ORIGINAL ARTICLE Production, purification and characterization of serraticin A, a novel cold-active antimicrobial produced by Serratia proteamaculans 136 L.A. Sa ´ nchez 1 , M. Hedstro ¨m 2 , M.A. Delgado 3 and O.D. Delgado 1 1 PROIMI – CONICET, Chacabuco, Tucuma ´ n, Argentina 2 Department of Biotechnology, Center for Chemistry and Chemical Engineering, Lund University, Lund, Sweden 3 INSIBIO – CONICET, Chacabuco, Tucuma ´ n, Argentina Introduction The need to decrease the use of chemical additives in food has triggered the search for natural antimicrobial substances produced by micro-organisms from different sources. Application of antimicrobial compounds has been well documented as a biotechnological advantage over different industrial sectors like food-processing, pharmaceutical chemistry and cosmetic industries. Conse- quently, screening and selection programs for antimicro- bial producing micro-organisms and bioproducts have increased in the last years, particularly focusing into improving the process control as well as the quality and safety of final products by inhibition of both, undesirable pathogenic and spoilage bacteria (Zahner and Fielder 1995; Davies and Webb 1998). Since the introduction of penicillin in the 1940s, antibi- otics have a history of success in controlling morbidity and mortality caused by infectious diseases. However, as a consequence of frequent use, bacterial resistance to known classes of antibiotics has become a severe global problem in recent years and presents a continuous chal- lenge (Pohlmann et al. 2005). Nowadays, it cannot be ignored that the antibiotic age is under adverse circum- stances. The high levels of antibiotic resistance among important pathogens along with an irregular supply of Keywords antimicrobials, bacteriocins, psychrophiles, psychrotolerant, Serratia proteamaculans. Correspondence Osvaldo Delgado. PROIMI, Av. Belgrano y Pje. Caseros (4000) Tucuman. Argentine. E-mail: [email protected] 2009 ⁄ 2139: received 13 December 2009, revised 10 March 2010 and accepted 10 March 2010. doi:10.1111/j.1365-2672.2010.04720.x Abstract Aim: This study focuses on the production, purification and characterization of serraticin A, a novel cold-active antimicrobial produced by Serratia proteamac- ulans 136. Methods and Results: A Ser. proteamaculans strain producing a novel cold- active antimicrobial was isolated from Isla de los Estados, Argentina. Antimi- crobial production was optimized in a BIOFLO 101 bioreactor under batch culture mode, with temperature, pH and dissolved oxygen controlled condi- tions. A purification protocol was developed including activated charcoal adsorption, solid-phase C18 extraction (SPE) and semi-preparative HPLC. The molecular weight was determined by LC ⁄ QTOF ⁄ MS ⁄ MS mass analysis. Conclusions: Serratia proteamaculans 136 produces a cold-active low molecular bacteriocin-like compound named serraticin A. In this work, it has been labo- ratory-scale produced, purified and partially characterized. Cross-immunity test revealed that serraticin A is very different from other well-known microcins assayed, with a wide inhibitory spectrum, showing an interesting biotechnology potential to be applied as a control agent against pathogenic bacteria. Significance and Impact of the Study: The present study is the first report of a cold-active compound with antimicrobial activity from Ser. proteamaculans. The work also highlights that cold environments could be a suitable source of micro-organisms with ability to produce cold-active biomolecules of biotechno- logical interest. Journal of Applied Microbiology ISSN 1364-5072 ª 2010 The Authors Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology 1

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL ARTICLE
Production, purification and characterization of serraticin A,a novel cold-active antimicrobial produced by Serratiaproteamaculans 136L.A. Sanchez1, M. Hedstrom2, M.A. Delgado3 and O.D. Delgado1
1 PROIMI – CONICET, Chacabuco, Tucuman, Argentina
2 Department of Biotechnology, Center for Chemistry and Chemical Engineering, Lund University, Lund, Sweden
3 INSIBIO – CONICET, Chacabuco, Tucuman, Argentina
Introduction
The need to decrease the use of chemical additives in
food has triggered the search for natural antimicrobial
substances produced by micro-organisms from different
sources. Application of antimicrobial compounds has
been well documented as a biotechnological advantage
over different industrial sectors like food-processing,
pharmaceutical chemistry and cosmetic industries. Conse-
quently, screening and selection programs for antimicro-
bial producing micro-organisms and bioproducts have
increased in the last years, particularly focusing into
improving the process control as well as the quality and
safety of final products by inhibition of both, undesirable
pathogenic and spoilage bacteria (Zahner and Fielder
1995; Davies and Webb 1998).
Since the introduction of penicillin in the 1940s, antibi-
otics have a history of success in controlling morbidity
and mortality caused by infectious diseases. However, as a
consequence of frequent use, bacterial resistance to
known classes of antibiotics has become a severe global
problem in recent years and presents a continuous chal-
lenge (Pohlmann et al. 2005). Nowadays, it cannot be
ignored that the antibiotic age is under adverse circum-
stances. The high levels of antibiotic resistance among
important pathogens along with an irregular supply of
Keywords
antimicrobials, bacteriocins, psychrophiles,
psychrotolerant, Serratia proteamaculans.
Correspondence
Osvaldo Delgado. PROIMI, Av. Belgrano y Pje.
Caseros (4000) Tucuman. Argentine.
E-mail: [email protected]
2009 ⁄ 2139: received 13 December 2009,
revised 10 March 2010 and accepted 10
March 2010.
doi:10.1111/j.1365-2672.2010.04720.x
Abstract
Aim: This study focuses on the production, purification and characterization of
serraticin A, a novel cold-active antimicrobial produced by Serratia proteamac-
ulans 136.
Methods and Results: A Ser. proteamaculans strain producing a novel cold-
active antimicrobial was isolated from Isla de los Estados, Argentina. Antimi-
crobial production was optimized in a BIOFLO 101 bioreactor under batch
culture mode, with temperature, pH and dissolved oxygen controlled condi-
tions. A purification protocol was developed including activated charcoal
adsorption, solid-phase C18 extraction (SPE) and semi-preparative HPLC. The
molecular weight was determined by LC ⁄ QTOF ⁄ MS ⁄ MS mass analysis.
Conclusions: Serratia proteamaculans 136 produces a cold-active low molecular
bacteriocin-like compound named serraticin A. In this work, it has been labo-
ratory-scale produced, purified and partially characterized. Cross-immunity test
revealed that serraticin A is very different from other well-known microcins
assayed, with a wide inhibitory spectrum, showing an interesting biotechnology
potential to be applied as a control agent against pathogenic bacteria.
Significance and Impact of the Study: The present study is the first report of a
cold-active compound with antimicrobial activity from Ser. proteamaculans.
The work also highlights that cold environments could be a suitable source of
micro-organisms with ability to produce cold-active biomolecules of biotechno-
logical interest.
Journal of Applied Microbiology ISSN 1364-5072
ª 2010 The Authors
Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology 1

novel classes of antibiotics, and the reduced number of
pharmaceutical companies involved in the discovery and
development of such compounds reveals an emergency
state into this scope (Wenzel 2004). The rate for drug dis-
covery ⁄ resistance has decreased in the latest 20 years with
evidence for novel resistance levels among pathogens such
as vancomycin-resistant enterococci, methicillin-resistant
and vancomycin-resistant Staphylococcus aureus and
multidrug-resistant Pseudomonas aeruginosa (Song 2008).
This current increase in drug resistance among several
common pathogenic bacteria highlights the necessity to
discover novel active compounds (Zahner and Fielder
1995; Davies and Webb 1998). Most new classes of anti-
biotics have arisen by testing natural sources (Gootz
1990), classically including several microbiological tools
like the isolation of novel micro-organisms; or otherwise
by means of the modification of well-known antibiotic-
producing micro-organisms as well as by metabolic path-
way diversion by engineering fermentation process (Souza
et al. 1982). The possibility to achieve more precise adap-
tations for a given industrial process, like low-temperature
antibiotic activity may contribute a significant biotechno-
logical improvement; with this aim, extremophilic micro-
organisms have been relatively unexplored group with
respect to mesophilic ones. Evidences from their study are
a thermophilic bacteriocin producing Bacillus (Kabuki
et al. 2007); a heat-labile b-lactamase purified from the
psychrophile Psychrobacter immobilis (Feller et al. 1997),
several archaea able to produce archaeocins which inhibit
closely related species (Aravalli et al. 1998; Prangishvili
et al. 2000) and recently described psychrophiles with
ability to produce antimicrobials at low temperatures
(O’Brien et al. 2004; Sanchez et al. 2009). Hence, the
extremophiles seem to exhibit a potential whose study is
growing day by day. This article introduces to the produc-
tion parameters, purification, biochemical characterization
and some important properties of serraticin A.
Materials and methods
Bacterial strains
Bacterial strains used in this work were obtained from
clinical isolations and collections (Table 1); Ser. protea-
maculans 136, natural antimicrobial producer, was
isolated in our laboratory (Sanchez et al. 2009).
Cross-streaking test
Cross-immunity or cross-inhibition against known micro-
cin producers was determined (Table 2) involving cross-
testing by the deferred (delayed) antagonism method
(Tagg et al. 1976).
Effect of Temperature, pH and media composition on
antimicrobial production
To determine the optimal medium for antimicrobial
production, different media compositions were evaluated:
LB, M9, M63 (Miller 1972) and R modified medium
(Reasoner and Geldreich 1985). Solid and soft media
contained 1Æ5 and 0Æ8% agar respectively and, when appro-
priate, l-amino acids (25 lg ml)1) were added.
Cell-free supernatants were evaluated for antimicrobial
production after 72 h incubation at 8�C. Optimal temper-
ature and pH values for antimicrobial production were
determined by incubating liquid cultures at both, differ-
ent temperatures and pH values.
Antimicrobial activity was determined by the critical
dilution method (Mayr-Harting et al. 1972), using Salmo-
nella enterica ser. Newport as indicator strain. Activity is
expressed either as antibiotic arbitrary units per millilitre
Table 1 Bacterial strains used in this study. Known microcin (Mcc)
producers are also listed
Strains Reference ⁄ Source
Shigella flexneri ATCC 29508
Staphylococcus aureus ATCC 29213;
ATCC 25923
Micrococcus luteus ATCC 10240
Bacillus subtilis 168 BGSC 1A1
Staphylococcus epidermidis ATCC 10390
Salmonella enterica
serovar Newport
Delgado et al. (2006)
Salmonella enterica
serovar typhimurium
Delgado et al. (2006)
Listeria monocytogenes ATCC 13932
Shigella sonnei Delgado et al. (2006)
Escherichia coli AB1133 CGSC
E. coli ATCC 35218;
ATCC 25922
E. coli O157:H7 (stx1,stx2);
O26:H11; O15:NM
Sanchez et al. (2009)
Citrobacter freundii ATCC 14135
Enterobacter aerogenes ATCC 10699
Enterobacter cloacae Sanchez et al. (2009)
Pseudomonas aeruginosa ATCC 27853
Salmonella enteritidis Delgado et al. (2006)
Proteus mirabilis Sanchez et al. (2009)
Proteus vulgaricus Sanchez et al. (2009)
Klebsiella pneumoniae
(Mcc E492)
De Lorenzo and
Pugsley (1985)
E. coli (Mcc J25) Salomon and Farias (1999)
E. coli (Mcc H47) Lavina et al. (1990)
E. coli (Mcc C7 ⁄ C51) Garcıa-Bustos et al. (1984);
Kurepina et al. (1993)
E. coli (Mcc V) Waters and Crosa (1991)
E. coli (Mcc B17) Baquero et al. (1978)
Serratia proteamaculans
136 (serraticin A)
This work
Serraticin A, a novel cold-active antimicrobial L.A. Sanchez et al.
2 Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology
ª 2010 The Authors
LEANDRO SANCHEZ
Highlight
(AU ml)1) or as micrograms per millilitre (lg ml)1)
(Sanchez et al. 2009).
Shake-flasks and batch cultivations
Shake-flask cultivations were conducted in 1-L Erlen-
meyer flasks, containing 300 ml of LB broth and inocu-
lated with 10% v ⁄ v of an overnight culture of
Ser. proteamaculans 136. Flasks were incubated at 8�C in
an orbital shaker at 200 rev min)1. A 7Æ5-l stirred tank
bioreactor (BioFlo 110 Modular Benchtop Fermen-
tor ⁄ Bioreactor; New Brunswick Scientific, Edison, NJ,
USA), with a working volume of 5 l (LB broth) and
10% inoculum was used. Agitation consisted of two
6-bladed Rushton-type impellers (52 mm), operating at
200 rev min)1. The culture pH was controlled at 7Æ0(initial pH was 6Æ8), while temperature was maintained
at 8�C. Air was injected at 0Æ5 v.v.m. flow rate. Growth
kinetic of both, shake-flasks and bioreactor was fol-
lowed by OD600nm measurements and antimicrobial
production was assayed along cultivation against
Salm. enterica ser. Newport as indicator strain. The cells
were removed by centrifugation at 8000 g at 4�C. Cell-
free supernatants were maintained at 4�C for further
antimicrobial purification.
Antimicrobial purification
Cell-free supernatant was mixed with granular activated
charcoal (6 g l)1), stirred and filtered through chromato-
graphy paper. Charcoal-bonded pigments from culture
medium were removed by resuspending in 300 ml of
30% (v ⁄ v) acetone and filtered as mentioned earlier. The
target material was suspended in 100 ml of 80% (v ⁄ v)
acetone and filtered. Acetone was evaporated from eluate
and original volume was corrected by water addition. The
antimicrobial was further adsorbed on a C18 Sep-Pak
SPE cartridge (Varian, Inc., Palo Alto, CA, USA) and
eluted by using 50% (v ⁄ v) acetonitrile. Further purifica-
tion steps were carried out by RP-HPLC by using a semi-
preparative C18 l Bondapak HPLC column (125A pore
size, 10l granulometry). A 0–100% water-acetonitrile
gradient (containing 0Æ1% TFA) at a flow rate of 6 ml
min)1 was established during 50 min, and optical density
was monitored by a 2998 Waters PDA detector (Waters
Co., Milford, MA, USA). Active fractions were pooled
and concentrated. Purity of the compound was re-evalu-
ated by analytical HPLC on the C18 column using similar
conditions. Protein concentration was assayed after each
purification step by using a Bicinchoninic acid protein kit
(Sigma Co., St Louis, MO, USA).
Minimal inhibitory concentration
Minimal inhibitory concentration (MIC) of the antimi-
crobial against different micro-organisms was determined
by the two-fold serial dilution assay (Yaron et al. 2003).
MIC was considered as the lowest concentration that
showed no turbidity increments (OD600nm) after 48 h of
incubation.
Antimicrobial mode of action on sensitive cell viability
Escherichia coli AB1133 and Salm. enterica ser. Newport
were exponentially grown in LB broth and to both
cultures, antimicrobial was added at MIC. The inhibition
of growth because of antimicrobial addition was followed
by OD600nm measurements at appropriate intervals of time.
To evaluate indicator strains viability, cells were counted
by the plate count method on LB and LB + antimicrobial
agar plates.
Antimicrobial effects on sensitive cell morphology
To establish an antimicrobial effect on sensitive cells
morphology, flask containing 10 ml of LB medium was
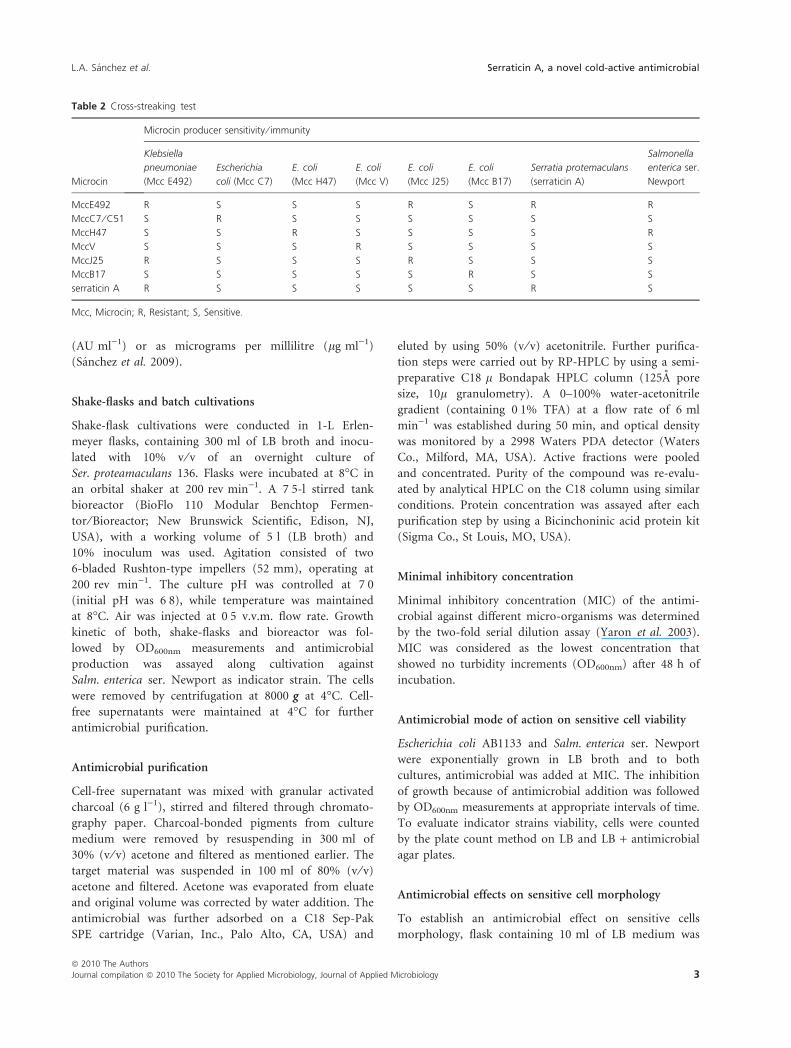
Table 2 Cross-streaking test
Microcin
Microcin producer sensitivity ⁄ immunity
Klebsiella
pneumoniae
(Mcc E492)
Escherichia
coli (Mcc C7)
E. coli
(Mcc H47)
E. coli
(Mcc V)
E. coli
(Mcc J25)
E. coli
(Mcc B17)
Serratia protemaculans
(serraticin A)
Salmonella
enterica ser.
Newport
MccE492 R S S S R S R R
MccC7 ⁄ C51 S R S S S S S S
MccH47 S S R S S S S R
MccV S S S R S S S S
MccJ25 R S S S R S S S
MccB17 S S S S S R S S
serraticin A R S S S S S R S
Mcc, Microcin; R, Resistant; S, Sensitive.
L.A. Sanchez et al. Serraticin A, a novel cold-active antimicrobial
ª 2010 The Authors
Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology 3

inoculated with approximately 108 cells of either E. coli
AB1133, ATCC 35218, DH5a or Salm. enterica ser.
Newport sensitive strains and incubated 2 h at 37�C, after
that, purified antimicrobial was added at MIC and
incubation was continued. After 12 h of culturing, cell
morphology was evaluated regarding to the control with
no antimicrobial addition. Sensitive strain cell morphology
was examined on a Nikon Eclipse 80i (Nikon GmbH,
Dusseldorf, Germany) microscope at · 1000 magnification.
Haemolytic, bio-emulsifier and bio-surfactant activities
Antimicrobial haemolytic activity was evaluated on blood
agar-plates (Banat 1993); LB soft-agar medium was sup-
plemented with 2% (v ⁄ v) of a fresh erythrocyte suspen-
sion (Moran et al. 2002). Ten microlitres of two-fold
serial dilutions of the antimicrobial stock solution
(500 lg ml)1) was prepared in physiological solution (pH
7Æ2) and deposited over LB-blood agar plates and incu-
bated at 37�C during 3 days. Staph. aureus ATCC 29213
was used as positive haemolytic control (Cooper et al.
1964).
Antimicrobial emulsifier property was also evaluated
according to Moran et al. (2000): 2 ml of a two-fold serial
dilution was prepared as described earlier, mixed with
3 ml of kerosene in a test tube and vortexed for 2 min.
After 24 h, the emulsified kerosene proportion was evalu-
ated. In the same way, the evaluation for bio-surfactant
activity was carried out by the drop-collapse test (Jain
et al. 1991) by using mineral, olive and corn oil.
Fluorescence spectrometry
To gain insight into the antimicrobial nature, fluorescence
of antimicrobial was measured. A purified solution
(5 lg ml)1) was prepared in sterile water, and fluores-
cence was measured in a PerkinElmer LS-55 fluorescence
spectrometer (PerkinElmer, Waltham, MA, USA), using
emission ⁄ excitation wavelengths associated with aromatic
aminoacids: tryptophan (kem = 348 nm; kex = 280 nm);
phenylalanine (kem = 282 nm; kex = 257 nm); and tyro-
sine (kem = 303 nm; kex = 274 nm).
Mass determination
Molecular mass of the compound was determined on a
Hybrid Quadrupole-TOF LC ⁄ MS ⁄ MS Mass Spectrometer
using a QSTAR� hybrid pulsar-i instrument (Applied
Biosystems, Foster City, CA, USA). Prior to, and in
sequence with MS, the sample was separated on RP
(C18)-HPLC Kromasil column (Eka, Sweden). Twenty
microlitres of sample (approximately 0Æ05 mg ml)1) was
injected in a linear gradient of water ⁄ acetonitrile at a
starting point of 80% water. The gradient was allowed to
proceed for 30 min to a final amount of 10% water. The
electrospray ionization (ESI) TurboIonSpray� source was
set to positive ion mode with a source voltage of +5500 V.
The quadrupole system was adjusted to scan between m ⁄ z50–3000 in TOF-MS mode, whereas for product ion mode
(i.e. MS ⁄ MS) a range of m ⁄ z 50–2000 was chosen. The
m ⁄ z value of individual precursor ions was automatically
selected in the information-dependent acquisition software
feature for fragmentation and was collided under argon
pressure using rolling collision energies ranging from 12
to 60 eV (i.e. collision induced dissociation).
Results
Cross-streaking test
Serratia proteamaculans 136 isolation was previously
described (Sanchez et al. 2009), and broad inhibition
spectra against both, Gram (+) and Gram ()) were
observed, with E. coli O26:H11 and Enterobacter aerogenes
the only exceptions, in which growth was not inhibited.
Cross-streaking test showed that the antimicrobial isolated
inhibited the growth of several up to date microcin-
producers (Table 2) with the only exception of MccE492
producer, Klebsiella pneumoniae.
Effect of media composition, pH and temperature on
serraticin A production
A significant dependence on media composition for anti-
microbial production was observed when Ser. proteamacu-
lans 136 was grown in several media with a maximum of
800 AU ml)1 in LB broth, although approximately
400 AU ml)1 was detected in other tested media.
Production of the antimicrobial compound was
observed only at 4–20�C range, while nonproduction was
observed at 30–37�C in spite of growth. Culture pH did
not affect significantly the antimicrobial production; only
at pH 5Æ0 or lower, a significant decrease in antimicrobial
production was observed.
Culture condition effects on Serratia proteamaculans
growth and antimicrobial production
Growth kinetics and antimicrobial production were dif-
ferent in shake-flasks and bioreactor cultivations (Fig. 1).
In shake-flask scale, culture growth was slower (lmax =
0Æ11), and the activity was detected in supernatants after
84 h of cultivation with a maximum of 600 AU ml)1 after
130 h of cultivation; on the other hand, a lmax = 0Æ18
and 800 AU ml)1 were reached at bioreactor scale after
45 h of cultivation; whereas the antimicrobial was
Serraticin A, a novel cold-active antimicrobial L.A. Sanchez et al.
4 Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology
ª 2010 The Authors
LEANDRO SANCHEZ
Highlight
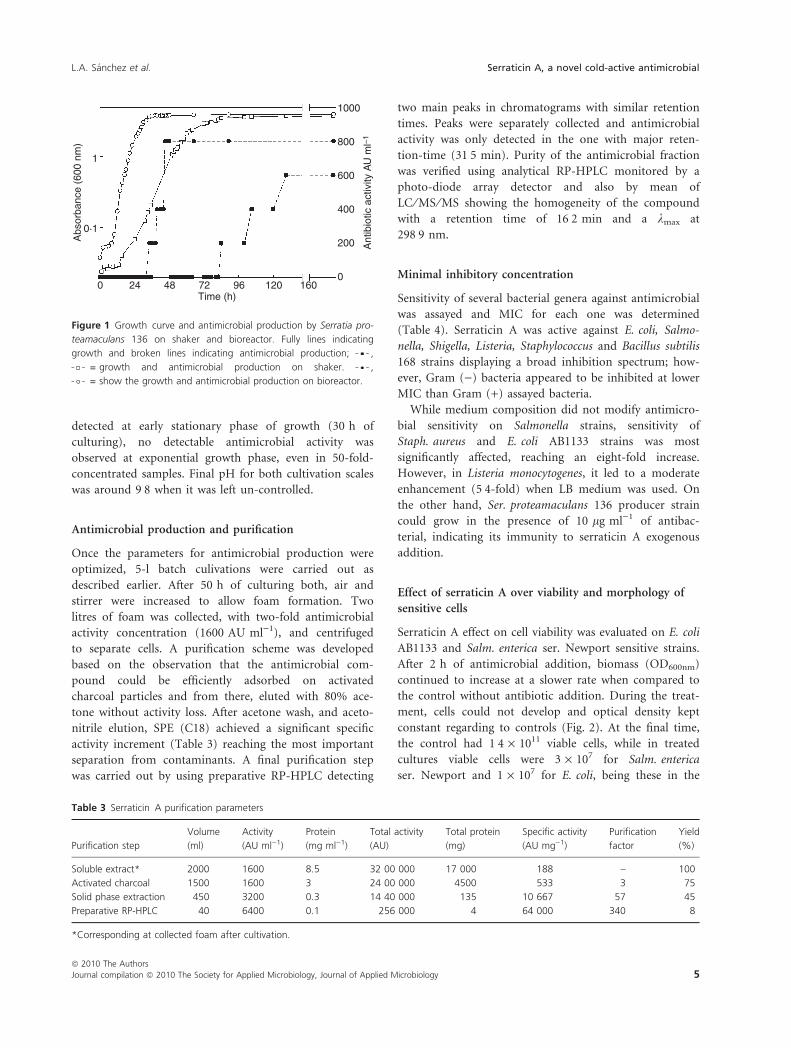
detected at early stationary phase of growth (30 h of
culturing), no detectable antimicrobial activity was
observed at exponential growth phase, even in 50-fold-
concentrated samples. Final pH for both cultivation scales
was around 9Æ8 when it was left un-controlled.
Antimicrobial production and purification
Once the parameters for antimicrobial production were
optimized, 5-l batch culivations were carried out as
described earlier. After 50 h of culturing both, air and
stirrer were increased to allow foam formation. Two
litres of foam was collected, with two-fold antimicrobial
activity concentration (1600 AU ml)1), and centrifuged
to separate cells. A purification scheme was developed
based on the observation that the antimicrobial com-
pound could be efficiently adsorbed on activated
charcoal particles and from there, eluted with 80% ace-
tone without activity loss. After acetone wash, and aceto-
nitrile elution, SPE (C18) achieved a significant specific
activity increment (Table 3) reaching the most important
separation from contaminants. A final purification step
was carried out by using preparative RP-HPLC detecting
two main peaks in chromatograms with similar retention
times. Peaks were separately collected and antimicrobial
activity was only detected in the one with major reten-
tion-time (31Æ5 min). Purity of the antimicrobial fraction
was verified using analytical RP-HPLC monitored by a
photo-diode array detector and also by mean of
LC ⁄ MS ⁄ MS showing the homogeneity of the compound
with a retention time of 16Æ2 min and a kmax at
298Æ9 nm.
Minimal inhibitory concentration
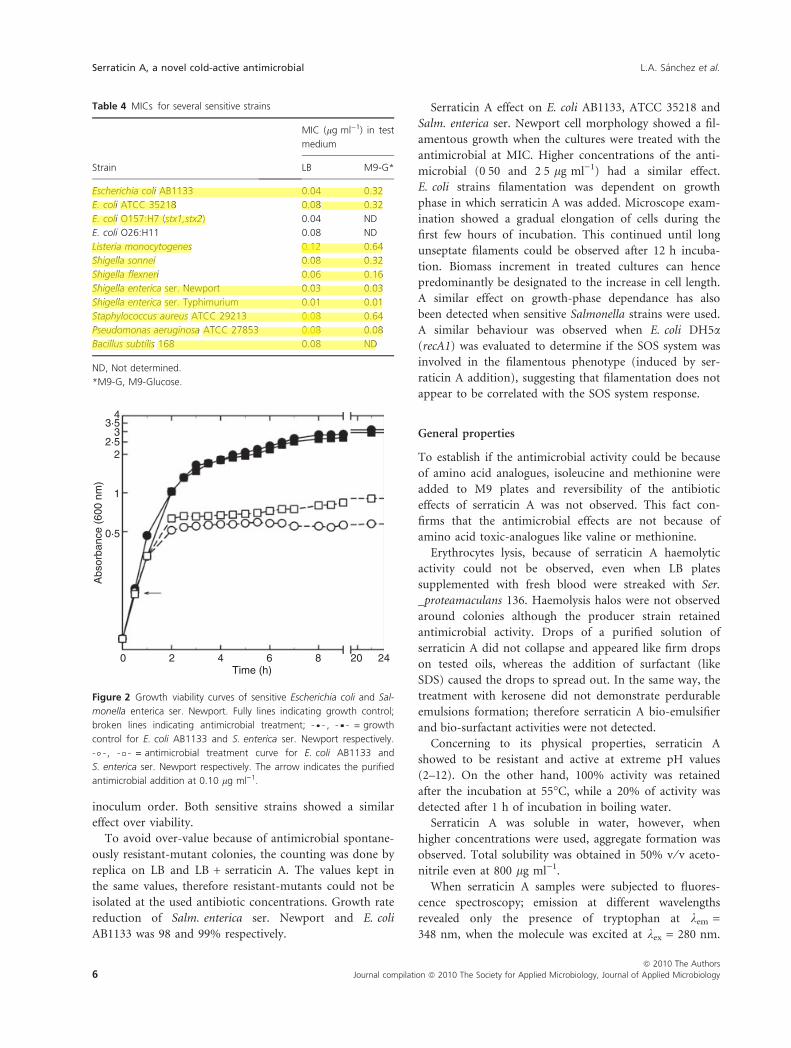
Sensitivity of several bacterial genera against antimicrobial
was assayed and MIC for each one was determined
(Table 4). Serraticin A was active against E. coli, Salmo-
nella, Shigella, Listeria, Staphylococcus and Bacillus subtilis
168 strains displaying a broad inhibition spectrum; how-
ever, Gram ()) bacteria appeared to be inhibited at lower
MIC than Gram (+) assayed bacteria.
While medium composition did not modify antimicro-
bial sensitivity on Salmonella strains, sensitivity of
Staph. aureus and E. coli AB1133 strains was most
significantly affected, reaching an eight-fold increase.
However, in Listeria monocytogenes, it led to a moderate
enhancement (5Æ4-fold) when LB medium was used. On
the other hand, Ser. proteamaculans 136 producer strain
could grow in the presence of 10 lg ml)1 of antibac-
terial, indicating its immunity to serraticin A exogenous
addition.
Effect of serraticin A over viability and morphology of
sensitive cells
Serraticin A effect on cell viability was evaluated on E. coli
AB1133 and Salm. enterica ser. Newport sensitive strains.
After 2 h of antimicrobial addition, biomass (OD600nm)
continued to increase at a slower rate when compared to
the control without antibiotic addition. During the treat-
ment, cells could not develop and optical density kept
constant regarding to controls (Fig. 2). At the final time,
the control had 1Æ4 · 1011 viable cells, while in treated
cultures viable cells were 3 · 107 for Salm. enterica
ser. Newport and 1 · 107 for E. coli, being these in the
0
0·1
1
1000
800
600
400
Ant
ibio
tic a
ctiv
ity A
U m
l–1
200
0
Abs
orba
nce
(600
nm
)
24 48 72 96Time (h)
120 160
Figure 1 Growth curve and antimicrobial production by Serratia pro-
teamaculans 136 on shaker and bioreactor. Fully lines indicating
growth and broken lines indicating antimicrobial production; ,
= growth and antimicrobial production on shaker. ,
= show the growth and antimicrobial production on bioreactor.
Table 3 Serraticin A purification parameters
Purification step
Volume
(ml)
Activity
(AU ml)1)
Protein
(mg ml)1)
Total activity
(AU)
Total protein
(mg)
Specific activity
(AU mg)1)
Purification
factor
Yield
(%)
Soluble extract* 2000 1600 8.5 32 00 000 17 000 188 – 100
Activated charcoal 1500 1600 3 24 00 000 4500 533 3 75
Solid phase extraction 450 3200 0.3 14 40 000 135 10 667 57 45
Preparative RP-HPLC 40 6400 0.1 256 000 4 64 000 340 8
*Corresponding at collected foam after cultivation.
L.A. Sanchez et al. Serraticin A, a novel cold-active antimicrobial
ª 2010 The Authors
Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology 5
inoculum order. Both sensitive strains showed a similar
effect over viability.
To avoid over-value because of antimicrobial spontane-
ously resistant-mutant colonies, the counting was done by
replica on LB and LB + serraticin A. The values kept in
the same values, therefore resistant-mutants could not be
isolated at the used antibiotic concentrations. Growth rate
reduction of Salm. enterica ser. Newport and E. coli
AB1133 was 98 and 99% respectively.
Serraticin A effect on E. coli AB1133, ATCC 35218 and
Salm. enterica ser. Newport cell morphology showed a fil-
amentous growth when the cultures were treated with the
antimicrobial at MIC. Higher concentrations of the anti-
microbial (0Æ50 and 2Æ5 lg ml)1) had a similar effect.
E. coli strains filamentation was dependent on growth
phase in which serraticin A was added. Microscope exam-
ination showed a gradual elongation of cells during the
first few hours of incubation. This continued until long
unseptate filaments could be observed after 12 h incuba-
tion. Biomass increment in treated cultures can hence
predominantly be designated to the increase in cell length.
A similar effect on growth-phase dependance has also
been detected when sensitive Salmonella strains were used.
A similar behaviour was observed when E. coli DH5a(recA1) was evaluated to determine if the SOS system was
involved in the filamentous phenotype (induced by ser-
raticin A addition), suggesting that filamentation does not
appear to be correlated with the SOS system response.
General properties
To establish if the antimicrobial activity could be because
of amino acid analogues, isoleucine and methionine were
added to M9 plates and reversibility of the antibiotic
effects of serraticin A was not observed. This fact con-
firms that the antimicrobial effects are not because of
amino acid toxic-analogues like valine or methionine.
Erythrocytes lysis, because of serraticin A haemolytic
activity could not be observed, even when LB plates
supplemented with fresh blood were streaked with Ser.
_proteamaculans 136. Haemolysis halos were not observed
around colonies although the producer strain retained
antimicrobial activity. Drops of a purified solution of
serraticin A did not collapse and appeared like firm drops
on tested oils, whereas the addition of surfactant (like
SDS) caused the drops to spread out. In the same way, the
treatment with kerosene did not demonstrate perdurable
emulsions formation; therefore serraticin A bio-emulsifier
and bio-surfactant activities were not detected.
Concerning to its physical properties, serraticin A
showed to be resistant and active at extreme pH values
(2–12). On the other hand, 100% activity was retained
after the incubation at 55�C, while a 20% of activity was
detected after 1 h of incubation in boiling water.
Serraticin A was soluble in water, however, when
higher concentrations were used, aggregate formation was
observed. Total solubility was obtained in 50% v ⁄ v aceto-
nitrile even at 800 lg ml)1.
When serraticin A samples were subjected to fluores-
cence spectroscopy; emission at different wavelengths
revealed only the presence of tryptophan at kem =
348 nm, when the molecule was excited at kex = 280 nm.
43·5
32·5
2
1
0·5
0 2 4 6Time (h)
8 20 24
Abs
orba
nce
(600
nm
)
Figure 2 Growth viability curves of sensitive Escherichia coli and Sal-
monella enterica ser. Newport. Fully lines indicating growth control;
broken lines indicating antimicrobial treatment; , = growth
control for E. coli AB1133 and S. enterica ser. Newport respectively.
, = antimicrobial treatment curve for E. coli AB1133 and
S. enterica ser. Newport respectively. The arrow indicates the purified
antimicrobial addition at 0.10 lg ml)1.
Table 4 MICs for several sensitive strains
Strain
MIC (lg ml)1) in test
medium
LB M9-G*
Escherichia coli AB1133 0.04 0.32
E. coli ATCC 35218 0.08 0.32
E. coli O157:H7 (stx1,stx2) 0.04 ND
E. coli O26:H11 0.08 ND
Listeria monocytogenes 0.12 0.64
Shigella sonnei 0.08 0.32
Shigella flexneri 0.06 0.16
Shigella enterica ser. Newport 0.03 0.03
Shigella enterica ser. Typhimurium 0.01 0.01
Staphylococcus aureus ATCC 29213 0.08 0.64
Pseudomonas aeruginosa ATCC 27853 0.08 0.08
Bacillus subtilis 168 0.08 ND
ND, Not determined.
*M9-G, M9-Glucose.
Serraticin A, a novel cold-active antimicrobial L.A. Sanchez et al.
6 Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology
ª 2010 The Authors
LEANDRO SANCHEZ
Highlight
LEANDRO SANCHEZ
Highlight
LEANDRO SANCHEZ
Highlight
LEANDRO SANCHEZ
Highlight
LEANDRO SANCHEZ
Highlight
LEANDRO SANCHEZ
Highlight
LEANDRO SANCHEZ
Highlight
LEANDRO SANCHEZ
Highlight
LEANDRO SANCHEZ
Highlight

In the same way, when samples were excited at kex = 274
and 257 nm; fluorescence was not observed at kem = 303
and 282 nm, suggesting tryptophan residues presence.
Mass spectrometry
The mass of the compound was evaluated by LC ⁄ ESI ⁄ Q-
TOF ⁄ MS ⁄ (MS), and spectra from both MS and MS ⁄ MS
(fragmentation pattern) were obtained. Purified serraticin
A subjected to MS revealed essentially three peaks with
identified mass-over-charges of 480Æ2, 959Æ4 and 1455Æ6,
the first one being the [M + H]+ ion form, hence with a
mass of the compound of 479Æ2 g mol)1. Other ions pres-
ent in the spectrum correspond to a dimer formation
[2M + H]+ = 959Æ4 m ⁄ z, and the mass of a multimeric
ion indicates that an additional ammonium molecule is
attached to the structure [3M + NH4]+ = 1455Æ6 m ⁄ z.
When the ions [2M + H]+ and [3M + NH4]+ were
subjected to MS ⁄ MS analysis, they showed the same
fragmentation pattern observed for the monomer. Also
the form [M + Na]+ = 502Æ2 m ⁄ z is conclusively identi-
fied with MS ⁄ MS.
Discussion
In a previous study, we have isolated a group of psychro-
philic and psychrotrophic micro-organisms from Isla de
los Estados-Ushuaia, Argentina which have the capability
to inhibit a wide range of pathogenic strains (Sanchez
et al. 2009). In this study, serraticin A, a cold-active anti-
microbial produced by Ser. proteamaculans 136 has been
produced, purified and partially characterized.
A cross-immunity test revealed that serraticin A is very
different from other well-known microcins assayed, show-
ing cross-immunity only with MccE492; however, they
differ in other important characteristics such as their
molecular weight, inhibition spectra, producer bacterium
and antimicrobial growth phase production. These
cross-sensitivity ⁄ inhibition, suggested us the novelty of
the antimicrobial discovered, named serraticin A. The
cross-immunity observed for both strains, could indicate
an affiliation between them or similarity in the cellular
structure where they are active. Several antibiotic-produc-
ing strains are sensitive to their own antibiotic in the
onset of antibiotic production (Martin and Demain
1980). From the cross-streak test, we have found that
Ser. proteamaculans 136 displayed a constitutive immu-
nity, as it was resistant to exogenous addition of high
concentrations of its own antibiotic. This fact is in accor-
dance with the statements for microcins, where genetic
studies show gene clusters coding immunity factors,
structural aminoacids and signal peptides (Duquesne
et al. 2007).
The isolated bacterium produced the antimicrobial
compound at low temperatures (4 and 8�C); while no
production was detected at higher temperatures (30, 37,
45�C), in spite of the fact that bacterium was able to
grow. This versatility demonstrates a compromise
between cold environments and antimicrobial production;
in recent years, more extensive studies have been carried
out about cold adaptation in both, prokaryote and
eukaryote organisms (Jones et al. 1987; Schindler et al.
1999; Yinghua et al. 2008; Schmid et al. 2009; Gocheva
et al. 2009).
Serratia proteamaculans 136 produced a heat-stable,
cold active, low molecular weight antimicrobial with a
broad inhibitory spectrum; being this, the first communi-
cation of this specie regarding antimicrobials production.
When different media were evaluated for serraticin A pro-
duction, our results indicated that in LB the maximum
production was obtained. This result differ significantly
from other well-known microcin producers, in which
maximum activity was obtained in a minimal or poor
media, and the role of ecological effectors in microbial
complex ecosystems has been postulated (Asensio 1976).
However, these data are in accordance with E. coli
producing MccC7 ⁄ C5 (Fomenko et al. 1996), the only
exception for which optimal antimicrobial production
was detected in rich media.
Serraticin A production was detected as soon as cul-
tures reached stationary phase of growth, this behaviour
was observed for other microcin synthesis, like MccB17
(Hernandez-Chico et al. 1986), MccC7 (Novoa et al.
1986) and MccJ25 (Salomon and Farias 1999) with the
exception of MccE492 which is produced in an active-
form during exponential phase of growth (Orellana and
Lagos 1996). In turn, several publications about this
dependence on growth-stage for synthesis are available for
other products of Serratia sp. producers (Braun and
Schmitz 1980; Givshov et al. 1988; Iusupova et al. 2002).
Hence, the relationship between serraticin A production
and the growth phase was strongly affected by dissolved
oxygen present in culture medium and indirectly the
antimicrobial production. The use of dissolved-oxygen
controlled fermentations to decrease the time of fermen-
tation from 82 to 32 h and also a significant increase in
serraticin A concentration up to 800 AU ml)1. This indi-
cates that antimicrobial synthesis could be associated with
an oxidative metabolic pathway. Similar results for bacte-
riocin production were previously reported for pediocin
SJ-1 (Schved et al. 1993), plantaricin C (Gonzales et al.
1994) and pediocin SA-1 (Anastasiadou et al. 2007).
An efficient method for antimicrobial recovery, based
on foam formation at stationary growth phase and
purification steps was elaborated. Literature describes the
peptide hydrophobicity as an important parameter which
L.A. Sanchez et al. Serraticin A, a novel cold-active antimicrobial
ª 2010 The Authors
Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology 7

has influence on foam stability as well as foaming prop-
erty studies of peptidic hydrolysates from several sources
(Popineau et al. 2002; Larre et al. 2006). The hydrophobic
nature of serraticin A clearly explains foam concentration,
activated charcoal and C-18 matrixes adsorption during
the purification steps.
The serraticin A antimicrobial activity because of
amino acid analogues was discarded, as the addition of
isoleucine, methionine and amino acid mixtures to a
minimal culture media did not antagonize the antibiotic
action. Evidences of natural isolates producing and
excreting amino acids which are toxic for E. coli K-12
family strains were previously reported (De Felice et al.
1979; Asensio 1976; Salomon and Farias 1999).
When sensitive cells treated with serraticin A were
microscopically observed, they seemed to be unseptated;
forming long filaments. This effect is commonly associ-
ated with a blocking of DNA replication and SOS system
response; an interruption in chromosome segregation or
inhibition on septation process (Martin and Demain
1980); similar data were reported for MccJ25 (Salomon
and Farias 1999).
Nowadays, it is well known that hydrophobic interac-
tions involving peptides and neutral lipid membranes of
erythrocytes play a significant role in the haemolytic
activity, while hydrophobic and charge interactions
between peptides and negatively charged lipid bacterial
membranes play an important role in antimicrobial acti-
vity (Matsuzaki 1999; Shai 2002). For this fact, haemolysis
provoked by serraticin A was evaluated showing no
haemolytic activity against fresh red blood cells at the
concentrations evaluated.
The purified fraction allows us to determine the mass
of the compound as 479Æ2 g mol)1. Noncovalent com-
plexes such as protonated dimeric ions [2M + H]+ and
multimeric ions [3M + NH4]+ was reported as a common
behaviour in LC ⁄ MS (Chen et al. 2007). Extensive
ESI-MS experiments designed to determine the underly-
ing principles in the formation of proton-bound dimer
ions and sodium-bridged dimer ions from different sub-
stances were reported (Hamilton and Chen 1988; Berthod
et al. 2005). Dimer and multimeric ion formation (959Æ4m ⁄ z, 1455Æ6 m ⁄ z) was observed; however, MS ⁄ MS spectra
come to a point in a single charged ion with 480Æ2 m ⁄ z.
Work is under way to characterize and to get sequence
and final structure of the antimicrobial compound, which
would help to clarify the mode of action of this antibiotic
compound over sensitive cells.
References
Anastasiadou, S., Papagianni, M., Filiousisa, G., Ambrosiadisa,
I. and Koidisa, P. (2007) Pediocin SA-1, an antimicrobial
peptide from Pediococcus acidilactici NRRL B5627:
Production conditions, purification and characterization.
Bioresour Technol 99, 5384–5390.
Aravalli, R.N., She, Q. and Garrett, R.A. (1998) Archaea and
the new age of microorganisms. Trends Ecol Evol 13,
190–194.
Asensio, C.. (1976) Molecular ecology. In Reflections on Bio-
chemistry ed. Kornberg, A., Horecker, B.L., Comudella, L.
and Oro, J.. pp. 235–240. Oxford: Pergamon Press.
Banat, I.M. (1993) The isolation of a thermophilic biosurfac-
tant producing Bacillus sp. Biotechnol Lett 15, 591–594.
Baquero, F., Bouanchaud, D., Martinez-Perez, M.C. and
Fernandez, C. (1978) Microcin Plasmids: a group of
extrachromosomal elements coding for Low-
Molecular-Weight antibiotics in Escherichia coli.
J Bacteriol 135, 342–347.
Berthod, A., Berthod, L. and Armstrong, D.W. (2005) Selectiv-
ity of a native b-cyclodextrin column in the separation of
catechins. J Liq Chromatogr Relat Technol 28, 1669.
Braun, V. and Schmitz, G. (1980) Excretion of a protease by
Serratia marcescens. Arch Microbiol 124, 55–61.
Chen, G., Zhang, L.K. and Pramanik, B.N. (2007) LC ⁄ MS:
theory, instrumentation and applications to small
molecules. In HPLC for Pharmaceutical Scientists ed.
Kazakevich, Y. and Lobrutto, R., pp. 281–346. New York:
John Wiley & Sons.
Cooper, L.Z., Madoff, M.A. and Weinstein, L. (1964) Hemoly-
sis of rabbit erythrocytes by purified Staphylococcal alpha–
toxin I. Kinetics of the lytic reaction. J Bacteriol 87,
127–135.
Davies, J. and Webb, V. (1998) Antibiotic resistance in bacte-
ria. In Emerging Infections ed. Krause, R.M. pp. 239–273.
New York: Academic Press.
De Felice, M., Levinthal, M., Iaccarino, M. and Guardiola, J.
(1979) Growth inhibition as a consequence of antagonism
between related amino acid: affect of valine in Escherichia
coli K-12. Microbiol Mol Biol Rev 43, 42–58.
De Lorenzo, V. and Pugsley, A.P. (1985) Microcin E492 a
Low-Molecular-Weight peptide antibiotic which causes
depolarization of the Escherichia coli cytoplasmic mem-
brane. Antimicrob Agents Chemother 27, 666–669.
Delgado, M., Mouslim, C. and Groisman, E.A. (2006) The
PmrA ⁄ PmrB and RcsC ⁄ YojN ⁄ RcsB systems control expres-
sion of the Salmonella O-antigen chain length determinant.
Mol Microbiol 60, 39–50.
Duquesne, S., Destoumieux-Garzon, D., Peduzzi, J. and
Rebuffat, S. (2007) Mirocins, gene-encoded antibacterial
peptides from enterobacteria. Nat Prod Rep 24, 708–734.
Feller, G., Zekhnini, Z., Lamotte-Brasseur, J. and Gerday, C.
(1997) Enzymes from cold-adapted microorganisms. The
class C ß-lactamase from the antarctic psychrophile
Psychrobacter immobilis A5. Eur J Biochem 244, 186–191.
Fomenko, D.E., Basyuk, E.I., Bezrukov, V.M., Volodin, A.A.,
Metlitskaya, A.Z. and Khmel, I.A. (1996) Identification
and expression of plasmid genes involved in synthesis of
Serraticin A, a novel cold-active antimicrobial L.A. Sanchez et al.
8 Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology
ª 2010 The Authors

microcin C51. Russ J Genet 32, 1149–1155. (Erratum
33:116, 1997).
Garcıa-Bustos, J.F., Pezzi, N. and Asensio, C. (1984) Microcin
7: purification and properties. Biochem Biophys Res
Commun 119, 779–785.
Givshov, M., Olsen, L. and Molin, S. (1988) Cloning and
expression in Escherichia coli of the gene for extracellular
phospholipase Al from Serratia liquefaciens. J Bacteriol 170,
5855–5862.
Gocheva, Y.G., Tosi, S., Krumova, E.T., Slokoska, L.S., Miteva,
J.G., Vassilev, S.V. and Angelova, M.B. (2009) Tempera-
ture downshift induces antioxidant response in fungi
isolated from Antarctica. Extremophiles 2, 273–281.
Gonzales, B., Arca, P., Mayo, B. and Suarez, J.E. (1994) Detec-
tion, purification and partial characterization of plantaricin
C, a bacteriocin produced by a Lactobacillus plantarum
strain of dairy origin. Appl Environ Microbiol 60, 2158–
2163.
Gootz, T.D. (1990) Discovery and development of new antimi-
crobial agents. Clin Microbiol Rev 1, 13–31.
Hamilton, J.A. and Chen, L. (1988) Crystal structures of inclu-
sion complexes of b-cyclodextrin with (S)-(+)- and (R)-
())-febpren. J Am Chem Soc 110, 4379.
Hernandez-Chico, C., San Millan, J.L., Kolter, R. and Moreno,
F. (1986) Growth phase and ompR regulation of transcrip-
tion of microcin B17 genes. J Bacteriol 167, 1058–1065.
Iusupova, D.V., Petukhova, E.V., Sokolova, R.B. and
Gabdrakhmanova, L.A. (2002) The accumulation of
proteins with chitinase activity in the culture media of the
parent and mutant Serratia marcescens strain grown in
the presence of mitomycin C. Mikrobiologiia 71, 635–638.
Jain, D.K., Collins-Thompson, D.L., Lee, H. and Trevors, J.T.
(1991) A drop-collapsing test for screening surfactant-pro-
ducing microorganisms. J Microbiol Methods 13, 271–279.
Jones, P.G., Van Bogelen, R.A. and Neidhardt, F.C. (1987)
Induction of proteins in response to low temperature
in Escheichia coli. J Bacteriol 169, 2092–2095.
Kabuki, T., Uenishi, H., Watanabe, M., Seto, Y. and Nakajima,
H. (2007) Characterization of a bacteriocin, Thermophilin
1277, produced by Streptococcus thermophilus SBT1277.
J Appl Microbiol 102, 971–980.
Kurepina, N.E., Basyuk, E.I., Metliskaya, A.Z., Zaitsev, D.A.
and Khmel, I.A. (1993) Cloning and mapping of genetic
determinants for microcin C51 production and immunity.
Mol Genet Genomics 241, 700–706.
Larre, C., Mulder, W., Sanchez-Vioque, R., Lazko, J., Berot, S.,
Gueguen, J. and Popineau, Y. (2006) Characterization and
foaming properties of hydrolysates derived from rapeseed
isolate. Colloids Surf B Biointerfaces 49, 40–52.
Lavina, M., Gaggero, C. and Moreno, F. (1990) Microcin H47,
a chromosome-encoded microcin antibiotic of Escherichia
coli. J Bacteriol 172, 6585–6588.
Martin, J.F. and Demain, A.L. (1980) Control of antibiotic
biosynthesis. Microbiol Mol Biol Rev 44, 230–251.
Matsuzaki, K. (1999) Why and how are peptide–lipid interac-
tions utilized for self-defense? Magainins and tachyplesins
as archetypes Biochem Biophys Acta – Biomembranes 1462,
1–10.
Mayr-Harting, A., Hedges, A. and Berkeley, R.C.W. (1972)
Methods for studying bacteriocins. In Methods Microbiol
7ª-7 ed. Norris, J.R. and Ribbons, D.W. pp. 315–422.
London ⁄ New York: Academic Press.
Miller, J.H. (1972) Cold Spring Harbor Laboratory. In Experi-
ments in Molecular Genetics. pp. 352–355. New York: Cold
Spring Harbor Laboratory Press.
Moran, A.C., Olivera, N., Commendatore, J., Esteves, J.L. and
Sineriz, F. (2000) Enhancement of hydrocarbon waste bio-
degradation by addition of a biosurfactant from Bacillus
subtilis O9. Biodegradation 11, 65–71.
Moran, A.C., Martinez, A.M. and Sineriz, F. (2002) Quantifica-
tion of surfactin in culture supernatants by hemolytic
activity. Biotechnol Lett 24, 177–180.
Novoa, M.A., Dıaz-Guerra, L., San Millan, J.L. and Moreno, F.
(1986) Cloning and mapping of the genetic determinants
for microcin C7 production and immunity. J Bacteriol 168,
1384–1391.
O’Brien, A., Sharp, R., Russell, N. and Roller, S. (2004)
Antarctic bacteria inhibit growth of food-borne microor-
ganisms at low temperatures. FEMS Microbiol Ecol 48,
157–167.
Orellana, C. and Lagos, R. (1996) The activity of microcin
E492 from Klebsiella pneumoniae is regulated by a micro-
cin-antagonist. FEMS Microbiol Lett 136, 297–303.
Pohlmann, J., Lampe, T., Shimada, M., Nell, P.G., Pernerstor-
fer, J., Svenstrup, N., Brunner, N., Schiffer, G. et al. (2005)
Pyrrolidindione derivatives as antibacterial agents with a
novel mode of action. Bioorg Med Chem Lett 15,
1189–1192.
Popineau, Y., Huchet, B., Larre, C. and Berot, S. (2002)
Foaming and emulsifying properties of fractions of gluten
peptides obtained by limited enzymatic hydrolysis and
ultrafiltration. J Cereal Sci 35, 327–335.
Prangishvili, D., Holz, I., Stieger, E., Nickell, S., Kristjansson,
J.K. and Zillig, W. (2000) Sulfolobicins, specific
proteinaceous toxins produced by strains of the extre-
mophilic archaeal genus Sulfolobus. J Bacteriol 182,
2985–2988.
Reasoner, D.J. and Geldreich, E.E. (1985) A new medium for
the enumeration and subculture of bacteria from potable
water. Appl Environ Microbiol 49, 1–7.
Salomon, R. and Farias, N. (1999) Microcin 25 a novel antimi-
crobial peptide produced by Escherichia coli. J Bacteriol
174, 7428–7435.
Sanchez, L.A., Gomez, F.F. and Delgado, O.D. (2009)
Cold-adapted microorganisms as a source of new antimi-
crobials. Extremophiles 13, 111–120.
Schindler, T., Graumann, P.L., Perl, D., Ma, S., Schmid, F.X.
and Marahiel, A. (1999) The family of cold shock proteins
L.A. Sanchez et al. Serraticin A, a novel cold-active antimicrobial
ª 2010 The Authors
Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology 9

of Bacillus subtilis stability and dynamics in vitro and in
vivo. J Biol Chem 274, 3407–3413.
Schmid, B., Klumpp, J., Raimann, E., Loessner, M.J., Stephan,
R. and Tasara, T. (2009) Role of cold shock proteins
(Csp) for growth of Listeria monocytogenes under cold and
osmotic stress conditions. Appl Environ Microbiol 75,
1621–1627.
Schved, F., Lalazar, A., Henis, Y. and Juven, B.J. (1993)
Purification, partial characterization and plasmid-
linkage of pediocin SJ-1, a bacteriocin produced by
Pediococcus acidilactici. J Appl Bacteriol 74,
67–77.
Shai, Y. (2002) From innate immunity to de-novo
designed antimicrobial peptides. Curr Pharm Des 8,
715–725.
Song, J.H. (2008) What’s new on the antimicrobial horizon?.
Int J Antimicrob Agents 32, S207–S213.
Souza, N.J., Ganguli, B.N. and Reden, J. (1982) Strategies in
the discovery of drugs from natural sources. Annu Rep
Med Chem 17, 301–310.
Tagg, J.R., Dajani, A.S. and Wannamaker, L.W. (1976) Bacte-
riocins of gram-positive bacteria. Microbiol Mol Biol Rev
40, 722–756.
Waters, V.L. and Crosa, J.H. (1991) Colicin V virulence plas-
mids. Microbiol Mol Biol Rev 55, 437–450.
Wenzel, R.P. (2004) The antibiotic pipeline challenges, costs,
and values. N Engl J Med 351, 523–526.
Yaron, S., Rydlo, T., Shachar, D. and Mor, A. (2003) Activity
of dermaseptin K4-S4 against foodborne pathogens.
Peptides 24, 1815–1821.
Yinghua, Z., Yuting, L. and Guicheng, H. (2008) Expression
of cold-shock-protein genes from Lactococcus lactis and
analysis of the cryoprotection function. Acta Microbiol Sin
48, 1203–1207.
Zahner, H. and Fielder, H.P. (1995) The need for new antibi-
otics: possible ways forward. In Fifty Years of Antimicro-
bials: Past Perspectives and Future Trends ed. Hunter, P.A.,
Darby, G.K. and Russell, N.J. pp. 67–83. SGM Symp 53
Cambridge: Cambridge University Press.
Serraticin A, a novel cold-active antimicrobial L.A. Sanchez et al.
10 Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology
ª 2010 The Authors
Related Documents











