak A (2000) WAY 100135, an antagonist of 5-HT1A sero-tonin receptors, attenuates psychotomimetic effects of MK-801. Neuropsychopharmacology 23:547–559
Wolkowitz OM, Reus VI, Canick J, Levin B, Lupien S (1997)Glucocorticoid medication, memory and steroid psychosis inmedical illness. Ann NY Acad Sci 823:81–96
Zahorodna A, Tokarski K, Bijak M (2000) Repeated cortico-sterone administration increases excitatory effect of 5-HT4 re-ceptor agonist in the rat hippocampus. Pol J Pharmacol 52:107–109
Zhong P, Ciaranello RD (1995) Transcriptional regulation of hip-pocampal 5-HT1a receptors by corticosteroid hormones. MolBrain Res 29:23–34
367
Related Documents











