Production of knockout mice by gene targeting in multipotent germline stem cells Masanori Takehashi a,1 , Mito Kanatsu-Shinohara a , Hiromi Miki b , Jiyoung Lee a , Yasuhiro Kazuki c , Kimiko Inoue b , Narumi Ogonuki b , Shinya Toyokuni d , Mitsuo Oshimura c , Atsuo Ogura b , Takashi Shinohara a, ⁎ a Department of Molecular Genetics, Graduate School of Medicine, Kyoto University, Yoshida-Konoe, Sakyo-ku, Kyoto 606-8501, Japan b The Institute of Physical and Chemical Research (RIKEN), Bioresource Center, Ibaraki 305-0074, Japan c Department of Molecular and Cell Genetics, School of Life Sciences, Faculty of Medicine, Tottori University, Yonago, Tottori 683-8503, Japan d Department of Pathology and Biology of Diseases, Graduate School of Medicine, Kyoto University, Kyoto 606-8501, Japan Received for publication 17 March 2007; revised 19 August 2007; accepted 18 September 2007 Available online 26 September 2007 Abstract Spermatogonial stem cells can convert into embryonic stem (ES) cell-like multipotent germline stem (mGS) cells in vitro and produce germline chimeras by blastocyst microinjection. Although homologous recombination was previously demonstrated in mGS cells, spermatogenesis was not found in chimeras, suggesting that they are not competent for germline modification. Here we conducted detailed analysis of chimeric animals to determine whether mGS cells retain germline potential after genetic manipulation. Spermatozoa that were deficient in the occludin gene could be recovered from animals that were chimeric with mGS cells that underwent homologous recombination. The phenotypes of the occludin knockout (KO) mice were similar to those reported for KO mice produced using ES cells, and the animals showed growth retardation, gastritis and male infertility. Furthermore, we found that heterozygous mGS cells acquire two copies of the G418-resistant genes and become homozygous for the targeted allele by culturing at high concentrations of G418. Cytogenetic analysis showed that the aneuploid mGS cells observed during genetic manipulation were trisomic for chromosome 8 or 11, which is a common chromosomal abnormality in ES cells. Thus, mGS cells can be used to produce KO animals, and this novel method of germline manipulation may prove useful in diverse mammalian species. © 2007 Elsevier Inc. All rights reserved. Keywords: Stem cell; Spermatogenesis; Pluripotency; Germ cell; Gene targeting; Microinsemination Introduction Germline cells are unique in that they transmit genetic information from parent to offspring (de Rooij and Russell, 2000; Meistrich and van Beek, 1993). In the postnatal period, only male germ cells initiate mitosis to become spermatogonial stem cells. These cells self-renew and also produce progenitor cells that eventually mature into spermatozoa. Female germline cells stop dividing after birth in most species, whereas spermatogonial stem cells continue to proliferate and support spermatogenesis throughout life in males. Although germ cells are normally committed to producing mature gametes, several studies have shown that these cells are able to change their fate and convert into pluripotent cell types under certain conditions. Primordial germ cells (PGCs) originate from the epiblast 7.5 days postcoitum (dpc) in mice (McLaren, 2003). These cells actively proliferate and migrate to the genital ridge in the middle of the gestation. By 11.5 dpc, most germ cells are located at the genital ridge, and female germ cells enter meiosis by 13.5 dpc. In contrast, by then male germ cells become proliferating gonocytes in the seminiferous cords, and the gonocytes stop proliferating at about 16 dpc. Between 8.5 and 12.5 dpc, mouse embryo PGCs can convert into Available online at www.sciencedirect.com Developmental Biology 312 (2007) 344 – 352 www.elsevier.com/developmentalbiology ⁎ Corresponding author. Fax: +81 75 751 4169. E-mail address: [email protected] (T. Shinohara). 1 Present address: Laboratory of Molecular Clinical Chemistry, Faculty of Pharmacy, Osaka Ohtani University, Tondabayashi, Osaka 584-8540, Japan. 0012-1606/$ - see front matter © 2007 Elsevier Inc. All rights reserved. doi:10.1016/j.ydbio.2007.09.029

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Available online at www.sciencedirect.com
12 (2007) 344–352www.elsevier.com/developmentalbiology
Developmental Biology 3
Production of knockout mice by gene targeting inmultipotent germline stem cells
Masanori Takehashi a,1, Mito Kanatsu-Shinohara a, Hiromi Miki b, Jiyoung Lee a,Yasuhiro Kazuki c, Kimiko Inoue b, Narumi Ogonuki b, Shinya Toyokuni d,
Mitsuo Oshimura c, Atsuo Ogura b, Takashi Shinohara a,⁎
a Department of Molecular Genetics, Graduate School of Medicine, Kyoto University, Yoshida-Konoe, Sakyo-ku, Kyoto 606-8501, Japanb The Institute of Physical and Chemical Research (RIKEN), Bioresource Center, Ibaraki 305-0074, Japan
c Department of Molecular and Cell Genetics, School of Life Sciences, Faculty of Medicine, Tottori University, Yonago, Tottori 683-8503, Japand Department of Pathology and Biology of Diseases, Graduate School of Medicine, Kyoto University, Kyoto 606-8501, Japan
Received for publication 17 March 2007; revised 19 August 2007; accepted 18 September 2007Available online 26 September 2007
Abstract
Spermatogonial stem cells can convert into embryonic stem (ES) cell-like multipotent germline stem (mGS) cells in vitro and produce germlinechimeras by blastocyst microinjection. Although homologous recombination was previously demonstrated in mGS cells, spermatogenesis was notfound in chimeras, suggesting that they are not competent for germline modification. Here we conducted detailed analysis of chimeric animals todetermine whether mGS cells retain germline potential after genetic manipulation. Spermatozoa that were deficient in the occludin gene could berecovered from animals that were chimeric with mGS cells that underwent homologous recombination. The phenotypes of the occludin knockout(KO) mice were similar to those reported for KO mice produced using ES cells, and the animals showed growth retardation, gastritis and maleinfertility. Furthermore, we found that heterozygous mGS cells acquire two copies of the G418-resistant genes and become homozygous for thetargeted allele by culturing at high concentrations of G418. Cytogenetic analysis showed that the aneuploid mGS cells observed during geneticmanipulation were trisomic for chromosome 8 or 11, which is a common chromosomal abnormality in ES cells. Thus, mGS cells can be used toproduce KO animals, and this novel method of germline manipulation may prove useful in diverse mammalian species.© 2007 Elsevier Inc. All rights reserved.
Keywords: Stem cell; Spermatogenesis; Pluripotency; Germ cell; Gene targeting; Microinsemination
Introduction
Germline cells are unique in that they transmit geneticinformation from parent to offspring (de Rooij and Russell,2000; Meistrich and van Beek, 1993). In the postnatal period,only male germ cells initiate mitosis to become spermatogonialstem cells. These cells self-renew and also produce progenitorcells that eventually mature into spermatozoa. Female germlinecells stop dividing after birth in most species, whereas
⁎ Corresponding author. Fax: +81 75 751 4169.E-mail address: [email protected] (T. Shinohara).
1 Present address: Laboratory of Molecular Clinical Chemistry, Faculty ofPharmacy, Osaka Ohtani University, Tondabayashi, Osaka 584-8540, Japan.
0012-1606/$ - see front matter © 2007 Elsevier Inc. All rights reserved.doi:10.1016/j.ydbio.2007.09.029
spermatogonial stem cells continue to proliferate and supportspermatogenesis throughout life in males.
Although germ cells are normally committed to producingmature gametes, several studies have shown that these cells areable to change their fate and convert into pluripotent cell typesunder certain conditions. Primordial germ cells (PGCs)originate from the epiblast 7.5 days postcoitum (dpc) in mice(McLaren, 2003). These cells actively proliferate and migrate tothe genital ridge in the middle of the gestation. By 11.5 dpc,most germ cells are located at the genital ridge, and female germcells enter meiosis by 13.5 dpc. In contrast, by then male germcells become proliferating gonocytes in the seminiferous cords,and the gonocytes stop proliferating at about 16 dpc. Between8.5 and 12.5 dpc, mouse embryo PGCs can convert into

345M. Takehashi et al. / Developmental Biology 312 (2007) 344–352
pluripotent stem cells, known as embryonic germ (EG) cells,when they are cultured in the presence of stem cell factor (SCF),leukemia inhibitory factor (LIF) and basic fibroblast growthfactor (bFGF) (Matsui et al., 1992; Resnick et al., 1992). Thephenotype and differentiation capacity of these cells are similarto those of ES cells derived from the inner cell mass (Evans andKaufman, 1981; Martin, 1981). EG cells form embryoid bodiesand produce differentiated somatic cell types. They alsocontribute to embryonic development and produce germlinechimeras when microinjected into blastocysts (Labosky et al.,1994; Stewart et al., 1994). These studies demonstrated for thefirst time that germline cells can de-differentiate and acquirepluripotency. EG cells have also been derived from non-murinespecies, including humans (Shim et al., 1997; Shamblott et al.,1998; Park and Han, 2000); however, since EG cells could notbe derived from germline cells at later developmental stages, ithas been suggested that pluripotency is lost during the late fetalstage.
The characteristics of EG cells suggest that they may beused as vehicles for germline transgenesis in a manner similarto ES cells. However, there are no reports of the productionof knockout (KO) mice using EG cells. The reason for thisfailure is unclear, but it may be related to genomic imprintingsince the only difference between ES and EG cells is thegenomic imprinting pattern; the level of site-specificmethylation for certain imprinted genes is reduced in EGcells derived from embryos 11.5 and 12.5 dpc (Labosky etal., 1994; Tada et al., 1998). Since genomic imprinting playsan important role in regulating the differentiation capacity ofthe cell, pluripotent cells derived from committed germ cellsmay be less stable than ES cells in terms of their germlinepotential.
We previously showed that spermatogonial stem cells frompostnatal testes could be expanded over the long-term in vitrounder culture conditions that included glial cell line-derivedneurotrophic factor (GDNF), LIF, bFGF and epidermal growthfactor (Kanatsu-Shinohara et al., 2003). These cultured cells,called germline stem (GS) cells, continued to proliferate for atleast 2 years in vitro, and they maintained their capacity toproduce normal sperm and fertile offspring after transplantationinto testes (Kanatsu-Shinohara et al., 2005a). GS cells exhibitthe characteristics of spermatogonial stem cells, and they arenot pluripotent. Surprisingly, however, we recently found thatES cell-like pluripotent stem cells appeared in a GS cell cultureduring the establishment phase from neonatal testes (Kanatsu-Shinohara et al., 2004a). Similar to ES/EG cells, these ES-likecells, which we designated as multipotent germline stem(mGS) cells, not only formed teratomas in vivo, but alsodifferentiated into a variety of somatic cell types in vitro. Thecells also produced germline chimeras by microinjection intoblastocysts, demonstrating that pluripotency persists in thesepostnatal germ cells. Although several lines of evidencesuggest that both cell types originated from spermatogonialstem cells, mGS cells cannot undergo spermatogenesis in thetestis, while GS cells produce sperm in the testis but do notcontribute to embryonic development (Kanatsu-Shinohara andShinohara, 2006).
Since GS and mGS cells can both be manipulated in vitroand produce offspring, the recent development of stem cellculture techniques for these cell types opens up newpossibilities for germline transgenesis. Recently, we succeededin producing KO mice by gene trapping and gene targetingusing GS cells, but we were unable to produce KO mice usingmGS cells (Kanatsu-Shinohara et al., 2006a). The mGS cellsthat underwent homologous recombination failed to contributeto the germline following blastocyst injection. The failure toproduce KO animals with EG and mGS cells suggests thatpluripotent stem cells derived from germ cells have lowergermline stability and limited potential for genetic modification.Here we report the successful production of KO mice usingmGS cells. The phenotypes of the mice were similar to thosemade by gene targeting in ES and GS cells.
Materials and methods
Cell culture
GS cells were cultured according to a previously established protocol(Kanatsu-Shinohara et al., 2003), whereas mGS cells were cultured on mouseembryonic fibroblasts (MEFs) in standard ES cell culture medium, whichconsisted of Dulbecco's modified Eagle's medium/15% fetal calf serum and103 U/ml LIF (ESGRO, Invitrogen, Carlsbad, CA)(Kanatsu-Shinohara et al.,2004a). For selection of the double KO cells, GS and mGS cells were culturedon MEFs with various concentrations of G418 (Wako, Osaka, Japan). Theselected clones were transferred to a 96-well plate and expanded for DNAanalysis. G418 resistance was assessed by plating 3×104 cells on G418-resistantMEFs in a 12-well culture plate, and the number of surviving colonies wascounted after 2 weeks of selection with 20 mg/ml G418. Embryoid bodies wereproduced by culturing mGS cells on bacterial dishes in the absence of LIF, asdescribed previously (Saitou et al., 1998).
Transplantation
For teratoma formation, approximately 4×106 double KO mGS cells wereinjected subcutaneously into KSN nude mice (Japan SLC, Shizuoka, Japan).The teratomas were recovered at 4–6 weeks after transplantation and analyzedby histological section. The Institutional Animal Care and Use Committee ofKyoto University approved all animal experimentation protocols.
Southern blot analysis
Genomic DNA was isolated by phenol/chloroform extraction, followed byethanol precipitation. DNA transfer and hybridization were performed asdescribed previously using probes for both exon 2 (internal) and intron 4(external) probes (Kanatsu-Shinohara et al., 2005b, 2006a). The clones werealso examined by hybridization with a cDNA fragment of the neo gene(nucleotides 187 to 804).
Combined bisulfite restriction analysis (COBRA)
The methylation status of the imprinted genes was assessed by COBRA, asdescribed previously (Kanatsu-Shinohara et al., 2005a). The polymerase chainreaction (PCR) products were digested with restriction enzymes that recognizethe CpG motif. The intensity of the digested DNAwas quantified using Mac Basversion 2.5 software (Fuji Photo Film, Tokyo, Japan).
Histological analysis
For histological analysis, the samples were fixed in 10% neutral bufferedformalin and processed for paraffin sectioning. All sections were stained withhematoxylin and eosin.

346 M. Takehashi et al. / Developmental Biology 312 (2007) 344–352
Reverse transcriptase-polymerase chain reaction (RT-PCR) andWestern blot analysis
Total RNA was prepared using TRIzol Reagent (Invitrogen). For RT-PCR,first strand cDNA was synthesized using Superscript™ II (RNase H− reversetranscriptase, Invitrogen). PCR was performed using primers specific for themouse occludin gene, as described previously (Saitou et al., 1998).
For the Western blots, the samples were lysed by homogenization with ice-cold lysis buffer (50 mM Tris–HCl, pH 6.8, 2% SDS, 20% glycerol, 2% 2-mercaptoethanol, 0.01% bromophenol blue) supplemented with a proteaseinhibitor cocktail (Sigma, St. Louis, MO). Whole-cell extracts were separated by10% SDS–polyacrylamide gel electrophoresis, transferred to Hybond-Pmembranes (Amersham Biosciences, Buckinghamshire, UK), blocked with5% skim milk in Tris-buffered saline containing 0.1% Triton X-100 and probedwith rabbit polyclonal anti-mouse occludin antibody (a gift from Dr. S. Tsukita,Kyoto University). Peroxidase-conjugated anti-rabbit IgG antibody (CellSignaling, Danvers, MA) was used as the secondary antibody, and the proteinbands were detected using the enhanced chemiluminescence detection method(LumiGLO™; Cell Signaling).
Microinsemination
The seminiferous tubules of the chimeric animals were dissected under UVlight. Enhanced green fluorescent protein (EGFP)-expressing seminiferoustubules were recovered, and the germ cells were collected mechanically usingfine forceps. Microinsemination was performed as described previously usinground spermatids (Kimura and Yanagimachi, 1995). Embryos that reached thetwo-cell stage after 24 h of culture were transferred to the oviducts of day 1pseudopregnant ICR female mice. Fetuses that were retrieved on day 19.5 wereraised by ICR foster mothers.
Chromosome analysis
The wild-type, heterozygous KO and double KO mGS cells harvested fromculture dishes were treated with 0.075 M KCl for 15 min and were fixed withmethanol/acetic acid (3:1). The chromosome slides were prepared using thestandard method. The slides were stained with quinacrine mustard and Hoechst33258. At least 20 metaphase cells were karyotyped for each cell type. Thechromosomal aberrations, trisomic for chromosome 8 and/or 11, were confirmedby fluorescence in situ hybridization (FISH) analysis using mouse chromosome8- and 11-specific probes (TAKARA BIO, Otsu, Japan).
Results
Production of KO mice by in vitro microinsemination usingtesticular sperm recovered from mGS chimeric mice
In a previous study, we generated chimeric animals using anEGFP-expressingmGS cell clone (clone 118) that was targeted tothe mouse occludin gene locus (Kanatsu-Shinohara et al., 2006a)(Figs. 1A–C). Of the 43 offspring (14 males and 29 females) thatmatured into adults, we sacrificed seven males and 15 females.Although we found EGFP-positive cells in the ovaries of twofemales, we were not able to determine whether the mGS cells inthe ovaries that contributed to the germ cells, somatic cells, orboth. Furthermore, none of the seven males had contributions inthe germline, as shown by EGFP fluorescence (Kanatsu-Shinohara et al., 2006a). We sacrificed the remaining sevenmales to examine whether germline transmission had occurredwith themGS cells and found that the seminiferous tubules of oneof the animals showed EGFP fluorescence under UV light (Fig.1D). This animal also had donor mGS cell contributions in itsheart, pancreas and other organs (data not shown).
To generate offspring from this chimeric male, we used invitro microinsemination, a technique commonly used toproduce offspring in infertile animals and humans (Palermo etal., 1992; Kimura and Yanagimachi, 1995). EGFP-expressingseminiferous tubules were dissociated using fine forceps, andthe round spermatids thus recovered were used for in vitromicroinsemination (Figs. 1E and F). Of the 220 embryosconstructed, 67 developed to the two-cell stage, and these weretransferred into pseudopregnant ICR mothers. Ten offspringwere born, and nine developed into normal mature adults (fourmales and five females). PCR analysis revealed that two of themales and two of the females were heterozygous for the targetedallele (Fig. 1G). COBRA revealed normal imprinting patterns inthe H19, Meg3 IG, Igf2r and Peg 10 regions in the offspring,indicating that erasure and acquisition of the normal imprint hadoccurred successfully during germline development in thechimeric male (Fig. 1H).
To examine whether gene targeting in spermatogonial stemcells influences the phenotypes of KO animals, we generatedtwo types of KO mice using mGS cells. One of theheterozygous males was bred with heterozygous femalesderived from gene targeting in GS cells (Kanatsu-Shinohara etal., 2006a), while the other heterozygous male was crossed withthe heterozygous females produced from mGS cells. Theintercrosses between the heterozygous mGS KO male and theGS KO females yielded offspring that segregated with theexpected Mendelian distribution: 12 wild-type, 23 heterozygousand 9 homozygous mutant (mGS/GS KO) pups were obtainedfrom 6 litters. Likewise, intercrosses between the mGSheterozygous offspring produced 14 wild-type, 29 heterozygousand 9 homozygous mutant (mGS/mGS KO) pups in 8 litters.The lack of occludin mRNA and protein was confirmed in bothtypes of KO mice (Figs. 1I and J). These results indicate thatmGS cells can be used to produce KO mice by homologousrecombination and germline transmission in a manner similar toES cells.
Phenotypes of the KO mice
Results from ES cell-based occludin gene-targeting experi-ments demonstrated that homozygous occludin KO mice hadpostnatal growth retardation but no gross macroscopic pheno-type (Saitou et al., 1998; Saitou et al., 2000). In agreement withthese observations, both the mGS/mGS and mGS/GS KO micewere normal in appearance, but they both showed growthretardation. Growth retardation was observed in both the malesand females; both mGS/mGS and mGS/GS KO mice were∼70% of the normal weight at 8 weeks of age (Fig. 2A).Similarly, the homozygous KO mice produced by genetargeting in GS cells (GS/GS KO) were ∼70% of the normalweight at 8 weeks of age (Fig. 2A). Even at more mature stages,the average weight of both types of KO mice was less than thatof the wild-type animals.
We then conducted histological analyses of several organsfrom the KO mice. Previously, we found that the striated ductsof the salivary glands of occludin KO (GS/GS KO) mice lackedcharacteristic cytoplasmic granules (Kanatsu-Shinohara et al.,

347M. Takehashi et al. / Developmental Biology 312 (2007) 344–352
2006a) (Figs. 2B and C). Additionally, in the gastric glandshowed predominant hyperplasia and numerous lymphocyteshad infiltrated the gastric epithelium, indicating chronic gastritis(Figs. 2D and E). Furthermore, the KO animals had osteoporo-sis and exhibited significantly thinner trabeculae than did the
wild-type animals (Figs. 2F and G). The animals also showedother pathological features, such as calcification in the brain(Figs. 2H and I). Similar abnormalities were found in all KOmice groups (mGS/mGS, mGS/GS and GS/GS), which isconsistent with the findings in ES cell-based KO mice (Saitou etal., 2000).
As the KO animals aged, their abnormalities graduallyworsened. All KO animals had regenerating gastric epitheliadue to constant inflammation. The most striking change,however, was observed in the testes. Although normalspermatogenesis was observed in young animals, germ cellswere progressively lost from the seminiferous tubules of olderanimals (Figs. 3A–F). When the testes were examined 9 monthsafter birth, the number of spermatogenic cells was decreasedpossibly due to decreased proliferation of the spermatogonialstem cells, and only a small number of postmeiotic germ cellswere found in these animals (Fig. 3E). The epididymides ofthese animals did not contain any spermatozoa (Fig. 3F), whichis consistent with previous observations in ES cell-based KOanimals (Saitou et al., 2000).
As expected from our histological findings, the male KOmice either produced significantly smaller numbers of offspringor were completely infertile. We analyzed the fertility of GS/GSand mGS/mGS KO males by mating with wild-type females. Inthe case of the GS/GS KO mice, while all three wild-type malesproduced a total of 57 litters during a 6-month period, two of thethree KO males produced 3 litters. The number of littersgradually decreased and the KO males were infertile when theyare older than 3 month of age (Fig. 3G). Likewise, while fourwild-type males produced a total of 19 litters, three of the fourmGS/mGS KO males produced a total of 3 litters during a 2-month period. As with GS/GS KO males, the mGS/mGS KOanimals were able to produce offspring only when the animalswere less than 3 month old. In contrast, all female KO micewere fertile, and germline transmission of the targeted allele wasconfirmed in all groups (Fig. 1K).
Fig. 1. Production of occludin KO mice using mGS cells. (A) Diagram of thetargeting strategy. B, BamHI; E, EcoRV; H, HindIII; N, NheI; P, PstI; X, XbaI.P1 and P2 represent the primers used for PCR. (B, C) Appearance of the targetedmGS cells (clone 118) (B), showing fluorescence under UV light (C). (D)Macroscopic appearance of the testis from a chimeric animal. Several EGFP-positive seminiferous tubules were found under UV illumination. (E, F)Spermatogenic cells released from part of a segment of seminiferous tubule thatshowed EGFP fluorescence. Nomarski (E) and fluorescent (F) images. Thearrowheads indicate the round spermatids that were used in microinsemination.(G) PCR analysis of gene targeting in the offspring. Tail DNA samples wereanalyzed by PCR to amplify a 2.4-kb fragment spanning the 3′ junction of thetargeted locus. (H) COBRA of genomic DNA from targeted mGS cells (clone118) and F1 offspring derived from microinsemination. DNA methylation statusof the target locus. DNA was amplified using specific primers and the PCRproducts were digested with the indicated restriction enzymes. The percentage ofmethylation, estimated by the intensity of the individual bands, is indicatedbelow the gels. U, uncleaved; C, cleaved. (I, J) The expression of occludinmRNA (I) and protein (J) in the lungs and the kidneys of wild-type andhomozygous mGS/mGS and mGS/GS KO mice. Both occludin mRNA (624-bpfragment) and protein (∼66 kDa) were absent in both types of homozygousmice, as detected by RT-PCR and Western blotting. L, lung; K, kidney.(K) Pedigree showing the transmission of the targeted allele through threegenerations. BDF1, C57BL/6×DBA/2. Scale bar=50 μm (B, C).
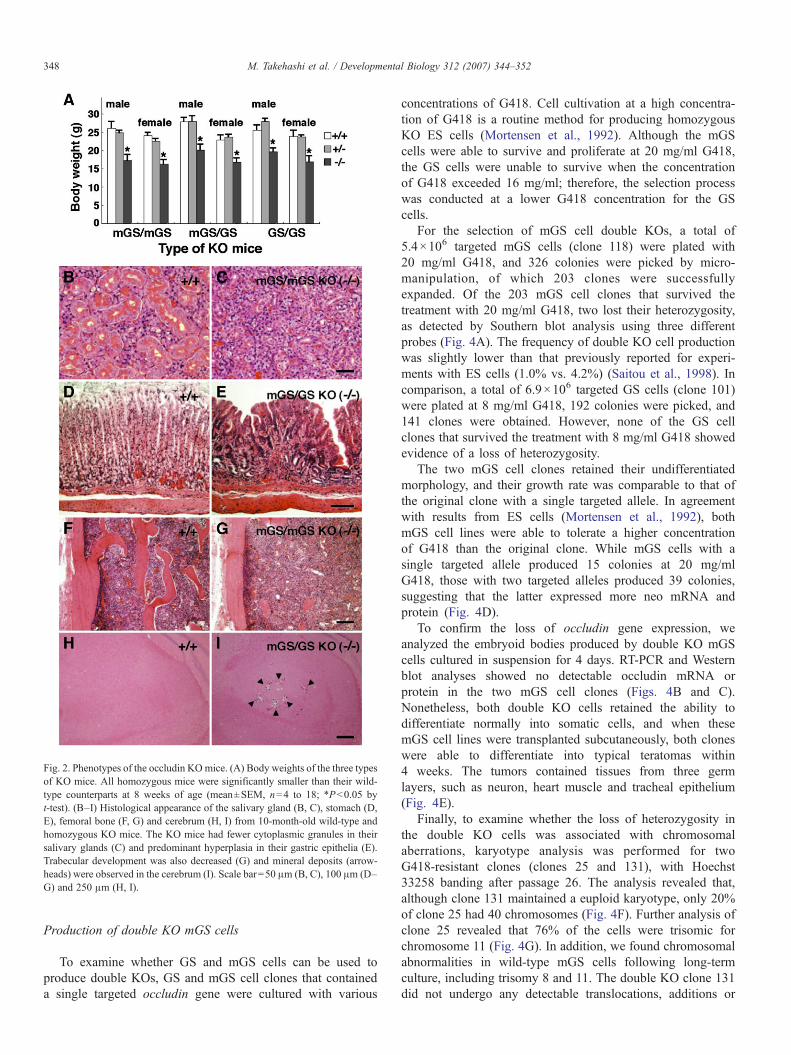
Fig. 2. Phenotypes of the occludin KOmice. (A) Body weights of the three typesof KO mice. All homozygous mice were significantly smaller than their wild-type counterparts at 8 weeks of age (mean±SEM, n=4 to 18; ∗Pb0.05 byt-test). (B–I) Histological appearance of the salivary gland (B, C), stomach (D,E), femoral bone (F, G) and cerebrum (H, I) from 10-month-old wild-type andhomozygous KO mice. The KO mice had fewer cytoplasmic granules in theirsalivary glands (C) and predominant hyperplasia in their gastric epithelia (E).Trabecular development was also decreased (G) and mineral deposits (arrow-heads) were observed in the cerebrum (I). Scale bar=50 μm (B, C), 100 μm (D–G) and 250 μm (H, I).
348 M. Takehashi et al. / Developmental Biology 312 (2007) 344–352
Production of double KO mGS cells
To examine whether GS and mGS cells can be used toproduce double KOs, GS and mGS cell clones that containeda single targeted occludin gene were cultured with various
concentrations of G418. Cell cultivation at a high concentra-tion of G418 is a routine method for producing homozygousKO ES cells (Mortensen et al., 1992). Although the mGScells were able to survive and proliferate at 20 mg/ml G418,the GS cells were unable to survive when the concentrationof G418 exceeded 16 mg/ml; therefore, the selection processwas conducted at a lower G418 concentration for the GScells.
For the selection of mGS cell double KOs, a total of5.4×106 targeted mGS cells (clone 118) were plated with20 mg/ml G418, and 326 colonies were picked by micro-manipulation, of which 203 clones were successfullyexpanded. Of the 203 mGS cell clones that survived thetreatment with 20 mg/ml G418, two lost their heterozygosity,as detected by Southern blot analysis using three differentprobes (Fig. 4A). The frequency of double KO cell productionwas slightly lower than that previously reported for experi-ments with ES cells (1.0% vs. 4.2%) (Saitou et al., 1998). Incomparison, a total of 6.9×106 targeted GS cells (clone 101)were plated at 8 mg/ml G418, 192 colonies were picked, and141 clones were obtained. However, none of the GS cellclones that survived the treatment with 8 mg/ml G418 showedevidence of a loss of heterozygosity.
The two mGS cell clones retained their undifferentiatedmorphology, and their growth rate was comparable to that ofthe original clone with a single targeted allele. In agreementwith results from ES cells (Mortensen et al., 1992), bothmGS cell lines were able to tolerate a higher concentrationof G418 than the original clone. While mGS cells with asingle targeted allele produced 15 colonies at 20 mg/mlG418, those with two targeted alleles produced 39 colonies,suggesting that the latter expressed more neo mRNA andprotein (Fig. 4D).
To confirm the loss of occludin gene expression, weanalyzed the embryoid bodies produced by double KO mGScells cultured in suspension for 4 days. RT-PCR and Westernblot analyses showed no detectable occludin mRNA orprotein in the two mGS cell clones (Figs. 4B and C).Nonetheless, both double KO cells retained the ability todifferentiate normally into somatic cells, and when thesemGS cell lines were transplanted subcutaneously, both cloneswere able to differentiate into typical teratomas within4 weeks. The tumors contained tissues from three germlayers, such as neuron, heart muscle and tracheal epithelium(Fig. 4E).
Finally, to examine whether the loss of heterozygosity inthe double KO cells was associated with chromosomalaberrations, karyotype analysis was performed for twoG418-resistant clones (clones 25 and 131), with Hoechst33258 banding after passage 26. The analysis revealed that,although clone 131 maintained a euploid karyotype, only 20%of clone 25 had 40 chromosomes (Fig. 4F). Further analysis ofclone 25 revealed that 76% of the cells were trisomic forchromosome 11 (Fig. 4G). In addition, we found chromosomalabnormalities in wild-type mGS cells following long-termculture, including trisomy 8 and 11. The double KO clone 131did not undergo any detectable translocations, additions or
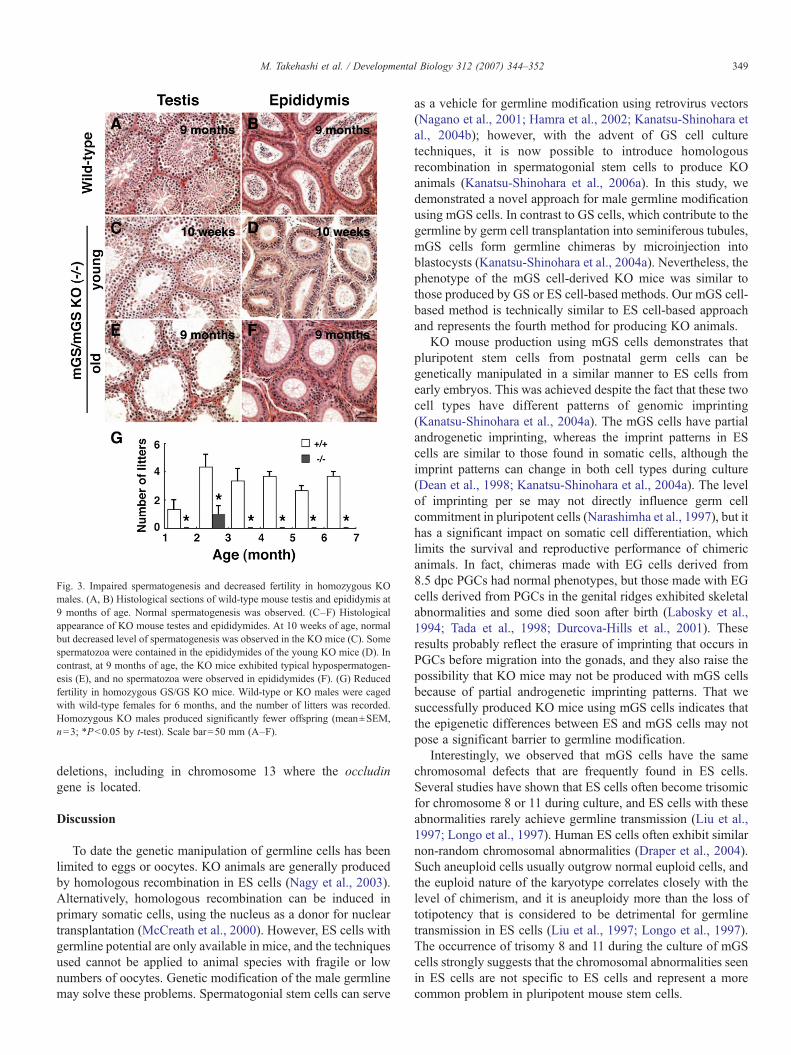
Fig. 3. Impaired spermatogenesis and decreased fertility in homozygous KOmales. (A, B) Histological sections of wild-type mouse testis and epididymis at9 months of age. Normal spermatogenesis was observed. (C–F) Histologicalappearance of KO mouse testes and epididymides. At 10 weeks of age, normalbut decreased level of spermatogenesis was observed in the KO mice (C). Somespermatozoa were contained in the epididymides of the young KO mice (D). Incontrast, at 9 months of age, the KO mice exhibited typical hypospermatogen-esis (E), and no spermatozoa were observed in epididymides (F). (G) Reducedfertility in homozygous GS/GS KO mice. Wild-type or KO males were cagedwith wild-type females for 6 months, and the number of litters was recorded.Homozygous KO males produced significantly fewer offspring (mean±SEM,n=3; ∗Pb0.05 by t-test). Scale bar=50 mm (A–F).
349M. Takehashi et al. / Developmental Biology 312 (2007) 344–352
deletions, including in chromosome 13 where the occludingene is located.
Discussion
To date the genetic manipulation of germline cells has beenlimited to eggs or oocytes. KO animals are generally producedby homologous recombination in ES cells (Nagy et al., 2003).Alternatively, homologous recombination can be induced inprimary somatic cells, using the nucleus as a donor for nucleartransplantation (McCreath et al., 2000). However, ES cells withgermline potential are only available in mice, and the techniquesused cannot be applied to animal species with fragile or lownumbers of oocytes. Genetic modification of the male germlinemay solve these problems. Spermatogonial stem cells can serve
as a vehicle for germline modification using retrovirus vectors(Nagano et al., 2001; Hamra et al., 2002; Kanatsu-Shinohara etal., 2004b); however, with the advent of GS cell culturetechniques, it is now possible to introduce homologousrecombination in spermatogonial stem cells to produce KOanimals (Kanatsu-Shinohara et al., 2006a). In this study, wedemonstrated a novel approach for male germline modificationusing mGS cells. In contrast to GS cells, which contribute to thegermline by germ cell transplantation into seminiferous tubules,mGS cells form germline chimeras by microinjection intoblastocysts (Kanatsu-Shinohara et al., 2004a). Nevertheless, thephenotype of the mGS cell-derived KO mice was similar tothose produced by GS or ES cell-based methods. Our mGS cell-based method is technically similar to ES cell-based approachand represents the fourth method for producing KO animals.
KO mouse production using mGS cells demonstrates thatpluripotent stem cells from postnatal germ cells can begenetically manipulated in a similar manner to ES cells fromearly embryos. This was achieved despite the fact that these twocell types have different patterns of genomic imprinting(Kanatsu-Shinohara et al., 2004a). The mGS cells have partialandrogenetic imprinting, whereas the imprint patterns in EScells are similar to those found in somatic cells, although theimprint patterns can change in both cell types during culture(Dean et al., 1998; Kanatsu-Shinohara et al., 2004a). The levelof imprinting per se may not directly influence germ cellcommitment in pluripotent cells (Narashimha et al., 1997), but ithas a significant impact on somatic cell differentiation, whichlimits the survival and reproductive performance of chimericanimals. In fact, chimeras made with EG cells derived from8.5 dpc PGCs had normal phenotypes, but those made with EGcells derived from PGCs in the genital ridges exhibited skeletalabnormalities and some died soon after birth (Labosky et al.,1994; Tada et al., 1998; Durcova-Hills et al., 2001). Theseresults probably reflect the erasure of imprinting that occurs inPGCs before migration into the gonads, and they also raise thepossibility that KO mice may not be produced with mGS cellsbecause of partial androgenetic imprinting patterns. That wesuccessfully produced KO mice using mGS cells indicates thatthe epigenetic differences between ES and mGS cells may notpose a significant barrier to germline modification.
Interestingly, we observed that mGS cells have the samechromosomal defects that are frequently found in ES cells.Several studies have shown that ES cells often become trisomicfor chromosome 8 or 11 during culture, and ES cells with theseabnormalities rarely achieve germline transmission (Liu et al.,1997; Longo et al., 1997). Human ES cells often exhibit similarnon-random chromosomal abnormalities (Draper et al., 2004).Such aneuploid cells usually outgrow normal euploid cells, andthe euploid nature of the karyotype correlates closely with thelevel of chimerism, and it is aneuploidy more than the loss oftotipotency that is considered to be detrimental for germlinetransmission in ES cells (Liu et al., 1997; Longo et al., 1997).The occurrence of trisomy 8 and 11 during the culture of mGScells strongly suggests that the chromosomal abnormalities seenin ES cells are not specific to ES cells and represent a morecommon problem in pluripotent mouse stem cells.

Fig. 4. Derivation of double KO mGS cells. A heterozygous KO clone (clone118) was used for double KO cell production. (A) Southern blot analysis ofDNA from the clones that survived G418 treatment. Genomic DNA wasdigested with the indicated restriction enzymes and hybridized with threedifferent probes. The bands expected from the endogenous gene and resultingfrom homologous recombination are indicated in Fig. 1A. (B, C) The lack ofoccludin mRNA (B) and protein (C) in the double KO embryoid bodies, asdetermined by RT-PCR and Western blotting, respectively. (D) Levels of G418resistance in wild-type, heterozygous and homozygous mGS cells. Equalnumbers of wild-type, heterozygous KO and double KO mGS cells were platedon G418-resistant MEFs and cultured with 20 mg/ml G418. The number ofsurviving colonies was determined after 14 days of culture (mean±SEM, n=16;Pb0.01 by t-test). (E) Histological section of a teratoma that developed from adouble KO mGS cells, showing tissues of three germ layers. Muscle (m), neural(n) and epithelial (e) tissues were found. (F) Karyotype analysis of the culturedcells. At least 20 cells were identified. (G) Types of aneuploid cells. At least 20cells were counted. The chromosomal aberrations were confirmed by FISHanalysis. Scale bar=100 μm (E).
350 M. Takehashi et al. / Developmental Biology 312 (2007) 344–352
The results of our double KO cell experiments also highlightthe close similarities between ES and mGS cells. Homozygouscells have been produced from heterozygous cultured cells by avariety of means. In this study, we followed the scheme for theselection of double KO ES cells production, and we found that
those mGS cells that survived exposure to a high concentrationof G418 produced double KO cells. Although the mechanism ofdouble KO cell production in ES cells has long been unknown,a recent report suggests that it occurs as a result of localuniparental disomy or gene conversion between the targeted andnon-targeted loci (Lefebvre et al., 2001), and the samemechanism probably applies to mGS cells. The feasibility ofdouble KO cell production from mGS cells may have importantimplications in clinical application because these cells may beused to produce differentiated somatic cells for regenerativemedicine. In particular, because mGS cells can be derived frompostnatal testes, this method may resolve the ethical problemssurrounding ES cell-based regenerative medicine.
On the other hand, GS cells were significantly more sensitiveto G418 than mGS cells, and we were unable to produce anydouble KO GS cells using the same procedure. One possiblereason for the failure is inefficient selection of the GS cells(Kanatsu-Shinohara et al., 2005b), which may be improved byoptimizing our growth conditions. However, given the fact thatGS cells maintain normal karyotypes after long-term culture(Kanatsu-Shinohara et al., 2005a), we speculate that theconfiguration and/or stability of the chromosomes are sig-nificantly different between pluripotent stem cells and tissue-specific stem cells, and that GS cells have greater chromosomalstability, thereby preventing the spontaneous loss of hetero-zygosity observed in pluripotent ES/mGS cells. Furthercharacterization of GS cells will be required to examine thispossibility.
Despite the close similarities between our approach and theES cell-based method, improvements must be made to increasethe usefulness of our technique. A major obstacle in thegermline modification of mGS cells was chimera production. Itis known that 129 ES cells contribute more to the somatic andgermline tissues in the chimeras than the C57BL/6 component(Papaioannou and Behringer, 2005). In this study, we micro-injected DBA/2 mGS cells into C57BL/6×DBA/2 blastocysts,and one of the seven animals analyzed was identified a germlinechimera. The donor cell contribution to coat color was low inthis animal; however, this was successfully overcome bymicroinsemination in our experiments. Finding more suitablehost will likely improve germline transmission. Anotherpotentially important parameter is the stability of the germlinepotential in vitro. Different levels of germline transmission areachieved using different ES cell lines (Nagy et al., 2003;Papaioannou and Behringer, 2005). While euploidy is onefactor that influences the germline stability, the cultureconditions of the ES cells also influences the maintenance ofgermline potential; serum-free medium generally producesbetter outcomes (Goldsborough et al., 1998). Additional studieswill provide important information to improve this technology.
A clear advantage of the current technique for KO mouseproduction is the use of different genetic backgrounds. WhileES cells are derived almost exclusively from preimplantationembryos in a 129 background, mGS cells as well as EG cellscan be derived not only from cells at later stages ofdevelopment, but they are also available in several backgrounds(Matsui et al., 1992; Resnick et al., 1992; Kanatsu-Shinohara et

351M. Takehashi et al. / Developmental Biology 312 (2007) 344–352
al., 2004a). Although improvements in ES culture conditionshave led to the derivation of ES cells in several other inbredbackgrounds (Schoonjans et al., 2003), only limited number ofgene targeting studies have used these cells, suggesting that 129has a more stable germline potential. The difference in germlinestability may simply reflect genetic differences, but it is alsopossible that the mechanism or origin of pluripotent cellderivation may significantly impact the stability of its germlinepotential. Indeed, it is possible that mGS cells achieve highergermline transmission rates than ES cells because of their germcell origin. Whatever the mechanism involved, the use ofdifferent genetic backgrounds is advantageous in mousefunctional genomics because otherwise substantial backcross-ing is necessary to obtain mutant animals with inbred geneticbackgrounds that can be used to study gene functions (Seonget al., 2004).
An important question that must be addressed in the future iswhether this technique can be extended to other animal species.ES-like cells from testes have been reported not only in mice butalso in other species, including humans (Cyranoski, 2006; Junget al., 2007). However, considering the close similarity betweenES and mGS cells and the fact that only mouse ES cells havegermline potential, the technology described here may not beapplicable to animals other than mice. Nevertheless, the resultsof a recent study suggest that spermatogonial stem cells havemore flexible differentiation potential than ES cells. Pluripotentcells from adult spermatogonia (i. e. multipotent adult germlinestem cells) achieved germline transmission following micro-injection either into the seminiferous tubules or the blastocystcavity (Guan et al., 2006). This result was unexpected becauseprevious attempts to demonstrate germline transmission byinjection into seminiferous tubules failed with ES and mGScells (Brinster and Avarbock, 1994; Kanatsu-Shinohara et al.,2004a). Therefore, a spermatogonia-based approach mayresolve critical problems with ES cell technology. Moreextensive studies are clearly required to determine whethermGS cells can also undergo germline transmission by germ celltransplantation and whether the pluripotent cells from sperma-togonia are a better vehicle for germline modification than EScells.
Genetic modification of spermatogonial stem cells providesa new opportunity for germline manipulation. Spermatogonialstem cells can be cultured in the absence of serum or feeder cells(Kanatsu-Shinohara et al., 2005c, 2006b), and they produceoffspring following xenogeneic transplantation into smallersurrogate animals (Shinohara et al., 2006). The current studydemonstrates the feasibility of KO mouse production usingmGS cells. Hopefully, this technique will be useful for thegermline modification in animal species, in which geneticmanipulation is very difficult or impossible.
Acknowledgments
We are grateful to Ms. A. Wada for technical assistance. Wealso thank Dr. M. Saitou for helpful discussions. Financialsupport for this research was provided by the Ministry ofEducation, Culture, Sports, Science, and Technology of Japan,
and by grants from CREST and the Human Science Foundation(Japanese). This work was also supported by the Program forPromotion of Fundamental Studies in Health Sciences of theNational Institute of Biomedical Innovation (NIBIO).
References
Brinster, R.L., Avarbock, M.R., 1994. Germ line transmission of donorhaplotype following spermatogonial transplantation. Proc. Natl. Acad. Sci.U. S. A. 91, 11303–11307.
Cyranoski, D., 2006. Stem cells from testes: could it work? Nature 440,586–587.
de Rooij, D.G., Russell, L.D., 2000. All you wanted to know aboutspermatogonia but were afraid to ask. J. Androl. 21, 776–798.
Dean, W., Bowden, L., Aitchison, A., Klose, J., Moore, T., Menesses, J.J., Reik,W., Feil, R., 1998. Altered imprinted gene methylation and expression incompletely ES cell-derived mouse fetuses: association with aberrantphenotypes. Development 125, 2273–2282.
Draper, J.S., Smith, K., Gokhale, P., Moore, H.D., Maltby, E., Johnson, J.,Meisner, L., Zwaka, T.P., Thomson, J.A., Andrews, P.W., 2004. Recurrentgain of chromosomes 17q and 12 in cultured human embryonic stem cells.Nat. Biotechnol. 22, 53–54.
Durcova-Hills, G., Ainscough, J., McLaren, A., 2001. Pluripotential stem cellsderived from migrating primordial germ cells. Differentiation 68, 220–226.
Evans, M.J., Kaufman, M.H., 1981. Establishment in culture of pluripotentialcells from mouse embryos. Nature 292, 154–156.
Goldsborough, M.D., Tilkins, M.L., Price, P.J., Lobo-Alfonso, J., Morrison,J.R., Stevens,M.E., 1998. Serum-free culture ofmurine embryonic stem (ES)cells. Focus 20, 8–12.
Guan, K., Nayernia, K., Maier, L.S., Wagner, S., Dressel, R., Lee, J.H., Nolte, J.,Wolf, F., Li, M., Engel, W., Hasenfuss, G., 2006. Pluripotency ofspermatogonial stem cells from adult mouse testis. Nature 440, 1199–1203.
Hamra, F.K., Gatlin, J., Chapman, K.M., Grellhes, I.D.M., Garcia, J.V.,Hammer, R.E., Garbers, D.L., 2002. Production of transgenic rats bylentiviral transduction of male germ-line stem cells. Proc. Natl. Acad. Sci.U. S. A. 99, 14931–14936.
Jung, J.G., Lee, Y.M., Park, T.S., Park, S.H., Lim, J.M., Han, J.Y., 2007.Identification, culture, and characterization of germline stem cell-like cells inchicken testes. Biol. Reprod. 76, 173–182.
Kanatsu-Shinohara, M., Shinohara, T., 2006. The germ of pluripotency. Nat.Biotechnol. 24, 663–664.
Kanatsu-Shinohara, M., Ogonuki, N., Inoue, K., Miki, H., Ogura, A.,Toyokuni, S., Shinohara, T., 2003. Long-term proliferation in culture andgermline transmission of mouse male germline stem cells. Biol. Reprod.69, 612–616.
Kanatsu-Shinohara, M., Inoue, K., Lee, J., Yoshimoto, M., Ogonuki, N., Miki,H., Baba, S., Kato, T., Kazuki, Y., Toyokuni, S., Toyoshima, M., Niwa, O.,Oshimura, M., Heike, T., Nakahata, T., Ishino, F., Ogura, A., Shinohara, T.,2004a. Generation of pluripotent stem cells from neonatal mouse testis. Cell119, 1001–1012.
Kanatsu-Shinohara, M., Toyokuni, S., Shinohara, T., 2004b. Transgenic miceproduced by retroviral transduction of male germ line stem cells in vivo.Biol. Reprod. 71, 1202–1207.
Kanatsu-Shinohara, M., Ogonuki, N., Iwano, T., Lee, J., Kazuki, Y., Inoue, K.,Miki, H., Takehashi, M., Toyokuni, S., Shinkai, Y., Oshimura, M., Ishino, F.,Ogura, A., Shinohara, T., 2005a. Genetic and epigenetic properties of mousemale germline stem cells during long-term culture. Development 132,4155–4163.
Kanatsu-Shinohara, M., Toyokuni, S., Shinohara, T., 2005b. Genetic selectionof mouse male germline stem cells in vitro: offspring from single stem cells.Biol. Reprod. 72, 236–240.
Kanatsu-Shinohara, M., Miki, H., Inoue, K., Ogonuki, N., Toyokuni, S.,Ogura, A., Shinohara, T., 2005c. Long-term culture of mouse malegermline stem cells under serum- or feeder-free conditions. Biol. Reprod.72, 985–991.
Kanatsu-Shinohara, M., Ikawa, M., Takehashi, M., Ogonuki, N., Miki, H.,Inoue, K., Kazuki, Y., Lee, J., Toyokuni, S., Oshimura, M., Ogura, A.,

352 M. Takehashi et al. / Developmental Biology 312 (2007) 344–352
Shinohara, T., 2006a. Production of knockout mice by random or targetedmutagenesis in spermatogonial stem cells. Proc. Natl. Acad. Sci. U. S. A.103, 8018–8023.
Kanatsu-Shinohara, M., Inoue, K., Lee, J., Miki, H., Ogonuki, N., Toyokuni, S.,Ogura, A., Shinohara, T., 2006b. Anchorage-independent growth of mousemale germline stem cells in vitro. Biol. Reprod. 74, 522–529.
Kimura, Y., Yanagimachi, R., 1995. Mouse oocytes injected with testicularspermatozoa or round spermatids can develop into normal offspring.Development 121, 2397–2405.
Labosky, P.A., Barlow, D.P., Hogan, B.L.M., 1994. Mouse embryonic germ(EG) cell lines: transmission through the germline and differences in themethylation imprint of insulin-like growth factor 2 receptor (Igf2r) genecompared with embryonic stem (ES) cell lines. Development 120,3197–3204.
Lefebvre, L., Dionne, N., Karaskova, J., Squire, J.A., Nagy, A., 2001. Selectionfor transgene homozygosity in embryonic stem cells results in extensive lossof heterozygosity. Nat. Genet. 27, 257–258.
Liu, X., Wu, H., Loring, J., Hormuzdi, S., Disteche, C.M., Bornstein, P.,Jaenisch, R., 1997. Trisomy eight in ES cells is a common potential problemin gene targeting and interferes with germ line transmission. Dev. Dyn. 209,85–91.
Longo, L., Bygrave, A., Grosveld, F.G., Pandolfi, P.P., 1997. The chromosomemake-up of mouse embryonic stem cells is predictive of somatic and germcell chimaerism. Transgenic Res. 6, 321–328.
Martin, G.R., 1981. Isolation of a pluripotent cell line from early mouse embryoscultured in medium conditioned by teratocarcinoma stem cells. Proc. Natl.Acad. Sci. U. S. A. 78, 7634–7638.
Matsui, Y., Zsebo, K., Hogan, B.L.M., 1992. Derivation of pluripotentialembryonic stem cells from murine primordial germ cells in culture. Cell 70,841–847.
McCreath, K.J., Howcroft, J., Campbell, K.H.S., Colman, A., Schnieke, A.E.,Kind, A.J., 2000. Production of gene-targeted sheep by nuclear transfer fromcultured somatic cells. Nature 405, 1066–1069.
McLaren, A., 2003. Primordial germ cells in the mouse. Dev. Biol. 262, 1–15.Meistrich, M.L., van Beek, M.E.A.B., 1993. Spermatogonial stem cells. In:
Desjardins, C., Ewing, L.L. (Eds.), Cell and Molecular Biology of the Testis.Oxford University Press, New York, pp. 266–295.
Mortensen, R.M., Conner, D.A., Chao, S., Geisterfer-Lowrance, A.A.T.,Seidman, J.G., 1992. Production of homozygous mutant ES cells with asingle targeting construct. Mol. Cell. Biol. 12, 2391–2395.
Nagano, M., Brinster, C.J., Orwig, K.E., Ryu, B.-Y., Avarbock, M.R.,Brinster, R.L., 2001. Transgenic mice produced by retroviral transductionof male germ-line stem cells. Proc. Natl. Acad. Sci. U. S. A. 98,13090–13095.
Nagy, A., Gertsenstein, M., Vintersten, K., Behringer, R., 2003. Introduction of
foreign DNA into embryonic stem cells, In: Nagy, A., Gertsenstein, M.,Vintersten, K., Behringer, R. (Eds.), Manipulating the Mouse Embryo, 3rded. Cold Spring Harbor Laboratory Press, New York, pp. 431–451.
Narashimha, M., Barton, S.C., Surani, M.A., 1997. The role of the paternalgenome in the development of the mouse germ line. Curr. Biol. 7, 881–884.
Palermo, G., Joris, H., Devroey, P., Van Steirteghem, A.C., 1992. Pregnanciesafter intracytoplasmic injection of single spermatozoon into an oocyte.Lancet 340, 17–18.
Papaioannou, V.E., Behringer, R.R., 2005. Getting to a phenotype. In:Papaionannou, V.E., Behringer, R.R. (Eds.), Mouse Phenotypes: AHandbook of Mutation Analysis. Cold Spring Harbor Laboratory Press,New York, pp. 29–47.
Park, T.S., Han, J.Y., 2000. Derivation and characterization of pluripotentembryonic germ cells in chicken. Mol. Reprod. Dev. 56, 475–482.
Resnick, J.L., Bixler, L.S., Cheng, L., Donovan, P.J., 1992. Long-termproliferation of mouse primordial germ cells in culture. Nature 359,550–551.
Saitou, M., Fujimoto, K., Doi, Y., Itoh, M., Fujimoto, T., Furuse, M., Takano, H.,Noda, T., Tsukita, S., 1998. Occludin-deficient embryonic stem cells candifferentiate into polarized epithelial cells bearing tight junctions. J. Cell.Biol. 141, 397–408.
Saitou, M., Furuse, M., Sasaki, H., Schulzke, J.D., Fromm, M., Takano, H.,Noda, T., Tsukita, S., 2000. Complex phenotype of mice lacking occludin, acomponent of tight junction strands. Mol. Biol. Cell 11, 4131–4142.
Schoonjans, L., Kreemers, V., Danloy, S., Moreadith, R.W., Laroche, Y., Collen,D., 2003. Improved generation of germline-competent embryonic stem celllines from inbred mouse strains. Stem Cells 21, 90–97.
Seong, E., Saunders, T.L., Stewart, C.L., Burmeister, M., 2004. To knockout in129 or in C57BL/6: that is the question. Trends Genet. 20, 59–62.
Shamblott, M.J., Axelman, J., Wang, S., Bugg, E.M., Littlefield, J.W., Donovan,P.J., Blumenthal, P.D., Huggins, G.R., Gearhart, J.D., 1998. Derivation ofpluripotent stem cells from cultured human primordial germ cells. Proc.Natl. Acad. Sci. U. S. A. 95, 13726–13731.
Shim, H., Gutierrez-Adan, A., Chen, L.R., BonDurant, R.H., Behboodi, E.,Anderson, G.B., 1997. Isolation of pluripotent stem cells from culturedporcine primordial germ cells. Biol. Reprod. 57, 1089–1095.
Shinohara, T., Kato, M., Takehashi, M., Lee, J., Chuma, S., Nakatsuji, N.,Kanatsu-Shinohara, M., Hirabayashi, M., 2006. Rats produced byinterspecies spermatogonial transplantation in mice and in vitromicroinsemination. Proc. Natl. Acad. Sci. U. S. A. 103, 13624–13628.
Stewart, C.L., Gadi, I., Bhatt, H., 1994. Stem cells from primordial germ cellscan reenter the germ line. Dev. Biol. 161, 626–628.
Tada, T., Tada, M., Hilton, K., Barton, S.C., Sado, T., Takagi, N., Surani, M.A.,1998. Epigenotype switching of imprintable loci in embryonic germ cells.Dev. Genes Evol. 207, 551–561.
Related Documents











