Production of Trametes pubescens Laccase under Submerged and Semi-Solid Culture Conditions on Agro- Industrial Wastes Juan C. Gonzalez 1☯ , Sandra C. Medina 1☯ , Alexander Rodriguez 2 , Johann F. Osma 3 , Carlos J. Alméciga- Díaz 2* , Oscar F. Sánchez 1* 1 Chemical Engineering Department, Universidad de los Andes, Bogotá, Colombia, 2 Proteins Expression and Purification Laboratory, Institute for the Study of Inborn Errors of Metabolism, Pontificia Universidad Javeriana, Bogotá, Colombia, 3 CMUA, Department of Electrical and Electronics Engineering, Universidad de los Andes, Bogotá, Colombia Abstract Laccases are copper-containing enzymes involved in the degradation of lignocellulosic materials and used in the treatment of phenol-containing wastewater. In this study we investigated the effect of culture conditions, i.e. submerged or semi-solid, and copper supplementation on laccase production by Trametes pubescens grown on coffee husk, soybean pod husk, or cedar sawdust. The highest specific laccase activity was achieved when the culture was conducted under submerged conditions supplemented with copper (5 mM), and using coffee husk as substrate. The crude extracts presented two laccase isoforms with molecular mass of 120 (Lac1) and 60 kDa (Lac2). Regardless of the substrate, enzymatic crude extract and purified fractions behaved similarly at different temperatures and pHs, most of them presented the maximum activity at 55 °C and a pH range between 2 and 3. In addition, they showed similar stability and electro-chemical properties. At optimal culture conditions laccase activity was 7.69±0.28 U mg -1 of protein for the crude extract, and 0.08±0.001 and 2.86±0.05 U mg -1 of protein for Lac1 and Lac2, respectively. In summary, these results show the potential of coffee husk as an important and economical growth medium to produce laccase, offering a new alternative use for this common agro-industrial byproduct. Citation: Gonzalez JC, Medina SC, Rodriguez A, Osma JF, Alméciga-Díaz CJ, et al. (2013) Production of Trametes pubescens Laccase under Submerged and Semi-Solid Culture Conditions on Agro-Industrial Wastes. PLoS ONE 8(9): e73721. doi:10.1371/journal.pone.0073721 Editor: Melanie R. Mormile, Missouri University of Science and Technology, United States of America Received May 12, 2013; Accepted July 19, 2013; Published September 3, 2013 Copyright: © 2013 Gonzalez et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited. Funding: This work was supported in part by Pontificia Universidad Javeriana (Grant ID 3400) and the Chemical Engineering Department at Universidad de Los Andes. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. Competing interests: The authors have declared that no competing interests exist. * E-mail: [email protected] (OFS); [email protected] (CJAD) ☯ These authors contributed equally to this work. Introduction Lignin, cellulose, and hemicellulose are the major compounds present in plant residues. Among them, cellulose and hemicellulose can be decomposed by a large number of aerobic and anaerobic microorganisms through the action of hydrolytic enzymes [1]. Conversely, lignin biodegradation occurs at a lower rate than plant cell wall polysaccharides [2]. Certain fungi, mostly belonging to Basidiomycetes, such as white- and brown-rot fungi, are known to be able to degrade lignin from lignocellulosic biomass [3]. The extracellular enzymatic system responsible for lignin degradation consists of lignin peroxidase (LiP, E.C. 1.11.1.14), manganese-dependent peroxidase (MnP, E.C. 1.11.1.13), and laccase (para-benzene- diol: oxygen oxidoreductase, EC 1.10.3.2I) [2,3]. While LiP catalyzes the oxidation of non-phenolic aromatic compounds such as veratryl alcohol, and MnP mainly oxidizes phenolic compounds [2], laccase catalyzes the oxidation of phenolic substrates with the concomitant reduction of oxygen to water [4]. However, the number of substrates that laccase can oxidize might be extended by using low molecular mass mediators (i.e. hydroxybenzotriazole, violuric acid, 2,2',6,6'- tetramethylpiperidine-1-oxyl), which are oxidized by laccase to organic radicals intermediates that in turn act as redox mediators [5]. In general, laccases are glycosylated and copper-containing enzymes with a molecular mass between 60 and 80 kDa, and an isoelectric point (pI) between 3.0 to 6.0 [6]. The role of laccases in lignin and phenolic compound degradation has been evaluated in a large number of biotechnological applications such as dye degradation, bioremediation of some toxic chemical wastes (e.g. chlorinated aromatic compounds, polycyclic aromatic hydrocarbons, nitroaromatics, and pesticides), and biosensor developments [3,7]. In addition, laccases have been used in food industry for PLOS ONE | www.plosone.org 1 September 2013 | Volume 8 | Issue 9 | e73721

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Production of Trametes pubescens Laccase underSubmerged and Semi-Solid Culture Conditions on Agro-Industrial WastesJuan C Gonzalez1 Sandra C Medina1 Alexander Rodriguez2 Johann F Osma3 Carlos J Almeacuteciga-Diacuteaz2 Oscar F Saacutenchez1
1 Chemical Engineering Department Universidad de los Andes Bogotaacute Colombia 2 Proteins Expression and Purification Laboratory Institute for the Study ofInborn Errors of Metabolism Pontificia Universidad Javeriana Bogotaacute Colombia 3 CMUA Department of Electrical and Electronics Engineering Universidadde los Andes Bogotaacute Colombia
Abstract
Laccases are copper-containing enzymes involved in the degradation of lignocellulosic materials and used in thetreatment of phenol-containing wastewater In this study we investigated the effect of culture conditions iesubmerged or semi-solid and copper supplementation on laccase production by Trametes pubescens grown oncoffee husk soybean pod husk or cedar sawdust The highest specific laccase activity was achieved when theculture was conducted under submerged conditions supplemented with copper (5 mM) and using coffee husk assubstrate The crude extracts presented two laccase isoforms with molecular mass of 120 (Lac1) and 60 kDa (Lac2)Regardless of the substrate enzymatic crude extract and purified fractions behaved similarly at differenttemperatures and pHs most of them presented the maximum activity at 55 degC and a pH range between 2 and 3 Inaddition they showed similar stability and electro-chemical properties At optimal culture conditions laccase activitywas 769plusmn028 U mg-1 of protein for the crude extract and 008plusmn0001 and 286plusmn005 U mg-1 of protein for Lac1 andLac2 respectively In summary these results show the potential of coffee husk as an important and economicalgrowth medium to produce laccase offering a new alternative use for this common agro-industrial byproduct
Citation Gonzalez JC Medina SC Rodriguez A Osma JF Almeacuteciga-Diacuteaz CJ et al (2013) Production of Trametes pubescens Laccase underSubmerged and Semi-Solid Culture Conditions on Agro-Industrial Wastes PLoS ONE 8(9) e73721 doi101371journalpone0073721
Editor Melanie R Mormile Missouri University of Science and Technology United States of America
Received May 12 2013 Accepted July 19 2013 Published September 3 2013
Copyright copy 2013 Gonzalez et al This is an open-access article distributed under the terms of the Creative Commons Attribution License which permitsunrestricted use distribution and reproduction in any medium provided the original author and source are credited
Funding This work was supported in part by Pontificia Universidad Javeriana (Grant ID 3400) and the Chemical Engineering Department at Universidadde Los Andes The funders had no role in study design data collection and analysis decision to publish or preparation of the manuscript
Competing interests The authors have declared that no competing interests exist
E-mail ofsanchezmunaleduco (OFS) cjalmecigajaverianaeduco (CJAD)
These authors contributed equally to this work
Introduction
Lignin cellulose and hemicellulose are the majorcompounds present in plant residues Among them celluloseand hemicellulose can be decomposed by a large number ofaerobic and anaerobic microorganisms through the action ofhydrolytic enzymes [1] Conversely lignin biodegradationoccurs at a lower rate than plant cell wall polysaccharides [2]Certain fungi mostly belonging to Basidiomycetes such aswhite- and brown-rot fungi are known to be able to degradelignin from lignocellulosic biomass [3] The extracellularenzymatic system responsible for lignin degradation consists oflignin peroxidase (LiP EC 111114) manganese-dependentperoxidase (MnP EC 111113) and laccase (para-benzene-diol oxygen oxidoreductase EC 11032I) [23] While LiPcatalyzes the oxidation of non-phenolic aromatic compoundssuch as veratryl alcohol and MnP mainly oxidizes phenolic
compounds [2] laccase catalyzes the oxidation of phenolicsubstrates with the concomitant reduction of oxygen to water[4] However the number of substrates that laccase canoxidize might be extended by using low molecular massmediators (ie hydroxybenzotriazole violuric acid 2266-tetramethylpiperidine-1-oxyl) which are oxidized by laccase toorganic radicals intermediates that in turn act as redoxmediators [5] In general laccases are glycosylated andcopper-containing enzymes with a molecular mass between 60and 80 kDa and an isoelectric point (pI) between 30 to 60 [6]
The role of laccases in lignin and phenolic compounddegradation has been evaluated in a large number ofbiotechnological applications such as dye degradationbioremediation of some toxic chemical wastes (eg chlorinatedaromatic compounds polycyclic aromatic hydrocarbonsnitroaromatics and pesticides) and biosensor developments[37] In addition laccases have been used in food industry for
PLOS ONE | wwwplosoneorg 1 September 2013 | Volume 8 | Issue 9 | e73721
wine and beer stabilization fruit juice processing and differentfood-related biosensors [8]
Use of industrial and agricultural wastes for laccaseproduction by white-rot fungi is an effective way to reduce thecost of production [910] In addition laccase-mediateddelignification allows to increase the nutritional value of agro-industrial byproducts for animal feed or soil fertilizer Toaccomplish the transformation of these agro-industrial wastessubmerged and solid state cultures have been conducted(Table 1) Submerged fermentations are commonly used inindustrial process although they can present drawbacks due tophysical space and energetic and water requirements [11] Onthe other hand solid state cultures present several advantageslike mimicking the natural habitat in which the microorganismgrows reduced water activity that reduce microbialcontaminations and limited water consumption and equipmentsize Also it has been reported that solid-state cultures presenthigher volumetric yield less energy requirements and higherend-product stability and concentration than submergedcultures [1112] Nevertheless there are important issuesrelated to heat and mass transfer that must be overcome inorder to scale up a solid-state culture in addition to accruedestimation of the biomass and recovery of the end product [12]Semi-solid culture has been studied as an alternative toaddress some of the drawbacks presented by submerged andsolid-state cultures The increment of water activity in this typeof culture allows to improve the nutrient and end-productavailability and culture control [13] It has been establishedthat nature and moisture of the agro-industrial waste used for asolid-state culture are critical factors [12] The evaluation ofthese parameters among other process conditions ie culturetemperature pH and aeration has encouraged the screeningof several agro-industrial wastes for the production of differentenzymes In spite that several studies have been reported forthe production of laccase in semi-solid culture usingPhanerochaete chrysosporium few studies are referred to itsproduction in semi-solid culture using Trametes pubescens[14ndash16]
Agriculture is one of the major economic activities inColombia Thus there is a large production of agro-industrialbyproducts and a concern about their disposal In this studycoffee husk soybean pod husk and cedar sawdust wereevaluated as growth substrates for laccase production by Tpubescens CBS 69694 under submerged (SmC) or semisolidcultures (SSC) The effect of Cu+2 supplementation and culturemethod on laccase production was also evaluated Thedifferent crude extracts were purified and the obtained laccasefractions were partially characterized Results showed coffeehusk as an alternative for laccase production regardless to theculture mode This probably owe to its content of caffeine andtannins that might act as laccase inducer In addition thedependence of the laccase production yield and the culturemode showed to be substrate dependent
Table 1 Agro-industrial byproducts used as substrates forlaccase production
Agro-industrialbyproduct Culture Organism
Activity (U g-1
or UL-1) Substrate ReferenceSugarcanebagasse
SFPleurotus
ostreatus112plusmn28 ABTS [56]
SFPhanerochaete
chrysosporium252plusmn15 ABTS [56]
SFPycnoporus
cinnabarinus2 ABTS [57]
SmF T versicolor 410 ABTS [58]Coffee pulp SF Ganoderma sp 142 ABTS [59]
SFPleurotus
ostreatus1 SGZ [60]
SFPleurotus
pulmonarius12 SGZ [60]
Wheat bran SFPleurotus sp
IE137~145 SGZ [61]
SFPleurotus
pulmonarius
CCB1920000 SGZ [62]
SmFPleurotus
ostreatus 18048033 ABTS [63]
SmFGanoderma
lucidum 44797340plusmn9460 ABTS [64]
Bananapeels
SF Ganoderma sp 974 ABTS [59]
SF Pleurotus florida 54 ABTS [65]
SFLentinula edodes
12239031 ABTS [66]
SFTrametes hirsuta
BT 25663550 ABTS [67]
Mandarinpeels
SmFGanoderma
lucidum 4473145plusmn370 ABTS [64]
SF Pleurotus florida 31 ABTS [65]
SmFGanoderma
lucidum 44735980plusmn3616 ABTS [64]
SmFTrametes
pubescens100 ABTS [68]
Cantaloupepeels
SmFTrametes
pubescens IBB663
1084plusmn92 ABTS [67]
Soy bran SF Pleurotus florida 40 ABTS [65]Vinasse +Cotton stalkextract
SmFGanoderma
lucidum 44793840plusmn9566 ABTS [64]
Tomatopomace
SmFCoriolus versicolor
ATCC 2008014406 SGZ [69]
Grape stalks SmFFunalia trogii
ATCC 2008004880 SGZ [70]
Barley straw SFPleurotus
ostreatus15 ABTS [70]
Grapeseeds
SFTrametes
versicolor35 ABTS [71]
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 2 September 2013 | Volume 8 | Issue 9 | e73721
Table 1 (continued)
Agro-industrialbyproduct Culture Organism
Activity (U g-1
or UL-1) Substrate Reference
SmFTrametes
versicolor~800 ABTS [71]
Wheat straw SmFTrametes
versicolor~500 ABTS [71]
SmFTrametes
versicolor~500 ABTS [66]
Reed grass SFTrametes hirsuta
BT 256622550 ABTS [72]
Bean stalk SFLentinula edodes
12254867 SGZ [67]
Barley bran SFTrametes
pubescens IBB663
162plusmn20 ABTS [72]
Barley bran+ Cu+2
SFLentinula edodes
12225769 SGZ [66]
Apple peels SFTrametes hirsuta
BT 256615740 ABTS [67]
SFTrametes
pubescens IBB663
188plusmn20 ABTS [67]
Tree leaves SmFTrametes
pubescens IBB663
1680plusmn190 ABTS [67]
SFTrametes
pubescens IBB663
280plusmn23 ABTS [67]
Olive millwastewater
SmFTrametes
pubescens IBB663
834plusmn61 ABTS [67]
SFTrametes
pubescens IBB663
205plusmn25 ABTS [67]
Woodshavings
SmFTrametes
pubescens IBB663
630plusmn54 ABTS [73]
SmFPanus tigrinus
CBS 577794600plusmn98 26-DMP [73]
SFPanus tigrinus
CBS 577791309plusmn20 26-DMP [74]
SmFTrametes
versicolor CBS10029
451 ABTS [75]
Woodshavings +Cu+2
SFCeriporiopsis
subvermispora~05 NI [75]
SF Ganoderma sp ~500 ABTS [59]
SFTrametes trogii
MYA 28-11901 ABTS [76]
SF Solid state fermentation SmF Submerged fermentationLaccase activity expressed as UL- 1 for these solid state culturesSGZ Syringaldazine 26-DMP 26-dimethoxyphenol NI No indicated
Materials and Methods
Microorganism and agro-industrial substratesLaccase was produced by using the white-rot fungus T
pubescens CBS 69694 Soybean pod and coffee husk wereobtained from Corporacioacuten Colombiana de InvestigacioacutenAgropecuaria (CORPOICA) and a local coffee-processingcompany respectively The cedar sawdust was obtained froma local market in Bogota (Colombia) The different byproductswere treated with a 05 vv hypochlorite solution for 5 h Afterthat they were rinsed with distilled water until obtaining aneutral pH Finally the byproducts were oven dried at 60 degCuntil constant weight The soybean pod and coffee husk werecrushed to an average particle size of about 08 x 05 mmwhile cedar sawdust was sieved The portion retained by a No100 sieve (opening 015 mm) was used for laccase production
Fungal culture maintenance and laccase productionT pubescens was cultured on malt extract agar (MEA) plates
for 10 days at 30 degC Colonized agar plugs were used forsubculture maintenance every 15 days Independent culturesusing each of the mentioned byproducts were conducted toproduce laccase under SmC or SSC conditions Each culturewas conducted in 1000 mL Erlenmeyer containing 15 g of thedesired dry byproduct and 150 or 50 mL of basal medium forSmC or SSC respectively [basal medium composition per liter05 g glucose 2 g KH2PO4 025 g MgSO4middot7H2O 09 g(NH4)2SO4 01 g CaCl2 and 05 g KCl in a citrate buffer 20mM pH 45 supplemented with 05 g L-1 thiamine [17]] Eachculture flask was inoculated with three 10-mm plugs of activefungus cultured on MEA and incubated for 25 days at 30degCwithout agitation Culture supernatant samples of 05 mL werewithdrawn to monitor laccase activity The effect of Cu+2 onlaccase production was evaluated within a 0 to 5 mMconcentration range (supplied as copper sulphate) The coppersolution was added to 4-days-old cultures A control wasprepared by growing the fungus in the basal medium withoutany agro-industrial waste under the described cultureconditions All the assays were performed in triplicate
Laccase purification and characterizationLaccase purification and characterization was conducted
using the crude extract that showed the highest enzyme activityfor each substrate under the evaluated culture conditions (SmCor SSC) The crude extract was obtained by vacuum filtrationthrough paper Whatman No 1 followed by centrifugation at 4degC and 4000 rpm for 15 min serial vacuum filtration through045 and 022 microm polyether sulphone membranes (Pall CorpPort Washington NY USA) and ultrafiltration (UF) through a10 kDa cut-off membrane of regenerated cellulose (MilliporeBillerica MA USA) The retentate (~10 mL) was analyzed bylow-pressure chromatography with a fraction collector(BioLogicTM LPC Biorad NJ US) using a Q-Sepharosecolumn (Pharmacia Biotech 10 x 10 cm glass chromatographycolumn filled with 6 mL media) equilibrated with 50 mM sodiumacetate buffer pH 50 Elution was performed by a linear NaClgradient (~20 mL) from 0 to 05 M All procedures were carriedout at 4 degC Fractions corresponding to protein peaks were
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 3 September 2013 | Volume 8 | Issue 9 | e73721
collected and analyzed by sodium dodecyl sulfate-polyacrylamide electrophoresis gel (SDS-PAGE) under non-reducing conditions and without heat denaturation Staining ofthe electrophoresis gel was done with 5 mM 22rsquo-azino-bis(3-ethylbenzothiazoline-6) sulphonic acid (ABTS) in succinic acid(25 mM pH 45) to detect laccase activity Fractions thatrevealed the same protein profile and laccase activity werecombined Samples of the combine fractions were analyzed bySDS-PAGE under mentioned conditions As a negative controlwere used a sample of the ultrafiltrated fraction which did notreveal laccase activity along the previous purification stepBands that revealed laccase activity were identified as Lac1and Lac2 These bands were excised from the electrophoresisgel and analyzed through nanoLC-MSMS (Applied BiomicsHayward CA USA) Peptides and protein sequences werealigned by using Clustal Omega [18] at EBI web server andBlast at NCBI web server Alignments figures were generatedusing CLC Sequence Viewer (CLC bio Denmark)
Enzyme activity was determined spectroscopically by usingeither 22rsquo-azino-bis(3-ethylbenzothiazoline-6) sulphonic acid(ABTS ε420 = 36mM-1 cm-1 Sigma-Aldrich) or 4-hydroxy-35-dimethoxybenzaldehyde (Syringaldazine ε530 = 65mM-1 cm-1Sigma-Aldrich) [19ndash21] ABTS activity assay was carried out aspreviously described [2122] Briefly 850 microL of 5 mM ABTS insuccinic acid (25 mM pH 45) were mixed with 150 microL of therespective dilution of culture extract and the absorbance was at420 nm Syringaldazine activity assay was carried out followingthe substrate manufacturerrsquos instructions (Sigma-Aldrich) Oneunit (U) was defined as the amount of enzyme required tooxidize 1 micromol of substrate (ABTS or syringaldazine) perminute Specific activity was expressed as U mgminus1 of protein asdetermined by Lowry assay [23]
Laccase production during the fungus culture was monitoredusing syringaldazine which has demonstrated to be a specificsubstrate for detecting extracellular laccase in the absence ofhydrogen peroxide [24] Absence of hydrogen peroxide wasconfirmed by incubation with catalase Crude extracts andpurified fractions were analyzed by SDS-PAGE withoutreduction by β-mercaptoethanol and heat denaturationProteins were visualized by silver staining while laccases wereidentified by placing the SDS-PAGE gel in an ABTS solution for5 min Temperature and pH effect on laccase activity wasdetermined for the UF retentate and the purified fractions byusing ABTS The temperature effect on laccase activity wasevaluated at 30 40 50 55 60 and 70 degC at pH 50 and the pHeffect on laccase activity was evaluated at 2 25 3 4 5 6 65and 7 at 55 degC Laccase stability was evaluated by using ABTSat optimal pH and temperature for 48 h
Cyclic voltammetry analysisCyclic voltammetry (CV) analysis was performed on a
computer-controlled potentiostat (Autolab PotentiostatGalvanostat PGSTAT 128N Metrohm USA) Data wereacquired by using the software Nova version 16 Aconventional three-electrode system was used to carry out thecyclic voltammograms The used electrodes were graphite-working electrode with a surface area of 023 cm2 AgAgCl KClsaturated reference electrode (Metrohm USA) and platinum
wire counter electrode Before running each experiment thesurface of the graphite electrode was polished with aluminaand thoroughly rinsed with distilled water CV analysis wasdone to the crude extract and purified laccase fractions thatpresented the highest activity for each substrate For the test20 microL of sample were added on the polished surface of theworking electrode and allowed to dry for 60 min at roomtemperature Each experiment was performed in triplicate usingeither a solution 11 of anhydride ethanol buffer acetate (100mM pH 50) and syringaldazine to a final concentration of 02mM or a solution of 100 mM acetate buffer (pH 30) and 05mM ABTS Cyclic voltammograms were carried out in 30 mL ofworking solution with continuous agitation Voltage wasscanned at 100 mV s-1 between -02 and 1 V vs Ag|AgCl|KClsat
for syringaldazine assays while for ABTS assays voltage wasscanned at 250 mV s-1 between -02 and 12 V vs Ag|AgCl|KClsat Assays were conducted in a Faraday cage at roomtemperature (20-22degC) Temperature and pH in the reactionsolution was monitored with the Unitrode with Pt 1000(Metrohm USA)
Statistical analysisAll experiments were conducted by triplicate Data are
presented as the mean plusmn standard deviation (SD) Statisticalcomparisons were conducted by using either t-test or Duncanrsquostest for one-factor design by using StatGraphics Centurionreg v16111 (2011) Statistical differences were established for p lt005
Results
SmC and SSC production of laccase by T pubescensCoffee husk soybean pod husk and cedar sawdust were
used as substrate for the production of laccase by Tpubescens under SmC or SSC conditions while syringaldazinewas used to monitor laccase production during the fungusculture under the studied conditions Hydrogen peroxide wasnot detected in any of the evaluated samples which discardthe interference of peroxidases in the syringaldazine assayLaccase activity and glucose concentration time course for Tpubescens grown on coffee husk under SmC and SSC atdifferent copper concentrations are presented in Figure 1Although a similar time course was observed for laccaseactivity and glucose concentration of soybean pod husk andcedar sawdust cultures differences on the day of maximumlacasse activity as well as in the maximum activity value wereobserved among the agro-industrial byproducts (Table 2) Ingeneral it was noticed that laccase activity was detectablearound the seventh and fourth day of culture for SmC andSSC respectively After the eighth culture day glucose wasdepleted from the culture medium and laccase activityincreased sharply
The substrate source seemed to be the most important factorfor laccase production (Table 2) T pubescens culture oncoffee husk allowed the maximum laccase activities for bothSmC (17723plusmn372 U L-1) and SSC (20400plusmn596 U L-1) atcopper concentrations of 5 and 0 microM respectively Laccaseactivity in SSC and SmC using coffee husk as substrate was
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 4 September 2013 | Volume 8 | Issue 9 | e73721
between 147- and 663-fold higher than that observed forcultures with soybean pod husk and cedar sawdust Laccaseactivity obtained in the control culture without agro-industrialwaste presented a maximum laccase activity of 222 plusmn 088 UL-1 after 14 days of culture while the glucose was depleted onthe seventh day of culture
Copper supplementation effect on laccase activity wasevaluated for both SmC and SSC Although in a non-linearmode the laccase activity was increased by the presence ofcopper with all substrates under SmC or SSC except withcoffee husks under SSC (Table 2) In addition the requiredcopper concentration to obtain the highest laccase activityvaried depending on the used agro-industrial waste Forexample laccase production was favored in cultures withsoybean pod husk at copper concentrations of 05 mM and 20mM for SmC and SSC respectively while for cultures with
Figure 1 Laccase activity profiles during T pubescensCBS 69694 culture under SmC (a) and SSC (b) on coffeehusk Culture media was enriched with different copperconcentrations 0 (Δ) 05 () 2 () and 5 (loz) mM and laccaseactivity was determined by using syringaldazine as substrateFilled symbols refer to the glucose concentration at therespective copper concentration All the assays wereperformed in triplicatedoi 101371journalpone0073721g001
coffee husk the highest laccase activity levels for SmC andSSC was observed at copper concentrations of 5 mM and 0mM respectively
Despite that culture mode ie SmC or SSC presented aneffect on laccase production this is substrate sensitive In theassays conducted without copper it was observed that laccaseactivity was 28- and 33-fold higher in SSC than in SmC for thesoybean pod husk and coffee husk respectively Howeverwhen cedar sawdust was used no significant difference inlaccase activity between the two culture types was obtained(Table 2) Due to the low volume of liquid media and highlaccase activity in SSC when compared to SmC the SSCpresented a higher volumetric yield For all agro-industrialwaste the SSC presented a shorter time than SmC to reachthe highest laccase activity which suggest a higher productivityunder SSC conditions
These results show that coffee husk under SmC or SSCconditions could be considered as a potential alternative forlaccase production by T pubescens However soybean podhusk and cedar sawdust could be also considered as importantalternatives for laccase production especially in comparisonwith synthetic culture media [10] Finally the optimal copperconcentration to obtain the highest laccase activity depends onthe agro-industrial byproduct used for the fungus culture
Laccase purification and characterizationLaccase purification was conducted using the crude extracts
that presented the highest activities For the SmC crude
Table 2 Effect of substrate copper concentration andculture mode on laccase activity (UL-1)
Substrate SmC SSCSoybean Pod Husk + 0 mM Cu+2 446 plusmn 138 (23) 1255 plusmn 052 (13)
Soybean Pod Husk + 05 mM Cu+2
8493 plusmn 846 (23)dagger 2028 plusmn 387 (21)
Soybean Pod Husk + 2 mM Cu+2 5761 plusmn 370 (19) 13785 plusmn 071 (18)dagger
Soybean Pod Husk + 5 mM Cu+2 4492 plusmn 487 (23) 2900 plusmn 407 (18)Coffee Husk + 0 mM Cu+2 6184 plusmn 176 (23) 20400 plusmn 596 (19)dagger
Coffee Husk + 05 mM Cu+2 13763 plusmn 568 (23) 11083 plusmn 940 (20)ab
Coffee Husk + 2 mM Cu+2 12304 plusmn 373 (23)b 10278 plusmn 814 (15)b
Coffee Husk + 5 mM Cu+2 17723 plusmn 372 (23)dagger 8266 plusmn 882 (15)Cedar Sawdust + 0 mM Cu+2 1355 plusmn 095 (17) 1082 plusmn 053 (15)
Cedar Sawdust + 05 mM Cu+2 2272 plusmn 084 (17) 1902 plusmn 002 (13)Cedar Sawdust + 2 mM Cu+2 2897 plusmn 127 (12)a 2638 plusmn 068 (8)Cedar Sawdust + 5 mM Cu+2 2936 plusmn 153 (13) abdagger 3073 plusmn 152 (12)bdagger
Data in parenthesis refers to the day where was observed the maximum laccaseactivity No Statistical Difference Statistical Difference (Evaluation of substrate effecton laccase activity for both culture modes submerged (SmC) and semisolid (SSC)no copper induction considered)Equal letters inside one agro-industrial waste group refers to No StatisticalDifference no letters refers to Statistical Difference inside the group (Evaluation ofcopper effect on laccase activity for both SmC and SSC)dagger Maximum laccase activity for the used agro-industrial wastes either in SmC orSSC culture
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 5 September 2013 | Volume 8 | Issue 9 | e73721
extracts obtained with soybean pod husk + Cu+2 05 mM coffeehusk + Cu+2 5 mM and cedar sawdust + Cu+2 5 mM wereselected while for SSC crude extracts obtained with soybeanpod husk + Cu+2 2 mM coffee husk + Cu+2 0 mM and cedarsawdust + Cu+2 5 mM were selected The electrophoreticanalysis of permeates from 045 and 022 microm filtrations and theUF retentate showed the presence of two proteins with laccaseactivity while no activity was observed within the ultrafiltratedsample (Figure 2a) These proteins presented a molecularmass of about 120 kDa and 60 kDa (Figure 2a) Thepurification of the UF retentate by ionic exchangechromatography confirmed the presence of two differentfractions with laccase activity (Figure 2b) The SDS-PAGEanalysis of these fractions under non-reducing conditionsshowed that one protein with laccase activity was presented oneach fraction (Figure 2b) and were named Lac1 (120 kDa) andLac2 (60 kDa) Considering that the conditions used tomeasure the enzyme activity were those suggested for laccaseactivity and that H2O2 or manganese were not added duringthe reaction for measuring the enzyme activity these resultssuggest that both proteins are laccases but not peroxidases
NanoLC-MSMS analysis of purified fractions showed thatthe identified peptides for Lac1 (SAGSTTYNYNDPIFR andRDTVSTGNSGDNVTIR) and Lac2 (SAGSTVYNYDNPIFR andANPNFGNVGFTGGINSAI) had identity scores between 80 to100 with laccases (over 100 sequences) available at the non-redundant GenBank dataset Proteins containing thesepeptides different that laccases were not identified Similarresults were observed when peptides were evaluated atUniprot Galhaup et al [17] described two laccase activefractions from T pubescens MB 89 (CBS 69694) Lap1(UniProtKBTrEMBL Q8TG93) and Lap2 (UniProtKBTrEMBLQ8TG94) which share a 70 identity (Figure 3a) Sequencealignment of Lac1 and Lac2 peptides with Lap1 and Lap2sequences respectively showed that the peptides arepresented within T pubescens MB 89 laccase sequencesthough some differences were observed for the Lac1 peptide(Figure 3b) This result suggests that the purified fractions arelaccase isoenzymes
Protein concentration and volumetric and specific laccaseactivities at different purification stages are presented in Table3 Lac1 presented lower laccase activity than Lac2 in a ratiothat ranges from 080- to 38-fold depending on the substratesource On the other hand laccase activity of the UF retentatefrom soybean pod husk culture was from 45- to 64-fold higherthan that of the purified fractions while for cedar sawdust andcoffee husk the laccase activity was from 46- to 120-fold andfrom 33- to 163-fold higher than the purified fractionsrespectively (Table 3) The purified Lac2 fractions from coffeehusk and cedar sawdust cultures showed the highest specificactivity levels (Table 3) The volumetric activities of UFretentates were higher than that obtained with purified fractions(Table 4)
Data in parenthesis shows the temperature or pH at whichrelative laccase activity reached 100
Temperature and pH effect on laccase activity wasdetermined for the UF retentate and Lac1 and Lac2 fractionsobtained along the purification of the enzymatic crude extracts
that resulted after the SSC of T pubescens on the selectedagro-industrial wastes Laccase activity for the UF retentateunder different temperature and pH conditions is presented inFigure 4 and Table 4 The maximum relative activity of theevaluated UF retentates was observed at 50-55 degC and at apH range from 20 to 40 However UF retentate from cedar
Figure 2 Purification of lacasses produced by submergedculture of T pubescens using coffee husk assubstrate All samples were analyzed by SDS-PAGE undernon-reducing conditions and stained with ABTS No heatdenaturation of the samples was conducted (a) The crudeextract (1) was centrifuged and filtered through a Whatman No1 filter (2) 045 microM membrane (3) 022 microm membrane (4) andultrafiltrated through a 10 kDa cut-off membrane permeate (5)and retentate (6) (b) UF retentate was purified by anionicexchange chromatography Lac1 represent the unboundfraction while Lac2 represent the eluted fraction The arrowshows the point when protein elution was started (c) Purifiedfractions (1) crude extract (2) Lac1 and (3) Lac2 MWMolecular Weight Similar results were observed for crudeextracts produced by using soybean pod husk and cedarsawdustdoi 101371journalpone0073721g002
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 6 September 2013 | Volume 8 | Issue 9 | e73721
sawdust culture behaved different to the UF retentate fromcoffee husk and soybean pod husk cultures In this sense the
UF retentate from cedar sawdust culture presented the lowestenzyme activity at temperatures le 40 degC but at 70 degC
Figure 3 Aligment of reported lacasses sequences and Lac1 and Lac2 peptides (a) Alignment of Lap1 (UniProtKBSwiss-Prot Q8TG93) and Lap2 (UniProtKBSwiss-Prot Q8TG94) from T pubescens CBS 69694 (b) Alignment of Lac1 and Lac2 peptideswith Lap1 and Lap2 respectively and tryptic peptides described by Shleev et al 2007 Alignments were carried out using ClustalOmega and figures were using CLC Sequence Viewer Different residues are colored in reddoi 101371journalpone0073721g003
Table 3 Laccase activity during the purification of crude extracts resulted from T pubescens culture on agro-industrialwastes under submerged (SmC) and semisolid (SSC) cultures
Soybean pod husk Coffee husk Cedar sawdust
Protein(mg mL-1)
VolumetricActivity (UL-1)
SpecificActivity (Umg-1
protein)Recovery()
Protein(mg mL-1)
VolumetricActivity (UL-1)
SpecificActivity (Umg-1
protein)Recovery()
Protein(mg mL-1)
VolumetricActivity (UL-1)
SpecificActivity (Umg-1
protein)Recover()
SmCCrudeExtract
0869 52888 0608 100 0414 152708 3688 100 0431 10735 0249 100
045 micromFiltration
0736 46549 0632 88 0334 117892 3529 77 0444 12053 0271 112
UF retentate 1010 78731 0779 74 0378 195263 5165 64 0510 25905 0507 120
Lac1 0494 5471 0110 1 0232 2399 0103 02 0222 838 0037 08
Lac2 0310 20661 0666 4 0116 107240 9252 7 0040 15234 3837 14 SSCCrudeExtract
1146 164966 1439 100 0596 154907 2601 100 1065 23203 0217 100
045 micromFiltration
0776 128392 1654 77 0474 115736 2439 75 0806 25824 0320 111
UF retentate 0973 335055 3443 101 0428 161750 3779 52 1134 51101 0450 110
Lac1 0097 8854 0912 2 0104 1514 0145 03 0042 1315 0313 2
Lac2 0168 48988 2915 10 0131 82298 6282 18 0072 16162 2244 23
740 ABTS was used to determine laccase activity
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 7 September 2013 | Volume 8 | Issue 9 | e73721
presented the highest enzyme activity ~90 of its initial valuein comparison to the other two UF retentates (Figure 4)
The temperature and pH effect on laccase activity for Lac1and Lac2 is presented in Figure 5 The maximum relativeactivity for both fractions was presented at a pH from 20 to 40regardless of the agro-industrial waste as observed for the UFretentate However at pH gt 40 Lac1 showed a sharperdecrease in its activity than that observed for Lac2Nevertheless Lac2 purified from cedar sawdust culturepresented a different performance This fraction showed themaximum activity at pH 50 while a sharp drop was observedat pHs above or below this value
Laccase activity profiles showed that Lac1 and Lac2presented the maximum enzyme activity at 55 and 60 degCrespectively (Table 4) It was observed that Lac1 from coffeehusk culture presented a lower decrease in enzyme activity attemperatures le 50 degC or ge 60 degC than Lac2 On the contraryLac2 from soybean husk or cedar sawdust cultures presenteda higher relative laccase activity than Lac1 at temperatures le55 degC The purified fractions from soybean pod husk and cedarsawdust cultures presented the lowest relative activity at 30 degCand 65 degC
Table 4 Optimum temperatures and pHs for maximumlaccase activities in UF retentate Lac1 and Lac2 obtainedfrom semisolid cultured T pubescens on agro-industrialwastes
EnzymaticFraction Temperature pH
VolumetricActivity (UL-1)
SpecificActivity (Umg-1 protein)
VolumetricActivity (UL-1)
SpecificActivity (Umg-1
protein)SoybeanPod Husk +2 mM Cu+2
148000plusmn1178(55)
1521plusmn0012182037plusmn3025(2-3)
1871plusmn0031
Lac122925plusmn917(60)
2363plusmn009536135plusmn831(23)
3725plusmn0085
Lac232638plusmn1558(55)
1942plusmn009228713plusmn274(23)
1709plusmn0016
CoffeeHusk + 0mM Cu+2
113740plusmn8289(50-55)
3008plusmn0219290666plusmn1039(2-3)
7689plusmn0275
Lac1 889plusmn039 (55) 0038plusmn0002 1779plusmn028 (23) 0077plusmn0001
Lac234194plusmn2789(60)
2947plusmn024033150plusmn526 (2ndash4)
2857plusmn0045
CedarSawdust +5 mM Cu+2
257666plusmn1201(55)
5052plusmn0024344634plusmn1043(2-3)
6757plusmn0204
Lac121370plusmn472(55)
0962plusmn002129117plusmn943(23)
1311plusmn0042
Lac255166plusmn1656(60)
1379plusmn0414 55166plusmn1656 (5) 1379plusmn0414
ABTS was used to determine laccase activity
Laccase stabilityFor selected enzymatic extracts and purified fractions (Table
4) a stability test for 48 h at their optimal pH and temperaturewas conducted Time course of the relative laccase activity forcrude extract and purified fractions from coffee husk and cedarsawdust cultures are presented in Figure 5 Similar resultswere observed for crude extract and purified fractions fromsoybean pod husk culture (data no shown)
It was observed that the activity loss depends on thesubstrate source used to grow T pubescens An activity loss of55 was observed for crude extracts and purified fractionsfrom coffee husk and soybean pod husk cultures while anactivity loss of 70 was observed for the crude extract andpurified fractions from cedar sawdust culture NeverthelessLac2 showed the lowest activity reduction with values of 10 40and 55 for coffee husk soybean pod husk and cedarsawdust respectively while Lac1 presented an activity loss of34 50 and 60 for coffee husk soybean pod husk and cedarsawdust respectively Despite the differences in laccaseactivity among agro-industrial wastes and between crudeextracts and purified fractions all the profiles showed a sharp
Figure 4 Effect of temperature (a) and pH (b) on laccaseactivity for the UF retentate obtained by semisolid cultureof T pubescens on coffee husk () soybean pod husk (loz)and cedar sawdust (Δ) Laccase stability was evaluated byusing ABTS All the assays were performed in triplicatedoi 101371journalpone0073721g004
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 8 September 2013 | Volume 8 | Issue 9 | e73721
decrease during the first 8 h while a reduced activity loss wasobserved during the last 24 h of the assay (Figure 6)
Laccase electrochemical characterizationFor selected crude extracts and purified fractions a cyclic
voltammetry characterization was conducted For eachmediator syringaldazine or ABTS similar cyclic
Figure 5 Effect of temperature () and pH () on enzymeactivity for Lac1 (solid line) and Lac2 (dashed line)obtained after the purification of the enzymatic crudeextract produced by semisolid culture of T pubescens oncoffee husk (a) soybean pod husk (b) and cedar sawdust(c) Laccase stability was evaluated by using ABTS All theassays were performed in triplicatedoi 101371journalpone0073721g005
voltammograms were obtained for the different crude extractsand purified fractions Voltammograms of the redox reaction forsyringaldazine (Figure 7a) and ABTS (Figure 6b) arepresented Two anionic and cationic peaks were observedwhen syringaldazine was used as mediator The first pair ofpeaks showed a reversible performance with a mean Ia Icratio of 109plusmn006 106plusmn005 and 098plusmn002 for the crudeextracts Lac1 and Lac2 respectively The second pair ofpeaks showed quasi-reversible performance and the cathodicpeak was presented at a potential of +062 V (vs Ag|AgCl)while the anodic peak was presented at a potential of +060 V(vs Ag|AgCl) No significant difference on the reductionpotential (Eo) for syringaldazine was observed for the crudeextracts or the purified fractions obtaining an Eo of0314plusmn0008 V (vs Ag|AgCl) However the peak potentialseparation (ΔEp) showed a different profile depending on thesubstrate used for the culture While not significant difference(p gt 005) on ΔEp value was observed between crude extract(0045plusmn0005 V vs Ag|AgCl) Lac1 (0046plusmn0005 V vs Ag|AgCl)and Lac2 (0028plusmn0001 V vs Ag|AgCl) from cultures with cedarsawdust and soybean pod husk the ΔEp was considerablyhigher for the crude extract (0083plusmn0004 V vs Ag|AgCl) Lac1(0068plusmn0003 V vs Ag|AgCl) and Lac2 (0051plusmn0005 V vs Ag|AgCl) from the coffee husk culture
For reactions conducted with ABTS and the different crudeextracts and purified fractions the voltammograms exhibitedone pair of cathodic and anodic peaks (Figure 7b) whichcorresponds to the redox of ABTSABTS+ (IaIc) with an Eo of0487plusmn0006 V (vs Ag|AgCl) This process was reversiblepresenting a mean Ia Ic ratio of 099plusmn002 In addition asecond cathodic peak was presented at +083 V (vs Ag|AgCl)when ABTS redox reaction was conducted using the crudeextract and the purified fractions This performance agrees withprevious reports in which the laccase activity cannot beobserved at potential values of the second reversible oxidation
Figure 6 Laccase stability profiles at optimal pH andtemperature conditions for crude extracts (solid line) Lac1(dashed line) and Lac2 (spotted line) obtained fromsemisolid culture of T pubescens on coffee husk (loz) andcedar sawdust (bull) Laccase stability was evaluated by usingABTS All the assays were performed in triplicatedoi 101371journalpone0073721g006
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 9 September 2013 | Volume 8 | Issue 9 | e73721
of ABTS ABTS+ABTS2+ due to this catalytic process cannotbe detected by cyclic voltammetry [2526] Neverthelessvoltammograms for laccase crude extract from a soybean podhusk culture presented the second anodic peak which inaddition to the second cathodic peak corresponds to the redoxof ABTS+ABTS2+ (IIaIIc) with an Eo of 0942plusmn0004 V (vs Ag|AgCl) Contrary to the reactions conducted usingsyringaldazine as substrate the ΔEp did not present asignificant difference for the crude extracts and purifiedfractions 0248plusmn0016 V (vs Ag|AgCl)
Discussion
In this study we evaluated the effect of the culture mode ieSmC and SSC on laccase production by T pubescens CBS69694 using three different agro-industrial wastes (coffeehusk soybean pod husk and cedar sawdust) Furthermore theeffect of Cu+2 supplementation on laccase production was alsoevaluated and the crude extracts and the purified fractionswere partially characterized
Figure 7 Cyclic voltammograms of (a) syringaldazine atpH 50 using the crude extract and purified laccasefractions from semisolid culture of T pubescens on coffeehusk or (b) ABTS at pH 30 using the crude extract andpurified laccase fractions from semisolid culture of Tpubescens on soybean pod husk Nomenclature onlymediator (dotted line) crude extract (solid line) Lac1 fraction(dashed line) and Lac2 fraction (dashed and dotted line)doi 101371journalpone0073721g007
Substrate source for laccase production showed to be one ofthe most important factors for laccase production (Table 2)Cultures with coffee husk as substrate presented the highestlaccase production levels The observed differences in laccaseactivity for each substrate could be owed to their chemicalcomposition Coffea arabica is the coffee variety raised inColombia and its husk has a reported composition of 2308cellulose (2076 glucose and 183 cellobiose) 2385hemicellulose (1356 xylose 523 arabinose 256 aceticacid and 195 glucuronic acid) 2828 total lignin and071 ashes [27] In addition coffee husk contents 13caffeine 45 tannins and 124 pectins [28] On the otherhand Glycine max is the soybean species cultured inColombia and its pod husk composition varies from 504 to62 crude protein 608 to 143 hemicellulose 3715 to4208 acid detergent fiber 4436 to 6015 neutral detergentfiber 850 to 893 acid detergent lignin and 2074 to 2106uronic acid [29ndash31] Finally for cedar sawdust the majorcomponents are glucose (426-444) xylose (46-195)cellulose (383-410) klason lignin (227-278) and acidlignin (40-43) [32ndash34] Although lignin is one of the mostimportant compounds associated with laccase production bywhite-rot fungi we did not observe a clear correlation betweenlignin composition percentage and laccase activity In thatsense the high laccase activity observed for the cultures thatuse coffee husk could be associated with its content of caffeineand tannins which have been identified as potent inducers oflaccase gene expression [173536] In fact caffeine has beenused as inducer for laccase production by Rhizoctonia solaniobtaining an increase of 41-fold in laccase activity [37] Similarresults were also reported for the production of Coloriopsisgallica laccase [38]
Laccase gene expression is mainly up regulated by Cu2+although Mn2+ Fe3+ heavy metals 26-dimethoxy-14-benzoquinone H2O2 amphotericin B syringic acid tannic acidTween 80 soybean oil aromatic compounds andmicroclimatic changes (ie lower temperature and osmoticpressure) are also recognized as potent inducers [173536] Ingeneral Cu2+ supplementation increased the laccase activity inboth SmC and SSC However Cu2+concentration required toobtain the highest laccase activity varied depending on agro-industrial waste These differences can be attributed to thecopper presented in the agro-industrial waste and the copperavailability in the medium Copper can be found in amountsthat range from 156 to 180 mg kg-1 of coffee husk [39] andfrom 26 to 67 mg kg-1 of soybean pod husk (or soybean hull)[3940] Despite that no reports indicate the copper content inuntreated cedar wood an average of 10 mg kg-1 of wood hasbeen reported for wood used as biomass in six different Europecountries [41] This allows to consider the cedar sawdust as thesubstrate with the lowest content of copper On the other handcopper adsorption by sawdust and coffee husk has beenreported obtaining a Langmuir based maximum adsorptioncapacity (Qmax mg g-1 adsorbent) of 845 and 75 respectively[4243] Although no adsorption studies has been reportedusing untreated soybean pod husk Šćiban et al [44] reporteda Qmax for copper adsorption of 0085 mmol g-1 adsorbent (~ 54mg g-1) using treated soybean straws Based on this
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 10 September 2013 | Volume 8 | Issue 9 | e73721
information it can be expected that a higher amount ofexogenous copper would be required for laccase induction oncultures with cedar sawdust than with soybean pod husk andcoffee husk due to its lower content of copper In addition dueto the high copper-adsorption capacity of sawdust the amountof available copper in the culture would be reduced allowingthe laccase production at higher copper concentrations withoutreaching levels that might be toxic for the fungus This fact canbe observed in the SSC (Table 4) However for the SmC thisbehavior is not maintained for the cultures with soybean podhusk and coffee husk (Table 4) where the maximum laccaseactivity was obtained with a copper concentration of 05 and 5mM respectively This could be owed to a lower contactbetween the fungus and the substrate in the SmC whencompared to the SSC which is close related to the transport ofnutrients presented in the substrates
Production of extracellular laccases has been reported in alarge number of species of white-rot fungi strains grown onnatural substrates (Table 1) Although a laccase activity profileis not clearly observed based on the agro-industrial wasteculture mode and fungus strain most cases in which SmC andSSC were conducted using the same substrate and fungusstrain showed a higher laccase activity under SmC conditionsIn this study was observed a substrate dependentperformance where the most favorable condition to obtain thehighest laccase activity was the SSC The laccase activitiesobtained in the present study are comparable and in somecases higher than those reported for Pleurotus ostreatusPhanerochaete chrysosporium Pycnoporus cinnabarinus Tversicolor and Ganoderma sp cultured under SC and SmCconditions and using different agro-industrial wastes (Table 1)Semi-solid cultures using different agro-industrial wastes havebeen reported using T versicolor Phlebia radiata Tpubescens P ostreatus and C unicolor Rodriguez et al [16]screened different supports for laccase production by Tversicolor under SSC They obtained the highest laccaseactivity (1200 U L-1) when in the culture medium was presentedbarley bran In addition they were able to obtain a laccaseactivity of 1700 U L-1 when the culture was supplemented withxylidine as an inducer These results were obtained after 17 or18 days of culture and ABTS was used to determine theenzyme activity Maumlkelauml et al [4546] have studied theexpression of the genes involved in laccase production Pr-lac1and Pr-lac2 presented in P radiata and evaluate laccaseproduction in SSC using milled alder wood (Alnus incana) assubstrate They reported a maximum laccase activity of 3 microkatL-1 (~180 U L-1) at the 14th day of culture and usingsyringaldazine for its detection Osma et al [15] have studiedthe morphological changes and laccase production of strains Tpubescens P ostreatus C unicolor and T versicolor grownon wheat bran flakes using SSC They obtained a maximumlaccase production between 1397 and 2778 U L-1 after 10 and13 days of culture This enzyme activity is strain dependentand ABTS was used for its quantification Our results show agood correlation with reported data The highest laccaseactivity was obtained when T pubescens was grown on coffeehusk under SSC without copper (204 U L-1 evaluated withsyringaldazine or 1549 U L-1 evaluated with ABTS) which is in
some cases higher than reported ones This shows that underthe studied conditions the production of the laccase crudeextract is favored in SSC using coffee husk without the additionof copper which could favors the cost of the productionprocess
It has been reported for white rot-fungi the production ofmultiple laccase isoenzymes which are encoded by differentgenes and show different expression profiles stabilitiessubstrate affinities and molecular masses (ranging from 35 to140 kDa) [3647ndash49] Two isoforms have been identified in Tpubescens strains two monomeric (Lac1 and Lac2) enzymesof 67 kDa but with different pI (53 and 51) [50] and twolaccase-activity fractions (Lap1 and Lap2) with different pI (gt30 and 26) from which Lap2 is a monomeric enzyme of 65kDa [17] In this study we identify two laccase isoenzymesfrom T pubescens CBS 69694 that have different molecularmasses (120 and 60 kDa) and pI The purifications resultssuggest a more acidic pI for Lac1 (le 4) than Lac2 (ge 6) whichdiffers from a previous report for this fungus in which twofractions with laccase activity were obtained Lap1 and Lap2with pI gt 3 and of 26 respectively [17] Since in both casessimilar purification protocols were used and Lac1 and Lac2were recovered from the unbound and bound fractionsrespectively as Lap1 and Lap2 [17] this difference could bedue to the culture conditions used in each case (ie thepresence of agro-industrial wastes) Nevertheless the nanoLC-MSMS analysis showed that peptides of Lac1 and Lac2 werepresented in Lap1 (UniProtKBSwiss-Prot Q8TG93) and Lap2(UniProtKBSwiss-Prot Q8TG94) which have a predicted pI of43 and 47 respectively Lac1 and Lac2 peptides werepresented in Laccase-2 (UniProtKBSwiss-Prot Q99046) andLaccase-1 (UniProtKBSwiss-Prot Q99044) from T villosarespectively with 80 to 100 identity These T villosa laccaseisoenzymes have a predicted pI of 47 (laccase-1) and 60(laccase-2) and a 77 identity among them In addition Tvillosa laccase-2 is reported as a homodimer protein whilelaccase-1 is reported as monomeric protein Alignment of Lac1-Lap2 and Lac2-Lap1 from T pubescens CBS 69694 and Tvillosa showed a 96 and 70 identity respectively Finallysimilar peptides were identified for Lac1 and Lac2 from Tpubescens (Schumach) Pilaacutet (Syn Coriolus pubescens(Schum ex Fr) Queacutel) BCB 923-2 (Figure 3b) [51] Theseresults suggest that the purified fractions correspond to laccaseisoenzymes that differ from previously reported ones for this Tpubescens strain
As mentioned above laccases isoenzymes present differentstabilities and substrate affinities This behavior was alsoobserved for the laccase isoenzymes purified from Tpubescens with Lac1 showing lower laccase activity thanLac2 with ABTS as substrate However the difference inactivity between these isoenzymes depended on the agro-industrial waste used for the T pubescens growth suggestinga probable effect of the substrate composition on the laccasephysicochemical properties In addition it was observed thatthe total laccase activity presented in the UF retentates ishigher than the laccase activity of the purified fractionsregardless of the agro-industrial byproduct used for the Tpubescens growth This suggests a synergistic effect of Lac1
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 11 September 2013 | Volume 8 | Issue 9 | e73721
and Lac2 as have been reported for laccases obtained fromTrametes and Pleurotus strains [175253]
The maximum relative activity for the UF retentate waspresented at a pH from 20 to 40 regardless of the agro-industrial byproduct with a monotonical decrease at pHs abovethis range The relation between pH and type of electron donormolecule explains this behavior on laccase activity It has beenreported that when the substrate is an organic hydrogen donorie phenols the optimal pH ranges from 35 to 60 while thelaccase activity monotonically decreases from pH 25 to 70 ifthe substrate is an electron donor like ABTS [54] Despite thatat different pHs most of the purified laccase fractionspresented a similar performance as the UF retentates the Lac2fraction obtained from cedar sawdust differed from the previousobservation For this fraction the maximum activity wasobserved at pH 50 and despite that at acidic pH (ge 30) therelative activity was higher than 80 at lower pHs (20) itdecreased almost in a 50 Nevertheless at basic pHs therelative laccase activity presented a sharp decreased that iswell correlate with the other laccase fractions We are not ableto attribute this particular trend to the presence of multiplelaccase isoenzymes in the fraction since the SDS-PAGErevealed only one band when stained with ABTS and from theNanoLC-MSMS analysis no difference in the number ofpeptides and their sequence was detected We speculate thatthis particular behavior can be owed to a compound added inthe pre-treatment of the cedar wood that might be used forpreserving it that might affect the activity of Lac2 and that wasnot completely removed during the purification
Temperature stability evaluation showed that Lac1 is themost thermolabile fraction while the laccase activity reductionin the crude extracts did not seem to be the result of anadditive effect of the activity loss of the purified fractionsAlthough there are no reports of laccase stability under theselected conditions these results agree with previous reportsthat state that fungal laccases rapidly decrease their activity attemperatures near 60 degC and thermal stability correlates withthe temperature range of the growth of the source organism[55] In addition we previously reported for a laccase crudeextract obtained from culturing T pubescens on coffee husk anactivity loss of 26 and 10 at 60 and 50 degC respectivelyafter 8 h at pH 60 using syringaldazine as substrate [48]
Finally an electrochemical characterization was carried outfor selected crude extracts and purified fractions The results
showed a difference between the electrochemicalperformances of extracts and purified fractions withsyringaldazine or ABTS These differences could be owed tothe different mechanisms for electron transfer as discussedabove However the overall performance of the obtained cyclicvoltammograms for ABTS and syringaldazine agrees withresults reported by Fernaacutendez-Saacutenchez et al [25] who used asaturated calomel reference electrode
Conclusions
We have shown the potential of coffee husk soybean podhusk and cedar sawdust as substrates for the laccaseproduction In several countries like Brazil and Colombia theproduction of coffee and soybean is of great importance andthe generated volume of residues is high The results showedthat semi-solid culture of T pubescens using coffee huskallows to obtain an important amount of laccase which couldbe favored by the phenolic-compounds ie tannins presentedin coffee husk While supplementation of the culture mediumwith copper favored the production of laccase in submergedand semi-solid culture using soybean pod husk and cedarsawdust semi-solid culture using coffee husk which presentedthe highest laccase volumetric yield did not require coppersupplementation This is an economic advantage that shouldbe considered when scaling up the process In this study Tpubescens showed its ability to produce two independentlaccases regardless of the used substrate However a higherlaccase activity was observed for crude extracts rather than forpurified fractions This suggests that Lac1 and Lac2 might havea synergistic effect on the final laccase activity rather than anadditive effect Based on this study the investigation of laccaseproduction by evaluating other agro-industrial wastes in whichphenolic-compounds constitute an important percentage oftheir composition is of great interest to avoid the use ofinducers that would increase the production cost
Author Contributions
Conceived and designed the experiments JFO CJAD OFSPerformed the experiments JCG SCM AR Analyzed the dataJCG SCM AR JFO CJAD OFS Wrote the manuscript AR JFOCJAD OFS
References
1 Vandamme EJ (2009) Agro-industrial residue utilization for industrialbiotechnology products In P NigamA Pandey Biotechnology for Agro-Industrial Residues Utilisation Netherlands Springer Verlag pp 3-11
2 Martiacutenez AT Speranza M Ruiz-Duentildeas FJ Ferreira P Camarero S etal (2005) Biodegradation of lignocellulosics microbial chemical andenzymatic aspects of the fungal attack of lignin Int Microbiol 8195-204 PubMed 16200498
3 Saacutenchez O Sierra R Almeacuteciga-Diacuteaz CJ (2011) Delignification processof agro-industrial aastes an alternative to obtain fermentablecarbohydrates for producing fuel In M Manzanera Alternative FuelIntech 111-154
4 Wesenberg D Kyriakides I Agathos SN (2003) White-rot fungi andtheir enzymes for the treatment of industrial dye effluents BiotechnolAdv 22 161-187 doi101016jbiotechadv200308011 PubMed14623049
5 Morozova OV Shumakovich GP Shleev SV Yaropolov YI (2007)Laccase-mediator systems and their applications A review ApplBiochem Microbiol 43 523-535 doi101134S0003683807050055
6 Baldrian P (2006) Fungal laccases - occurrence and properties FEMSMicrobiol Rev 30 215-242 doi101111j1574-4976200500010xPubMed 16472305
7 Rodriacuteguez Couto S Herrera JLT (2006) Industrial and biotechnologicalapplications of laccases A review Biotechnol Adv 24 500-513 doi101016jbiotechadv200604003 PubMed 16716556
8 Osma JF Toca-Herrera JL Rodriacuteguez-Couto S (2010) Uses oflaccases in the food industry Enzyme Res 2010 2010 918761PubMed 21048873
9 Rodriguez S (2008) Exploitation of biological wastes for the productionof value-added products under solid-state fermentation conditionsBiotechnol J 3 859-870 doi101002biot200800031 PubMed18543242
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 12 September 2013 | Volume 8 | Issue 9 | e73721
10 Osma JF Toca-Herrera JL Rodriacuteguez-Couto S (2011) Cost analysis inlaccase production J Environ Manage 92 2907-2912 doi101016jjenvman201106052 PubMed 21775046
11 Jain A Morlok CK Henson JM (2013) Comparison of solid-state andsubmerged-state fermentation for the bioprocessing of switchgrass toethanol and acetate by Clostridium phytofermentans Appl MicrobiolBiotechnol 97 905-917 doi101007s00253-012-4511-4 PubMed23111595
12 Singhania RR Patel AK Soccol CR Pandey A (2009) Recentadvances in solid-state fermentation Biochem Eng J 44 13-18 doi101016jbej200810019
13 Economou CN Makri A Aggelis G Pavlou S Vayenas DV (2010)Semi-solid state fermentation of sweet sorghum for the biotechnologicalproduction of single cell oil Bioresour Technol 101 1385-1388 doi101016jbiortech200909028 PubMed 19781936
14 Rodriacuteguez CS Santoro R Cameselle C Sanroman A (1997) Laccaseproduction in semi-solid cultures of Phanerochaete chrysosporiumBiotechnol Lett 19 995-998 doi101023A1018495216946
15 Osma JF Moilanen U Toca-Herrera JL Rodriacuteguez-Couto S (2011)Morphology and laccase production of white-rot fungi grown on wheatbran flakes under semi-solid-state fermentation conditions FEMSMicrobiol Lett 318 27-34 doi101111j1574-6968201102234xPubMed 21291496
16 Couto SR Gundin M Lorenzo M Sanroman MN (2002) Screening ofsupports and inducers for laccase production by Trametes versicolor insemi-solid-state conditions Proc Biochem 38 249-255 doi101016S0032-9592(02)00087-0
17 Galhaup C Goller S Peterbauer CK Strauss J Haltrich D (2002)Characterization of the major laccase isoenzyme from Trametespubescens and regulation of its synthesis by metal ions Microbiology148 2159-2169 PubMed 12101303
18 Sievers F Wilm A Dineen D Gibson TJ Karplus K et al (2011) Fastscalable generation of high-quality protein multiple sequencealignments using Clustal Omega Mol Syst Biol 7 539 PubMed21988835
19 Childs RE Bardsley WG (1975) The steady-state kinetics of peroxidasewith 22-azino-di-(3-ethyl-benzthiazoline-6-sulphonic acid) aschromogen Biochem J 145 93-103 PubMed 1191252
20 Harkin JM Obst JR (1973) Syringaldazine an Effective Reagent forDetecting Laccase and Peroxidase in Fungi Experientia 29 382-387
21 Nikupaavola ML Raaska L Itavaara M (1990) Detection of white-rotfungi by a non-roxic stain Mycol Res 94 27-31 doi101016S0953-7562(09)81260-4
22 Gonzalez LF Sarria V Sanchez OF (2010) Degradation ofchlorophenols by sequential biological-advanced oxidative processusing Trametes pubescens and TiO(2)UV Bioresour Technol 1013493-3499
23 Waterborg JH (2009) The Lowry method for protein quantitation In JWalker The Protein Protocols Handbook Humana Press pp 7-10
24 Harkin JM Obst JR (1973) Syringaldazine an effective reagent fordetecting laccase and peroxidase in fungi Cell Mol Life Sci 29381-387 doi101007BF01926734
25 Fernaacutendez-Saacutenchez C Tzanov T Guumlbitz GM Cavaco-Paulo A (2002)Voltammetric monitoring of laccase-catalysed mediated reactionsBioelectrochemistry 58 149-156 doi101016S1567-5394(02)00119-6PubMed 12414320
26 Bourbonnais R Leech D Paice MG (1998) Electrochemical analysis ofthe interactions of laccase mediators with lignin model compoundsBiochim Biophys Acta 1379 381ndash390 doi101016S0304-4165(97)00117-7 PubMed 9545600
27 Ferraz F Silva S (2009) Characterization of coffee husk biomass forbiotechnological purposes N Biotechnol 25 S256-S256 doi101016jnbt200906573
28 Pandey A Soccol CR Nigam P Brand D Mohan R et al (2000)Biotechnological potential of coffee pulp and coffee husk forbioprocesses Biochem Eng J 6 153-162 doi101016S1369-703X(00)00084-X PubMed 10959086
29 Sruamsiri S Silman P (2008) Nutritive composition of soybean by-products and nutrient digestibility of soybean pod husk Maejo Int J SciTechnol 2 568-576
30 Romkaew J Nagaya Y Goto M Suzuki J Umezaki T (2008) Poddehiscence in relation to chemical components of pod shell in soybeanPlant Prod Sci 11 278-282 doi101626pps11278
31 Nakamanee G Polbumroong T Phaikaew C (2003) Alternative use ofcrop waste to increase milk production Proceedings of the 8th Meetingof the Regional Working Group on Grazing and Feed Resources forSoutheast Asia Kuala Lumpur Malaysia 66 - 70
32 Kurek B Gaudard F (2000) Oxidation of spruce wood sawdust byMnO(2) plus oxalate a biochemical investigation J Agric Food Chem48 3058-3062 doi101021jf000015g PubMed 10898665
33 Weil J Brewer M Hendrickson R Sarikaya A Ladisc MR (1998)Continuous pH monitoring during pretreatment of yellow poplar woodsawdust by pressure cooking in water Appl Biochem Biotechnol 70-7299-111 doi101007BF02920127
34 Weil J Sarikaya A Rau S-L Goetz J Ladisch CM et al (1997)Pretreatment of yellow poplar sawdust by pressure cooking in waterAppl Biochem Biotechnol 68 21-40 doi101007BF02785978
35 Dekker RF Barbosa AM Giese EC Godoy SD Covizzi LG (2007)Influence of nutrients on enhancing laccase production byBotryosphaeria rhodina MAMB-05 Int Microbiol 10 177-185 PubMed18075999
36 Xiao YZ Hong YZ Li JF Hang J Tong PG et al (2006) Cloning ofnovel laccase isozyme genes from Trametes sp AH28-2 and analysesof their differential expression Appl Microbiol Biotechnol 71 493-501doi101007s00253-005-0188-2 PubMed 16283298
37 Crowe JD Olsson S (2001) Induction of laccase activity in Rhizoctoniasolani by antagonistic Pseudomonas fluorescens strains and a range ofchemical treatments Appl Environ Microbiol 67 2088-2094 doi101128AEM6752088-20942001 PubMed 11319086
38 Yaguumle S Terroacuten MC Gonzaacutelez T Zapico E Bocchini P et al (2000)Biotreatment of tannin-rich beer-factory wastewater with white-rotbasidiomycete Coriolopsis gallica monitored by pyrolysisgaschromatographymass spectrometry Rapid Commun Mass Spectrom14 905-910 doi101002(SICI)1097-0231(20000530)1410 PubMed10825255
39 Gowda NKS Ramana JV Prasad CS Singh K (2004) Micronutrientcontent of certain tropical conventional and unconventional feedresources of southern India Trop Anim Heatlth Pro 36 77-94 doi101023BTROP0000009522309491d PubMed 14979561
40 DePeters EJ Fadel JG Arana MJ Ohanesian N Etchebarne MA et al(2000) Variability in the chemical composition of seventeen selectedby-product feedstuffs used by the California dairy industry Prof AnimSci 16 69-99
41 Obernberger I Thek G (2004) Physical characterisation and chemicalcomposition of densified biomass fuels with regard to their combustionbehaviour Biomass Bioenerg 27 653-669 doi101016jbiombioe200307006
42 Larous S Meniai A-H Lehocine MB (2005) Experimental study of theremoval of copper from aqueous solutions by adsorption usingsawdust Desalination 185 483ndash490 doi101016jdesal200503090
43 Oliveira WE Franca AS Oliveira LS Rocha SD (2008) Untreatedcoffee husks as biosorbents for the removal of heavy metals fromaqueous solutions J Hazard Mater 152 1073ndash1081 doi101016jjhazmat200707085 PubMed 17804159
44 Šciban M Klašnja M Škrbic B (2008) Adsorption of copper ions fromwater by modified agricultural by-products Desalination 229 170ndash180doi101016jdesal200708017
45 Maumlkelauml MR Hildeacuten KS Hakala TK Hatakka A Lundell TK (2006)Expression and molecular properties of a new laccase of the white rotfungus Phlebia radiata grown on wood Curr Genet 50 323-333 doi101007s00294-006-0090-1 PubMed 16927090
46 Maumlkelauml MR Lundell T Hatakka A Hildeacuten K (2013) Effect of coppernutrient nitrogen and wood-supplement on the production of lignin-modifying enzymes by the white-rot fungus Phlebia radiata Fungal Biol117 62-70 doi101016jfunbio201211006 PubMed 23332834
47 Dwivedi UN Singha P Pandeya VP Kumara A (2011) Structurendashfunction relationship among bacterial fungal and plant laccases J MolCatal B Enzym 68 117-128 doi101016jmolcatb201011002
48 Gaitan IJ Medina SC Gonzaacutelez JC Rodriacuteguez A Espejo AacuteJ et al(2011) Evaluation of toxicity and degradation of a chlorophenol mixtureby the laccase produced by Trametes pubescens Bioresour Technol102 3632ndash3635 doi101016jbiortech201011040 PubMed21115244
49 Karp SG Faraco V Amore A Birolo L Giangrande C et al (2012)Characterization of laccase isoforms produced by Pleurotus ostreatusin solid state fermentation of sugarcane bagasse Bioresour Technol114 735-739 doi101016jbiortech201203058 PubMed 22487128
50 Nikitina OV Shleev SV Gorshina ES Rusinova TV Serezhenkov VAet al (2005) Isolation and purification of enzymes from ligninolyticcomplex of the basidial fungus Trametes pubescens (Schumach) Pilatand study of their properties Biochemistry (Mosc) 70 1274-1279 doi101007s10541-005-0259-0
51 Shleev S Nikitina O Christenson A Reimann CT Yaropolov AI et al(2007) Characterization of two new multiforms of Trametes pubescenslaccase Bioorg Chem 35 35-49 doi101016jbioorg200608001PubMed 16989887
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 13 September 2013 | Volume 8 | Issue 9 | e73721
52 Xiao YZ Chen Q Hang J Shi YY Wu J et al (2004) Selectiveinduction purification and characterization of a laccase isozyme fromthe basidiomycete Trametes sp AH28-2 Mycologia 96 26-35 doi1023073761984 PubMed 21148825
53 Muntildeoz C Guilleacuten F Martiacutenez AT Martiacutenez MJ (1997) Laccaseisoenzymes of Pleurotus eryngii characterization catalytic propertiesand participation in activation of molecular oxygen and Mn2+ oxidationAppl Environ Microbiol 63 2166-2174 PubMed 9172335
54 Morozova OV Shumakovich GP Gorbacheva MA Shleev SVYaropolov AI (2007) Blue laccases Biochemistry (Mosc) 721136-1150 doi101134S0006297907100112
55 Hildeacuten K Hakala TK Lundell T (2009) Thermotolerant andthermostable laccases Biotechnol Lett 31 1117-1128 doi101007s10529-009-9998-0 PubMed 19360388
56 Verma P Madamwar D (2002) Production of ligninolytic enzymes fordye decolorization by cocultivation of white-rot fungi Pleurotusostreatus and Phanerochaete chrysosporium under solid-statefermentation Appl Biochem Biotechnol 102-103 109-118 doi101385ABAB102-1031-6109 PubMed 12396115
57 Meza JC Lomascolo A Casalot L Sigoillot J-C Auria R (2005)Laccase production by Pycnoporus cinnabarinus grown on sugar-canebagasse Influence of ethanol vapours as inducer Proc Biochem 403365ndash3371 doi101016jprocbio200503004
58 Hossain SM Anantharaman N (2006) Activity enhancement ofligninolytic enzymes of Trametes versicolor with bagasse powder Afr JBiotechnol 5 189-194
59 Revankar MS Desai KM Lele SS (2007) Solid-state fermentation forenhanced production of laccase using indigenously isolatedGanoderma sp Appl Biochem Biotechnol 143 16-26 doi101007s12010-007-0029-0 PubMed 18025593
60 Velazquez-Cedeno MA Mata G Savoie J-M (2002) Waste-reducingcultivation of Pleurotus ostreatus and Pleurotus pulmonarius on coffeepulp changes in the production of some lignocellulolytic enzymesWorld J Microbiol Biotechnol 18 201ndash207 doi101023A1014999616381
61 Mata G Murrieta D Andreu LI (2005) Changes in lignocellulolyticenzyme activities in six Pleurotus spp strains cultivated on coffee pulpin confrontation with Trichoderma spp World J Microbiol Biotechnol 21143ndash150 doi101007s11274-004-3041-3
62 de Souza DF Tychanowicz GK de Souza CG Peralta RM (2006) Co-production of ligninolytic enzymes by Pleurotus pulmonarius on wheatbran solid state cultures J Basic Microbiol 46 126-134 doi101002jobm200510014 PubMed 16598826
63 Prasad KK Mohan SV Bhaskar YV Ramanaiah SV Babu VL et al(2005) Laccase production using Pleurotus ostreatus 1804 immobilizedon PUF cubes in batch and packed bed reactors influence of cultureconditions J Microbiol 43 301-307 PubMed 15995650
64 Songulashvili G Elisashvili V Wasser SP Nevo E Hadar Y (2007)Basidiomycetes laccase and manganese peroxidase activity in
submerged fermentation of food industry wastes Enzyme MicrobTechnol 41 57ndash61 doi101016jenzmictec200611024
65 Sathishkumar P Murugesan K Palvannan T (2010) Production oflaccase from Pleurotus florida using agro-wastes and efficientdecolorization of Reactive blue 198 J Basic Microbiol 50 360-367 doi101002jobm200900407 PubMed 20586068
66 Moldes D Gallego PP Rodriacuteguez Couto S Sanromaacuten A (2003) Grapeseeds the best lignocellulosic waste to produce laccase by solid statecultures of Trametes hirsuta Biotechnol Lett 25 491-495 doi101023A1022660230653 PubMed 12882277
67 Elisashvili V Kachlishvili E Penninckx M (2008) Effect of growthsubstrate method of fermentation and nitrogen source onlignocellulose-degrading enzymes production by white-rotbasidiomycetes J Ind Microbiol Biotechnol 35 1531-1538 doi101007s10295-008-0454-2 PubMed 18716810
68 Osma JF Saravia V Herrera JL Couto SR (2007) Mandarin peelingsthe best carbon source to produce laccase by static cultures ofTrametes pubescens Chemosphere 67 1677-1680 doi101016jchemosphere200611051 PubMed 17234250
69 Kahraman SS Gurdal IH (2002) Effect of synthetic and natural culturemedia on laccase production by white rot fungi Bioresour Technol 82215-217 doi101016S0960-8524(01)00193-6 PubMed 11991068
70 Iandolo D Piscitelli A Sannia G Faraco V (2011) Enzyme Productionby Solid Substrate Fermentation of Pleurotus ostreatus and Trametesversicolor on Tomato Pomace Appl Biochem Biotechnol 163 40-51doi101007s12010-010-9014-0 PubMed 20582639
71 Moldes D Lorenzo M Sanromaacuten MA (2004) Different proportions oflaccase isoenzymes produced by submerged cultures of Trametesversicolor grown on lignocellulosic wastes Biotechnol Lett 26 327-330doi101023BBILE000001545240213bf PubMed 15055770
72 Philippoussis A Diamantopoulou P Papadopoulou K Lakhtar HRoussos S et al (2010) Biomass laccase and endoglucanaseproduction by Lentinula edodes during solid state fermentation of reedgrass bean stalks and wheat straw residues World J MicrobiolBiotechnol 27 285-297
73 Fenice M Giovannozzi Sermanni G Federici F DrsquoAnnibale A (2003)Submerged and solid-state production of laccase and Mn-peroxidaseby Panus tigrinus on olive mill wastewater-based media J Biotechnol100 77-85 doi101016S0168-1656(02)00241-9 PubMed 12413788
74 Rodriguez-Couto S GundIn M Lorenzo M Sanromaacuten MAacute (2002)Screening of supports and inducers for laccase production by Trametesversicolor in semi-solid-state conditions Proc Biochem 38 249-255doi101016S0032-9592(02)00087-0
75 Fackler K Gradinger C Schmutzer M Tavzes C Burgert I et al (2007)Biotechnological wood modification with selective white-rot fungi and itsmolecular mechanisms Food Technol Biotechnol 45 269ndash276
76 Levin L Herrmann C Papinutti VL (2008) Optimization oflignocellulolytic enzyme production by the white-rot fungus Trametestrogii in solid-state fermentation using response surface methodologyBiochem Eng J 39 207-214 doi101016jbej200709004
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 14 September 2013 | Volume 8 | Issue 9 | e73721

wine and beer stabilization fruit juice processing and differentfood-related biosensors [8]
Use of industrial and agricultural wastes for laccaseproduction by white-rot fungi is an effective way to reduce thecost of production [910] In addition laccase-mediateddelignification allows to increase the nutritional value of agro-industrial byproducts for animal feed or soil fertilizer Toaccomplish the transformation of these agro-industrial wastessubmerged and solid state cultures have been conducted(Table 1) Submerged fermentations are commonly used inindustrial process although they can present drawbacks due tophysical space and energetic and water requirements [11] Onthe other hand solid state cultures present several advantageslike mimicking the natural habitat in which the microorganismgrows reduced water activity that reduce microbialcontaminations and limited water consumption and equipmentsize Also it has been reported that solid-state cultures presenthigher volumetric yield less energy requirements and higherend-product stability and concentration than submergedcultures [1112] Nevertheless there are important issuesrelated to heat and mass transfer that must be overcome inorder to scale up a solid-state culture in addition to accruedestimation of the biomass and recovery of the end product [12]Semi-solid culture has been studied as an alternative toaddress some of the drawbacks presented by submerged andsolid-state cultures The increment of water activity in this typeof culture allows to improve the nutrient and end-productavailability and culture control [13] It has been establishedthat nature and moisture of the agro-industrial waste used for asolid-state culture are critical factors [12] The evaluation ofthese parameters among other process conditions ie culturetemperature pH and aeration has encouraged the screeningof several agro-industrial wastes for the production of differentenzymes In spite that several studies have been reported forthe production of laccase in semi-solid culture usingPhanerochaete chrysosporium few studies are referred to itsproduction in semi-solid culture using Trametes pubescens[14ndash16]
Agriculture is one of the major economic activities inColombia Thus there is a large production of agro-industrialbyproducts and a concern about their disposal In this studycoffee husk soybean pod husk and cedar sawdust wereevaluated as growth substrates for laccase production by Tpubescens CBS 69694 under submerged (SmC) or semisolidcultures (SSC) The effect of Cu+2 supplementation and culturemethod on laccase production was also evaluated Thedifferent crude extracts were purified and the obtained laccasefractions were partially characterized Results showed coffeehusk as an alternative for laccase production regardless to theculture mode This probably owe to its content of caffeine andtannins that might act as laccase inducer In addition thedependence of the laccase production yield and the culturemode showed to be substrate dependent
Table 1 Agro-industrial byproducts used as substrates forlaccase production
Agro-industrialbyproduct Culture Organism
Activity (U g-1
or UL-1) Substrate ReferenceSugarcanebagasse
SFPleurotus
ostreatus112plusmn28 ABTS [56]
SFPhanerochaete
chrysosporium252plusmn15 ABTS [56]
SFPycnoporus
cinnabarinus2 ABTS [57]
SmF T versicolor 410 ABTS [58]Coffee pulp SF Ganoderma sp 142 ABTS [59]
SFPleurotus
ostreatus1 SGZ [60]
SFPleurotus
pulmonarius12 SGZ [60]
Wheat bran SFPleurotus sp
IE137~145 SGZ [61]
SFPleurotus
pulmonarius
CCB1920000 SGZ [62]
SmFPleurotus
ostreatus 18048033 ABTS [63]
SmFGanoderma
lucidum 44797340plusmn9460 ABTS [64]
Bananapeels
SF Ganoderma sp 974 ABTS [59]
SF Pleurotus florida 54 ABTS [65]
SFLentinula edodes
12239031 ABTS [66]
SFTrametes hirsuta
BT 25663550 ABTS [67]
Mandarinpeels
SmFGanoderma
lucidum 4473145plusmn370 ABTS [64]
SF Pleurotus florida 31 ABTS [65]
SmFGanoderma
lucidum 44735980plusmn3616 ABTS [64]
SmFTrametes
pubescens100 ABTS [68]
Cantaloupepeels
SmFTrametes
pubescens IBB663
1084plusmn92 ABTS [67]
Soy bran SF Pleurotus florida 40 ABTS [65]Vinasse +Cotton stalkextract
SmFGanoderma
lucidum 44793840plusmn9566 ABTS [64]
Tomatopomace
SmFCoriolus versicolor
ATCC 2008014406 SGZ [69]
Grape stalks SmFFunalia trogii
ATCC 2008004880 SGZ [70]
Barley straw SFPleurotus
ostreatus15 ABTS [70]
Grapeseeds
SFTrametes
versicolor35 ABTS [71]
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 2 September 2013 | Volume 8 | Issue 9 | e73721
Table 1 (continued)
Agro-industrialbyproduct Culture Organism
Activity (U g-1
or UL-1) Substrate Reference
SmFTrametes
versicolor~800 ABTS [71]
Wheat straw SmFTrametes
versicolor~500 ABTS [71]
SmFTrametes
versicolor~500 ABTS [66]
Reed grass SFTrametes hirsuta
BT 256622550 ABTS [72]
Bean stalk SFLentinula edodes
12254867 SGZ [67]
Barley bran SFTrametes
pubescens IBB663
162plusmn20 ABTS [72]
Barley bran+ Cu+2
SFLentinula edodes
12225769 SGZ [66]
Apple peels SFTrametes hirsuta
BT 256615740 ABTS [67]
SFTrametes
pubescens IBB663
188plusmn20 ABTS [67]
Tree leaves SmFTrametes
pubescens IBB663
1680plusmn190 ABTS [67]
SFTrametes
pubescens IBB663
280plusmn23 ABTS [67]
Olive millwastewater
SmFTrametes
pubescens IBB663
834plusmn61 ABTS [67]
SFTrametes
pubescens IBB663
205plusmn25 ABTS [67]
Woodshavings
SmFTrametes
pubescens IBB663
630plusmn54 ABTS [73]
SmFPanus tigrinus
CBS 577794600plusmn98 26-DMP [73]
SFPanus tigrinus
CBS 577791309plusmn20 26-DMP [74]
SmFTrametes
versicolor CBS10029
451 ABTS [75]
Woodshavings +Cu+2
SFCeriporiopsis
subvermispora~05 NI [75]
SF Ganoderma sp ~500 ABTS [59]
SFTrametes trogii
MYA 28-11901 ABTS [76]
SF Solid state fermentation SmF Submerged fermentationLaccase activity expressed as UL- 1 for these solid state culturesSGZ Syringaldazine 26-DMP 26-dimethoxyphenol NI No indicated
Materials and Methods
Microorganism and agro-industrial substratesLaccase was produced by using the white-rot fungus T
pubescens CBS 69694 Soybean pod and coffee husk wereobtained from Corporacioacuten Colombiana de InvestigacioacutenAgropecuaria (CORPOICA) and a local coffee-processingcompany respectively The cedar sawdust was obtained froma local market in Bogota (Colombia) The different byproductswere treated with a 05 vv hypochlorite solution for 5 h Afterthat they were rinsed with distilled water until obtaining aneutral pH Finally the byproducts were oven dried at 60 degCuntil constant weight The soybean pod and coffee husk werecrushed to an average particle size of about 08 x 05 mmwhile cedar sawdust was sieved The portion retained by a No100 sieve (opening 015 mm) was used for laccase production
Fungal culture maintenance and laccase productionT pubescens was cultured on malt extract agar (MEA) plates
for 10 days at 30 degC Colonized agar plugs were used forsubculture maintenance every 15 days Independent culturesusing each of the mentioned byproducts were conducted toproduce laccase under SmC or SSC conditions Each culturewas conducted in 1000 mL Erlenmeyer containing 15 g of thedesired dry byproduct and 150 or 50 mL of basal medium forSmC or SSC respectively [basal medium composition per liter05 g glucose 2 g KH2PO4 025 g MgSO4middot7H2O 09 g(NH4)2SO4 01 g CaCl2 and 05 g KCl in a citrate buffer 20mM pH 45 supplemented with 05 g L-1 thiamine [17]] Eachculture flask was inoculated with three 10-mm plugs of activefungus cultured on MEA and incubated for 25 days at 30degCwithout agitation Culture supernatant samples of 05 mL werewithdrawn to monitor laccase activity The effect of Cu+2 onlaccase production was evaluated within a 0 to 5 mMconcentration range (supplied as copper sulphate) The coppersolution was added to 4-days-old cultures A control wasprepared by growing the fungus in the basal medium withoutany agro-industrial waste under the described cultureconditions All the assays were performed in triplicate
Laccase purification and characterizationLaccase purification and characterization was conducted
using the crude extract that showed the highest enzyme activityfor each substrate under the evaluated culture conditions (SmCor SSC) The crude extract was obtained by vacuum filtrationthrough paper Whatman No 1 followed by centrifugation at 4degC and 4000 rpm for 15 min serial vacuum filtration through045 and 022 microm polyether sulphone membranes (Pall CorpPort Washington NY USA) and ultrafiltration (UF) through a10 kDa cut-off membrane of regenerated cellulose (MilliporeBillerica MA USA) The retentate (~10 mL) was analyzed bylow-pressure chromatography with a fraction collector(BioLogicTM LPC Biorad NJ US) using a Q-Sepharosecolumn (Pharmacia Biotech 10 x 10 cm glass chromatographycolumn filled with 6 mL media) equilibrated with 50 mM sodiumacetate buffer pH 50 Elution was performed by a linear NaClgradient (~20 mL) from 0 to 05 M All procedures were carriedout at 4 degC Fractions corresponding to protein peaks were
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 3 September 2013 | Volume 8 | Issue 9 | e73721
collected and analyzed by sodium dodecyl sulfate-polyacrylamide electrophoresis gel (SDS-PAGE) under non-reducing conditions and without heat denaturation Staining ofthe electrophoresis gel was done with 5 mM 22rsquo-azino-bis(3-ethylbenzothiazoline-6) sulphonic acid (ABTS) in succinic acid(25 mM pH 45) to detect laccase activity Fractions thatrevealed the same protein profile and laccase activity werecombined Samples of the combine fractions were analyzed bySDS-PAGE under mentioned conditions As a negative controlwere used a sample of the ultrafiltrated fraction which did notreveal laccase activity along the previous purification stepBands that revealed laccase activity were identified as Lac1and Lac2 These bands were excised from the electrophoresisgel and analyzed through nanoLC-MSMS (Applied BiomicsHayward CA USA) Peptides and protein sequences werealigned by using Clustal Omega [18] at EBI web server andBlast at NCBI web server Alignments figures were generatedusing CLC Sequence Viewer (CLC bio Denmark)
Enzyme activity was determined spectroscopically by usingeither 22rsquo-azino-bis(3-ethylbenzothiazoline-6) sulphonic acid(ABTS ε420 = 36mM-1 cm-1 Sigma-Aldrich) or 4-hydroxy-35-dimethoxybenzaldehyde (Syringaldazine ε530 = 65mM-1 cm-1Sigma-Aldrich) [19ndash21] ABTS activity assay was carried out aspreviously described [2122] Briefly 850 microL of 5 mM ABTS insuccinic acid (25 mM pH 45) were mixed with 150 microL of therespective dilution of culture extract and the absorbance was at420 nm Syringaldazine activity assay was carried out followingthe substrate manufacturerrsquos instructions (Sigma-Aldrich) Oneunit (U) was defined as the amount of enzyme required tooxidize 1 micromol of substrate (ABTS or syringaldazine) perminute Specific activity was expressed as U mgminus1 of protein asdetermined by Lowry assay [23]
Laccase production during the fungus culture was monitoredusing syringaldazine which has demonstrated to be a specificsubstrate for detecting extracellular laccase in the absence ofhydrogen peroxide [24] Absence of hydrogen peroxide wasconfirmed by incubation with catalase Crude extracts andpurified fractions were analyzed by SDS-PAGE withoutreduction by β-mercaptoethanol and heat denaturationProteins were visualized by silver staining while laccases wereidentified by placing the SDS-PAGE gel in an ABTS solution for5 min Temperature and pH effect on laccase activity wasdetermined for the UF retentate and the purified fractions byusing ABTS The temperature effect on laccase activity wasevaluated at 30 40 50 55 60 and 70 degC at pH 50 and the pHeffect on laccase activity was evaluated at 2 25 3 4 5 6 65and 7 at 55 degC Laccase stability was evaluated by using ABTSat optimal pH and temperature for 48 h
Cyclic voltammetry analysisCyclic voltammetry (CV) analysis was performed on a
computer-controlled potentiostat (Autolab PotentiostatGalvanostat PGSTAT 128N Metrohm USA) Data wereacquired by using the software Nova version 16 Aconventional three-electrode system was used to carry out thecyclic voltammograms The used electrodes were graphite-working electrode with a surface area of 023 cm2 AgAgCl KClsaturated reference electrode (Metrohm USA) and platinum
wire counter electrode Before running each experiment thesurface of the graphite electrode was polished with aluminaand thoroughly rinsed with distilled water CV analysis wasdone to the crude extract and purified laccase fractions thatpresented the highest activity for each substrate For the test20 microL of sample were added on the polished surface of theworking electrode and allowed to dry for 60 min at roomtemperature Each experiment was performed in triplicate usingeither a solution 11 of anhydride ethanol buffer acetate (100mM pH 50) and syringaldazine to a final concentration of 02mM or a solution of 100 mM acetate buffer (pH 30) and 05mM ABTS Cyclic voltammograms were carried out in 30 mL ofworking solution with continuous agitation Voltage wasscanned at 100 mV s-1 between -02 and 1 V vs Ag|AgCl|KClsat
for syringaldazine assays while for ABTS assays voltage wasscanned at 250 mV s-1 between -02 and 12 V vs Ag|AgCl|KClsat Assays were conducted in a Faraday cage at roomtemperature (20-22degC) Temperature and pH in the reactionsolution was monitored with the Unitrode with Pt 1000(Metrohm USA)
Statistical analysisAll experiments were conducted by triplicate Data are
presented as the mean plusmn standard deviation (SD) Statisticalcomparisons were conducted by using either t-test or Duncanrsquostest for one-factor design by using StatGraphics Centurionreg v16111 (2011) Statistical differences were established for p lt005
Results
SmC and SSC production of laccase by T pubescensCoffee husk soybean pod husk and cedar sawdust were
used as substrate for the production of laccase by Tpubescens under SmC or SSC conditions while syringaldazinewas used to monitor laccase production during the fungusculture under the studied conditions Hydrogen peroxide wasnot detected in any of the evaluated samples which discardthe interference of peroxidases in the syringaldazine assayLaccase activity and glucose concentration time course for Tpubescens grown on coffee husk under SmC and SSC atdifferent copper concentrations are presented in Figure 1Although a similar time course was observed for laccaseactivity and glucose concentration of soybean pod husk andcedar sawdust cultures differences on the day of maximumlacasse activity as well as in the maximum activity value wereobserved among the agro-industrial byproducts (Table 2) Ingeneral it was noticed that laccase activity was detectablearound the seventh and fourth day of culture for SmC andSSC respectively After the eighth culture day glucose wasdepleted from the culture medium and laccase activityincreased sharply
The substrate source seemed to be the most important factorfor laccase production (Table 2) T pubescens culture oncoffee husk allowed the maximum laccase activities for bothSmC (17723plusmn372 U L-1) and SSC (20400plusmn596 U L-1) atcopper concentrations of 5 and 0 microM respectively Laccaseactivity in SSC and SmC using coffee husk as substrate was
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 4 September 2013 | Volume 8 | Issue 9 | e73721
between 147- and 663-fold higher than that observed forcultures with soybean pod husk and cedar sawdust Laccaseactivity obtained in the control culture without agro-industrialwaste presented a maximum laccase activity of 222 plusmn 088 UL-1 after 14 days of culture while the glucose was depleted onthe seventh day of culture
Copper supplementation effect on laccase activity wasevaluated for both SmC and SSC Although in a non-linearmode the laccase activity was increased by the presence ofcopper with all substrates under SmC or SSC except withcoffee husks under SSC (Table 2) In addition the requiredcopper concentration to obtain the highest laccase activityvaried depending on the used agro-industrial waste Forexample laccase production was favored in cultures withsoybean pod husk at copper concentrations of 05 mM and 20mM for SmC and SSC respectively while for cultures with
Figure 1 Laccase activity profiles during T pubescensCBS 69694 culture under SmC (a) and SSC (b) on coffeehusk Culture media was enriched with different copperconcentrations 0 (Δ) 05 () 2 () and 5 (loz) mM and laccaseactivity was determined by using syringaldazine as substrateFilled symbols refer to the glucose concentration at therespective copper concentration All the assays wereperformed in triplicatedoi 101371journalpone0073721g001
coffee husk the highest laccase activity levels for SmC andSSC was observed at copper concentrations of 5 mM and 0mM respectively
Despite that culture mode ie SmC or SSC presented aneffect on laccase production this is substrate sensitive In theassays conducted without copper it was observed that laccaseactivity was 28- and 33-fold higher in SSC than in SmC for thesoybean pod husk and coffee husk respectively Howeverwhen cedar sawdust was used no significant difference inlaccase activity between the two culture types was obtained(Table 2) Due to the low volume of liquid media and highlaccase activity in SSC when compared to SmC the SSCpresented a higher volumetric yield For all agro-industrialwaste the SSC presented a shorter time than SmC to reachthe highest laccase activity which suggest a higher productivityunder SSC conditions
These results show that coffee husk under SmC or SSCconditions could be considered as a potential alternative forlaccase production by T pubescens However soybean podhusk and cedar sawdust could be also considered as importantalternatives for laccase production especially in comparisonwith synthetic culture media [10] Finally the optimal copperconcentration to obtain the highest laccase activity depends onthe agro-industrial byproduct used for the fungus culture
Laccase purification and characterizationLaccase purification was conducted using the crude extracts
that presented the highest activities For the SmC crude
Table 2 Effect of substrate copper concentration andculture mode on laccase activity (UL-1)
Substrate SmC SSCSoybean Pod Husk + 0 mM Cu+2 446 plusmn 138 (23) 1255 plusmn 052 (13)
Soybean Pod Husk + 05 mM Cu+2
8493 plusmn 846 (23)dagger 2028 plusmn 387 (21)
Soybean Pod Husk + 2 mM Cu+2 5761 plusmn 370 (19) 13785 plusmn 071 (18)dagger
Soybean Pod Husk + 5 mM Cu+2 4492 plusmn 487 (23) 2900 plusmn 407 (18)Coffee Husk + 0 mM Cu+2 6184 plusmn 176 (23) 20400 plusmn 596 (19)dagger
Coffee Husk + 05 mM Cu+2 13763 plusmn 568 (23) 11083 plusmn 940 (20)ab
Coffee Husk + 2 mM Cu+2 12304 plusmn 373 (23)b 10278 plusmn 814 (15)b
Coffee Husk + 5 mM Cu+2 17723 plusmn 372 (23)dagger 8266 plusmn 882 (15)Cedar Sawdust + 0 mM Cu+2 1355 plusmn 095 (17) 1082 plusmn 053 (15)
Cedar Sawdust + 05 mM Cu+2 2272 plusmn 084 (17) 1902 plusmn 002 (13)Cedar Sawdust + 2 mM Cu+2 2897 plusmn 127 (12)a 2638 plusmn 068 (8)Cedar Sawdust + 5 mM Cu+2 2936 plusmn 153 (13) abdagger 3073 plusmn 152 (12)bdagger
Data in parenthesis refers to the day where was observed the maximum laccaseactivity No Statistical Difference Statistical Difference (Evaluation of substrate effecton laccase activity for both culture modes submerged (SmC) and semisolid (SSC)no copper induction considered)Equal letters inside one agro-industrial waste group refers to No StatisticalDifference no letters refers to Statistical Difference inside the group (Evaluation ofcopper effect on laccase activity for both SmC and SSC)dagger Maximum laccase activity for the used agro-industrial wastes either in SmC orSSC culture
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 5 September 2013 | Volume 8 | Issue 9 | e73721
extracts obtained with soybean pod husk + Cu+2 05 mM coffeehusk + Cu+2 5 mM and cedar sawdust + Cu+2 5 mM wereselected while for SSC crude extracts obtained with soybeanpod husk + Cu+2 2 mM coffee husk + Cu+2 0 mM and cedarsawdust + Cu+2 5 mM were selected The electrophoreticanalysis of permeates from 045 and 022 microm filtrations and theUF retentate showed the presence of two proteins with laccaseactivity while no activity was observed within the ultrafiltratedsample (Figure 2a) These proteins presented a molecularmass of about 120 kDa and 60 kDa (Figure 2a) Thepurification of the UF retentate by ionic exchangechromatography confirmed the presence of two differentfractions with laccase activity (Figure 2b) The SDS-PAGEanalysis of these fractions under non-reducing conditionsshowed that one protein with laccase activity was presented oneach fraction (Figure 2b) and were named Lac1 (120 kDa) andLac2 (60 kDa) Considering that the conditions used tomeasure the enzyme activity were those suggested for laccaseactivity and that H2O2 or manganese were not added duringthe reaction for measuring the enzyme activity these resultssuggest that both proteins are laccases but not peroxidases
NanoLC-MSMS analysis of purified fractions showed thatthe identified peptides for Lac1 (SAGSTTYNYNDPIFR andRDTVSTGNSGDNVTIR) and Lac2 (SAGSTVYNYDNPIFR andANPNFGNVGFTGGINSAI) had identity scores between 80 to100 with laccases (over 100 sequences) available at the non-redundant GenBank dataset Proteins containing thesepeptides different that laccases were not identified Similarresults were observed when peptides were evaluated atUniprot Galhaup et al [17] described two laccase activefractions from T pubescens MB 89 (CBS 69694) Lap1(UniProtKBTrEMBL Q8TG93) and Lap2 (UniProtKBTrEMBLQ8TG94) which share a 70 identity (Figure 3a) Sequencealignment of Lac1 and Lac2 peptides with Lap1 and Lap2sequences respectively showed that the peptides arepresented within T pubescens MB 89 laccase sequencesthough some differences were observed for the Lac1 peptide(Figure 3b) This result suggests that the purified fractions arelaccase isoenzymes
Protein concentration and volumetric and specific laccaseactivities at different purification stages are presented in Table3 Lac1 presented lower laccase activity than Lac2 in a ratiothat ranges from 080- to 38-fold depending on the substratesource On the other hand laccase activity of the UF retentatefrom soybean pod husk culture was from 45- to 64-fold higherthan that of the purified fractions while for cedar sawdust andcoffee husk the laccase activity was from 46- to 120-fold andfrom 33- to 163-fold higher than the purified fractionsrespectively (Table 3) The purified Lac2 fractions from coffeehusk and cedar sawdust cultures showed the highest specificactivity levels (Table 3) The volumetric activities of UFretentates were higher than that obtained with purified fractions(Table 4)
Data in parenthesis shows the temperature or pH at whichrelative laccase activity reached 100
Temperature and pH effect on laccase activity wasdetermined for the UF retentate and Lac1 and Lac2 fractionsobtained along the purification of the enzymatic crude extracts
that resulted after the SSC of T pubescens on the selectedagro-industrial wastes Laccase activity for the UF retentateunder different temperature and pH conditions is presented inFigure 4 and Table 4 The maximum relative activity of theevaluated UF retentates was observed at 50-55 degC and at apH range from 20 to 40 However UF retentate from cedar
Figure 2 Purification of lacasses produced by submergedculture of T pubescens using coffee husk assubstrate All samples were analyzed by SDS-PAGE undernon-reducing conditions and stained with ABTS No heatdenaturation of the samples was conducted (a) The crudeextract (1) was centrifuged and filtered through a Whatman No1 filter (2) 045 microM membrane (3) 022 microm membrane (4) andultrafiltrated through a 10 kDa cut-off membrane permeate (5)and retentate (6) (b) UF retentate was purified by anionicexchange chromatography Lac1 represent the unboundfraction while Lac2 represent the eluted fraction The arrowshows the point when protein elution was started (c) Purifiedfractions (1) crude extract (2) Lac1 and (3) Lac2 MWMolecular Weight Similar results were observed for crudeextracts produced by using soybean pod husk and cedarsawdustdoi 101371journalpone0073721g002
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 6 September 2013 | Volume 8 | Issue 9 | e73721
sawdust culture behaved different to the UF retentate fromcoffee husk and soybean pod husk cultures In this sense the
UF retentate from cedar sawdust culture presented the lowestenzyme activity at temperatures le 40 degC but at 70 degC
Figure 3 Aligment of reported lacasses sequences and Lac1 and Lac2 peptides (a) Alignment of Lap1 (UniProtKBSwiss-Prot Q8TG93) and Lap2 (UniProtKBSwiss-Prot Q8TG94) from T pubescens CBS 69694 (b) Alignment of Lac1 and Lac2 peptideswith Lap1 and Lap2 respectively and tryptic peptides described by Shleev et al 2007 Alignments were carried out using ClustalOmega and figures were using CLC Sequence Viewer Different residues are colored in reddoi 101371journalpone0073721g003
Table 3 Laccase activity during the purification of crude extracts resulted from T pubescens culture on agro-industrialwastes under submerged (SmC) and semisolid (SSC) cultures
Soybean pod husk Coffee husk Cedar sawdust
Protein(mg mL-1)
VolumetricActivity (UL-1)
SpecificActivity (Umg-1
protein)Recovery()
Protein(mg mL-1)
VolumetricActivity (UL-1)
SpecificActivity (Umg-1
protein)Recovery()
Protein(mg mL-1)
VolumetricActivity (UL-1)
SpecificActivity (Umg-1
protein)Recover()
SmCCrudeExtract
0869 52888 0608 100 0414 152708 3688 100 0431 10735 0249 100
045 micromFiltration
0736 46549 0632 88 0334 117892 3529 77 0444 12053 0271 112
UF retentate 1010 78731 0779 74 0378 195263 5165 64 0510 25905 0507 120
Lac1 0494 5471 0110 1 0232 2399 0103 02 0222 838 0037 08
Lac2 0310 20661 0666 4 0116 107240 9252 7 0040 15234 3837 14 SSCCrudeExtract
1146 164966 1439 100 0596 154907 2601 100 1065 23203 0217 100
045 micromFiltration
0776 128392 1654 77 0474 115736 2439 75 0806 25824 0320 111
UF retentate 0973 335055 3443 101 0428 161750 3779 52 1134 51101 0450 110
Lac1 0097 8854 0912 2 0104 1514 0145 03 0042 1315 0313 2
Lac2 0168 48988 2915 10 0131 82298 6282 18 0072 16162 2244 23
740 ABTS was used to determine laccase activity
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 7 September 2013 | Volume 8 | Issue 9 | e73721
presented the highest enzyme activity ~90 of its initial valuein comparison to the other two UF retentates (Figure 4)
The temperature and pH effect on laccase activity for Lac1and Lac2 is presented in Figure 5 The maximum relativeactivity for both fractions was presented at a pH from 20 to 40regardless of the agro-industrial waste as observed for the UFretentate However at pH gt 40 Lac1 showed a sharperdecrease in its activity than that observed for Lac2Nevertheless Lac2 purified from cedar sawdust culturepresented a different performance This fraction showed themaximum activity at pH 50 while a sharp drop was observedat pHs above or below this value
Laccase activity profiles showed that Lac1 and Lac2presented the maximum enzyme activity at 55 and 60 degCrespectively (Table 4) It was observed that Lac1 from coffeehusk culture presented a lower decrease in enzyme activity attemperatures le 50 degC or ge 60 degC than Lac2 On the contraryLac2 from soybean husk or cedar sawdust cultures presenteda higher relative laccase activity than Lac1 at temperatures le55 degC The purified fractions from soybean pod husk and cedarsawdust cultures presented the lowest relative activity at 30 degCand 65 degC
Table 4 Optimum temperatures and pHs for maximumlaccase activities in UF retentate Lac1 and Lac2 obtainedfrom semisolid cultured T pubescens on agro-industrialwastes
EnzymaticFraction Temperature pH
VolumetricActivity (UL-1)
SpecificActivity (Umg-1 protein)
VolumetricActivity (UL-1)
SpecificActivity (Umg-1
protein)SoybeanPod Husk +2 mM Cu+2
148000plusmn1178(55)
1521plusmn0012182037plusmn3025(2-3)
1871plusmn0031
Lac122925plusmn917(60)
2363plusmn009536135plusmn831(23)
3725plusmn0085
Lac232638plusmn1558(55)
1942plusmn009228713plusmn274(23)
1709plusmn0016
CoffeeHusk + 0mM Cu+2
113740plusmn8289(50-55)
3008plusmn0219290666plusmn1039(2-3)
7689plusmn0275
Lac1 889plusmn039 (55) 0038plusmn0002 1779plusmn028 (23) 0077plusmn0001
Lac234194plusmn2789(60)
2947plusmn024033150plusmn526 (2ndash4)
2857plusmn0045
CedarSawdust +5 mM Cu+2
257666plusmn1201(55)
5052plusmn0024344634plusmn1043(2-3)
6757plusmn0204
Lac121370plusmn472(55)
0962plusmn002129117plusmn943(23)
1311plusmn0042
Lac255166plusmn1656(60)
1379plusmn0414 55166plusmn1656 (5) 1379plusmn0414
ABTS was used to determine laccase activity
Laccase stabilityFor selected enzymatic extracts and purified fractions (Table
4) a stability test for 48 h at their optimal pH and temperaturewas conducted Time course of the relative laccase activity forcrude extract and purified fractions from coffee husk and cedarsawdust cultures are presented in Figure 5 Similar resultswere observed for crude extract and purified fractions fromsoybean pod husk culture (data no shown)
It was observed that the activity loss depends on thesubstrate source used to grow T pubescens An activity loss of55 was observed for crude extracts and purified fractionsfrom coffee husk and soybean pod husk cultures while anactivity loss of 70 was observed for the crude extract andpurified fractions from cedar sawdust culture NeverthelessLac2 showed the lowest activity reduction with values of 10 40and 55 for coffee husk soybean pod husk and cedarsawdust respectively while Lac1 presented an activity loss of34 50 and 60 for coffee husk soybean pod husk and cedarsawdust respectively Despite the differences in laccaseactivity among agro-industrial wastes and between crudeextracts and purified fractions all the profiles showed a sharp
Figure 4 Effect of temperature (a) and pH (b) on laccaseactivity for the UF retentate obtained by semisolid cultureof T pubescens on coffee husk () soybean pod husk (loz)and cedar sawdust (Δ) Laccase stability was evaluated byusing ABTS All the assays were performed in triplicatedoi 101371journalpone0073721g004
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 8 September 2013 | Volume 8 | Issue 9 | e73721
decrease during the first 8 h while a reduced activity loss wasobserved during the last 24 h of the assay (Figure 6)
Laccase electrochemical characterizationFor selected crude extracts and purified fractions a cyclic
voltammetry characterization was conducted For eachmediator syringaldazine or ABTS similar cyclic
Figure 5 Effect of temperature () and pH () on enzymeactivity for Lac1 (solid line) and Lac2 (dashed line)obtained after the purification of the enzymatic crudeextract produced by semisolid culture of T pubescens oncoffee husk (a) soybean pod husk (b) and cedar sawdust(c) Laccase stability was evaluated by using ABTS All theassays were performed in triplicatedoi 101371journalpone0073721g005
voltammograms were obtained for the different crude extractsand purified fractions Voltammograms of the redox reaction forsyringaldazine (Figure 7a) and ABTS (Figure 6b) arepresented Two anionic and cationic peaks were observedwhen syringaldazine was used as mediator The first pair ofpeaks showed a reversible performance with a mean Ia Icratio of 109plusmn006 106plusmn005 and 098plusmn002 for the crudeextracts Lac1 and Lac2 respectively The second pair ofpeaks showed quasi-reversible performance and the cathodicpeak was presented at a potential of +062 V (vs Ag|AgCl)while the anodic peak was presented at a potential of +060 V(vs Ag|AgCl) No significant difference on the reductionpotential (Eo) for syringaldazine was observed for the crudeextracts or the purified fractions obtaining an Eo of0314plusmn0008 V (vs Ag|AgCl) However the peak potentialseparation (ΔEp) showed a different profile depending on thesubstrate used for the culture While not significant difference(p gt 005) on ΔEp value was observed between crude extract(0045plusmn0005 V vs Ag|AgCl) Lac1 (0046plusmn0005 V vs Ag|AgCl)and Lac2 (0028plusmn0001 V vs Ag|AgCl) from cultures with cedarsawdust and soybean pod husk the ΔEp was considerablyhigher for the crude extract (0083plusmn0004 V vs Ag|AgCl) Lac1(0068plusmn0003 V vs Ag|AgCl) and Lac2 (0051plusmn0005 V vs Ag|AgCl) from the coffee husk culture
For reactions conducted with ABTS and the different crudeextracts and purified fractions the voltammograms exhibitedone pair of cathodic and anodic peaks (Figure 7b) whichcorresponds to the redox of ABTSABTS+ (IaIc) with an Eo of0487plusmn0006 V (vs Ag|AgCl) This process was reversiblepresenting a mean Ia Ic ratio of 099plusmn002 In addition asecond cathodic peak was presented at +083 V (vs Ag|AgCl)when ABTS redox reaction was conducted using the crudeextract and the purified fractions This performance agrees withprevious reports in which the laccase activity cannot beobserved at potential values of the second reversible oxidation
Figure 6 Laccase stability profiles at optimal pH andtemperature conditions for crude extracts (solid line) Lac1(dashed line) and Lac2 (spotted line) obtained fromsemisolid culture of T pubescens on coffee husk (loz) andcedar sawdust (bull) Laccase stability was evaluated by usingABTS All the assays were performed in triplicatedoi 101371journalpone0073721g006
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 9 September 2013 | Volume 8 | Issue 9 | e73721
of ABTS ABTS+ABTS2+ due to this catalytic process cannotbe detected by cyclic voltammetry [2526] Neverthelessvoltammograms for laccase crude extract from a soybean podhusk culture presented the second anodic peak which inaddition to the second cathodic peak corresponds to the redoxof ABTS+ABTS2+ (IIaIIc) with an Eo of 0942plusmn0004 V (vs Ag|AgCl) Contrary to the reactions conducted usingsyringaldazine as substrate the ΔEp did not present asignificant difference for the crude extracts and purifiedfractions 0248plusmn0016 V (vs Ag|AgCl)
Discussion
In this study we evaluated the effect of the culture mode ieSmC and SSC on laccase production by T pubescens CBS69694 using three different agro-industrial wastes (coffeehusk soybean pod husk and cedar sawdust) Furthermore theeffect of Cu+2 supplementation on laccase production was alsoevaluated and the crude extracts and the purified fractionswere partially characterized
Figure 7 Cyclic voltammograms of (a) syringaldazine atpH 50 using the crude extract and purified laccasefractions from semisolid culture of T pubescens on coffeehusk or (b) ABTS at pH 30 using the crude extract andpurified laccase fractions from semisolid culture of Tpubescens on soybean pod husk Nomenclature onlymediator (dotted line) crude extract (solid line) Lac1 fraction(dashed line) and Lac2 fraction (dashed and dotted line)doi 101371journalpone0073721g007
Substrate source for laccase production showed to be one ofthe most important factors for laccase production (Table 2)Cultures with coffee husk as substrate presented the highestlaccase production levels The observed differences in laccaseactivity for each substrate could be owed to their chemicalcomposition Coffea arabica is the coffee variety raised inColombia and its husk has a reported composition of 2308cellulose (2076 glucose and 183 cellobiose) 2385hemicellulose (1356 xylose 523 arabinose 256 aceticacid and 195 glucuronic acid) 2828 total lignin and071 ashes [27] In addition coffee husk contents 13caffeine 45 tannins and 124 pectins [28] On the otherhand Glycine max is the soybean species cultured inColombia and its pod husk composition varies from 504 to62 crude protein 608 to 143 hemicellulose 3715 to4208 acid detergent fiber 4436 to 6015 neutral detergentfiber 850 to 893 acid detergent lignin and 2074 to 2106uronic acid [29ndash31] Finally for cedar sawdust the majorcomponents are glucose (426-444) xylose (46-195)cellulose (383-410) klason lignin (227-278) and acidlignin (40-43) [32ndash34] Although lignin is one of the mostimportant compounds associated with laccase production bywhite-rot fungi we did not observe a clear correlation betweenlignin composition percentage and laccase activity In thatsense the high laccase activity observed for the cultures thatuse coffee husk could be associated with its content of caffeineand tannins which have been identified as potent inducers oflaccase gene expression [173536] In fact caffeine has beenused as inducer for laccase production by Rhizoctonia solaniobtaining an increase of 41-fold in laccase activity [37] Similarresults were also reported for the production of Coloriopsisgallica laccase [38]
Laccase gene expression is mainly up regulated by Cu2+although Mn2+ Fe3+ heavy metals 26-dimethoxy-14-benzoquinone H2O2 amphotericin B syringic acid tannic acidTween 80 soybean oil aromatic compounds andmicroclimatic changes (ie lower temperature and osmoticpressure) are also recognized as potent inducers [173536] Ingeneral Cu2+ supplementation increased the laccase activity inboth SmC and SSC However Cu2+concentration required toobtain the highest laccase activity varied depending on agro-industrial waste These differences can be attributed to thecopper presented in the agro-industrial waste and the copperavailability in the medium Copper can be found in amountsthat range from 156 to 180 mg kg-1 of coffee husk [39] andfrom 26 to 67 mg kg-1 of soybean pod husk (or soybean hull)[3940] Despite that no reports indicate the copper content inuntreated cedar wood an average of 10 mg kg-1 of wood hasbeen reported for wood used as biomass in six different Europecountries [41] This allows to consider the cedar sawdust as thesubstrate with the lowest content of copper On the other handcopper adsorption by sawdust and coffee husk has beenreported obtaining a Langmuir based maximum adsorptioncapacity (Qmax mg g-1 adsorbent) of 845 and 75 respectively[4243] Although no adsorption studies has been reportedusing untreated soybean pod husk Šćiban et al [44] reporteda Qmax for copper adsorption of 0085 mmol g-1 adsorbent (~ 54mg g-1) using treated soybean straws Based on this
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 10 September 2013 | Volume 8 | Issue 9 | e73721
information it can be expected that a higher amount ofexogenous copper would be required for laccase induction oncultures with cedar sawdust than with soybean pod husk andcoffee husk due to its lower content of copper In addition dueto the high copper-adsorption capacity of sawdust the amountof available copper in the culture would be reduced allowingthe laccase production at higher copper concentrations withoutreaching levels that might be toxic for the fungus This fact canbe observed in the SSC (Table 4) However for the SmC thisbehavior is not maintained for the cultures with soybean podhusk and coffee husk (Table 4) where the maximum laccaseactivity was obtained with a copper concentration of 05 and 5mM respectively This could be owed to a lower contactbetween the fungus and the substrate in the SmC whencompared to the SSC which is close related to the transport ofnutrients presented in the substrates
Production of extracellular laccases has been reported in alarge number of species of white-rot fungi strains grown onnatural substrates (Table 1) Although a laccase activity profileis not clearly observed based on the agro-industrial wasteculture mode and fungus strain most cases in which SmC andSSC were conducted using the same substrate and fungusstrain showed a higher laccase activity under SmC conditionsIn this study was observed a substrate dependentperformance where the most favorable condition to obtain thehighest laccase activity was the SSC The laccase activitiesobtained in the present study are comparable and in somecases higher than those reported for Pleurotus ostreatusPhanerochaete chrysosporium Pycnoporus cinnabarinus Tversicolor and Ganoderma sp cultured under SC and SmCconditions and using different agro-industrial wastes (Table 1)Semi-solid cultures using different agro-industrial wastes havebeen reported using T versicolor Phlebia radiata Tpubescens P ostreatus and C unicolor Rodriguez et al [16]screened different supports for laccase production by Tversicolor under SSC They obtained the highest laccaseactivity (1200 U L-1) when in the culture medium was presentedbarley bran In addition they were able to obtain a laccaseactivity of 1700 U L-1 when the culture was supplemented withxylidine as an inducer These results were obtained after 17 or18 days of culture and ABTS was used to determine theenzyme activity Maumlkelauml et al [4546] have studied theexpression of the genes involved in laccase production Pr-lac1and Pr-lac2 presented in P radiata and evaluate laccaseproduction in SSC using milled alder wood (Alnus incana) assubstrate They reported a maximum laccase activity of 3 microkatL-1 (~180 U L-1) at the 14th day of culture and usingsyringaldazine for its detection Osma et al [15] have studiedthe morphological changes and laccase production of strains Tpubescens P ostreatus C unicolor and T versicolor grownon wheat bran flakes using SSC They obtained a maximumlaccase production between 1397 and 2778 U L-1 after 10 and13 days of culture This enzyme activity is strain dependentand ABTS was used for its quantification Our results show agood correlation with reported data The highest laccaseactivity was obtained when T pubescens was grown on coffeehusk under SSC without copper (204 U L-1 evaluated withsyringaldazine or 1549 U L-1 evaluated with ABTS) which is in
some cases higher than reported ones This shows that underthe studied conditions the production of the laccase crudeextract is favored in SSC using coffee husk without the additionof copper which could favors the cost of the productionprocess
It has been reported for white rot-fungi the production ofmultiple laccase isoenzymes which are encoded by differentgenes and show different expression profiles stabilitiessubstrate affinities and molecular masses (ranging from 35 to140 kDa) [3647ndash49] Two isoforms have been identified in Tpubescens strains two monomeric (Lac1 and Lac2) enzymesof 67 kDa but with different pI (53 and 51) [50] and twolaccase-activity fractions (Lap1 and Lap2) with different pI (gt30 and 26) from which Lap2 is a monomeric enzyme of 65kDa [17] In this study we identify two laccase isoenzymesfrom T pubescens CBS 69694 that have different molecularmasses (120 and 60 kDa) and pI The purifications resultssuggest a more acidic pI for Lac1 (le 4) than Lac2 (ge 6) whichdiffers from a previous report for this fungus in which twofractions with laccase activity were obtained Lap1 and Lap2with pI gt 3 and of 26 respectively [17] Since in both casessimilar purification protocols were used and Lac1 and Lac2were recovered from the unbound and bound fractionsrespectively as Lap1 and Lap2 [17] this difference could bedue to the culture conditions used in each case (ie thepresence of agro-industrial wastes) Nevertheless the nanoLC-MSMS analysis showed that peptides of Lac1 and Lac2 werepresented in Lap1 (UniProtKBSwiss-Prot Q8TG93) and Lap2(UniProtKBSwiss-Prot Q8TG94) which have a predicted pI of43 and 47 respectively Lac1 and Lac2 peptides werepresented in Laccase-2 (UniProtKBSwiss-Prot Q99046) andLaccase-1 (UniProtKBSwiss-Prot Q99044) from T villosarespectively with 80 to 100 identity These T villosa laccaseisoenzymes have a predicted pI of 47 (laccase-1) and 60(laccase-2) and a 77 identity among them In addition Tvillosa laccase-2 is reported as a homodimer protein whilelaccase-1 is reported as monomeric protein Alignment of Lac1-Lap2 and Lac2-Lap1 from T pubescens CBS 69694 and Tvillosa showed a 96 and 70 identity respectively Finallysimilar peptides were identified for Lac1 and Lac2 from Tpubescens (Schumach) Pilaacutet (Syn Coriolus pubescens(Schum ex Fr) Queacutel) BCB 923-2 (Figure 3b) [51] Theseresults suggest that the purified fractions correspond to laccaseisoenzymes that differ from previously reported ones for this Tpubescens strain
As mentioned above laccases isoenzymes present differentstabilities and substrate affinities This behavior was alsoobserved for the laccase isoenzymes purified from Tpubescens with Lac1 showing lower laccase activity thanLac2 with ABTS as substrate However the difference inactivity between these isoenzymes depended on the agro-industrial waste used for the T pubescens growth suggestinga probable effect of the substrate composition on the laccasephysicochemical properties In addition it was observed thatthe total laccase activity presented in the UF retentates ishigher than the laccase activity of the purified fractionsregardless of the agro-industrial byproduct used for the Tpubescens growth This suggests a synergistic effect of Lac1
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 11 September 2013 | Volume 8 | Issue 9 | e73721
and Lac2 as have been reported for laccases obtained fromTrametes and Pleurotus strains [175253]
The maximum relative activity for the UF retentate waspresented at a pH from 20 to 40 regardless of the agro-industrial byproduct with a monotonical decrease at pHs abovethis range The relation between pH and type of electron donormolecule explains this behavior on laccase activity It has beenreported that when the substrate is an organic hydrogen donorie phenols the optimal pH ranges from 35 to 60 while thelaccase activity monotonically decreases from pH 25 to 70 ifthe substrate is an electron donor like ABTS [54] Despite thatat different pHs most of the purified laccase fractionspresented a similar performance as the UF retentates the Lac2fraction obtained from cedar sawdust differed from the previousobservation For this fraction the maximum activity wasobserved at pH 50 and despite that at acidic pH (ge 30) therelative activity was higher than 80 at lower pHs (20) itdecreased almost in a 50 Nevertheless at basic pHs therelative laccase activity presented a sharp decreased that iswell correlate with the other laccase fractions We are not ableto attribute this particular trend to the presence of multiplelaccase isoenzymes in the fraction since the SDS-PAGErevealed only one band when stained with ABTS and from theNanoLC-MSMS analysis no difference in the number ofpeptides and their sequence was detected We speculate thatthis particular behavior can be owed to a compound added inthe pre-treatment of the cedar wood that might be used forpreserving it that might affect the activity of Lac2 and that wasnot completely removed during the purification
Temperature stability evaluation showed that Lac1 is themost thermolabile fraction while the laccase activity reductionin the crude extracts did not seem to be the result of anadditive effect of the activity loss of the purified fractionsAlthough there are no reports of laccase stability under theselected conditions these results agree with previous reportsthat state that fungal laccases rapidly decrease their activity attemperatures near 60 degC and thermal stability correlates withthe temperature range of the growth of the source organism[55] In addition we previously reported for a laccase crudeextract obtained from culturing T pubescens on coffee husk anactivity loss of 26 and 10 at 60 and 50 degC respectivelyafter 8 h at pH 60 using syringaldazine as substrate [48]
Finally an electrochemical characterization was carried outfor selected crude extracts and purified fractions The results
showed a difference between the electrochemicalperformances of extracts and purified fractions withsyringaldazine or ABTS These differences could be owed tothe different mechanisms for electron transfer as discussedabove However the overall performance of the obtained cyclicvoltammograms for ABTS and syringaldazine agrees withresults reported by Fernaacutendez-Saacutenchez et al [25] who used asaturated calomel reference electrode
Conclusions
We have shown the potential of coffee husk soybean podhusk and cedar sawdust as substrates for the laccaseproduction In several countries like Brazil and Colombia theproduction of coffee and soybean is of great importance andthe generated volume of residues is high The results showedthat semi-solid culture of T pubescens using coffee huskallows to obtain an important amount of laccase which couldbe favored by the phenolic-compounds ie tannins presentedin coffee husk While supplementation of the culture mediumwith copper favored the production of laccase in submergedand semi-solid culture using soybean pod husk and cedarsawdust semi-solid culture using coffee husk which presentedthe highest laccase volumetric yield did not require coppersupplementation This is an economic advantage that shouldbe considered when scaling up the process In this study Tpubescens showed its ability to produce two independentlaccases regardless of the used substrate However a higherlaccase activity was observed for crude extracts rather than forpurified fractions This suggests that Lac1 and Lac2 might havea synergistic effect on the final laccase activity rather than anadditive effect Based on this study the investigation of laccaseproduction by evaluating other agro-industrial wastes in whichphenolic-compounds constitute an important percentage oftheir composition is of great interest to avoid the use ofinducers that would increase the production cost
Author Contributions
Conceived and designed the experiments JFO CJAD OFSPerformed the experiments JCG SCM AR Analyzed the dataJCG SCM AR JFO CJAD OFS Wrote the manuscript AR JFOCJAD OFS
References
1 Vandamme EJ (2009) Agro-industrial residue utilization for industrialbiotechnology products In P NigamA Pandey Biotechnology for Agro-Industrial Residues Utilisation Netherlands Springer Verlag pp 3-11
2 Martiacutenez AT Speranza M Ruiz-Duentildeas FJ Ferreira P Camarero S etal (2005) Biodegradation of lignocellulosics microbial chemical andenzymatic aspects of the fungal attack of lignin Int Microbiol 8195-204 PubMed 16200498
3 Saacutenchez O Sierra R Almeacuteciga-Diacuteaz CJ (2011) Delignification processof agro-industrial aastes an alternative to obtain fermentablecarbohydrates for producing fuel In M Manzanera Alternative FuelIntech 111-154
4 Wesenberg D Kyriakides I Agathos SN (2003) White-rot fungi andtheir enzymes for the treatment of industrial dye effluents BiotechnolAdv 22 161-187 doi101016jbiotechadv200308011 PubMed14623049
5 Morozova OV Shumakovich GP Shleev SV Yaropolov YI (2007)Laccase-mediator systems and their applications A review ApplBiochem Microbiol 43 523-535 doi101134S0003683807050055
6 Baldrian P (2006) Fungal laccases - occurrence and properties FEMSMicrobiol Rev 30 215-242 doi101111j1574-4976200500010xPubMed 16472305
7 Rodriacuteguez Couto S Herrera JLT (2006) Industrial and biotechnologicalapplications of laccases A review Biotechnol Adv 24 500-513 doi101016jbiotechadv200604003 PubMed 16716556
8 Osma JF Toca-Herrera JL Rodriacuteguez-Couto S (2010) Uses oflaccases in the food industry Enzyme Res 2010 2010 918761PubMed 21048873
9 Rodriguez S (2008) Exploitation of biological wastes for the productionof value-added products under solid-state fermentation conditionsBiotechnol J 3 859-870 doi101002biot200800031 PubMed18543242
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 12 September 2013 | Volume 8 | Issue 9 | e73721
10 Osma JF Toca-Herrera JL Rodriacuteguez-Couto S (2011) Cost analysis inlaccase production J Environ Manage 92 2907-2912 doi101016jjenvman201106052 PubMed 21775046
11 Jain A Morlok CK Henson JM (2013) Comparison of solid-state andsubmerged-state fermentation for the bioprocessing of switchgrass toethanol and acetate by Clostridium phytofermentans Appl MicrobiolBiotechnol 97 905-917 doi101007s00253-012-4511-4 PubMed23111595
12 Singhania RR Patel AK Soccol CR Pandey A (2009) Recentadvances in solid-state fermentation Biochem Eng J 44 13-18 doi101016jbej200810019
13 Economou CN Makri A Aggelis G Pavlou S Vayenas DV (2010)Semi-solid state fermentation of sweet sorghum for the biotechnologicalproduction of single cell oil Bioresour Technol 101 1385-1388 doi101016jbiortech200909028 PubMed 19781936
14 Rodriacuteguez CS Santoro R Cameselle C Sanroman A (1997) Laccaseproduction in semi-solid cultures of Phanerochaete chrysosporiumBiotechnol Lett 19 995-998 doi101023A1018495216946
15 Osma JF Moilanen U Toca-Herrera JL Rodriacuteguez-Couto S (2011)Morphology and laccase production of white-rot fungi grown on wheatbran flakes under semi-solid-state fermentation conditions FEMSMicrobiol Lett 318 27-34 doi101111j1574-6968201102234xPubMed 21291496
16 Couto SR Gundin M Lorenzo M Sanroman MN (2002) Screening ofsupports and inducers for laccase production by Trametes versicolor insemi-solid-state conditions Proc Biochem 38 249-255 doi101016S0032-9592(02)00087-0
17 Galhaup C Goller S Peterbauer CK Strauss J Haltrich D (2002)Characterization of the major laccase isoenzyme from Trametespubescens and regulation of its synthesis by metal ions Microbiology148 2159-2169 PubMed 12101303
18 Sievers F Wilm A Dineen D Gibson TJ Karplus K et al (2011) Fastscalable generation of high-quality protein multiple sequencealignments using Clustal Omega Mol Syst Biol 7 539 PubMed21988835
19 Childs RE Bardsley WG (1975) The steady-state kinetics of peroxidasewith 22-azino-di-(3-ethyl-benzthiazoline-6-sulphonic acid) aschromogen Biochem J 145 93-103 PubMed 1191252
20 Harkin JM Obst JR (1973) Syringaldazine an Effective Reagent forDetecting Laccase and Peroxidase in Fungi Experientia 29 382-387
21 Nikupaavola ML Raaska L Itavaara M (1990) Detection of white-rotfungi by a non-roxic stain Mycol Res 94 27-31 doi101016S0953-7562(09)81260-4
22 Gonzalez LF Sarria V Sanchez OF (2010) Degradation ofchlorophenols by sequential biological-advanced oxidative processusing Trametes pubescens and TiO(2)UV Bioresour Technol 1013493-3499
23 Waterborg JH (2009) The Lowry method for protein quantitation In JWalker The Protein Protocols Handbook Humana Press pp 7-10
24 Harkin JM Obst JR (1973) Syringaldazine an effective reagent fordetecting laccase and peroxidase in fungi Cell Mol Life Sci 29381-387 doi101007BF01926734
25 Fernaacutendez-Saacutenchez C Tzanov T Guumlbitz GM Cavaco-Paulo A (2002)Voltammetric monitoring of laccase-catalysed mediated reactionsBioelectrochemistry 58 149-156 doi101016S1567-5394(02)00119-6PubMed 12414320
26 Bourbonnais R Leech D Paice MG (1998) Electrochemical analysis ofthe interactions of laccase mediators with lignin model compoundsBiochim Biophys Acta 1379 381ndash390 doi101016S0304-4165(97)00117-7 PubMed 9545600
27 Ferraz F Silva S (2009) Characterization of coffee husk biomass forbiotechnological purposes N Biotechnol 25 S256-S256 doi101016jnbt200906573
28 Pandey A Soccol CR Nigam P Brand D Mohan R et al (2000)Biotechnological potential of coffee pulp and coffee husk forbioprocesses Biochem Eng J 6 153-162 doi101016S1369-703X(00)00084-X PubMed 10959086
29 Sruamsiri S Silman P (2008) Nutritive composition of soybean by-products and nutrient digestibility of soybean pod husk Maejo Int J SciTechnol 2 568-576
30 Romkaew J Nagaya Y Goto M Suzuki J Umezaki T (2008) Poddehiscence in relation to chemical components of pod shell in soybeanPlant Prod Sci 11 278-282 doi101626pps11278
31 Nakamanee G Polbumroong T Phaikaew C (2003) Alternative use ofcrop waste to increase milk production Proceedings of the 8th Meetingof the Regional Working Group on Grazing and Feed Resources forSoutheast Asia Kuala Lumpur Malaysia 66 - 70
32 Kurek B Gaudard F (2000) Oxidation of spruce wood sawdust byMnO(2) plus oxalate a biochemical investigation J Agric Food Chem48 3058-3062 doi101021jf000015g PubMed 10898665
33 Weil J Brewer M Hendrickson R Sarikaya A Ladisc MR (1998)Continuous pH monitoring during pretreatment of yellow poplar woodsawdust by pressure cooking in water Appl Biochem Biotechnol 70-7299-111 doi101007BF02920127
34 Weil J Sarikaya A Rau S-L Goetz J Ladisch CM et al (1997)Pretreatment of yellow poplar sawdust by pressure cooking in waterAppl Biochem Biotechnol 68 21-40 doi101007BF02785978
35 Dekker RF Barbosa AM Giese EC Godoy SD Covizzi LG (2007)Influence of nutrients on enhancing laccase production byBotryosphaeria rhodina MAMB-05 Int Microbiol 10 177-185 PubMed18075999
36 Xiao YZ Hong YZ Li JF Hang J Tong PG et al (2006) Cloning ofnovel laccase isozyme genes from Trametes sp AH28-2 and analysesof their differential expression Appl Microbiol Biotechnol 71 493-501doi101007s00253-005-0188-2 PubMed 16283298
37 Crowe JD Olsson S (2001) Induction of laccase activity in Rhizoctoniasolani by antagonistic Pseudomonas fluorescens strains and a range ofchemical treatments Appl Environ Microbiol 67 2088-2094 doi101128AEM6752088-20942001 PubMed 11319086
38 Yaguumle S Terroacuten MC Gonzaacutelez T Zapico E Bocchini P et al (2000)Biotreatment of tannin-rich beer-factory wastewater with white-rotbasidiomycete Coriolopsis gallica monitored by pyrolysisgaschromatographymass spectrometry Rapid Commun Mass Spectrom14 905-910 doi101002(SICI)1097-0231(20000530)1410 PubMed10825255
39 Gowda NKS Ramana JV Prasad CS Singh K (2004) Micronutrientcontent of certain tropical conventional and unconventional feedresources of southern India Trop Anim Heatlth Pro 36 77-94 doi101023BTROP0000009522309491d PubMed 14979561
40 DePeters EJ Fadel JG Arana MJ Ohanesian N Etchebarne MA et al(2000) Variability in the chemical composition of seventeen selectedby-product feedstuffs used by the California dairy industry Prof AnimSci 16 69-99
41 Obernberger I Thek G (2004) Physical characterisation and chemicalcomposition of densified biomass fuels with regard to their combustionbehaviour Biomass Bioenerg 27 653-669 doi101016jbiombioe200307006
42 Larous S Meniai A-H Lehocine MB (2005) Experimental study of theremoval of copper from aqueous solutions by adsorption usingsawdust Desalination 185 483ndash490 doi101016jdesal200503090
43 Oliveira WE Franca AS Oliveira LS Rocha SD (2008) Untreatedcoffee husks as biosorbents for the removal of heavy metals fromaqueous solutions J Hazard Mater 152 1073ndash1081 doi101016jjhazmat200707085 PubMed 17804159
44 Šciban M Klašnja M Škrbic B (2008) Adsorption of copper ions fromwater by modified agricultural by-products Desalination 229 170ndash180doi101016jdesal200708017
45 Maumlkelauml MR Hildeacuten KS Hakala TK Hatakka A Lundell TK (2006)Expression and molecular properties of a new laccase of the white rotfungus Phlebia radiata grown on wood Curr Genet 50 323-333 doi101007s00294-006-0090-1 PubMed 16927090
46 Maumlkelauml MR Lundell T Hatakka A Hildeacuten K (2013) Effect of coppernutrient nitrogen and wood-supplement on the production of lignin-modifying enzymes by the white-rot fungus Phlebia radiata Fungal Biol117 62-70 doi101016jfunbio201211006 PubMed 23332834
47 Dwivedi UN Singha P Pandeya VP Kumara A (2011) Structurendashfunction relationship among bacterial fungal and plant laccases J MolCatal B Enzym 68 117-128 doi101016jmolcatb201011002
48 Gaitan IJ Medina SC Gonzaacutelez JC Rodriacuteguez A Espejo AacuteJ et al(2011) Evaluation of toxicity and degradation of a chlorophenol mixtureby the laccase produced by Trametes pubescens Bioresour Technol102 3632ndash3635 doi101016jbiortech201011040 PubMed21115244
49 Karp SG Faraco V Amore A Birolo L Giangrande C et al (2012)Characterization of laccase isoforms produced by Pleurotus ostreatusin solid state fermentation of sugarcane bagasse Bioresour Technol114 735-739 doi101016jbiortech201203058 PubMed 22487128
50 Nikitina OV Shleev SV Gorshina ES Rusinova TV Serezhenkov VAet al (2005) Isolation and purification of enzymes from ligninolyticcomplex of the basidial fungus Trametes pubescens (Schumach) Pilatand study of their properties Biochemistry (Mosc) 70 1274-1279 doi101007s10541-005-0259-0
51 Shleev S Nikitina O Christenson A Reimann CT Yaropolov AI et al(2007) Characterization of two new multiforms of Trametes pubescenslaccase Bioorg Chem 35 35-49 doi101016jbioorg200608001PubMed 16989887
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 13 September 2013 | Volume 8 | Issue 9 | e73721
52 Xiao YZ Chen Q Hang J Shi YY Wu J et al (2004) Selectiveinduction purification and characterization of a laccase isozyme fromthe basidiomycete Trametes sp AH28-2 Mycologia 96 26-35 doi1023073761984 PubMed 21148825
53 Muntildeoz C Guilleacuten F Martiacutenez AT Martiacutenez MJ (1997) Laccaseisoenzymes of Pleurotus eryngii characterization catalytic propertiesand participation in activation of molecular oxygen and Mn2+ oxidationAppl Environ Microbiol 63 2166-2174 PubMed 9172335
54 Morozova OV Shumakovich GP Gorbacheva MA Shleev SVYaropolov AI (2007) Blue laccases Biochemistry (Mosc) 721136-1150 doi101134S0006297907100112
55 Hildeacuten K Hakala TK Lundell T (2009) Thermotolerant andthermostable laccases Biotechnol Lett 31 1117-1128 doi101007s10529-009-9998-0 PubMed 19360388
56 Verma P Madamwar D (2002) Production of ligninolytic enzymes fordye decolorization by cocultivation of white-rot fungi Pleurotusostreatus and Phanerochaete chrysosporium under solid-statefermentation Appl Biochem Biotechnol 102-103 109-118 doi101385ABAB102-1031-6109 PubMed 12396115
57 Meza JC Lomascolo A Casalot L Sigoillot J-C Auria R (2005)Laccase production by Pycnoporus cinnabarinus grown on sugar-canebagasse Influence of ethanol vapours as inducer Proc Biochem 403365ndash3371 doi101016jprocbio200503004
58 Hossain SM Anantharaman N (2006) Activity enhancement ofligninolytic enzymes of Trametes versicolor with bagasse powder Afr JBiotechnol 5 189-194
59 Revankar MS Desai KM Lele SS (2007) Solid-state fermentation forenhanced production of laccase using indigenously isolatedGanoderma sp Appl Biochem Biotechnol 143 16-26 doi101007s12010-007-0029-0 PubMed 18025593
60 Velazquez-Cedeno MA Mata G Savoie J-M (2002) Waste-reducingcultivation of Pleurotus ostreatus and Pleurotus pulmonarius on coffeepulp changes in the production of some lignocellulolytic enzymesWorld J Microbiol Biotechnol 18 201ndash207 doi101023A1014999616381
61 Mata G Murrieta D Andreu LI (2005) Changes in lignocellulolyticenzyme activities in six Pleurotus spp strains cultivated on coffee pulpin confrontation with Trichoderma spp World J Microbiol Biotechnol 21143ndash150 doi101007s11274-004-3041-3
62 de Souza DF Tychanowicz GK de Souza CG Peralta RM (2006) Co-production of ligninolytic enzymes by Pleurotus pulmonarius on wheatbran solid state cultures J Basic Microbiol 46 126-134 doi101002jobm200510014 PubMed 16598826
63 Prasad KK Mohan SV Bhaskar YV Ramanaiah SV Babu VL et al(2005) Laccase production using Pleurotus ostreatus 1804 immobilizedon PUF cubes in batch and packed bed reactors influence of cultureconditions J Microbiol 43 301-307 PubMed 15995650
64 Songulashvili G Elisashvili V Wasser SP Nevo E Hadar Y (2007)Basidiomycetes laccase and manganese peroxidase activity in
submerged fermentation of food industry wastes Enzyme MicrobTechnol 41 57ndash61 doi101016jenzmictec200611024
65 Sathishkumar P Murugesan K Palvannan T (2010) Production oflaccase from Pleurotus florida using agro-wastes and efficientdecolorization of Reactive blue 198 J Basic Microbiol 50 360-367 doi101002jobm200900407 PubMed 20586068
66 Moldes D Gallego PP Rodriacuteguez Couto S Sanromaacuten A (2003) Grapeseeds the best lignocellulosic waste to produce laccase by solid statecultures of Trametes hirsuta Biotechnol Lett 25 491-495 doi101023A1022660230653 PubMed 12882277
67 Elisashvili V Kachlishvili E Penninckx M (2008) Effect of growthsubstrate method of fermentation and nitrogen source onlignocellulose-degrading enzymes production by white-rotbasidiomycetes J Ind Microbiol Biotechnol 35 1531-1538 doi101007s10295-008-0454-2 PubMed 18716810
68 Osma JF Saravia V Herrera JL Couto SR (2007) Mandarin peelingsthe best carbon source to produce laccase by static cultures ofTrametes pubescens Chemosphere 67 1677-1680 doi101016jchemosphere200611051 PubMed 17234250
69 Kahraman SS Gurdal IH (2002) Effect of synthetic and natural culturemedia on laccase production by white rot fungi Bioresour Technol 82215-217 doi101016S0960-8524(01)00193-6 PubMed 11991068
70 Iandolo D Piscitelli A Sannia G Faraco V (2011) Enzyme Productionby Solid Substrate Fermentation of Pleurotus ostreatus and Trametesversicolor on Tomato Pomace Appl Biochem Biotechnol 163 40-51doi101007s12010-010-9014-0 PubMed 20582639
71 Moldes D Lorenzo M Sanromaacuten MA (2004) Different proportions oflaccase isoenzymes produced by submerged cultures of Trametesversicolor grown on lignocellulosic wastes Biotechnol Lett 26 327-330doi101023BBILE000001545240213bf PubMed 15055770
72 Philippoussis A Diamantopoulou P Papadopoulou K Lakhtar HRoussos S et al (2010) Biomass laccase and endoglucanaseproduction by Lentinula edodes during solid state fermentation of reedgrass bean stalks and wheat straw residues World J MicrobiolBiotechnol 27 285-297
73 Fenice M Giovannozzi Sermanni G Federici F DrsquoAnnibale A (2003)Submerged and solid-state production of laccase and Mn-peroxidaseby Panus tigrinus on olive mill wastewater-based media J Biotechnol100 77-85 doi101016S0168-1656(02)00241-9 PubMed 12413788
74 Rodriguez-Couto S GundIn M Lorenzo M Sanromaacuten MAacute (2002)Screening of supports and inducers for laccase production by Trametesversicolor in semi-solid-state conditions Proc Biochem 38 249-255doi101016S0032-9592(02)00087-0
75 Fackler K Gradinger C Schmutzer M Tavzes C Burgert I et al (2007)Biotechnological wood modification with selective white-rot fungi and itsmolecular mechanisms Food Technol Biotechnol 45 269ndash276
76 Levin L Herrmann C Papinutti VL (2008) Optimization oflignocellulolytic enzyme production by the white-rot fungus Trametestrogii in solid-state fermentation using response surface methodologyBiochem Eng J 39 207-214 doi101016jbej200709004
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 14 September 2013 | Volume 8 | Issue 9 | e73721
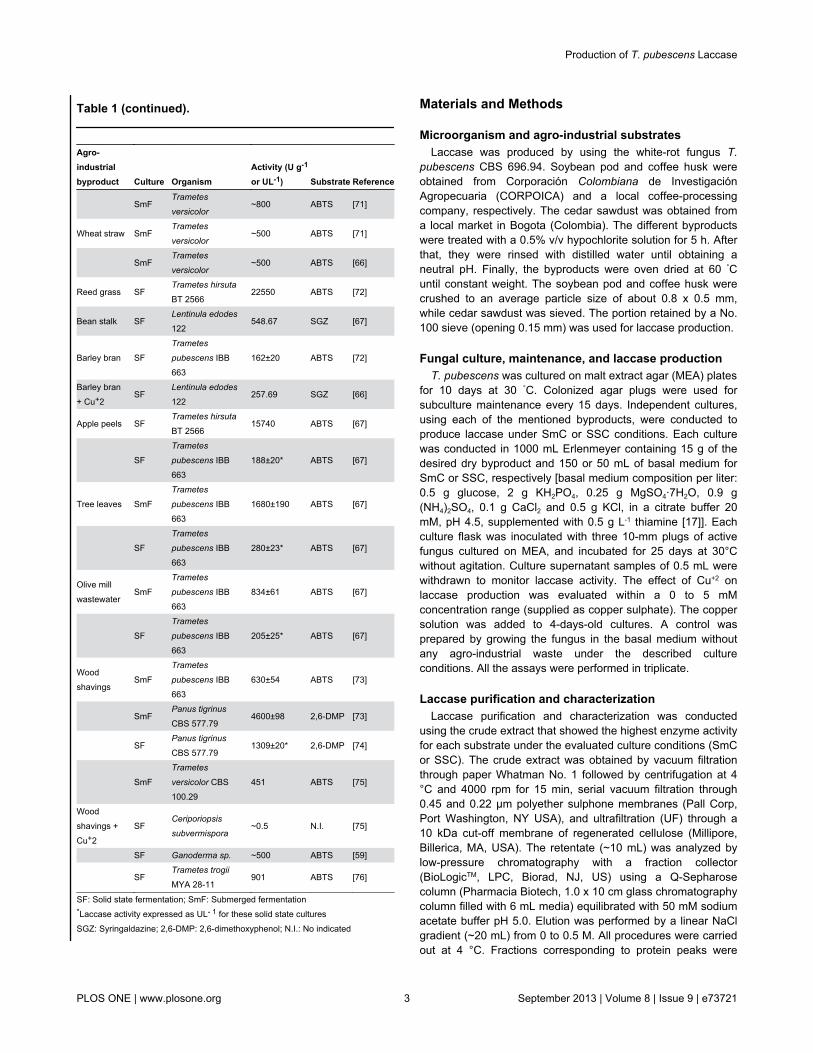
Table 1 (continued)
Agro-industrialbyproduct Culture Organism
Activity (U g-1
or UL-1) Substrate Reference
SmFTrametes
versicolor~800 ABTS [71]
Wheat straw SmFTrametes
versicolor~500 ABTS [71]
SmFTrametes
versicolor~500 ABTS [66]
Reed grass SFTrametes hirsuta
BT 256622550 ABTS [72]
Bean stalk SFLentinula edodes
12254867 SGZ [67]
Barley bran SFTrametes
pubescens IBB663
162plusmn20 ABTS [72]
Barley bran+ Cu+2
SFLentinula edodes
12225769 SGZ [66]
Apple peels SFTrametes hirsuta
BT 256615740 ABTS [67]
SFTrametes
pubescens IBB663
188plusmn20 ABTS [67]
Tree leaves SmFTrametes
pubescens IBB663
1680plusmn190 ABTS [67]
SFTrametes
pubescens IBB663
280plusmn23 ABTS [67]
Olive millwastewater
SmFTrametes
pubescens IBB663
834plusmn61 ABTS [67]
SFTrametes
pubescens IBB663
205plusmn25 ABTS [67]
Woodshavings
SmFTrametes
pubescens IBB663
630plusmn54 ABTS [73]
SmFPanus tigrinus
CBS 577794600plusmn98 26-DMP [73]
SFPanus tigrinus
CBS 577791309plusmn20 26-DMP [74]
SmFTrametes
versicolor CBS10029
451 ABTS [75]
Woodshavings +Cu+2
SFCeriporiopsis
subvermispora~05 NI [75]
SF Ganoderma sp ~500 ABTS [59]
SFTrametes trogii
MYA 28-11901 ABTS [76]
SF Solid state fermentation SmF Submerged fermentationLaccase activity expressed as UL- 1 for these solid state culturesSGZ Syringaldazine 26-DMP 26-dimethoxyphenol NI No indicated
Materials and Methods
Microorganism and agro-industrial substratesLaccase was produced by using the white-rot fungus T
pubescens CBS 69694 Soybean pod and coffee husk wereobtained from Corporacioacuten Colombiana de InvestigacioacutenAgropecuaria (CORPOICA) and a local coffee-processingcompany respectively The cedar sawdust was obtained froma local market in Bogota (Colombia) The different byproductswere treated with a 05 vv hypochlorite solution for 5 h Afterthat they were rinsed with distilled water until obtaining aneutral pH Finally the byproducts were oven dried at 60 degCuntil constant weight The soybean pod and coffee husk werecrushed to an average particle size of about 08 x 05 mmwhile cedar sawdust was sieved The portion retained by a No100 sieve (opening 015 mm) was used for laccase production
Fungal culture maintenance and laccase productionT pubescens was cultured on malt extract agar (MEA) plates
for 10 days at 30 degC Colonized agar plugs were used forsubculture maintenance every 15 days Independent culturesusing each of the mentioned byproducts were conducted toproduce laccase under SmC or SSC conditions Each culturewas conducted in 1000 mL Erlenmeyer containing 15 g of thedesired dry byproduct and 150 or 50 mL of basal medium forSmC or SSC respectively [basal medium composition per liter05 g glucose 2 g KH2PO4 025 g MgSO4middot7H2O 09 g(NH4)2SO4 01 g CaCl2 and 05 g KCl in a citrate buffer 20mM pH 45 supplemented with 05 g L-1 thiamine [17]] Eachculture flask was inoculated with three 10-mm plugs of activefungus cultured on MEA and incubated for 25 days at 30degCwithout agitation Culture supernatant samples of 05 mL werewithdrawn to monitor laccase activity The effect of Cu+2 onlaccase production was evaluated within a 0 to 5 mMconcentration range (supplied as copper sulphate) The coppersolution was added to 4-days-old cultures A control wasprepared by growing the fungus in the basal medium withoutany agro-industrial waste under the described cultureconditions All the assays were performed in triplicate
Laccase purification and characterizationLaccase purification and characterization was conducted
using the crude extract that showed the highest enzyme activityfor each substrate under the evaluated culture conditions (SmCor SSC) The crude extract was obtained by vacuum filtrationthrough paper Whatman No 1 followed by centrifugation at 4degC and 4000 rpm for 15 min serial vacuum filtration through045 and 022 microm polyether sulphone membranes (Pall CorpPort Washington NY USA) and ultrafiltration (UF) through a10 kDa cut-off membrane of regenerated cellulose (MilliporeBillerica MA USA) The retentate (~10 mL) was analyzed bylow-pressure chromatography with a fraction collector(BioLogicTM LPC Biorad NJ US) using a Q-Sepharosecolumn (Pharmacia Biotech 10 x 10 cm glass chromatographycolumn filled with 6 mL media) equilibrated with 50 mM sodiumacetate buffer pH 50 Elution was performed by a linear NaClgradient (~20 mL) from 0 to 05 M All procedures were carriedout at 4 degC Fractions corresponding to protein peaks were
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 3 September 2013 | Volume 8 | Issue 9 | e73721
collected and analyzed by sodium dodecyl sulfate-polyacrylamide electrophoresis gel (SDS-PAGE) under non-reducing conditions and without heat denaturation Staining ofthe electrophoresis gel was done with 5 mM 22rsquo-azino-bis(3-ethylbenzothiazoline-6) sulphonic acid (ABTS) in succinic acid(25 mM pH 45) to detect laccase activity Fractions thatrevealed the same protein profile and laccase activity werecombined Samples of the combine fractions were analyzed bySDS-PAGE under mentioned conditions As a negative controlwere used a sample of the ultrafiltrated fraction which did notreveal laccase activity along the previous purification stepBands that revealed laccase activity were identified as Lac1and Lac2 These bands were excised from the electrophoresisgel and analyzed through nanoLC-MSMS (Applied BiomicsHayward CA USA) Peptides and protein sequences werealigned by using Clustal Omega [18] at EBI web server andBlast at NCBI web server Alignments figures were generatedusing CLC Sequence Viewer (CLC bio Denmark)
Enzyme activity was determined spectroscopically by usingeither 22rsquo-azino-bis(3-ethylbenzothiazoline-6) sulphonic acid(ABTS ε420 = 36mM-1 cm-1 Sigma-Aldrich) or 4-hydroxy-35-dimethoxybenzaldehyde (Syringaldazine ε530 = 65mM-1 cm-1Sigma-Aldrich) [19ndash21] ABTS activity assay was carried out aspreviously described [2122] Briefly 850 microL of 5 mM ABTS insuccinic acid (25 mM pH 45) were mixed with 150 microL of therespective dilution of culture extract and the absorbance was at420 nm Syringaldazine activity assay was carried out followingthe substrate manufacturerrsquos instructions (Sigma-Aldrich) Oneunit (U) was defined as the amount of enzyme required tooxidize 1 micromol of substrate (ABTS or syringaldazine) perminute Specific activity was expressed as U mgminus1 of protein asdetermined by Lowry assay [23]
Laccase production during the fungus culture was monitoredusing syringaldazine which has demonstrated to be a specificsubstrate for detecting extracellular laccase in the absence ofhydrogen peroxide [24] Absence of hydrogen peroxide wasconfirmed by incubation with catalase Crude extracts andpurified fractions were analyzed by SDS-PAGE withoutreduction by β-mercaptoethanol and heat denaturationProteins were visualized by silver staining while laccases wereidentified by placing the SDS-PAGE gel in an ABTS solution for5 min Temperature and pH effect on laccase activity wasdetermined for the UF retentate and the purified fractions byusing ABTS The temperature effect on laccase activity wasevaluated at 30 40 50 55 60 and 70 degC at pH 50 and the pHeffect on laccase activity was evaluated at 2 25 3 4 5 6 65and 7 at 55 degC Laccase stability was evaluated by using ABTSat optimal pH and temperature for 48 h
Cyclic voltammetry analysisCyclic voltammetry (CV) analysis was performed on a
computer-controlled potentiostat (Autolab PotentiostatGalvanostat PGSTAT 128N Metrohm USA) Data wereacquired by using the software Nova version 16 Aconventional three-electrode system was used to carry out thecyclic voltammograms The used electrodes were graphite-working electrode with a surface area of 023 cm2 AgAgCl KClsaturated reference electrode (Metrohm USA) and platinum
wire counter electrode Before running each experiment thesurface of the graphite electrode was polished with aluminaand thoroughly rinsed with distilled water CV analysis wasdone to the crude extract and purified laccase fractions thatpresented the highest activity for each substrate For the test20 microL of sample were added on the polished surface of theworking electrode and allowed to dry for 60 min at roomtemperature Each experiment was performed in triplicate usingeither a solution 11 of anhydride ethanol buffer acetate (100mM pH 50) and syringaldazine to a final concentration of 02mM or a solution of 100 mM acetate buffer (pH 30) and 05mM ABTS Cyclic voltammograms were carried out in 30 mL ofworking solution with continuous agitation Voltage wasscanned at 100 mV s-1 between -02 and 1 V vs Ag|AgCl|KClsat
for syringaldazine assays while for ABTS assays voltage wasscanned at 250 mV s-1 between -02 and 12 V vs Ag|AgCl|KClsat Assays were conducted in a Faraday cage at roomtemperature (20-22degC) Temperature and pH in the reactionsolution was monitored with the Unitrode with Pt 1000(Metrohm USA)
Statistical analysisAll experiments were conducted by triplicate Data are
presented as the mean plusmn standard deviation (SD) Statisticalcomparisons were conducted by using either t-test or Duncanrsquostest for one-factor design by using StatGraphics Centurionreg v16111 (2011) Statistical differences were established for p lt005
Results
SmC and SSC production of laccase by T pubescensCoffee husk soybean pod husk and cedar sawdust were
used as substrate for the production of laccase by Tpubescens under SmC or SSC conditions while syringaldazinewas used to monitor laccase production during the fungusculture under the studied conditions Hydrogen peroxide wasnot detected in any of the evaluated samples which discardthe interference of peroxidases in the syringaldazine assayLaccase activity and glucose concentration time course for Tpubescens grown on coffee husk under SmC and SSC atdifferent copper concentrations are presented in Figure 1Although a similar time course was observed for laccaseactivity and glucose concentration of soybean pod husk andcedar sawdust cultures differences on the day of maximumlacasse activity as well as in the maximum activity value wereobserved among the agro-industrial byproducts (Table 2) Ingeneral it was noticed that laccase activity was detectablearound the seventh and fourth day of culture for SmC andSSC respectively After the eighth culture day glucose wasdepleted from the culture medium and laccase activityincreased sharply
The substrate source seemed to be the most important factorfor laccase production (Table 2) T pubescens culture oncoffee husk allowed the maximum laccase activities for bothSmC (17723plusmn372 U L-1) and SSC (20400plusmn596 U L-1) atcopper concentrations of 5 and 0 microM respectively Laccaseactivity in SSC and SmC using coffee husk as substrate was
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 4 September 2013 | Volume 8 | Issue 9 | e73721
between 147- and 663-fold higher than that observed forcultures with soybean pod husk and cedar sawdust Laccaseactivity obtained in the control culture without agro-industrialwaste presented a maximum laccase activity of 222 plusmn 088 UL-1 after 14 days of culture while the glucose was depleted onthe seventh day of culture
Copper supplementation effect on laccase activity wasevaluated for both SmC and SSC Although in a non-linearmode the laccase activity was increased by the presence ofcopper with all substrates under SmC or SSC except withcoffee husks under SSC (Table 2) In addition the requiredcopper concentration to obtain the highest laccase activityvaried depending on the used agro-industrial waste Forexample laccase production was favored in cultures withsoybean pod husk at copper concentrations of 05 mM and 20mM for SmC and SSC respectively while for cultures with
Figure 1 Laccase activity profiles during T pubescensCBS 69694 culture under SmC (a) and SSC (b) on coffeehusk Culture media was enriched with different copperconcentrations 0 (Δ) 05 () 2 () and 5 (loz) mM and laccaseactivity was determined by using syringaldazine as substrateFilled symbols refer to the glucose concentration at therespective copper concentration All the assays wereperformed in triplicatedoi 101371journalpone0073721g001
coffee husk the highest laccase activity levels for SmC andSSC was observed at copper concentrations of 5 mM and 0mM respectively
Despite that culture mode ie SmC or SSC presented aneffect on laccase production this is substrate sensitive In theassays conducted without copper it was observed that laccaseactivity was 28- and 33-fold higher in SSC than in SmC for thesoybean pod husk and coffee husk respectively Howeverwhen cedar sawdust was used no significant difference inlaccase activity between the two culture types was obtained(Table 2) Due to the low volume of liquid media and highlaccase activity in SSC when compared to SmC the SSCpresented a higher volumetric yield For all agro-industrialwaste the SSC presented a shorter time than SmC to reachthe highest laccase activity which suggest a higher productivityunder SSC conditions
These results show that coffee husk under SmC or SSCconditions could be considered as a potential alternative forlaccase production by T pubescens However soybean podhusk and cedar sawdust could be also considered as importantalternatives for laccase production especially in comparisonwith synthetic culture media [10] Finally the optimal copperconcentration to obtain the highest laccase activity depends onthe agro-industrial byproduct used for the fungus culture
Laccase purification and characterizationLaccase purification was conducted using the crude extracts
that presented the highest activities For the SmC crude
Table 2 Effect of substrate copper concentration andculture mode on laccase activity (UL-1)
Substrate SmC SSCSoybean Pod Husk + 0 mM Cu+2 446 plusmn 138 (23) 1255 plusmn 052 (13)
Soybean Pod Husk + 05 mM Cu+2
8493 plusmn 846 (23)dagger 2028 plusmn 387 (21)
Soybean Pod Husk + 2 mM Cu+2 5761 plusmn 370 (19) 13785 plusmn 071 (18)dagger
Soybean Pod Husk + 5 mM Cu+2 4492 plusmn 487 (23) 2900 plusmn 407 (18)Coffee Husk + 0 mM Cu+2 6184 plusmn 176 (23) 20400 plusmn 596 (19)dagger
Coffee Husk + 05 mM Cu+2 13763 plusmn 568 (23) 11083 plusmn 940 (20)ab
Coffee Husk + 2 mM Cu+2 12304 plusmn 373 (23)b 10278 plusmn 814 (15)b
Coffee Husk + 5 mM Cu+2 17723 plusmn 372 (23)dagger 8266 plusmn 882 (15)Cedar Sawdust + 0 mM Cu+2 1355 plusmn 095 (17) 1082 plusmn 053 (15)
Cedar Sawdust + 05 mM Cu+2 2272 plusmn 084 (17) 1902 plusmn 002 (13)Cedar Sawdust + 2 mM Cu+2 2897 plusmn 127 (12)a 2638 plusmn 068 (8)Cedar Sawdust + 5 mM Cu+2 2936 plusmn 153 (13) abdagger 3073 plusmn 152 (12)bdagger
Data in parenthesis refers to the day where was observed the maximum laccaseactivity No Statistical Difference Statistical Difference (Evaluation of substrate effecton laccase activity for both culture modes submerged (SmC) and semisolid (SSC)no copper induction considered)Equal letters inside one agro-industrial waste group refers to No StatisticalDifference no letters refers to Statistical Difference inside the group (Evaluation ofcopper effect on laccase activity for both SmC and SSC)dagger Maximum laccase activity for the used agro-industrial wastes either in SmC orSSC culture
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 5 September 2013 | Volume 8 | Issue 9 | e73721
extracts obtained with soybean pod husk + Cu+2 05 mM coffeehusk + Cu+2 5 mM and cedar sawdust + Cu+2 5 mM wereselected while for SSC crude extracts obtained with soybeanpod husk + Cu+2 2 mM coffee husk + Cu+2 0 mM and cedarsawdust + Cu+2 5 mM were selected The electrophoreticanalysis of permeates from 045 and 022 microm filtrations and theUF retentate showed the presence of two proteins with laccaseactivity while no activity was observed within the ultrafiltratedsample (Figure 2a) These proteins presented a molecularmass of about 120 kDa and 60 kDa (Figure 2a) Thepurification of the UF retentate by ionic exchangechromatography confirmed the presence of two differentfractions with laccase activity (Figure 2b) The SDS-PAGEanalysis of these fractions under non-reducing conditionsshowed that one protein with laccase activity was presented oneach fraction (Figure 2b) and were named Lac1 (120 kDa) andLac2 (60 kDa) Considering that the conditions used tomeasure the enzyme activity were those suggested for laccaseactivity and that H2O2 or manganese were not added duringthe reaction for measuring the enzyme activity these resultssuggest that both proteins are laccases but not peroxidases
NanoLC-MSMS analysis of purified fractions showed thatthe identified peptides for Lac1 (SAGSTTYNYNDPIFR andRDTVSTGNSGDNVTIR) and Lac2 (SAGSTVYNYDNPIFR andANPNFGNVGFTGGINSAI) had identity scores between 80 to100 with laccases (over 100 sequences) available at the non-redundant GenBank dataset Proteins containing thesepeptides different that laccases were not identified Similarresults were observed when peptides were evaluated atUniprot Galhaup et al [17] described two laccase activefractions from T pubescens MB 89 (CBS 69694) Lap1(UniProtKBTrEMBL Q8TG93) and Lap2 (UniProtKBTrEMBLQ8TG94) which share a 70 identity (Figure 3a) Sequencealignment of Lac1 and Lac2 peptides with Lap1 and Lap2sequences respectively showed that the peptides arepresented within T pubescens MB 89 laccase sequencesthough some differences were observed for the Lac1 peptide(Figure 3b) This result suggests that the purified fractions arelaccase isoenzymes
Protein concentration and volumetric and specific laccaseactivities at different purification stages are presented in Table3 Lac1 presented lower laccase activity than Lac2 in a ratiothat ranges from 080- to 38-fold depending on the substratesource On the other hand laccase activity of the UF retentatefrom soybean pod husk culture was from 45- to 64-fold higherthan that of the purified fractions while for cedar sawdust andcoffee husk the laccase activity was from 46- to 120-fold andfrom 33- to 163-fold higher than the purified fractionsrespectively (Table 3) The purified Lac2 fractions from coffeehusk and cedar sawdust cultures showed the highest specificactivity levels (Table 3) The volumetric activities of UFretentates were higher than that obtained with purified fractions(Table 4)
Data in parenthesis shows the temperature or pH at whichrelative laccase activity reached 100
Temperature and pH effect on laccase activity wasdetermined for the UF retentate and Lac1 and Lac2 fractionsobtained along the purification of the enzymatic crude extracts
that resulted after the SSC of T pubescens on the selectedagro-industrial wastes Laccase activity for the UF retentateunder different temperature and pH conditions is presented inFigure 4 and Table 4 The maximum relative activity of theevaluated UF retentates was observed at 50-55 degC and at apH range from 20 to 40 However UF retentate from cedar
Figure 2 Purification of lacasses produced by submergedculture of T pubescens using coffee husk assubstrate All samples were analyzed by SDS-PAGE undernon-reducing conditions and stained with ABTS No heatdenaturation of the samples was conducted (a) The crudeextract (1) was centrifuged and filtered through a Whatman No1 filter (2) 045 microM membrane (3) 022 microm membrane (4) andultrafiltrated through a 10 kDa cut-off membrane permeate (5)and retentate (6) (b) UF retentate was purified by anionicexchange chromatography Lac1 represent the unboundfraction while Lac2 represent the eluted fraction The arrowshows the point when protein elution was started (c) Purifiedfractions (1) crude extract (2) Lac1 and (3) Lac2 MWMolecular Weight Similar results were observed for crudeextracts produced by using soybean pod husk and cedarsawdustdoi 101371journalpone0073721g002
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 6 September 2013 | Volume 8 | Issue 9 | e73721
sawdust culture behaved different to the UF retentate fromcoffee husk and soybean pod husk cultures In this sense the
UF retentate from cedar sawdust culture presented the lowestenzyme activity at temperatures le 40 degC but at 70 degC
Figure 3 Aligment of reported lacasses sequences and Lac1 and Lac2 peptides (a) Alignment of Lap1 (UniProtKBSwiss-Prot Q8TG93) and Lap2 (UniProtKBSwiss-Prot Q8TG94) from T pubescens CBS 69694 (b) Alignment of Lac1 and Lac2 peptideswith Lap1 and Lap2 respectively and tryptic peptides described by Shleev et al 2007 Alignments were carried out using ClustalOmega and figures were using CLC Sequence Viewer Different residues are colored in reddoi 101371journalpone0073721g003
Table 3 Laccase activity during the purification of crude extracts resulted from T pubescens culture on agro-industrialwastes under submerged (SmC) and semisolid (SSC) cultures
Soybean pod husk Coffee husk Cedar sawdust
Protein(mg mL-1)
VolumetricActivity (UL-1)
SpecificActivity (Umg-1
protein)Recovery()
Protein(mg mL-1)
VolumetricActivity (UL-1)
SpecificActivity (Umg-1
protein)Recovery()
Protein(mg mL-1)
VolumetricActivity (UL-1)
SpecificActivity (Umg-1
protein)Recover()
SmCCrudeExtract
0869 52888 0608 100 0414 152708 3688 100 0431 10735 0249 100
045 micromFiltration
0736 46549 0632 88 0334 117892 3529 77 0444 12053 0271 112
UF retentate 1010 78731 0779 74 0378 195263 5165 64 0510 25905 0507 120
Lac1 0494 5471 0110 1 0232 2399 0103 02 0222 838 0037 08
Lac2 0310 20661 0666 4 0116 107240 9252 7 0040 15234 3837 14 SSCCrudeExtract
1146 164966 1439 100 0596 154907 2601 100 1065 23203 0217 100
045 micromFiltration
0776 128392 1654 77 0474 115736 2439 75 0806 25824 0320 111
UF retentate 0973 335055 3443 101 0428 161750 3779 52 1134 51101 0450 110
Lac1 0097 8854 0912 2 0104 1514 0145 03 0042 1315 0313 2
Lac2 0168 48988 2915 10 0131 82298 6282 18 0072 16162 2244 23
740 ABTS was used to determine laccase activity
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 7 September 2013 | Volume 8 | Issue 9 | e73721
presented the highest enzyme activity ~90 of its initial valuein comparison to the other two UF retentates (Figure 4)
The temperature and pH effect on laccase activity for Lac1and Lac2 is presented in Figure 5 The maximum relativeactivity for both fractions was presented at a pH from 20 to 40regardless of the agro-industrial waste as observed for the UFretentate However at pH gt 40 Lac1 showed a sharperdecrease in its activity than that observed for Lac2Nevertheless Lac2 purified from cedar sawdust culturepresented a different performance This fraction showed themaximum activity at pH 50 while a sharp drop was observedat pHs above or below this value
Laccase activity profiles showed that Lac1 and Lac2presented the maximum enzyme activity at 55 and 60 degCrespectively (Table 4) It was observed that Lac1 from coffeehusk culture presented a lower decrease in enzyme activity attemperatures le 50 degC or ge 60 degC than Lac2 On the contraryLac2 from soybean husk or cedar sawdust cultures presenteda higher relative laccase activity than Lac1 at temperatures le55 degC The purified fractions from soybean pod husk and cedarsawdust cultures presented the lowest relative activity at 30 degCand 65 degC
Table 4 Optimum temperatures and pHs for maximumlaccase activities in UF retentate Lac1 and Lac2 obtainedfrom semisolid cultured T pubescens on agro-industrialwastes
EnzymaticFraction Temperature pH
VolumetricActivity (UL-1)
SpecificActivity (Umg-1 protein)
VolumetricActivity (UL-1)
SpecificActivity (Umg-1
protein)SoybeanPod Husk +2 mM Cu+2
148000plusmn1178(55)
1521plusmn0012182037plusmn3025(2-3)
1871plusmn0031
Lac122925plusmn917(60)
2363plusmn009536135plusmn831(23)
3725plusmn0085
Lac232638plusmn1558(55)
1942plusmn009228713plusmn274(23)
1709plusmn0016
CoffeeHusk + 0mM Cu+2
113740plusmn8289(50-55)
3008plusmn0219290666plusmn1039(2-3)
7689plusmn0275
Lac1 889plusmn039 (55) 0038plusmn0002 1779plusmn028 (23) 0077plusmn0001
Lac234194plusmn2789(60)
2947plusmn024033150plusmn526 (2ndash4)
2857plusmn0045
CedarSawdust +5 mM Cu+2
257666plusmn1201(55)
5052plusmn0024344634plusmn1043(2-3)
6757plusmn0204
Lac121370plusmn472(55)
0962plusmn002129117plusmn943(23)
1311plusmn0042
Lac255166plusmn1656(60)
1379plusmn0414 55166plusmn1656 (5) 1379plusmn0414
ABTS was used to determine laccase activity
Laccase stabilityFor selected enzymatic extracts and purified fractions (Table
4) a stability test for 48 h at their optimal pH and temperaturewas conducted Time course of the relative laccase activity forcrude extract and purified fractions from coffee husk and cedarsawdust cultures are presented in Figure 5 Similar resultswere observed for crude extract and purified fractions fromsoybean pod husk culture (data no shown)
It was observed that the activity loss depends on thesubstrate source used to grow T pubescens An activity loss of55 was observed for crude extracts and purified fractionsfrom coffee husk and soybean pod husk cultures while anactivity loss of 70 was observed for the crude extract andpurified fractions from cedar sawdust culture NeverthelessLac2 showed the lowest activity reduction with values of 10 40and 55 for coffee husk soybean pod husk and cedarsawdust respectively while Lac1 presented an activity loss of34 50 and 60 for coffee husk soybean pod husk and cedarsawdust respectively Despite the differences in laccaseactivity among agro-industrial wastes and between crudeextracts and purified fractions all the profiles showed a sharp
Figure 4 Effect of temperature (a) and pH (b) on laccaseactivity for the UF retentate obtained by semisolid cultureof T pubescens on coffee husk () soybean pod husk (loz)and cedar sawdust (Δ) Laccase stability was evaluated byusing ABTS All the assays were performed in triplicatedoi 101371journalpone0073721g004
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 8 September 2013 | Volume 8 | Issue 9 | e73721
decrease during the first 8 h while a reduced activity loss wasobserved during the last 24 h of the assay (Figure 6)
Laccase electrochemical characterizationFor selected crude extracts and purified fractions a cyclic
voltammetry characterization was conducted For eachmediator syringaldazine or ABTS similar cyclic
Figure 5 Effect of temperature () and pH () on enzymeactivity for Lac1 (solid line) and Lac2 (dashed line)obtained after the purification of the enzymatic crudeextract produced by semisolid culture of T pubescens oncoffee husk (a) soybean pod husk (b) and cedar sawdust(c) Laccase stability was evaluated by using ABTS All theassays were performed in triplicatedoi 101371journalpone0073721g005
voltammograms were obtained for the different crude extractsand purified fractions Voltammograms of the redox reaction forsyringaldazine (Figure 7a) and ABTS (Figure 6b) arepresented Two anionic and cationic peaks were observedwhen syringaldazine was used as mediator The first pair ofpeaks showed a reversible performance with a mean Ia Icratio of 109plusmn006 106plusmn005 and 098plusmn002 for the crudeextracts Lac1 and Lac2 respectively The second pair ofpeaks showed quasi-reversible performance and the cathodicpeak was presented at a potential of +062 V (vs Ag|AgCl)while the anodic peak was presented at a potential of +060 V(vs Ag|AgCl) No significant difference on the reductionpotential (Eo) for syringaldazine was observed for the crudeextracts or the purified fractions obtaining an Eo of0314plusmn0008 V (vs Ag|AgCl) However the peak potentialseparation (ΔEp) showed a different profile depending on thesubstrate used for the culture While not significant difference(p gt 005) on ΔEp value was observed between crude extract(0045plusmn0005 V vs Ag|AgCl) Lac1 (0046plusmn0005 V vs Ag|AgCl)and Lac2 (0028plusmn0001 V vs Ag|AgCl) from cultures with cedarsawdust and soybean pod husk the ΔEp was considerablyhigher for the crude extract (0083plusmn0004 V vs Ag|AgCl) Lac1(0068plusmn0003 V vs Ag|AgCl) and Lac2 (0051plusmn0005 V vs Ag|AgCl) from the coffee husk culture
For reactions conducted with ABTS and the different crudeextracts and purified fractions the voltammograms exhibitedone pair of cathodic and anodic peaks (Figure 7b) whichcorresponds to the redox of ABTSABTS+ (IaIc) with an Eo of0487plusmn0006 V (vs Ag|AgCl) This process was reversiblepresenting a mean Ia Ic ratio of 099plusmn002 In addition asecond cathodic peak was presented at +083 V (vs Ag|AgCl)when ABTS redox reaction was conducted using the crudeextract and the purified fractions This performance agrees withprevious reports in which the laccase activity cannot beobserved at potential values of the second reversible oxidation
Figure 6 Laccase stability profiles at optimal pH andtemperature conditions for crude extracts (solid line) Lac1(dashed line) and Lac2 (spotted line) obtained fromsemisolid culture of T pubescens on coffee husk (loz) andcedar sawdust (bull) Laccase stability was evaluated by usingABTS All the assays were performed in triplicatedoi 101371journalpone0073721g006
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 9 September 2013 | Volume 8 | Issue 9 | e73721
of ABTS ABTS+ABTS2+ due to this catalytic process cannotbe detected by cyclic voltammetry [2526] Neverthelessvoltammograms for laccase crude extract from a soybean podhusk culture presented the second anodic peak which inaddition to the second cathodic peak corresponds to the redoxof ABTS+ABTS2+ (IIaIIc) with an Eo of 0942plusmn0004 V (vs Ag|AgCl) Contrary to the reactions conducted usingsyringaldazine as substrate the ΔEp did not present asignificant difference for the crude extracts and purifiedfractions 0248plusmn0016 V (vs Ag|AgCl)
Discussion
In this study we evaluated the effect of the culture mode ieSmC and SSC on laccase production by T pubescens CBS69694 using three different agro-industrial wastes (coffeehusk soybean pod husk and cedar sawdust) Furthermore theeffect of Cu+2 supplementation on laccase production was alsoevaluated and the crude extracts and the purified fractionswere partially characterized
Figure 7 Cyclic voltammograms of (a) syringaldazine atpH 50 using the crude extract and purified laccasefractions from semisolid culture of T pubescens on coffeehusk or (b) ABTS at pH 30 using the crude extract andpurified laccase fractions from semisolid culture of Tpubescens on soybean pod husk Nomenclature onlymediator (dotted line) crude extract (solid line) Lac1 fraction(dashed line) and Lac2 fraction (dashed and dotted line)doi 101371journalpone0073721g007
Substrate source for laccase production showed to be one ofthe most important factors for laccase production (Table 2)Cultures with coffee husk as substrate presented the highestlaccase production levels The observed differences in laccaseactivity for each substrate could be owed to their chemicalcomposition Coffea arabica is the coffee variety raised inColombia and its husk has a reported composition of 2308cellulose (2076 glucose and 183 cellobiose) 2385hemicellulose (1356 xylose 523 arabinose 256 aceticacid and 195 glucuronic acid) 2828 total lignin and071 ashes [27] In addition coffee husk contents 13caffeine 45 tannins and 124 pectins [28] On the otherhand Glycine max is the soybean species cultured inColombia and its pod husk composition varies from 504 to62 crude protein 608 to 143 hemicellulose 3715 to4208 acid detergent fiber 4436 to 6015 neutral detergentfiber 850 to 893 acid detergent lignin and 2074 to 2106uronic acid [29ndash31] Finally for cedar sawdust the majorcomponents are glucose (426-444) xylose (46-195)cellulose (383-410) klason lignin (227-278) and acidlignin (40-43) [32ndash34] Although lignin is one of the mostimportant compounds associated with laccase production bywhite-rot fungi we did not observe a clear correlation betweenlignin composition percentage and laccase activity In thatsense the high laccase activity observed for the cultures thatuse coffee husk could be associated with its content of caffeineand tannins which have been identified as potent inducers oflaccase gene expression [173536] In fact caffeine has beenused as inducer for laccase production by Rhizoctonia solaniobtaining an increase of 41-fold in laccase activity [37] Similarresults were also reported for the production of Coloriopsisgallica laccase [38]
Laccase gene expression is mainly up regulated by Cu2+although Mn2+ Fe3+ heavy metals 26-dimethoxy-14-benzoquinone H2O2 amphotericin B syringic acid tannic acidTween 80 soybean oil aromatic compounds andmicroclimatic changes (ie lower temperature and osmoticpressure) are also recognized as potent inducers [173536] Ingeneral Cu2+ supplementation increased the laccase activity inboth SmC and SSC However Cu2+concentration required toobtain the highest laccase activity varied depending on agro-industrial waste These differences can be attributed to thecopper presented in the agro-industrial waste and the copperavailability in the medium Copper can be found in amountsthat range from 156 to 180 mg kg-1 of coffee husk [39] andfrom 26 to 67 mg kg-1 of soybean pod husk (or soybean hull)[3940] Despite that no reports indicate the copper content inuntreated cedar wood an average of 10 mg kg-1 of wood hasbeen reported for wood used as biomass in six different Europecountries [41] This allows to consider the cedar sawdust as thesubstrate with the lowest content of copper On the other handcopper adsorption by sawdust and coffee husk has beenreported obtaining a Langmuir based maximum adsorptioncapacity (Qmax mg g-1 adsorbent) of 845 and 75 respectively[4243] Although no adsorption studies has been reportedusing untreated soybean pod husk Šćiban et al [44] reporteda Qmax for copper adsorption of 0085 mmol g-1 adsorbent (~ 54mg g-1) using treated soybean straws Based on this
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 10 September 2013 | Volume 8 | Issue 9 | e73721
information it can be expected that a higher amount ofexogenous copper would be required for laccase induction oncultures with cedar sawdust than with soybean pod husk andcoffee husk due to its lower content of copper In addition dueto the high copper-adsorption capacity of sawdust the amountof available copper in the culture would be reduced allowingthe laccase production at higher copper concentrations withoutreaching levels that might be toxic for the fungus This fact canbe observed in the SSC (Table 4) However for the SmC thisbehavior is not maintained for the cultures with soybean podhusk and coffee husk (Table 4) where the maximum laccaseactivity was obtained with a copper concentration of 05 and 5mM respectively This could be owed to a lower contactbetween the fungus and the substrate in the SmC whencompared to the SSC which is close related to the transport ofnutrients presented in the substrates
Production of extracellular laccases has been reported in alarge number of species of white-rot fungi strains grown onnatural substrates (Table 1) Although a laccase activity profileis not clearly observed based on the agro-industrial wasteculture mode and fungus strain most cases in which SmC andSSC were conducted using the same substrate and fungusstrain showed a higher laccase activity under SmC conditionsIn this study was observed a substrate dependentperformance where the most favorable condition to obtain thehighest laccase activity was the SSC The laccase activitiesobtained in the present study are comparable and in somecases higher than those reported for Pleurotus ostreatusPhanerochaete chrysosporium Pycnoporus cinnabarinus Tversicolor and Ganoderma sp cultured under SC and SmCconditions and using different agro-industrial wastes (Table 1)Semi-solid cultures using different agro-industrial wastes havebeen reported using T versicolor Phlebia radiata Tpubescens P ostreatus and C unicolor Rodriguez et al [16]screened different supports for laccase production by Tversicolor under SSC They obtained the highest laccaseactivity (1200 U L-1) when in the culture medium was presentedbarley bran In addition they were able to obtain a laccaseactivity of 1700 U L-1 when the culture was supplemented withxylidine as an inducer These results were obtained after 17 or18 days of culture and ABTS was used to determine theenzyme activity Maumlkelauml et al [4546] have studied theexpression of the genes involved in laccase production Pr-lac1and Pr-lac2 presented in P radiata and evaluate laccaseproduction in SSC using milled alder wood (Alnus incana) assubstrate They reported a maximum laccase activity of 3 microkatL-1 (~180 U L-1) at the 14th day of culture and usingsyringaldazine for its detection Osma et al [15] have studiedthe morphological changes and laccase production of strains Tpubescens P ostreatus C unicolor and T versicolor grownon wheat bran flakes using SSC They obtained a maximumlaccase production between 1397 and 2778 U L-1 after 10 and13 days of culture This enzyme activity is strain dependentand ABTS was used for its quantification Our results show agood correlation with reported data The highest laccaseactivity was obtained when T pubescens was grown on coffeehusk under SSC without copper (204 U L-1 evaluated withsyringaldazine or 1549 U L-1 evaluated with ABTS) which is in
some cases higher than reported ones This shows that underthe studied conditions the production of the laccase crudeextract is favored in SSC using coffee husk without the additionof copper which could favors the cost of the productionprocess
It has been reported for white rot-fungi the production ofmultiple laccase isoenzymes which are encoded by differentgenes and show different expression profiles stabilitiessubstrate affinities and molecular masses (ranging from 35 to140 kDa) [3647ndash49] Two isoforms have been identified in Tpubescens strains two monomeric (Lac1 and Lac2) enzymesof 67 kDa but with different pI (53 and 51) [50] and twolaccase-activity fractions (Lap1 and Lap2) with different pI (gt30 and 26) from which Lap2 is a monomeric enzyme of 65kDa [17] In this study we identify two laccase isoenzymesfrom T pubescens CBS 69694 that have different molecularmasses (120 and 60 kDa) and pI The purifications resultssuggest a more acidic pI for Lac1 (le 4) than Lac2 (ge 6) whichdiffers from a previous report for this fungus in which twofractions with laccase activity were obtained Lap1 and Lap2with pI gt 3 and of 26 respectively [17] Since in both casessimilar purification protocols were used and Lac1 and Lac2were recovered from the unbound and bound fractionsrespectively as Lap1 and Lap2 [17] this difference could bedue to the culture conditions used in each case (ie thepresence of agro-industrial wastes) Nevertheless the nanoLC-MSMS analysis showed that peptides of Lac1 and Lac2 werepresented in Lap1 (UniProtKBSwiss-Prot Q8TG93) and Lap2(UniProtKBSwiss-Prot Q8TG94) which have a predicted pI of43 and 47 respectively Lac1 and Lac2 peptides werepresented in Laccase-2 (UniProtKBSwiss-Prot Q99046) andLaccase-1 (UniProtKBSwiss-Prot Q99044) from T villosarespectively with 80 to 100 identity These T villosa laccaseisoenzymes have a predicted pI of 47 (laccase-1) and 60(laccase-2) and a 77 identity among them In addition Tvillosa laccase-2 is reported as a homodimer protein whilelaccase-1 is reported as monomeric protein Alignment of Lac1-Lap2 and Lac2-Lap1 from T pubescens CBS 69694 and Tvillosa showed a 96 and 70 identity respectively Finallysimilar peptides were identified for Lac1 and Lac2 from Tpubescens (Schumach) Pilaacutet (Syn Coriolus pubescens(Schum ex Fr) Queacutel) BCB 923-2 (Figure 3b) [51] Theseresults suggest that the purified fractions correspond to laccaseisoenzymes that differ from previously reported ones for this Tpubescens strain
As mentioned above laccases isoenzymes present differentstabilities and substrate affinities This behavior was alsoobserved for the laccase isoenzymes purified from Tpubescens with Lac1 showing lower laccase activity thanLac2 with ABTS as substrate However the difference inactivity between these isoenzymes depended on the agro-industrial waste used for the T pubescens growth suggestinga probable effect of the substrate composition on the laccasephysicochemical properties In addition it was observed thatthe total laccase activity presented in the UF retentates ishigher than the laccase activity of the purified fractionsregardless of the agro-industrial byproduct used for the Tpubescens growth This suggests a synergistic effect of Lac1
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 11 September 2013 | Volume 8 | Issue 9 | e73721
and Lac2 as have been reported for laccases obtained fromTrametes and Pleurotus strains [175253]
The maximum relative activity for the UF retentate waspresented at a pH from 20 to 40 regardless of the agro-industrial byproduct with a monotonical decrease at pHs abovethis range The relation between pH and type of electron donormolecule explains this behavior on laccase activity It has beenreported that when the substrate is an organic hydrogen donorie phenols the optimal pH ranges from 35 to 60 while thelaccase activity monotonically decreases from pH 25 to 70 ifthe substrate is an electron donor like ABTS [54] Despite thatat different pHs most of the purified laccase fractionspresented a similar performance as the UF retentates the Lac2fraction obtained from cedar sawdust differed from the previousobservation For this fraction the maximum activity wasobserved at pH 50 and despite that at acidic pH (ge 30) therelative activity was higher than 80 at lower pHs (20) itdecreased almost in a 50 Nevertheless at basic pHs therelative laccase activity presented a sharp decreased that iswell correlate with the other laccase fractions We are not ableto attribute this particular trend to the presence of multiplelaccase isoenzymes in the fraction since the SDS-PAGErevealed only one band when stained with ABTS and from theNanoLC-MSMS analysis no difference in the number ofpeptides and their sequence was detected We speculate thatthis particular behavior can be owed to a compound added inthe pre-treatment of the cedar wood that might be used forpreserving it that might affect the activity of Lac2 and that wasnot completely removed during the purification
Temperature stability evaluation showed that Lac1 is themost thermolabile fraction while the laccase activity reductionin the crude extracts did not seem to be the result of anadditive effect of the activity loss of the purified fractionsAlthough there are no reports of laccase stability under theselected conditions these results agree with previous reportsthat state that fungal laccases rapidly decrease their activity attemperatures near 60 degC and thermal stability correlates withthe temperature range of the growth of the source organism[55] In addition we previously reported for a laccase crudeextract obtained from culturing T pubescens on coffee husk anactivity loss of 26 and 10 at 60 and 50 degC respectivelyafter 8 h at pH 60 using syringaldazine as substrate [48]
Finally an electrochemical characterization was carried outfor selected crude extracts and purified fractions The results
showed a difference between the electrochemicalperformances of extracts and purified fractions withsyringaldazine or ABTS These differences could be owed tothe different mechanisms for electron transfer as discussedabove However the overall performance of the obtained cyclicvoltammograms for ABTS and syringaldazine agrees withresults reported by Fernaacutendez-Saacutenchez et al [25] who used asaturated calomel reference electrode
Conclusions
We have shown the potential of coffee husk soybean podhusk and cedar sawdust as substrates for the laccaseproduction In several countries like Brazil and Colombia theproduction of coffee and soybean is of great importance andthe generated volume of residues is high The results showedthat semi-solid culture of T pubescens using coffee huskallows to obtain an important amount of laccase which couldbe favored by the phenolic-compounds ie tannins presentedin coffee husk While supplementation of the culture mediumwith copper favored the production of laccase in submergedand semi-solid culture using soybean pod husk and cedarsawdust semi-solid culture using coffee husk which presentedthe highest laccase volumetric yield did not require coppersupplementation This is an economic advantage that shouldbe considered when scaling up the process In this study Tpubescens showed its ability to produce two independentlaccases regardless of the used substrate However a higherlaccase activity was observed for crude extracts rather than forpurified fractions This suggests that Lac1 and Lac2 might havea synergistic effect on the final laccase activity rather than anadditive effect Based on this study the investigation of laccaseproduction by evaluating other agro-industrial wastes in whichphenolic-compounds constitute an important percentage oftheir composition is of great interest to avoid the use ofinducers that would increase the production cost
Author Contributions
Conceived and designed the experiments JFO CJAD OFSPerformed the experiments JCG SCM AR Analyzed the dataJCG SCM AR JFO CJAD OFS Wrote the manuscript AR JFOCJAD OFS
References
1 Vandamme EJ (2009) Agro-industrial residue utilization for industrialbiotechnology products In P NigamA Pandey Biotechnology for Agro-Industrial Residues Utilisation Netherlands Springer Verlag pp 3-11
2 Martiacutenez AT Speranza M Ruiz-Duentildeas FJ Ferreira P Camarero S etal (2005) Biodegradation of lignocellulosics microbial chemical andenzymatic aspects of the fungal attack of lignin Int Microbiol 8195-204 PubMed 16200498
3 Saacutenchez O Sierra R Almeacuteciga-Diacuteaz CJ (2011) Delignification processof agro-industrial aastes an alternative to obtain fermentablecarbohydrates for producing fuel In M Manzanera Alternative FuelIntech 111-154
4 Wesenberg D Kyriakides I Agathos SN (2003) White-rot fungi andtheir enzymes for the treatment of industrial dye effluents BiotechnolAdv 22 161-187 doi101016jbiotechadv200308011 PubMed14623049
5 Morozova OV Shumakovich GP Shleev SV Yaropolov YI (2007)Laccase-mediator systems and their applications A review ApplBiochem Microbiol 43 523-535 doi101134S0003683807050055
6 Baldrian P (2006) Fungal laccases - occurrence and properties FEMSMicrobiol Rev 30 215-242 doi101111j1574-4976200500010xPubMed 16472305
7 Rodriacuteguez Couto S Herrera JLT (2006) Industrial and biotechnologicalapplications of laccases A review Biotechnol Adv 24 500-513 doi101016jbiotechadv200604003 PubMed 16716556
8 Osma JF Toca-Herrera JL Rodriacuteguez-Couto S (2010) Uses oflaccases in the food industry Enzyme Res 2010 2010 918761PubMed 21048873
9 Rodriguez S (2008) Exploitation of biological wastes for the productionof value-added products under solid-state fermentation conditionsBiotechnol J 3 859-870 doi101002biot200800031 PubMed18543242
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 12 September 2013 | Volume 8 | Issue 9 | e73721
10 Osma JF Toca-Herrera JL Rodriacuteguez-Couto S (2011) Cost analysis inlaccase production J Environ Manage 92 2907-2912 doi101016jjenvman201106052 PubMed 21775046
11 Jain A Morlok CK Henson JM (2013) Comparison of solid-state andsubmerged-state fermentation for the bioprocessing of switchgrass toethanol and acetate by Clostridium phytofermentans Appl MicrobiolBiotechnol 97 905-917 doi101007s00253-012-4511-4 PubMed23111595
12 Singhania RR Patel AK Soccol CR Pandey A (2009) Recentadvances in solid-state fermentation Biochem Eng J 44 13-18 doi101016jbej200810019
13 Economou CN Makri A Aggelis G Pavlou S Vayenas DV (2010)Semi-solid state fermentation of sweet sorghum for the biotechnologicalproduction of single cell oil Bioresour Technol 101 1385-1388 doi101016jbiortech200909028 PubMed 19781936
14 Rodriacuteguez CS Santoro R Cameselle C Sanroman A (1997) Laccaseproduction in semi-solid cultures of Phanerochaete chrysosporiumBiotechnol Lett 19 995-998 doi101023A1018495216946
15 Osma JF Moilanen U Toca-Herrera JL Rodriacuteguez-Couto S (2011)Morphology and laccase production of white-rot fungi grown on wheatbran flakes under semi-solid-state fermentation conditions FEMSMicrobiol Lett 318 27-34 doi101111j1574-6968201102234xPubMed 21291496
16 Couto SR Gundin M Lorenzo M Sanroman MN (2002) Screening ofsupports and inducers for laccase production by Trametes versicolor insemi-solid-state conditions Proc Biochem 38 249-255 doi101016S0032-9592(02)00087-0
17 Galhaup C Goller S Peterbauer CK Strauss J Haltrich D (2002)Characterization of the major laccase isoenzyme from Trametespubescens and regulation of its synthesis by metal ions Microbiology148 2159-2169 PubMed 12101303
18 Sievers F Wilm A Dineen D Gibson TJ Karplus K et al (2011) Fastscalable generation of high-quality protein multiple sequencealignments using Clustal Omega Mol Syst Biol 7 539 PubMed21988835
19 Childs RE Bardsley WG (1975) The steady-state kinetics of peroxidasewith 22-azino-di-(3-ethyl-benzthiazoline-6-sulphonic acid) aschromogen Biochem J 145 93-103 PubMed 1191252
20 Harkin JM Obst JR (1973) Syringaldazine an Effective Reagent forDetecting Laccase and Peroxidase in Fungi Experientia 29 382-387
21 Nikupaavola ML Raaska L Itavaara M (1990) Detection of white-rotfungi by a non-roxic stain Mycol Res 94 27-31 doi101016S0953-7562(09)81260-4
22 Gonzalez LF Sarria V Sanchez OF (2010) Degradation ofchlorophenols by sequential biological-advanced oxidative processusing Trametes pubescens and TiO(2)UV Bioresour Technol 1013493-3499
23 Waterborg JH (2009) The Lowry method for protein quantitation In JWalker The Protein Protocols Handbook Humana Press pp 7-10
24 Harkin JM Obst JR (1973) Syringaldazine an effective reagent fordetecting laccase and peroxidase in fungi Cell Mol Life Sci 29381-387 doi101007BF01926734
25 Fernaacutendez-Saacutenchez C Tzanov T Guumlbitz GM Cavaco-Paulo A (2002)Voltammetric monitoring of laccase-catalysed mediated reactionsBioelectrochemistry 58 149-156 doi101016S1567-5394(02)00119-6PubMed 12414320
26 Bourbonnais R Leech D Paice MG (1998) Electrochemical analysis ofthe interactions of laccase mediators with lignin model compoundsBiochim Biophys Acta 1379 381ndash390 doi101016S0304-4165(97)00117-7 PubMed 9545600
27 Ferraz F Silva S (2009) Characterization of coffee husk biomass forbiotechnological purposes N Biotechnol 25 S256-S256 doi101016jnbt200906573
28 Pandey A Soccol CR Nigam P Brand D Mohan R et al (2000)Biotechnological potential of coffee pulp and coffee husk forbioprocesses Biochem Eng J 6 153-162 doi101016S1369-703X(00)00084-X PubMed 10959086
29 Sruamsiri S Silman P (2008) Nutritive composition of soybean by-products and nutrient digestibility of soybean pod husk Maejo Int J SciTechnol 2 568-576
30 Romkaew J Nagaya Y Goto M Suzuki J Umezaki T (2008) Poddehiscence in relation to chemical components of pod shell in soybeanPlant Prod Sci 11 278-282 doi101626pps11278
31 Nakamanee G Polbumroong T Phaikaew C (2003) Alternative use ofcrop waste to increase milk production Proceedings of the 8th Meetingof the Regional Working Group on Grazing and Feed Resources forSoutheast Asia Kuala Lumpur Malaysia 66 - 70
32 Kurek B Gaudard F (2000) Oxidation of spruce wood sawdust byMnO(2) plus oxalate a biochemical investigation J Agric Food Chem48 3058-3062 doi101021jf000015g PubMed 10898665
33 Weil J Brewer M Hendrickson R Sarikaya A Ladisc MR (1998)Continuous pH monitoring during pretreatment of yellow poplar woodsawdust by pressure cooking in water Appl Biochem Biotechnol 70-7299-111 doi101007BF02920127
34 Weil J Sarikaya A Rau S-L Goetz J Ladisch CM et al (1997)Pretreatment of yellow poplar sawdust by pressure cooking in waterAppl Biochem Biotechnol 68 21-40 doi101007BF02785978
35 Dekker RF Barbosa AM Giese EC Godoy SD Covizzi LG (2007)Influence of nutrients on enhancing laccase production byBotryosphaeria rhodina MAMB-05 Int Microbiol 10 177-185 PubMed18075999
36 Xiao YZ Hong YZ Li JF Hang J Tong PG et al (2006) Cloning ofnovel laccase isozyme genes from Trametes sp AH28-2 and analysesof their differential expression Appl Microbiol Biotechnol 71 493-501doi101007s00253-005-0188-2 PubMed 16283298
37 Crowe JD Olsson S (2001) Induction of laccase activity in Rhizoctoniasolani by antagonistic Pseudomonas fluorescens strains and a range ofchemical treatments Appl Environ Microbiol 67 2088-2094 doi101128AEM6752088-20942001 PubMed 11319086
38 Yaguumle S Terroacuten MC Gonzaacutelez T Zapico E Bocchini P et al (2000)Biotreatment of tannin-rich beer-factory wastewater with white-rotbasidiomycete Coriolopsis gallica monitored by pyrolysisgaschromatographymass spectrometry Rapid Commun Mass Spectrom14 905-910 doi101002(SICI)1097-0231(20000530)1410 PubMed10825255
39 Gowda NKS Ramana JV Prasad CS Singh K (2004) Micronutrientcontent of certain tropical conventional and unconventional feedresources of southern India Trop Anim Heatlth Pro 36 77-94 doi101023BTROP0000009522309491d PubMed 14979561
40 DePeters EJ Fadel JG Arana MJ Ohanesian N Etchebarne MA et al(2000) Variability in the chemical composition of seventeen selectedby-product feedstuffs used by the California dairy industry Prof AnimSci 16 69-99
41 Obernberger I Thek G (2004) Physical characterisation and chemicalcomposition of densified biomass fuels with regard to their combustionbehaviour Biomass Bioenerg 27 653-669 doi101016jbiombioe200307006
42 Larous S Meniai A-H Lehocine MB (2005) Experimental study of theremoval of copper from aqueous solutions by adsorption usingsawdust Desalination 185 483ndash490 doi101016jdesal200503090
43 Oliveira WE Franca AS Oliveira LS Rocha SD (2008) Untreatedcoffee husks as biosorbents for the removal of heavy metals fromaqueous solutions J Hazard Mater 152 1073ndash1081 doi101016jjhazmat200707085 PubMed 17804159
44 Šciban M Klašnja M Škrbic B (2008) Adsorption of copper ions fromwater by modified agricultural by-products Desalination 229 170ndash180doi101016jdesal200708017
45 Maumlkelauml MR Hildeacuten KS Hakala TK Hatakka A Lundell TK (2006)Expression and molecular properties of a new laccase of the white rotfungus Phlebia radiata grown on wood Curr Genet 50 323-333 doi101007s00294-006-0090-1 PubMed 16927090
46 Maumlkelauml MR Lundell T Hatakka A Hildeacuten K (2013) Effect of coppernutrient nitrogen and wood-supplement on the production of lignin-modifying enzymes by the white-rot fungus Phlebia radiata Fungal Biol117 62-70 doi101016jfunbio201211006 PubMed 23332834
47 Dwivedi UN Singha P Pandeya VP Kumara A (2011) Structurendashfunction relationship among bacterial fungal and plant laccases J MolCatal B Enzym 68 117-128 doi101016jmolcatb201011002
48 Gaitan IJ Medina SC Gonzaacutelez JC Rodriacuteguez A Espejo AacuteJ et al(2011) Evaluation of toxicity and degradation of a chlorophenol mixtureby the laccase produced by Trametes pubescens Bioresour Technol102 3632ndash3635 doi101016jbiortech201011040 PubMed21115244
49 Karp SG Faraco V Amore A Birolo L Giangrande C et al (2012)Characterization of laccase isoforms produced by Pleurotus ostreatusin solid state fermentation of sugarcane bagasse Bioresour Technol114 735-739 doi101016jbiortech201203058 PubMed 22487128
50 Nikitina OV Shleev SV Gorshina ES Rusinova TV Serezhenkov VAet al (2005) Isolation and purification of enzymes from ligninolyticcomplex of the basidial fungus Trametes pubescens (Schumach) Pilatand study of their properties Biochemistry (Mosc) 70 1274-1279 doi101007s10541-005-0259-0
51 Shleev S Nikitina O Christenson A Reimann CT Yaropolov AI et al(2007) Characterization of two new multiforms of Trametes pubescenslaccase Bioorg Chem 35 35-49 doi101016jbioorg200608001PubMed 16989887
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 13 September 2013 | Volume 8 | Issue 9 | e73721
52 Xiao YZ Chen Q Hang J Shi YY Wu J et al (2004) Selectiveinduction purification and characterization of a laccase isozyme fromthe basidiomycete Trametes sp AH28-2 Mycologia 96 26-35 doi1023073761984 PubMed 21148825
53 Muntildeoz C Guilleacuten F Martiacutenez AT Martiacutenez MJ (1997) Laccaseisoenzymes of Pleurotus eryngii characterization catalytic propertiesand participation in activation of molecular oxygen and Mn2+ oxidationAppl Environ Microbiol 63 2166-2174 PubMed 9172335
54 Morozova OV Shumakovich GP Gorbacheva MA Shleev SVYaropolov AI (2007) Blue laccases Biochemistry (Mosc) 721136-1150 doi101134S0006297907100112
55 Hildeacuten K Hakala TK Lundell T (2009) Thermotolerant andthermostable laccases Biotechnol Lett 31 1117-1128 doi101007s10529-009-9998-0 PubMed 19360388
56 Verma P Madamwar D (2002) Production of ligninolytic enzymes fordye decolorization by cocultivation of white-rot fungi Pleurotusostreatus and Phanerochaete chrysosporium under solid-statefermentation Appl Biochem Biotechnol 102-103 109-118 doi101385ABAB102-1031-6109 PubMed 12396115
57 Meza JC Lomascolo A Casalot L Sigoillot J-C Auria R (2005)Laccase production by Pycnoporus cinnabarinus grown on sugar-canebagasse Influence of ethanol vapours as inducer Proc Biochem 403365ndash3371 doi101016jprocbio200503004
58 Hossain SM Anantharaman N (2006) Activity enhancement ofligninolytic enzymes of Trametes versicolor with bagasse powder Afr JBiotechnol 5 189-194
59 Revankar MS Desai KM Lele SS (2007) Solid-state fermentation forenhanced production of laccase using indigenously isolatedGanoderma sp Appl Biochem Biotechnol 143 16-26 doi101007s12010-007-0029-0 PubMed 18025593
60 Velazquez-Cedeno MA Mata G Savoie J-M (2002) Waste-reducingcultivation of Pleurotus ostreatus and Pleurotus pulmonarius on coffeepulp changes in the production of some lignocellulolytic enzymesWorld J Microbiol Biotechnol 18 201ndash207 doi101023A1014999616381
61 Mata G Murrieta D Andreu LI (2005) Changes in lignocellulolyticenzyme activities in six Pleurotus spp strains cultivated on coffee pulpin confrontation with Trichoderma spp World J Microbiol Biotechnol 21143ndash150 doi101007s11274-004-3041-3
62 de Souza DF Tychanowicz GK de Souza CG Peralta RM (2006) Co-production of ligninolytic enzymes by Pleurotus pulmonarius on wheatbran solid state cultures J Basic Microbiol 46 126-134 doi101002jobm200510014 PubMed 16598826
63 Prasad KK Mohan SV Bhaskar YV Ramanaiah SV Babu VL et al(2005) Laccase production using Pleurotus ostreatus 1804 immobilizedon PUF cubes in batch and packed bed reactors influence of cultureconditions J Microbiol 43 301-307 PubMed 15995650
64 Songulashvili G Elisashvili V Wasser SP Nevo E Hadar Y (2007)Basidiomycetes laccase and manganese peroxidase activity in
submerged fermentation of food industry wastes Enzyme MicrobTechnol 41 57ndash61 doi101016jenzmictec200611024
65 Sathishkumar P Murugesan K Palvannan T (2010) Production oflaccase from Pleurotus florida using agro-wastes and efficientdecolorization of Reactive blue 198 J Basic Microbiol 50 360-367 doi101002jobm200900407 PubMed 20586068
66 Moldes D Gallego PP Rodriacuteguez Couto S Sanromaacuten A (2003) Grapeseeds the best lignocellulosic waste to produce laccase by solid statecultures of Trametes hirsuta Biotechnol Lett 25 491-495 doi101023A1022660230653 PubMed 12882277
67 Elisashvili V Kachlishvili E Penninckx M (2008) Effect of growthsubstrate method of fermentation and nitrogen source onlignocellulose-degrading enzymes production by white-rotbasidiomycetes J Ind Microbiol Biotechnol 35 1531-1538 doi101007s10295-008-0454-2 PubMed 18716810
68 Osma JF Saravia V Herrera JL Couto SR (2007) Mandarin peelingsthe best carbon source to produce laccase by static cultures ofTrametes pubescens Chemosphere 67 1677-1680 doi101016jchemosphere200611051 PubMed 17234250
69 Kahraman SS Gurdal IH (2002) Effect of synthetic and natural culturemedia on laccase production by white rot fungi Bioresour Technol 82215-217 doi101016S0960-8524(01)00193-6 PubMed 11991068
70 Iandolo D Piscitelli A Sannia G Faraco V (2011) Enzyme Productionby Solid Substrate Fermentation of Pleurotus ostreatus and Trametesversicolor on Tomato Pomace Appl Biochem Biotechnol 163 40-51doi101007s12010-010-9014-0 PubMed 20582639
71 Moldes D Lorenzo M Sanromaacuten MA (2004) Different proportions oflaccase isoenzymes produced by submerged cultures of Trametesversicolor grown on lignocellulosic wastes Biotechnol Lett 26 327-330doi101023BBILE000001545240213bf PubMed 15055770
72 Philippoussis A Diamantopoulou P Papadopoulou K Lakhtar HRoussos S et al (2010) Biomass laccase and endoglucanaseproduction by Lentinula edodes during solid state fermentation of reedgrass bean stalks and wheat straw residues World J MicrobiolBiotechnol 27 285-297
73 Fenice M Giovannozzi Sermanni G Federici F DrsquoAnnibale A (2003)Submerged and solid-state production of laccase and Mn-peroxidaseby Panus tigrinus on olive mill wastewater-based media J Biotechnol100 77-85 doi101016S0168-1656(02)00241-9 PubMed 12413788
74 Rodriguez-Couto S GundIn M Lorenzo M Sanromaacuten MAacute (2002)Screening of supports and inducers for laccase production by Trametesversicolor in semi-solid-state conditions Proc Biochem 38 249-255doi101016S0032-9592(02)00087-0
75 Fackler K Gradinger C Schmutzer M Tavzes C Burgert I et al (2007)Biotechnological wood modification with selective white-rot fungi and itsmolecular mechanisms Food Technol Biotechnol 45 269ndash276
76 Levin L Herrmann C Papinutti VL (2008) Optimization oflignocellulolytic enzyme production by the white-rot fungus Trametestrogii in solid-state fermentation using response surface methodologyBiochem Eng J 39 207-214 doi101016jbej200709004
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 14 September 2013 | Volume 8 | Issue 9 | e73721

collected and analyzed by sodium dodecyl sulfate-polyacrylamide electrophoresis gel (SDS-PAGE) under non-reducing conditions and without heat denaturation Staining ofthe electrophoresis gel was done with 5 mM 22rsquo-azino-bis(3-ethylbenzothiazoline-6) sulphonic acid (ABTS) in succinic acid(25 mM pH 45) to detect laccase activity Fractions thatrevealed the same protein profile and laccase activity werecombined Samples of the combine fractions were analyzed bySDS-PAGE under mentioned conditions As a negative controlwere used a sample of the ultrafiltrated fraction which did notreveal laccase activity along the previous purification stepBands that revealed laccase activity were identified as Lac1and Lac2 These bands were excised from the electrophoresisgel and analyzed through nanoLC-MSMS (Applied BiomicsHayward CA USA) Peptides and protein sequences werealigned by using Clustal Omega [18] at EBI web server andBlast at NCBI web server Alignments figures were generatedusing CLC Sequence Viewer (CLC bio Denmark)
Enzyme activity was determined spectroscopically by usingeither 22rsquo-azino-bis(3-ethylbenzothiazoline-6) sulphonic acid(ABTS ε420 = 36mM-1 cm-1 Sigma-Aldrich) or 4-hydroxy-35-dimethoxybenzaldehyde (Syringaldazine ε530 = 65mM-1 cm-1Sigma-Aldrich) [19ndash21] ABTS activity assay was carried out aspreviously described [2122] Briefly 850 microL of 5 mM ABTS insuccinic acid (25 mM pH 45) were mixed with 150 microL of therespective dilution of culture extract and the absorbance was at420 nm Syringaldazine activity assay was carried out followingthe substrate manufacturerrsquos instructions (Sigma-Aldrich) Oneunit (U) was defined as the amount of enzyme required tooxidize 1 micromol of substrate (ABTS or syringaldazine) perminute Specific activity was expressed as U mgminus1 of protein asdetermined by Lowry assay [23]
Laccase production during the fungus culture was monitoredusing syringaldazine which has demonstrated to be a specificsubstrate for detecting extracellular laccase in the absence ofhydrogen peroxide [24] Absence of hydrogen peroxide wasconfirmed by incubation with catalase Crude extracts andpurified fractions were analyzed by SDS-PAGE withoutreduction by β-mercaptoethanol and heat denaturationProteins were visualized by silver staining while laccases wereidentified by placing the SDS-PAGE gel in an ABTS solution for5 min Temperature and pH effect on laccase activity wasdetermined for the UF retentate and the purified fractions byusing ABTS The temperature effect on laccase activity wasevaluated at 30 40 50 55 60 and 70 degC at pH 50 and the pHeffect on laccase activity was evaluated at 2 25 3 4 5 6 65and 7 at 55 degC Laccase stability was evaluated by using ABTSat optimal pH and temperature for 48 h
Cyclic voltammetry analysisCyclic voltammetry (CV) analysis was performed on a
computer-controlled potentiostat (Autolab PotentiostatGalvanostat PGSTAT 128N Metrohm USA) Data wereacquired by using the software Nova version 16 Aconventional three-electrode system was used to carry out thecyclic voltammograms The used electrodes were graphite-working electrode with a surface area of 023 cm2 AgAgCl KClsaturated reference electrode (Metrohm USA) and platinum
wire counter electrode Before running each experiment thesurface of the graphite electrode was polished with aluminaand thoroughly rinsed with distilled water CV analysis wasdone to the crude extract and purified laccase fractions thatpresented the highest activity for each substrate For the test20 microL of sample were added on the polished surface of theworking electrode and allowed to dry for 60 min at roomtemperature Each experiment was performed in triplicate usingeither a solution 11 of anhydride ethanol buffer acetate (100mM pH 50) and syringaldazine to a final concentration of 02mM or a solution of 100 mM acetate buffer (pH 30) and 05mM ABTS Cyclic voltammograms were carried out in 30 mL ofworking solution with continuous agitation Voltage wasscanned at 100 mV s-1 between -02 and 1 V vs Ag|AgCl|KClsat
for syringaldazine assays while for ABTS assays voltage wasscanned at 250 mV s-1 between -02 and 12 V vs Ag|AgCl|KClsat Assays were conducted in a Faraday cage at roomtemperature (20-22degC) Temperature and pH in the reactionsolution was monitored with the Unitrode with Pt 1000(Metrohm USA)
Statistical analysisAll experiments were conducted by triplicate Data are
presented as the mean plusmn standard deviation (SD) Statisticalcomparisons were conducted by using either t-test or Duncanrsquostest for one-factor design by using StatGraphics Centurionreg v16111 (2011) Statistical differences were established for p lt005
Results
SmC and SSC production of laccase by T pubescensCoffee husk soybean pod husk and cedar sawdust were
used as substrate for the production of laccase by Tpubescens under SmC or SSC conditions while syringaldazinewas used to monitor laccase production during the fungusculture under the studied conditions Hydrogen peroxide wasnot detected in any of the evaluated samples which discardthe interference of peroxidases in the syringaldazine assayLaccase activity and glucose concentration time course for Tpubescens grown on coffee husk under SmC and SSC atdifferent copper concentrations are presented in Figure 1Although a similar time course was observed for laccaseactivity and glucose concentration of soybean pod husk andcedar sawdust cultures differences on the day of maximumlacasse activity as well as in the maximum activity value wereobserved among the agro-industrial byproducts (Table 2) Ingeneral it was noticed that laccase activity was detectablearound the seventh and fourth day of culture for SmC andSSC respectively After the eighth culture day glucose wasdepleted from the culture medium and laccase activityincreased sharply
The substrate source seemed to be the most important factorfor laccase production (Table 2) T pubescens culture oncoffee husk allowed the maximum laccase activities for bothSmC (17723plusmn372 U L-1) and SSC (20400plusmn596 U L-1) atcopper concentrations of 5 and 0 microM respectively Laccaseactivity in SSC and SmC using coffee husk as substrate was
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 4 September 2013 | Volume 8 | Issue 9 | e73721
between 147- and 663-fold higher than that observed forcultures with soybean pod husk and cedar sawdust Laccaseactivity obtained in the control culture without agro-industrialwaste presented a maximum laccase activity of 222 plusmn 088 UL-1 after 14 days of culture while the glucose was depleted onthe seventh day of culture
Copper supplementation effect on laccase activity wasevaluated for both SmC and SSC Although in a non-linearmode the laccase activity was increased by the presence ofcopper with all substrates under SmC or SSC except withcoffee husks under SSC (Table 2) In addition the requiredcopper concentration to obtain the highest laccase activityvaried depending on the used agro-industrial waste Forexample laccase production was favored in cultures withsoybean pod husk at copper concentrations of 05 mM and 20mM for SmC and SSC respectively while for cultures with
Figure 1 Laccase activity profiles during T pubescensCBS 69694 culture under SmC (a) and SSC (b) on coffeehusk Culture media was enriched with different copperconcentrations 0 (Δ) 05 () 2 () and 5 (loz) mM and laccaseactivity was determined by using syringaldazine as substrateFilled symbols refer to the glucose concentration at therespective copper concentration All the assays wereperformed in triplicatedoi 101371journalpone0073721g001
coffee husk the highest laccase activity levels for SmC andSSC was observed at copper concentrations of 5 mM and 0mM respectively
Despite that culture mode ie SmC or SSC presented aneffect on laccase production this is substrate sensitive In theassays conducted without copper it was observed that laccaseactivity was 28- and 33-fold higher in SSC than in SmC for thesoybean pod husk and coffee husk respectively Howeverwhen cedar sawdust was used no significant difference inlaccase activity between the two culture types was obtained(Table 2) Due to the low volume of liquid media and highlaccase activity in SSC when compared to SmC the SSCpresented a higher volumetric yield For all agro-industrialwaste the SSC presented a shorter time than SmC to reachthe highest laccase activity which suggest a higher productivityunder SSC conditions
These results show that coffee husk under SmC or SSCconditions could be considered as a potential alternative forlaccase production by T pubescens However soybean podhusk and cedar sawdust could be also considered as importantalternatives for laccase production especially in comparisonwith synthetic culture media [10] Finally the optimal copperconcentration to obtain the highest laccase activity depends onthe agro-industrial byproduct used for the fungus culture
Laccase purification and characterizationLaccase purification was conducted using the crude extracts
that presented the highest activities For the SmC crude
Table 2 Effect of substrate copper concentration andculture mode on laccase activity (UL-1)
Substrate SmC SSCSoybean Pod Husk + 0 mM Cu+2 446 plusmn 138 (23) 1255 plusmn 052 (13)
Soybean Pod Husk + 05 mM Cu+2
8493 plusmn 846 (23)dagger 2028 plusmn 387 (21)
Soybean Pod Husk + 2 mM Cu+2 5761 plusmn 370 (19) 13785 plusmn 071 (18)dagger
Soybean Pod Husk + 5 mM Cu+2 4492 plusmn 487 (23) 2900 plusmn 407 (18)Coffee Husk + 0 mM Cu+2 6184 plusmn 176 (23) 20400 plusmn 596 (19)dagger
Coffee Husk + 05 mM Cu+2 13763 plusmn 568 (23) 11083 plusmn 940 (20)ab
Coffee Husk + 2 mM Cu+2 12304 plusmn 373 (23)b 10278 plusmn 814 (15)b
Coffee Husk + 5 mM Cu+2 17723 plusmn 372 (23)dagger 8266 plusmn 882 (15)Cedar Sawdust + 0 mM Cu+2 1355 plusmn 095 (17) 1082 plusmn 053 (15)
Cedar Sawdust + 05 mM Cu+2 2272 plusmn 084 (17) 1902 plusmn 002 (13)Cedar Sawdust + 2 mM Cu+2 2897 plusmn 127 (12)a 2638 plusmn 068 (8)Cedar Sawdust + 5 mM Cu+2 2936 plusmn 153 (13) abdagger 3073 plusmn 152 (12)bdagger
Data in parenthesis refers to the day where was observed the maximum laccaseactivity No Statistical Difference Statistical Difference (Evaluation of substrate effecton laccase activity for both culture modes submerged (SmC) and semisolid (SSC)no copper induction considered)Equal letters inside one agro-industrial waste group refers to No StatisticalDifference no letters refers to Statistical Difference inside the group (Evaluation ofcopper effect on laccase activity for both SmC and SSC)dagger Maximum laccase activity for the used agro-industrial wastes either in SmC orSSC culture
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 5 September 2013 | Volume 8 | Issue 9 | e73721
extracts obtained with soybean pod husk + Cu+2 05 mM coffeehusk + Cu+2 5 mM and cedar sawdust + Cu+2 5 mM wereselected while for SSC crude extracts obtained with soybeanpod husk + Cu+2 2 mM coffee husk + Cu+2 0 mM and cedarsawdust + Cu+2 5 mM were selected The electrophoreticanalysis of permeates from 045 and 022 microm filtrations and theUF retentate showed the presence of two proteins with laccaseactivity while no activity was observed within the ultrafiltratedsample (Figure 2a) These proteins presented a molecularmass of about 120 kDa and 60 kDa (Figure 2a) Thepurification of the UF retentate by ionic exchangechromatography confirmed the presence of two differentfractions with laccase activity (Figure 2b) The SDS-PAGEanalysis of these fractions under non-reducing conditionsshowed that one protein with laccase activity was presented oneach fraction (Figure 2b) and were named Lac1 (120 kDa) andLac2 (60 kDa) Considering that the conditions used tomeasure the enzyme activity were those suggested for laccaseactivity and that H2O2 or manganese were not added duringthe reaction for measuring the enzyme activity these resultssuggest that both proteins are laccases but not peroxidases
NanoLC-MSMS analysis of purified fractions showed thatthe identified peptides for Lac1 (SAGSTTYNYNDPIFR andRDTVSTGNSGDNVTIR) and Lac2 (SAGSTVYNYDNPIFR andANPNFGNVGFTGGINSAI) had identity scores between 80 to100 with laccases (over 100 sequences) available at the non-redundant GenBank dataset Proteins containing thesepeptides different that laccases were not identified Similarresults were observed when peptides were evaluated atUniprot Galhaup et al [17] described two laccase activefractions from T pubescens MB 89 (CBS 69694) Lap1(UniProtKBTrEMBL Q8TG93) and Lap2 (UniProtKBTrEMBLQ8TG94) which share a 70 identity (Figure 3a) Sequencealignment of Lac1 and Lac2 peptides with Lap1 and Lap2sequences respectively showed that the peptides arepresented within T pubescens MB 89 laccase sequencesthough some differences were observed for the Lac1 peptide(Figure 3b) This result suggests that the purified fractions arelaccase isoenzymes
Protein concentration and volumetric and specific laccaseactivities at different purification stages are presented in Table3 Lac1 presented lower laccase activity than Lac2 in a ratiothat ranges from 080- to 38-fold depending on the substratesource On the other hand laccase activity of the UF retentatefrom soybean pod husk culture was from 45- to 64-fold higherthan that of the purified fractions while for cedar sawdust andcoffee husk the laccase activity was from 46- to 120-fold andfrom 33- to 163-fold higher than the purified fractionsrespectively (Table 3) The purified Lac2 fractions from coffeehusk and cedar sawdust cultures showed the highest specificactivity levels (Table 3) The volumetric activities of UFretentates were higher than that obtained with purified fractions(Table 4)
Data in parenthesis shows the temperature or pH at whichrelative laccase activity reached 100
Temperature and pH effect on laccase activity wasdetermined for the UF retentate and Lac1 and Lac2 fractionsobtained along the purification of the enzymatic crude extracts
that resulted after the SSC of T pubescens on the selectedagro-industrial wastes Laccase activity for the UF retentateunder different temperature and pH conditions is presented inFigure 4 and Table 4 The maximum relative activity of theevaluated UF retentates was observed at 50-55 degC and at apH range from 20 to 40 However UF retentate from cedar
Figure 2 Purification of lacasses produced by submergedculture of T pubescens using coffee husk assubstrate All samples were analyzed by SDS-PAGE undernon-reducing conditions and stained with ABTS No heatdenaturation of the samples was conducted (a) The crudeextract (1) was centrifuged and filtered through a Whatman No1 filter (2) 045 microM membrane (3) 022 microm membrane (4) andultrafiltrated through a 10 kDa cut-off membrane permeate (5)and retentate (6) (b) UF retentate was purified by anionicexchange chromatography Lac1 represent the unboundfraction while Lac2 represent the eluted fraction The arrowshows the point when protein elution was started (c) Purifiedfractions (1) crude extract (2) Lac1 and (3) Lac2 MWMolecular Weight Similar results were observed for crudeextracts produced by using soybean pod husk and cedarsawdustdoi 101371journalpone0073721g002
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 6 September 2013 | Volume 8 | Issue 9 | e73721
sawdust culture behaved different to the UF retentate fromcoffee husk and soybean pod husk cultures In this sense the
UF retentate from cedar sawdust culture presented the lowestenzyme activity at temperatures le 40 degC but at 70 degC
Figure 3 Aligment of reported lacasses sequences and Lac1 and Lac2 peptides (a) Alignment of Lap1 (UniProtKBSwiss-Prot Q8TG93) and Lap2 (UniProtKBSwiss-Prot Q8TG94) from T pubescens CBS 69694 (b) Alignment of Lac1 and Lac2 peptideswith Lap1 and Lap2 respectively and tryptic peptides described by Shleev et al 2007 Alignments were carried out using ClustalOmega and figures were using CLC Sequence Viewer Different residues are colored in reddoi 101371journalpone0073721g003
Table 3 Laccase activity during the purification of crude extracts resulted from T pubescens culture on agro-industrialwastes under submerged (SmC) and semisolid (SSC) cultures
Soybean pod husk Coffee husk Cedar sawdust
Protein(mg mL-1)
VolumetricActivity (UL-1)
SpecificActivity (Umg-1
protein)Recovery()
Protein(mg mL-1)
VolumetricActivity (UL-1)
SpecificActivity (Umg-1
protein)Recovery()
Protein(mg mL-1)
VolumetricActivity (UL-1)
SpecificActivity (Umg-1
protein)Recover()
SmCCrudeExtract
0869 52888 0608 100 0414 152708 3688 100 0431 10735 0249 100
045 micromFiltration
0736 46549 0632 88 0334 117892 3529 77 0444 12053 0271 112
UF retentate 1010 78731 0779 74 0378 195263 5165 64 0510 25905 0507 120
Lac1 0494 5471 0110 1 0232 2399 0103 02 0222 838 0037 08
Lac2 0310 20661 0666 4 0116 107240 9252 7 0040 15234 3837 14 SSCCrudeExtract
1146 164966 1439 100 0596 154907 2601 100 1065 23203 0217 100
045 micromFiltration
0776 128392 1654 77 0474 115736 2439 75 0806 25824 0320 111
UF retentate 0973 335055 3443 101 0428 161750 3779 52 1134 51101 0450 110
Lac1 0097 8854 0912 2 0104 1514 0145 03 0042 1315 0313 2
Lac2 0168 48988 2915 10 0131 82298 6282 18 0072 16162 2244 23
740 ABTS was used to determine laccase activity
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 7 September 2013 | Volume 8 | Issue 9 | e73721
presented the highest enzyme activity ~90 of its initial valuein comparison to the other two UF retentates (Figure 4)
The temperature and pH effect on laccase activity for Lac1and Lac2 is presented in Figure 5 The maximum relativeactivity for both fractions was presented at a pH from 20 to 40regardless of the agro-industrial waste as observed for the UFretentate However at pH gt 40 Lac1 showed a sharperdecrease in its activity than that observed for Lac2Nevertheless Lac2 purified from cedar sawdust culturepresented a different performance This fraction showed themaximum activity at pH 50 while a sharp drop was observedat pHs above or below this value
Laccase activity profiles showed that Lac1 and Lac2presented the maximum enzyme activity at 55 and 60 degCrespectively (Table 4) It was observed that Lac1 from coffeehusk culture presented a lower decrease in enzyme activity attemperatures le 50 degC or ge 60 degC than Lac2 On the contraryLac2 from soybean husk or cedar sawdust cultures presenteda higher relative laccase activity than Lac1 at temperatures le55 degC The purified fractions from soybean pod husk and cedarsawdust cultures presented the lowest relative activity at 30 degCand 65 degC
Table 4 Optimum temperatures and pHs for maximumlaccase activities in UF retentate Lac1 and Lac2 obtainedfrom semisolid cultured T pubescens on agro-industrialwastes
EnzymaticFraction Temperature pH
VolumetricActivity (UL-1)
SpecificActivity (Umg-1 protein)
VolumetricActivity (UL-1)
SpecificActivity (Umg-1
protein)SoybeanPod Husk +2 mM Cu+2
148000plusmn1178(55)
1521plusmn0012182037plusmn3025(2-3)
1871plusmn0031
Lac122925plusmn917(60)
2363plusmn009536135plusmn831(23)
3725plusmn0085
Lac232638plusmn1558(55)
1942plusmn009228713plusmn274(23)
1709plusmn0016
CoffeeHusk + 0mM Cu+2
113740plusmn8289(50-55)
3008plusmn0219290666plusmn1039(2-3)
7689plusmn0275
Lac1 889plusmn039 (55) 0038plusmn0002 1779plusmn028 (23) 0077plusmn0001
Lac234194plusmn2789(60)
2947plusmn024033150plusmn526 (2ndash4)
2857plusmn0045
CedarSawdust +5 mM Cu+2
257666plusmn1201(55)
5052plusmn0024344634plusmn1043(2-3)
6757plusmn0204
Lac121370plusmn472(55)
0962plusmn002129117plusmn943(23)
1311plusmn0042
Lac255166plusmn1656(60)
1379plusmn0414 55166plusmn1656 (5) 1379plusmn0414
ABTS was used to determine laccase activity
Laccase stabilityFor selected enzymatic extracts and purified fractions (Table
4) a stability test for 48 h at their optimal pH and temperaturewas conducted Time course of the relative laccase activity forcrude extract and purified fractions from coffee husk and cedarsawdust cultures are presented in Figure 5 Similar resultswere observed for crude extract and purified fractions fromsoybean pod husk culture (data no shown)
It was observed that the activity loss depends on thesubstrate source used to grow T pubescens An activity loss of55 was observed for crude extracts and purified fractionsfrom coffee husk and soybean pod husk cultures while anactivity loss of 70 was observed for the crude extract andpurified fractions from cedar sawdust culture NeverthelessLac2 showed the lowest activity reduction with values of 10 40and 55 for coffee husk soybean pod husk and cedarsawdust respectively while Lac1 presented an activity loss of34 50 and 60 for coffee husk soybean pod husk and cedarsawdust respectively Despite the differences in laccaseactivity among agro-industrial wastes and between crudeextracts and purified fractions all the profiles showed a sharp
Figure 4 Effect of temperature (a) and pH (b) on laccaseactivity for the UF retentate obtained by semisolid cultureof T pubescens on coffee husk () soybean pod husk (loz)and cedar sawdust (Δ) Laccase stability was evaluated byusing ABTS All the assays were performed in triplicatedoi 101371journalpone0073721g004
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 8 September 2013 | Volume 8 | Issue 9 | e73721
decrease during the first 8 h while a reduced activity loss wasobserved during the last 24 h of the assay (Figure 6)
Laccase electrochemical characterizationFor selected crude extracts and purified fractions a cyclic
voltammetry characterization was conducted For eachmediator syringaldazine or ABTS similar cyclic
Figure 5 Effect of temperature () and pH () on enzymeactivity for Lac1 (solid line) and Lac2 (dashed line)obtained after the purification of the enzymatic crudeextract produced by semisolid culture of T pubescens oncoffee husk (a) soybean pod husk (b) and cedar sawdust(c) Laccase stability was evaluated by using ABTS All theassays were performed in triplicatedoi 101371journalpone0073721g005
voltammograms were obtained for the different crude extractsand purified fractions Voltammograms of the redox reaction forsyringaldazine (Figure 7a) and ABTS (Figure 6b) arepresented Two anionic and cationic peaks were observedwhen syringaldazine was used as mediator The first pair ofpeaks showed a reversible performance with a mean Ia Icratio of 109plusmn006 106plusmn005 and 098plusmn002 for the crudeextracts Lac1 and Lac2 respectively The second pair ofpeaks showed quasi-reversible performance and the cathodicpeak was presented at a potential of +062 V (vs Ag|AgCl)while the anodic peak was presented at a potential of +060 V(vs Ag|AgCl) No significant difference on the reductionpotential (Eo) for syringaldazine was observed for the crudeextracts or the purified fractions obtaining an Eo of0314plusmn0008 V (vs Ag|AgCl) However the peak potentialseparation (ΔEp) showed a different profile depending on thesubstrate used for the culture While not significant difference(p gt 005) on ΔEp value was observed between crude extract(0045plusmn0005 V vs Ag|AgCl) Lac1 (0046plusmn0005 V vs Ag|AgCl)and Lac2 (0028plusmn0001 V vs Ag|AgCl) from cultures with cedarsawdust and soybean pod husk the ΔEp was considerablyhigher for the crude extract (0083plusmn0004 V vs Ag|AgCl) Lac1(0068plusmn0003 V vs Ag|AgCl) and Lac2 (0051plusmn0005 V vs Ag|AgCl) from the coffee husk culture
For reactions conducted with ABTS and the different crudeextracts and purified fractions the voltammograms exhibitedone pair of cathodic and anodic peaks (Figure 7b) whichcorresponds to the redox of ABTSABTS+ (IaIc) with an Eo of0487plusmn0006 V (vs Ag|AgCl) This process was reversiblepresenting a mean Ia Ic ratio of 099plusmn002 In addition asecond cathodic peak was presented at +083 V (vs Ag|AgCl)when ABTS redox reaction was conducted using the crudeextract and the purified fractions This performance agrees withprevious reports in which the laccase activity cannot beobserved at potential values of the second reversible oxidation
Figure 6 Laccase stability profiles at optimal pH andtemperature conditions for crude extracts (solid line) Lac1(dashed line) and Lac2 (spotted line) obtained fromsemisolid culture of T pubescens on coffee husk (loz) andcedar sawdust (bull) Laccase stability was evaluated by usingABTS All the assays were performed in triplicatedoi 101371journalpone0073721g006
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 9 September 2013 | Volume 8 | Issue 9 | e73721
of ABTS ABTS+ABTS2+ due to this catalytic process cannotbe detected by cyclic voltammetry [2526] Neverthelessvoltammograms for laccase crude extract from a soybean podhusk culture presented the second anodic peak which inaddition to the second cathodic peak corresponds to the redoxof ABTS+ABTS2+ (IIaIIc) with an Eo of 0942plusmn0004 V (vs Ag|AgCl) Contrary to the reactions conducted usingsyringaldazine as substrate the ΔEp did not present asignificant difference for the crude extracts and purifiedfractions 0248plusmn0016 V (vs Ag|AgCl)
Discussion
In this study we evaluated the effect of the culture mode ieSmC and SSC on laccase production by T pubescens CBS69694 using three different agro-industrial wastes (coffeehusk soybean pod husk and cedar sawdust) Furthermore theeffect of Cu+2 supplementation on laccase production was alsoevaluated and the crude extracts and the purified fractionswere partially characterized
Figure 7 Cyclic voltammograms of (a) syringaldazine atpH 50 using the crude extract and purified laccasefractions from semisolid culture of T pubescens on coffeehusk or (b) ABTS at pH 30 using the crude extract andpurified laccase fractions from semisolid culture of Tpubescens on soybean pod husk Nomenclature onlymediator (dotted line) crude extract (solid line) Lac1 fraction(dashed line) and Lac2 fraction (dashed and dotted line)doi 101371journalpone0073721g007
Substrate source for laccase production showed to be one ofthe most important factors for laccase production (Table 2)Cultures with coffee husk as substrate presented the highestlaccase production levels The observed differences in laccaseactivity for each substrate could be owed to their chemicalcomposition Coffea arabica is the coffee variety raised inColombia and its husk has a reported composition of 2308cellulose (2076 glucose and 183 cellobiose) 2385hemicellulose (1356 xylose 523 arabinose 256 aceticacid and 195 glucuronic acid) 2828 total lignin and071 ashes [27] In addition coffee husk contents 13caffeine 45 tannins and 124 pectins [28] On the otherhand Glycine max is the soybean species cultured inColombia and its pod husk composition varies from 504 to62 crude protein 608 to 143 hemicellulose 3715 to4208 acid detergent fiber 4436 to 6015 neutral detergentfiber 850 to 893 acid detergent lignin and 2074 to 2106uronic acid [29ndash31] Finally for cedar sawdust the majorcomponents are glucose (426-444) xylose (46-195)cellulose (383-410) klason lignin (227-278) and acidlignin (40-43) [32ndash34] Although lignin is one of the mostimportant compounds associated with laccase production bywhite-rot fungi we did not observe a clear correlation betweenlignin composition percentage and laccase activity In thatsense the high laccase activity observed for the cultures thatuse coffee husk could be associated with its content of caffeineand tannins which have been identified as potent inducers oflaccase gene expression [173536] In fact caffeine has beenused as inducer for laccase production by Rhizoctonia solaniobtaining an increase of 41-fold in laccase activity [37] Similarresults were also reported for the production of Coloriopsisgallica laccase [38]
Laccase gene expression is mainly up regulated by Cu2+although Mn2+ Fe3+ heavy metals 26-dimethoxy-14-benzoquinone H2O2 amphotericin B syringic acid tannic acidTween 80 soybean oil aromatic compounds andmicroclimatic changes (ie lower temperature and osmoticpressure) are also recognized as potent inducers [173536] Ingeneral Cu2+ supplementation increased the laccase activity inboth SmC and SSC However Cu2+concentration required toobtain the highest laccase activity varied depending on agro-industrial waste These differences can be attributed to thecopper presented in the agro-industrial waste and the copperavailability in the medium Copper can be found in amountsthat range from 156 to 180 mg kg-1 of coffee husk [39] andfrom 26 to 67 mg kg-1 of soybean pod husk (or soybean hull)[3940] Despite that no reports indicate the copper content inuntreated cedar wood an average of 10 mg kg-1 of wood hasbeen reported for wood used as biomass in six different Europecountries [41] This allows to consider the cedar sawdust as thesubstrate with the lowest content of copper On the other handcopper adsorption by sawdust and coffee husk has beenreported obtaining a Langmuir based maximum adsorptioncapacity (Qmax mg g-1 adsorbent) of 845 and 75 respectively[4243] Although no adsorption studies has been reportedusing untreated soybean pod husk Šćiban et al [44] reporteda Qmax for copper adsorption of 0085 mmol g-1 adsorbent (~ 54mg g-1) using treated soybean straws Based on this
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 10 September 2013 | Volume 8 | Issue 9 | e73721
information it can be expected that a higher amount ofexogenous copper would be required for laccase induction oncultures with cedar sawdust than with soybean pod husk andcoffee husk due to its lower content of copper In addition dueto the high copper-adsorption capacity of sawdust the amountof available copper in the culture would be reduced allowingthe laccase production at higher copper concentrations withoutreaching levels that might be toxic for the fungus This fact canbe observed in the SSC (Table 4) However for the SmC thisbehavior is not maintained for the cultures with soybean podhusk and coffee husk (Table 4) where the maximum laccaseactivity was obtained with a copper concentration of 05 and 5mM respectively This could be owed to a lower contactbetween the fungus and the substrate in the SmC whencompared to the SSC which is close related to the transport ofnutrients presented in the substrates
Production of extracellular laccases has been reported in alarge number of species of white-rot fungi strains grown onnatural substrates (Table 1) Although a laccase activity profileis not clearly observed based on the agro-industrial wasteculture mode and fungus strain most cases in which SmC andSSC were conducted using the same substrate and fungusstrain showed a higher laccase activity under SmC conditionsIn this study was observed a substrate dependentperformance where the most favorable condition to obtain thehighest laccase activity was the SSC The laccase activitiesobtained in the present study are comparable and in somecases higher than those reported for Pleurotus ostreatusPhanerochaete chrysosporium Pycnoporus cinnabarinus Tversicolor and Ganoderma sp cultured under SC and SmCconditions and using different agro-industrial wastes (Table 1)Semi-solid cultures using different agro-industrial wastes havebeen reported using T versicolor Phlebia radiata Tpubescens P ostreatus and C unicolor Rodriguez et al [16]screened different supports for laccase production by Tversicolor under SSC They obtained the highest laccaseactivity (1200 U L-1) when in the culture medium was presentedbarley bran In addition they were able to obtain a laccaseactivity of 1700 U L-1 when the culture was supplemented withxylidine as an inducer These results were obtained after 17 or18 days of culture and ABTS was used to determine theenzyme activity Maumlkelauml et al [4546] have studied theexpression of the genes involved in laccase production Pr-lac1and Pr-lac2 presented in P radiata and evaluate laccaseproduction in SSC using milled alder wood (Alnus incana) assubstrate They reported a maximum laccase activity of 3 microkatL-1 (~180 U L-1) at the 14th day of culture and usingsyringaldazine for its detection Osma et al [15] have studiedthe morphological changes and laccase production of strains Tpubescens P ostreatus C unicolor and T versicolor grownon wheat bran flakes using SSC They obtained a maximumlaccase production between 1397 and 2778 U L-1 after 10 and13 days of culture This enzyme activity is strain dependentand ABTS was used for its quantification Our results show agood correlation with reported data The highest laccaseactivity was obtained when T pubescens was grown on coffeehusk under SSC without copper (204 U L-1 evaluated withsyringaldazine or 1549 U L-1 evaluated with ABTS) which is in
some cases higher than reported ones This shows that underthe studied conditions the production of the laccase crudeextract is favored in SSC using coffee husk without the additionof copper which could favors the cost of the productionprocess
It has been reported for white rot-fungi the production ofmultiple laccase isoenzymes which are encoded by differentgenes and show different expression profiles stabilitiessubstrate affinities and molecular masses (ranging from 35 to140 kDa) [3647ndash49] Two isoforms have been identified in Tpubescens strains two monomeric (Lac1 and Lac2) enzymesof 67 kDa but with different pI (53 and 51) [50] and twolaccase-activity fractions (Lap1 and Lap2) with different pI (gt30 and 26) from which Lap2 is a monomeric enzyme of 65kDa [17] In this study we identify two laccase isoenzymesfrom T pubescens CBS 69694 that have different molecularmasses (120 and 60 kDa) and pI The purifications resultssuggest a more acidic pI for Lac1 (le 4) than Lac2 (ge 6) whichdiffers from a previous report for this fungus in which twofractions with laccase activity were obtained Lap1 and Lap2with pI gt 3 and of 26 respectively [17] Since in both casessimilar purification protocols were used and Lac1 and Lac2were recovered from the unbound and bound fractionsrespectively as Lap1 and Lap2 [17] this difference could bedue to the culture conditions used in each case (ie thepresence of agro-industrial wastes) Nevertheless the nanoLC-MSMS analysis showed that peptides of Lac1 and Lac2 werepresented in Lap1 (UniProtKBSwiss-Prot Q8TG93) and Lap2(UniProtKBSwiss-Prot Q8TG94) which have a predicted pI of43 and 47 respectively Lac1 and Lac2 peptides werepresented in Laccase-2 (UniProtKBSwiss-Prot Q99046) andLaccase-1 (UniProtKBSwiss-Prot Q99044) from T villosarespectively with 80 to 100 identity These T villosa laccaseisoenzymes have a predicted pI of 47 (laccase-1) and 60(laccase-2) and a 77 identity among them In addition Tvillosa laccase-2 is reported as a homodimer protein whilelaccase-1 is reported as monomeric protein Alignment of Lac1-Lap2 and Lac2-Lap1 from T pubescens CBS 69694 and Tvillosa showed a 96 and 70 identity respectively Finallysimilar peptides were identified for Lac1 and Lac2 from Tpubescens (Schumach) Pilaacutet (Syn Coriolus pubescens(Schum ex Fr) Queacutel) BCB 923-2 (Figure 3b) [51] Theseresults suggest that the purified fractions correspond to laccaseisoenzymes that differ from previously reported ones for this Tpubescens strain
As mentioned above laccases isoenzymes present differentstabilities and substrate affinities This behavior was alsoobserved for the laccase isoenzymes purified from Tpubescens with Lac1 showing lower laccase activity thanLac2 with ABTS as substrate However the difference inactivity between these isoenzymes depended on the agro-industrial waste used for the T pubescens growth suggestinga probable effect of the substrate composition on the laccasephysicochemical properties In addition it was observed thatthe total laccase activity presented in the UF retentates ishigher than the laccase activity of the purified fractionsregardless of the agro-industrial byproduct used for the Tpubescens growth This suggests a synergistic effect of Lac1
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 11 September 2013 | Volume 8 | Issue 9 | e73721
and Lac2 as have been reported for laccases obtained fromTrametes and Pleurotus strains [175253]
The maximum relative activity for the UF retentate waspresented at a pH from 20 to 40 regardless of the agro-industrial byproduct with a monotonical decrease at pHs abovethis range The relation between pH and type of electron donormolecule explains this behavior on laccase activity It has beenreported that when the substrate is an organic hydrogen donorie phenols the optimal pH ranges from 35 to 60 while thelaccase activity monotonically decreases from pH 25 to 70 ifthe substrate is an electron donor like ABTS [54] Despite thatat different pHs most of the purified laccase fractionspresented a similar performance as the UF retentates the Lac2fraction obtained from cedar sawdust differed from the previousobservation For this fraction the maximum activity wasobserved at pH 50 and despite that at acidic pH (ge 30) therelative activity was higher than 80 at lower pHs (20) itdecreased almost in a 50 Nevertheless at basic pHs therelative laccase activity presented a sharp decreased that iswell correlate with the other laccase fractions We are not ableto attribute this particular trend to the presence of multiplelaccase isoenzymes in the fraction since the SDS-PAGErevealed only one band when stained with ABTS and from theNanoLC-MSMS analysis no difference in the number ofpeptides and their sequence was detected We speculate thatthis particular behavior can be owed to a compound added inthe pre-treatment of the cedar wood that might be used forpreserving it that might affect the activity of Lac2 and that wasnot completely removed during the purification
Temperature stability evaluation showed that Lac1 is themost thermolabile fraction while the laccase activity reductionin the crude extracts did not seem to be the result of anadditive effect of the activity loss of the purified fractionsAlthough there are no reports of laccase stability under theselected conditions these results agree with previous reportsthat state that fungal laccases rapidly decrease their activity attemperatures near 60 degC and thermal stability correlates withthe temperature range of the growth of the source organism[55] In addition we previously reported for a laccase crudeextract obtained from culturing T pubescens on coffee husk anactivity loss of 26 and 10 at 60 and 50 degC respectivelyafter 8 h at pH 60 using syringaldazine as substrate [48]
Finally an electrochemical characterization was carried outfor selected crude extracts and purified fractions The results
showed a difference between the electrochemicalperformances of extracts and purified fractions withsyringaldazine or ABTS These differences could be owed tothe different mechanisms for electron transfer as discussedabove However the overall performance of the obtained cyclicvoltammograms for ABTS and syringaldazine agrees withresults reported by Fernaacutendez-Saacutenchez et al [25] who used asaturated calomel reference electrode
Conclusions
We have shown the potential of coffee husk soybean podhusk and cedar sawdust as substrates for the laccaseproduction In several countries like Brazil and Colombia theproduction of coffee and soybean is of great importance andthe generated volume of residues is high The results showedthat semi-solid culture of T pubescens using coffee huskallows to obtain an important amount of laccase which couldbe favored by the phenolic-compounds ie tannins presentedin coffee husk While supplementation of the culture mediumwith copper favored the production of laccase in submergedand semi-solid culture using soybean pod husk and cedarsawdust semi-solid culture using coffee husk which presentedthe highest laccase volumetric yield did not require coppersupplementation This is an economic advantage that shouldbe considered when scaling up the process In this study Tpubescens showed its ability to produce two independentlaccases regardless of the used substrate However a higherlaccase activity was observed for crude extracts rather than forpurified fractions This suggests that Lac1 and Lac2 might havea synergistic effect on the final laccase activity rather than anadditive effect Based on this study the investigation of laccaseproduction by evaluating other agro-industrial wastes in whichphenolic-compounds constitute an important percentage oftheir composition is of great interest to avoid the use ofinducers that would increase the production cost
Author Contributions
Conceived and designed the experiments JFO CJAD OFSPerformed the experiments JCG SCM AR Analyzed the dataJCG SCM AR JFO CJAD OFS Wrote the manuscript AR JFOCJAD OFS
References
1 Vandamme EJ (2009) Agro-industrial residue utilization for industrialbiotechnology products In P NigamA Pandey Biotechnology for Agro-Industrial Residues Utilisation Netherlands Springer Verlag pp 3-11
2 Martiacutenez AT Speranza M Ruiz-Duentildeas FJ Ferreira P Camarero S etal (2005) Biodegradation of lignocellulosics microbial chemical andenzymatic aspects of the fungal attack of lignin Int Microbiol 8195-204 PubMed 16200498
3 Saacutenchez O Sierra R Almeacuteciga-Diacuteaz CJ (2011) Delignification processof agro-industrial aastes an alternative to obtain fermentablecarbohydrates for producing fuel In M Manzanera Alternative FuelIntech 111-154
4 Wesenberg D Kyriakides I Agathos SN (2003) White-rot fungi andtheir enzymes for the treatment of industrial dye effluents BiotechnolAdv 22 161-187 doi101016jbiotechadv200308011 PubMed14623049
5 Morozova OV Shumakovich GP Shleev SV Yaropolov YI (2007)Laccase-mediator systems and their applications A review ApplBiochem Microbiol 43 523-535 doi101134S0003683807050055
6 Baldrian P (2006) Fungal laccases - occurrence and properties FEMSMicrobiol Rev 30 215-242 doi101111j1574-4976200500010xPubMed 16472305
7 Rodriacuteguez Couto S Herrera JLT (2006) Industrial and biotechnologicalapplications of laccases A review Biotechnol Adv 24 500-513 doi101016jbiotechadv200604003 PubMed 16716556
8 Osma JF Toca-Herrera JL Rodriacuteguez-Couto S (2010) Uses oflaccases in the food industry Enzyme Res 2010 2010 918761PubMed 21048873
9 Rodriguez S (2008) Exploitation of biological wastes for the productionof value-added products under solid-state fermentation conditionsBiotechnol J 3 859-870 doi101002biot200800031 PubMed18543242
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 12 September 2013 | Volume 8 | Issue 9 | e73721
10 Osma JF Toca-Herrera JL Rodriacuteguez-Couto S (2011) Cost analysis inlaccase production J Environ Manage 92 2907-2912 doi101016jjenvman201106052 PubMed 21775046
11 Jain A Morlok CK Henson JM (2013) Comparison of solid-state andsubmerged-state fermentation for the bioprocessing of switchgrass toethanol and acetate by Clostridium phytofermentans Appl MicrobiolBiotechnol 97 905-917 doi101007s00253-012-4511-4 PubMed23111595
12 Singhania RR Patel AK Soccol CR Pandey A (2009) Recentadvances in solid-state fermentation Biochem Eng J 44 13-18 doi101016jbej200810019
13 Economou CN Makri A Aggelis G Pavlou S Vayenas DV (2010)Semi-solid state fermentation of sweet sorghum for the biotechnologicalproduction of single cell oil Bioresour Technol 101 1385-1388 doi101016jbiortech200909028 PubMed 19781936
14 Rodriacuteguez CS Santoro R Cameselle C Sanroman A (1997) Laccaseproduction in semi-solid cultures of Phanerochaete chrysosporiumBiotechnol Lett 19 995-998 doi101023A1018495216946
15 Osma JF Moilanen U Toca-Herrera JL Rodriacuteguez-Couto S (2011)Morphology and laccase production of white-rot fungi grown on wheatbran flakes under semi-solid-state fermentation conditions FEMSMicrobiol Lett 318 27-34 doi101111j1574-6968201102234xPubMed 21291496
16 Couto SR Gundin M Lorenzo M Sanroman MN (2002) Screening ofsupports and inducers for laccase production by Trametes versicolor insemi-solid-state conditions Proc Biochem 38 249-255 doi101016S0032-9592(02)00087-0
17 Galhaup C Goller S Peterbauer CK Strauss J Haltrich D (2002)Characterization of the major laccase isoenzyme from Trametespubescens and regulation of its synthesis by metal ions Microbiology148 2159-2169 PubMed 12101303
18 Sievers F Wilm A Dineen D Gibson TJ Karplus K et al (2011) Fastscalable generation of high-quality protein multiple sequencealignments using Clustal Omega Mol Syst Biol 7 539 PubMed21988835
19 Childs RE Bardsley WG (1975) The steady-state kinetics of peroxidasewith 22-azino-di-(3-ethyl-benzthiazoline-6-sulphonic acid) aschromogen Biochem J 145 93-103 PubMed 1191252
20 Harkin JM Obst JR (1973) Syringaldazine an Effective Reagent forDetecting Laccase and Peroxidase in Fungi Experientia 29 382-387
21 Nikupaavola ML Raaska L Itavaara M (1990) Detection of white-rotfungi by a non-roxic stain Mycol Res 94 27-31 doi101016S0953-7562(09)81260-4
22 Gonzalez LF Sarria V Sanchez OF (2010) Degradation ofchlorophenols by sequential biological-advanced oxidative processusing Trametes pubescens and TiO(2)UV Bioresour Technol 1013493-3499
23 Waterborg JH (2009) The Lowry method for protein quantitation In JWalker The Protein Protocols Handbook Humana Press pp 7-10
24 Harkin JM Obst JR (1973) Syringaldazine an effective reagent fordetecting laccase and peroxidase in fungi Cell Mol Life Sci 29381-387 doi101007BF01926734
25 Fernaacutendez-Saacutenchez C Tzanov T Guumlbitz GM Cavaco-Paulo A (2002)Voltammetric monitoring of laccase-catalysed mediated reactionsBioelectrochemistry 58 149-156 doi101016S1567-5394(02)00119-6PubMed 12414320
26 Bourbonnais R Leech D Paice MG (1998) Electrochemical analysis ofthe interactions of laccase mediators with lignin model compoundsBiochim Biophys Acta 1379 381ndash390 doi101016S0304-4165(97)00117-7 PubMed 9545600
27 Ferraz F Silva S (2009) Characterization of coffee husk biomass forbiotechnological purposes N Biotechnol 25 S256-S256 doi101016jnbt200906573
28 Pandey A Soccol CR Nigam P Brand D Mohan R et al (2000)Biotechnological potential of coffee pulp and coffee husk forbioprocesses Biochem Eng J 6 153-162 doi101016S1369-703X(00)00084-X PubMed 10959086
29 Sruamsiri S Silman P (2008) Nutritive composition of soybean by-products and nutrient digestibility of soybean pod husk Maejo Int J SciTechnol 2 568-576
30 Romkaew J Nagaya Y Goto M Suzuki J Umezaki T (2008) Poddehiscence in relation to chemical components of pod shell in soybeanPlant Prod Sci 11 278-282 doi101626pps11278
31 Nakamanee G Polbumroong T Phaikaew C (2003) Alternative use ofcrop waste to increase milk production Proceedings of the 8th Meetingof the Regional Working Group on Grazing and Feed Resources forSoutheast Asia Kuala Lumpur Malaysia 66 - 70
32 Kurek B Gaudard F (2000) Oxidation of spruce wood sawdust byMnO(2) plus oxalate a biochemical investigation J Agric Food Chem48 3058-3062 doi101021jf000015g PubMed 10898665
33 Weil J Brewer M Hendrickson R Sarikaya A Ladisc MR (1998)Continuous pH monitoring during pretreatment of yellow poplar woodsawdust by pressure cooking in water Appl Biochem Biotechnol 70-7299-111 doi101007BF02920127
34 Weil J Sarikaya A Rau S-L Goetz J Ladisch CM et al (1997)Pretreatment of yellow poplar sawdust by pressure cooking in waterAppl Biochem Biotechnol 68 21-40 doi101007BF02785978
35 Dekker RF Barbosa AM Giese EC Godoy SD Covizzi LG (2007)Influence of nutrients on enhancing laccase production byBotryosphaeria rhodina MAMB-05 Int Microbiol 10 177-185 PubMed18075999
36 Xiao YZ Hong YZ Li JF Hang J Tong PG et al (2006) Cloning ofnovel laccase isozyme genes from Trametes sp AH28-2 and analysesof their differential expression Appl Microbiol Biotechnol 71 493-501doi101007s00253-005-0188-2 PubMed 16283298
37 Crowe JD Olsson S (2001) Induction of laccase activity in Rhizoctoniasolani by antagonistic Pseudomonas fluorescens strains and a range ofchemical treatments Appl Environ Microbiol 67 2088-2094 doi101128AEM6752088-20942001 PubMed 11319086
38 Yaguumle S Terroacuten MC Gonzaacutelez T Zapico E Bocchini P et al (2000)Biotreatment of tannin-rich beer-factory wastewater with white-rotbasidiomycete Coriolopsis gallica monitored by pyrolysisgaschromatographymass spectrometry Rapid Commun Mass Spectrom14 905-910 doi101002(SICI)1097-0231(20000530)1410 PubMed10825255
39 Gowda NKS Ramana JV Prasad CS Singh K (2004) Micronutrientcontent of certain tropical conventional and unconventional feedresources of southern India Trop Anim Heatlth Pro 36 77-94 doi101023BTROP0000009522309491d PubMed 14979561
40 DePeters EJ Fadel JG Arana MJ Ohanesian N Etchebarne MA et al(2000) Variability in the chemical composition of seventeen selectedby-product feedstuffs used by the California dairy industry Prof AnimSci 16 69-99
41 Obernberger I Thek G (2004) Physical characterisation and chemicalcomposition of densified biomass fuels with regard to their combustionbehaviour Biomass Bioenerg 27 653-669 doi101016jbiombioe200307006
42 Larous S Meniai A-H Lehocine MB (2005) Experimental study of theremoval of copper from aqueous solutions by adsorption usingsawdust Desalination 185 483ndash490 doi101016jdesal200503090
43 Oliveira WE Franca AS Oliveira LS Rocha SD (2008) Untreatedcoffee husks as biosorbents for the removal of heavy metals fromaqueous solutions J Hazard Mater 152 1073ndash1081 doi101016jjhazmat200707085 PubMed 17804159
44 Šciban M Klašnja M Škrbic B (2008) Adsorption of copper ions fromwater by modified agricultural by-products Desalination 229 170ndash180doi101016jdesal200708017
45 Maumlkelauml MR Hildeacuten KS Hakala TK Hatakka A Lundell TK (2006)Expression and molecular properties of a new laccase of the white rotfungus Phlebia radiata grown on wood Curr Genet 50 323-333 doi101007s00294-006-0090-1 PubMed 16927090
46 Maumlkelauml MR Lundell T Hatakka A Hildeacuten K (2013) Effect of coppernutrient nitrogen and wood-supplement on the production of lignin-modifying enzymes by the white-rot fungus Phlebia radiata Fungal Biol117 62-70 doi101016jfunbio201211006 PubMed 23332834
47 Dwivedi UN Singha P Pandeya VP Kumara A (2011) Structurendashfunction relationship among bacterial fungal and plant laccases J MolCatal B Enzym 68 117-128 doi101016jmolcatb201011002
48 Gaitan IJ Medina SC Gonzaacutelez JC Rodriacuteguez A Espejo AacuteJ et al(2011) Evaluation of toxicity and degradation of a chlorophenol mixtureby the laccase produced by Trametes pubescens Bioresour Technol102 3632ndash3635 doi101016jbiortech201011040 PubMed21115244
49 Karp SG Faraco V Amore A Birolo L Giangrande C et al (2012)Characterization of laccase isoforms produced by Pleurotus ostreatusin solid state fermentation of sugarcane bagasse Bioresour Technol114 735-739 doi101016jbiortech201203058 PubMed 22487128
50 Nikitina OV Shleev SV Gorshina ES Rusinova TV Serezhenkov VAet al (2005) Isolation and purification of enzymes from ligninolyticcomplex of the basidial fungus Trametes pubescens (Schumach) Pilatand study of their properties Biochemistry (Mosc) 70 1274-1279 doi101007s10541-005-0259-0
51 Shleev S Nikitina O Christenson A Reimann CT Yaropolov AI et al(2007) Characterization of two new multiforms of Trametes pubescenslaccase Bioorg Chem 35 35-49 doi101016jbioorg200608001PubMed 16989887
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 13 September 2013 | Volume 8 | Issue 9 | e73721
52 Xiao YZ Chen Q Hang J Shi YY Wu J et al (2004) Selectiveinduction purification and characterization of a laccase isozyme fromthe basidiomycete Trametes sp AH28-2 Mycologia 96 26-35 doi1023073761984 PubMed 21148825
53 Muntildeoz C Guilleacuten F Martiacutenez AT Martiacutenez MJ (1997) Laccaseisoenzymes of Pleurotus eryngii characterization catalytic propertiesand participation in activation of molecular oxygen and Mn2+ oxidationAppl Environ Microbiol 63 2166-2174 PubMed 9172335
54 Morozova OV Shumakovich GP Gorbacheva MA Shleev SVYaropolov AI (2007) Blue laccases Biochemistry (Mosc) 721136-1150 doi101134S0006297907100112
55 Hildeacuten K Hakala TK Lundell T (2009) Thermotolerant andthermostable laccases Biotechnol Lett 31 1117-1128 doi101007s10529-009-9998-0 PubMed 19360388
56 Verma P Madamwar D (2002) Production of ligninolytic enzymes fordye decolorization by cocultivation of white-rot fungi Pleurotusostreatus and Phanerochaete chrysosporium under solid-statefermentation Appl Biochem Biotechnol 102-103 109-118 doi101385ABAB102-1031-6109 PubMed 12396115
57 Meza JC Lomascolo A Casalot L Sigoillot J-C Auria R (2005)Laccase production by Pycnoporus cinnabarinus grown on sugar-canebagasse Influence of ethanol vapours as inducer Proc Biochem 403365ndash3371 doi101016jprocbio200503004
58 Hossain SM Anantharaman N (2006) Activity enhancement ofligninolytic enzymes of Trametes versicolor with bagasse powder Afr JBiotechnol 5 189-194
59 Revankar MS Desai KM Lele SS (2007) Solid-state fermentation forenhanced production of laccase using indigenously isolatedGanoderma sp Appl Biochem Biotechnol 143 16-26 doi101007s12010-007-0029-0 PubMed 18025593
60 Velazquez-Cedeno MA Mata G Savoie J-M (2002) Waste-reducingcultivation of Pleurotus ostreatus and Pleurotus pulmonarius on coffeepulp changes in the production of some lignocellulolytic enzymesWorld J Microbiol Biotechnol 18 201ndash207 doi101023A1014999616381
61 Mata G Murrieta D Andreu LI (2005) Changes in lignocellulolyticenzyme activities in six Pleurotus spp strains cultivated on coffee pulpin confrontation with Trichoderma spp World J Microbiol Biotechnol 21143ndash150 doi101007s11274-004-3041-3
62 de Souza DF Tychanowicz GK de Souza CG Peralta RM (2006) Co-production of ligninolytic enzymes by Pleurotus pulmonarius on wheatbran solid state cultures J Basic Microbiol 46 126-134 doi101002jobm200510014 PubMed 16598826
63 Prasad KK Mohan SV Bhaskar YV Ramanaiah SV Babu VL et al(2005) Laccase production using Pleurotus ostreatus 1804 immobilizedon PUF cubes in batch and packed bed reactors influence of cultureconditions J Microbiol 43 301-307 PubMed 15995650
64 Songulashvili G Elisashvili V Wasser SP Nevo E Hadar Y (2007)Basidiomycetes laccase and manganese peroxidase activity in
submerged fermentation of food industry wastes Enzyme MicrobTechnol 41 57ndash61 doi101016jenzmictec200611024
65 Sathishkumar P Murugesan K Palvannan T (2010) Production oflaccase from Pleurotus florida using agro-wastes and efficientdecolorization of Reactive blue 198 J Basic Microbiol 50 360-367 doi101002jobm200900407 PubMed 20586068
66 Moldes D Gallego PP Rodriacuteguez Couto S Sanromaacuten A (2003) Grapeseeds the best lignocellulosic waste to produce laccase by solid statecultures of Trametes hirsuta Biotechnol Lett 25 491-495 doi101023A1022660230653 PubMed 12882277
67 Elisashvili V Kachlishvili E Penninckx M (2008) Effect of growthsubstrate method of fermentation and nitrogen source onlignocellulose-degrading enzymes production by white-rotbasidiomycetes J Ind Microbiol Biotechnol 35 1531-1538 doi101007s10295-008-0454-2 PubMed 18716810
68 Osma JF Saravia V Herrera JL Couto SR (2007) Mandarin peelingsthe best carbon source to produce laccase by static cultures ofTrametes pubescens Chemosphere 67 1677-1680 doi101016jchemosphere200611051 PubMed 17234250
69 Kahraman SS Gurdal IH (2002) Effect of synthetic and natural culturemedia on laccase production by white rot fungi Bioresour Technol 82215-217 doi101016S0960-8524(01)00193-6 PubMed 11991068
70 Iandolo D Piscitelli A Sannia G Faraco V (2011) Enzyme Productionby Solid Substrate Fermentation of Pleurotus ostreatus and Trametesversicolor on Tomato Pomace Appl Biochem Biotechnol 163 40-51doi101007s12010-010-9014-0 PubMed 20582639
71 Moldes D Lorenzo M Sanromaacuten MA (2004) Different proportions oflaccase isoenzymes produced by submerged cultures of Trametesversicolor grown on lignocellulosic wastes Biotechnol Lett 26 327-330doi101023BBILE000001545240213bf PubMed 15055770
72 Philippoussis A Diamantopoulou P Papadopoulou K Lakhtar HRoussos S et al (2010) Biomass laccase and endoglucanaseproduction by Lentinula edodes during solid state fermentation of reedgrass bean stalks and wheat straw residues World J MicrobiolBiotechnol 27 285-297
73 Fenice M Giovannozzi Sermanni G Federici F DrsquoAnnibale A (2003)Submerged and solid-state production of laccase and Mn-peroxidaseby Panus tigrinus on olive mill wastewater-based media J Biotechnol100 77-85 doi101016S0168-1656(02)00241-9 PubMed 12413788
74 Rodriguez-Couto S GundIn M Lorenzo M Sanromaacuten MAacute (2002)Screening of supports and inducers for laccase production by Trametesversicolor in semi-solid-state conditions Proc Biochem 38 249-255doi101016S0032-9592(02)00087-0
75 Fackler K Gradinger C Schmutzer M Tavzes C Burgert I et al (2007)Biotechnological wood modification with selective white-rot fungi and itsmolecular mechanisms Food Technol Biotechnol 45 269ndash276
76 Levin L Herrmann C Papinutti VL (2008) Optimization oflignocellulolytic enzyme production by the white-rot fungus Trametestrogii in solid-state fermentation using response surface methodologyBiochem Eng J 39 207-214 doi101016jbej200709004
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 14 September 2013 | Volume 8 | Issue 9 | e73721

between 147- and 663-fold higher than that observed forcultures with soybean pod husk and cedar sawdust Laccaseactivity obtained in the control culture without agro-industrialwaste presented a maximum laccase activity of 222 plusmn 088 UL-1 after 14 days of culture while the glucose was depleted onthe seventh day of culture
Copper supplementation effect on laccase activity wasevaluated for both SmC and SSC Although in a non-linearmode the laccase activity was increased by the presence ofcopper with all substrates under SmC or SSC except withcoffee husks under SSC (Table 2) In addition the requiredcopper concentration to obtain the highest laccase activityvaried depending on the used agro-industrial waste Forexample laccase production was favored in cultures withsoybean pod husk at copper concentrations of 05 mM and 20mM for SmC and SSC respectively while for cultures with
Figure 1 Laccase activity profiles during T pubescensCBS 69694 culture under SmC (a) and SSC (b) on coffeehusk Culture media was enriched with different copperconcentrations 0 (Δ) 05 () 2 () and 5 (loz) mM and laccaseactivity was determined by using syringaldazine as substrateFilled symbols refer to the glucose concentration at therespective copper concentration All the assays wereperformed in triplicatedoi 101371journalpone0073721g001
coffee husk the highest laccase activity levels for SmC andSSC was observed at copper concentrations of 5 mM and 0mM respectively
Despite that culture mode ie SmC or SSC presented aneffect on laccase production this is substrate sensitive In theassays conducted without copper it was observed that laccaseactivity was 28- and 33-fold higher in SSC than in SmC for thesoybean pod husk and coffee husk respectively Howeverwhen cedar sawdust was used no significant difference inlaccase activity between the two culture types was obtained(Table 2) Due to the low volume of liquid media and highlaccase activity in SSC when compared to SmC the SSCpresented a higher volumetric yield For all agro-industrialwaste the SSC presented a shorter time than SmC to reachthe highest laccase activity which suggest a higher productivityunder SSC conditions
These results show that coffee husk under SmC or SSCconditions could be considered as a potential alternative forlaccase production by T pubescens However soybean podhusk and cedar sawdust could be also considered as importantalternatives for laccase production especially in comparisonwith synthetic culture media [10] Finally the optimal copperconcentration to obtain the highest laccase activity depends onthe agro-industrial byproduct used for the fungus culture
Laccase purification and characterizationLaccase purification was conducted using the crude extracts
that presented the highest activities For the SmC crude
Table 2 Effect of substrate copper concentration andculture mode on laccase activity (UL-1)
Substrate SmC SSCSoybean Pod Husk + 0 mM Cu+2 446 plusmn 138 (23) 1255 plusmn 052 (13)
Soybean Pod Husk + 05 mM Cu+2
8493 plusmn 846 (23)dagger 2028 plusmn 387 (21)
Soybean Pod Husk + 2 mM Cu+2 5761 plusmn 370 (19) 13785 plusmn 071 (18)dagger
Soybean Pod Husk + 5 mM Cu+2 4492 plusmn 487 (23) 2900 plusmn 407 (18)Coffee Husk + 0 mM Cu+2 6184 plusmn 176 (23) 20400 plusmn 596 (19)dagger
Coffee Husk + 05 mM Cu+2 13763 plusmn 568 (23) 11083 plusmn 940 (20)ab
Coffee Husk + 2 mM Cu+2 12304 plusmn 373 (23)b 10278 plusmn 814 (15)b
Coffee Husk + 5 mM Cu+2 17723 plusmn 372 (23)dagger 8266 plusmn 882 (15)Cedar Sawdust + 0 mM Cu+2 1355 plusmn 095 (17) 1082 plusmn 053 (15)
Cedar Sawdust + 05 mM Cu+2 2272 plusmn 084 (17) 1902 plusmn 002 (13)Cedar Sawdust + 2 mM Cu+2 2897 plusmn 127 (12)a 2638 plusmn 068 (8)Cedar Sawdust + 5 mM Cu+2 2936 plusmn 153 (13) abdagger 3073 plusmn 152 (12)bdagger
Data in parenthesis refers to the day where was observed the maximum laccaseactivity No Statistical Difference Statistical Difference (Evaluation of substrate effecton laccase activity for both culture modes submerged (SmC) and semisolid (SSC)no copper induction considered)Equal letters inside one agro-industrial waste group refers to No StatisticalDifference no letters refers to Statistical Difference inside the group (Evaluation ofcopper effect on laccase activity for both SmC and SSC)dagger Maximum laccase activity for the used agro-industrial wastes either in SmC orSSC culture
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 5 September 2013 | Volume 8 | Issue 9 | e73721
extracts obtained with soybean pod husk + Cu+2 05 mM coffeehusk + Cu+2 5 mM and cedar sawdust + Cu+2 5 mM wereselected while for SSC crude extracts obtained with soybeanpod husk + Cu+2 2 mM coffee husk + Cu+2 0 mM and cedarsawdust + Cu+2 5 mM were selected The electrophoreticanalysis of permeates from 045 and 022 microm filtrations and theUF retentate showed the presence of two proteins with laccaseactivity while no activity was observed within the ultrafiltratedsample (Figure 2a) These proteins presented a molecularmass of about 120 kDa and 60 kDa (Figure 2a) Thepurification of the UF retentate by ionic exchangechromatography confirmed the presence of two differentfractions with laccase activity (Figure 2b) The SDS-PAGEanalysis of these fractions under non-reducing conditionsshowed that one protein with laccase activity was presented oneach fraction (Figure 2b) and were named Lac1 (120 kDa) andLac2 (60 kDa) Considering that the conditions used tomeasure the enzyme activity were those suggested for laccaseactivity and that H2O2 or manganese were not added duringthe reaction for measuring the enzyme activity these resultssuggest that both proteins are laccases but not peroxidases
NanoLC-MSMS analysis of purified fractions showed thatthe identified peptides for Lac1 (SAGSTTYNYNDPIFR andRDTVSTGNSGDNVTIR) and Lac2 (SAGSTVYNYDNPIFR andANPNFGNVGFTGGINSAI) had identity scores between 80 to100 with laccases (over 100 sequences) available at the non-redundant GenBank dataset Proteins containing thesepeptides different that laccases were not identified Similarresults were observed when peptides were evaluated atUniprot Galhaup et al [17] described two laccase activefractions from T pubescens MB 89 (CBS 69694) Lap1(UniProtKBTrEMBL Q8TG93) and Lap2 (UniProtKBTrEMBLQ8TG94) which share a 70 identity (Figure 3a) Sequencealignment of Lac1 and Lac2 peptides with Lap1 and Lap2sequences respectively showed that the peptides arepresented within T pubescens MB 89 laccase sequencesthough some differences were observed for the Lac1 peptide(Figure 3b) This result suggests that the purified fractions arelaccase isoenzymes
Protein concentration and volumetric and specific laccaseactivities at different purification stages are presented in Table3 Lac1 presented lower laccase activity than Lac2 in a ratiothat ranges from 080- to 38-fold depending on the substratesource On the other hand laccase activity of the UF retentatefrom soybean pod husk culture was from 45- to 64-fold higherthan that of the purified fractions while for cedar sawdust andcoffee husk the laccase activity was from 46- to 120-fold andfrom 33- to 163-fold higher than the purified fractionsrespectively (Table 3) The purified Lac2 fractions from coffeehusk and cedar sawdust cultures showed the highest specificactivity levels (Table 3) The volumetric activities of UFretentates were higher than that obtained with purified fractions(Table 4)
Data in parenthesis shows the temperature or pH at whichrelative laccase activity reached 100
Temperature and pH effect on laccase activity wasdetermined for the UF retentate and Lac1 and Lac2 fractionsobtained along the purification of the enzymatic crude extracts
that resulted after the SSC of T pubescens on the selectedagro-industrial wastes Laccase activity for the UF retentateunder different temperature and pH conditions is presented inFigure 4 and Table 4 The maximum relative activity of theevaluated UF retentates was observed at 50-55 degC and at apH range from 20 to 40 However UF retentate from cedar
Figure 2 Purification of lacasses produced by submergedculture of T pubescens using coffee husk assubstrate All samples were analyzed by SDS-PAGE undernon-reducing conditions and stained with ABTS No heatdenaturation of the samples was conducted (a) The crudeextract (1) was centrifuged and filtered through a Whatman No1 filter (2) 045 microM membrane (3) 022 microm membrane (4) andultrafiltrated through a 10 kDa cut-off membrane permeate (5)and retentate (6) (b) UF retentate was purified by anionicexchange chromatography Lac1 represent the unboundfraction while Lac2 represent the eluted fraction The arrowshows the point when protein elution was started (c) Purifiedfractions (1) crude extract (2) Lac1 and (3) Lac2 MWMolecular Weight Similar results were observed for crudeextracts produced by using soybean pod husk and cedarsawdustdoi 101371journalpone0073721g002
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 6 September 2013 | Volume 8 | Issue 9 | e73721
sawdust culture behaved different to the UF retentate fromcoffee husk and soybean pod husk cultures In this sense the
UF retentate from cedar sawdust culture presented the lowestenzyme activity at temperatures le 40 degC but at 70 degC
Figure 3 Aligment of reported lacasses sequences and Lac1 and Lac2 peptides (a) Alignment of Lap1 (UniProtKBSwiss-Prot Q8TG93) and Lap2 (UniProtKBSwiss-Prot Q8TG94) from T pubescens CBS 69694 (b) Alignment of Lac1 and Lac2 peptideswith Lap1 and Lap2 respectively and tryptic peptides described by Shleev et al 2007 Alignments were carried out using ClustalOmega and figures were using CLC Sequence Viewer Different residues are colored in reddoi 101371journalpone0073721g003
Table 3 Laccase activity during the purification of crude extracts resulted from T pubescens culture on agro-industrialwastes under submerged (SmC) and semisolid (SSC) cultures
Soybean pod husk Coffee husk Cedar sawdust
Protein(mg mL-1)
VolumetricActivity (UL-1)
SpecificActivity (Umg-1
protein)Recovery()
Protein(mg mL-1)
VolumetricActivity (UL-1)
SpecificActivity (Umg-1
protein)Recovery()
Protein(mg mL-1)
VolumetricActivity (UL-1)
SpecificActivity (Umg-1
protein)Recover()
SmCCrudeExtract
0869 52888 0608 100 0414 152708 3688 100 0431 10735 0249 100
045 micromFiltration
0736 46549 0632 88 0334 117892 3529 77 0444 12053 0271 112
UF retentate 1010 78731 0779 74 0378 195263 5165 64 0510 25905 0507 120
Lac1 0494 5471 0110 1 0232 2399 0103 02 0222 838 0037 08
Lac2 0310 20661 0666 4 0116 107240 9252 7 0040 15234 3837 14 SSCCrudeExtract
1146 164966 1439 100 0596 154907 2601 100 1065 23203 0217 100
045 micromFiltration
0776 128392 1654 77 0474 115736 2439 75 0806 25824 0320 111
UF retentate 0973 335055 3443 101 0428 161750 3779 52 1134 51101 0450 110
Lac1 0097 8854 0912 2 0104 1514 0145 03 0042 1315 0313 2
Lac2 0168 48988 2915 10 0131 82298 6282 18 0072 16162 2244 23
740 ABTS was used to determine laccase activity
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 7 September 2013 | Volume 8 | Issue 9 | e73721
presented the highest enzyme activity ~90 of its initial valuein comparison to the other two UF retentates (Figure 4)
The temperature and pH effect on laccase activity for Lac1and Lac2 is presented in Figure 5 The maximum relativeactivity for both fractions was presented at a pH from 20 to 40regardless of the agro-industrial waste as observed for the UFretentate However at pH gt 40 Lac1 showed a sharperdecrease in its activity than that observed for Lac2Nevertheless Lac2 purified from cedar sawdust culturepresented a different performance This fraction showed themaximum activity at pH 50 while a sharp drop was observedat pHs above or below this value
Laccase activity profiles showed that Lac1 and Lac2presented the maximum enzyme activity at 55 and 60 degCrespectively (Table 4) It was observed that Lac1 from coffeehusk culture presented a lower decrease in enzyme activity attemperatures le 50 degC or ge 60 degC than Lac2 On the contraryLac2 from soybean husk or cedar sawdust cultures presenteda higher relative laccase activity than Lac1 at temperatures le55 degC The purified fractions from soybean pod husk and cedarsawdust cultures presented the lowest relative activity at 30 degCand 65 degC
Table 4 Optimum temperatures and pHs for maximumlaccase activities in UF retentate Lac1 and Lac2 obtainedfrom semisolid cultured T pubescens on agro-industrialwastes
EnzymaticFraction Temperature pH
VolumetricActivity (UL-1)
SpecificActivity (Umg-1 protein)
VolumetricActivity (UL-1)
SpecificActivity (Umg-1
protein)SoybeanPod Husk +2 mM Cu+2
148000plusmn1178(55)
1521plusmn0012182037plusmn3025(2-3)
1871plusmn0031
Lac122925plusmn917(60)
2363plusmn009536135plusmn831(23)
3725plusmn0085
Lac232638plusmn1558(55)
1942plusmn009228713plusmn274(23)
1709plusmn0016
CoffeeHusk + 0mM Cu+2
113740plusmn8289(50-55)
3008plusmn0219290666plusmn1039(2-3)
7689plusmn0275
Lac1 889plusmn039 (55) 0038plusmn0002 1779plusmn028 (23) 0077plusmn0001
Lac234194plusmn2789(60)
2947plusmn024033150plusmn526 (2ndash4)
2857plusmn0045
CedarSawdust +5 mM Cu+2
257666plusmn1201(55)
5052plusmn0024344634plusmn1043(2-3)
6757plusmn0204
Lac121370plusmn472(55)
0962plusmn002129117plusmn943(23)
1311plusmn0042
Lac255166plusmn1656(60)
1379plusmn0414 55166plusmn1656 (5) 1379plusmn0414
ABTS was used to determine laccase activity
Laccase stabilityFor selected enzymatic extracts and purified fractions (Table
4) a stability test for 48 h at their optimal pH and temperaturewas conducted Time course of the relative laccase activity forcrude extract and purified fractions from coffee husk and cedarsawdust cultures are presented in Figure 5 Similar resultswere observed for crude extract and purified fractions fromsoybean pod husk culture (data no shown)
It was observed that the activity loss depends on thesubstrate source used to grow T pubescens An activity loss of55 was observed for crude extracts and purified fractionsfrom coffee husk and soybean pod husk cultures while anactivity loss of 70 was observed for the crude extract andpurified fractions from cedar sawdust culture NeverthelessLac2 showed the lowest activity reduction with values of 10 40and 55 for coffee husk soybean pod husk and cedarsawdust respectively while Lac1 presented an activity loss of34 50 and 60 for coffee husk soybean pod husk and cedarsawdust respectively Despite the differences in laccaseactivity among agro-industrial wastes and between crudeextracts and purified fractions all the profiles showed a sharp
Figure 4 Effect of temperature (a) and pH (b) on laccaseactivity for the UF retentate obtained by semisolid cultureof T pubescens on coffee husk () soybean pod husk (loz)and cedar sawdust (Δ) Laccase stability was evaluated byusing ABTS All the assays were performed in triplicatedoi 101371journalpone0073721g004
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 8 September 2013 | Volume 8 | Issue 9 | e73721
decrease during the first 8 h while a reduced activity loss wasobserved during the last 24 h of the assay (Figure 6)
Laccase electrochemical characterizationFor selected crude extracts and purified fractions a cyclic
voltammetry characterization was conducted For eachmediator syringaldazine or ABTS similar cyclic
Figure 5 Effect of temperature () and pH () on enzymeactivity for Lac1 (solid line) and Lac2 (dashed line)obtained after the purification of the enzymatic crudeextract produced by semisolid culture of T pubescens oncoffee husk (a) soybean pod husk (b) and cedar sawdust(c) Laccase stability was evaluated by using ABTS All theassays were performed in triplicatedoi 101371journalpone0073721g005
voltammograms were obtained for the different crude extractsand purified fractions Voltammograms of the redox reaction forsyringaldazine (Figure 7a) and ABTS (Figure 6b) arepresented Two anionic and cationic peaks were observedwhen syringaldazine was used as mediator The first pair ofpeaks showed a reversible performance with a mean Ia Icratio of 109plusmn006 106plusmn005 and 098plusmn002 for the crudeextracts Lac1 and Lac2 respectively The second pair ofpeaks showed quasi-reversible performance and the cathodicpeak was presented at a potential of +062 V (vs Ag|AgCl)while the anodic peak was presented at a potential of +060 V(vs Ag|AgCl) No significant difference on the reductionpotential (Eo) for syringaldazine was observed for the crudeextracts or the purified fractions obtaining an Eo of0314plusmn0008 V (vs Ag|AgCl) However the peak potentialseparation (ΔEp) showed a different profile depending on thesubstrate used for the culture While not significant difference(p gt 005) on ΔEp value was observed between crude extract(0045plusmn0005 V vs Ag|AgCl) Lac1 (0046plusmn0005 V vs Ag|AgCl)and Lac2 (0028plusmn0001 V vs Ag|AgCl) from cultures with cedarsawdust and soybean pod husk the ΔEp was considerablyhigher for the crude extract (0083plusmn0004 V vs Ag|AgCl) Lac1(0068plusmn0003 V vs Ag|AgCl) and Lac2 (0051plusmn0005 V vs Ag|AgCl) from the coffee husk culture
For reactions conducted with ABTS and the different crudeextracts and purified fractions the voltammograms exhibitedone pair of cathodic and anodic peaks (Figure 7b) whichcorresponds to the redox of ABTSABTS+ (IaIc) with an Eo of0487plusmn0006 V (vs Ag|AgCl) This process was reversiblepresenting a mean Ia Ic ratio of 099plusmn002 In addition asecond cathodic peak was presented at +083 V (vs Ag|AgCl)when ABTS redox reaction was conducted using the crudeextract and the purified fractions This performance agrees withprevious reports in which the laccase activity cannot beobserved at potential values of the second reversible oxidation
Figure 6 Laccase stability profiles at optimal pH andtemperature conditions for crude extracts (solid line) Lac1(dashed line) and Lac2 (spotted line) obtained fromsemisolid culture of T pubescens on coffee husk (loz) andcedar sawdust (bull) Laccase stability was evaluated by usingABTS All the assays were performed in triplicatedoi 101371journalpone0073721g006
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 9 September 2013 | Volume 8 | Issue 9 | e73721
of ABTS ABTS+ABTS2+ due to this catalytic process cannotbe detected by cyclic voltammetry [2526] Neverthelessvoltammograms for laccase crude extract from a soybean podhusk culture presented the second anodic peak which inaddition to the second cathodic peak corresponds to the redoxof ABTS+ABTS2+ (IIaIIc) with an Eo of 0942plusmn0004 V (vs Ag|AgCl) Contrary to the reactions conducted usingsyringaldazine as substrate the ΔEp did not present asignificant difference for the crude extracts and purifiedfractions 0248plusmn0016 V (vs Ag|AgCl)
Discussion
In this study we evaluated the effect of the culture mode ieSmC and SSC on laccase production by T pubescens CBS69694 using three different agro-industrial wastes (coffeehusk soybean pod husk and cedar sawdust) Furthermore theeffect of Cu+2 supplementation on laccase production was alsoevaluated and the crude extracts and the purified fractionswere partially characterized
Figure 7 Cyclic voltammograms of (a) syringaldazine atpH 50 using the crude extract and purified laccasefractions from semisolid culture of T pubescens on coffeehusk or (b) ABTS at pH 30 using the crude extract andpurified laccase fractions from semisolid culture of Tpubescens on soybean pod husk Nomenclature onlymediator (dotted line) crude extract (solid line) Lac1 fraction(dashed line) and Lac2 fraction (dashed and dotted line)doi 101371journalpone0073721g007
Substrate source for laccase production showed to be one ofthe most important factors for laccase production (Table 2)Cultures with coffee husk as substrate presented the highestlaccase production levels The observed differences in laccaseactivity for each substrate could be owed to their chemicalcomposition Coffea arabica is the coffee variety raised inColombia and its husk has a reported composition of 2308cellulose (2076 glucose and 183 cellobiose) 2385hemicellulose (1356 xylose 523 arabinose 256 aceticacid and 195 glucuronic acid) 2828 total lignin and071 ashes [27] In addition coffee husk contents 13caffeine 45 tannins and 124 pectins [28] On the otherhand Glycine max is the soybean species cultured inColombia and its pod husk composition varies from 504 to62 crude protein 608 to 143 hemicellulose 3715 to4208 acid detergent fiber 4436 to 6015 neutral detergentfiber 850 to 893 acid detergent lignin and 2074 to 2106uronic acid [29ndash31] Finally for cedar sawdust the majorcomponents are glucose (426-444) xylose (46-195)cellulose (383-410) klason lignin (227-278) and acidlignin (40-43) [32ndash34] Although lignin is one of the mostimportant compounds associated with laccase production bywhite-rot fungi we did not observe a clear correlation betweenlignin composition percentage and laccase activity In thatsense the high laccase activity observed for the cultures thatuse coffee husk could be associated with its content of caffeineand tannins which have been identified as potent inducers oflaccase gene expression [173536] In fact caffeine has beenused as inducer for laccase production by Rhizoctonia solaniobtaining an increase of 41-fold in laccase activity [37] Similarresults were also reported for the production of Coloriopsisgallica laccase [38]
Laccase gene expression is mainly up regulated by Cu2+although Mn2+ Fe3+ heavy metals 26-dimethoxy-14-benzoquinone H2O2 amphotericin B syringic acid tannic acidTween 80 soybean oil aromatic compounds andmicroclimatic changes (ie lower temperature and osmoticpressure) are also recognized as potent inducers [173536] Ingeneral Cu2+ supplementation increased the laccase activity inboth SmC and SSC However Cu2+concentration required toobtain the highest laccase activity varied depending on agro-industrial waste These differences can be attributed to thecopper presented in the agro-industrial waste and the copperavailability in the medium Copper can be found in amountsthat range from 156 to 180 mg kg-1 of coffee husk [39] andfrom 26 to 67 mg kg-1 of soybean pod husk (or soybean hull)[3940] Despite that no reports indicate the copper content inuntreated cedar wood an average of 10 mg kg-1 of wood hasbeen reported for wood used as biomass in six different Europecountries [41] This allows to consider the cedar sawdust as thesubstrate with the lowest content of copper On the other handcopper adsorption by sawdust and coffee husk has beenreported obtaining a Langmuir based maximum adsorptioncapacity (Qmax mg g-1 adsorbent) of 845 and 75 respectively[4243] Although no adsorption studies has been reportedusing untreated soybean pod husk Šćiban et al [44] reporteda Qmax for copper adsorption of 0085 mmol g-1 adsorbent (~ 54mg g-1) using treated soybean straws Based on this
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 10 September 2013 | Volume 8 | Issue 9 | e73721
information it can be expected that a higher amount ofexogenous copper would be required for laccase induction oncultures with cedar sawdust than with soybean pod husk andcoffee husk due to its lower content of copper In addition dueto the high copper-adsorption capacity of sawdust the amountof available copper in the culture would be reduced allowingthe laccase production at higher copper concentrations withoutreaching levels that might be toxic for the fungus This fact canbe observed in the SSC (Table 4) However for the SmC thisbehavior is not maintained for the cultures with soybean podhusk and coffee husk (Table 4) where the maximum laccaseactivity was obtained with a copper concentration of 05 and 5mM respectively This could be owed to a lower contactbetween the fungus and the substrate in the SmC whencompared to the SSC which is close related to the transport ofnutrients presented in the substrates
Production of extracellular laccases has been reported in alarge number of species of white-rot fungi strains grown onnatural substrates (Table 1) Although a laccase activity profileis not clearly observed based on the agro-industrial wasteculture mode and fungus strain most cases in which SmC andSSC were conducted using the same substrate and fungusstrain showed a higher laccase activity under SmC conditionsIn this study was observed a substrate dependentperformance where the most favorable condition to obtain thehighest laccase activity was the SSC The laccase activitiesobtained in the present study are comparable and in somecases higher than those reported for Pleurotus ostreatusPhanerochaete chrysosporium Pycnoporus cinnabarinus Tversicolor and Ganoderma sp cultured under SC and SmCconditions and using different agro-industrial wastes (Table 1)Semi-solid cultures using different agro-industrial wastes havebeen reported using T versicolor Phlebia radiata Tpubescens P ostreatus and C unicolor Rodriguez et al [16]screened different supports for laccase production by Tversicolor under SSC They obtained the highest laccaseactivity (1200 U L-1) when in the culture medium was presentedbarley bran In addition they were able to obtain a laccaseactivity of 1700 U L-1 when the culture was supplemented withxylidine as an inducer These results were obtained after 17 or18 days of culture and ABTS was used to determine theenzyme activity Maumlkelauml et al [4546] have studied theexpression of the genes involved in laccase production Pr-lac1and Pr-lac2 presented in P radiata and evaluate laccaseproduction in SSC using milled alder wood (Alnus incana) assubstrate They reported a maximum laccase activity of 3 microkatL-1 (~180 U L-1) at the 14th day of culture and usingsyringaldazine for its detection Osma et al [15] have studiedthe morphological changes and laccase production of strains Tpubescens P ostreatus C unicolor and T versicolor grownon wheat bran flakes using SSC They obtained a maximumlaccase production between 1397 and 2778 U L-1 after 10 and13 days of culture This enzyme activity is strain dependentand ABTS was used for its quantification Our results show agood correlation with reported data The highest laccaseactivity was obtained when T pubescens was grown on coffeehusk under SSC without copper (204 U L-1 evaluated withsyringaldazine or 1549 U L-1 evaluated with ABTS) which is in
some cases higher than reported ones This shows that underthe studied conditions the production of the laccase crudeextract is favored in SSC using coffee husk without the additionof copper which could favors the cost of the productionprocess
It has been reported for white rot-fungi the production ofmultiple laccase isoenzymes which are encoded by differentgenes and show different expression profiles stabilitiessubstrate affinities and molecular masses (ranging from 35 to140 kDa) [3647ndash49] Two isoforms have been identified in Tpubescens strains two monomeric (Lac1 and Lac2) enzymesof 67 kDa but with different pI (53 and 51) [50] and twolaccase-activity fractions (Lap1 and Lap2) with different pI (gt30 and 26) from which Lap2 is a monomeric enzyme of 65kDa [17] In this study we identify two laccase isoenzymesfrom T pubescens CBS 69694 that have different molecularmasses (120 and 60 kDa) and pI The purifications resultssuggest a more acidic pI for Lac1 (le 4) than Lac2 (ge 6) whichdiffers from a previous report for this fungus in which twofractions with laccase activity were obtained Lap1 and Lap2with pI gt 3 and of 26 respectively [17] Since in both casessimilar purification protocols were used and Lac1 and Lac2were recovered from the unbound and bound fractionsrespectively as Lap1 and Lap2 [17] this difference could bedue to the culture conditions used in each case (ie thepresence of agro-industrial wastes) Nevertheless the nanoLC-MSMS analysis showed that peptides of Lac1 and Lac2 werepresented in Lap1 (UniProtKBSwiss-Prot Q8TG93) and Lap2(UniProtKBSwiss-Prot Q8TG94) which have a predicted pI of43 and 47 respectively Lac1 and Lac2 peptides werepresented in Laccase-2 (UniProtKBSwiss-Prot Q99046) andLaccase-1 (UniProtKBSwiss-Prot Q99044) from T villosarespectively with 80 to 100 identity These T villosa laccaseisoenzymes have a predicted pI of 47 (laccase-1) and 60(laccase-2) and a 77 identity among them In addition Tvillosa laccase-2 is reported as a homodimer protein whilelaccase-1 is reported as monomeric protein Alignment of Lac1-Lap2 and Lac2-Lap1 from T pubescens CBS 69694 and Tvillosa showed a 96 and 70 identity respectively Finallysimilar peptides were identified for Lac1 and Lac2 from Tpubescens (Schumach) Pilaacutet (Syn Coriolus pubescens(Schum ex Fr) Queacutel) BCB 923-2 (Figure 3b) [51] Theseresults suggest that the purified fractions correspond to laccaseisoenzymes that differ from previously reported ones for this Tpubescens strain
As mentioned above laccases isoenzymes present differentstabilities and substrate affinities This behavior was alsoobserved for the laccase isoenzymes purified from Tpubescens with Lac1 showing lower laccase activity thanLac2 with ABTS as substrate However the difference inactivity between these isoenzymes depended on the agro-industrial waste used for the T pubescens growth suggestinga probable effect of the substrate composition on the laccasephysicochemical properties In addition it was observed thatthe total laccase activity presented in the UF retentates ishigher than the laccase activity of the purified fractionsregardless of the agro-industrial byproduct used for the Tpubescens growth This suggests a synergistic effect of Lac1
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 11 September 2013 | Volume 8 | Issue 9 | e73721
and Lac2 as have been reported for laccases obtained fromTrametes and Pleurotus strains [175253]
The maximum relative activity for the UF retentate waspresented at a pH from 20 to 40 regardless of the agro-industrial byproduct with a monotonical decrease at pHs abovethis range The relation between pH and type of electron donormolecule explains this behavior on laccase activity It has beenreported that when the substrate is an organic hydrogen donorie phenols the optimal pH ranges from 35 to 60 while thelaccase activity monotonically decreases from pH 25 to 70 ifthe substrate is an electron donor like ABTS [54] Despite thatat different pHs most of the purified laccase fractionspresented a similar performance as the UF retentates the Lac2fraction obtained from cedar sawdust differed from the previousobservation For this fraction the maximum activity wasobserved at pH 50 and despite that at acidic pH (ge 30) therelative activity was higher than 80 at lower pHs (20) itdecreased almost in a 50 Nevertheless at basic pHs therelative laccase activity presented a sharp decreased that iswell correlate with the other laccase fractions We are not ableto attribute this particular trend to the presence of multiplelaccase isoenzymes in the fraction since the SDS-PAGErevealed only one band when stained with ABTS and from theNanoLC-MSMS analysis no difference in the number ofpeptides and their sequence was detected We speculate thatthis particular behavior can be owed to a compound added inthe pre-treatment of the cedar wood that might be used forpreserving it that might affect the activity of Lac2 and that wasnot completely removed during the purification
Temperature stability evaluation showed that Lac1 is themost thermolabile fraction while the laccase activity reductionin the crude extracts did not seem to be the result of anadditive effect of the activity loss of the purified fractionsAlthough there are no reports of laccase stability under theselected conditions these results agree with previous reportsthat state that fungal laccases rapidly decrease their activity attemperatures near 60 degC and thermal stability correlates withthe temperature range of the growth of the source organism[55] In addition we previously reported for a laccase crudeextract obtained from culturing T pubescens on coffee husk anactivity loss of 26 and 10 at 60 and 50 degC respectivelyafter 8 h at pH 60 using syringaldazine as substrate [48]
Finally an electrochemical characterization was carried outfor selected crude extracts and purified fractions The results
showed a difference between the electrochemicalperformances of extracts and purified fractions withsyringaldazine or ABTS These differences could be owed tothe different mechanisms for electron transfer as discussedabove However the overall performance of the obtained cyclicvoltammograms for ABTS and syringaldazine agrees withresults reported by Fernaacutendez-Saacutenchez et al [25] who used asaturated calomel reference electrode
Conclusions
We have shown the potential of coffee husk soybean podhusk and cedar sawdust as substrates for the laccaseproduction In several countries like Brazil and Colombia theproduction of coffee and soybean is of great importance andthe generated volume of residues is high The results showedthat semi-solid culture of T pubescens using coffee huskallows to obtain an important amount of laccase which couldbe favored by the phenolic-compounds ie tannins presentedin coffee husk While supplementation of the culture mediumwith copper favored the production of laccase in submergedand semi-solid culture using soybean pod husk and cedarsawdust semi-solid culture using coffee husk which presentedthe highest laccase volumetric yield did not require coppersupplementation This is an economic advantage that shouldbe considered when scaling up the process In this study Tpubescens showed its ability to produce two independentlaccases regardless of the used substrate However a higherlaccase activity was observed for crude extracts rather than forpurified fractions This suggests that Lac1 and Lac2 might havea synergistic effect on the final laccase activity rather than anadditive effect Based on this study the investigation of laccaseproduction by evaluating other agro-industrial wastes in whichphenolic-compounds constitute an important percentage oftheir composition is of great interest to avoid the use ofinducers that would increase the production cost
Author Contributions
Conceived and designed the experiments JFO CJAD OFSPerformed the experiments JCG SCM AR Analyzed the dataJCG SCM AR JFO CJAD OFS Wrote the manuscript AR JFOCJAD OFS
References
1 Vandamme EJ (2009) Agro-industrial residue utilization for industrialbiotechnology products In P NigamA Pandey Biotechnology for Agro-Industrial Residues Utilisation Netherlands Springer Verlag pp 3-11
2 Martiacutenez AT Speranza M Ruiz-Duentildeas FJ Ferreira P Camarero S etal (2005) Biodegradation of lignocellulosics microbial chemical andenzymatic aspects of the fungal attack of lignin Int Microbiol 8195-204 PubMed 16200498
3 Saacutenchez O Sierra R Almeacuteciga-Diacuteaz CJ (2011) Delignification processof agro-industrial aastes an alternative to obtain fermentablecarbohydrates for producing fuel In M Manzanera Alternative FuelIntech 111-154
4 Wesenberg D Kyriakides I Agathos SN (2003) White-rot fungi andtheir enzymes for the treatment of industrial dye effluents BiotechnolAdv 22 161-187 doi101016jbiotechadv200308011 PubMed14623049
5 Morozova OV Shumakovich GP Shleev SV Yaropolov YI (2007)Laccase-mediator systems and their applications A review ApplBiochem Microbiol 43 523-535 doi101134S0003683807050055
6 Baldrian P (2006) Fungal laccases - occurrence and properties FEMSMicrobiol Rev 30 215-242 doi101111j1574-4976200500010xPubMed 16472305
7 Rodriacuteguez Couto S Herrera JLT (2006) Industrial and biotechnologicalapplications of laccases A review Biotechnol Adv 24 500-513 doi101016jbiotechadv200604003 PubMed 16716556
8 Osma JF Toca-Herrera JL Rodriacuteguez-Couto S (2010) Uses oflaccases in the food industry Enzyme Res 2010 2010 918761PubMed 21048873
9 Rodriguez S (2008) Exploitation of biological wastes for the productionof value-added products under solid-state fermentation conditionsBiotechnol J 3 859-870 doi101002biot200800031 PubMed18543242
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 12 September 2013 | Volume 8 | Issue 9 | e73721
10 Osma JF Toca-Herrera JL Rodriacuteguez-Couto S (2011) Cost analysis inlaccase production J Environ Manage 92 2907-2912 doi101016jjenvman201106052 PubMed 21775046
11 Jain A Morlok CK Henson JM (2013) Comparison of solid-state andsubmerged-state fermentation for the bioprocessing of switchgrass toethanol and acetate by Clostridium phytofermentans Appl MicrobiolBiotechnol 97 905-917 doi101007s00253-012-4511-4 PubMed23111595
12 Singhania RR Patel AK Soccol CR Pandey A (2009) Recentadvances in solid-state fermentation Biochem Eng J 44 13-18 doi101016jbej200810019
13 Economou CN Makri A Aggelis G Pavlou S Vayenas DV (2010)Semi-solid state fermentation of sweet sorghum for the biotechnologicalproduction of single cell oil Bioresour Technol 101 1385-1388 doi101016jbiortech200909028 PubMed 19781936
14 Rodriacuteguez CS Santoro R Cameselle C Sanroman A (1997) Laccaseproduction in semi-solid cultures of Phanerochaete chrysosporiumBiotechnol Lett 19 995-998 doi101023A1018495216946
15 Osma JF Moilanen U Toca-Herrera JL Rodriacuteguez-Couto S (2011)Morphology and laccase production of white-rot fungi grown on wheatbran flakes under semi-solid-state fermentation conditions FEMSMicrobiol Lett 318 27-34 doi101111j1574-6968201102234xPubMed 21291496
16 Couto SR Gundin M Lorenzo M Sanroman MN (2002) Screening ofsupports and inducers for laccase production by Trametes versicolor insemi-solid-state conditions Proc Biochem 38 249-255 doi101016S0032-9592(02)00087-0
17 Galhaup C Goller S Peterbauer CK Strauss J Haltrich D (2002)Characterization of the major laccase isoenzyme from Trametespubescens and regulation of its synthesis by metal ions Microbiology148 2159-2169 PubMed 12101303
18 Sievers F Wilm A Dineen D Gibson TJ Karplus K et al (2011) Fastscalable generation of high-quality protein multiple sequencealignments using Clustal Omega Mol Syst Biol 7 539 PubMed21988835
19 Childs RE Bardsley WG (1975) The steady-state kinetics of peroxidasewith 22-azino-di-(3-ethyl-benzthiazoline-6-sulphonic acid) aschromogen Biochem J 145 93-103 PubMed 1191252
20 Harkin JM Obst JR (1973) Syringaldazine an Effective Reagent forDetecting Laccase and Peroxidase in Fungi Experientia 29 382-387
21 Nikupaavola ML Raaska L Itavaara M (1990) Detection of white-rotfungi by a non-roxic stain Mycol Res 94 27-31 doi101016S0953-7562(09)81260-4
22 Gonzalez LF Sarria V Sanchez OF (2010) Degradation ofchlorophenols by sequential biological-advanced oxidative processusing Trametes pubescens and TiO(2)UV Bioresour Technol 1013493-3499
23 Waterborg JH (2009) The Lowry method for protein quantitation In JWalker The Protein Protocols Handbook Humana Press pp 7-10
24 Harkin JM Obst JR (1973) Syringaldazine an effective reagent fordetecting laccase and peroxidase in fungi Cell Mol Life Sci 29381-387 doi101007BF01926734
25 Fernaacutendez-Saacutenchez C Tzanov T Guumlbitz GM Cavaco-Paulo A (2002)Voltammetric monitoring of laccase-catalysed mediated reactionsBioelectrochemistry 58 149-156 doi101016S1567-5394(02)00119-6PubMed 12414320
26 Bourbonnais R Leech D Paice MG (1998) Electrochemical analysis ofthe interactions of laccase mediators with lignin model compoundsBiochim Biophys Acta 1379 381ndash390 doi101016S0304-4165(97)00117-7 PubMed 9545600
27 Ferraz F Silva S (2009) Characterization of coffee husk biomass forbiotechnological purposes N Biotechnol 25 S256-S256 doi101016jnbt200906573
28 Pandey A Soccol CR Nigam P Brand D Mohan R et al (2000)Biotechnological potential of coffee pulp and coffee husk forbioprocesses Biochem Eng J 6 153-162 doi101016S1369-703X(00)00084-X PubMed 10959086
29 Sruamsiri S Silman P (2008) Nutritive composition of soybean by-products and nutrient digestibility of soybean pod husk Maejo Int J SciTechnol 2 568-576
30 Romkaew J Nagaya Y Goto M Suzuki J Umezaki T (2008) Poddehiscence in relation to chemical components of pod shell in soybeanPlant Prod Sci 11 278-282 doi101626pps11278
31 Nakamanee G Polbumroong T Phaikaew C (2003) Alternative use ofcrop waste to increase milk production Proceedings of the 8th Meetingof the Regional Working Group on Grazing and Feed Resources forSoutheast Asia Kuala Lumpur Malaysia 66 - 70
32 Kurek B Gaudard F (2000) Oxidation of spruce wood sawdust byMnO(2) plus oxalate a biochemical investigation J Agric Food Chem48 3058-3062 doi101021jf000015g PubMed 10898665
33 Weil J Brewer M Hendrickson R Sarikaya A Ladisc MR (1998)Continuous pH monitoring during pretreatment of yellow poplar woodsawdust by pressure cooking in water Appl Biochem Biotechnol 70-7299-111 doi101007BF02920127
34 Weil J Sarikaya A Rau S-L Goetz J Ladisch CM et al (1997)Pretreatment of yellow poplar sawdust by pressure cooking in waterAppl Biochem Biotechnol 68 21-40 doi101007BF02785978
35 Dekker RF Barbosa AM Giese EC Godoy SD Covizzi LG (2007)Influence of nutrients on enhancing laccase production byBotryosphaeria rhodina MAMB-05 Int Microbiol 10 177-185 PubMed18075999
36 Xiao YZ Hong YZ Li JF Hang J Tong PG et al (2006) Cloning ofnovel laccase isozyme genes from Trametes sp AH28-2 and analysesof their differential expression Appl Microbiol Biotechnol 71 493-501doi101007s00253-005-0188-2 PubMed 16283298
37 Crowe JD Olsson S (2001) Induction of laccase activity in Rhizoctoniasolani by antagonistic Pseudomonas fluorescens strains and a range ofchemical treatments Appl Environ Microbiol 67 2088-2094 doi101128AEM6752088-20942001 PubMed 11319086
38 Yaguumle S Terroacuten MC Gonzaacutelez T Zapico E Bocchini P et al (2000)Biotreatment of tannin-rich beer-factory wastewater with white-rotbasidiomycete Coriolopsis gallica monitored by pyrolysisgaschromatographymass spectrometry Rapid Commun Mass Spectrom14 905-910 doi101002(SICI)1097-0231(20000530)1410 PubMed10825255
39 Gowda NKS Ramana JV Prasad CS Singh K (2004) Micronutrientcontent of certain tropical conventional and unconventional feedresources of southern India Trop Anim Heatlth Pro 36 77-94 doi101023BTROP0000009522309491d PubMed 14979561
40 DePeters EJ Fadel JG Arana MJ Ohanesian N Etchebarne MA et al(2000) Variability in the chemical composition of seventeen selectedby-product feedstuffs used by the California dairy industry Prof AnimSci 16 69-99
41 Obernberger I Thek G (2004) Physical characterisation and chemicalcomposition of densified biomass fuels with regard to their combustionbehaviour Biomass Bioenerg 27 653-669 doi101016jbiombioe200307006
42 Larous S Meniai A-H Lehocine MB (2005) Experimental study of theremoval of copper from aqueous solutions by adsorption usingsawdust Desalination 185 483ndash490 doi101016jdesal200503090
43 Oliveira WE Franca AS Oliveira LS Rocha SD (2008) Untreatedcoffee husks as biosorbents for the removal of heavy metals fromaqueous solutions J Hazard Mater 152 1073ndash1081 doi101016jjhazmat200707085 PubMed 17804159
44 Šciban M Klašnja M Škrbic B (2008) Adsorption of copper ions fromwater by modified agricultural by-products Desalination 229 170ndash180doi101016jdesal200708017
45 Maumlkelauml MR Hildeacuten KS Hakala TK Hatakka A Lundell TK (2006)Expression and molecular properties of a new laccase of the white rotfungus Phlebia radiata grown on wood Curr Genet 50 323-333 doi101007s00294-006-0090-1 PubMed 16927090
46 Maumlkelauml MR Lundell T Hatakka A Hildeacuten K (2013) Effect of coppernutrient nitrogen and wood-supplement on the production of lignin-modifying enzymes by the white-rot fungus Phlebia radiata Fungal Biol117 62-70 doi101016jfunbio201211006 PubMed 23332834
47 Dwivedi UN Singha P Pandeya VP Kumara A (2011) Structurendashfunction relationship among bacterial fungal and plant laccases J MolCatal B Enzym 68 117-128 doi101016jmolcatb201011002
48 Gaitan IJ Medina SC Gonzaacutelez JC Rodriacuteguez A Espejo AacuteJ et al(2011) Evaluation of toxicity and degradation of a chlorophenol mixtureby the laccase produced by Trametes pubescens Bioresour Technol102 3632ndash3635 doi101016jbiortech201011040 PubMed21115244
49 Karp SG Faraco V Amore A Birolo L Giangrande C et al (2012)Characterization of laccase isoforms produced by Pleurotus ostreatusin solid state fermentation of sugarcane bagasse Bioresour Technol114 735-739 doi101016jbiortech201203058 PubMed 22487128
50 Nikitina OV Shleev SV Gorshina ES Rusinova TV Serezhenkov VAet al (2005) Isolation and purification of enzymes from ligninolyticcomplex of the basidial fungus Trametes pubescens (Schumach) Pilatand study of their properties Biochemistry (Mosc) 70 1274-1279 doi101007s10541-005-0259-0
51 Shleev S Nikitina O Christenson A Reimann CT Yaropolov AI et al(2007) Characterization of two new multiforms of Trametes pubescenslaccase Bioorg Chem 35 35-49 doi101016jbioorg200608001PubMed 16989887
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 13 September 2013 | Volume 8 | Issue 9 | e73721
52 Xiao YZ Chen Q Hang J Shi YY Wu J et al (2004) Selectiveinduction purification and characterization of a laccase isozyme fromthe basidiomycete Trametes sp AH28-2 Mycologia 96 26-35 doi1023073761984 PubMed 21148825
53 Muntildeoz C Guilleacuten F Martiacutenez AT Martiacutenez MJ (1997) Laccaseisoenzymes of Pleurotus eryngii characterization catalytic propertiesand participation in activation of molecular oxygen and Mn2+ oxidationAppl Environ Microbiol 63 2166-2174 PubMed 9172335
54 Morozova OV Shumakovich GP Gorbacheva MA Shleev SVYaropolov AI (2007) Blue laccases Biochemistry (Mosc) 721136-1150 doi101134S0006297907100112
55 Hildeacuten K Hakala TK Lundell T (2009) Thermotolerant andthermostable laccases Biotechnol Lett 31 1117-1128 doi101007s10529-009-9998-0 PubMed 19360388
56 Verma P Madamwar D (2002) Production of ligninolytic enzymes fordye decolorization by cocultivation of white-rot fungi Pleurotusostreatus and Phanerochaete chrysosporium under solid-statefermentation Appl Biochem Biotechnol 102-103 109-118 doi101385ABAB102-1031-6109 PubMed 12396115
57 Meza JC Lomascolo A Casalot L Sigoillot J-C Auria R (2005)Laccase production by Pycnoporus cinnabarinus grown on sugar-canebagasse Influence of ethanol vapours as inducer Proc Biochem 403365ndash3371 doi101016jprocbio200503004
58 Hossain SM Anantharaman N (2006) Activity enhancement ofligninolytic enzymes of Trametes versicolor with bagasse powder Afr JBiotechnol 5 189-194
59 Revankar MS Desai KM Lele SS (2007) Solid-state fermentation forenhanced production of laccase using indigenously isolatedGanoderma sp Appl Biochem Biotechnol 143 16-26 doi101007s12010-007-0029-0 PubMed 18025593
60 Velazquez-Cedeno MA Mata G Savoie J-M (2002) Waste-reducingcultivation of Pleurotus ostreatus and Pleurotus pulmonarius on coffeepulp changes in the production of some lignocellulolytic enzymesWorld J Microbiol Biotechnol 18 201ndash207 doi101023A1014999616381
61 Mata G Murrieta D Andreu LI (2005) Changes in lignocellulolyticenzyme activities in six Pleurotus spp strains cultivated on coffee pulpin confrontation with Trichoderma spp World J Microbiol Biotechnol 21143ndash150 doi101007s11274-004-3041-3
62 de Souza DF Tychanowicz GK de Souza CG Peralta RM (2006) Co-production of ligninolytic enzymes by Pleurotus pulmonarius on wheatbran solid state cultures J Basic Microbiol 46 126-134 doi101002jobm200510014 PubMed 16598826
63 Prasad KK Mohan SV Bhaskar YV Ramanaiah SV Babu VL et al(2005) Laccase production using Pleurotus ostreatus 1804 immobilizedon PUF cubes in batch and packed bed reactors influence of cultureconditions J Microbiol 43 301-307 PubMed 15995650
64 Songulashvili G Elisashvili V Wasser SP Nevo E Hadar Y (2007)Basidiomycetes laccase and manganese peroxidase activity in
submerged fermentation of food industry wastes Enzyme MicrobTechnol 41 57ndash61 doi101016jenzmictec200611024
65 Sathishkumar P Murugesan K Palvannan T (2010) Production oflaccase from Pleurotus florida using agro-wastes and efficientdecolorization of Reactive blue 198 J Basic Microbiol 50 360-367 doi101002jobm200900407 PubMed 20586068
66 Moldes D Gallego PP Rodriacuteguez Couto S Sanromaacuten A (2003) Grapeseeds the best lignocellulosic waste to produce laccase by solid statecultures of Trametes hirsuta Biotechnol Lett 25 491-495 doi101023A1022660230653 PubMed 12882277
67 Elisashvili V Kachlishvili E Penninckx M (2008) Effect of growthsubstrate method of fermentation and nitrogen source onlignocellulose-degrading enzymes production by white-rotbasidiomycetes J Ind Microbiol Biotechnol 35 1531-1538 doi101007s10295-008-0454-2 PubMed 18716810
68 Osma JF Saravia V Herrera JL Couto SR (2007) Mandarin peelingsthe best carbon source to produce laccase by static cultures ofTrametes pubescens Chemosphere 67 1677-1680 doi101016jchemosphere200611051 PubMed 17234250
69 Kahraman SS Gurdal IH (2002) Effect of synthetic and natural culturemedia on laccase production by white rot fungi Bioresour Technol 82215-217 doi101016S0960-8524(01)00193-6 PubMed 11991068
70 Iandolo D Piscitelli A Sannia G Faraco V (2011) Enzyme Productionby Solid Substrate Fermentation of Pleurotus ostreatus and Trametesversicolor on Tomato Pomace Appl Biochem Biotechnol 163 40-51doi101007s12010-010-9014-0 PubMed 20582639
71 Moldes D Lorenzo M Sanromaacuten MA (2004) Different proportions oflaccase isoenzymes produced by submerged cultures of Trametesversicolor grown on lignocellulosic wastes Biotechnol Lett 26 327-330doi101023BBILE000001545240213bf PubMed 15055770
72 Philippoussis A Diamantopoulou P Papadopoulou K Lakhtar HRoussos S et al (2010) Biomass laccase and endoglucanaseproduction by Lentinula edodes during solid state fermentation of reedgrass bean stalks and wheat straw residues World J MicrobiolBiotechnol 27 285-297
73 Fenice M Giovannozzi Sermanni G Federici F DrsquoAnnibale A (2003)Submerged and solid-state production of laccase and Mn-peroxidaseby Panus tigrinus on olive mill wastewater-based media J Biotechnol100 77-85 doi101016S0168-1656(02)00241-9 PubMed 12413788
74 Rodriguez-Couto S GundIn M Lorenzo M Sanromaacuten MAacute (2002)Screening of supports and inducers for laccase production by Trametesversicolor in semi-solid-state conditions Proc Biochem 38 249-255doi101016S0032-9592(02)00087-0
75 Fackler K Gradinger C Schmutzer M Tavzes C Burgert I et al (2007)Biotechnological wood modification with selective white-rot fungi and itsmolecular mechanisms Food Technol Biotechnol 45 269ndash276
76 Levin L Herrmann C Papinutti VL (2008) Optimization oflignocellulolytic enzyme production by the white-rot fungus Trametestrogii in solid-state fermentation using response surface methodologyBiochem Eng J 39 207-214 doi101016jbej200709004
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 14 September 2013 | Volume 8 | Issue 9 | e73721
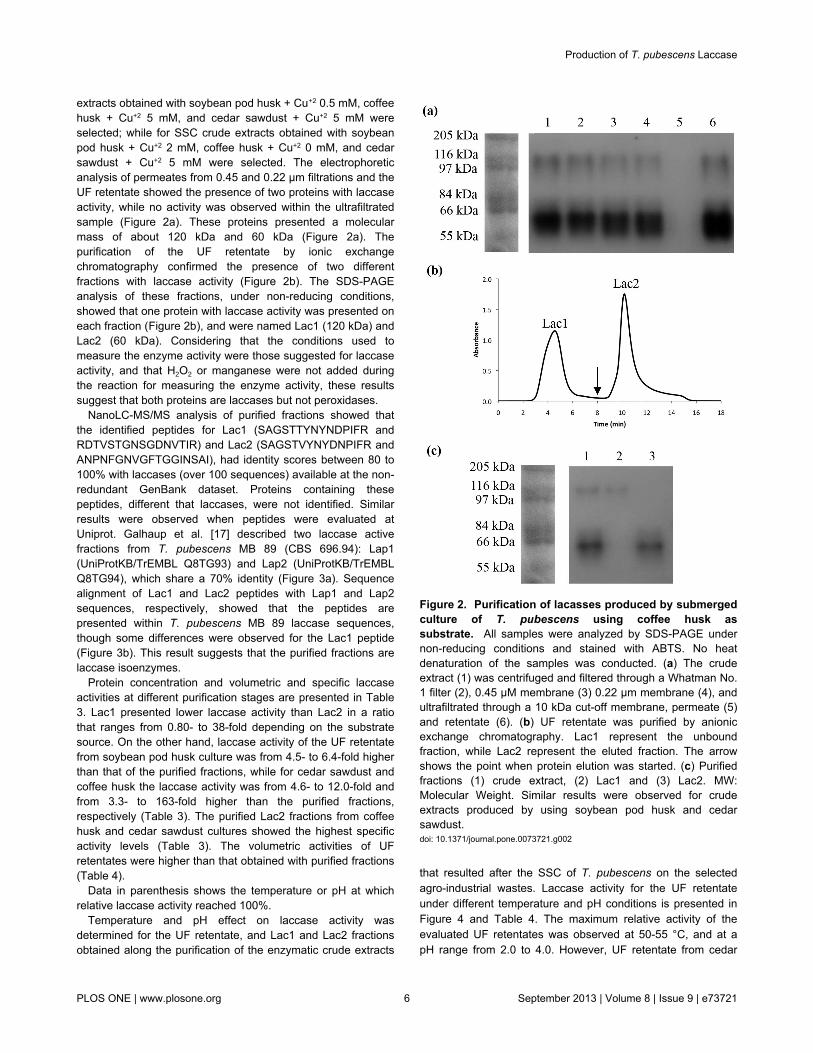
extracts obtained with soybean pod husk + Cu+2 05 mM coffeehusk + Cu+2 5 mM and cedar sawdust + Cu+2 5 mM wereselected while for SSC crude extracts obtained with soybeanpod husk + Cu+2 2 mM coffee husk + Cu+2 0 mM and cedarsawdust + Cu+2 5 mM were selected The electrophoreticanalysis of permeates from 045 and 022 microm filtrations and theUF retentate showed the presence of two proteins with laccaseactivity while no activity was observed within the ultrafiltratedsample (Figure 2a) These proteins presented a molecularmass of about 120 kDa and 60 kDa (Figure 2a) Thepurification of the UF retentate by ionic exchangechromatography confirmed the presence of two differentfractions with laccase activity (Figure 2b) The SDS-PAGEanalysis of these fractions under non-reducing conditionsshowed that one protein with laccase activity was presented oneach fraction (Figure 2b) and were named Lac1 (120 kDa) andLac2 (60 kDa) Considering that the conditions used tomeasure the enzyme activity were those suggested for laccaseactivity and that H2O2 or manganese were not added duringthe reaction for measuring the enzyme activity these resultssuggest that both proteins are laccases but not peroxidases
NanoLC-MSMS analysis of purified fractions showed thatthe identified peptides for Lac1 (SAGSTTYNYNDPIFR andRDTVSTGNSGDNVTIR) and Lac2 (SAGSTVYNYDNPIFR andANPNFGNVGFTGGINSAI) had identity scores between 80 to100 with laccases (over 100 sequences) available at the non-redundant GenBank dataset Proteins containing thesepeptides different that laccases were not identified Similarresults were observed when peptides were evaluated atUniprot Galhaup et al [17] described two laccase activefractions from T pubescens MB 89 (CBS 69694) Lap1(UniProtKBTrEMBL Q8TG93) and Lap2 (UniProtKBTrEMBLQ8TG94) which share a 70 identity (Figure 3a) Sequencealignment of Lac1 and Lac2 peptides with Lap1 and Lap2sequences respectively showed that the peptides arepresented within T pubescens MB 89 laccase sequencesthough some differences were observed for the Lac1 peptide(Figure 3b) This result suggests that the purified fractions arelaccase isoenzymes
Protein concentration and volumetric and specific laccaseactivities at different purification stages are presented in Table3 Lac1 presented lower laccase activity than Lac2 in a ratiothat ranges from 080- to 38-fold depending on the substratesource On the other hand laccase activity of the UF retentatefrom soybean pod husk culture was from 45- to 64-fold higherthan that of the purified fractions while for cedar sawdust andcoffee husk the laccase activity was from 46- to 120-fold andfrom 33- to 163-fold higher than the purified fractionsrespectively (Table 3) The purified Lac2 fractions from coffeehusk and cedar sawdust cultures showed the highest specificactivity levels (Table 3) The volumetric activities of UFretentates were higher than that obtained with purified fractions(Table 4)
Data in parenthesis shows the temperature or pH at whichrelative laccase activity reached 100
Temperature and pH effect on laccase activity wasdetermined for the UF retentate and Lac1 and Lac2 fractionsobtained along the purification of the enzymatic crude extracts
that resulted after the SSC of T pubescens on the selectedagro-industrial wastes Laccase activity for the UF retentateunder different temperature and pH conditions is presented inFigure 4 and Table 4 The maximum relative activity of theevaluated UF retentates was observed at 50-55 degC and at apH range from 20 to 40 However UF retentate from cedar
Figure 2 Purification of lacasses produced by submergedculture of T pubescens using coffee husk assubstrate All samples were analyzed by SDS-PAGE undernon-reducing conditions and stained with ABTS No heatdenaturation of the samples was conducted (a) The crudeextract (1) was centrifuged and filtered through a Whatman No1 filter (2) 045 microM membrane (3) 022 microm membrane (4) andultrafiltrated through a 10 kDa cut-off membrane permeate (5)and retentate (6) (b) UF retentate was purified by anionicexchange chromatography Lac1 represent the unboundfraction while Lac2 represent the eluted fraction The arrowshows the point when protein elution was started (c) Purifiedfractions (1) crude extract (2) Lac1 and (3) Lac2 MWMolecular Weight Similar results were observed for crudeextracts produced by using soybean pod husk and cedarsawdustdoi 101371journalpone0073721g002
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 6 September 2013 | Volume 8 | Issue 9 | e73721
sawdust culture behaved different to the UF retentate fromcoffee husk and soybean pod husk cultures In this sense the
UF retentate from cedar sawdust culture presented the lowestenzyme activity at temperatures le 40 degC but at 70 degC
Figure 3 Aligment of reported lacasses sequences and Lac1 and Lac2 peptides (a) Alignment of Lap1 (UniProtKBSwiss-Prot Q8TG93) and Lap2 (UniProtKBSwiss-Prot Q8TG94) from T pubescens CBS 69694 (b) Alignment of Lac1 and Lac2 peptideswith Lap1 and Lap2 respectively and tryptic peptides described by Shleev et al 2007 Alignments were carried out using ClustalOmega and figures were using CLC Sequence Viewer Different residues are colored in reddoi 101371journalpone0073721g003
Table 3 Laccase activity during the purification of crude extracts resulted from T pubescens culture on agro-industrialwastes under submerged (SmC) and semisolid (SSC) cultures
Soybean pod husk Coffee husk Cedar sawdust
Protein(mg mL-1)
VolumetricActivity (UL-1)
SpecificActivity (Umg-1
protein)Recovery()
Protein(mg mL-1)
VolumetricActivity (UL-1)
SpecificActivity (Umg-1
protein)Recovery()
Protein(mg mL-1)
VolumetricActivity (UL-1)
SpecificActivity (Umg-1
protein)Recover()
SmCCrudeExtract
0869 52888 0608 100 0414 152708 3688 100 0431 10735 0249 100
045 micromFiltration
0736 46549 0632 88 0334 117892 3529 77 0444 12053 0271 112
UF retentate 1010 78731 0779 74 0378 195263 5165 64 0510 25905 0507 120
Lac1 0494 5471 0110 1 0232 2399 0103 02 0222 838 0037 08
Lac2 0310 20661 0666 4 0116 107240 9252 7 0040 15234 3837 14 SSCCrudeExtract
1146 164966 1439 100 0596 154907 2601 100 1065 23203 0217 100
045 micromFiltration
0776 128392 1654 77 0474 115736 2439 75 0806 25824 0320 111
UF retentate 0973 335055 3443 101 0428 161750 3779 52 1134 51101 0450 110
Lac1 0097 8854 0912 2 0104 1514 0145 03 0042 1315 0313 2
Lac2 0168 48988 2915 10 0131 82298 6282 18 0072 16162 2244 23
740 ABTS was used to determine laccase activity
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 7 September 2013 | Volume 8 | Issue 9 | e73721
presented the highest enzyme activity ~90 of its initial valuein comparison to the other two UF retentates (Figure 4)
The temperature and pH effect on laccase activity for Lac1and Lac2 is presented in Figure 5 The maximum relativeactivity for both fractions was presented at a pH from 20 to 40regardless of the agro-industrial waste as observed for the UFretentate However at pH gt 40 Lac1 showed a sharperdecrease in its activity than that observed for Lac2Nevertheless Lac2 purified from cedar sawdust culturepresented a different performance This fraction showed themaximum activity at pH 50 while a sharp drop was observedat pHs above or below this value
Laccase activity profiles showed that Lac1 and Lac2presented the maximum enzyme activity at 55 and 60 degCrespectively (Table 4) It was observed that Lac1 from coffeehusk culture presented a lower decrease in enzyme activity attemperatures le 50 degC or ge 60 degC than Lac2 On the contraryLac2 from soybean husk or cedar sawdust cultures presenteda higher relative laccase activity than Lac1 at temperatures le55 degC The purified fractions from soybean pod husk and cedarsawdust cultures presented the lowest relative activity at 30 degCand 65 degC
Table 4 Optimum temperatures and pHs for maximumlaccase activities in UF retentate Lac1 and Lac2 obtainedfrom semisolid cultured T pubescens on agro-industrialwastes
EnzymaticFraction Temperature pH
VolumetricActivity (UL-1)
SpecificActivity (Umg-1 protein)
VolumetricActivity (UL-1)
SpecificActivity (Umg-1
protein)SoybeanPod Husk +2 mM Cu+2
148000plusmn1178(55)
1521plusmn0012182037plusmn3025(2-3)
1871plusmn0031
Lac122925plusmn917(60)
2363plusmn009536135plusmn831(23)
3725plusmn0085
Lac232638plusmn1558(55)
1942plusmn009228713plusmn274(23)
1709plusmn0016
CoffeeHusk + 0mM Cu+2
113740plusmn8289(50-55)
3008plusmn0219290666plusmn1039(2-3)
7689plusmn0275
Lac1 889plusmn039 (55) 0038plusmn0002 1779plusmn028 (23) 0077plusmn0001
Lac234194plusmn2789(60)
2947plusmn024033150plusmn526 (2ndash4)
2857plusmn0045
CedarSawdust +5 mM Cu+2
257666plusmn1201(55)
5052plusmn0024344634plusmn1043(2-3)
6757plusmn0204
Lac121370plusmn472(55)
0962plusmn002129117plusmn943(23)
1311plusmn0042
Lac255166plusmn1656(60)
1379plusmn0414 55166plusmn1656 (5) 1379plusmn0414
ABTS was used to determine laccase activity
Laccase stabilityFor selected enzymatic extracts and purified fractions (Table
4) a stability test for 48 h at their optimal pH and temperaturewas conducted Time course of the relative laccase activity forcrude extract and purified fractions from coffee husk and cedarsawdust cultures are presented in Figure 5 Similar resultswere observed for crude extract and purified fractions fromsoybean pod husk culture (data no shown)
It was observed that the activity loss depends on thesubstrate source used to grow T pubescens An activity loss of55 was observed for crude extracts and purified fractionsfrom coffee husk and soybean pod husk cultures while anactivity loss of 70 was observed for the crude extract andpurified fractions from cedar sawdust culture NeverthelessLac2 showed the lowest activity reduction with values of 10 40and 55 for coffee husk soybean pod husk and cedarsawdust respectively while Lac1 presented an activity loss of34 50 and 60 for coffee husk soybean pod husk and cedarsawdust respectively Despite the differences in laccaseactivity among agro-industrial wastes and between crudeextracts and purified fractions all the profiles showed a sharp
Figure 4 Effect of temperature (a) and pH (b) on laccaseactivity for the UF retentate obtained by semisolid cultureof T pubescens on coffee husk () soybean pod husk (loz)and cedar sawdust (Δ) Laccase stability was evaluated byusing ABTS All the assays were performed in triplicatedoi 101371journalpone0073721g004
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 8 September 2013 | Volume 8 | Issue 9 | e73721
decrease during the first 8 h while a reduced activity loss wasobserved during the last 24 h of the assay (Figure 6)
Laccase electrochemical characterizationFor selected crude extracts and purified fractions a cyclic
voltammetry characterization was conducted For eachmediator syringaldazine or ABTS similar cyclic
Figure 5 Effect of temperature () and pH () on enzymeactivity for Lac1 (solid line) and Lac2 (dashed line)obtained after the purification of the enzymatic crudeextract produced by semisolid culture of T pubescens oncoffee husk (a) soybean pod husk (b) and cedar sawdust(c) Laccase stability was evaluated by using ABTS All theassays were performed in triplicatedoi 101371journalpone0073721g005
voltammograms were obtained for the different crude extractsand purified fractions Voltammograms of the redox reaction forsyringaldazine (Figure 7a) and ABTS (Figure 6b) arepresented Two anionic and cationic peaks were observedwhen syringaldazine was used as mediator The first pair ofpeaks showed a reversible performance with a mean Ia Icratio of 109plusmn006 106plusmn005 and 098plusmn002 for the crudeextracts Lac1 and Lac2 respectively The second pair ofpeaks showed quasi-reversible performance and the cathodicpeak was presented at a potential of +062 V (vs Ag|AgCl)while the anodic peak was presented at a potential of +060 V(vs Ag|AgCl) No significant difference on the reductionpotential (Eo) for syringaldazine was observed for the crudeextracts or the purified fractions obtaining an Eo of0314plusmn0008 V (vs Ag|AgCl) However the peak potentialseparation (ΔEp) showed a different profile depending on thesubstrate used for the culture While not significant difference(p gt 005) on ΔEp value was observed between crude extract(0045plusmn0005 V vs Ag|AgCl) Lac1 (0046plusmn0005 V vs Ag|AgCl)and Lac2 (0028plusmn0001 V vs Ag|AgCl) from cultures with cedarsawdust and soybean pod husk the ΔEp was considerablyhigher for the crude extract (0083plusmn0004 V vs Ag|AgCl) Lac1(0068plusmn0003 V vs Ag|AgCl) and Lac2 (0051plusmn0005 V vs Ag|AgCl) from the coffee husk culture
For reactions conducted with ABTS and the different crudeextracts and purified fractions the voltammograms exhibitedone pair of cathodic and anodic peaks (Figure 7b) whichcorresponds to the redox of ABTSABTS+ (IaIc) with an Eo of0487plusmn0006 V (vs Ag|AgCl) This process was reversiblepresenting a mean Ia Ic ratio of 099plusmn002 In addition asecond cathodic peak was presented at +083 V (vs Ag|AgCl)when ABTS redox reaction was conducted using the crudeextract and the purified fractions This performance agrees withprevious reports in which the laccase activity cannot beobserved at potential values of the second reversible oxidation
Figure 6 Laccase stability profiles at optimal pH andtemperature conditions for crude extracts (solid line) Lac1(dashed line) and Lac2 (spotted line) obtained fromsemisolid culture of T pubescens on coffee husk (loz) andcedar sawdust (bull) Laccase stability was evaluated by usingABTS All the assays were performed in triplicatedoi 101371journalpone0073721g006
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 9 September 2013 | Volume 8 | Issue 9 | e73721
of ABTS ABTS+ABTS2+ due to this catalytic process cannotbe detected by cyclic voltammetry [2526] Neverthelessvoltammograms for laccase crude extract from a soybean podhusk culture presented the second anodic peak which inaddition to the second cathodic peak corresponds to the redoxof ABTS+ABTS2+ (IIaIIc) with an Eo of 0942plusmn0004 V (vs Ag|AgCl) Contrary to the reactions conducted usingsyringaldazine as substrate the ΔEp did not present asignificant difference for the crude extracts and purifiedfractions 0248plusmn0016 V (vs Ag|AgCl)
Discussion
In this study we evaluated the effect of the culture mode ieSmC and SSC on laccase production by T pubescens CBS69694 using three different agro-industrial wastes (coffeehusk soybean pod husk and cedar sawdust) Furthermore theeffect of Cu+2 supplementation on laccase production was alsoevaluated and the crude extracts and the purified fractionswere partially characterized
Figure 7 Cyclic voltammograms of (a) syringaldazine atpH 50 using the crude extract and purified laccasefractions from semisolid culture of T pubescens on coffeehusk or (b) ABTS at pH 30 using the crude extract andpurified laccase fractions from semisolid culture of Tpubescens on soybean pod husk Nomenclature onlymediator (dotted line) crude extract (solid line) Lac1 fraction(dashed line) and Lac2 fraction (dashed and dotted line)doi 101371journalpone0073721g007
Substrate source for laccase production showed to be one ofthe most important factors for laccase production (Table 2)Cultures with coffee husk as substrate presented the highestlaccase production levels The observed differences in laccaseactivity for each substrate could be owed to their chemicalcomposition Coffea arabica is the coffee variety raised inColombia and its husk has a reported composition of 2308cellulose (2076 glucose and 183 cellobiose) 2385hemicellulose (1356 xylose 523 arabinose 256 aceticacid and 195 glucuronic acid) 2828 total lignin and071 ashes [27] In addition coffee husk contents 13caffeine 45 tannins and 124 pectins [28] On the otherhand Glycine max is the soybean species cultured inColombia and its pod husk composition varies from 504 to62 crude protein 608 to 143 hemicellulose 3715 to4208 acid detergent fiber 4436 to 6015 neutral detergentfiber 850 to 893 acid detergent lignin and 2074 to 2106uronic acid [29ndash31] Finally for cedar sawdust the majorcomponents are glucose (426-444) xylose (46-195)cellulose (383-410) klason lignin (227-278) and acidlignin (40-43) [32ndash34] Although lignin is one of the mostimportant compounds associated with laccase production bywhite-rot fungi we did not observe a clear correlation betweenlignin composition percentage and laccase activity In thatsense the high laccase activity observed for the cultures thatuse coffee husk could be associated with its content of caffeineand tannins which have been identified as potent inducers oflaccase gene expression [173536] In fact caffeine has beenused as inducer for laccase production by Rhizoctonia solaniobtaining an increase of 41-fold in laccase activity [37] Similarresults were also reported for the production of Coloriopsisgallica laccase [38]
Laccase gene expression is mainly up regulated by Cu2+although Mn2+ Fe3+ heavy metals 26-dimethoxy-14-benzoquinone H2O2 amphotericin B syringic acid tannic acidTween 80 soybean oil aromatic compounds andmicroclimatic changes (ie lower temperature and osmoticpressure) are also recognized as potent inducers [173536] Ingeneral Cu2+ supplementation increased the laccase activity inboth SmC and SSC However Cu2+concentration required toobtain the highest laccase activity varied depending on agro-industrial waste These differences can be attributed to thecopper presented in the agro-industrial waste and the copperavailability in the medium Copper can be found in amountsthat range from 156 to 180 mg kg-1 of coffee husk [39] andfrom 26 to 67 mg kg-1 of soybean pod husk (or soybean hull)[3940] Despite that no reports indicate the copper content inuntreated cedar wood an average of 10 mg kg-1 of wood hasbeen reported for wood used as biomass in six different Europecountries [41] This allows to consider the cedar sawdust as thesubstrate with the lowest content of copper On the other handcopper adsorption by sawdust and coffee husk has beenreported obtaining a Langmuir based maximum adsorptioncapacity (Qmax mg g-1 adsorbent) of 845 and 75 respectively[4243] Although no adsorption studies has been reportedusing untreated soybean pod husk Šćiban et al [44] reporteda Qmax for copper adsorption of 0085 mmol g-1 adsorbent (~ 54mg g-1) using treated soybean straws Based on this
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 10 September 2013 | Volume 8 | Issue 9 | e73721
information it can be expected that a higher amount ofexogenous copper would be required for laccase induction oncultures with cedar sawdust than with soybean pod husk andcoffee husk due to its lower content of copper In addition dueto the high copper-adsorption capacity of sawdust the amountof available copper in the culture would be reduced allowingthe laccase production at higher copper concentrations withoutreaching levels that might be toxic for the fungus This fact canbe observed in the SSC (Table 4) However for the SmC thisbehavior is not maintained for the cultures with soybean podhusk and coffee husk (Table 4) where the maximum laccaseactivity was obtained with a copper concentration of 05 and 5mM respectively This could be owed to a lower contactbetween the fungus and the substrate in the SmC whencompared to the SSC which is close related to the transport ofnutrients presented in the substrates
Production of extracellular laccases has been reported in alarge number of species of white-rot fungi strains grown onnatural substrates (Table 1) Although a laccase activity profileis not clearly observed based on the agro-industrial wasteculture mode and fungus strain most cases in which SmC andSSC were conducted using the same substrate and fungusstrain showed a higher laccase activity under SmC conditionsIn this study was observed a substrate dependentperformance where the most favorable condition to obtain thehighest laccase activity was the SSC The laccase activitiesobtained in the present study are comparable and in somecases higher than those reported for Pleurotus ostreatusPhanerochaete chrysosporium Pycnoporus cinnabarinus Tversicolor and Ganoderma sp cultured under SC and SmCconditions and using different agro-industrial wastes (Table 1)Semi-solid cultures using different agro-industrial wastes havebeen reported using T versicolor Phlebia radiata Tpubescens P ostreatus and C unicolor Rodriguez et al [16]screened different supports for laccase production by Tversicolor under SSC They obtained the highest laccaseactivity (1200 U L-1) when in the culture medium was presentedbarley bran In addition they were able to obtain a laccaseactivity of 1700 U L-1 when the culture was supplemented withxylidine as an inducer These results were obtained after 17 or18 days of culture and ABTS was used to determine theenzyme activity Maumlkelauml et al [4546] have studied theexpression of the genes involved in laccase production Pr-lac1and Pr-lac2 presented in P radiata and evaluate laccaseproduction in SSC using milled alder wood (Alnus incana) assubstrate They reported a maximum laccase activity of 3 microkatL-1 (~180 U L-1) at the 14th day of culture and usingsyringaldazine for its detection Osma et al [15] have studiedthe morphological changes and laccase production of strains Tpubescens P ostreatus C unicolor and T versicolor grownon wheat bran flakes using SSC They obtained a maximumlaccase production between 1397 and 2778 U L-1 after 10 and13 days of culture This enzyme activity is strain dependentand ABTS was used for its quantification Our results show agood correlation with reported data The highest laccaseactivity was obtained when T pubescens was grown on coffeehusk under SSC without copper (204 U L-1 evaluated withsyringaldazine or 1549 U L-1 evaluated with ABTS) which is in
some cases higher than reported ones This shows that underthe studied conditions the production of the laccase crudeextract is favored in SSC using coffee husk without the additionof copper which could favors the cost of the productionprocess
It has been reported for white rot-fungi the production ofmultiple laccase isoenzymes which are encoded by differentgenes and show different expression profiles stabilitiessubstrate affinities and molecular masses (ranging from 35 to140 kDa) [3647ndash49] Two isoforms have been identified in Tpubescens strains two monomeric (Lac1 and Lac2) enzymesof 67 kDa but with different pI (53 and 51) [50] and twolaccase-activity fractions (Lap1 and Lap2) with different pI (gt30 and 26) from which Lap2 is a monomeric enzyme of 65kDa [17] In this study we identify two laccase isoenzymesfrom T pubescens CBS 69694 that have different molecularmasses (120 and 60 kDa) and pI The purifications resultssuggest a more acidic pI for Lac1 (le 4) than Lac2 (ge 6) whichdiffers from a previous report for this fungus in which twofractions with laccase activity were obtained Lap1 and Lap2with pI gt 3 and of 26 respectively [17] Since in both casessimilar purification protocols were used and Lac1 and Lac2were recovered from the unbound and bound fractionsrespectively as Lap1 and Lap2 [17] this difference could bedue to the culture conditions used in each case (ie thepresence of agro-industrial wastes) Nevertheless the nanoLC-MSMS analysis showed that peptides of Lac1 and Lac2 werepresented in Lap1 (UniProtKBSwiss-Prot Q8TG93) and Lap2(UniProtKBSwiss-Prot Q8TG94) which have a predicted pI of43 and 47 respectively Lac1 and Lac2 peptides werepresented in Laccase-2 (UniProtKBSwiss-Prot Q99046) andLaccase-1 (UniProtKBSwiss-Prot Q99044) from T villosarespectively with 80 to 100 identity These T villosa laccaseisoenzymes have a predicted pI of 47 (laccase-1) and 60(laccase-2) and a 77 identity among them In addition Tvillosa laccase-2 is reported as a homodimer protein whilelaccase-1 is reported as monomeric protein Alignment of Lac1-Lap2 and Lac2-Lap1 from T pubescens CBS 69694 and Tvillosa showed a 96 and 70 identity respectively Finallysimilar peptides were identified for Lac1 and Lac2 from Tpubescens (Schumach) Pilaacutet (Syn Coriolus pubescens(Schum ex Fr) Queacutel) BCB 923-2 (Figure 3b) [51] Theseresults suggest that the purified fractions correspond to laccaseisoenzymes that differ from previously reported ones for this Tpubescens strain
As mentioned above laccases isoenzymes present differentstabilities and substrate affinities This behavior was alsoobserved for the laccase isoenzymes purified from Tpubescens with Lac1 showing lower laccase activity thanLac2 with ABTS as substrate However the difference inactivity between these isoenzymes depended on the agro-industrial waste used for the T pubescens growth suggestinga probable effect of the substrate composition on the laccasephysicochemical properties In addition it was observed thatthe total laccase activity presented in the UF retentates ishigher than the laccase activity of the purified fractionsregardless of the agro-industrial byproduct used for the Tpubescens growth This suggests a synergistic effect of Lac1
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 11 September 2013 | Volume 8 | Issue 9 | e73721
and Lac2 as have been reported for laccases obtained fromTrametes and Pleurotus strains [175253]
The maximum relative activity for the UF retentate waspresented at a pH from 20 to 40 regardless of the agro-industrial byproduct with a monotonical decrease at pHs abovethis range The relation between pH and type of electron donormolecule explains this behavior on laccase activity It has beenreported that when the substrate is an organic hydrogen donorie phenols the optimal pH ranges from 35 to 60 while thelaccase activity monotonically decreases from pH 25 to 70 ifthe substrate is an electron donor like ABTS [54] Despite thatat different pHs most of the purified laccase fractionspresented a similar performance as the UF retentates the Lac2fraction obtained from cedar sawdust differed from the previousobservation For this fraction the maximum activity wasobserved at pH 50 and despite that at acidic pH (ge 30) therelative activity was higher than 80 at lower pHs (20) itdecreased almost in a 50 Nevertheless at basic pHs therelative laccase activity presented a sharp decreased that iswell correlate with the other laccase fractions We are not ableto attribute this particular trend to the presence of multiplelaccase isoenzymes in the fraction since the SDS-PAGErevealed only one band when stained with ABTS and from theNanoLC-MSMS analysis no difference in the number ofpeptides and their sequence was detected We speculate thatthis particular behavior can be owed to a compound added inthe pre-treatment of the cedar wood that might be used forpreserving it that might affect the activity of Lac2 and that wasnot completely removed during the purification
Temperature stability evaluation showed that Lac1 is themost thermolabile fraction while the laccase activity reductionin the crude extracts did not seem to be the result of anadditive effect of the activity loss of the purified fractionsAlthough there are no reports of laccase stability under theselected conditions these results agree with previous reportsthat state that fungal laccases rapidly decrease their activity attemperatures near 60 degC and thermal stability correlates withthe temperature range of the growth of the source organism[55] In addition we previously reported for a laccase crudeextract obtained from culturing T pubescens on coffee husk anactivity loss of 26 and 10 at 60 and 50 degC respectivelyafter 8 h at pH 60 using syringaldazine as substrate [48]
Finally an electrochemical characterization was carried outfor selected crude extracts and purified fractions The results
showed a difference between the electrochemicalperformances of extracts and purified fractions withsyringaldazine or ABTS These differences could be owed tothe different mechanisms for electron transfer as discussedabove However the overall performance of the obtained cyclicvoltammograms for ABTS and syringaldazine agrees withresults reported by Fernaacutendez-Saacutenchez et al [25] who used asaturated calomel reference electrode
Conclusions
We have shown the potential of coffee husk soybean podhusk and cedar sawdust as substrates for the laccaseproduction In several countries like Brazil and Colombia theproduction of coffee and soybean is of great importance andthe generated volume of residues is high The results showedthat semi-solid culture of T pubescens using coffee huskallows to obtain an important amount of laccase which couldbe favored by the phenolic-compounds ie tannins presentedin coffee husk While supplementation of the culture mediumwith copper favored the production of laccase in submergedand semi-solid culture using soybean pod husk and cedarsawdust semi-solid culture using coffee husk which presentedthe highest laccase volumetric yield did not require coppersupplementation This is an economic advantage that shouldbe considered when scaling up the process In this study Tpubescens showed its ability to produce two independentlaccases regardless of the used substrate However a higherlaccase activity was observed for crude extracts rather than forpurified fractions This suggests that Lac1 and Lac2 might havea synergistic effect on the final laccase activity rather than anadditive effect Based on this study the investigation of laccaseproduction by evaluating other agro-industrial wastes in whichphenolic-compounds constitute an important percentage oftheir composition is of great interest to avoid the use ofinducers that would increase the production cost
Author Contributions
Conceived and designed the experiments JFO CJAD OFSPerformed the experiments JCG SCM AR Analyzed the dataJCG SCM AR JFO CJAD OFS Wrote the manuscript AR JFOCJAD OFS
References
1 Vandamme EJ (2009) Agro-industrial residue utilization for industrialbiotechnology products In P NigamA Pandey Biotechnology for Agro-Industrial Residues Utilisation Netherlands Springer Verlag pp 3-11
2 Martiacutenez AT Speranza M Ruiz-Duentildeas FJ Ferreira P Camarero S etal (2005) Biodegradation of lignocellulosics microbial chemical andenzymatic aspects of the fungal attack of lignin Int Microbiol 8195-204 PubMed 16200498
3 Saacutenchez O Sierra R Almeacuteciga-Diacuteaz CJ (2011) Delignification processof agro-industrial aastes an alternative to obtain fermentablecarbohydrates for producing fuel In M Manzanera Alternative FuelIntech 111-154
4 Wesenberg D Kyriakides I Agathos SN (2003) White-rot fungi andtheir enzymes for the treatment of industrial dye effluents BiotechnolAdv 22 161-187 doi101016jbiotechadv200308011 PubMed14623049
5 Morozova OV Shumakovich GP Shleev SV Yaropolov YI (2007)Laccase-mediator systems and their applications A review ApplBiochem Microbiol 43 523-535 doi101134S0003683807050055
6 Baldrian P (2006) Fungal laccases - occurrence and properties FEMSMicrobiol Rev 30 215-242 doi101111j1574-4976200500010xPubMed 16472305
7 Rodriacuteguez Couto S Herrera JLT (2006) Industrial and biotechnologicalapplications of laccases A review Biotechnol Adv 24 500-513 doi101016jbiotechadv200604003 PubMed 16716556
8 Osma JF Toca-Herrera JL Rodriacuteguez-Couto S (2010) Uses oflaccases in the food industry Enzyme Res 2010 2010 918761PubMed 21048873
9 Rodriguez S (2008) Exploitation of biological wastes for the productionof value-added products under solid-state fermentation conditionsBiotechnol J 3 859-870 doi101002biot200800031 PubMed18543242
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 12 September 2013 | Volume 8 | Issue 9 | e73721
10 Osma JF Toca-Herrera JL Rodriacuteguez-Couto S (2011) Cost analysis inlaccase production J Environ Manage 92 2907-2912 doi101016jjenvman201106052 PubMed 21775046
11 Jain A Morlok CK Henson JM (2013) Comparison of solid-state andsubmerged-state fermentation for the bioprocessing of switchgrass toethanol and acetate by Clostridium phytofermentans Appl MicrobiolBiotechnol 97 905-917 doi101007s00253-012-4511-4 PubMed23111595
12 Singhania RR Patel AK Soccol CR Pandey A (2009) Recentadvances in solid-state fermentation Biochem Eng J 44 13-18 doi101016jbej200810019
13 Economou CN Makri A Aggelis G Pavlou S Vayenas DV (2010)Semi-solid state fermentation of sweet sorghum for the biotechnologicalproduction of single cell oil Bioresour Technol 101 1385-1388 doi101016jbiortech200909028 PubMed 19781936
14 Rodriacuteguez CS Santoro R Cameselle C Sanroman A (1997) Laccaseproduction in semi-solid cultures of Phanerochaete chrysosporiumBiotechnol Lett 19 995-998 doi101023A1018495216946
15 Osma JF Moilanen U Toca-Herrera JL Rodriacuteguez-Couto S (2011)Morphology and laccase production of white-rot fungi grown on wheatbran flakes under semi-solid-state fermentation conditions FEMSMicrobiol Lett 318 27-34 doi101111j1574-6968201102234xPubMed 21291496
16 Couto SR Gundin M Lorenzo M Sanroman MN (2002) Screening ofsupports and inducers for laccase production by Trametes versicolor insemi-solid-state conditions Proc Biochem 38 249-255 doi101016S0032-9592(02)00087-0
17 Galhaup C Goller S Peterbauer CK Strauss J Haltrich D (2002)Characterization of the major laccase isoenzyme from Trametespubescens and regulation of its synthesis by metal ions Microbiology148 2159-2169 PubMed 12101303
18 Sievers F Wilm A Dineen D Gibson TJ Karplus K et al (2011) Fastscalable generation of high-quality protein multiple sequencealignments using Clustal Omega Mol Syst Biol 7 539 PubMed21988835
19 Childs RE Bardsley WG (1975) The steady-state kinetics of peroxidasewith 22-azino-di-(3-ethyl-benzthiazoline-6-sulphonic acid) aschromogen Biochem J 145 93-103 PubMed 1191252
20 Harkin JM Obst JR (1973) Syringaldazine an Effective Reagent forDetecting Laccase and Peroxidase in Fungi Experientia 29 382-387
21 Nikupaavola ML Raaska L Itavaara M (1990) Detection of white-rotfungi by a non-roxic stain Mycol Res 94 27-31 doi101016S0953-7562(09)81260-4
22 Gonzalez LF Sarria V Sanchez OF (2010) Degradation ofchlorophenols by sequential biological-advanced oxidative processusing Trametes pubescens and TiO(2)UV Bioresour Technol 1013493-3499
23 Waterborg JH (2009) The Lowry method for protein quantitation In JWalker The Protein Protocols Handbook Humana Press pp 7-10
24 Harkin JM Obst JR (1973) Syringaldazine an effective reagent fordetecting laccase and peroxidase in fungi Cell Mol Life Sci 29381-387 doi101007BF01926734
25 Fernaacutendez-Saacutenchez C Tzanov T Guumlbitz GM Cavaco-Paulo A (2002)Voltammetric monitoring of laccase-catalysed mediated reactionsBioelectrochemistry 58 149-156 doi101016S1567-5394(02)00119-6PubMed 12414320
26 Bourbonnais R Leech D Paice MG (1998) Electrochemical analysis ofthe interactions of laccase mediators with lignin model compoundsBiochim Biophys Acta 1379 381ndash390 doi101016S0304-4165(97)00117-7 PubMed 9545600
27 Ferraz F Silva S (2009) Characterization of coffee husk biomass forbiotechnological purposes N Biotechnol 25 S256-S256 doi101016jnbt200906573
28 Pandey A Soccol CR Nigam P Brand D Mohan R et al (2000)Biotechnological potential of coffee pulp and coffee husk forbioprocesses Biochem Eng J 6 153-162 doi101016S1369-703X(00)00084-X PubMed 10959086
29 Sruamsiri S Silman P (2008) Nutritive composition of soybean by-products and nutrient digestibility of soybean pod husk Maejo Int J SciTechnol 2 568-576
30 Romkaew J Nagaya Y Goto M Suzuki J Umezaki T (2008) Poddehiscence in relation to chemical components of pod shell in soybeanPlant Prod Sci 11 278-282 doi101626pps11278
31 Nakamanee G Polbumroong T Phaikaew C (2003) Alternative use ofcrop waste to increase milk production Proceedings of the 8th Meetingof the Regional Working Group on Grazing and Feed Resources forSoutheast Asia Kuala Lumpur Malaysia 66 - 70
32 Kurek B Gaudard F (2000) Oxidation of spruce wood sawdust byMnO(2) plus oxalate a biochemical investigation J Agric Food Chem48 3058-3062 doi101021jf000015g PubMed 10898665
33 Weil J Brewer M Hendrickson R Sarikaya A Ladisc MR (1998)Continuous pH monitoring during pretreatment of yellow poplar woodsawdust by pressure cooking in water Appl Biochem Biotechnol 70-7299-111 doi101007BF02920127
34 Weil J Sarikaya A Rau S-L Goetz J Ladisch CM et al (1997)Pretreatment of yellow poplar sawdust by pressure cooking in waterAppl Biochem Biotechnol 68 21-40 doi101007BF02785978
35 Dekker RF Barbosa AM Giese EC Godoy SD Covizzi LG (2007)Influence of nutrients on enhancing laccase production byBotryosphaeria rhodina MAMB-05 Int Microbiol 10 177-185 PubMed18075999
36 Xiao YZ Hong YZ Li JF Hang J Tong PG et al (2006) Cloning ofnovel laccase isozyme genes from Trametes sp AH28-2 and analysesof their differential expression Appl Microbiol Biotechnol 71 493-501doi101007s00253-005-0188-2 PubMed 16283298
37 Crowe JD Olsson S (2001) Induction of laccase activity in Rhizoctoniasolani by antagonistic Pseudomonas fluorescens strains and a range ofchemical treatments Appl Environ Microbiol 67 2088-2094 doi101128AEM6752088-20942001 PubMed 11319086
38 Yaguumle S Terroacuten MC Gonzaacutelez T Zapico E Bocchini P et al (2000)Biotreatment of tannin-rich beer-factory wastewater with white-rotbasidiomycete Coriolopsis gallica monitored by pyrolysisgaschromatographymass spectrometry Rapid Commun Mass Spectrom14 905-910 doi101002(SICI)1097-0231(20000530)1410 PubMed10825255
39 Gowda NKS Ramana JV Prasad CS Singh K (2004) Micronutrientcontent of certain tropical conventional and unconventional feedresources of southern India Trop Anim Heatlth Pro 36 77-94 doi101023BTROP0000009522309491d PubMed 14979561
40 DePeters EJ Fadel JG Arana MJ Ohanesian N Etchebarne MA et al(2000) Variability in the chemical composition of seventeen selectedby-product feedstuffs used by the California dairy industry Prof AnimSci 16 69-99
41 Obernberger I Thek G (2004) Physical characterisation and chemicalcomposition of densified biomass fuels with regard to their combustionbehaviour Biomass Bioenerg 27 653-669 doi101016jbiombioe200307006
42 Larous S Meniai A-H Lehocine MB (2005) Experimental study of theremoval of copper from aqueous solutions by adsorption usingsawdust Desalination 185 483ndash490 doi101016jdesal200503090
43 Oliveira WE Franca AS Oliveira LS Rocha SD (2008) Untreatedcoffee husks as biosorbents for the removal of heavy metals fromaqueous solutions J Hazard Mater 152 1073ndash1081 doi101016jjhazmat200707085 PubMed 17804159
44 Šciban M Klašnja M Škrbic B (2008) Adsorption of copper ions fromwater by modified agricultural by-products Desalination 229 170ndash180doi101016jdesal200708017
45 Maumlkelauml MR Hildeacuten KS Hakala TK Hatakka A Lundell TK (2006)Expression and molecular properties of a new laccase of the white rotfungus Phlebia radiata grown on wood Curr Genet 50 323-333 doi101007s00294-006-0090-1 PubMed 16927090
46 Maumlkelauml MR Lundell T Hatakka A Hildeacuten K (2013) Effect of coppernutrient nitrogen and wood-supplement on the production of lignin-modifying enzymes by the white-rot fungus Phlebia radiata Fungal Biol117 62-70 doi101016jfunbio201211006 PubMed 23332834
47 Dwivedi UN Singha P Pandeya VP Kumara A (2011) Structurendashfunction relationship among bacterial fungal and plant laccases J MolCatal B Enzym 68 117-128 doi101016jmolcatb201011002
48 Gaitan IJ Medina SC Gonzaacutelez JC Rodriacuteguez A Espejo AacuteJ et al(2011) Evaluation of toxicity and degradation of a chlorophenol mixtureby the laccase produced by Trametes pubescens Bioresour Technol102 3632ndash3635 doi101016jbiortech201011040 PubMed21115244
49 Karp SG Faraco V Amore A Birolo L Giangrande C et al (2012)Characterization of laccase isoforms produced by Pleurotus ostreatusin solid state fermentation of sugarcane bagasse Bioresour Technol114 735-739 doi101016jbiortech201203058 PubMed 22487128
50 Nikitina OV Shleev SV Gorshina ES Rusinova TV Serezhenkov VAet al (2005) Isolation and purification of enzymes from ligninolyticcomplex of the basidial fungus Trametes pubescens (Schumach) Pilatand study of their properties Biochemistry (Mosc) 70 1274-1279 doi101007s10541-005-0259-0
51 Shleev S Nikitina O Christenson A Reimann CT Yaropolov AI et al(2007) Characterization of two new multiforms of Trametes pubescenslaccase Bioorg Chem 35 35-49 doi101016jbioorg200608001PubMed 16989887
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 13 September 2013 | Volume 8 | Issue 9 | e73721
52 Xiao YZ Chen Q Hang J Shi YY Wu J et al (2004) Selectiveinduction purification and characterization of a laccase isozyme fromthe basidiomycete Trametes sp AH28-2 Mycologia 96 26-35 doi1023073761984 PubMed 21148825
53 Muntildeoz C Guilleacuten F Martiacutenez AT Martiacutenez MJ (1997) Laccaseisoenzymes of Pleurotus eryngii characterization catalytic propertiesand participation in activation of molecular oxygen and Mn2+ oxidationAppl Environ Microbiol 63 2166-2174 PubMed 9172335
54 Morozova OV Shumakovich GP Gorbacheva MA Shleev SVYaropolov AI (2007) Blue laccases Biochemistry (Mosc) 721136-1150 doi101134S0006297907100112
55 Hildeacuten K Hakala TK Lundell T (2009) Thermotolerant andthermostable laccases Biotechnol Lett 31 1117-1128 doi101007s10529-009-9998-0 PubMed 19360388
56 Verma P Madamwar D (2002) Production of ligninolytic enzymes fordye decolorization by cocultivation of white-rot fungi Pleurotusostreatus and Phanerochaete chrysosporium under solid-statefermentation Appl Biochem Biotechnol 102-103 109-118 doi101385ABAB102-1031-6109 PubMed 12396115
57 Meza JC Lomascolo A Casalot L Sigoillot J-C Auria R (2005)Laccase production by Pycnoporus cinnabarinus grown on sugar-canebagasse Influence of ethanol vapours as inducer Proc Biochem 403365ndash3371 doi101016jprocbio200503004
58 Hossain SM Anantharaman N (2006) Activity enhancement ofligninolytic enzymes of Trametes versicolor with bagasse powder Afr JBiotechnol 5 189-194
59 Revankar MS Desai KM Lele SS (2007) Solid-state fermentation forenhanced production of laccase using indigenously isolatedGanoderma sp Appl Biochem Biotechnol 143 16-26 doi101007s12010-007-0029-0 PubMed 18025593
60 Velazquez-Cedeno MA Mata G Savoie J-M (2002) Waste-reducingcultivation of Pleurotus ostreatus and Pleurotus pulmonarius on coffeepulp changes in the production of some lignocellulolytic enzymesWorld J Microbiol Biotechnol 18 201ndash207 doi101023A1014999616381
61 Mata G Murrieta D Andreu LI (2005) Changes in lignocellulolyticenzyme activities in six Pleurotus spp strains cultivated on coffee pulpin confrontation with Trichoderma spp World J Microbiol Biotechnol 21143ndash150 doi101007s11274-004-3041-3
62 de Souza DF Tychanowicz GK de Souza CG Peralta RM (2006) Co-production of ligninolytic enzymes by Pleurotus pulmonarius on wheatbran solid state cultures J Basic Microbiol 46 126-134 doi101002jobm200510014 PubMed 16598826
63 Prasad KK Mohan SV Bhaskar YV Ramanaiah SV Babu VL et al(2005) Laccase production using Pleurotus ostreatus 1804 immobilizedon PUF cubes in batch and packed bed reactors influence of cultureconditions J Microbiol 43 301-307 PubMed 15995650
64 Songulashvili G Elisashvili V Wasser SP Nevo E Hadar Y (2007)Basidiomycetes laccase and manganese peroxidase activity in
submerged fermentation of food industry wastes Enzyme MicrobTechnol 41 57ndash61 doi101016jenzmictec200611024
65 Sathishkumar P Murugesan K Palvannan T (2010) Production oflaccase from Pleurotus florida using agro-wastes and efficientdecolorization of Reactive blue 198 J Basic Microbiol 50 360-367 doi101002jobm200900407 PubMed 20586068
66 Moldes D Gallego PP Rodriacuteguez Couto S Sanromaacuten A (2003) Grapeseeds the best lignocellulosic waste to produce laccase by solid statecultures of Trametes hirsuta Biotechnol Lett 25 491-495 doi101023A1022660230653 PubMed 12882277
67 Elisashvili V Kachlishvili E Penninckx M (2008) Effect of growthsubstrate method of fermentation and nitrogen source onlignocellulose-degrading enzymes production by white-rotbasidiomycetes J Ind Microbiol Biotechnol 35 1531-1538 doi101007s10295-008-0454-2 PubMed 18716810
68 Osma JF Saravia V Herrera JL Couto SR (2007) Mandarin peelingsthe best carbon source to produce laccase by static cultures ofTrametes pubescens Chemosphere 67 1677-1680 doi101016jchemosphere200611051 PubMed 17234250
69 Kahraman SS Gurdal IH (2002) Effect of synthetic and natural culturemedia on laccase production by white rot fungi Bioresour Technol 82215-217 doi101016S0960-8524(01)00193-6 PubMed 11991068
70 Iandolo D Piscitelli A Sannia G Faraco V (2011) Enzyme Productionby Solid Substrate Fermentation of Pleurotus ostreatus and Trametesversicolor on Tomato Pomace Appl Biochem Biotechnol 163 40-51doi101007s12010-010-9014-0 PubMed 20582639
71 Moldes D Lorenzo M Sanromaacuten MA (2004) Different proportions oflaccase isoenzymes produced by submerged cultures of Trametesversicolor grown on lignocellulosic wastes Biotechnol Lett 26 327-330doi101023BBILE000001545240213bf PubMed 15055770
72 Philippoussis A Diamantopoulou P Papadopoulou K Lakhtar HRoussos S et al (2010) Biomass laccase and endoglucanaseproduction by Lentinula edodes during solid state fermentation of reedgrass bean stalks and wheat straw residues World J MicrobiolBiotechnol 27 285-297
73 Fenice M Giovannozzi Sermanni G Federici F DrsquoAnnibale A (2003)Submerged and solid-state production of laccase and Mn-peroxidaseby Panus tigrinus on olive mill wastewater-based media J Biotechnol100 77-85 doi101016S0168-1656(02)00241-9 PubMed 12413788
74 Rodriguez-Couto S GundIn M Lorenzo M Sanromaacuten MAacute (2002)Screening of supports and inducers for laccase production by Trametesversicolor in semi-solid-state conditions Proc Biochem 38 249-255doi101016S0032-9592(02)00087-0
75 Fackler K Gradinger C Schmutzer M Tavzes C Burgert I et al (2007)Biotechnological wood modification with selective white-rot fungi and itsmolecular mechanisms Food Technol Biotechnol 45 269ndash276
76 Levin L Herrmann C Papinutti VL (2008) Optimization oflignocellulolytic enzyme production by the white-rot fungus Trametestrogii in solid-state fermentation using response surface methodologyBiochem Eng J 39 207-214 doi101016jbej200709004
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 14 September 2013 | Volume 8 | Issue 9 | e73721

sawdust culture behaved different to the UF retentate fromcoffee husk and soybean pod husk cultures In this sense the
UF retentate from cedar sawdust culture presented the lowestenzyme activity at temperatures le 40 degC but at 70 degC
Figure 3 Aligment of reported lacasses sequences and Lac1 and Lac2 peptides (a) Alignment of Lap1 (UniProtKBSwiss-Prot Q8TG93) and Lap2 (UniProtKBSwiss-Prot Q8TG94) from T pubescens CBS 69694 (b) Alignment of Lac1 and Lac2 peptideswith Lap1 and Lap2 respectively and tryptic peptides described by Shleev et al 2007 Alignments were carried out using ClustalOmega and figures were using CLC Sequence Viewer Different residues are colored in reddoi 101371journalpone0073721g003
Table 3 Laccase activity during the purification of crude extracts resulted from T pubescens culture on agro-industrialwastes under submerged (SmC) and semisolid (SSC) cultures
Soybean pod husk Coffee husk Cedar sawdust
Protein(mg mL-1)
VolumetricActivity (UL-1)
SpecificActivity (Umg-1
protein)Recovery()
Protein(mg mL-1)
VolumetricActivity (UL-1)
SpecificActivity (Umg-1
protein)Recovery()
Protein(mg mL-1)
VolumetricActivity (UL-1)
SpecificActivity (Umg-1
protein)Recover()
SmCCrudeExtract
0869 52888 0608 100 0414 152708 3688 100 0431 10735 0249 100
045 micromFiltration
0736 46549 0632 88 0334 117892 3529 77 0444 12053 0271 112
UF retentate 1010 78731 0779 74 0378 195263 5165 64 0510 25905 0507 120
Lac1 0494 5471 0110 1 0232 2399 0103 02 0222 838 0037 08
Lac2 0310 20661 0666 4 0116 107240 9252 7 0040 15234 3837 14 SSCCrudeExtract
1146 164966 1439 100 0596 154907 2601 100 1065 23203 0217 100
045 micromFiltration
0776 128392 1654 77 0474 115736 2439 75 0806 25824 0320 111
UF retentate 0973 335055 3443 101 0428 161750 3779 52 1134 51101 0450 110
Lac1 0097 8854 0912 2 0104 1514 0145 03 0042 1315 0313 2
Lac2 0168 48988 2915 10 0131 82298 6282 18 0072 16162 2244 23
740 ABTS was used to determine laccase activity
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 7 September 2013 | Volume 8 | Issue 9 | e73721
presented the highest enzyme activity ~90 of its initial valuein comparison to the other two UF retentates (Figure 4)
The temperature and pH effect on laccase activity for Lac1and Lac2 is presented in Figure 5 The maximum relativeactivity for both fractions was presented at a pH from 20 to 40regardless of the agro-industrial waste as observed for the UFretentate However at pH gt 40 Lac1 showed a sharperdecrease in its activity than that observed for Lac2Nevertheless Lac2 purified from cedar sawdust culturepresented a different performance This fraction showed themaximum activity at pH 50 while a sharp drop was observedat pHs above or below this value
Laccase activity profiles showed that Lac1 and Lac2presented the maximum enzyme activity at 55 and 60 degCrespectively (Table 4) It was observed that Lac1 from coffeehusk culture presented a lower decrease in enzyme activity attemperatures le 50 degC or ge 60 degC than Lac2 On the contraryLac2 from soybean husk or cedar sawdust cultures presenteda higher relative laccase activity than Lac1 at temperatures le55 degC The purified fractions from soybean pod husk and cedarsawdust cultures presented the lowest relative activity at 30 degCand 65 degC
Table 4 Optimum temperatures and pHs for maximumlaccase activities in UF retentate Lac1 and Lac2 obtainedfrom semisolid cultured T pubescens on agro-industrialwastes
EnzymaticFraction Temperature pH
VolumetricActivity (UL-1)
SpecificActivity (Umg-1 protein)
VolumetricActivity (UL-1)
SpecificActivity (Umg-1
protein)SoybeanPod Husk +2 mM Cu+2
148000plusmn1178(55)
1521plusmn0012182037plusmn3025(2-3)
1871plusmn0031
Lac122925plusmn917(60)
2363plusmn009536135plusmn831(23)
3725plusmn0085
Lac232638plusmn1558(55)
1942plusmn009228713plusmn274(23)
1709plusmn0016
CoffeeHusk + 0mM Cu+2
113740plusmn8289(50-55)
3008plusmn0219290666plusmn1039(2-3)
7689plusmn0275
Lac1 889plusmn039 (55) 0038plusmn0002 1779plusmn028 (23) 0077plusmn0001
Lac234194plusmn2789(60)
2947plusmn024033150plusmn526 (2ndash4)
2857plusmn0045
CedarSawdust +5 mM Cu+2
257666plusmn1201(55)
5052plusmn0024344634plusmn1043(2-3)
6757plusmn0204
Lac121370plusmn472(55)
0962plusmn002129117plusmn943(23)
1311plusmn0042
Lac255166plusmn1656(60)
1379plusmn0414 55166plusmn1656 (5) 1379plusmn0414
ABTS was used to determine laccase activity
Laccase stabilityFor selected enzymatic extracts and purified fractions (Table
4) a stability test for 48 h at their optimal pH and temperaturewas conducted Time course of the relative laccase activity forcrude extract and purified fractions from coffee husk and cedarsawdust cultures are presented in Figure 5 Similar resultswere observed for crude extract and purified fractions fromsoybean pod husk culture (data no shown)
It was observed that the activity loss depends on thesubstrate source used to grow T pubescens An activity loss of55 was observed for crude extracts and purified fractionsfrom coffee husk and soybean pod husk cultures while anactivity loss of 70 was observed for the crude extract andpurified fractions from cedar sawdust culture NeverthelessLac2 showed the lowest activity reduction with values of 10 40and 55 for coffee husk soybean pod husk and cedarsawdust respectively while Lac1 presented an activity loss of34 50 and 60 for coffee husk soybean pod husk and cedarsawdust respectively Despite the differences in laccaseactivity among agro-industrial wastes and between crudeextracts and purified fractions all the profiles showed a sharp
Figure 4 Effect of temperature (a) and pH (b) on laccaseactivity for the UF retentate obtained by semisolid cultureof T pubescens on coffee husk () soybean pod husk (loz)and cedar sawdust (Δ) Laccase stability was evaluated byusing ABTS All the assays were performed in triplicatedoi 101371journalpone0073721g004
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 8 September 2013 | Volume 8 | Issue 9 | e73721
decrease during the first 8 h while a reduced activity loss wasobserved during the last 24 h of the assay (Figure 6)
Laccase electrochemical characterizationFor selected crude extracts and purified fractions a cyclic
voltammetry characterization was conducted For eachmediator syringaldazine or ABTS similar cyclic
Figure 5 Effect of temperature () and pH () on enzymeactivity for Lac1 (solid line) and Lac2 (dashed line)obtained after the purification of the enzymatic crudeextract produced by semisolid culture of T pubescens oncoffee husk (a) soybean pod husk (b) and cedar sawdust(c) Laccase stability was evaluated by using ABTS All theassays were performed in triplicatedoi 101371journalpone0073721g005
voltammograms were obtained for the different crude extractsand purified fractions Voltammograms of the redox reaction forsyringaldazine (Figure 7a) and ABTS (Figure 6b) arepresented Two anionic and cationic peaks were observedwhen syringaldazine was used as mediator The first pair ofpeaks showed a reversible performance with a mean Ia Icratio of 109plusmn006 106plusmn005 and 098plusmn002 for the crudeextracts Lac1 and Lac2 respectively The second pair ofpeaks showed quasi-reversible performance and the cathodicpeak was presented at a potential of +062 V (vs Ag|AgCl)while the anodic peak was presented at a potential of +060 V(vs Ag|AgCl) No significant difference on the reductionpotential (Eo) for syringaldazine was observed for the crudeextracts or the purified fractions obtaining an Eo of0314plusmn0008 V (vs Ag|AgCl) However the peak potentialseparation (ΔEp) showed a different profile depending on thesubstrate used for the culture While not significant difference(p gt 005) on ΔEp value was observed between crude extract(0045plusmn0005 V vs Ag|AgCl) Lac1 (0046plusmn0005 V vs Ag|AgCl)and Lac2 (0028plusmn0001 V vs Ag|AgCl) from cultures with cedarsawdust and soybean pod husk the ΔEp was considerablyhigher for the crude extract (0083plusmn0004 V vs Ag|AgCl) Lac1(0068plusmn0003 V vs Ag|AgCl) and Lac2 (0051plusmn0005 V vs Ag|AgCl) from the coffee husk culture
For reactions conducted with ABTS and the different crudeextracts and purified fractions the voltammograms exhibitedone pair of cathodic and anodic peaks (Figure 7b) whichcorresponds to the redox of ABTSABTS+ (IaIc) with an Eo of0487plusmn0006 V (vs Ag|AgCl) This process was reversiblepresenting a mean Ia Ic ratio of 099plusmn002 In addition asecond cathodic peak was presented at +083 V (vs Ag|AgCl)when ABTS redox reaction was conducted using the crudeextract and the purified fractions This performance agrees withprevious reports in which the laccase activity cannot beobserved at potential values of the second reversible oxidation
Figure 6 Laccase stability profiles at optimal pH andtemperature conditions for crude extracts (solid line) Lac1(dashed line) and Lac2 (spotted line) obtained fromsemisolid culture of T pubescens on coffee husk (loz) andcedar sawdust (bull) Laccase stability was evaluated by usingABTS All the assays were performed in triplicatedoi 101371journalpone0073721g006
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 9 September 2013 | Volume 8 | Issue 9 | e73721
of ABTS ABTS+ABTS2+ due to this catalytic process cannotbe detected by cyclic voltammetry [2526] Neverthelessvoltammograms for laccase crude extract from a soybean podhusk culture presented the second anodic peak which inaddition to the second cathodic peak corresponds to the redoxof ABTS+ABTS2+ (IIaIIc) with an Eo of 0942plusmn0004 V (vs Ag|AgCl) Contrary to the reactions conducted usingsyringaldazine as substrate the ΔEp did not present asignificant difference for the crude extracts and purifiedfractions 0248plusmn0016 V (vs Ag|AgCl)
Discussion
In this study we evaluated the effect of the culture mode ieSmC and SSC on laccase production by T pubescens CBS69694 using three different agro-industrial wastes (coffeehusk soybean pod husk and cedar sawdust) Furthermore theeffect of Cu+2 supplementation on laccase production was alsoevaluated and the crude extracts and the purified fractionswere partially characterized
Figure 7 Cyclic voltammograms of (a) syringaldazine atpH 50 using the crude extract and purified laccasefractions from semisolid culture of T pubescens on coffeehusk or (b) ABTS at pH 30 using the crude extract andpurified laccase fractions from semisolid culture of Tpubescens on soybean pod husk Nomenclature onlymediator (dotted line) crude extract (solid line) Lac1 fraction(dashed line) and Lac2 fraction (dashed and dotted line)doi 101371journalpone0073721g007
Substrate source for laccase production showed to be one ofthe most important factors for laccase production (Table 2)Cultures with coffee husk as substrate presented the highestlaccase production levels The observed differences in laccaseactivity for each substrate could be owed to their chemicalcomposition Coffea arabica is the coffee variety raised inColombia and its husk has a reported composition of 2308cellulose (2076 glucose and 183 cellobiose) 2385hemicellulose (1356 xylose 523 arabinose 256 aceticacid and 195 glucuronic acid) 2828 total lignin and071 ashes [27] In addition coffee husk contents 13caffeine 45 tannins and 124 pectins [28] On the otherhand Glycine max is the soybean species cultured inColombia and its pod husk composition varies from 504 to62 crude protein 608 to 143 hemicellulose 3715 to4208 acid detergent fiber 4436 to 6015 neutral detergentfiber 850 to 893 acid detergent lignin and 2074 to 2106uronic acid [29ndash31] Finally for cedar sawdust the majorcomponents are glucose (426-444) xylose (46-195)cellulose (383-410) klason lignin (227-278) and acidlignin (40-43) [32ndash34] Although lignin is one of the mostimportant compounds associated with laccase production bywhite-rot fungi we did not observe a clear correlation betweenlignin composition percentage and laccase activity In thatsense the high laccase activity observed for the cultures thatuse coffee husk could be associated with its content of caffeineand tannins which have been identified as potent inducers oflaccase gene expression [173536] In fact caffeine has beenused as inducer for laccase production by Rhizoctonia solaniobtaining an increase of 41-fold in laccase activity [37] Similarresults were also reported for the production of Coloriopsisgallica laccase [38]
Laccase gene expression is mainly up regulated by Cu2+although Mn2+ Fe3+ heavy metals 26-dimethoxy-14-benzoquinone H2O2 amphotericin B syringic acid tannic acidTween 80 soybean oil aromatic compounds andmicroclimatic changes (ie lower temperature and osmoticpressure) are also recognized as potent inducers [173536] Ingeneral Cu2+ supplementation increased the laccase activity inboth SmC and SSC However Cu2+concentration required toobtain the highest laccase activity varied depending on agro-industrial waste These differences can be attributed to thecopper presented in the agro-industrial waste and the copperavailability in the medium Copper can be found in amountsthat range from 156 to 180 mg kg-1 of coffee husk [39] andfrom 26 to 67 mg kg-1 of soybean pod husk (or soybean hull)[3940] Despite that no reports indicate the copper content inuntreated cedar wood an average of 10 mg kg-1 of wood hasbeen reported for wood used as biomass in six different Europecountries [41] This allows to consider the cedar sawdust as thesubstrate with the lowest content of copper On the other handcopper adsorption by sawdust and coffee husk has beenreported obtaining a Langmuir based maximum adsorptioncapacity (Qmax mg g-1 adsorbent) of 845 and 75 respectively[4243] Although no adsorption studies has been reportedusing untreated soybean pod husk Šćiban et al [44] reporteda Qmax for copper adsorption of 0085 mmol g-1 adsorbent (~ 54mg g-1) using treated soybean straws Based on this
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 10 September 2013 | Volume 8 | Issue 9 | e73721
information it can be expected that a higher amount ofexogenous copper would be required for laccase induction oncultures with cedar sawdust than with soybean pod husk andcoffee husk due to its lower content of copper In addition dueto the high copper-adsorption capacity of sawdust the amountof available copper in the culture would be reduced allowingthe laccase production at higher copper concentrations withoutreaching levels that might be toxic for the fungus This fact canbe observed in the SSC (Table 4) However for the SmC thisbehavior is not maintained for the cultures with soybean podhusk and coffee husk (Table 4) where the maximum laccaseactivity was obtained with a copper concentration of 05 and 5mM respectively This could be owed to a lower contactbetween the fungus and the substrate in the SmC whencompared to the SSC which is close related to the transport ofnutrients presented in the substrates
Production of extracellular laccases has been reported in alarge number of species of white-rot fungi strains grown onnatural substrates (Table 1) Although a laccase activity profileis not clearly observed based on the agro-industrial wasteculture mode and fungus strain most cases in which SmC andSSC were conducted using the same substrate and fungusstrain showed a higher laccase activity under SmC conditionsIn this study was observed a substrate dependentperformance where the most favorable condition to obtain thehighest laccase activity was the SSC The laccase activitiesobtained in the present study are comparable and in somecases higher than those reported for Pleurotus ostreatusPhanerochaete chrysosporium Pycnoporus cinnabarinus Tversicolor and Ganoderma sp cultured under SC and SmCconditions and using different agro-industrial wastes (Table 1)Semi-solid cultures using different agro-industrial wastes havebeen reported using T versicolor Phlebia radiata Tpubescens P ostreatus and C unicolor Rodriguez et al [16]screened different supports for laccase production by Tversicolor under SSC They obtained the highest laccaseactivity (1200 U L-1) when in the culture medium was presentedbarley bran In addition they were able to obtain a laccaseactivity of 1700 U L-1 when the culture was supplemented withxylidine as an inducer These results were obtained after 17 or18 days of culture and ABTS was used to determine theenzyme activity Maumlkelauml et al [4546] have studied theexpression of the genes involved in laccase production Pr-lac1and Pr-lac2 presented in P radiata and evaluate laccaseproduction in SSC using milled alder wood (Alnus incana) assubstrate They reported a maximum laccase activity of 3 microkatL-1 (~180 U L-1) at the 14th day of culture and usingsyringaldazine for its detection Osma et al [15] have studiedthe morphological changes and laccase production of strains Tpubescens P ostreatus C unicolor and T versicolor grownon wheat bran flakes using SSC They obtained a maximumlaccase production between 1397 and 2778 U L-1 after 10 and13 days of culture This enzyme activity is strain dependentand ABTS was used for its quantification Our results show agood correlation with reported data The highest laccaseactivity was obtained when T pubescens was grown on coffeehusk under SSC without copper (204 U L-1 evaluated withsyringaldazine or 1549 U L-1 evaluated with ABTS) which is in
some cases higher than reported ones This shows that underthe studied conditions the production of the laccase crudeextract is favored in SSC using coffee husk without the additionof copper which could favors the cost of the productionprocess
It has been reported for white rot-fungi the production ofmultiple laccase isoenzymes which are encoded by differentgenes and show different expression profiles stabilitiessubstrate affinities and molecular masses (ranging from 35 to140 kDa) [3647ndash49] Two isoforms have been identified in Tpubescens strains two monomeric (Lac1 and Lac2) enzymesof 67 kDa but with different pI (53 and 51) [50] and twolaccase-activity fractions (Lap1 and Lap2) with different pI (gt30 and 26) from which Lap2 is a monomeric enzyme of 65kDa [17] In this study we identify two laccase isoenzymesfrom T pubescens CBS 69694 that have different molecularmasses (120 and 60 kDa) and pI The purifications resultssuggest a more acidic pI for Lac1 (le 4) than Lac2 (ge 6) whichdiffers from a previous report for this fungus in which twofractions with laccase activity were obtained Lap1 and Lap2with pI gt 3 and of 26 respectively [17] Since in both casessimilar purification protocols were used and Lac1 and Lac2were recovered from the unbound and bound fractionsrespectively as Lap1 and Lap2 [17] this difference could bedue to the culture conditions used in each case (ie thepresence of agro-industrial wastes) Nevertheless the nanoLC-MSMS analysis showed that peptides of Lac1 and Lac2 werepresented in Lap1 (UniProtKBSwiss-Prot Q8TG93) and Lap2(UniProtKBSwiss-Prot Q8TG94) which have a predicted pI of43 and 47 respectively Lac1 and Lac2 peptides werepresented in Laccase-2 (UniProtKBSwiss-Prot Q99046) andLaccase-1 (UniProtKBSwiss-Prot Q99044) from T villosarespectively with 80 to 100 identity These T villosa laccaseisoenzymes have a predicted pI of 47 (laccase-1) and 60(laccase-2) and a 77 identity among them In addition Tvillosa laccase-2 is reported as a homodimer protein whilelaccase-1 is reported as monomeric protein Alignment of Lac1-Lap2 and Lac2-Lap1 from T pubescens CBS 69694 and Tvillosa showed a 96 and 70 identity respectively Finallysimilar peptides were identified for Lac1 and Lac2 from Tpubescens (Schumach) Pilaacutet (Syn Coriolus pubescens(Schum ex Fr) Queacutel) BCB 923-2 (Figure 3b) [51] Theseresults suggest that the purified fractions correspond to laccaseisoenzymes that differ from previously reported ones for this Tpubescens strain
As mentioned above laccases isoenzymes present differentstabilities and substrate affinities This behavior was alsoobserved for the laccase isoenzymes purified from Tpubescens with Lac1 showing lower laccase activity thanLac2 with ABTS as substrate However the difference inactivity between these isoenzymes depended on the agro-industrial waste used for the T pubescens growth suggestinga probable effect of the substrate composition on the laccasephysicochemical properties In addition it was observed thatthe total laccase activity presented in the UF retentates ishigher than the laccase activity of the purified fractionsregardless of the agro-industrial byproduct used for the Tpubescens growth This suggests a synergistic effect of Lac1
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 11 September 2013 | Volume 8 | Issue 9 | e73721
and Lac2 as have been reported for laccases obtained fromTrametes and Pleurotus strains [175253]
The maximum relative activity for the UF retentate waspresented at a pH from 20 to 40 regardless of the agro-industrial byproduct with a monotonical decrease at pHs abovethis range The relation between pH and type of electron donormolecule explains this behavior on laccase activity It has beenreported that when the substrate is an organic hydrogen donorie phenols the optimal pH ranges from 35 to 60 while thelaccase activity monotonically decreases from pH 25 to 70 ifthe substrate is an electron donor like ABTS [54] Despite thatat different pHs most of the purified laccase fractionspresented a similar performance as the UF retentates the Lac2fraction obtained from cedar sawdust differed from the previousobservation For this fraction the maximum activity wasobserved at pH 50 and despite that at acidic pH (ge 30) therelative activity was higher than 80 at lower pHs (20) itdecreased almost in a 50 Nevertheless at basic pHs therelative laccase activity presented a sharp decreased that iswell correlate with the other laccase fractions We are not ableto attribute this particular trend to the presence of multiplelaccase isoenzymes in the fraction since the SDS-PAGErevealed only one band when stained with ABTS and from theNanoLC-MSMS analysis no difference in the number ofpeptides and their sequence was detected We speculate thatthis particular behavior can be owed to a compound added inthe pre-treatment of the cedar wood that might be used forpreserving it that might affect the activity of Lac2 and that wasnot completely removed during the purification
Temperature stability evaluation showed that Lac1 is themost thermolabile fraction while the laccase activity reductionin the crude extracts did not seem to be the result of anadditive effect of the activity loss of the purified fractionsAlthough there are no reports of laccase stability under theselected conditions these results agree with previous reportsthat state that fungal laccases rapidly decrease their activity attemperatures near 60 degC and thermal stability correlates withthe temperature range of the growth of the source organism[55] In addition we previously reported for a laccase crudeextract obtained from culturing T pubescens on coffee husk anactivity loss of 26 and 10 at 60 and 50 degC respectivelyafter 8 h at pH 60 using syringaldazine as substrate [48]
Finally an electrochemical characterization was carried outfor selected crude extracts and purified fractions The results
showed a difference between the electrochemicalperformances of extracts and purified fractions withsyringaldazine or ABTS These differences could be owed tothe different mechanisms for electron transfer as discussedabove However the overall performance of the obtained cyclicvoltammograms for ABTS and syringaldazine agrees withresults reported by Fernaacutendez-Saacutenchez et al [25] who used asaturated calomel reference electrode
Conclusions
We have shown the potential of coffee husk soybean podhusk and cedar sawdust as substrates for the laccaseproduction In several countries like Brazil and Colombia theproduction of coffee and soybean is of great importance andthe generated volume of residues is high The results showedthat semi-solid culture of T pubescens using coffee huskallows to obtain an important amount of laccase which couldbe favored by the phenolic-compounds ie tannins presentedin coffee husk While supplementation of the culture mediumwith copper favored the production of laccase in submergedand semi-solid culture using soybean pod husk and cedarsawdust semi-solid culture using coffee husk which presentedthe highest laccase volumetric yield did not require coppersupplementation This is an economic advantage that shouldbe considered when scaling up the process In this study Tpubescens showed its ability to produce two independentlaccases regardless of the used substrate However a higherlaccase activity was observed for crude extracts rather than forpurified fractions This suggests that Lac1 and Lac2 might havea synergistic effect on the final laccase activity rather than anadditive effect Based on this study the investigation of laccaseproduction by evaluating other agro-industrial wastes in whichphenolic-compounds constitute an important percentage oftheir composition is of great interest to avoid the use ofinducers that would increase the production cost
Author Contributions
Conceived and designed the experiments JFO CJAD OFSPerformed the experiments JCG SCM AR Analyzed the dataJCG SCM AR JFO CJAD OFS Wrote the manuscript AR JFOCJAD OFS
References
1 Vandamme EJ (2009) Agro-industrial residue utilization for industrialbiotechnology products In P NigamA Pandey Biotechnology for Agro-Industrial Residues Utilisation Netherlands Springer Verlag pp 3-11
2 Martiacutenez AT Speranza M Ruiz-Duentildeas FJ Ferreira P Camarero S etal (2005) Biodegradation of lignocellulosics microbial chemical andenzymatic aspects of the fungal attack of lignin Int Microbiol 8195-204 PubMed 16200498
3 Saacutenchez O Sierra R Almeacuteciga-Diacuteaz CJ (2011) Delignification processof agro-industrial aastes an alternative to obtain fermentablecarbohydrates for producing fuel In M Manzanera Alternative FuelIntech 111-154
4 Wesenberg D Kyriakides I Agathos SN (2003) White-rot fungi andtheir enzymes for the treatment of industrial dye effluents BiotechnolAdv 22 161-187 doi101016jbiotechadv200308011 PubMed14623049
5 Morozova OV Shumakovich GP Shleev SV Yaropolov YI (2007)Laccase-mediator systems and their applications A review ApplBiochem Microbiol 43 523-535 doi101134S0003683807050055
6 Baldrian P (2006) Fungal laccases - occurrence and properties FEMSMicrobiol Rev 30 215-242 doi101111j1574-4976200500010xPubMed 16472305
7 Rodriacuteguez Couto S Herrera JLT (2006) Industrial and biotechnologicalapplications of laccases A review Biotechnol Adv 24 500-513 doi101016jbiotechadv200604003 PubMed 16716556
8 Osma JF Toca-Herrera JL Rodriacuteguez-Couto S (2010) Uses oflaccases in the food industry Enzyme Res 2010 2010 918761PubMed 21048873
9 Rodriguez S (2008) Exploitation of biological wastes for the productionof value-added products under solid-state fermentation conditionsBiotechnol J 3 859-870 doi101002biot200800031 PubMed18543242
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 12 September 2013 | Volume 8 | Issue 9 | e73721
10 Osma JF Toca-Herrera JL Rodriacuteguez-Couto S (2011) Cost analysis inlaccase production J Environ Manage 92 2907-2912 doi101016jjenvman201106052 PubMed 21775046
11 Jain A Morlok CK Henson JM (2013) Comparison of solid-state andsubmerged-state fermentation for the bioprocessing of switchgrass toethanol and acetate by Clostridium phytofermentans Appl MicrobiolBiotechnol 97 905-917 doi101007s00253-012-4511-4 PubMed23111595
12 Singhania RR Patel AK Soccol CR Pandey A (2009) Recentadvances in solid-state fermentation Biochem Eng J 44 13-18 doi101016jbej200810019
13 Economou CN Makri A Aggelis G Pavlou S Vayenas DV (2010)Semi-solid state fermentation of sweet sorghum for the biotechnologicalproduction of single cell oil Bioresour Technol 101 1385-1388 doi101016jbiortech200909028 PubMed 19781936
14 Rodriacuteguez CS Santoro R Cameselle C Sanroman A (1997) Laccaseproduction in semi-solid cultures of Phanerochaete chrysosporiumBiotechnol Lett 19 995-998 doi101023A1018495216946
15 Osma JF Moilanen U Toca-Herrera JL Rodriacuteguez-Couto S (2011)Morphology and laccase production of white-rot fungi grown on wheatbran flakes under semi-solid-state fermentation conditions FEMSMicrobiol Lett 318 27-34 doi101111j1574-6968201102234xPubMed 21291496
16 Couto SR Gundin M Lorenzo M Sanroman MN (2002) Screening ofsupports and inducers for laccase production by Trametes versicolor insemi-solid-state conditions Proc Biochem 38 249-255 doi101016S0032-9592(02)00087-0
17 Galhaup C Goller S Peterbauer CK Strauss J Haltrich D (2002)Characterization of the major laccase isoenzyme from Trametespubescens and regulation of its synthesis by metal ions Microbiology148 2159-2169 PubMed 12101303
18 Sievers F Wilm A Dineen D Gibson TJ Karplus K et al (2011) Fastscalable generation of high-quality protein multiple sequencealignments using Clustal Omega Mol Syst Biol 7 539 PubMed21988835
19 Childs RE Bardsley WG (1975) The steady-state kinetics of peroxidasewith 22-azino-di-(3-ethyl-benzthiazoline-6-sulphonic acid) aschromogen Biochem J 145 93-103 PubMed 1191252
20 Harkin JM Obst JR (1973) Syringaldazine an Effective Reagent forDetecting Laccase and Peroxidase in Fungi Experientia 29 382-387
21 Nikupaavola ML Raaska L Itavaara M (1990) Detection of white-rotfungi by a non-roxic stain Mycol Res 94 27-31 doi101016S0953-7562(09)81260-4
22 Gonzalez LF Sarria V Sanchez OF (2010) Degradation ofchlorophenols by sequential biological-advanced oxidative processusing Trametes pubescens and TiO(2)UV Bioresour Technol 1013493-3499
23 Waterborg JH (2009) The Lowry method for protein quantitation In JWalker The Protein Protocols Handbook Humana Press pp 7-10
24 Harkin JM Obst JR (1973) Syringaldazine an effective reagent fordetecting laccase and peroxidase in fungi Cell Mol Life Sci 29381-387 doi101007BF01926734
25 Fernaacutendez-Saacutenchez C Tzanov T Guumlbitz GM Cavaco-Paulo A (2002)Voltammetric monitoring of laccase-catalysed mediated reactionsBioelectrochemistry 58 149-156 doi101016S1567-5394(02)00119-6PubMed 12414320
26 Bourbonnais R Leech D Paice MG (1998) Electrochemical analysis ofthe interactions of laccase mediators with lignin model compoundsBiochim Biophys Acta 1379 381ndash390 doi101016S0304-4165(97)00117-7 PubMed 9545600
27 Ferraz F Silva S (2009) Characterization of coffee husk biomass forbiotechnological purposes N Biotechnol 25 S256-S256 doi101016jnbt200906573
28 Pandey A Soccol CR Nigam P Brand D Mohan R et al (2000)Biotechnological potential of coffee pulp and coffee husk forbioprocesses Biochem Eng J 6 153-162 doi101016S1369-703X(00)00084-X PubMed 10959086
29 Sruamsiri S Silman P (2008) Nutritive composition of soybean by-products and nutrient digestibility of soybean pod husk Maejo Int J SciTechnol 2 568-576
30 Romkaew J Nagaya Y Goto M Suzuki J Umezaki T (2008) Poddehiscence in relation to chemical components of pod shell in soybeanPlant Prod Sci 11 278-282 doi101626pps11278
31 Nakamanee G Polbumroong T Phaikaew C (2003) Alternative use ofcrop waste to increase milk production Proceedings of the 8th Meetingof the Regional Working Group on Grazing and Feed Resources forSoutheast Asia Kuala Lumpur Malaysia 66 - 70
32 Kurek B Gaudard F (2000) Oxidation of spruce wood sawdust byMnO(2) plus oxalate a biochemical investigation J Agric Food Chem48 3058-3062 doi101021jf000015g PubMed 10898665
33 Weil J Brewer M Hendrickson R Sarikaya A Ladisc MR (1998)Continuous pH monitoring during pretreatment of yellow poplar woodsawdust by pressure cooking in water Appl Biochem Biotechnol 70-7299-111 doi101007BF02920127
34 Weil J Sarikaya A Rau S-L Goetz J Ladisch CM et al (1997)Pretreatment of yellow poplar sawdust by pressure cooking in waterAppl Biochem Biotechnol 68 21-40 doi101007BF02785978
35 Dekker RF Barbosa AM Giese EC Godoy SD Covizzi LG (2007)Influence of nutrients on enhancing laccase production byBotryosphaeria rhodina MAMB-05 Int Microbiol 10 177-185 PubMed18075999
36 Xiao YZ Hong YZ Li JF Hang J Tong PG et al (2006) Cloning ofnovel laccase isozyme genes from Trametes sp AH28-2 and analysesof their differential expression Appl Microbiol Biotechnol 71 493-501doi101007s00253-005-0188-2 PubMed 16283298
37 Crowe JD Olsson S (2001) Induction of laccase activity in Rhizoctoniasolani by antagonistic Pseudomonas fluorescens strains and a range ofchemical treatments Appl Environ Microbiol 67 2088-2094 doi101128AEM6752088-20942001 PubMed 11319086
38 Yaguumle S Terroacuten MC Gonzaacutelez T Zapico E Bocchini P et al (2000)Biotreatment of tannin-rich beer-factory wastewater with white-rotbasidiomycete Coriolopsis gallica monitored by pyrolysisgaschromatographymass spectrometry Rapid Commun Mass Spectrom14 905-910 doi101002(SICI)1097-0231(20000530)1410 PubMed10825255
39 Gowda NKS Ramana JV Prasad CS Singh K (2004) Micronutrientcontent of certain tropical conventional and unconventional feedresources of southern India Trop Anim Heatlth Pro 36 77-94 doi101023BTROP0000009522309491d PubMed 14979561
40 DePeters EJ Fadel JG Arana MJ Ohanesian N Etchebarne MA et al(2000) Variability in the chemical composition of seventeen selectedby-product feedstuffs used by the California dairy industry Prof AnimSci 16 69-99
41 Obernberger I Thek G (2004) Physical characterisation and chemicalcomposition of densified biomass fuels with regard to their combustionbehaviour Biomass Bioenerg 27 653-669 doi101016jbiombioe200307006
42 Larous S Meniai A-H Lehocine MB (2005) Experimental study of theremoval of copper from aqueous solutions by adsorption usingsawdust Desalination 185 483ndash490 doi101016jdesal200503090
43 Oliveira WE Franca AS Oliveira LS Rocha SD (2008) Untreatedcoffee husks as biosorbents for the removal of heavy metals fromaqueous solutions J Hazard Mater 152 1073ndash1081 doi101016jjhazmat200707085 PubMed 17804159
44 Šciban M Klašnja M Škrbic B (2008) Adsorption of copper ions fromwater by modified agricultural by-products Desalination 229 170ndash180doi101016jdesal200708017
45 Maumlkelauml MR Hildeacuten KS Hakala TK Hatakka A Lundell TK (2006)Expression and molecular properties of a new laccase of the white rotfungus Phlebia radiata grown on wood Curr Genet 50 323-333 doi101007s00294-006-0090-1 PubMed 16927090
46 Maumlkelauml MR Lundell T Hatakka A Hildeacuten K (2013) Effect of coppernutrient nitrogen and wood-supplement on the production of lignin-modifying enzymes by the white-rot fungus Phlebia radiata Fungal Biol117 62-70 doi101016jfunbio201211006 PubMed 23332834
47 Dwivedi UN Singha P Pandeya VP Kumara A (2011) Structurendashfunction relationship among bacterial fungal and plant laccases J MolCatal B Enzym 68 117-128 doi101016jmolcatb201011002
48 Gaitan IJ Medina SC Gonzaacutelez JC Rodriacuteguez A Espejo AacuteJ et al(2011) Evaluation of toxicity and degradation of a chlorophenol mixtureby the laccase produced by Trametes pubescens Bioresour Technol102 3632ndash3635 doi101016jbiortech201011040 PubMed21115244
49 Karp SG Faraco V Amore A Birolo L Giangrande C et al (2012)Characterization of laccase isoforms produced by Pleurotus ostreatusin solid state fermentation of sugarcane bagasse Bioresour Technol114 735-739 doi101016jbiortech201203058 PubMed 22487128
50 Nikitina OV Shleev SV Gorshina ES Rusinova TV Serezhenkov VAet al (2005) Isolation and purification of enzymes from ligninolyticcomplex of the basidial fungus Trametes pubescens (Schumach) Pilatand study of their properties Biochemistry (Mosc) 70 1274-1279 doi101007s10541-005-0259-0
51 Shleev S Nikitina O Christenson A Reimann CT Yaropolov AI et al(2007) Characterization of two new multiforms of Trametes pubescenslaccase Bioorg Chem 35 35-49 doi101016jbioorg200608001PubMed 16989887
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 13 September 2013 | Volume 8 | Issue 9 | e73721
52 Xiao YZ Chen Q Hang J Shi YY Wu J et al (2004) Selectiveinduction purification and characterization of a laccase isozyme fromthe basidiomycete Trametes sp AH28-2 Mycologia 96 26-35 doi1023073761984 PubMed 21148825
53 Muntildeoz C Guilleacuten F Martiacutenez AT Martiacutenez MJ (1997) Laccaseisoenzymes of Pleurotus eryngii characterization catalytic propertiesand participation in activation of molecular oxygen and Mn2+ oxidationAppl Environ Microbiol 63 2166-2174 PubMed 9172335
54 Morozova OV Shumakovich GP Gorbacheva MA Shleev SVYaropolov AI (2007) Blue laccases Biochemistry (Mosc) 721136-1150 doi101134S0006297907100112
55 Hildeacuten K Hakala TK Lundell T (2009) Thermotolerant andthermostable laccases Biotechnol Lett 31 1117-1128 doi101007s10529-009-9998-0 PubMed 19360388
56 Verma P Madamwar D (2002) Production of ligninolytic enzymes fordye decolorization by cocultivation of white-rot fungi Pleurotusostreatus and Phanerochaete chrysosporium under solid-statefermentation Appl Biochem Biotechnol 102-103 109-118 doi101385ABAB102-1031-6109 PubMed 12396115
57 Meza JC Lomascolo A Casalot L Sigoillot J-C Auria R (2005)Laccase production by Pycnoporus cinnabarinus grown on sugar-canebagasse Influence of ethanol vapours as inducer Proc Biochem 403365ndash3371 doi101016jprocbio200503004
58 Hossain SM Anantharaman N (2006) Activity enhancement ofligninolytic enzymes of Trametes versicolor with bagasse powder Afr JBiotechnol 5 189-194
59 Revankar MS Desai KM Lele SS (2007) Solid-state fermentation forenhanced production of laccase using indigenously isolatedGanoderma sp Appl Biochem Biotechnol 143 16-26 doi101007s12010-007-0029-0 PubMed 18025593
60 Velazquez-Cedeno MA Mata G Savoie J-M (2002) Waste-reducingcultivation of Pleurotus ostreatus and Pleurotus pulmonarius on coffeepulp changes in the production of some lignocellulolytic enzymesWorld J Microbiol Biotechnol 18 201ndash207 doi101023A1014999616381
61 Mata G Murrieta D Andreu LI (2005) Changes in lignocellulolyticenzyme activities in six Pleurotus spp strains cultivated on coffee pulpin confrontation with Trichoderma spp World J Microbiol Biotechnol 21143ndash150 doi101007s11274-004-3041-3
62 de Souza DF Tychanowicz GK de Souza CG Peralta RM (2006) Co-production of ligninolytic enzymes by Pleurotus pulmonarius on wheatbran solid state cultures J Basic Microbiol 46 126-134 doi101002jobm200510014 PubMed 16598826
63 Prasad KK Mohan SV Bhaskar YV Ramanaiah SV Babu VL et al(2005) Laccase production using Pleurotus ostreatus 1804 immobilizedon PUF cubes in batch and packed bed reactors influence of cultureconditions J Microbiol 43 301-307 PubMed 15995650
64 Songulashvili G Elisashvili V Wasser SP Nevo E Hadar Y (2007)Basidiomycetes laccase and manganese peroxidase activity in
submerged fermentation of food industry wastes Enzyme MicrobTechnol 41 57ndash61 doi101016jenzmictec200611024
65 Sathishkumar P Murugesan K Palvannan T (2010) Production oflaccase from Pleurotus florida using agro-wastes and efficientdecolorization of Reactive blue 198 J Basic Microbiol 50 360-367 doi101002jobm200900407 PubMed 20586068
66 Moldes D Gallego PP Rodriacuteguez Couto S Sanromaacuten A (2003) Grapeseeds the best lignocellulosic waste to produce laccase by solid statecultures of Trametes hirsuta Biotechnol Lett 25 491-495 doi101023A1022660230653 PubMed 12882277
67 Elisashvili V Kachlishvili E Penninckx M (2008) Effect of growthsubstrate method of fermentation and nitrogen source onlignocellulose-degrading enzymes production by white-rotbasidiomycetes J Ind Microbiol Biotechnol 35 1531-1538 doi101007s10295-008-0454-2 PubMed 18716810
68 Osma JF Saravia V Herrera JL Couto SR (2007) Mandarin peelingsthe best carbon source to produce laccase by static cultures ofTrametes pubescens Chemosphere 67 1677-1680 doi101016jchemosphere200611051 PubMed 17234250
69 Kahraman SS Gurdal IH (2002) Effect of synthetic and natural culturemedia on laccase production by white rot fungi Bioresour Technol 82215-217 doi101016S0960-8524(01)00193-6 PubMed 11991068
70 Iandolo D Piscitelli A Sannia G Faraco V (2011) Enzyme Productionby Solid Substrate Fermentation of Pleurotus ostreatus and Trametesversicolor on Tomato Pomace Appl Biochem Biotechnol 163 40-51doi101007s12010-010-9014-0 PubMed 20582639
71 Moldes D Lorenzo M Sanromaacuten MA (2004) Different proportions oflaccase isoenzymes produced by submerged cultures of Trametesversicolor grown on lignocellulosic wastes Biotechnol Lett 26 327-330doi101023BBILE000001545240213bf PubMed 15055770
72 Philippoussis A Diamantopoulou P Papadopoulou K Lakhtar HRoussos S et al (2010) Biomass laccase and endoglucanaseproduction by Lentinula edodes during solid state fermentation of reedgrass bean stalks and wheat straw residues World J MicrobiolBiotechnol 27 285-297
73 Fenice M Giovannozzi Sermanni G Federici F DrsquoAnnibale A (2003)Submerged and solid-state production of laccase and Mn-peroxidaseby Panus tigrinus on olive mill wastewater-based media J Biotechnol100 77-85 doi101016S0168-1656(02)00241-9 PubMed 12413788
74 Rodriguez-Couto S GundIn M Lorenzo M Sanromaacuten MAacute (2002)Screening of supports and inducers for laccase production by Trametesversicolor in semi-solid-state conditions Proc Biochem 38 249-255doi101016S0032-9592(02)00087-0
75 Fackler K Gradinger C Schmutzer M Tavzes C Burgert I et al (2007)Biotechnological wood modification with selective white-rot fungi and itsmolecular mechanisms Food Technol Biotechnol 45 269ndash276
76 Levin L Herrmann C Papinutti VL (2008) Optimization oflignocellulolytic enzyme production by the white-rot fungus Trametestrogii in solid-state fermentation using response surface methodologyBiochem Eng J 39 207-214 doi101016jbej200709004
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 14 September 2013 | Volume 8 | Issue 9 | e73721

presented the highest enzyme activity ~90 of its initial valuein comparison to the other two UF retentates (Figure 4)
The temperature and pH effect on laccase activity for Lac1and Lac2 is presented in Figure 5 The maximum relativeactivity for both fractions was presented at a pH from 20 to 40regardless of the agro-industrial waste as observed for the UFretentate However at pH gt 40 Lac1 showed a sharperdecrease in its activity than that observed for Lac2Nevertheless Lac2 purified from cedar sawdust culturepresented a different performance This fraction showed themaximum activity at pH 50 while a sharp drop was observedat pHs above or below this value
Laccase activity profiles showed that Lac1 and Lac2presented the maximum enzyme activity at 55 and 60 degCrespectively (Table 4) It was observed that Lac1 from coffeehusk culture presented a lower decrease in enzyme activity attemperatures le 50 degC or ge 60 degC than Lac2 On the contraryLac2 from soybean husk or cedar sawdust cultures presenteda higher relative laccase activity than Lac1 at temperatures le55 degC The purified fractions from soybean pod husk and cedarsawdust cultures presented the lowest relative activity at 30 degCand 65 degC
Table 4 Optimum temperatures and pHs for maximumlaccase activities in UF retentate Lac1 and Lac2 obtainedfrom semisolid cultured T pubescens on agro-industrialwastes
EnzymaticFraction Temperature pH
VolumetricActivity (UL-1)
SpecificActivity (Umg-1 protein)
VolumetricActivity (UL-1)
SpecificActivity (Umg-1
protein)SoybeanPod Husk +2 mM Cu+2
148000plusmn1178(55)
1521plusmn0012182037plusmn3025(2-3)
1871plusmn0031
Lac122925plusmn917(60)
2363plusmn009536135plusmn831(23)
3725plusmn0085
Lac232638plusmn1558(55)
1942plusmn009228713plusmn274(23)
1709plusmn0016
CoffeeHusk + 0mM Cu+2
113740plusmn8289(50-55)
3008plusmn0219290666plusmn1039(2-3)
7689plusmn0275
Lac1 889plusmn039 (55) 0038plusmn0002 1779plusmn028 (23) 0077plusmn0001
Lac234194plusmn2789(60)
2947plusmn024033150plusmn526 (2ndash4)
2857plusmn0045
CedarSawdust +5 mM Cu+2
257666plusmn1201(55)
5052plusmn0024344634plusmn1043(2-3)
6757plusmn0204
Lac121370plusmn472(55)
0962plusmn002129117plusmn943(23)
1311plusmn0042
Lac255166plusmn1656(60)
1379plusmn0414 55166plusmn1656 (5) 1379plusmn0414
ABTS was used to determine laccase activity
Laccase stabilityFor selected enzymatic extracts and purified fractions (Table
4) a stability test for 48 h at their optimal pH and temperaturewas conducted Time course of the relative laccase activity forcrude extract and purified fractions from coffee husk and cedarsawdust cultures are presented in Figure 5 Similar resultswere observed for crude extract and purified fractions fromsoybean pod husk culture (data no shown)
It was observed that the activity loss depends on thesubstrate source used to grow T pubescens An activity loss of55 was observed for crude extracts and purified fractionsfrom coffee husk and soybean pod husk cultures while anactivity loss of 70 was observed for the crude extract andpurified fractions from cedar sawdust culture NeverthelessLac2 showed the lowest activity reduction with values of 10 40and 55 for coffee husk soybean pod husk and cedarsawdust respectively while Lac1 presented an activity loss of34 50 and 60 for coffee husk soybean pod husk and cedarsawdust respectively Despite the differences in laccaseactivity among agro-industrial wastes and between crudeextracts and purified fractions all the profiles showed a sharp
Figure 4 Effect of temperature (a) and pH (b) on laccaseactivity for the UF retentate obtained by semisolid cultureof T pubescens on coffee husk () soybean pod husk (loz)and cedar sawdust (Δ) Laccase stability was evaluated byusing ABTS All the assays were performed in triplicatedoi 101371journalpone0073721g004
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 8 September 2013 | Volume 8 | Issue 9 | e73721
decrease during the first 8 h while a reduced activity loss wasobserved during the last 24 h of the assay (Figure 6)
Laccase electrochemical characterizationFor selected crude extracts and purified fractions a cyclic
voltammetry characterization was conducted For eachmediator syringaldazine or ABTS similar cyclic
Figure 5 Effect of temperature () and pH () on enzymeactivity for Lac1 (solid line) and Lac2 (dashed line)obtained after the purification of the enzymatic crudeextract produced by semisolid culture of T pubescens oncoffee husk (a) soybean pod husk (b) and cedar sawdust(c) Laccase stability was evaluated by using ABTS All theassays were performed in triplicatedoi 101371journalpone0073721g005
voltammograms were obtained for the different crude extractsand purified fractions Voltammograms of the redox reaction forsyringaldazine (Figure 7a) and ABTS (Figure 6b) arepresented Two anionic and cationic peaks were observedwhen syringaldazine was used as mediator The first pair ofpeaks showed a reversible performance with a mean Ia Icratio of 109plusmn006 106plusmn005 and 098plusmn002 for the crudeextracts Lac1 and Lac2 respectively The second pair ofpeaks showed quasi-reversible performance and the cathodicpeak was presented at a potential of +062 V (vs Ag|AgCl)while the anodic peak was presented at a potential of +060 V(vs Ag|AgCl) No significant difference on the reductionpotential (Eo) for syringaldazine was observed for the crudeextracts or the purified fractions obtaining an Eo of0314plusmn0008 V (vs Ag|AgCl) However the peak potentialseparation (ΔEp) showed a different profile depending on thesubstrate used for the culture While not significant difference(p gt 005) on ΔEp value was observed between crude extract(0045plusmn0005 V vs Ag|AgCl) Lac1 (0046plusmn0005 V vs Ag|AgCl)and Lac2 (0028plusmn0001 V vs Ag|AgCl) from cultures with cedarsawdust and soybean pod husk the ΔEp was considerablyhigher for the crude extract (0083plusmn0004 V vs Ag|AgCl) Lac1(0068plusmn0003 V vs Ag|AgCl) and Lac2 (0051plusmn0005 V vs Ag|AgCl) from the coffee husk culture
For reactions conducted with ABTS and the different crudeextracts and purified fractions the voltammograms exhibitedone pair of cathodic and anodic peaks (Figure 7b) whichcorresponds to the redox of ABTSABTS+ (IaIc) with an Eo of0487plusmn0006 V (vs Ag|AgCl) This process was reversiblepresenting a mean Ia Ic ratio of 099plusmn002 In addition asecond cathodic peak was presented at +083 V (vs Ag|AgCl)when ABTS redox reaction was conducted using the crudeextract and the purified fractions This performance agrees withprevious reports in which the laccase activity cannot beobserved at potential values of the second reversible oxidation
Figure 6 Laccase stability profiles at optimal pH andtemperature conditions for crude extracts (solid line) Lac1(dashed line) and Lac2 (spotted line) obtained fromsemisolid culture of T pubescens on coffee husk (loz) andcedar sawdust (bull) Laccase stability was evaluated by usingABTS All the assays were performed in triplicatedoi 101371journalpone0073721g006
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 9 September 2013 | Volume 8 | Issue 9 | e73721
of ABTS ABTS+ABTS2+ due to this catalytic process cannotbe detected by cyclic voltammetry [2526] Neverthelessvoltammograms for laccase crude extract from a soybean podhusk culture presented the second anodic peak which inaddition to the second cathodic peak corresponds to the redoxof ABTS+ABTS2+ (IIaIIc) with an Eo of 0942plusmn0004 V (vs Ag|AgCl) Contrary to the reactions conducted usingsyringaldazine as substrate the ΔEp did not present asignificant difference for the crude extracts and purifiedfractions 0248plusmn0016 V (vs Ag|AgCl)
Discussion
In this study we evaluated the effect of the culture mode ieSmC and SSC on laccase production by T pubescens CBS69694 using three different agro-industrial wastes (coffeehusk soybean pod husk and cedar sawdust) Furthermore theeffect of Cu+2 supplementation on laccase production was alsoevaluated and the crude extracts and the purified fractionswere partially characterized
Figure 7 Cyclic voltammograms of (a) syringaldazine atpH 50 using the crude extract and purified laccasefractions from semisolid culture of T pubescens on coffeehusk or (b) ABTS at pH 30 using the crude extract andpurified laccase fractions from semisolid culture of Tpubescens on soybean pod husk Nomenclature onlymediator (dotted line) crude extract (solid line) Lac1 fraction(dashed line) and Lac2 fraction (dashed and dotted line)doi 101371journalpone0073721g007
Substrate source for laccase production showed to be one ofthe most important factors for laccase production (Table 2)Cultures with coffee husk as substrate presented the highestlaccase production levels The observed differences in laccaseactivity for each substrate could be owed to their chemicalcomposition Coffea arabica is the coffee variety raised inColombia and its husk has a reported composition of 2308cellulose (2076 glucose and 183 cellobiose) 2385hemicellulose (1356 xylose 523 arabinose 256 aceticacid and 195 glucuronic acid) 2828 total lignin and071 ashes [27] In addition coffee husk contents 13caffeine 45 tannins and 124 pectins [28] On the otherhand Glycine max is the soybean species cultured inColombia and its pod husk composition varies from 504 to62 crude protein 608 to 143 hemicellulose 3715 to4208 acid detergent fiber 4436 to 6015 neutral detergentfiber 850 to 893 acid detergent lignin and 2074 to 2106uronic acid [29ndash31] Finally for cedar sawdust the majorcomponents are glucose (426-444) xylose (46-195)cellulose (383-410) klason lignin (227-278) and acidlignin (40-43) [32ndash34] Although lignin is one of the mostimportant compounds associated with laccase production bywhite-rot fungi we did not observe a clear correlation betweenlignin composition percentage and laccase activity In thatsense the high laccase activity observed for the cultures thatuse coffee husk could be associated with its content of caffeineand tannins which have been identified as potent inducers oflaccase gene expression [173536] In fact caffeine has beenused as inducer for laccase production by Rhizoctonia solaniobtaining an increase of 41-fold in laccase activity [37] Similarresults were also reported for the production of Coloriopsisgallica laccase [38]
Laccase gene expression is mainly up regulated by Cu2+although Mn2+ Fe3+ heavy metals 26-dimethoxy-14-benzoquinone H2O2 amphotericin B syringic acid tannic acidTween 80 soybean oil aromatic compounds andmicroclimatic changes (ie lower temperature and osmoticpressure) are also recognized as potent inducers [173536] Ingeneral Cu2+ supplementation increased the laccase activity inboth SmC and SSC However Cu2+concentration required toobtain the highest laccase activity varied depending on agro-industrial waste These differences can be attributed to thecopper presented in the agro-industrial waste and the copperavailability in the medium Copper can be found in amountsthat range from 156 to 180 mg kg-1 of coffee husk [39] andfrom 26 to 67 mg kg-1 of soybean pod husk (or soybean hull)[3940] Despite that no reports indicate the copper content inuntreated cedar wood an average of 10 mg kg-1 of wood hasbeen reported for wood used as biomass in six different Europecountries [41] This allows to consider the cedar sawdust as thesubstrate with the lowest content of copper On the other handcopper adsorption by sawdust and coffee husk has beenreported obtaining a Langmuir based maximum adsorptioncapacity (Qmax mg g-1 adsorbent) of 845 and 75 respectively[4243] Although no adsorption studies has been reportedusing untreated soybean pod husk Šćiban et al [44] reporteda Qmax for copper adsorption of 0085 mmol g-1 adsorbent (~ 54mg g-1) using treated soybean straws Based on this
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 10 September 2013 | Volume 8 | Issue 9 | e73721
information it can be expected that a higher amount ofexogenous copper would be required for laccase induction oncultures with cedar sawdust than with soybean pod husk andcoffee husk due to its lower content of copper In addition dueto the high copper-adsorption capacity of sawdust the amountof available copper in the culture would be reduced allowingthe laccase production at higher copper concentrations withoutreaching levels that might be toxic for the fungus This fact canbe observed in the SSC (Table 4) However for the SmC thisbehavior is not maintained for the cultures with soybean podhusk and coffee husk (Table 4) where the maximum laccaseactivity was obtained with a copper concentration of 05 and 5mM respectively This could be owed to a lower contactbetween the fungus and the substrate in the SmC whencompared to the SSC which is close related to the transport ofnutrients presented in the substrates
Production of extracellular laccases has been reported in alarge number of species of white-rot fungi strains grown onnatural substrates (Table 1) Although a laccase activity profileis not clearly observed based on the agro-industrial wasteculture mode and fungus strain most cases in which SmC andSSC were conducted using the same substrate and fungusstrain showed a higher laccase activity under SmC conditionsIn this study was observed a substrate dependentperformance where the most favorable condition to obtain thehighest laccase activity was the SSC The laccase activitiesobtained in the present study are comparable and in somecases higher than those reported for Pleurotus ostreatusPhanerochaete chrysosporium Pycnoporus cinnabarinus Tversicolor and Ganoderma sp cultured under SC and SmCconditions and using different agro-industrial wastes (Table 1)Semi-solid cultures using different agro-industrial wastes havebeen reported using T versicolor Phlebia radiata Tpubescens P ostreatus and C unicolor Rodriguez et al [16]screened different supports for laccase production by Tversicolor under SSC They obtained the highest laccaseactivity (1200 U L-1) when in the culture medium was presentedbarley bran In addition they were able to obtain a laccaseactivity of 1700 U L-1 when the culture was supplemented withxylidine as an inducer These results were obtained after 17 or18 days of culture and ABTS was used to determine theenzyme activity Maumlkelauml et al [4546] have studied theexpression of the genes involved in laccase production Pr-lac1and Pr-lac2 presented in P radiata and evaluate laccaseproduction in SSC using milled alder wood (Alnus incana) assubstrate They reported a maximum laccase activity of 3 microkatL-1 (~180 U L-1) at the 14th day of culture and usingsyringaldazine for its detection Osma et al [15] have studiedthe morphological changes and laccase production of strains Tpubescens P ostreatus C unicolor and T versicolor grownon wheat bran flakes using SSC They obtained a maximumlaccase production between 1397 and 2778 U L-1 after 10 and13 days of culture This enzyme activity is strain dependentand ABTS was used for its quantification Our results show agood correlation with reported data The highest laccaseactivity was obtained when T pubescens was grown on coffeehusk under SSC without copper (204 U L-1 evaluated withsyringaldazine or 1549 U L-1 evaluated with ABTS) which is in
some cases higher than reported ones This shows that underthe studied conditions the production of the laccase crudeextract is favored in SSC using coffee husk without the additionof copper which could favors the cost of the productionprocess
It has been reported for white rot-fungi the production ofmultiple laccase isoenzymes which are encoded by differentgenes and show different expression profiles stabilitiessubstrate affinities and molecular masses (ranging from 35 to140 kDa) [3647ndash49] Two isoforms have been identified in Tpubescens strains two monomeric (Lac1 and Lac2) enzymesof 67 kDa but with different pI (53 and 51) [50] and twolaccase-activity fractions (Lap1 and Lap2) with different pI (gt30 and 26) from which Lap2 is a monomeric enzyme of 65kDa [17] In this study we identify two laccase isoenzymesfrom T pubescens CBS 69694 that have different molecularmasses (120 and 60 kDa) and pI The purifications resultssuggest a more acidic pI for Lac1 (le 4) than Lac2 (ge 6) whichdiffers from a previous report for this fungus in which twofractions with laccase activity were obtained Lap1 and Lap2with pI gt 3 and of 26 respectively [17] Since in both casessimilar purification protocols were used and Lac1 and Lac2were recovered from the unbound and bound fractionsrespectively as Lap1 and Lap2 [17] this difference could bedue to the culture conditions used in each case (ie thepresence of agro-industrial wastes) Nevertheless the nanoLC-MSMS analysis showed that peptides of Lac1 and Lac2 werepresented in Lap1 (UniProtKBSwiss-Prot Q8TG93) and Lap2(UniProtKBSwiss-Prot Q8TG94) which have a predicted pI of43 and 47 respectively Lac1 and Lac2 peptides werepresented in Laccase-2 (UniProtKBSwiss-Prot Q99046) andLaccase-1 (UniProtKBSwiss-Prot Q99044) from T villosarespectively with 80 to 100 identity These T villosa laccaseisoenzymes have a predicted pI of 47 (laccase-1) and 60(laccase-2) and a 77 identity among them In addition Tvillosa laccase-2 is reported as a homodimer protein whilelaccase-1 is reported as monomeric protein Alignment of Lac1-Lap2 and Lac2-Lap1 from T pubescens CBS 69694 and Tvillosa showed a 96 and 70 identity respectively Finallysimilar peptides were identified for Lac1 and Lac2 from Tpubescens (Schumach) Pilaacutet (Syn Coriolus pubescens(Schum ex Fr) Queacutel) BCB 923-2 (Figure 3b) [51] Theseresults suggest that the purified fractions correspond to laccaseisoenzymes that differ from previously reported ones for this Tpubescens strain
As mentioned above laccases isoenzymes present differentstabilities and substrate affinities This behavior was alsoobserved for the laccase isoenzymes purified from Tpubescens with Lac1 showing lower laccase activity thanLac2 with ABTS as substrate However the difference inactivity between these isoenzymes depended on the agro-industrial waste used for the T pubescens growth suggestinga probable effect of the substrate composition on the laccasephysicochemical properties In addition it was observed thatthe total laccase activity presented in the UF retentates ishigher than the laccase activity of the purified fractionsregardless of the agro-industrial byproduct used for the Tpubescens growth This suggests a synergistic effect of Lac1
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 11 September 2013 | Volume 8 | Issue 9 | e73721
and Lac2 as have been reported for laccases obtained fromTrametes and Pleurotus strains [175253]
The maximum relative activity for the UF retentate waspresented at a pH from 20 to 40 regardless of the agro-industrial byproduct with a monotonical decrease at pHs abovethis range The relation between pH and type of electron donormolecule explains this behavior on laccase activity It has beenreported that when the substrate is an organic hydrogen donorie phenols the optimal pH ranges from 35 to 60 while thelaccase activity monotonically decreases from pH 25 to 70 ifthe substrate is an electron donor like ABTS [54] Despite thatat different pHs most of the purified laccase fractionspresented a similar performance as the UF retentates the Lac2fraction obtained from cedar sawdust differed from the previousobservation For this fraction the maximum activity wasobserved at pH 50 and despite that at acidic pH (ge 30) therelative activity was higher than 80 at lower pHs (20) itdecreased almost in a 50 Nevertheless at basic pHs therelative laccase activity presented a sharp decreased that iswell correlate with the other laccase fractions We are not ableto attribute this particular trend to the presence of multiplelaccase isoenzymes in the fraction since the SDS-PAGErevealed only one band when stained with ABTS and from theNanoLC-MSMS analysis no difference in the number ofpeptides and their sequence was detected We speculate thatthis particular behavior can be owed to a compound added inthe pre-treatment of the cedar wood that might be used forpreserving it that might affect the activity of Lac2 and that wasnot completely removed during the purification
Temperature stability evaluation showed that Lac1 is themost thermolabile fraction while the laccase activity reductionin the crude extracts did not seem to be the result of anadditive effect of the activity loss of the purified fractionsAlthough there are no reports of laccase stability under theselected conditions these results agree with previous reportsthat state that fungal laccases rapidly decrease their activity attemperatures near 60 degC and thermal stability correlates withthe temperature range of the growth of the source organism[55] In addition we previously reported for a laccase crudeextract obtained from culturing T pubescens on coffee husk anactivity loss of 26 and 10 at 60 and 50 degC respectivelyafter 8 h at pH 60 using syringaldazine as substrate [48]
Finally an electrochemical characterization was carried outfor selected crude extracts and purified fractions The results
showed a difference between the electrochemicalperformances of extracts and purified fractions withsyringaldazine or ABTS These differences could be owed tothe different mechanisms for electron transfer as discussedabove However the overall performance of the obtained cyclicvoltammograms for ABTS and syringaldazine agrees withresults reported by Fernaacutendez-Saacutenchez et al [25] who used asaturated calomel reference electrode
Conclusions
We have shown the potential of coffee husk soybean podhusk and cedar sawdust as substrates for the laccaseproduction In several countries like Brazil and Colombia theproduction of coffee and soybean is of great importance andthe generated volume of residues is high The results showedthat semi-solid culture of T pubescens using coffee huskallows to obtain an important amount of laccase which couldbe favored by the phenolic-compounds ie tannins presentedin coffee husk While supplementation of the culture mediumwith copper favored the production of laccase in submergedand semi-solid culture using soybean pod husk and cedarsawdust semi-solid culture using coffee husk which presentedthe highest laccase volumetric yield did not require coppersupplementation This is an economic advantage that shouldbe considered when scaling up the process In this study Tpubescens showed its ability to produce two independentlaccases regardless of the used substrate However a higherlaccase activity was observed for crude extracts rather than forpurified fractions This suggests that Lac1 and Lac2 might havea synergistic effect on the final laccase activity rather than anadditive effect Based on this study the investigation of laccaseproduction by evaluating other agro-industrial wastes in whichphenolic-compounds constitute an important percentage oftheir composition is of great interest to avoid the use ofinducers that would increase the production cost
Author Contributions
Conceived and designed the experiments JFO CJAD OFSPerformed the experiments JCG SCM AR Analyzed the dataJCG SCM AR JFO CJAD OFS Wrote the manuscript AR JFOCJAD OFS
References
1 Vandamme EJ (2009) Agro-industrial residue utilization for industrialbiotechnology products In P NigamA Pandey Biotechnology for Agro-Industrial Residues Utilisation Netherlands Springer Verlag pp 3-11
2 Martiacutenez AT Speranza M Ruiz-Duentildeas FJ Ferreira P Camarero S etal (2005) Biodegradation of lignocellulosics microbial chemical andenzymatic aspects of the fungal attack of lignin Int Microbiol 8195-204 PubMed 16200498
3 Saacutenchez O Sierra R Almeacuteciga-Diacuteaz CJ (2011) Delignification processof agro-industrial aastes an alternative to obtain fermentablecarbohydrates for producing fuel In M Manzanera Alternative FuelIntech 111-154
4 Wesenberg D Kyriakides I Agathos SN (2003) White-rot fungi andtheir enzymes for the treatment of industrial dye effluents BiotechnolAdv 22 161-187 doi101016jbiotechadv200308011 PubMed14623049
5 Morozova OV Shumakovich GP Shleev SV Yaropolov YI (2007)Laccase-mediator systems and their applications A review ApplBiochem Microbiol 43 523-535 doi101134S0003683807050055
6 Baldrian P (2006) Fungal laccases - occurrence and properties FEMSMicrobiol Rev 30 215-242 doi101111j1574-4976200500010xPubMed 16472305
7 Rodriacuteguez Couto S Herrera JLT (2006) Industrial and biotechnologicalapplications of laccases A review Biotechnol Adv 24 500-513 doi101016jbiotechadv200604003 PubMed 16716556
8 Osma JF Toca-Herrera JL Rodriacuteguez-Couto S (2010) Uses oflaccases in the food industry Enzyme Res 2010 2010 918761PubMed 21048873
9 Rodriguez S (2008) Exploitation of biological wastes for the productionof value-added products under solid-state fermentation conditionsBiotechnol J 3 859-870 doi101002biot200800031 PubMed18543242
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 12 September 2013 | Volume 8 | Issue 9 | e73721
10 Osma JF Toca-Herrera JL Rodriacuteguez-Couto S (2011) Cost analysis inlaccase production J Environ Manage 92 2907-2912 doi101016jjenvman201106052 PubMed 21775046
11 Jain A Morlok CK Henson JM (2013) Comparison of solid-state andsubmerged-state fermentation for the bioprocessing of switchgrass toethanol and acetate by Clostridium phytofermentans Appl MicrobiolBiotechnol 97 905-917 doi101007s00253-012-4511-4 PubMed23111595
12 Singhania RR Patel AK Soccol CR Pandey A (2009) Recentadvances in solid-state fermentation Biochem Eng J 44 13-18 doi101016jbej200810019
13 Economou CN Makri A Aggelis G Pavlou S Vayenas DV (2010)Semi-solid state fermentation of sweet sorghum for the biotechnologicalproduction of single cell oil Bioresour Technol 101 1385-1388 doi101016jbiortech200909028 PubMed 19781936
14 Rodriacuteguez CS Santoro R Cameselle C Sanroman A (1997) Laccaseproduction in semi-solid cultures of Phanerochaete chrysosporiumBiotechnol Lett 19 995-998 doi101023A1018495216946
15 Osma JF Moilanen U Toca-Herrera JL Rodriacuteguez-Couto S (2011)Morphology and laccase production of white-rot fungi grown on wheatbran flakes under semi-solid-state fermentation conditions FEMSMicrobiol Lett 318 27-34 doi101111j1574-6968201102234xPubMed 21291496
16 Couto SR Gundin M Lorenzo M Sanroman MN (2002) Screening ofsupports and inducers for laccase production by Trametes versicolor insemi-solid-state conditions Proc Biochem 38 249-255 doi101016S0032-9592(02)00087-0
17 Galhaup C Goller S Peterbauer CK Strauss J Haltrich D (2002)Characterization of the major laccase isoenzyme from Trametespubescens and regulation of its synthesis by metal ions Microbiology148 2159-2169 PubMed 12101303
18 Sievers F Wilm A Dineen D Gibson TJ Karplus K et al (2011) Fastscalable generation of high-quality protein multiple sequencealignments using Clustal Omega Mol Syst Biol 7 539 PubMed21988835
19 Childs RE Bardsley WG (1975) The steady-state kinetics of peroxidasewith 22-azino-di-(3-ethyl-benzthiazoline-6-sulphonic acid) aschromogen Biochem J 145 93-103 PubMed 1191252
20 Harkin JM Obst JR (1973) Syringaldazine an Effective Reagent forDetecting Laccase and Peroxidase in Fungi Experientia 29 382-387
21 Nikupaavola ML Raaska L Itavaara M (1990) Detection of white-rotfungi by a non-roxic stain Mycol Res 94 27-31 doi101016S0953-7562(09)81260-4
22 Gonzalez LF Sarria V Sanchez OF (2010) Degradation ofchlorophenols by sequential biological-advanced oxidative processusing Trametes pubescens and TiO(2)UV Bioresour Technol 1013493-3499
23 Waterborg JH (2009) The Lowry method for protein quantitation In JWalker The Protein Protocols Handbook Humana Press pp 7-10
24 Harkin JM Obst JR (1973) Syringaldazine an effective reagent fordetecting laccase and peroxidase in fungi Cell Mol Life Sci 29381-387 doi101007BF01926734
25 Fernaacutendez-Saacutenchez C Tzanov T Guumlbitz GM Cavaco-Paulo A (2002)Voltammetric monitoring of laccase-catalysed mediated reactionsBioelectrochemistry 58 149-156 doi101016S1567-5394(02)00119-6PubMed 12414320
26 Bourbonnais R Leech D Paice MG (1998) Electrochemical analysis ofthe interactions of laccase mediators with lignin model compoundsBiochim Biophys Acta 1379 381ndash390 doi101016S0304-4165(97)00117-7 PubMed 9545600
27 Ferraz F Silva S (2009) Characterization of coffee husk biomass forbiotechnological purposes N Biotechnol 25 S256-S256 doi101016jnbt200906573
28 Pandey A Soccol CR Nigam P Brand D Mohan R et al (2000)Biotechnological potential of coffee pulp and coffee husk forbioprocesses Biochem Eng J 6 153-162 doi101016S1369-703X(00)00084-X PubMed 10959086
29 Sruamsiri S Silman P (2008) Nutritive composition of soybean by-products and nutrient digestibility of soybean pod husk Maejo Int J SciTechnol 2 568-576
30 Romkaew J Nagaya Y Goto M Suzuki J Umezaki T (2008) Poddehiscence in relation to chemical components of pod shell in soybeanPlant Prod Sci 11 278-282 doi101626pps11278
31 Nakamanee G Polbumroong T Phaikaew C (2003) Alternative use ofcrop waste to increase milk production Proceedings of the 8th Meetingof the Regional Working Group on Grazing and Feed Resources forSoutheast Asia Kuala Lumpur Malaysia 66 - 70
32 Kurek B Gaudard F (2000) Oxidation of spruce wood sawdust byMnO(2) plus oxalate a biochemical investigation J Agric Food Chem48 3058-3062 doi101021jf000015g PubMed 10898665
33 Weil J Brewer M Hendrickson R Sarikaya A Ladisc MR (1998)Continuous pH monitoring during pretreatment of yellow poplar woodsawdust by pressure cooking in water Appl Biochem Biotechnol 70-7299-111 doi101007BF02920127
34 Weil J Sarikaya A Rau S-L Goetz J Ladisch CM et al (1997)Pretreatment of yellow poplar sawdust by pressure cooking in waterAppl Biochem Biotechnol 68 21-40 doi101007BF02785978
35 Dekker RF Barbosa AM Giese EC Godoy SD Covizzi LG (2007)Influence of nutrients on enhancing laccase production byBotryosphaeria rhodina MAMB-05 Int Microbiol 10 177-185 PubMed18075999
36 Xiao YZ Hong YZ Li JF Hang J Tong PG et al (2006) Cloning ofnovel laccase isozyme genes from Trametes sp AH28-2 and analysesof their differential expression Appl Microbiol Biotechnol 71 493-501doi101007s00253-005-0188-2 PubMed 16283298
37 Crowe JD Olsson S (2001) Induction of laccase activity in Rhizoctoniasolani by antagonistic Pseudomonas fluorescens strains and a range ofchemical treatments Appl Environ Microbiol 67 2088-2094 doi101128AEM6752088-20942001 PubMed 11319086
38 Yaguumle S Terroacuten MC Gonzaacutelez T Zapico E Bocchini P et al (2000)Biotreatment of tannin-rich beer-factory wastewater with white-rotbasidiomycete Coriolopsis gallica monitored by pyrolysisgaschromatographymass spectrometry Rapid Commun Mass Spectrom14 905-910 doi101002(SICI)1097-0231(20000530)1410 PubMed10825255
39 Gowda NKS Ramana JV Prasad CS Singh K (2004) Micronutrientcontent of certain tropical conventional and unconventional feedresources of southern India Trop Anim Heatlth Pro 36 77-94 doi101023BTROP0000009522309491d PubMed 14979561
40 DePeters EJ Fadel JG Arana MJ Ohanesian N Etchebarne MA et al(2000) Variability in the chemical composition of seventeen selectedby-product feedstuffs used by the California dairy industry Prof AnimSci 16 69-99
41 Obernberger I Thek G (2004) Physical characterisation and chemicalcomposition of densified biomass fuels with regard to their combustionbehaviour Biomass Bioenerg 27 653-669 doi101016jbiombioe200307006
42 Larous S Meniai A-H Lehocine MB (2005) Experimental study of theremoval of copper from aqueous solutions by adsorption usingsawdust Desalination 185 483ndash490 doi101016jdesal200503090
43 Oliveira WE Franca AS Oliveira LS Rocha SD (2008) Untreatedcoffee husks as biosorbents for the removal of heavy metals fromaqueous solutions J Hazard Mater 152 1073ndash1081 doi101016jjhazmat200707085 PubMed 17804159
44 Šciban M Klašnja M Škrbic B (2008) Adsorption of copper ions fromwater by modified agricultural by-products Desalination 229 170ndash180doi101016jdesal200708017
45 Maumlkelauml MR Hildeacuten KS Hakala TK Hatakka A Lundell TK (2006)Expression and molecular properties of a new laccase of the white rotfungus Phlebia radiata grown on wood Curr Genet 50 323-333 doi101007s00294-006-0090-1 PubMed 16927090
46 Maumlkelauml MR Lundell T Hatakka A Hildeacuten K (2013) Effect of coppernutrient nitrogen and wood-supplement on the production of lignin-modifying enzymes by the white-rot fungus Phlebia radiata Fungal Biol117 62-70 doi101016jfunbio201211006 PubMed 23332834
47 Dwivedi UN Singha P Pandeya VP Kumara A (2011) Structurendashfunction relationship among bacterial fungal and plant laccases J MolCatal B Enzym 68 117-128 doi101016jmolcatb201011002
48 Gaitan IJ Medina SC Gonzaacutelez JC Rodriacuteguez A Espejo AacuteJ et al(2011) Evaluation of toxicity and degradation of a chlorophenol mixtureby the laccase produced by Trametes pubescens Bioresour Technol102 3632ndash3635 doi101016jbiortech201011040 PubMed21115244
49 Karp SG Faraco V Amore A Birolo L Giangrande C et al (2012)Characterization of laccase isoforms produced by Pleurotus ostreatusin solid state fermentation of sugarcane bagasse Bioresour Technol114 735-739 doi101016jbiortech201203058 PubMed 22487128
50 Nikitina OV Shleev SV Gorshina ES Rusinova TV Serezhenkov VAet al (2005) Isolation and purification of enzymes from ligninolyticcomplex of the basidial fungus Trametes pubescens (Schumach) Pilatand study of their properties Biochemistry (Mosc) 70 1274-1279 doi101007s10541-005-0259-0
51 Shleev S Nikitina O Christenson A Reimann CT Yaropolov AI et al(2007) Characterization of two new multiforms of Trametes pubescenslaccase Bioorg Chem 35 35-49 doi101016jbioorg200608001PubMed 16989887
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 13 September 2013 | Volume 8 | Issue 9 | e73721
52 Xiao YZ Chen Q Hang J Shi YY Wu J et al (2004) Selectiveinduction purification and characterization of a laccase isozyme fromthe basidiomycete Trametes sp AH28-2 Mycologia 96 26-35 doi1023073761984 PubMed 21148825
53 Muntildeoz C Guilleacuten F Martiacutenez AT Martiacutenez MJ (1997) Laccaseisoenzymes of Pleurotus eryngii characterization catalytic propertiesand participation in activation of molecular oxygen and Mn2+ oxidationAppl Environ Microbiol 63 2166-2174 PubMed 9172335
54 Morozova OV Shumakovich GP Gorbacheva MA Shleev SVYaropolov AI (2007) Blue laccases Biochemistry (Mosc) 721136-1150 doi101134S0006297907100112
55 Hildeacuten K Hakala TK Lundell T (2009) Thermotolerant andthermostable laccases Biotechnol Lett 31 1117-1128 doi101007s10529-009-9998-0 PubMed 19360388
56 Verma P Madamwar D (2002) Production of ligninolytic enzymes fordye decolorization by cocultivation of white-rot fungi Pleurotusostreatus and Phanerochaete chrysosporium under solid-statefermentation Appl Biochem Biotechnol 102-103 109-118 doi101385ABAB102-1031-6109 PubMed 12396115
57 Meza JC Lomascolo A Casalot L Sigoillot J-C Auria R (2005)Laccase production by Pycnoporus cinnabarinus grown on sugar-canebagasse Influence of ethanol vapours as inducer Proc Biochem 403365ndash3371 doi101016jprocbio200503004
58 Hossain SM Anantharaman N (2006) Activity enhancement ofligninolytic enzymes of Trametes versicolor with bagasse powder Afr JBiotechnol 5 189-194
59 Revankar MS Desai KM Lele SS (2007) Solid-state fermentation forenhanced production of laccase using indigenously isolatedGanoderma sp Appl Biochem Biotechnol 143 16-26 doi101007s12010-007-0029-0 PubMed 18025593
60 Velazquez-Cedeno MA Mata G Savoie J-M (2002) Waste-reducingcultivation of Pleurotus ostreatus and Pleurotus pulmonarius on coffeepulp changes in the production of some lignocellulolytic enzymesWorld J Microbiol Biotechnol 18 201ndash207 doi101023A1014999616381
61 Mata G Murrieta D Andreu LI (2005) Changes in lignocellulolyticenzyme activities in six Pleurotus spp strains cultivated on coffee pulpin confrontation with Trichoderma spp World J Microbiol Biotechnol 21143ndash150 doi101007s11274-004-3041-3
62 de Souza DF Tychanowicz GK de Souza CG Peralta RM (2006) Co-production of ligninolytic enzymes by Pleurotus pulmonarius on wheatbran solid state cultures J Basic Microbiol 46 126-134 doi101002jobm200510014 PubMed 16598826
63 Prasad KK Mohan SV Bhaskar YV Ramanaiah SV Babu VL et al(2005) Laccase production using Pleurotus ostreatus 1804 immobilizedon PUF cubes in batch and packed bed reactors influence of cultureconditions J Microbiol 43 301-307 PubMed 15995650
64 Songulashvili G Elisashvili V Wasser SP Nevo E Hadar Y (2007)Basidiomycetes laccase and manganese peroxidase activity in
submerged fermentation of food industry wastes Enzyme MicrobTechnol 41 57ndash61 doi101016jenzmictec200611024
65 Sathishkumar P Murugesan K Palvannan T (2010) Production oflaccase from Pleurotus florida using agro-wastes and efficientdecolorization of Reactive blue 198 J Basic Microbiol 50 360-367 doi101002jobm200900407 PubMed 20586068
66 Moldes D Gallego PP Rodriacuteguez Couto S Sanromaacuten A (2003) Grapeseeds the best lignocellulosic waste to produce laccase by solid statecultures of Trametes hirsuta Biotechnol Lett 25 491-495 doi101023A1022660230653 PubMed 12882277
67 Elisashvili V Kachlishvili E Penninckx M (2008) Effect of growthsubstrate method of fermentation and nitrogen source onlignocellulose-degrading enzymes production by white-rotbasidiomycetes J Ind Microbiol Biotechnol 35 1531-1538 doi101007s10295-008-0454-2 PubMed 18716810
68 Osma JF Saravia V Herrera JL Couto SR (2007) Mandarin peelingsthe best carbon source to produce laccase by static cultures ofTrametes pubescens Chemosphere 67 1677-1680 doi101016jchemosphere200611051 PubMed 17234250
69 Kahraman SS Gurdal IH (2002) Effect of synthetic and natural culturemedia on laccase production by white rot fungi Bioresour Technol 82215-217 doi101016S0960-8524(01)00193-6 PubMed 11991068
70 Iandolo D Piscitelli A Sannia G Faraco V (2011) Enzyme Productionby Solid Substrate Fermentation of Pleurotus ostreatus and Trametesversicolor on Tomato Pomace Appl Biochem Biotechnol 163 40-51doi101007s12010-010-9014-0 PubMed 20582639
71 Moldes D Lorenzo M Sanromaacuten MA (2004) Different proportions oflaccase isoenzymes produced by submerged cultures of Trametesversicolor grown on lignocellulosic wastes Biotechnol Lett 26 327-330doi101023BBILE000001545240213bf PubMed 15055770
72 Philippoussis A Diamantopoulou P Papadopoulou K Lakhtar HRoussos S et al (2010) Biomass laccase and endoglucanaseproduction by Lentinula edodes during solid state fermentation of reedgrass bean stalks and wheat straw residues World J MicrobiolBiotechnol 27 285-297
73 Fenice M Giovannozzi Sermanni G Federici F DrsquoAnnibale A (2003)Submerged and solid-state production of laccase and Mn-peroxidaseby Panus tigrinus on olive mill wastewater-based media J Biotechnol100 77-85 doi101016S0168-1656(02)00241-9 PubMed 12413788
74 Rodriguez-Couto S GundIn M Lorenzo M Sanromaacuten MAacute (2002)Screening of supports and inducers for laccase production by Trametesversicolor in semi-solid-state conditions Proc Biochem 38 249-255doi101016S0032-9592(02)00087-0
75 Fackler K Gradinger C Schmutzer M Tavzes C Burgert I et al (2007)Biotechnological wood modification with selective white-rot fungi and itsmolecular mechanisms Food Technol Biotechnol 45 269ndash276
76 Levin L Herrmann C Papinutti VL (2008) Optimization oflignocellulolytic enzyme production by the white-rot fungus Trametestrogii in solid-state fermentation using response surface methodologyBiochem Eng J 39 207-214 doi101016jbej200709004
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 14 September 2013 | Volume 8 | Issue 9 | e73721
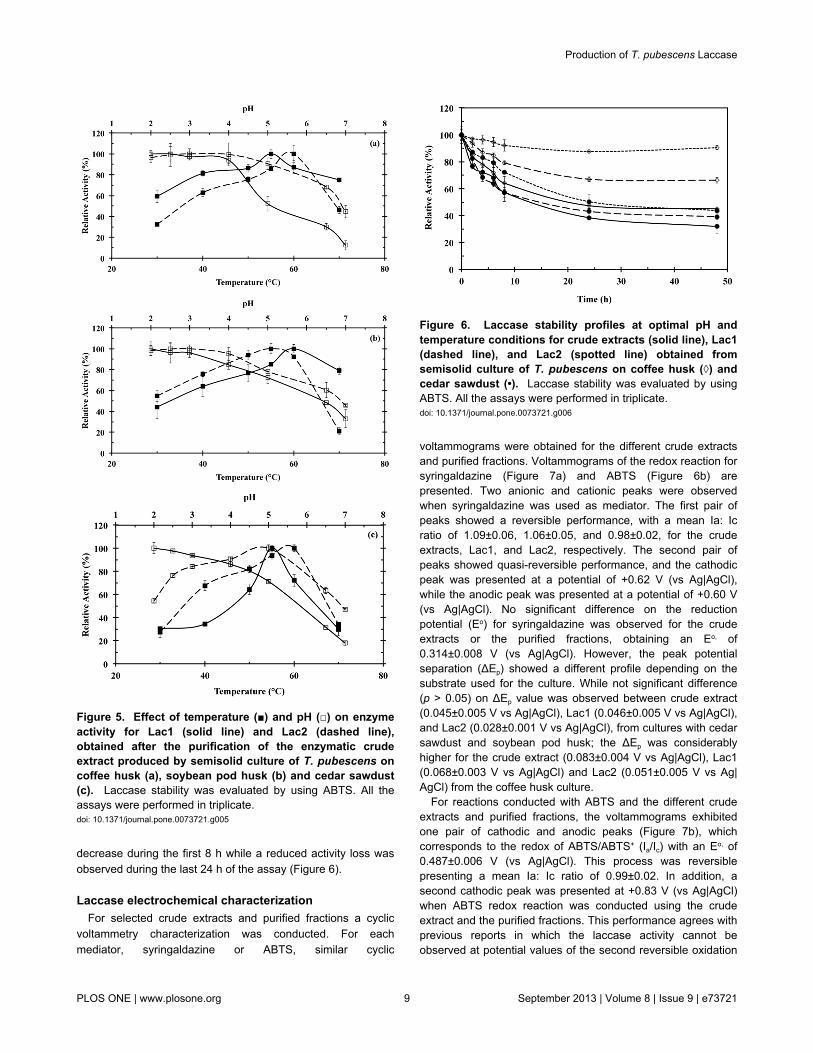
decrease during the first 8 h while a reduced activity loss wasobserved during the last 24 h of the assay (Figure 6)
Laccase electrochemical characterizationFor selected crude extracts and purified fractions a cyclic
voltammetry characterization was conducted For eachmediator syringaldazine or ABTS similar cyclic
Figure 5 Effect of temperature () and pH () on enzymeactivity for Lac1 (solid line) and Lac2 (dashed line)obtained after the purification of the enzymatic crudeextract produced by semisolid culture of T pubescens oncoffee husk (a) soybean pod husk (b) and cedar sawdust(c) Laccase stability was evaluated by using ABTS All theassays were performed in triplicatedoi 101371journalpone0073721g005
voltammograms were obtained for the different crude extractsand purified fractions Voltammograms of the redox reaction forsyringaldazine (Figure 7a) and ABTS (Figure 6b) arepresented Two anionic and cationic peaks were observedwhen syringaldazine was used as mediator The first pair ofpeaks showed a reversible performance with a mean Ia Icratio of 109plusmn006 106plusmn005 and 098plusmn002 for the crudeextracts Lac1 and Lac2 respectively The second pair ofpeaks showed quasi-reversible performance and the cathodicpeak was presented at a potential of +062 V (vs Ag|AgCl)while the anodic peak was presented at a potential of +060 V(vs Ag|AgCl) No significant difference on the reductionpotential (Eo) for syringaldazine was observed for the crudeextracts or the purified fractions obtaining an Eo of0314plusmn0008 V (vs Ag|AgCl) However the peak potentialseparation (ΔEp) showed a different profile depending on thesubstrate used for the culture While not significant difference(p gt 005) on ΔEp value was observed between crude extract(0045plusmn0005 V vs Ag|AgCl) Lac1 (0046plusmn0005 V vs Ag|AgCl)and Lac2 (0028plusmn0001 V vs Ag|AgCl) from cultures with cedarsawdust and soybean pod husk the ΔEp was considerablyhigher for the crude extract (0083plusmn0004 V vs Ag|AgCl) Lac1(0068plusmn0003 V vs Ag|AgCl) and Lac2 (0051plusmn0005 V vs Ag|AgCl) from the coffee husk culture
For reactions conducted with ABTS and the different crudeextracts and purified fractions the voltammograms exhibitedone pair of cathodic and anodic peaks (Figure 7b) whichcorresponds to the redox of ABTSABTS+ (IaIc) with an Eo of0487plusmn0006 V (vs Ag|AgCl) This process was reversiblepresenting a mean Ia Ic ratio of 099plusmn002 In addition asecond cathodic peak was presented at +083 V (vs Ag|AgCl)when ABTS redox reaction was conducted using the crudeextract and the purified fractions This performance agrees withprevious reports in which the laccase activity cannot beobserved at potential values of the second reversible oxidation
Figure 6 Laccase stability profiles at optimal pH andtemperature conditions for crude extracts (solid line) Lac1(dashed line) and Lac2 (spotted line) obtained fromsemisolid culture of T pubescens on coffee husk (loz) andcedar sawdust (bull) Laccase stability was evaluated by usingABTS All the assays were performed in triplicatedoi 101371journalpone0073721g006
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 9 September 2013 | Volume 8 | Issue 9 | e73721
of ABTS ABTS+ABTS2+ due to this catalytic process cannotbe detected by cyclic voltammetry [2526] Neverthelessvoltammograms for laccase crude extract from a soybean podhusk culture presented the second anodic peak which inaddition to the second cathodic peak corresponds to the redoxof ABTS+ABTS2+ (IIaIIc) with an Eo of 0942plusmn0004 V (vs Ag|AgCl) Contrary to the reactions conducted usingsyringaldazine as substrate the ΔEp did not present asignificant difference for the crude extracts and purifiedfractions 0248plusmn0016 V (vs Ag|AgCl)
Discussion
In this study we evaluated the effect of the culture mode ieSmC and SSC on laccase production by T pubescens CBS69694 using three different agro-industrial wastes (coffeehusk soybean pod husk and cedar sawdust) Furthermore theeffect of Cu+2 supplementation on laccase production was alsoevaluated and the crude extracts and the purified fractionswere partially characterized
Figure 7 Cyclic voltammograms of (a) syringaldazine atpH 50 using the crude extract and purified laccasefractions from semisolid culture of T pubescens on coffeehusk or (b) ABTS at pH 30 using the crude extract andpurified laccase fractions from semisolid culture of Tpubescens on soybean pod husk Nomenclature onlymediator (dotted line) crude extract (solid line) Lac1 fraction(dashed line) and Lac2 fraction (dashed and dotted line)doi 101371journalpone0073721g007
Substrate source for laccase production showed to be one ofthe most important factors for laccase production (Table 2)Cultures with coffee husk as substrate presented the highestlaccase production levels The observed differences in laccaseactivity for each substrate could be owed to their chemicalcomposition Coffea arabica is the coffee variety raised inColombia and its husk has a reported composition of 2308cellulose (2076 glucose and 183 cellobiose) 2385hemicellulose (1356 xylose 523 arabinose 256 aceticacid and 195 glucuronic acid) 2828 total lignin and071 ashes [27] In addition coffee husk contents 13caffeine 45 tannins and 124 pectins [28] On the otherhand Glycine max is the soybean species cultured inColombia and its pod husk composition varies from 504 to62 crude protein 608 to 143 hemicellulose 3715 to4208 acid detergent fiber 4436 to 6015 neutral detergentfiber 850 to 893 acid detergent lignin and 2074 to 2106uronic acid [29ndash31] Finally for cedar sawdust the majorcomponents are glucose (426-444) xylose (46-195)cellulose (383-410) klason lignin (227-278) and acidlignin (40-43) [32ndash34] Although lignin is one of the mostimportant compounds associated with laccase production bywhite-rot fungi we did not observe a clear correlation betweenlignin composition percentage and laccase activity In thatsense the high laccase activity observed for the cultures thatuse coffee husk could be associated with its content of caffeineand tannins which have been identified as potent inducers oflaccase gene expression [173536] In fact caffeine has beenused as inducer for laccase production by Rhizoctonia solaniobtaining an increase of 41-fold in laccase activity [37] Similarresults were also reported for the production of Coloriopsisgallica laccase [38]
Laccase gene expression is mainly up regulated by Cu2+although Mn2+ Fe3+ heavy metals 26-dimethoxy-14-benzoquinone H2O2 amphotericin B syringic acid tannic acidTween 80 soybean oil aromatic compounds andmicroclimatic changes (ie lower temperature and osmoticpressure) are also recognized as potent inducers [173536] Ingeneral Cu2+ supplementation increased the laccase activity inboth SmC and SSC However Cu2+concentration required toobtain the highest laccase activity varied depending on agro-industrial waste These differences can be attributed to thecopper presented in the agro-industrial waste and the copperavailability in the medium Copper can be found in amountsthat range from 156 to 180 mg kg-1 of coffee husk [39] andfrom 26 to 67 mg kg-1 of soybean pod husk (or soybean hull)[3940] Despite that no reports indicate the copper content inuntreated cedar wood an average of 10 mg kg-1 of wood hasbeen reported for wood used as biomass in six different Europecountries [41] This allows to consider the cedar sawdust as thesubstrate with the lowest content of copper On the other handcopper adsorption by sawdust and coffee husk has beenreported obtaining a Langmuir based maximum adsorptioncapacity (Qmax mg g-1 adsorbent) of 845 and 75 respectively[4243] Although no adsorption studies has been reportedusing untreated soybean pod husk Šćiban et al [44] reporteda Qmax for copper adsorption of 0085 mmol g-1 adsorbent (~ 54mg g-1) using treated soybean straws Based on this
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 10 September 2013 | Volume 8 | Issue 9 | e73721
information it can be expected that a higher amount ofexogenous copper would be required for laccase induction oncultures with cedar sawdust than with soybean pod husk andcoffee husk due to its lower content of copper In addition dueto the high copper-adsorption capacity of sawdust the amountof available copper in the culture would be reduced allowingthe laccase production at higher copper concentrations withoutreaching levels that might be toxic for the fungus This fact canbe observed in the SSC (Table 4) However for the SmC thisbehavior is not maintained for the cultures with soybean podhusk and coffee husk (Table 4) where the maximum laccaseactivity was obtained with a copper concentration of 05 and 5mM respectively This could be owed to a lower contactbetween the fungus and the substrate in the SmC whencompared to the SSC which is close related to the transport ofnutrients presented in the substrates
Production of extracellular laccases has been reported in alarge number of species of white-rot fungi strains grown onnatural substrates (Table 1) Although a laccase activity profileis not clearly observed based on the agro-industrial wasteculture mode and fungus strain most cases in which SmC andSSC were conducted using the same substrate and fungusstrain showed a higher laccase activity under SmC conditionsIn this study was observed a substrate dependentperformance where the most favorable condition to obtain thehighest laccase activity was the SSC The laccase activitiesobtained in the present study are comparable and in somecases higher than those reported for Pleurotus ostreatusPhanerochaete chrysosporium Pycnoporus cinnabarinus Tversicolor and Ganoderma sp cultured under SC and SmCconditions and using different agro-industrial wastes (Table 1)Semi-solid cultures using different agro-industrial wastes havebeen reported using T versicolor Phlebia radiata Tpubescens P ostreatus and C unicolor Rodriguez et al [16]screened different supports for laccase production by Tversicolor under SSC They obtained the highest laccaseactivity (1200 U L-1) when in the culture medium was presentedbarley bran In addition they were able to obtain a laccaseactivity of 1700 U L-1 when the culture was supplemented withxylidine as an inducer These results were obtained after 17 or18 days of culture and ABTS was used to determine theenzyme activity Maumlkelauml et al [4546] have studied theexpression of the genes involved in laccase production Pr-lac1and Pr-lac2 presented in P radiata and evaluate laccaseproduction in SSC using milled alder wood (Alnus incana) assubstrate They reported a maximum laccase activity of 3 microkatL-1 (~180 U L-1) at the 14th day of culture and usingsyringaldazine for its detection Osma et al [15] have studiedthe morphological changes and laccase production of strains Tpubescens P ostreatus C unicolor and T versicolor grownon wheat bran flakes using SSC They obtained a maximumlaccase production between 1397 and 2778 U L-1 after 10 and13 days of culture This enzyme activity is strain dependentand ABTS was used for its quantification Our results show agood correlation with reported data The highest laccaseactivity was obtained when T pubescens was grown on coffeehusk under SSC without copper (204 U L-1 evaluated withsyringaldazine or 1549 U L-1 evaluated with ABTS) which is in
some cases higher than reported ones This shows that underthe studied conditions the production of the laccase crudeextract is favored in SSC using coffee husk without the additionof copper which could favors the cost of the productionprocess
It has been reported for white rot-fungi the production ofmultiple laccase isoenzymes which are encoded by differentgenes and show different expression profiles stabilitiessubstrate affinities and molecular masses (ranging from 35 to140 kDa) [3647ndash49] Two isoforms have been identified in Tpubescens strains two monomeric (Lac1 and Lac2) enzymesof 67 kDa but with different pI (53 and 51) [50] and twolaccase-activity fractions (Lap1 and Lap2) with different pI (gt30 and 26) from which Lap2 is a monomeric enzyme of 65kDa [17] In this study we identify two laccase isoenzymesfrom T pubescens CBS 69694 that have different molecularmasses (120 and 60 kDa) and pI The purifications resultssuggest a more acidic pI for Lac1 (le 4) than Lac2 (ge 6) whichdiffers from a previous report for this fungus in which twofractions with laccase activity were obtained Lap1 and Lap2with pI gt 3 and of 26 respectively [17] Since in both casessimilar purification protocols were used and Lac1 and Lac2were recovered from the unbound and bound fractionsrespectively as Lap1 and Lap2 [17] this difference could bedue to the culture conditions used in each case (ie thepresence of agro-industrial wastes) Nevertheless the nanoLC-MSMS analysis showed that peptides of Lac1 and Lac2 werepresented in Lap1 (UniProtKBSwiss-Prot Q8TG93) and Lap2(UniProtKBSwiss-Prot Q8TG94) which have a predicted pI of43 and 47 respectively Lac1 and Lac2 peptides werepresented in Laccase-2 (UniProtKBSwiss-Prot Q99046) andLaccase-1 (UniProtKBSwiss-Prot Q99044) from T villosarespectively with 80 to 100 identity These T villosa laccaseisoenzymes have a predicted pI of 47 (laccase-1) and 60(laccase-2) and a 77 identity among them In addition Tvillosa laccase-2 is reported as a homodimer protein whilelaccase-1 is reported as monomeric protein Alignment of Lac1-Lap2 and Lac2-Lap1 from T pubescens CBS 69694 and Tvillosa showed a 96 and 70 identity respectively Finallysimilar peptides were identified for Lac1 and Lac2 from Tpubescens (Schumach) Pilaacutet (Syn Coriolus pubescens(Schum ex Fr) Queacutel) BCB 923-2 (Figure 3b) [51] Theseresults suggest that the purified fractions correspond to laccaseisoenzymes that differ from previously reported ones for this Tpubescens strain
As mentioned above laccases isoenzymes present differentstabilities and substrate affinities This behavior was alsoobserved for the laccase isoenzymes purified from Tpubescens with Lac1 showing lower laccase activity thanLac2 with ABTS as substrate However the difference inactivity between these isoenzymes depended on the agro-industrial waste used for the T pubescens growth suggestinga probable effect of the substrate composition on the laccasephysicochemical properties In addition it was observed thatthe total laccase activity presented in the UF retentates ishigher than the laccase activity of the purified fractionsregardless of the agro-industrial byproduct used for the Tpubescens growth This suggests a synergistic effect of Lac1
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 11 September 2013 | Volume 8 | Issue 9 | e73721
and Lac2 as have been reported for laccases obtained fromTrametes and Pleurotus strains [175253]
The maximum relative activity for the UF retentate waspresented at a pH from 20 to 40 regardless of the agro-industrial byproduct with a monotonical decrease at pHs abovethis range The relation between pH and type of electron donormolecule explains this behavior on laccase activity It has beenreported that when the substrate is an organic hydrogen donorie phenols the optimal pH ranges from 35 to 60 while thelaccase activity monotonically decreases from pH 25 to 70 ifthe substrate is an electron donor like ABTS [54] Despite thatat different pHs most of the purified laccase fractionspresented a similar performance as the UF retentates the Lac2fraction obtained from cedar sawdust differed from the previousobservation For this fraction the maximum activity wasobserved at pH 50 and despite that at acidic pH (ge 30) therelative activity was higher than 80 at lower pHs (20) itdecreased almost in a 50 Nevertheless at basic pHs therelative laccase activity presented a sharp decreased that iswell correlate with the other laccase fractions We are not ableto attribute this particular trend to the presence of multiplelaccase isoenzymes in the fraction since the SDS-PAGErevealed only one band when stained with ABTS and from theNanoLC-MSMS analysis no difference in the number ofpeptides and their sequence was detected We speculate thatthis particular behavior can be owed to a compound added inthe pre-treatment of the cedar wood that might be used forpreserving it that might affect the activity of Lac2 and that wasnot completely removed during the purification
Temperature stability evaluation showed that Lac1 is themost thermolabile fraction while the laccase activity reductionin the crude extracts did not seem to be the result of anadditive effect of the activity loss of the purified fractionsAlthough there are no reports of laccase stability under theselected conditions these results agree with previous reportsthat state that fungal laccases rapidly decrease their activity attemperatures near 60 degC and thermal stability correlates withthe temperature range of the growth of the source organism[55] In addition we previously reported for a laccase crudeextract obtained from culturing T pubescens on coffee husk anactivity loss of 26 and 10 at 60 and 50 degC respectivelyafter 8 h at pH 60 using syringaldazine as substrate [48]
Finally an electrochemical characterization was carried outfor selected crude extracts and purified fractions The results
showed a difference between the electrochemicalperformances of extracts and purified fractions withsyringaldazine or ABTS These differences could be owed tothe different mechanisms for electron transfer as discussedabove However the overall performance of the obtained cyclicvoltammograms for ABTS and syringaldazine agrees withresults reported by Fernaacutendez-Saacutenchez et al [25] who used asaturated calomel reference electrode
Conclusions
We have shown the potential of coffee husk soybean podhusk and cedar sawdust as substrates for the laccaseproduction In several countries like Brazil and Colombia theproduction of coffee and soybean is of great importance andthe generated volume of residues is high The results showedthat semi-solid culture of T pubescens using coffee huskallows to obtain an important amount of laccase which couldbe favored by the phenolic-compounds ie tannins presentedin coffee husk While supplementation of the culture mediumwith copper favored the production of laccase in submergedand semi-solid culture using soybean pod husk and cedarsawdust semi-solid culture using coffee husk which presentedthe highest laccase volumetric yield did not require coppersupplementation This is an economic advantage that shouldbe considered when scaling up the process In this study Tpubescens showed its ability to produce two independentlaccases regardless of the used substrate However a higherlaccase activity was observed for crude extracts rather than forpurified fractions This suggests that Lac1 and Lac2 might havea synergistic effect on the final laccase activity rather than anadditive effect Based on this study the investigation of laccaseproduction by evaluating other agro-industrial wastes in whichphenolic-compounds constitute an important percentage oftheir composition is of great interest to avoid the use ofinducers that would increase the production cost
Author Contributions
Conceived and designed the experiments JFO CJAD OFSPerformed the experiments JCG SCM AR Analyzed the dataJCG SCM AR JFO CJAD OFS Wrote the manuscript AR JFOCJAD OFS
References
1 Vandamme EJ (2009) Agro-industrial residue utilization for industrialbiotechnology products In P NigamA Pandey Biotechnology for Agro-Industrial Residues Utilisation Netherlands Springer Verlag pp 3-11
2 Martiacutenez AT Speranza M Ruiz-Duentildeas FJ Ferreira P Camarero S etal (2005) Biodegradation of lignocellulosics microbial chemical andenzymatic aspects of the fungal attack of lignin Int Microbiol 8195-204 PubMed 16200498
3 Saacutenchez O Sierra R Almeacuteciga-Diacuteaz CJ (2011) Delignification processof agro-industrial aastes an alternative to obtain fermentablecarbohydrates for producing fuel In M Manzanera Alternative FuelIntech 111-154
4 Wesenberg D Kyriakides I Agathos SN (2003) White-rot fungi andtheir enzymes for the treatment of industrial dye effluents BiotechnolAdv 22 161-187 doi101016jbiotechadv200308011 PubMed14623049
5 Morozova OV Shumakovich GP Shleev SV Yaropolov YI (2007)Laccase-mediator systems and their applications A review ApplBiochem Microbiol 43 523-535 doi101134S0003683807050055
6 Baldrian P (2006) Fungal laccases - occurrence and properties FEMSMicrobiol Rev 30 215-242 doi101111j1574-4976200500010xPubMed 16472305
7 Rodriacuteguez Couto S Herrera JLT (2006) Industrial and biotechnologicalapplications of laccases A review Biotechnol Adv 24 500-513 doi101016jbiotechadv200604003 PubMed 16716556
8 Osma JF Toca-Herrera JL Rodriacuteguez-Couto S (2010) Uses oflaccases in the food industry Enzyme Res 2010 2010 918761PubMed 21048873
9 Rodriguez S (2008) Exploitation of biological wastes for the productionof value-added products under solid-state fermentation conditionsBiotechnol J 3 859-870 doi101002biot200800031 PubMed18543242
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 12 September 2013 | Volume 8 | Issue 9 | e73721
10 Osma JF Toca-Herrera JL Rodriacuteguez-Couto S (2011) Cost analysis inlaccase production J Environ Manage 92 2907-2912 doi101016jjenvman201106052 PubMed 21775046
11 Jain A Morlok CK Henson JM (2013) Comparison of solid-state andsubmerged-state fermentation for the bioprocessing of switchgrass toethanol and acetate by Clostridium phytofermentans Appl MicrobiolBiotechnol 97 905-917 doi101007s00253-012-4511-4 PubMed23111595
12 Singhania RR Patel AK Soccol CR Pandey A (2009) Recentadvances in solid-state fermentation Biochem Eng J 44 13-18 doi101016jbej200810019
13 Economou CN Makri A Aggelis G Pavlou S Vayenas DV (2010)Semi-solid state fermentation of sweet sorghum for the biotechnologicalproduction of single cell oil Bioresour Technol 101 1385-1388 doi101016jbiortech200909028 PubMed 19781936
14 Rodriacuteguez CS Santoro R Cameselle C Sanroman A (1997) Laccaseproduction in semi-solid cultures of Phanerochaete chrysosporiumBiotechnol Lett 19 995-998 doi101023A1018495216946
15 Osma JF Moilanen U Toca-Herrera JL Rodriacuteguez-Couto S (2011)Morphology and laccase production of white-rot fungi grown on wheatbran flakes under semi-solid-state fermentation conditions FEMSMicrobiol Lett 318 27-34 doi101111j1574-6968201102234xPubMed 21291496
16 Couto SR Gundin M Lorenzo M Sanroman MN (2002) Screening ofsupports and inducers for laccase production by Trametes versicolor insemi-solid-state conditions Proc Biochem 38 249-255 doi101016S0032-9592(02)00087-0
17 Galhaup C Goller S Peterbauer CK Strauss J Haltrich D (2002)Characterization of the major laccase isoenzyme from Trametespubescens and regulation of its synthesis by metal ions Microbiology148 2159-2169 PubMed 12101303
18 Sievers F Wilm A Dineen D Gibson TJ Karplus K et al (2011) Fastscalable generation of high-quality protein multiple sequencealignments using Clustal Omega Mol Syst Biol 7 539 PubMed21988835
19 Childs RE Bardsley WG (1975) The steady-state kinetics of peroxidasewith 22-azino-di-(3-ethyl-benzthiazoline-6-sulphonic acid) aschromogen Biochem J 145 93-103 PubMed 1191252
20 Harkin JM Obst JR (1973) Syringaldazine an Effective Reagent forDetecting Laccase and Peroxidase in Fungi Experientia 29 382-387
21 Nikupaavola ML Raaska L Itavaara M (1990) Detection of white-rotfungi by a non-roxic stain Mycol Res 94 27-31 doi101016S0953-7562(09)81260-4
22 Gonzalez LF Sarria V Sanchez OF (2010) Degradation ofchlorophenols by sequential biological-advanced oxidative processusing Trametes pubescens and TiO(2)UV Bioresour Technol 1013493-3499
23 Waterborg JH (2009) The Lowry method for protein quantitation In JWalker The Protein Protocols Handbook Humana Press pp 7-10
24 Harkin JM Obst JR (1973) Syringaldazine an effective reagent fordetecting laccase and peroxidase in fungi Cell Mol Life Sci 29381-387 doi101007BF01926734
25 Fernaacutendez-Saacutenchez C Tzanov T Guumlbitz GM Cavaco-Paulo A (2002)Voltammetric monitoring of laccase-catalysed mediated reactionsBioelectrochemistry 58 149-156 doi101016S1567-5394(02)00119-6PubMed 12414320
26 Bourbonnais R Leech D Paice MG (1998) Electrochemical analysis ofthe interactions of laccase mediators with lignin model compoundsBiochim Biophys Acta 1379 381ndash390 doi101016S0304-4165(97)00117-7 PubMed 9545600
27 Ferraz F Silva S (2009) Characterization of coffee husk biomass forbiotechnological purposes N Biotechnol 25 S256-S256 doi101016jnbt200906573
28 Pandey A Soccol CR Nigam P Brand D Mohan R et al (2000)Biotechnological potential of coffee pulp and coffee husk forbioprocesses Biochem Eng J 6 153-162 doi101016S1369-703X(00)00084-X PubMed 10959086
29 Sruamsiri S Silman P (2008) Nutritive composition of soybean by-products and nutrient digestibility of soybean pod husk Maejo Int J SciTechnol 2 568-576
30 Romkaew J Nagaya Y Goto M Suzuki J Umezaki T (2008) Poddehiscence in relation to chemical components of pod shell in soybeanPlant Prod Sci 11 278-282 doi101626pps11278
31 Nakamanee G Polbumroong T Phaikaew C (2003) Alternative use ofcrop waste to increase milk production Proceedings of the 8th Meetingof the Regional Working Group on Grazing and Feed Resources forSoutheast Asia Kuala Lumpur Malaysia 66 - 70
32 Kurek B Gaudard F (2000) Oxidation of spruce wood sawdust byMnO(2) plus oxalate a biochemical investigation J Agric Food Chem48 3058-3062 doi101021jf000015g PubMed 10898665
33 Weil J Brewer M Hendrickson R Sarikaya A Ladisc MR (1998)Continuous pH monitoring during pretreatment of yellow poplar woodsawdust by pressure cooking in water Appl Biochem Biotechnol 70-7299-111 doi101007BF02920127
34 Weil J Sarikaya A Rau S-L Goetz J Ladisch CM et al (1997)Pretreatment of yellow poplar sawdust by pressure cooking in waterAppl Biochem Biotechnol 68 21-40 doi101007BF02785978
35 Dekker RF Barbosa AM Giese EC Godoy SD Covizzi LG (2007)Influence of nutrients on enhancing laccase production byBotryosphaeria rhodina MAMB-05 Int Microbiol 10 177-185 PubMed18075999
36 Xiao YZ Hong YZ Li JF Hang J Tong PG et al (2006) Cloning ofnovel laccase isozyme genes from Trametes sp AH28-2 and analysesof their differential expression Appl Microbiol Biotechnol 71 493-501doi101007s00253-005-0188-2 PubMed 16283298
37 Crowe JD Olsson S (2001) Induction of laccase activity in Rhizoctoniasolani by antagonistic Pseudomonas fluorescens strains and a range ofchemical treatments Appl Environ Microbiol 67 2088-2094 doi101128AEM6752088-20942001 PubMed 11319086
38 Yaguumle S Terroacuten MC Gonzaacutelez T Zapico E Bocchini P et al (2000)Biotreatment of tannin-rich beer-factory wastewater with white-rotbasidiomycete Coriolopsis gallica monitored by pyrolysisgaschromatographymass spectrometry Rapid Commun Mass Spectrom14 905-910 doi101002(SICI)1097-0231(20000530)1410 PubMed10825255
39 Gowda NKS Ramana JV Prasad CS Singh K (2004) Micronutrientcontent of certain tropical conventional and unconventional feedresources of southern India Trop Anim Heatlth Pro 36 77-94 doi101023BTROP0000009522309491d PubMed 14979561
40 DePeters EJ Fadel JG Arana MJ Ohanesian N Etchebarne MA et al(2000) Variability in the chemical composition of seventeen selectedby-product feedstuffs used by the California dairy industry Prof AnimSci 16 69-99
41 Obernberger I Thek G (2004) Physical characterisation and chemicalcomposition of densified biomass fuels with regard to their combustionbehaviour Biomass Bioenerg 27 653-669 doi101016jbiombioe200307006
42 Larous S Meniai A-H Lehocine MB (2005) Experimental study of theremoval of copper from aqueous solutions by adsorption usingsawdust Desalination 185 483ndash490 doi101016jdesal200503090
43 Oliveira WE Franca AS Oliveira LS Rocha SD (2008) Untreatedcoffee husks as biosorbents for the removal of heavy metals fromaqueous solutions J Hazard Mater 152 1073ndash1081 doi101016jjhazmat200707085 PubMed 17804159
44 Šciban M Klašnja M Škrbic B (2008) Adsorption of copper ions fromwater by modified agricultural by-products Desalination 229 170ndash180doi101016jdesal200708017
45 Maumlkelauml MR Hildeacuten KS Hakala TK Hatakka A Lundell TK (2006)Expression and molecular properties of a new laccase of the white rotfungus Phlebia radiata grown on wood Curr Genet 50 323-333 doi101007s00294-006-0090-1 PubMed 16927090
46 Maumlkelauml MR Lundell T Hatakka A Hildeacuten K (2013) Effect of coppernutrient nitrogen and wood-supplement on the production of lignin-modifying enzymes by the white-rot fungus Phlebia radiata Fungal Biol117 62-70 doi101016jfunbio201211006 PubMed 23332834
47 Dwivedi UN Singha P Pandeya VP Kumara A (2011) Structurendashfunction relationship among bacterial fungal and plant laccases J MolCatal B Enzym 68 117-128 doi101016jmolcatb201011002
48 Gaitan IJ Medina SC Gonzaacutelez JC Rodriacuteguez A Espejo AacuteJ et al(2011) Evaluation of toxicity and degradation of a chlorophenol mixtureby the laccase produced by Trametes pubescens Bioresour Technol102 3632ndash3635 doi101016jbiortech201011040 PubMed21115244
49 Karp SG Faraco V Amore A Birolo L Giangrande C et al (2012)Characterization of laccase isoforms produced by Pleurotus ostreatusin solid state fermentation of sugarcane bagasse Bioresour Technol114 735-739 doi101016jbiortech201203058 PubMed 22487128
50 Nikitina OV Shleev SV Gorshina ES Rusinova TV Serezhenkov VAet al (2005) Isolation and purification of enzymes from ligninolyticcomplex of the basidial fungus Trametes pubescens (Schumach) Pilatand study of their properties Biochemistry (Mosc) 70 1274-1279 doi101007s10541-005-0259-0
51 Shleev S Nikitina O Christenson A Reimann CT Yaropolov AI et al(2007) Characterization of two new multiforms of Trametes pubescenslaccase Bioorg Chem 35 35-49 doi101016jbioorg200608001PubMed 16989887
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 13 September 2013 | Volume 8 | Issue 9 | e73721
52 Xiao YZ Chen Q Hang J Shi YY Wu J et al (2004) Selectiveinduction purification and characterization of a laccase isozyme fromthe basidiomycete Trametes sp AH28-2 Mycologia 96 26-35 doi1023073761984 PubMed 21148825
53 Muntildeoz C Guilleacuten F Martiacutenez AT Martiacutenez MJ (1997) Laccaseisoenzymes of Pleurotus eryngii characterization catalytic propertiesand participation in activation of molecular oxygen and Mn2+ oxidationAppl Environ Microbiol 63 2166-2174 PubMed 9172335
54 Morozova OV Shumakovich GP Gorbacheva MA Shleev SVYaropolov AI (2007) Blue laccases Biochemistry (Mosc) 721136-1150 doi101134S0006297907100112
55 Hildeacuten K Hakala TK Lundell T (2009) Thermotolerant andthermostable laccases Biotechnol Lett 31 1117-1128 doi101007s10529-009-9998-0 PubMed 19360388
56 Verma P Madamwar D (2002) Production of ligninolytic enzymes fordye decolorization by cocultivation of white-rot fungi Pleurotusostreatus and Phanerochaete chrysosporium under solid-statefermentation Appl Biochem Biotechnol 102-103 109-118 doi101385ABAB102-1031-6109 PubMed 12396115
57 Meza JC Lomascolo A Casalot L Sigoillot J-C Auria R (2005)Laccase production by Pycnoporus cinnabarinus grown on sugar-canebagasse Influence of ethanol vapours as inducer Proc Biochem 403365ndash3371 doi101016jprocbio200503004
58 Hossain SM Anantharaman N (2006) Activity enhancement ofligninolytic enzymes of Trametes versicolor with bagasse powder Afr JBiotechnol 5 189-194
59 Revankar MS Desai KM Lele SS (2007) Solid-state fermentation forenhanced production of laccase using indigenously isolatedGanoderma sp Appl Biochem Biotechnol 143 16-26 doi101007s12010-007-0029-0 PubMed 18025593
60 Velazquez-Cedeno MA Mata G Savoie J-M (2002) Waste-reducingcultivation of Pleurotus ostreatus and Pleurotus pulmonarius on coffeepulp changes in the production of some lignocellulolytic enzymesWorld J Microbiol Biotechnol 18 201ndash207 doi101023A1014999616381
61 Mata G Murrieta D Andreu LI (2005) Changes in lignocellulolyticenzyme activities in six Pleurotus spp strains cultivated on coffee pulpin confrontation with Trichoderma spp World J Microbiol Biotechnol 21143ndash150 doi101007s11274-004-3041-3
62 de Souza DF Tychanowicz GK de Souza CG Peralta RM (2006) Co-production of ligninolytic enzymes by Pleurotus pulmonarius on wheatbran solid state cultures J Basic Microbiol 46 126-134 doi101002jobm200510014 PubMed 16598826
63 Prasad KK Mohan SV Bhaskar YV Ramanaiah SV Babu VL et al(2005) Laccase production using Pleurotus ostreatus 1804 immobilizedon PUF cubes in batch and packed bed reactors influence of cultureconditions J Microbiol 43 301-307 PubMed 15995650
64 Songulashvili G Elisashvili V Wasser SP Nevo E Hadar Y (2007)Basidiomycetes laccase and manganese peroxidase activity in
submerged fermentation of food industry wastes Enzyme MicrobTechnol 41 57ndash61 doi101016jenzmictec200611024
65 Sathishkumar P Murugesan K Palvannan T (2010) Production oflaccase from Pleurotus florida using agro-wastes and efficientdecolorization of Reactive blue 198 J Basic Microbiol 50 360-367 doi101002jobm200900407 PubMed 20586068
66 Moldes D Gallego PP Rodriacuteguez Couto S Sanromaacuten A (2003) Grapeseeds the best lignocellulosic waste to produce laccase by solid statecultures of Trametes hirsuta Biotechnol Lett 25 491-495 doi101023A1022660230653 PubMed 12882277
67 Elisashvili V Kachlishvili E Penninckx M (2008) Effect of growthsubstrate method of fermentation and nitrogen source onlignocellulose-degrading enzymes production by white-rotbasidiomycetes J Ind Microbiol Biotechnol 35 1531-1538 doi101007s10295-008-0454-2 PubMed 18716810
68 Osma JF Saravia V Herrera JL Couto SR (2007) Mandarin peelingsthe best carbon source to produce laccase by static cultures ofTrametes pubescens Chemosphere 67 1677-1680 doi101016jchemosphere200611051 PubMed 17234250
69 Kahraman SS Gurdal IH (2002) Effect of synthetic and natural culturemedia on laccase production by white rot fungi Bioresour Technol 82215-217 doi101016S0960-8524(01)00193-6 PubMed 11991068
70 Iandolo D Piscitelli A Sannia G Faraco V (2011) Enzyme Productionby Solid Substrate Fermentation of Pleurotus ostreatus and Trametesversicolor on Tomato Pomace Appl Biochem Biotechnol 163 40-51doi101007s12010-010-9014-0 PubMed 20582639
71 Moldes D Lorenzo M Sanromaacuten MA (2004) Different proportions oflaccase isoenzymes produced by submerged cultures of Trametesversicolor grown on lignocellulosic wastes Biotechnol Lett 26 327-330doi101023BBILE000001545240213bf PubMed 15055770
72 Philippoussis A Diamantopoulou P Papadopoulou K Lakhtar HRoussos S et al (2010) Biomass laccase and endoglucanaseproduction by Lentinula edodes during solid state fermentation of reedgrass bean stalks and wheat straw residues World J MicrobiolBiotechnol 27 285-297
73 Fenice M Giovannozzi Sermanni G Federici F DrsquoAnnibale A (2003)Submerged and solid-state production of laccase and Mn-peroxidaseby Panus tigrinus on olive mill wastewater-based media J Biotechnol100 77-85 doi101016S0168-1656(02)00241-9 PubMed 12413788
74 Rodriguez-Couto S GundIn M Lorenzo M Sanromaacuten MAacute (2002)Screening of supports and inducers for laccase production by Trametesversicolor in semi-solid-state conditions Proc Biochem 38 249-255doi101016S0032-9592(02)00087-0
75 Fackler K Gradinger C Schmutzer M Tavzes C Burgert I et al (2007)Biotechnological wood modification with selective white-rot fungi and itsmolecular mechanisms Food Technol Biotechnol 45 269ndash276
76 Levin L Herrmann C Papinutti VL (2008) Optimization oflignocellulolytic enzyme production by the white-rot fungus Trametestrogii in solid-state fermentation using response surface methodologyBiochem Eng J 39 207-214 doi101016jbej200709004
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 14 September 2013 | Volume 8 | Issue 9 | e73721

of ABTS ABTS+ABTS2+ due to this catalytic process cannotbe detected by cyclic voltammetry [2526] Neverthelessvoltammograms for laccase crude extract from a soybean podhusk culture presented the second anodic peak which inaddition to the second cathodic peak corresponds to the redoxof ABTS+ABTS2+ (IIaIIc) with an Eo of 0942plusmn0004 V (vs Ag|AgCl) Contrary to the reactions conducted usingsyringaldazine as substrate the ΔEp did not present asignificant difference for the crude extracts and purifiedfractions 0248plusmn0016 V (vs Ag|AgCl)
Discussion
In this study we evaluated the effect of the culture mode ieSmC and SSC on laccase production by T pubescens CBS69694 using three different agro-industrial wastes (coffeehusk soybean pod husk and cedar sawdust) Furthermore theeffect of Cu+2 supplementation on laccase production was alsoevaluated and the crude extracts and the purified fractionswere partially characterized
Figure 7 Cyclic voltammograms of (a) syringaldazine atpH 50 using the crude extract and purified laccasefractions from semisolid culture of T pubescens on coffeehusk or (b) ABTS at pH 30 using the crude extract andpurified laccase fractions from semisolid culture of Tpubescens on soybean pod husk Nomenclature onlymediator (dotted line) crude extract (solid line) Lac1 fraction(dashed line) and Lac2 fraction (dashed and dotted line)doi 101371journalpone0073721g007
Substrate source for laccase production showed to be one ofthe most important factors for laccase production (Table 2)Cultures with coffee husk as substrate presented the highestlaccase production levels The observed differences in laccaseactivity for each substrate could be owed to their chemicalcomposition Coffea arabica is the coffee variety raised inColombia and its husk has a reported composition of 2308cellulose (2076 glucose and 183 cellobiose) 2385hemicellulose (1356 xylose 523 arabinose 256 aceticacid and 195 glucuronic acid) 2828 total lignin and071 ashes [27] In addition coffee husk contents 13caffeine 45 tannins and 124 pectins [28] On the otherhand Glycine max is the soybean species cultured inColombia and its pod husk composition varies from 504 to62 crude protein 608 to 143 hemicellulose 3715 to4208 acid detergent fiber 4436 to 6015 neutral detergentfiber 850 to 893 acid detergent lignin and 2074 to 2106uronic acid [29ndash31] Finally for cedar sawdust the majorcomponents are glucose (426-444) xylose (46-195)cellulose (383-410) klason lignin (227-278) and acidlignin (40-43) [32ndash34] Although lignin is one of the mostimportant compounds associated with laccase production bywhite-rot fungi we did not observe a clear correlation betweenlignin composition percentage and laccase activity In thatsense the high laccase activity observed for the cultures thatuse coffee husk could be associated with its content of caffeineand tannins which have been identified as potent inducers oflaccase gene expression [173536] In fact caffeine has beenused as inducer for laccase production by Rhizoctonia solaniobtaining an increase of 41-fold in laccase activity [37] Similarresults were also reported for the production of Coloriopsisgallica laccase [38]
Laccase gene expression is mainly up regulated by Cu2+although Mn2+ Fe3+ heavy metals 26-dimethoxy-14-benzoquinone H2O2 amphotericin B syringic acid tannic acidTween 80 soybean oil aromatic compounds andmicroclimatic changes (ie lower temperature and osmoticpressure) are also recognized as potent inducers [173536] Ingeneral Cu2+ supplementation increased the laccase activity inboth SmC and SSC However Cu2+concentration required toobtain the highest laccase activity varied depending on agro-industrial waste These differences can be attributed to thecopper presented in the agro-industrial waste and the copperavailability in the medium Copper can be found in amountsthat range from 156 to 180 mg kg-1 of coffee husk [39] andfrom 26 to 67 mg kg-1 of soybean pod husk (or soybean hull)[3940] Despite that no reports indicate the copper content inuntreated cedar wood an average of 10 mg kg-1 of wood hasbeen reported for wood used as biomass in six different Europecountries [41] This allows to consider the cedar sawdust as thesubstrate with the lowest content of copper On the other handcopper adsorption by sawdust and coffee husk has beenreported obtaining a Langmuir based maximum adsorptioncapacity (Qmax mg g-1 adsorbent) of 845 and 75 respectively[4243] Although no adsorption studies has been reportedusing untreated soybean pod husk Šćiban et al [44] reporteda Qmax for copper adsorption of 0085 mmol g-1 adsorbent (~ 54mg g-1) using treated soybean straws Based on this
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 10 September 2013 | Volume 8 | Issue 9 | e73721
information it can be expected that a higher amount ofexogenous copper would be required for laccase induction oncultures with cedar sawdust than with soybean pod husk andcoffee husk due to its lower content of copper In addition dueto the high copper-adsorption capacity of sawdust the amountof available copper in the culture would be reduced allowingthe laccase production at higher copper concentrations withoutreaching levels that might be toxic for the fungus This fact canbe observed in the SSC (Table 4) However for the SmC thisbehavior is not maintained for the cultures with soybean podhusk and coffee husk (Table 4) where the maximum laccaseactivity was obtained with a copper concentration of 05 and 5mM respectively This could be owed to a lower contactbetween the fungus and the substrate in the SmC whencompared to the SSC which is close related to the transport ofnutrients presented in the substrates
Production of extracellular laccases has been reported in alarge number of species of white-rot fungi strains grown onnatural substrates (Table 1) Although a laccase activity profileis not clearly observed based on the agro-industrial wasteculture mode and fungus strain most cases in which SmC andSSC were conducted using the same substrate and fungusstrain showed a higher laccase activity under SmC conditionsIn this study was observed a substrate dependentperformance where the most favorable condition to obtain thehighest laccase activity was the SSC The laccase activitiesobtained in the present study are comparable and in somecases higher than those reported for Pleurotus ostreatusPhanerochaete chrysosporium Pycnoporus cinnabarinus Tversicolor and Ganoderma sp cultured under SC and SmCconditions and using different agro-industrial wastes (Table 1)Semi-solid cultures using different agro-industrial wastes havebeen reported using T versicolor Phlebia radiata Tpubescens P ostreatus and C unicolor Rodriguez et al [16]screened different supports for laccase production by Tversicolor under SSC They obtained the highest laccaseactivity (1200 U L-1) when in the culture medium was presentedbarley bran In addition they were able to obtain a laccaseactivity of 1700 U L-1 when the culture was supplemented withxylidine as an inducer These results were obtained after 17 or18 days of culture and ABTS was used to determine theenzyme activity Maumlkelauml et al [4546] have studied theexpression of the genes involved in laccase production Pr-lac1and Pr-lac2 presented in P radiata and evaluate laccaseproduction in SSC using milled alder wood (Alnus incana) assubstrate They reported a maximum laccase activity of 3 microkatL-1 (~180 U L-1) at the 14th day of culture and usingsyringaldazine for its detection Osma et al [15] have studiedthe morphological changes and laccase production of strains Tpubescens P ostreatus C unicolor and T versicolor grownon wheat bran flakes using SSC They obtained a maximumlaccase production between 1397 and 2778 U L-1 after 10 and13 days of culture This enzyme activity is strain dependentand ABTS was used for its quantification Our results show agood correlation with reported data The highest laccaseactivity was obtained when T pubescens was grown on coffeehusk under SSC without copper (204 U L-1 evaluated withsyringaldazine or 1549 U L-1 evaluated with ABTS) which is in
some cases higher than reported ones This shows that underthe studied conditions the production of the laccase crudeextract is favored in SSC using coffee husk without the additionof copper which could favors the cost of the productionprocess
It has been reported for white rot-fungi the production ofmultiple laccase isoenzymes which are encoded by differentgenes and show different expression profiles stabilitiessubstrate affinities and molecular masses (ranging from 35 to140 kDa) [3647ndash49] Two isoforms have been identified in Tpubescens strains two monomeric (Lac1 and Lac2) enzymesof 67 kDa but with different pI (53 and 51) [50] and twolaccase-activity fractions (Lap1 and Lap2) with different pI (gt30 and 26) from which Lap2 is a monomeric enzyme of 65kDa [17] In this study we identify two laccase isoenzymesfrom T pubescens CBS 69694 that have different molecularmasses (120 and 60 kDa) and pI The purifications resultssuggest a more acidic pI for Lac1 (le 4) than Lac2 (ge 6) whichdiffers from a previous report for this fungus in which twofractions with laccase activity were obtained Lap1 and Lap2with pI gt 3 and of 26 respectively [17] Since in both casessimilar purification protocols were used and Lac1 and Lac2were recovered from the unbound and bound fractionsrespectively as Lap1 and Lap2 [17] this difference could bedue to the culture conditions used in each case (ie thepresence of agro-industrial wastes) Nevertheless the nanoLC-MSMS analysis showed that peptides of Lac1 and Lac2 werepresented in Lap1 (UniProtKBSwiss-Prot Q8TG93) and Lap2(UniProtKBSwiss-Prot Q8TG94) which have a predicted pI of43 and 47 respectively Lac1 and Lac2 peptides werepresented in Laccase-2 (UniProtKBSwiss-Prot Q99046) andLaccase-1 (UniProtKBSwiss-Prot Q99044) from T villosarespectively with 80 to 100 identity These T villosa laccaseisoenzymes have a predicted pI of 47 (laccase-1) and 60(laccase-2) and a 77 identity among them In addition Tvillosa laccase-2 is reported as a homodimer protein whilelaccase-1 is reported as monomeric protein Alignment of Lac1-Lap2 and Lac2-Lap1 from T pubescens CBS 69694 and Tvillosa showed a 96 and 70 identity respectively Finallysimilar peptides were identified for Lac1 and Lac2 from Tpubescens (Schumach) Pilaacutet (Syn Coriolus pubescens(Schum ex Fr) Queacutel) BCB 923-2 (Figure 3b) [51] Theseresults suggest that the purified fractions correspond to laccaseisoenzymes that differ from previously reported ones for this Tpubescens strain
As mentioned above laccases isoenzymes present differentstabilities and substrate affinities This behavior was alsoobserved for the laccase isoenzymes purified from Tpubescens with Lac1 showing lower laccase activity thanLac2 with ABTS as substrate However the difference inactivity between these isoenzymes depended on the agro-industrial waste used for the T pubescens growth suggestinga probable effect of the substrate composition on the laccasephysicochemical properties In addition it was observed thatthe total laccase activity presented in the UF retentates ishigher than the laccase activity of the purified fractionsregardless of the agro-industrial byproduct used for the Tpubescens growth This suggests a synergistic effect of Lac1
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 11 September 2013 | Volume 8 | Issue 9 | e73721
and Lac2 as have been reported for laccases obtained fromTrametes and Pleurotus strains [175253]
The maximum relative activity for the UF retentate waspresented at a pH from 20 to 40 regardless of the agro-industrial byproduct with a monotonical decrease at pHs abovethis range The relation between pH and type of electron donormolecule explains this behavior on laccase activity It has beenreported that when the substrate is an organic hydrogen donorie phenols the optimal pH ranges from 35 to 60 while thelaccase activity monotonically decreases from pH 25 to 70 ifthe substrate is an electron donor like ABTS [54] Despite thatat different pHs most of the purified laccase fractionspresented a similar performance as the UF retentates the Lac2fraction obtained from cedar sawdust differed from the previousobservation For this fraction the maximum activity wasobserved at pH 50 and despite that at acidic pH (ge 30) therelative activity was higher than 80 at lower pHs (20) itdecreased almost in a 50 Nevertheless at basic pHs therelative laccase activity presented a sharp decreased that iswell correlate with the other laccase fractions We are not ableto attribute this particular trend to the presence of multiplelaccase isoenzymes in the fraction since the SDS-PAGErevealed only one band when stained with ABTS and from theNanoLC-MSMS analysis no difference in the number ofpeptides and their sequence was detected We speculate thatthis particular behavior can be owed to a compound added inthe pre-treatment of the cedar wood that might be used forpreserving it that might affect the activity of Lac2 and that wasnot completely removed during the purification
Temperature stability evaluation showed that Lac1 is themost thermolabile fraction while the laccase activity reductionin the crude extracts did not seem to be the result of anadditive effect of the activity loss of the purified fractionsAlthough there are no reports of laccase stability under theselected conditions these results agree with previous reportsthat state that fungal laccases rapidly decrease their activity attemperatures near 60 degC and thermal stability correlates withthe temperature range of the growth of the source organism[55] In addition we previously reported for a laccase crudeextract obtained from culturing T pubescens on coffee husk anactivity loss of 26 and 10 at 60 and 50 degC respectivelyafter 8 h at pH 60 using syringaldazine as substrate [48]
Finally an electrochemical characterization was carried outfor selected crude extracts and purified fractions The results
showed a difference between the electrochemicalperformances of extracts and purified fractions withsyringaldazine or ABTS These differences could be owed tothe different mechanisms for electron transfer as discussedabove However the overall performance of the obtained cyclicvoltammograms for ABTS and syringaldazine agrees withresults reported by Fernaacutendez-Saacutenchez et al [25] who used asaturated calomel reference electrode
Conclusions
We have shown the potential of coffee husk soybean podhusk and cedar sawdust as substrates for the laccaseproduction In several countries like Brazil and Colombia theproduction of coffee and soybean is of great importance andthe generated volume of residues is high The results showedthat semi-solid culture of T pubescens using coffee huskallows to obtain an important amount of laccase which couldbe favored by the phenolic-compounds ie tannins presentedin coffee husk While supplementation of the culture mediumwith copper favored the production of laccase in submergedand semi-solid culture using soybean pod husk and cedarsawdust semi-solid culture using coffee husk which presentedthe highest laccase volumetric yield did not require coppersupplementation This is an economic advantage that shouldbe considered when scaling up the process In this study Tpubescens showed its ability to produce two independentlaccases regardless of the used substrate However a higherlaccase activity was observed for crude extracts rather than forpurified fractions This suggests that Lac1 and Lac2 might havea synergistic effect on the final laccase activity rather than anadditive effect Based on this study the investigation of laccaseproduction by evaluating other agro-industrial wastes in whichphenolic-compounds constitute an important percentage oftheir composition is of great interest to avoid the use ofinducers that would increase the production cost
Author Contributions
Conceived and designed the experiments JFO CJAD OFSPerformed the experiments JCG SCM AR Analyzed the dataJCG SCM AR JFO CJAD OFS Wrote the manuscript AR JFOCJAD OFS
References
1 Vandamme EJ (2009) Agro-industrial residue utilization for industrialbiotechnology products In P NigamA Pandey Biotechnology for Agro-Industrial Residues Utilisation Netherlands Springer Verlag pp 3-11
2 Martiacutenez AT Speranza M Ruiz-Duentildeas FJ Ferreira P Camarero S etal (2005) Biodegradation of lignocellulosics microbial chemical andenzymatic aspects of the fungal attack of lignin Int Microbiol 8195-204 PubMed 16200498
3 Saacutenchez O Sierra R Almeacuteciga-Diacuteaz CJ (2011) Delignification processof agro-industrial aastes an alternative to obtain fermentablecarbohydrates for producing fuel In M Manzanera Alternative FuelIntech 111-154
4 Wesenberg D Kyriakides I Agathos SN (2003) White-rot fungi andtheir enzymes for the treatment of industrial dye effluents BiotechnolAdv 22 161-187 doi101016jbiotechadv200308011 PubMed14623049
5 Morozova OV Shumakovich GP Shleev SV Yaropolov YI (2007)Laccase-mediator systems and their applications A review ApplBiochem Microbiol 43 523-535 doi101134S0003683807050055
6 Baldrian P (2006) Fungal laccases - occurrence and properties FEMSMicrobiol Rev 30 215-242 doi101111j1574-4976200500010xPubMed 16472305
7 Rodriacuteguez Couto S Herrera JLT (2006) Industrial and biotechnologicalapplications of laccases A review Biotechnol Adv 24 500-513 doi101016jbiotechadv200604003 PubMed 16716556
8 Osma JF Toca-Herrera JL Rodriacuteguez-Couto S (2010) Uses oflaccases in the food industry Enzyme Res 2010 2010 918761PubMed 21048873
9 Rodriguez S (2008) Exploitation of biological wastes for the productionof value-added products under solid-state fermentation conditionsBiotechnol J 3 859-870 doi101002biot200800031 PubMed18543242
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 12 September 2013 | Volume 8 | Issue 9 | e73721
10 Osma JF Toca-Herrera JL Rodriacuteguez-Couto S (2011) Cost analysis inlaccase production J Environ Manage 92 2907-2912 doi101016jjenvman201106052 PubMed 21775046
11 Jain A Morlok CK Henson JM (2013) Comparison of solid-state andsubmerged-state fermentation for the bioprocessing of switchgrass toethanol and acetate by Clostridium phytofermentans Appl MicrobiolBiotechnol 97 905-917 doi101007s00253-012-4511-4 PubMed23111595
12 Singhania RR Patel AK Soccol CR Pandey A (2009) Recentadvances in solid-state fermentation Biochem Eng J 44 13-18 doi101016jbej200810019
13 Economou CN Makri A Aggelis G Pavlou S Vayenas DV (2010)Semi-solid state fermentation of sweet sorghum for the biotechnologicalproduction of single cell oil Bioresour Technol 101 1385-1388 doi101016jbiortech200909028 PubMed 19781936
14 Rodriacuteguez CS Santoro R Cameselle C Sanroman A (1997) Laccaseproduction in semi-solid cultures of Phanerochaete chrysosporiumBiotechnol Lett 19 995-998 doi101023A1018495216946
15 Osma JF Moilanen U Toca-Herrera JL Rodriacuteguez-Couto S (2011)Morphology and laccase production of white-rot fungi grown on wheatbran flakes under semi-solid-state fermentation conditions FEMSMicrobiol Lett 318 27-34 doi101111j1574-6968201102234xPubMed 21291496
16 Couto SR Gundin M Lorenzo M Sanroman MN (2002) Screening ofsupports and inducers for laccase production by Trametes versicolor insemi-solid-state conditions Proc Biochem 38 249-255 doi101016S0032-9592(02)00087-0
17 Galhaup C Goller S Peterbauer CK Strauss J Haltrich D (2002)Characterization of the major laccase isoenzyme from Trametespubescens and regulation of its synthesis by metal ions Microbiology148 2159-2169 PubMed 12101303
18 Sievers F Wilm A Dineen D Gibson TJ Karplus K et al (2011) Fastscalable generation of high-quality protein multiple sequencealignments using Clustal Omega Mol Syst Biol 7 539 PubMed21988835
19 Childs RE Bardsley WG (1975) The steady-state kinetics of peroxidasewith 22-azino-di-(3-ethyl-benzthiazoline-6-sulphonic acid) aschromogen Biochem J 145 93-103 PubMed 1191252
20 Harkin JM Obst JR (1973) Syringaldazine an Effective Reagent forDetecting Laccase and Peroxidase in Fungi Experientia 29 382-387
21 Nikupaavola ML Raaska L Itavaara M (1990) Detection of white-rotfungi by a non-roxic stain Mycol Res 94 27-31 doi101016S0953-7562(09)81260-4
22 Gonzalez LF Sarria V Sanchez OF (2010) Degradation ofchlorophenols by sequential biological-advanced oxidative processusing Trametes pubescens and TiO(2)UV Bioresour Technol 1013493-3499
23 Waterborg JH (2009) The Lowry method for protein quantitation In JWalker The Protein Protocols Handbook Humana Press pp 7-10
24 Harkin JM Obst JR (1973) Syringaldazine an effective reagent fordetecting laccase and peroxidase in fungi Cell Mol Life Sci 29381-387 doi101007BF01926734
25 Fernaacutendez-Saacutenchez C Tzanov T Guumlbitz GM Cavaco-Paulo A (2002)Voltammetric monitoring of laccase-catalysed mediated reactionsBioelectrochemistry 58 149-156 doi101016S1567-5394(02)00119-6PubMed 12414320
26 Bourbonnais R Leech D Paice MG (1998) Electrochemical analysis ofthe interactions of laccase mediators with lignin model compoundsBiochim Biophys Acta 1379 381ndash390 doi101016S0304-4165(97)00117-7 PubMed 9545600
27 Ferraz F Silva S (2009) Characterization of coffee husk biomass forbiotechnological purposes N Biotechnol 25 S256-S256 doi101016jnbt200906573
28 Pandey A Soccol CR Nigam P Brand D Mohan R et al (2000)Biotechnological potential of coffee pulp and coffee husk forbioprocesses Biochem Eng J 6 153-162 doi101016S1369-703X(00)00084-X PubMed 10959086
29 Sruamsiri S Silman P (2008) Nutritive composition of soybean by-products and nutrient digestibility of soybean pod husk Maejo Int J SciTechnol 2 568-576
30 Romkaew J Nagaya Y Goto M Suzuki J Umezaki T (2008) Poddehiscence in relation to chemical components of pod shell in soybeanPlant Prod Sci 11 278-282 doi101626pps11278
31 Nakamanee G Polbumroong T Phaikaew C (2003) Alternative use ofcrop waste to increase milk production Proceedings of the 8th Meetingof the Regional Working Group on Grazing and Feed Resources forSoutheast Asia Kuala Lumpur Malaysia 66 - 70
32 Kurek B Gaudard F (2000) Oxidation of spruce wood sawdust byMnO(2) plus oxalate a biochemical investigation J Agric Food Chem48 3058-3062 doi101021jf000015g PubMed 10898665
33 Weil J Brewer M Hendrickson R Sarikaya A Ladisc MR (1998)Continuous pH monitoring during pretreatment of yellow poplar woodsawdust by pressure cooking in water Appl Biochem Biotechnol 70-7299-111 doi101007BF02920127
34 Weil J Sarikaya A Rau S-L Goetz J Ladisch CM et al (1997)Pretreatment of yellow poplar sawdust by pressure cooking in waterAppl Biochem Biotechnol 68 21-40 doi101007BF02785978
35 Dekker RF Barbosa AM Giese EC Godoy SD Covizzi LG (2007)Influence of nutrients on enhancing laccase production byBotryosphaeria rhodina MAMB-05 Int Microbiol 10 177-185 PubMed18075999
36 Xiao YZ Hong YZ Li JF Hang J Tong PG et al (2006) Cloning ofnovel laccase isozyme genes from Trametes sp AH28-2 and analysesof their differential expression Appl Microbiol Biotechnol 71 493-501doi101007s00253-005-0188-2 PubMed 16283298
37 Crowe JD Olsson S (2001) Induction of laccase activity in Rhizoctoniasolani by antagonistic Pseudomonas fluorescens strains and a range ofchemical treatments Appl Environ Microbiol 67 2088-2094 doi101128AEM6752088-20942001 PubMed 11319086
38 Yaguumle S Terroacuten MC Gonzaacutelez T Zapico E Bocchini P et al (2000)Biotreatment of tannin-rich beer-factory wastewater with white-rotbasidiomycete Coriolopsis gallica monitored by pyrolysisgaschromatographymass spectrometry Rapid Commun Mass Spectrom14 905-910 doi101002(SICI)1097-0231(20000530)1410 PubMed10825255
39 Gowda NKS Ramana JV Prasad CS Singh K (2004) Micronutrientcontent of certain tropical conventional and unconventional feedresources of southern India Trop Anim Heatlth Pro 36 77-94 doi101023BTROP0000009522309491d PubMed 14979561
40 DePeters EJ Fadel JG Arana MJ Ohanesian N Etchebarne MA et al(2000) Variability in the chemical composition of seventeen selectedby-product feedstuffs used by the California dairy industry Prof AnimSci 16 69-99
41 Obernberger I Thek G (2004) Physical characterisation and chemicalcomposition of densified biomass fuels with regard to their combustionbehaviour Biomass Bioenerg 27 653-669 doi101016jbiombioe200307006
42 Larous S Meniai A-H Lehocine MB (2005) Experimental study of theremoval of copper from aqueous solutions by adsorption usingsawdust Desalination 185 483ndash490 doi101016jdesal200503090
43 Oliveira WE Franca AS Oliveira LS Rocha SD (2008) Untreatedcoffee husks as biosorbents for the removal of heavy metals fromaqueous solutions J Hazard Mater 152 1073ndash1081 doi101016jjhazmat200707085 PubMed 17804159
44 Šciban M Klašnja M Škrbic B (2008) Adsorption of copper ions fromwater by modified agricultural by-products Desalination 229 170ndash180doi101016jdesal200708017
45 Maumlkelauml MR Hildeacuten KS Hakala TK Hatakka A Lundell TK (2006)Expression and molecular properties of a new laccase of the white rotfungus Phlebia radiata grown on wood Curr Genet 50 323-333 doi101007s00294-006-0090-1 PubMed 16927090
46 Maumlkelauml MR Lundell T Hatakka A Hildeacuten K (2013) Effect of coppernutrient nitrogen and wood-supplement on the production of lignin-modifying enzymes by the white-rot fungus Phlebia radiata Fungal Biol117 62-70 doi101016jfunbio201211006 PubMed 23332834
47 Dwivedi UN Singha P Pandeya VP Kumara A (2011) Structurendashfunction relationship among bacterial fungal and plant laccases J MolCatal B Enzym 68 117-128 doi101016jmolcatb201011002
48 Gaitan IJ Medina SC Gonzaacutelez JC Rodriacuteguez A Espejo AacuteJ et al(2011) Evaluation of toxicity and degradation of a chlorophenol mixtureby the laccase produced by Trametes pubescens Bioresour Technol102 3632ndash3635 doi101016jbiortech201011040 PubMed21115244
49 Karp SG Faraco V Amore A Birolo L Giangrande C et al (2012)Characterization of laccase isoforms produced by Pleurotus ostreatusin solid state fermentation of sugarcane bagasse Bioresour Technol114 735-739 doi101016jbiortech201203058 PubMed 22487128
50 Nikitina OV Shleev SV Gorshina ES Rusinova TV Serezhenkov VAet al (2005) Isolation and purification of enzymes from ligninolyticcomplex of the basidial fungus Trametes pubescens (Schumach) Pilatand study of their properties Biochemistry (Mosc) 70 1274-1279 doi101007s10541-005-0259-0
51 Shleev S Nikitina O Christenson A Reimann CT Yaropolov AI et al(2007) Characterization of two new multiforms of Trametes pubescenslaccase Bioorg Chem 35 35-49 doi101016jbioorg200608001PubMed 16989887
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 13 September 2013 | Volume 8 | Issue 9 | e73721
52 Xiao YZ Chen Q Hang J Shi YY Wu J et al (2004) Selectiveinduction purification and characterization of a laccase isozyme fromthe basidiomycete Trametes sp AH28-2 Mycologia 96 26-35 doi1023073761984 PubMed 21148825
53 Muntildeoz C Guilleacuten F Martiacutenez AT Martiacutenez MJ (1997) Laccaseisoenzymes of Pleurotus eryngii characterization catalytic propertiesand participation in activation of molecular oxygen and Mn2+ oxidationAppl Environ Microbiol 63 2166-2174 PubMed 9172335
54 Morozova OV Shumakovich GP Gorbacheva MA Shleev SVYaropolov AI (2007) Blue laccases Biochemistry (Mosc) 721136-1150 doi101134S0006297907100112
55 Hildeacuten K Hakala TK Lundell T (2009) Thermotolerant andthermostable laccases Biotechnol Lett 31 1117-1128 doi101007s10529-009-9998-0 PubMed 19360388
56 Verma P Madamwar D (2002) Production of ligninolytic enzymes fordye decolorization by cocultivation of white-rot fungi Pleurotusostreatus and Phanerochaete chrysosporium under solid-statefermentation Appl Biochem Biotechnol 102-103 109-118 doi101385ABAB102-1031-6109 PubMed 12396115
57 Meza JC Lomascolo A Casalot L Sigoillot J-C Auria R (2005)Laccase production by Pycnoporus cinnabarinus grown on sugar-canebagasse Influence of ethanol vapours as inducer Proc Biochem 403365ndash3371 doi101016jprocbio200503004
58 Hossain SM Anantharaman N (2006) Activity enhancement ofligninolytic enzymes of Trametes versicolor with bagasse powder Afr JBiotechnol 5 189-194
59 Revankar MS Desai KM Lele SS (2007) Solid-state fermentation forenhanced production of laccase using indigenously isolatedGanoderma sp Appl Biochem Biotechnol 143 16-26 doi101007s12010-007-0029-0 PubMed 18025593
60 Velazquez-Cedeno MA Mata G Savoie J-M (2002) Waste-reducingcultivation of Pleurotus ostreatus and Pleurotus pulmonarius on coffeepulp changes in the production of some lignocellulolytic enzymesWorld J Microbiol Biotechnol 18 201ndash207 doi101023A1014999616381
61 Mata G Murrieta D Andreu LI (2005) Changes in lignocellulolyticenzyme activities in six Pleurotus spp strains cultivated on coffee pulpin confrontation with Trichoderma spp World J Microbiol Biotechnol 21143ndash150 doi101007s11274-004-3041-3
62 de Souza DF Tychanowicz GK de Souza CG Peralta RM (2006) Co-production of ligninolytic enzymes by Pleurotus pulmonarius on wheatbran solid state cultures J Basic Microbiol 46 126-134 doi101002jobm200510014 PubMed 16598826
63 Prasad KK Mohan SV Bhaskar YV Ramanaiah SV Babu VL et al(2005) Laccase production using Pleurotus ostreatus 1804 immobilizedon PUF cubes in batch and packed bed reactors influence of cultureconditions J Microbiol 43 301-307 PubMed 15995650
64 Songulashvili G Elisashvili V Wasser SP Nevo E Hadar Y (2007)Basidiomycetes laccase and manganese peroxidase activity in
submerged fermentation of food industry wastes Enzyme MicrobTechnol 41 57ndash61 doi101016jenzmictec200611024
65 Sathishkumar P Murugesan K Palvannan T (2010) Production oflaccase from Pleurotus florida using agro-wastes and efficientdecolorization of Reactive blue 198 J Basic Microbiol 50 360-367 doi101002jobm200900407 PubMed 20586068
66 Moldes D Gallego PP Rodriacuteguez Couto S Sanromaacuten A (2003) Grapeseeds the best lignocellulosic waste to produce laccase by solid statecultures of Trametes hirsuta Biotechnol Lett 25 491-495 doi101023A1022660230653 PubMed 12882277
67 Elisashvili V Kachlishvili E Penninckx M (2008) Effect of growthsubstrate method of fermentation and nitrogen source onlignocellulose-degrading enzymes production by white-rotbasidiomycetes J Ind Microbiol Biotechnol 35 1531-1538 doi101007s10295-008-0454-2 PubMed 18716810
68 Osma JF Saravia V Herrera JL Couto SR (2007) Mandarin peelingsthe best carbon source to produce laccase by static cultures ofTrametes pubescens Chemosphere 67 1677-1680 doi101016jchemosphere200611051 PubMed 17234250
69 Kahraman SS Gurdal IH (2002) Effect of synthetic and natural culturemedia on laccase production by white rot fungi Bioresour Technol 82215-217 doi101016S0960-8524(01)00193-6 PubMed 11991068
70 Iandolo D Piscitelli A Sannia G Faraco V (2011) Enzyme Productionby Solid Substrate Fermentation of Pleurotus ostreatus and Trametesversicolor on Tomato Pomace Appl Biochem Biotechnol 163 40-51doi101007s12010-010-9014-0 PubMed 20582639
71 Moldes D Lorenzo M Sanromaacuten MA (2004) Different proportions oflaccase isoenzymes produced by submerged cultures of Trametesversicolor grown on lignocellulosic wastes Biotechnol Lett 26 327-330doi101023BBILE000001545240213bf PubMed 15055770
72 Philippoussis A Diamantopoulou P Papadopoulou K Lakhtar HRoussos S et al (2010) Biomass laccase and endoglucanaseproduction by Lentinula edodes during solid state fermentation of reedgrass bean stalks and wheat straw residues World J MicrobiolBiotechnol 27 285-297
73 Fenice M Giovannozzi Sermanni G Federici F DrsquoAnnibale A (2003)Submerged and solid-state production of laccase and Mn-peroxidaseby Panus tigrinus on olive mill wastewater-based media J Biotechnol100 77-85 doi101016S0168-1656(02)00241-9 PubMed 12413788
74 Rodriguez-Couto S GundIn M Lorenzo M Sanromaacuten MAacute (2002)Screening of supports and inducers for laccase production by Trametesversicolor in semi-solid-state conditions Proc Biochem 38 249-255doi101016S0032-9592(02)00087-0
75 Fackler K Gradinger C Schmutzer M Tavzes C Burgert I et al (2007)Biotechnological wood modification with selective white-rot fungi and itsmolecular mechanisms Food Technol Biotechnol 45 269ndash276
76 Levin L Herrmann C Papinutti VL (2008) Optimization oflignocellulolytic enzyme production by the white-rot fungus Trametestrogii in solid-state fermentation using response surface methodologyBiochem Eng J 39 207-214 doi101016jbej200709004
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 14 September 2013 | Volume 8 | Issue 9 | e73721

information it can be expected that a higher amount ofexogenous copper would be required for laccase induction oncultures with cedar sawdust than with soybean pod husk andcoffee husk due to its lower content of copper In addition dueto the high copper-adsorption capacity of sawdust the amountof available copper in the culture would be reduced allowingthe laccase production at higher copper concentrations withoutreaching levels that might be toxic for the fungus This fact canbe observed in the SSC (Table 4) However for the SmC thisbehavior is not maintained for the cultures with soybean podhusk and coffee husk (Table 4) where the maximum laccaseactivity was obtained with a copper concentration of 05 and 5mM respectively This could be owed to a lower contactbetween the fungus and the substrate in the SmC whencompared to the SSC which is close related to the transport ofnutrients presented in the substrates
Production of extracellular laccases has been reported in alarge number of species of white-rot fungi strains grown onnatural substrates (Table 1) Although a laccase activity profileis not clearly observed based on the agro-industrial wasteculture mode and fungus strain most cases in which SmC andSSC were conducted using the same substrate and fungusstrain showed a higher laccase activity under SmC conditionsIn this study was observed a substrate dependentperformance where the most favorable condition to obtain thehighest laccase activity was the SSC The laccase activitiesobtained in the present study are comparable and in somecases higher than those reported for Pleurotus ostreatusPhanerochaete chrysosporium Pycnoporus cinnabarinus Tversicolor and Ganoderma sp cultured under SC and SmCconditions and using different agro-industrial wastes (Table 1)Semi-solid cultures using different agro-industrial wastes havebeen reported using T versicolor Phlebia radiata Tpubescens P ostreatus and C unicolor Rodriguez et al [16]screened different supports for laccase production by Tversicolor under SSC They obtained the highest laccaseactivity (1200 U L-1) when in the culture medium was presentedbarley bran In addition they were able to obtain a laccaseactivity of 1700 U L-1 when the culture was supplemented withxylidine as an inducer These results were obtained after 17 or18 days of culture and ABTS was used to determine theenzyme activity Maumlkelauml et al [4546] have studied theexpression of the genes involved in laccase production Pr-lac1and Pr-lac2 presented in P radiata and evaluate laccaseproduction in SSC using milled alder wood (Alnus incana) assubstrate They reported a maximum laccase activity of 3 microkatL-1 (~180 U L-1) at the 14th day of culture and usingsyringaldazine for its detection Osma et al [15] have studiedthe morphological changes and laccase production of strains Tpubescens P ostreatus C unicolor and T versicolor grownon wheat bran flakes using SSC They obtained a maximumlaccase production between 1397 and 2778 U L-1 after 10 and13 days of culture This enzyme activity is strain dependentand ABTS was used for its quantification Our results show agood correlation with reported data The highest laccaseactivity was obtained when T pubescens was grown on coffeehusk under SSC without copper (204 U L-1 evaluated withsyringaldazine or 1549 U L-1 evaluated with ABTS) which is in
some cases higher than reported ones This shows that underthe studied conditions the production of the laccase crudeextract is favored in SSC using coffee husk without the additionof copper which could favors the cost of the productionprocess
It has been reported for white rot-fungi the production ofmultiple laccase isoenzymes which are encoded by differentgenes and show different expression profiles stabilitiessubstrate affinities and molecular masses (ranging from 35 to140 kDa) [3647ndash49] Two isoforms have been identified in Tpubescens strains two monomeric (Lac1 and Lac2) enzymesof 67 kDa but with different pI (53 and 51) [50] and twolaccase-activity fractions (Lap1 and Lap2) with different pI (gt30 and 26) from which Lap2 is a monomeric enzyme of 65kDa [17] In this study we identify two laccase isoenzymesfrom T pubescens CBS 69694 that have different molecularmasses (120 and 60 kDa) and pI The purifications resultssuggest a more acidic pI for Lac1 (le 4) than Lac2 (ge 6) whichdiffers from a previous report for this fungus in which twofractions with laccase activity were obtained Lap1 and Lap2with pI gt 3 and of 26 respectively [17] Since in both casessimilar purification protocols were used and Lac1 and Lac2were recovered from the unbound and bound fractionsrespectively as Lap1 and Lap2 [17] this difference could bedue to the culture conditions used in each case (ie thepresence of agro-industrial wastes) Nevertheless the nanoLC-MSMS analysis showed that peptides of Lac1 and Lac2 werepresented in Lap1 (UniProtKBSwiss-Prot Q8TG93) and Lap2(UniProtKBSwiss-Prot Q8TG94) which have a predicted pI of43 and 47 respectively Lac1 and Lac2 peptides werepresented in Laccase-2 (UniProtKBSwiss-Prot Q99046) andLaccase-1 (UniProtKBSwiss-Prot Q99044) from T villosarespectively with 80 to 100 identity These T villosa laccaseisoenzymes have a predicted pI of 47 (laccase-1) and 60(laccase-2) and a 77 identity among them In addition Tvillosa laccase-2 is reported as a homodimer protein whilelaccase-1 is reported as monomeric protein Alignment of Lac1-Lap2 and Lac2-Lap1 from T pubescens CBS 69694 and Tvillosa showed a 96 and 70 identity respectively Finallysimilar peptides were identified for Lac1 and Lac2 from Tpubescens (Schumach) Pilaacutet (Syn Coriolus pubescens(Schum ex Fr) Queacutel) BCB 923-2 (Figure 3b) [51] Theseresults suggest that the purified fractions correspond to laccaseisoenzymes that differ from previously reported ones for this Tpubescens strain
As mentioned above laccases isoenzymes present differentstabilities and substrate affinities This behavior was alsoobserved for the laccase isoenzymes purified from Tpubescens with Lac1 showing lower laccase activity thanLac2 with ABTS as substrate However the difference inactivity between these isoenzymes depended on the agro-industrial waste used for the T pubescens growth suggestinga probable effect of the substrate composition on the laccasephysicochemical properties In addition it was observed thatthe total laccase activity presented in the UF retentates ishigher than the laccase activity of the purified fractionsregardless of the agro-industrial byproduct used for the Tpubescens growth This suggests a synergistic effect of Lac1
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 11 September 2013 | Volume 8 | Issue 9 | e73721
and Lac2 as have been reported for laccases obtained fromTrametes and Pleurotus strains [175253]
The maximum relative activity for the UF retentate waspresented at a pH from 20 to 40 regardless of the agro-industrial byproduct with a monotonical decrease at pHs abovethis range The relation between pH and type of electron donormolecule explains this behavior on laccase activity It has beenreported that when the substrate is an organic hydrogen donorie phenols the optimal pH ranges from 35 to 60 while thelaccase activity monotonically decreases from pH 25 to 70 ifthe substrate is an electron donor like ABTS [54] Despite thatat different pHs most of the purified laccase fractionspresented a similar performance as the UF retentates the Lac2fraction obtained from cedar sawdust differed from the previousobservation For this fraction the maximum activity wasobserved at pH 50 and despite that at acidic pH (ge 30) therelative activity was higher than 80 at lower pHs (20) itdecreased almost in a 50 Nevertheless at basic pHs therelative laccase activity presented a sharp decreased that iswell correlate with the other laccase fractions We are not ableto attribute this particular trend to the presence of multiplelaccase isoenzymes in the fraction since the SDS-PAGErevealed only one band when stained with ABTS and from theNanoLC-MSMS analysis no difference in the number ofpeptides and their sequence was detected We speculate thatthis particular behavior can be owed to a compound added inthe pre-treatment of the cedar wood that might be used forpreserving it that might affect the activity of Lac2 and that wasnot completely removed during the purification
Temperature stability evaluation showed that Lac1 is themost thermolabile fraction while the laccase activity reductionin the crude extracts did not seem to be the result of anadditive effect of the activity loss of the purified fractionsAlthough there are no reports of laccase stability under theselected conditions these results agree with previous reportsthat state that fungal laccases rapidly decrease their activity attemperatures near 60 degC and thermal stability correlates withthe temperature range of the growth of the source organism[55] In addition we previously reported for a laccase crudeextract obtained from culturing T pubescens on coffee husk anactivity loss of 26 and 10 at 60 and 50 degC respectivelyafter 8 h at pH 60 using syringaldazine as substrate [48]
Finally an electrochemical characterization was carried outfor selected crude extracts and purified fractions The results
showed a difference between the electrochemicalperformances of extracts and purified fractions withsyringaldazine or ABTS These differences could be owed tothe different mechanisms for electron transfer as discussedabove However the overall performance of the obtained cyclicvoltammograms for ABTS and syringaldazine agrees withresults reported by Fernaacutendez-Saacutenchez et al [25] who used asaturated calomel reference electrode
Conclusions
We have shown the potential of coffee husk soybean podhusk and cedar sawdust as substrates for the laccaseproduction In several countries like Brazil and Colombia theproduction of coffee and soybean is of great importance andthe generated volume of residues is high The results showedthat semi-solid culture of T pubescens using coffee huskallows to obtain an important amount of laccase which couldbe favored by the phenolic-compounds ie tannins presentedin coffee husk While supplementation of the culture mediumwith copper favored the production of laccase in submergedand semi-solid culture using soybean pod husk and cedarsawdust semi-solid culture using coffee husk which presentedthe highest laccase volumetric yield did not require coppersupplementation This is an economic advantage that shouldbe considered when scaling up the process In this study Tpubescens showed its ability to produce two independentlaccases regardless of the used substrate However a higherlaccase activity was observed for crude extracts rather than forpurified fractions This suggests that Lac1 and Lac2 might havea synergistic effect on the final laccase activity rather than anadditive effect Based on this study the investigation of laccaseproduction by evaluating other agro-industrial wastes in whichphenolic-compounds constitute an important percentage oftheir composition is of great interest to avoid the use ofinducers that would increase the production cost
Author Contributions
Conceived and designed the experiments JFO CJAD OFSPerformed the experiments JCG SCM AR Analyzed the dataJCG SCM AR JFO CJAD OFS Wrote the manuscript AR JFOCJAD OFS
References
1 Vandamme EJ (2009) Agro-industrial residue utilization for industrialbiotechnology products In P NigamA Pandey Biotechnology for Agro-Industrial Residues Utilisation Netherlands Springer Verlag pp 3-11
2 Martiacutenez AT Speranza M Ruiz-Duentildeas FJ Ferreira P Camarero S etal (2005) Biodegradation of lignocellulosics microbial chemical andenzymatic aspects of the fungal attack of lignin Int Microbiol 8195-204 PubMed 16200498
3 Saacutenchez O Sierra R Almeacuteciga-Diacuteaz CJ (2011) Delignification processof agro-industrial aastes an alternative to obtain fermentablecarbohydrates for producing fuel In M Manzanera Alternative FuelIntech 111-154
4 Wesenberg D Kyriakides I Agathos SN (2003) White-rot fungi andtheir enzymes for the treatment of industrial dye effluents BiotechnolAdv 22 161-187 doi101016jbiotechadv200308011 PubMed14623049
5 Morozova OV Shumakovich GP Shleev SV Yaropolov YI (2007)Laccase-mediator systems and their applications A review ApplBiochem Microbiol 43 523-535 doi101134S0003683807050055
6 Baldrian P (2006) Fungal laccases - occurrence and properties FEMSMicrobiol Rev 30 215-242 doi101111j1574-4976200500010xPubMed 16472305
7 Rodriacuteguez Couto S Herrera JLT (2006) Industrial and biotechnologicalapplications of laccases A review Biotechnol Adv 24 500-513 doi101016jbiotechadv200604003 PubMed 16716556
8 Osma JF Toca-Herrera JL Rodriacuteguez-Couto S (2010) Uses oflaccases in the food industry Enzyme Res 2010 2010 918761PubMed 21048873
9 Rodriguez S (2008) Exploitation of biological wastes for the productionof value-added products under solid-state fermentation conditionsBiotechnol J 3 859-870 doi101002biot200800031 PubMed18543242
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 12 September 2013 | Volume 8 | Issue 9 | e73721
10 Osma JF Toca-Herrera JL Rodriacuteguez-Couto S (2011) Cost analysis inlaccase production J Environ Manage 92 2907-2912 doi101016jjenvman201106052 PubMed 21775046
11 Jain A Morlok CK Henson JM (2013) Comparison of solid-state andsubmerged-state fermentation for the bioprocessing of switchgrass toethanol and acetate by Clostridium phytofermentans Appl MicrobiolBiotechnol 97 905-917 doi101007s00253-012-4511-4 PubMed23111595
12 Singhania RR Patel AK Soccol CR Pandey A (2009) Recentadvances in solid-state fermentation Biochem Eng J 44 13-18 doi101016jbej200810019
13 Economou CN Makri A Aggelis G Pavlou S Vayenas DV (2010)Semi-solid state fermentation of sweet sorghum for the biotechnologicalproduction of single cell oil Bioresour Technol 101 1385-1388 doi101016jbiortech200909028 PubMed 19781936
14 Rodriacuteguez CS Santoro R Cameselle C Sanroman A (1997) Laccaseproduction in semi-solid cultures of Phanerochaete chrysosporiumBiotechnol Lett 19 995-998 doi101023A1018495216946
15 Osma JF Moilanen U Toca-Herrera JL Rodriacuteguez-Couto S (2011)Morphology and laccase production of white-rot fungi grown on wheatbran flakes under semi-solid-state fermentation conditions FEMSMicrobiol Lett 318 27-34 doi101111j1574-6968201102234xPubMed 21291496
16 Couto SR Gundin M Lorenzo M Sanroman MN (2002) Screening ofsupports and inducers for laccase production by Trametes versicolor insemi-solid-state conditions Proc Biochem 38 249-255 doi101016S0032-9592(02)00087-0
17 Galhaup C Goller S Peterbauer CK Strauss J Haltrich D (2002)Characterization of the major laccase isoenzyme from Trametespubescens and regulation of its synthesis by metal ions Microbiology148 2159-2169 PubMed 12101303
18 Sievers F Wilm A Dineen D Gibson TJ Karplus K et al (2011) Fastscalable generation of high-quality protein multiple sequencealignments using Clustal Omega Mol Syst Biol 7 539 PubMed21988835
19 Childs RE Bardsley WG (1975) The steady-state kinetics of peroxidasewith 22-azino-di-(3-ethyl-benzthiazoline-6-sulphonic acid) aschromogen Biochem J 145 93-103 PubMed 1191252
20 Harkin JM Obst JR (1973) Syringaldazine an Effective Reagent forDetecting Laccase and Peroxidase in Fungi Experientia 29 382-387
21 Nikupaavola ML Raaska L Itavaara M (1990) Detection of white-rotfungi by a non-roxic stain Mycol Res 94 27-31 doi101016S0953-7562(09)81260-4
22 Gonzalez LF Sarria V Sanchez OF (2010) Degradation ofchlorophenols by sequential biological-advanced oxidative processusing Trametes pubescens and TiO(2)UV Bioresour Technol 1013493-3499
23 Waterborg JH (2009) The Lowry method for protein quantitation In JWalker The Protein Protocols Handbook Humana Press pp 7-10
24 Harkin JM Obst JR (1973) Syringaldazine an effective reagent fordetecting laccase and peroxidase in fungi Cell Mol Life Sci 29381-387 doi101007BF01926734
25 Fernaacutendez-Saacutenchez C Tzanov T Guumlbitz GM Cavaco-Paulo A (2002)Voltammetric monitoring of laccase-catalysed mediated reactionsBioelectrochemistry 58 149-156 doi101016S1567-5394(02)00119-6PubMed 12414320
26 Bourbonnais R Leech D Paice MG (1998) Electrochemical analysis ofthe interactions of laccase mediators with lignin model compoundsBiochim Biophys Acta 1379 381ndash390 doi101016S0304-4165(97)00117-7 PubMed 9545600
27 Ferraz F Silva S (2009) Characterization of coffee husk biomass forbiotechnological purposes N Biotechnol 25 S256-S256 doi101016jnbt200906573
28 Pandey A Soccol CR Nigam P Brand D Mohan R et al (2000)Biotechnological potential of coffee pulp and coffee husk forbioprocesses Biochem Eng J 6 153-162 doi101016S1369-703X(00)00084-X PubMed 10959086
29 Sruamsiri S Silman P (2008) Nutritive composition of soybean by-products and nutrient digestibility of soybean pod husk Maejo Int J SciTechnol 2 568-576
30 Romkaew J Nagaya Y Goto M Suzuki J Umezaki T (2008) Poddehiscence in relation to chemical components of pod shell in soybeanPlant Prod Sci 11 278-282 doi101626pps11278
31 Nakamanee G Polbumroong T Phaikaew C (2003) Alternative use ofcrop waste to increase milk production Proceedings of the 8th Meetingof the Regional Working Group on Grazing and Feed Resources forSoutheast Asia Kuala Lumpur Malaysia 66 - 70
32 Kurek B Gaudard F (2000) Oxidation of spruce wood sawdust byMnO(2) plus oxalate a biochemical investigation J Agric Food Chem48 3058-3062 doi101021jf000015g PubMed 10898665
33 Weil J Brewer M Hendrickson R Sarikaya A Ladisc MR (1998)Continuous pH monitoring during pretreatment of yellow poplar woodsawdust by pressure cooking in water Appl Biochem Biotechnol 70-7299-111 doi101007BF02920127
34 Weil J Sarikaya A Rau S-L Goetz J Ladisch CM et al (1997)Pretreatment of yellow poplar sawdust by pressure cooking in waterAppl Biochem Biotechnol 68 21-40 doi101007BF02785978
35 Dekker RF Barbosa AM Giese EC Godoy SD Covizzi LG (2007)Influence of nutrients on enhancing laccase production byBotryosphaeria rhodina MAMB-05 Int Microbiol 10 177-185 PubMed18075999
36 Xiao YZ Hong YZ Li JF Hang J Tong PG et al (2006) Cloning ofnovel laccase isozyme genes from Trametes sp AH28-2 and analysesof their differential expression Appl Microbiol Biotechnol 71 493-501doi101007s00253-005-0188-2 PubMed 16283298
37 Crowe JD Olsson S (2001) Induction of laccase activity in Rhizoctoniasolani by antagonistic Pseudomonas fluorescens strains and a range ofchemical treatments Appl Environ Microbiol 67 2088-2094 doi101128AEM6752088-20942001 PubMed 11319086
38 Yaguumle S Terroacuten MC Gonzaacutelez T Zapico E Bocchini P et al (2000)Biotreatment of tannin-rich beer-factory wastewater with white-rotbasidiomycete Coriolopsis gallica monitored by pyrolysisgaschromatographymass spectrometry Rapid Commun Mass Spectrom14 905-910 doi101002(SICI)1097-0231(20000530)1410 PubMed10825255
39 Gowda NKS Ramana JV Prasad CS Singh K (2004) Micronutrientcontent of certain tropical conventional and unconventional feedresources of southern India Trop Anim Heatlth Pro 36 77-94 doi101023BTROP0000009522309491d PubMed 14979561
40 DePeters EJ Fadel JG Arana MJ Ohanesian N Etchebarne MA et al(2000) Variability in the chemical composition of seventeen selectedby-product feedstuffs used by the California dairy industry Prof AnimSci 16 69-99
41 Obernberger I Thek G (2004) Physical characterisation and chemicalcomposition of densified biomass fuels with regard to their combustionbehaviour Biomass Bioenerg 27 653-669 doi101016jbiombioe200307006
42 Larous S Meniai A-H Lehocine MB (2005) Experimental study of theremoval of copper from aqueous solutions by adsorption usingsawdust Desalination 185 483ndash490 doi101016jdesal200503090
43 Oliveira WE Franca AS Oliveira LS Rocha SD (2008) Untreatedcoffee husks as biosorbents for the removal of heavy metals fromaqueous solutions J Hazard Mater 152 1073ndash1081 doi101016jjhazmat200707085 PubMed 17804159
44 Šciban M Klašnja M Škrbic B (2008) Adsorption of copper ions fromwater by modified agricultural by-products Desalination 229 170ndash180doi101016jdesal200708017
45 Maumlkelauml MR Hildeacuten KS Hakala TK Hatakka A Lundell TK (2006)Expression and molecular properties of a new laccase of the white rotfungus Phlebia radiata grown on wood Curr Genet 50 323-333 doi101007s00294-006-0090-1 PubMed 16927090
46 Maumlkelauml MR Lundell T Hatakka A Hildeacuten K (2013) Effect of coppernutrient nitrogen and wood-supplement on the production of lignin-modifying enzymes by the white-rot fungus Phlebia radiata Fungal Biol117 62-70 doi101016jfunbio201211006 PubMed 23332834
47 Dwivedi UN Singha P Pandeya VP Kumara A (2011) Structurendashfunction relationship among bacterial fungal and plant laccases J MolCatal B Enzym 68 117-128 doi101016jmolcatb201011002
48 Gaitan IJ Medina SC Gonzaacutelez JC Rodriacuteguez A Espejo AacuteJ et al(2011) Evaluation of toxicity and degradation of a chlorophenol mixtureby the laccase produced by Trametes pubescens Bioresour Technol102 3632ndash3635 doi101016jbiortech201011040 PubMed21115244
49 Karp SG Faraco V Amore A Birolo L Giangrande C et al (2012)Characterization of laccase isoforms produced by Pleurotus ostreatusin solid state fermentation of sugarcane bagasse Bioresour Technol114 735-739 doi101016jbiortech201203058 PubMed 22487128
50 Nikitina OV Shleev SV Gorshina ES Rusinova TV Serezhenkov VAet al (2005) Isolation and purification of enzymes from ligninolyticcomplex of the basidial fungus Trametes pubescens (Schumach) Pilatand study of their properties Biochemistry (Mosc) 70 1274-1279 doi101007s10541-005-0259-0
51 Shleev S Nikitina O Christenson A Reimann CT Yaropolov AI et al(2007) Characterization of two new multiforms of Trametes pubescenslaccase Bioorg Chem 35 35-49 doi101016jbioorg200608001PubMed 16989887
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 13 September 2013 | Volume 8 | Issue 9 | e73721
52 Xiao YZ Chen Q Hang J Shi YY Wu J et al (2004) Selectiveinduction purification and characterization of a laccase isozyme fromthe basidiomycete Trametes sp AH28-2 Mycologia 96 26-35 doi1023073761984 PubMed 21148825
53 Muntildeoz C Guilleacuten F Martiacutenez AT Martiacutenez MJ (1997) Laccaseisoenzymes of Pleurotus eryngii characterization catalytic propertiesand participation in activation of molecular oxygen and Mn2+ oxidationAppl Environ Microbiol 63 2166-2174 PubMed 9172335
54 Morozova OV Shumakovich GP Gorbacheva MA Shleev SVYaropolov AI (2007) Blue laccases Biochemistry (Mosc) 721136-1150 doi101134S0006297907100112
55 Hildeacuten K Hakala TK Lundell T (2009) Thermotolerant andthermostable laccases Biotechnol Lett 31 1117-1128 doi101007s10529-009-9998-0 PubMed 19360388
56 Verma P Madamwar D (2002) Production of ligninolytic enzymes fordye decolorization by cocultivation of white-rot fungi Pleurotusostreatus and Phanerochaete chrysosporium under solid-statefermentation Appl Biochem Biotechnol 102-103 109-118 doi101385ABAB102-1031-6109 PubMed 12396115
57 Meza JC Lomascolo A Casalot L Sigoillot J-C Auria R (2005)Laccase production by Pycnoporus cinnabarinus grown on sugar-canebagasse Influence of ethanol vapours as inducer Proc Biochem 403365ndash3371 doi101016jprocbio200503004
58 Hossain SM Anantharaman N (2006) Activity enhancement ofligninolytic enzymes of Trametes versicolor with bagasse powder Afr JBiotechnol 5 189-194
59 Revankar MS Desai KM Lele SS (2007) Solid-state fermentation forenhanced production of laccase using indigenously isolatedGanoderma sp Appl Biochem Biotechnol 143 16-26 doi101007s12010-007-0029-0 PubMed 18025593
60 Velazquez-Cedeno MA Mata G Savoie J-M (2002) Waste-reducingcultivation of Pleurotus ostreatus and Pleurotus pulmonarius on coffeepulp changes in the production of some lignocellulolytic enzymesWorld J Microbiol Biotechnol 18 201ndash207 doi101023A1014999616381
61 Mata G Murrieta D Andreu LI (2005) Changes in lignocellulolyticenzyme activities in six Pleurotus spp strains cultivated on coffee pulpin confrontation with Trichoderma spp World J Microbiol Biotechnol 21143ndash150 doi101007s11274-004-3041-3
62 de Souza DF Tychanowicz GK de Souza CG Peralta RM (2006) Co-production of ligninolytic enzymes by Pleurotus pulmonarius on wheatbran solid state cultures J Basic Microbiol 46 126-134 doi101002jobm200510014 PubMed 16598826
63 Prasad KK Mohan SV Bhaskar YV Ramanaiah SV Babu VL et al(2005) Laccase production using Pleurotus ostreatus 1804 immobilizedon PUF cubes in batch and packed bed reactors influence of cultureconditions J Microbiol 43 301-307 PubMed 15995650
64 Songulashvili G Elisashvili V Wasser SP Nevo E Hadar Y (2007)Basidiomycetes laccase and manganese peroxidase activity in
submerged fermentation of food industry wastes Enzyme MicrobTechnol 41 57ndash61 doi101016jenzmictec200611024
65 Sathishkumar P Murugesan K Palvannan T (2010) Production oflaccase from Pleurotus florida using agro-wastes and efficientdecolorization of Reactive blue 198 J Basic Microbiol 50 360-367 doi101002jobm200900407 PubMed 20586068
66 Moldes D Gallego PP Rodriacuteguez Couto S Sanromaacuten A (2003) Grapeseeds the best lignocellulosic waste to produce laccase by solid statecultures of Trametes hirsuta Biotechnol Lett 25 491-495 doi101023A1022660230653 PubMed 12882277
67 Elisashvili V Kachlishvili E Penninckx M (2008) Effect of growthsubstrate method of fermentation and nitrogen source onlignocellulose-degrading enzymes production by white-rotbasidiomycetes J Ind Microbiol Biotechnol 35 1531-1538 doi101007s10295-008-0454-2 PubMed 18716810
68 Osma JF Saravia V Herrera JL Couto SR (2007) Mandarin peelingsthe best carbon source to produce laccase by static cultures ofTrametes pubescens Chemosphere 67 1677-1680 doi101016jchemosphere200611051 PubMed 17234250
69 Kahraman SS Gurdal IH (2002) Effect of synthetic and natural culturemedia on laccase production by white rot fungi Bioresour Technol 82215-217 doi101016S0960-8524(01)00193-6 PubMed 11991068
70 Iandolo D Piscitelli A Sannia G Faraco V (2011) Enzyme Productionby Solid Substrate Fermentation of Pleurotus ostreatus and Trametesversicolor on Tomato Pomace Appl Biochem Biotechnol 163 40-51doi101007s12010-010-9014-0 PubMed 20582639
71 Moldes D Lorenzo M Sanromaacuten MA (2004) Different proportions oflaccase isoenzymes produced by submerged cultures of Trametesversicolor grown on lignocellulosic wastes Biotechnol Lett 26 327-330doi101023BBILE000001545240213bf PubMed 15055770
72 Philippoussis A Diamantopoulou P Papadopoulou K Lakhtar HRoussos S et al (2010) Biomass laccase and endoglucanaseproduction by Lentinula edodes during solid state fermentation of reedgrass bean stalks and wheat straw residues World J MicrobiolBiotechnol 27 285-297
73 Fenice M Giovannozzi Sermanni G Federici F DrsquoAnnibale A (2003)Submerged and solid-state production of laccase and Mn-peroxidaseby Panus tigrinus on olive mill wastewater-based media J Biotechnol100 77-85 doi101016S0168-1656(02)00241-9 PubMed 12413788
74 Rodriguez-Couto S GundIn M Lorenzo M Sanromaacuten MAacute (2002)Screening of supports and inducers for laccase production by Trametesversicolor in semi-solid-state conditions Proc Biochem 38 249-255doi101016S0032-9592(02)00087-0
75 Fackler K Gradinger C Schmutzer M Tavzes C Burgert I et al (2007)Biotechnological wood modification with selective white-rot fungi and itsmolecular mechanisms Food Technol Biotechnol 45 269ndash276
76 Levin L Herrmann C Papinutti VL (2008) Optimization oflignocellulolytic enzyme production by the white-rot fungus Trametestrogii in solid-state fermentation using response surface methodologyBiochem Eng J 39 207-214 doi101016jbej200709004
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 14 September 2013 | Volume 8 | Issue 9 | e73721

and Lac2 as have been reported for laccases obtained fromTrametes and Pleurotus strains [175253]
The maximum relative activity for the UF retentate waspresented at a pH from 20 to 40 regardless of the agro-industrial byproduct with a monotonical decrease at pHs abovethis range The relation between pH and type of electron donormolecule explains this behavior on laccase activity It has beenreported that when the substrate is an organic hydrogen donorie phenols the optimal pH ranges from 35 to 60 while thelaccase activity monotonically decreases from pH 25 to 70 ifthe substrate is an electron donor like ABTS [54] Despite thatat different pHs most of the purified laccase fractionspresented a similar performance as the UF retentates the Lac2fraction obtained from cedar sawdust differed from the previousobservation For this fraction the maximum activity wasobserved at pH 50 and despite that at acidic pH (ge 30) therelative activity was higher than 80 at lower pHs (20) itdecreased almost in a 50 Nevertheless at basic pHs therelative laccase activity presented a sharp decreased that iswell correlate with the other laccase fractions We are not ableto attribute this particular trend to the presence of multiplelaccase isoenzymes in the fraction since the SDS-PAGErevealed only one band when stained with ABTS and from theNanoLC-MSMS analysis no difference in the number ofpeptides and their sequence was detected We speculate thatthis particular behavior can be owed to a compound added inthe pre-treatment of the cedar wood that might be used forpreserving it that might affect the activity of Lac2 and that wasnot completely removed during the purification
Temperature stability evaluation showed that Lac1 is themost thermolabile fraction while the laccase activity reductionin the crude extracts did not seem to be the result of anadditive effect of the activity loss of the purified fractionsAlthough there are no reports of laccase stability under theselected conditions these results agree with previous reportsthat state that fungal laccases rapidly decrease their activity attemperatures near 60 degC and thermal stability correlates withthe temperature range of the growth of the source organism[55] In addition we previously reported for a laccase crudeextract obtained from culturing T pubescens on coffee husk anactivity loss of 26 and 10 at 60 and 50 degC respectivelyafter 8 h at pH 60 using syringaldazine as substrate [48]
Finally an electrochemical characterization was carried outfor selected crude extracts and purified fractions The results
showed a difference between the electrochemicalperformances of extracts and purified fractions withsyringaldazine or ABTS These differences could be owed tothe different mechanisms for electron transfer as discussedabove However the overall performance of the obtained cyclicvoltammograms for ABTS and syringaldazine agrees withresults reported by Fernaacutendez-Saacutenchez et al [25] who used asaturated calomel reference electrode
Conclusions
We have shown the potential of coffee husk soybean podhusk and cedar sawdust as substrates for the laccaseproduction In several countries like Brazil and Colombia theproduction of coffee and soybean is of great importance andthe generated volume of residues is high The results showedthat semi-solid culture of T pubescens using coffee huskallows to obtain an important amount of laccase which couldbe favored by the phenolic-compounds ie tannins presentedin coffee husk While supplementation of the culture mediumwith copper favored the production of laccase in submergedand semi-solid culture using soybean pod husk and cedarsawdust semi-solid culture using coffee husk which presentedthe highest laccase volumetric yield did not require coppersupplementation This is an economic advantage that shouldbe considered when scaling up the process In this study Tpubescens showed its ability to produce two independentlaccases regardless of the used substrate However a higherlaccase activity was observed for crude extracts rather than forpurified fractions This suggests that Lac1 and Lac2 might havea synergistic effect on the final laccase activity rather than anadditive effect Based on this study the investigation of laccaseproduction by evaluating other agro-industrial wastes in whichphenolic-compounds constitute an important percentage oftheir composition is of great interest to avoid the use ofinducers that would increase the production cost
Author Contributions
Conceived and designed the experiments JFO CJAD OFSPerformed the experiments JCG SCM AR Analyzed the dataJCG SCM AR JFO CJAD OFS Wrote the manuscript AR JFOCJAD OFS
References
1 Vandamme EJ (2009) Agro-industrial residue utilization for industrialbiotechnology products In P NigamA Pandey Biotechnology for Agro-Industrial Residues Utilisation Netherlands Springer Verlag pp 3-11
2 Martiacutenez AT Speranza M Ruiz-Duentildeas FJ Ferreira P Camarero S etal (2005) Biodegradation of lignocellulosics microbial chemical andenzymatic aspects of the fungal attack of lignin Int Microbiol 8195-204 PubMed 16200498
3 Saacutenchez O Sierra R Almeacuteciga-Diacuteaz CJ (2011) Delignification processof agro-industrial aastes an alternative to obtain fermentablecarbohydrates for producing fuel In M Manzanera Alternative FuelIntech 111-154
4 Wesenberg D Kyriakides I Agathos SN (2003) White-rot fungi andtheir enzymes for the treatment of industrial dye effluents BiotechnolAdv 22 161-187 doi101016jbiotechadv200308011 PubMed14623049
5 Morozova OV Shumakovich GP Shleev SV Yaropolov YI (2007)Laccase-mediator systems and their applications A review ApplBiochem Microbiol 43 523-535 doi101134S0003683807050055
6 Baldrian P (2006) Fungal laccases - occurrence and properties FEMSMicrobiol Rev 30 215-242 doi101111j1574-4976200500010xPubMed 16472305
7 Rodriacuteguez Couto S Herrera JLT (2006) Industrial and biotechnologicalapplications of laccases A review Biotechnol Adv 24 500-513 doi101016jbiotechadv200604003 PubMed 16716556
8 Osma JF Toca-Herrera JL Rodriacuteguez-Couto S (2010) Uses oflaccases in the food industry Enzyme Res 2010 2010 918761PubMed 21048873
9 Rodriguez S (2008) Exploitation of biological wastes for the productionof value-added products under solid-state fermentation conditionsBiotechnol J 3 859-870 doi101002biot200800031 PubMed18543242
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 12 September 2013 | Volume 8 | Issue 9 | e73721
10 Osma JF Toca-Herrera JL Rodriacuteguez-Couto S (2011) Cost analysis inlaccase production J Environ Manage 92 2907-2912 doi101016jjenvman201106052 PubMed 21775046
11 Jain A Morlok CK Henson JM (2013) Comparison of solid-state andsubmerged-state fermentation for the bioprocessing of switchgrass toethanol and acetate by Clostridium phytofermentans Appl MicrobiolBiotechnol 97 905-917 doi101007s00253-012-4511-4 PubMed23111595
12 Singhania RR Patel AK Soccol CR Pandey A (2009) Recentadvances in solid-state fermentation Biochem Eng J 44 13-18 doi101016jbej200810019
13 Economou CN Makri A Aggelis G Pavlou S Vayenas DV (2010)Semi-solid state fermentation of sweet sorghum for the biotechnologicalproduction of single cell oil Bioresour Technol 101 1385-1388 doi101016jbiortech200909028 PubMed 19781936
14 Rodriacuteguez CS Santoro R Cameselle C Sanroman A (1997) Laccaseproduction in semi-solid cultures of Phanerochaete chrysosporiumBiotechnol Lett 19 995-998 doi101023A1018495216946
15 Osma JF Moilanen U Toca-Herrera JL Rodriacuteguez-Couto S (2011)Morphology and laccase production of white-rot fungi grown on wheatbran flakes under semi-solid-state fermentation conditions FEMSMicrobiol Lett 318 27-34 doi101111j1574-6968201102234xPubMed 21291496
16 Couto SR Gundin M Lorenzo M Sanroman MN (2002) Screening ofsupports and inducers for laccase production by Trametes versicolor insemi-solid-state conditions Proc Biochem 38 249-255 doi101016S0032-9592(02)00087-0
17 Galhaup C Goller S Peterbauer CK Strauss J Haltrich D (2002)Characterization of the major laccase isoenzyme from Trametespubescens and regulation of its synthesis by metal ions Microbiology148 2159-2169 PubMed 12101303
18 Sievers F Wilm A Dineen D Gibson TJ Karplus K et al (2011) Fastscalable generation of high-quality protein multiple sequencealignments using Clustal Omega Mol Syst Biol 7 539 PubMed21988835
19 Childs RE Bardsley WG (1975) The steady-state kinetics of peroxidasewith 22-azino-di-(3-ethyl-benzthiazoline-6-sulphonic acid) aschromogen Biochem J 145 93-103 PubMed 1191252
20 Harkin JM Obst JR (1973) Syringaldazine an Effective Reagent forDetecting Laccase and Peroxidase in Fungi Experientia 29 382-387
21 Nikupaavola ML Raaska L Itavaara M (1990) Detection of white-rotfungi by a non-roxic stain Mycol Res 94 27-31 doi101016S0953-7562(09)81260-4
22 Gonzalez LF Sarria V Sanchez OF (2010) Degradation ofchlorophenols by sequential biological-advanced oxidative processusing Trametes pubescens and TiO(2)UV Bioresour Technol 1013493-3499
23 Waterborg JH (2009) The Lowry method for protein quantitation In JWalker The Protein Protocols Handbook Humana Press pp 7-10
24 Harkin JM Obst JR (1973) Syringaldazine an effective reagent fordetecting laccase and peroxidase in fungi Cell Mol Life Sci 29381-387 doi101007BF01926734
25 Fernaacutendez-Saacutenchez C Tzanov T Guumlbitz GM Cavaco-Paulo A (2002)Voltammetric monitoring of laccase-catalysed mediated reactionsBioelectrochemistry 58 149-156 doi101016S1567-5394(02)00119-6PubMed 12414320
26 Bourbonnais R Leech D Paice MG (1998) Electrochemical analysis ofthe interactions of laccase mediators with lignin model compoundsBiochim Biophys Acta 1379 381ndash390 doi101016S0304-4165(97)00117-7 PubMed 9545600
27 Ferraz F Silva S (2009) Characterization of coffee husk biomass forbiotechnological purposes N Biotechnol 25 S256-S256 doi101016jnbt200906573
28 Pandey A Soccol CR Nigam P Brand D Mohan R et al (2000)Biotechnological potential of coffee pulp and coffee husk forbioprocesses Biochem Eng J 6 153-162 doi101016S1369-703X(00)00084-X PubMed 10959086
29 Sruamsiri S Silman P (2008) Nutritive composition of soybean by-products and nutrient digestibility of soybean pod husk Maejo Int J SciTechnol 2 568-576
30 Romkaew J Nagaya Y Goto M Suzuki J Umezaki T (2008) Poddehiscence in relation to chemical components of pod shell in soybeanPlant Prod Sci 11 278-282 doi101626pps11278
31 Nakamanee G Polbumroong T Phaikaew C (2003) Alternative use ofcrop waste to increase milk production Proceedings of the 8th Meetingof the Regional Working Group on Grazing and Feed Resources forSoutheast Asia Kuala Lumpur Malaysia 66 - 70
32 Kurek B Gaudard F (2000) Oxidation of spruce wood sawdust byMnO(2) plus oxalate a biochemical investigation J Agric Food Chem48 3058-3062 doi101021jf000015g PubMed 10898665
33 Weil J Brewer M Hendrickson R Sarikaya A Ladisc MR (1998)Continuous pH monitoring during pretreatment of yellow poplar woodsawdust by pressure cooking in water Appl Biochem Biotechnol 70-7299-111 doi101007BF02920127
34 Weil J Sarikaya A Rau S-L Goetz J Ladisch CM et al (1997)Pretreatment of yellow poplar sawdust by pressure cooking in waterAppl Biochem Biotechnol 68 21-40 doi101007BF02785978
35 Dekker RF Barbosa AM Giese EC Godoy SD Covizzi LG (2007)Influence of nutrients on enhancing laccase production byBotryosphaeria rhodina MAMB-05 Int Microbiol 10 177-185 PubMed18075999
36 Xiao YZ Hong YZ Li JF Hang J Tong PG et al (2006) Cloning ofnovel laccase isozyme genes from Trametes sp AH28-2 and analysesof their differential expression Appl Microbiol Biotechnol 71 493-501doi101007s00253-005-0188-2 PubMed 16283298
37 Crowe JD Olsson S (2001) Induction of laccase activity in Rhizoctoniasolani by antagonistic Pseudomonas fluorescens strains and a range ofchemical treatments Appl Environ Microbiol 67 2088-2094 doi101128AEM6752088-20942001 PubMed 11319086
38 Yaguumle S Terroacuten MC Gonzaacutelez T Zapico E Bocchini P et al (2000)Biotreatment of tannin-rich beer-factory wastewater with white-rotbasidiomycete Coriolopsis gallica monitored by pyrolysisgaschromatographymass spectrometry Rapid Commun Mass Spectrom14 905-910 doi101002(SICI)1097-0231(20000530)1410 PubMed10825255
39 Gowda NKS Ramana JV Prasad CS Singh K (2004) Micronutrientcontent of certain tropical conventional and unconventional feedresources of southern India Trop Anim Heatlth Pro 36 77-94 doi101023BTROP0000009522309491d PubMed 14979561
40 DePeters EJ Fadel JG Arana MJ Ohanesian N Etchebarne MA et al(2000) Variability in the chemical composition of seventeen selectedby-product feedstuffs used by the California dairy industry Prof AnimSci 16 69-99
41 Obernberger I Thek G (2004) Physical characterisation and chemicalcomposition of densified biomass fuels with regard to their combustionbehaviour Biomass Bioenerg 27 653-669 doi101016jbiombioe200307006
42 Larous S Meniai A-H Lehocine MB (2005) Experimental study of theremoval of copper from aqueous solutions by adsorption usingsawdust Desalination 185 483ndash490 doi101016jdesal200503090
43 Oliveira WE Franca AS Oliveira LS Rocha SD (2008) Untreatedcoffee husks as biosorbents for the removal of heavy metals fromaqueous solutions J Hazard Mater 152 1073ndash1081 doi101016jjhazmat200707085 PubMed 17804159
44 Šciban M Klašnja M Škrbic B (2008) Adsorption of copper ions fromwater by modified agricultural by-products Desalination 229 170ndash180doi101016jdesal200708017
45 Maumlkelauml MR Hildeacuten KS Hakala TK Hatakka A Lundell TK (2006)Expression and molecular properties of a new laccase of the white rotfungus Phlebia radiata grown on wood Curr Genet 50 323-333 doi101007s00294-006-0090-1 PubMed 16927090
46 Maumlkelauml MR Lundell T Hatakka A Hildeacuten K (2013) Effect of coppernutrient nitrogen and wood-supplement on the production of lignin-modifying enzymes by the white-rot fungus Phlebia radiata Fungal Biol117 62-70 doi101016jfunbio201211006 PubMed 23332834
47 Dwivedi UN Singha P Pandeya VP Kumara A (2011) Structurendashfunction relationship among bacterial fungal and plant laccases J MolCatal B Enzym 68 117-128 doi101016jmolcatb201011002
48 Gaitan IJ Medina SC Gonzaacutelez JC Rodriacuteguez A Espejo AacuteJ et al(2011) Evaluation of toxicity and degradation of a chlorophenol mixtureby the laccase produced by Trametes pubescens Bioresour Technol102 3632ndash3635 doi101016jbiortech201011040 PubMed21115244
49 Karp SG Faraco V Amore A Birolo L Giangrande C et al (2012)Characterization of laccase isoforms produced by Pleurotus ostreatusin solid state fermentation of sugarcane bagasse Bioresour Technol114 735-739 doi101016jbiortech201203058 PubMed 22487128
50 Nikitina OV Shleev SV Gorshina ES Rusinova TV Serezhenkov VAet al (2005) Isolation and purification of enzymes from ligninolyticcomplex of the basidial fungus Trametes pubescens (Schumach) Pilatand study of their properties Biochemistry (Mosc) 70 1274-1279 doi101007s10541-005-0259-0
51 Shleev S Nikitina O Christenson A Reimann CT Yaropolov AI et al(2007) Characterization of two new multiforms of Trametes pubescenslaccase Bioorg Chem 35 35-49 doi101016jbioorg200608001PubMed 16989887
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 13 September 2013 | Volume 8 | Issue 9 | e73721
52 Xiao YZ Chen Q Hang J Shi YY Wu J et al (2004) Selectiveinduction purification and characterization of a laccase isozyme fromthe basidiomycete Trametes sp AH28-2 Mycologia 96 26-35 doi1023073761984 PubMed 21148825
53 Muntildeoz C Guilleacuten F Martiacutenez AT Martiacutenez MJ (1997) Laccaseisoenzymes of Pleurotus eryngii characterization catalytic propertiesand participation in activation of molecular oxygen and Mn2+ oxidationAppl Environ Microbiol 63 2166-2174 PubMed 9172335
54 Morozova OV Shumakovich GP Gorbacheva MA Shleev SVYaropolov AI (2007) Blue laccases Biochemistry (Mosc) 721136-1150 doi101134S0006297907100112
55 Hildeacuten K Hakala TK Lundell T (2009) Thermotolerant andthermostable laccases Biotechnol Lett 31 1117-1128 doi101007s10529-009-9998-0 PubMed 19360388
56 Verma P Madamwar D (2002) Production of ligninolytic enzymes fordye decolorization by cocultivation of white-rot fungi Pleurotusostreatus and Phanerochaete chrysosporium under solid-statefermentation Appl Biochem Biotechnol 102-103 109-118 doi101385ABAB102-1031-6109 PubMed 12396115
57 Meza JC Lomascolo A Casalot L Sigoillot J-C Auria R (2005)Laccase production by Pycnoporus cinnabarinus grown on sugar-canebagasse Influence of ethanol vapours as inducer Proc Biochem 403365ndash3371 doi101016jprocbio200503004
58 Hossain SM Anantharaman N (2006) Activity enhancement ofligninolytic enzymes of Trametes versicolor with bagasse powder Afr JBiotechnol 5 189-194
59 Revankar MS Desai KM Lele SS (2007) Solid-state fermentation forenhanced production of laccase using indigenously isolatedGanoderma sp Appl Biochem Biotechnol 143 16-26 doi101007s12010-007-0029-0 PubMed 18025593
60 Velazquez-Cedeno MA Mata G Savoie J-M (2002) Waste-reducingcultivation of Pleurotus ostreatus and Pleurotus pulmonarius on coffeepulp changes in the production of some lignocellulolytic enzymesWorld J Microbiol Biotechnol 18 201ndash207 doi101023A1014999616381
61 Mata G Murrieta D Andreu LI (2005) Changes in lignocellulolyticenzyme activities in six Pleurotus spp strains cultivated on coffee pulpin confrontation with Trichoderma spp World J Microbiol Biotechnol 21143ndash150 doi101007s11274-004-3041-3
62 de Souza DF Tychanowicz GK de Souza CG Peralta RM (2006) Co-production of ligninolytic enzymes by Pleurotus pulmonarius on wheatbran solid state cultures J Basic Microbiol 46 126-134 doi101002jobm200510014 PubMed 16598826
63 Prasad KK Mohan SV Bhaskar YV Ramanaiah SV Babu VL et al(2005) Laccase production using Pleurotus ostreatus 1804 immobilizedon PUF cubes in batch and packed bed reactors influence of cultureconditions J Microbiol 43 301-307 PubMed 15995650
64 Songulashvili G Elisashvili V Wasser SP Nevo E Hadar Y (2007)Basidiomycetes laccase and manganese peroxidase activity in
submerged fermentation of food industry wastes Enzyme MicrobTechnol 41 57ndash61 doi101016jenzmictec200611024
65 Sathishkumar P Murugesan K Palvannan T (2010) Production oflaccase from Pleurotus florida using agro-wastes and efficientdecolorization of Reactive blue 198 J Basic Microbiol 50 360-367 doi101002jobm200900407 PubMed 20586068
66 Moldes D Gallego PP Rodriacuteguez Couto S Sanromaacuten A (2003) Grapeseeds the best lignocellulosic waste to produce laccase by solid statecultures of Trametes hirsuta Biotechnol Lett 25 491-495 doi101023A1022660230653 PubMed 12882277
67 Elisashvili V Kachlishvili E Penninckx M (2008) Effect of growthsubstrate method of fermentation and nitrogen source onlignocellulose-degrading enzymes production by white-rotbasidiomycetes J Ind Microbiol Biotechnol 35 1531-1538 doi101007s10295-008-0454-2 PubMed 18716810
68 Osma JF Saravia V Herrera JL Couto SR (2007) Mandarin peelingsthe best carbon source to produce laccase by static cultures ofTrametes pubescens Chemosphere 67 1677-1680 doi101016jchemosphere200611051 PubMed 17234250
69 Kahraman SS Gurdal IH (2002) Effect of synthetic and natural culturemedia on laccase production by white rot fungi Bioresour Technol 82215-217 doi101016S0960-8524(01)00193-6 PubMed 11991068
70 Iandolo D Piscitelli A Sannia G Faraco V (2011) Enzyme Productionby Solid Substrate Fermentation of Pleurotus ostreatus and Trametesversicolor on Tomato Pomace Appl Biochem Biotechnol 163 40-51doi101007s12010-010-9014-0 PubMed 20582639
71 Moldes D Lorenzo M Sanromaacuten MA (2004) Different proportions oflaccase isoenzymes produced by submerged cultures of Trametesversicolor grown on lignocellulosic wastes Biotechnol Lett 26 327-330doi101023BBILE000001545240213bf PubMed 15055770
72 Philippoussis A Diamantopoulou P Papadopoulou K Lakhtar HRoussos S et al (2010) Biomass laccase and endoglucanaseproduction by Lentinula edodes during solid state fermentation of reedgrass bean stalks and wheat straw residues World J MicrobiolBiotechnol 27 285-297
73 Fenice M Giovannozzi Sermanni G Federici F DrsquoAnnibale A (2003)Submerged and solid-state production of laccase and Mn-peroxidaseby Panus tigrinus on olive mill wastewater-based media J Biotechnol100 77-85 doi101016S0168-1656(02)00241-9 PubMed 12413788
74 Rodriguez-Couto S GundIn M Lorenzo M Sanromaacuten MAacute (2002)Screening of supports and inducers for laccase production by Trametesversicolor in semi-solid-state conditions Proc Biochem 38 249-255doi101016S0032-9592(02)00087-0
75 Fackler K Gradinger C Schmutzer M Tavzes C Burgert I et al (2007)Biotechnological wood modification with selective white-rot fungi and itsmolecular mechanisms Food Technol Biotechnol 45 269ndash276
76 Levin L Herrmann C Papinutti VL (2008) Optimization oflignocellulolytic enzyme production by the white-rot fungus Trametestrogii in solid-state fermentation using response surface methodologyBiochem Eng J 39 207-214 doi101016jbej200709004
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 14 September 2013 | Volume 8 | Issue 9 | e73721

10 Osma JF Toca-Herrera JL Rodriacuteguez-Couto S (2011) Cost analysis inlaccase production J Environ Manage 92 2907-2912 doi101016jjenvman201106052 PubMed 21775046
11 Jain A Morlok CK Henson JM (2013) Comparison of solid-state andsubmerged-state fermentation for the bioprocessing of switchgrass toethanol and acetate by Clostridium phytofermentans Appl MicrobiolBiotechnol 97 905-917 doi101007s00253-012-4511-4 PubMed23111595
12 Singhania RR Patel AK Soccol CR Pandey A (2009) Recentadvances in solid-state fermentation Biochem Eng J 44 13-18 doi101016jbej200810019
13 Economou CN Makri A Aggelis G Pavlou S Vayenas DV (2010)Semi-solid state fermentation of sweet sorghum for the biotechnologicalproduction of single cell oil Bioresour Technol 101 1385-1388 doi101016jbiortech200909028 PubMed 19781936
14 Rodriacuteguez CS Santoro R Cameselle C Sanroman A (1997) Laccaseproduction in semi-solid cultures of Phanerochaete chrysosporiumBiotechnol Lett 19 995-998 doi101023A1018495216946
15 Osma JF Moilanen U Toca-Herrera JL Rodriacuteguez-Couto S (2011)Morphology and laccase production of white-rot fungi grown on wheatbran flakes under semi-solid-state fermentation conditions FEMSMicrobiol Lett 318 27-34 doi101111j1574-6968201102234xPubMed 21291496
16 Couto SR Gundin M Lorenzo M Sanroman MN (2002) Screening ofsupports and inducers for laccase production by Trametes versicolor insemi-solid-state conditions Proc Biochem 38 249-255 doi101016S0032-9592(02)00087-0
17 Galhaup C Goller S Peterbauer CK Strauss J Haltrich D (2002)Characterization of the major laccase isoenzyme from Trametespubescens and regulation of its synthesis by metal ions Microbiology148 2159-2169 PubMed 12101303
18 Sievers F Wilm A Dineen D Gibson TJ Karplus K et al (2011) Fastscalable generation of high-quality protein multiple sequencealignments using Clustal Omega Mol Syst Biol 7 539 PubMed21988835
19 Childs RE Bardsley WG (1975) The steady-state kinetics of peroxidasewith 22-azino-di-(3-ethyl-benzthiazoline-6-sulphonic acid) aschromogen Biochem J 145 93-103 PubMed 1191252
20 Harkin JM Obst JR (1973) Syringaldazine an Effective Reagent forDetecting Laccase and Peroxidase in Fungi Experientia 29 382-387
21 Nikupaavola ML Raaska L Itavaara M (1990) Detection of white-rotfungi by a non-roxic stain Mycol Res 94 27-31 doi101016S0953-7562(09)81260-4
22 Gonzalez LF Sarria V Sanchez OF (2010) Degradation ofchlorophenols by sequential biological-advanced oxidative processusing Trametes pubescens and TiO(2)UV Bioresour Technol 1013493-3499
23 Waterborg JH (2009) The Lowry method for protein quantitation In JWalker The Protein Protocols Handbook Humana Press pp 7-10
24 Harkin JM Obst JR (1973) Syringaldazine an effective reagent fordetecting laccase and peroxidase in fungi Cell Mol Life Sci 29381-387 doi101007BF01926734
25 Fernaacutendez-Saacutenchez C Tzanov T Guumlbitz GM Cavaco-Paulo A (2002)Voltammetric monitoring of laccase-catalysed mediated reactionsBioelectrochemistry 58 149-156 doi101016S1567-5394(02)00119-6PubMed 12414320
26 Bourbonnais R Leech D Paice MG (1998) Electrochemical analysis ofthe interactions of laccase mediators with lignin model compoundsBiochim Biophys Acta 1379 381ndash390 doi101016S0304-4165(97)00117-7 PubMed 9545600
27 Ferraz F Silva S (2009) Characterization of coffee husk biomass forbiotechnological purposes N Biotechnol 25 S256-S256 doi101016jnbt200906573
28 Pandey A Soccol CR Nigam P Brand D Mohan R et al (2000)Biotechnological potential of coffee pulp and coffee husk forbioprocesses Biochem Eng J 6 153-162 doi101016S1369-703X(00)00084-X PubMed 10959086
29 Sruamsiri S Silman P (2008) Nutritive composition of soybean by-products and nutrient digestibility of soybean pod husk Maejo Int J SciTechnol 2 568-576
30 Romkaew J Nagaya Y Goto M Suzuki J Umezaki T (2008) Poddehiscence in relation to chemical components of pod shell in soybeanPlant Prod Sci 11 278-282 doi101626pps11278
31 Nakamanee G Polbumroong T Phaikaew C (2003) Alternative use ofcrop waste to increase milk production Proceedings of the 8th Meetingof the Regional Working Group on Grazing and Feed Resources forSoutheast Asia Kuala Lumpur Malaysia 66 - 70
32 Kurek B Gaudard F (2000) Oxidation of spruce wood sawdust byMnO(2) plus oxalate a biochemical investigation J Agric Food Chem48 3058-3062 doi101021jf000015g PubMed 10898665
33 Weil J Brewer M Hendrickson R Sarikaya A Ladisc MR (1998)Continuous pH monitoring during pretreatment of yellow poplar woodsawdust by pressure cooking in water Appl Biochem Biotechnol 70-7299-111 doi101007BF02920127
34 Weil J Sarikaya A Rau S-L Goetz J Ladisch CM et al (1997)Pretreatment of yellow poplar sawdust by pressure cooking in waterAppl Biochem Biotechnol 68 21-40 doi101007BF02785978
35 Dekker RF Barbosa AM Giese EC Godoy SD Covizzi LG (2007)Influence of nutrients on enhancing laccase production byBotryosphaeria rhodina MAMB-05 Int Microbiol 10 177-185 PubMed18075999
36 Xiao YZ Hong YZ Li JF Hang J Tong PG et al (2006) Cloning ofnovel laccase isozyme genes from Trametes sp AH28-2 and analysesof their differential expression Appl Microbiol Biotechnol 71 493-501doi101007s00253-005-0188-2 PubMed 16283298
37 Crowe JD Olsson S (2001) Induction of laccase activity in Rhizoctoniasolani by antagonistic Pseudomonas fluorescens strains and a range ofchemical treatments Appl Environ Microbiol 67 2088-2094 doi101128AEM6752088-20942001 PubMed 11319086
38 Yaguumle S Terroacuten MC Gonzaacutelez T Zapico E Bocchini P et al (2000)Biotreatment of tannin-rich beer-factory wastewater with white-rotbasidiomycete Coriolopsis gallica monitored by pyrolysisgaschromatographymass spectrometry Rapid Commun Mass Spectrom14 905-910 doi101002(SICI)1097-0231(20000530)1410 PubMed10825255
39 Gowda NKS Ramana JV Prasad CS Singh K (2004) Micronutrientcontent of certain tropical conventional and unconventional feedresources of southern India Trop Anim Heatlth Pro 36 77-94 doi101023BTROP0000009522309491d PubMed 14979561
40 DePeters EJ Fadel JG Arana MJ Ohanesian N Etchebarne MA et al(2000) Variability in the chemical composition of seventeen selectedby-product feedstuffs used by the California dairy industry Prof AnimSci 16 69-99
41 Obernberger I Thek G (2004) Physical characterisation and chemicalcomposition of densified biomass fuels with regard to their combustionbehaviour Biomass Bioenerg 27 653-669 doi101016jbiombioe200307006
42 Larous S Meniai A-H Lehocine MB (2005) Experimental study of theremoval of copper from aqueous solutions by adsorption usingsawdust Desalination 185 483ndash490 doi101016jdesal200503090
43 Oliveira WE Franca AS Oliveira LS Rocha SD (2008) Untreatedcoffee husks as biosorbents for the removal of heavy metals fromaqueous solutions J Hazard Mater 152 1073ndash1081 doi101016jjhazmat200707085 PubMed 17804159
44 Šciban M Klašnja M Škrbic B (2008) Adsorption of copper ions fromwater by modified agricultural by-products Desalination 229 170ndash180doi101016jdesal200708017
45 Maumlkelauml MR Hildeacuten KS Hakala TK Hatakka A Lundell TK (2006)Expression and molecular properties of a new laccase of the white rotfungus Phlebia radiata grown on wood Curr Genet 50 323-333 doi101007s00294-006-0090-1 PubMed 16927090
46 Maumlkelauml MR Lundell T Hatakka A Hildeacuten K (2013) Effect of coppernutrient nitrogen and wood-supplement on the production of lignin-modifying enzymes by the white-rot fungus Phlebia radiata Fungal Biol117 62-70 doi101016jfunbio201211006 PubMed 23332834
47 Dwivedi UN Singha P Pandeya VP Kumara A (2011) Structurendashfunction relationship among bacterial fungal and plant laccases J MolCatal B Enzym 68 117-128 doi101016jmolcatb201011002
48 Gaitan IJ Medina SC Gonzaacutelez JC Rodriacuteguez A Espejo AacuteJ et al(2011) Evaluation of toxicity and degradation of a chlorophenol mixtureby the laccase produced by Trametes pubescens Bioresour Technol102 3632ndash3635 doi101016jbiortech201011040 PubMed21115244
49 Karp SG Faraco V Amore A Birolo L Giangrande C et al (2012)Characterization of laccase isoforms produced by Pleurotus ostreatusin solid state fermentation of sugarcane bagasse Bioresour Technol114 735-739 doi101016jbiortech201203058 PubMed 22487128
50 Nikitina OV Shleev SV Gorshina ES Rusinova TV Serezhenkov VAet al (2005) Isolation and purification of enzymes from ligninolyticcomplex of the basidial fungus Trametes pubescens (Schumach) Pilatand study of their properties Biochemistry (Mosc) 70 1274-1279 doi101007s10541-005-0259-0
51 Shleev S Nikitina O Christenson A Reimann CT Yaropolov AI et al(2007) Characterization of two new multiforms of Trametes pubescenslaccase Bioorg Chem 35 35-49 doi101016jbioorg200608001PubMed 16989887
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 13 September 2013 | Volume 8 | Issue 9 | e73721
52 Xiao YZ Chen Q Hang J Shi YY Wu J et al (2004) Selectiveinduction purification and characterization of a laccase isozyme fromthe basidiomycete Trametes sp AH28-2 Mycologia 96 26-35 doi1023073761984 PubMed 21148825
53 Muntildeoz C Guilleacuten F Martiacutenez AT Martiacutenez MJ (1997) Laccaseisoenzymes of Pleurotus eryngii characterization catalytic propertiesand participation in activation of molecular oxygen and Mn2+ oxidationAppl Environ Microbiol 63 2166-2174 PubMed 9172335
54 Morozova OV Shumakovich GP Gorbacheva MA Shleev SVYaropolov AI (2007) Blue laccases Biochemistry (Mosc) 721136-1150 doi101134S0006297907100112
55 Hildeacuten K Hakala TK Lundell T (2009) Thermotolerant andthermostable laccases Biotechnol Lett 31 1117-1128 doi101007s10529-009-9998-0 PubMed 19360388
56 Verma P Madamwar D (2002) Production of ligninolytic enzymes fordye decolorization by cocultivation of white-rot fungi Pleurotusostreatus and Phanerochaete chrysosporium under solid-statefermentation Appl Biochem Biotechnol 102-103 109-118 doi101385ABAB102-1031-6109 PubMed 12396115
57 Meza JC Lomascolo A Casalot L Sigoillot J-C Auria R (2005)Laccase production by Pycnoporus cinnabarinus grown on sugar-canebagasse Influence of ethanol vapours as inducer Proc Biochem 403365ndash3371 doi101016jprocbio200503004
58 Hossain SM Anantharaman N (2006) Activity enhancement ofligninolytic enzymes of Trametes versicolor with bagasse powder Afr JBiotechnol 5 189-194
59 Revankar MS Desai KM Lele SS (2007) Solid-state fermentation forenhanced production of laccase using indigenously isolatedGanoderma sp Appl Biochem Biotechnol 143 16-26 doi101007s12010-007-0029-0 PubMed 18025593
60 Velazquez-Cedeno MA Mata G Savoie J-M (2002) Waste-reducingcultivation of Pleurotus ostreatus and Pleurotus pulmonarius on coffeepulp changes in the production of some lignocellulolytic enzymesWorld J Microbiol Biotechnol 18 201ndash207 doi101023A1014999616381
61 Mata G Murrieta D Andreu LI (2005) Changes in lignocellulolyticenzyme activities in six Pleurotus spp strains cultivated on coffee pulpin confrontation with Trichoderma spp World J Microbiol Biotechnol 21143ndash150 doi101007s11274-004-3041-3
62 de Souza DF Tychanowicz GK de Souza CG Peralta RM (2006) Co-production of ligninolytic enzymes by Pleurotus pulmonarius on wheatbran solid state cultures J Basic Microbiol 46 126-134 doi101002jobm200510014 PubMed 16598826
63 Prasad KK Mohan SV Bhaskar YV Ramanaiah SV Babu VL et al(2005) Laccase production using Pleurotus ostreatus 1804 immobilizedon PUF cubes in batch and packed bed reactors influence of cultureconditions J Microbiol 43 301-307 PubMed 15995650
64 Songulashvili G Elisashvili V Wasser SP Nevo E Hadar Y (2007)Basidiomycetes laccase and manganese peroxidase activity in
submerged fermentation of food industry wastes Enzyme MicrobTechnol 41 57ndash61 doi101016jenzmictec200611024
65 Sathishkumar P Murugesan K Palvannan T (2010) Production oflaccase from Pleurotus florida using agro-wastes and efficientdecolorization of Reactive blue 198 J Basic Microbiol 50 360-367 doi101002jobm200900407 PubMed 20586068
66 Moldes D Gallego PP Rodriacuteguez Couto S Sanromaacuten A (2003) Grapeseeds the best lignocellulosic waste to produce laccase by solid statecultures of Trametes hirsuta Biotechnol Lett 25 491-495 doi101023A1022660230653 PubMed 12882277
67 Elisashvili V Kachlishvili E Penninckx M (2008) Effect of growthsubstrate method of fermentation and nitrogen source onlignocellulose-degrading enzymes production by white-rotbasidiomycetes J Ind Microbiol Biotechnol 35 1531-1538 doi101007s10295-008-0454-2 PubMed 18716810
68 Osma JF Saravia V Herrera JL Couto SR (2007) Mandarin peelingsthe best carbon source to produce laccase by static cultures ofTrametes pubescens Chemosphere 67 1677-1680 doi101016jchemosphere200611051 PubMed 17234250
69 Kahraman SS Gurdal IH (2002) Effect of synthetic and natural culturemedia on laccase production by white rot fungi Bioresour Technol 82215-217 doi101016S0960-8524(01)00193-6 PubMed 11991068
70 Iandolo D Piscitelli A Sannia G Faraco V (2011) Enzyme Productionby Solid Substrate Fermentation of Pleurotus ostreatus and Trametesversicolor on Tomato Pomace Appl Biochem Biotechnol 163 40-51doi101007s12010-010-9014-0 PubMed 20582639
71 Moldes D Lorenzo M Sanromaacuten MA (2004) Different proportions oflaccase isoenzymes produced by submerged cultures of Trametesversicolor grown on lignocellulosic wastes Biotechnol Lett 26 327-330doi101023BBILE000001545240213bf PubMed 15055770
72 Philippoussis A Diamantopoulou P Papadopoulou K Lakhtar HRoussos S et al (2010) Biomass laccase and endoglucanaseproduction by Lentinula edodes during solid state fermentation of reedgrass bean stalks and wheat straw residues World J MicrobiolBiotechnol 27 285-297
73 Fenice M Giovannozzi Sermanni G Federici F DrsquoAnnibale A (2003)Submerged and solid-state production of laccase and Mn-peroxidaseby Panus tigrinus on olive mill wastewater-based media J Biotechnol100 77-85 doi101016S0168-1656(02)00241-9 PubMed 12413788
74 Rodriguez-Couto S GundIn M Lorenzo M Sanromaacuten MAacute (2002)Screening of supports and inducers for laccase production by Trametesversicolor in semi-solid-state conditions Proc Biochem 38 249-255doi101016S0032-9592(02)00087-0
75 Fackler K Gradinger C Schmutzer M Tavzes C Burgert I et al (2007)Biotechnological wood modification with selective white-rot fungi and itsmolecular mechanisms Food Technol Biotechnol 45 269ndash276
76 Levin L Herrmann C Papinutti VL (2008) Optimization oflignocellulolytic enzyme production by the white-rot fungus Trametestrogii in solid-state fermentation using response surface methodologyBiochem Eng J 39 207-214 doi101016jbej200709004
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 14 September 2013 | Volume 8 | Issue 9 | e73721

52 Xiao YZ Chen Q Hang J Shi YY Wu J et al (2004) Selectiveinduction purification and characterization of a laccase isozyme fromthe basidiomycete Trametes sp AH28-2 Mycologia 96 26-35 doi1023073761984 PubMed 21148825
53 Muntildeoz C Guilleacuten F Martiacutenez AT Martiacutenez MJ (1997) Laccaseisoenzymes of Pleurotus eryngii characterization catalytic propertiesand participation in activation of molecular oxygen and Mn2+ oxidationAppl Environ Microbiol 63 2166-2174 PubMed 9172335
54 Morozova OV Shumakovich GP Gorbacheva MA Shleev SVYaropolov AI (2007) Blue laccases Biochemistry (Mosc) 721136-1150 doi101134S0006297907100112
55 Hildeacuten K Hakala TK Lundell T (2009) Thermotolerant andthermostable laccases Biotechnol Lett 31 1117-1128 doi101007s10529-009-9998-0 PubMed 19360388
56 Verma P Madamwar D (2002) Production of ligninolytic enzymes fordye decolorization by cocultivation of white-rot fungi Pleurotusostreatus and Phanerochaete chrysosporium under solid-statefermentation Appl Biochem Biotechnol 102-103 109-118 doi101385ABAB102-1031-6109 PubMed 12396115
57 Meza JC Lomascolo A Casalot L Sigoillot J-C Auria R (2005)Laccase production by Pycnoporus cinnabarinus grown on sugar-canebagasse Influence of ethanol vapours as inducer Proc Biochem 403365ndash3371 doi101016jprocbio200503004
58 Hossain SM Anantharaman N (2006) Activity enhancement ofligninolytic enzymes of Trametes versicolor with bagasse powder Afr JBiotechnol 5 189-194
59 Revankar MS Desai KM Lele SS (2007) Solid-state fermentation forenhanced production of laccase using indigenously isolatedGanoderma sp Appl Biochem Biotechnol 143 16-26 doi101007s12010-007-0029-0 PubMed 18025593
60 Velazquez-Cedeno MA Mata G Savoie J-M (2002) Waste-reducingcultivation of Pleurotus ostreatus and Pleurotus pulmonarius on coffeepulp changes in the production of some lignocellulolytic enzymesWorld J Microbiol Biotechnol 18 201ndash207 doi101023A1014999616381
61 Mata G Murrieta D Andreu LI (2005) Changes in lignocellulolyticenzyme activities in six Pleurotus spp strains cultivated on coffee pulpin confrontation with Trichoderma spp World J Microbiol Biotechnol 21143ndash150 doi101007s11274-004-3041-3
62 de Souza DF Tychanowicz GK de Souza CG Peralta RM (2006) Co-production of ligninolytic enzymes by Pleurotus pulmonarius on wheatbran solid state cultures J Basic Microbiol 46 126-134 doi101002jobm200510014 PubMed 16598826
63 Prasad KK Mohan SV Bhaskar YV Ramanaiah SV Babu VL et al(2005) Laccase production using Pleurotus ostreatus 1804 immobilizedon PUF cubes in batch and packed bed reactors influence of cultureconditions J Microbiol 43 301-307 PubMed 15995650
64 Songulashvili G Elisashvili V Wasser SP Nevo E Hadar Y (2007)Basidiomycetes laccase and manganese peroxidase activity in
submerged fermentation of food industry wastes Enzyme MicrobTechnol 41 57ndash61 doi101016jenzmictec200611024
65 Sathishkumar P Murugesan K Palvannan T (2010) Production oflaccase from Pleurotus florida using agro-wastes and efficientdecolorization of Reactive blue 198 J Basic Microbiol 50 360-367 doi101002jobm200900407 PubMed 20586068
66 Moldes D Gallego PP Rodriacuteguez Couto S Sanromaacuten A (2003) Grapeseeds the best lignocellulosic waste to produce laccase by solid statecultures of Trametes hirsuta Biotechnol Lett 25 491-495 doi101023A1022660230653 PubMed 12882277
67 Elisashvili V Kachlishvili E Penninckx M (2008) Effect of growthsubstrate method of fermentation and nitrogen source onlignocellulose-degrading enzymes production by white-rotbasidiomycetes J Ind Microbiol Biotechnol 35 1531-1538 doi101007s10295-008-0454-2 PubMed 18716810
68 Osma JF Saravia V Herrera JL Couto SR (2007) Mandarin peelingsthe best carbon source to produce laccase by static cultures ofTrametes pubescens Chemosphere 67 1677-1680 doi101016jchemosphere200611051 PubMed 17234250
69 Kahraman SS Gurdal IH (2002) Effect of synthetic and natural culturemedia on laccase production by white rot fungi Bioresour Technol 82215-217 doi101016S0960-8524(01)00193-6 PubMed 11991068
70 Iandolo D Piscitelli A Sannia G Faraco V (2011) Enzyme Productionby Solid Substrate Fermentation of Pleurotus ostreatus and Trametesversicolor on Tomato Pomace Appl Biochem Biotechnol 163 40-51doi101007s12010-010-9014-0 PubMed 20582639
71 Moldes D Lorenzo M Sanromaacuten MA (2004) Different proportions oflaccase isoenzymes produced by submerged cultures of Trametesversicolor grown on lignocellulosic wastes Biotechnol Lett 26 327-330doi101023BBILE000001545240213bf PubMed 15055770
72 Philippoussis A Diamantopoulou P Papadopoulou K Lakhtar HRoussos S et al (2010) Biomass laccase and endoglucanaseproduction by Lentinula edodes during solid state fermentation of reedgrass bean stalks and wheat straw residues World J MicrobiolBiotechnol 27 285-297
73 Fenice M Giovannozzi Sermanni G Federici F DrsquoAnnibale A (2003)Submerged and solid-state production of laccase and Mn-peroxidaseby Panus tigrinus on olive mill wastewater-based media J Biotechnol100 77-85 doi101016S0168-1656(02)00241-9 PubMed 12413788
74 Rodriguez-Couto S GundIn M Lorenzo M Sanromaacuten MAacute (2002)Screening of supports and inducers for laccase production by Trametesversicolor in semi-solid-state conditions Proc Biochem 38 249-255doi101016S0032-9592(02)00087-0
75 Fackler K Gradinger C Schmutzer M Tavzes C Burgert I et al (2007)Biotechnological wood modification with selective white-rot fungi and itsmolecular mechanisms Food Technol Biotechnol 45 269ndash276
76 Levin L Herrmann C Papinutti VL (2008) Optimization oflignocellulolytic enzyme production by the white-rot fungus Trametestrogii in solid-state fermentation using response surface methodologyBiochem Eng J 39 207-214 doi101016jbej200709004
Production of T pubescens Laccase
PLOS ONE | wwwplosoneorg 14 September 2013 | Volume 8 | Issue 9 | e73721
Related Documents











