1998 4: 1069-1082 RNA M Sha, T Levy, P Kois, et al. splice site RNA oligonucleotides. Probing of the spliceosome with site-specifically derivatized 5' References http://rnajournal.cshlp.org/content/4/9/1069#related-urls Article cited in: service Email alerting click here top right corner of the article or Receive free email alerts when new articles cite this article - sign up in the box at the http://rnajournal.cshlp.org/subscriptions go to: RNA To subscribe to © 1998 RNA Society Cold Spring Harbor Laboratory Press on July 13, 2011 - Published by rnajournal.cshlp.org Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1998 4: 1069-1082RNA M Sha, T Levy, P Kois, et al. splice site RNA oligonucleotides.Probing of the spliceosome with site-specifically derivatized 5'
References http://rnajournal.cshlp.org/content/4/9/1069#related-urls
Article cited in:
serviceEmail alerting
click heretop right corner of the article orReceive free email alerts when new articles cite this article - sign up in the box at the
http://rnajournal.cshlp.org/subscriptions go to: RNATo subscribe to
© 1998 RNA Society
Cold Spring Harbor Laboratory Press on July 13, 2011 - Published by rnajournal.cshlp.orgDownloaded from

Probing of the spliceosome with site-specificallyderivatized 59 splice site RNA oligonucleotides
MA SHA, TAO LEVY, PAVOL KOIS, 1 and MARIA M. KONARSKAThe Rockefeller University, New York, New York 10021, USA
ABSTRACT
We have developed a site-specific chemical modification technique to incorporate a photoreactive azidophenacyl(APA) group at designated internal positions along the RNA phosphodiester backbone. Using this technique, we haveanalyzed interactions of the 59 splice site (59SS) RNA within the spliceosome. Several crosslinked products can bedetected within complex B using the derivatized 5 9SS RNAs, including U6 snRNA, hPrp8p, and 114-, 90-, 70-, 54-, and27-kDa proteins. The 5 9SS RNAs derivatized at intron positions +4 to +8 crosslink to U6 snRNA, confirming thepreviously reported pairing interaction between these sequences. hPrp8p and p70 are crosslinked to the 5 9SS RNAwhen the APA is placed within the 5 9 exon. Finally, a set of unidentified proteins, including p114, p54, and p27, isdetected with the 5 9SS RNA derivatized at intron positions +4 to +8. Introduction of the bulky APA group near the 5 9SSjunction (positions –2 to +3) strongly interferes with complex B formation and thus no APA crosslinks are observedat these positions. Together with our earlier observation that hPrp8p crosslinks to the GU dinucleotide at the 5 9 endof the intron, these results suggest that the inhibitory effect of APA results from steric hindrance of the hPrp8p:5 9SSinteraction. Unexpectedly, thio-modifications within the region of the 5 9SS RNA that is involved in base pairing toU6 snRNA strongly stimulate complex B formation.
Keywords: pre-mRNA splicing; spliceosome; 5 9 splice site; snRNP; UV crosslinking
INTRODUCTION
Splicing of nuclear pre-mRNAs takes place in a large,dynamic complex, the spliceosome (reviewed in Mooreet al+, 1993; Krämer, 1996)+ Its assembly involves aseries of intermediate complexes composed of smallnuclear ribonucleoprotein particles (snRNPs) and splic-ing factors that interact with pre-mRNA and each other+Several models of the catalytic center of the splice-osome have been proposed recently (Moore et al+, 1993;Nilsen, 1994; Staley & Guthrie, 1998)+ Branch site rec-ognition involves a pairing interaction with a segmentof U2 snRNA within complex A (Moore et al+, 1993)+ Atthe stage of complex B formation, U1 snRNP bound tothe 59SS is replaced by U4/U5/U6 snRNP, resulting inpairing of U6 snRNA with the intron sequences at the59SS (Sawa & Abelson, 1992; Sawa & Shimura, 1992;Wassarman & Steitz, 1992; Konforti et al+, 1993)+ Sub-sequently, the extensive pairing within U4/U6 snRNP isdisrupted and replaced by new helices between U2and U6 snRNAs (Moore et al+, 1993)+ Thus, interactionbetween U2 snRNA, paired to the branch site, with U6
snRNA, paired to the intron sequence near the 59SS, isthought to bring together and properly position the tworeactants for the first catalytic step+ The central loop ofU5 snRNA is thought to contact exon sequences (New-man & Norman, 1992; Sontheimer & Steitz, 1993; New-man et al+, 1995), although this interaction is not strictlynecessary because deletion of the U5 snRNA centralloop does not block splicing (O’Keefe et al+, 1996)+ Thisconclusion is consistent with the lack of sequence con-servation within exons+
Systematic analysis of mammalian factors based onaffinity purification of spliceosomes followed by 2D gelelectrophoresis has identified a number of spliceosome-associated polypeptides (SAPs, Gozani et al+, 1994)+This approach, combined with extensive biochemicalpurification and characterization of snRNPs (e+g+, Beh-rens & Lührmann, 1991; Fabrizio et al+, 1994, 1997;Lauber et al+, 1996, 1997), provides an important basisfor further analysis of spliceosomal proteins+ Finally,the recently identified SR family of proteins representsan important group of mammalian splicing factors that,by mediating both RNA–protein and protein–protein in-teractions, is thought to facilitate the selection of splicesites (reviewed in Fu, 1995)+ Several spliceosomal pro-teins are known to interact directly with pre-mRNA+Oneof these, the yeast U5 snRNP-specific protein Prp8
Reprint requests to: Maria M+ Konarska, The Rockefeller Univer-sity, 1230 York Avenue, New York, New York 10021, USA+
1Present address: Comenius University, Organic Chemistry De-partment, SK-84215 Bratislava, Slovakia+
RNA (1998), 4:1069–1082+ Cambridge University Press+ Printed in the USA+Copyright © 1998 RNA Society+
1069
Cold Spring Harbor Laboratory Press on July 13, 2011 - Published by rnajournal.cshlp.orgDownloaded from

and its mammalian homologue, hPrp8p, also termedp220 (Anderson et al+, 1989; Pinto & Steitz, 1989), canbe crosslinked to pre-mRNA at the 59SS, branch site,polypyrimidine tract, and the 39 SS region (MacMillanet al+, 1994; Teigelkamp et al+, 1995; Umen & Guthrie,1995, 1996)+ hPrp8p/Prp8p forms crosslinks with theintron lariat intermediate, indicating that this protein re-mains in close proximity to the splice sites at later stagesof the reaction (Sontheimer & Steitz, 1993; Teigelkampet al+, 1995; Umen & Guthrie, 1995)+ A number of othermammalian factors have been identified that directlycontact pre-mRNA at different stages of splicing+ Forexample, several proteins have been detected in prox-imity of the 59SS, 39SS, and the branch site by site-specific crosslinking (MacMillan et al+, 1994; Gauret al+, 1995; Singh et al+, 1995; Chiara et al+, 1996; Wu& Green, 1997)+
We previously developed a simplified in vitro system(Konforti & Konarska, 1995) in which a short 59SS RNAtogether with a longer 39SS RNA containing the branchsite, polypyrimidine tract, and the 39 splice site are usedas substrates in a bimolecular reaction+ The 59SS RNAinduces spliceosome formation and undergoes bothsteps of splicing when the 59 end of U1 snRNA is blockedor removed (Konforti & Konarska, 1994; Konforti et al+,1993)+ Two spliceosome-specific UV crosslinks in-volving the 59SS RNA are detected in this system: the59SS RNA:U6 snRNA and the 59SS RNA:hPrp8p (Reyeset al+, 1996)+ The former crosslink is consistent withthe pairing between intron positions 15 to 18 and hU6snRNA positions 39–42+ The latter crosslink productreflects the proximity of hPrp8p to the 59 exon andthe 59SS+ The hPrp8p crosslink maps to the GU at the59 end of the intron, the most highly conserved elementwithin the 59SS consensus+ This result suggests thatinteraction of Prp8 with the 59SS is highly specific, be-cause modifications of the uridine within the GU inter-fere with spliceosome formation and splicing (Reyeset al+, 1996)+ Using this system, we have now intro-duced a photoreactive azidophenacyl (APA) group atinternal positions along the 59SS RNA to probe thedistribution of spliceosome components in the prox-imity of the 59SS+ In addition to the expected 59SS RNAcrosslink to U6 snRNA, several protein crosslinks, in-cluding hPrp8p and a set of previously unidentifiedproteins of apparent molecular weights of 114 kDa,70 kDa, 54 kDa, and 27 kDa, were detected in thisanalysis+
RESULTS
Synthesis and characterization of thesite-specifically modified 5 9 splice site RNA
A simple two-step chemical modification strategy wasused to couple selected adducts at specific internalpositions within the RNA+ The strategy involves a post-
synthetic derivatization of RNA by coupling the halo-genated adduct to a thiol residue in the phosphodiesterbackbone+ Similar strategies for studies of RNA–proteinand RNA–RNA interactions were employed previouslyin analysis of tRNA:synthetase (Musier-Forsyth &Schimmel, 1994), tRNA:RNase P (Burgin & Pace, 1990;Harris et al+, 1994), and internal guide sequence:group Iintron (Wang et al+, 1993) associations+ We first testedthis technique by incorporating a photoreactive APAgroup (Fig+ 1) into the 59SS RNA, but a number of otheradducts can be used in this method (e+g+, Fig+ 1B,D)+First, the 59SS RNA that contains a thiol group at aselected position (Fig+ 1A) is prepared using standardprocedures of chemical synthesis+ Subsequently, cou-pling of the thiol with azidophenacyl bromide (APB,Hixson & Hixson, 1975; Burgin & Pace, 1990) is per-formed by nucleophilic displacement of bromine(Fig+ 1C)+ In the context of RNA, alkylation of the phos-phorothioate positioned adjacent to a 29OH group gen-erates a highly unstable triester, leading to site-specificdegradation of RNA (Gish & Eckstein, 1988)+ Thus, tostabilize the resulting triester, all RNAs used in this
FIGURE 1. Covalent attachment of adducts to thiol-RNAs+ A: Struc-ture of a three-nucleotide RNA fragment with a phosphorothioatelinkage adjacent to a 29-O-Me group (RNA-SH)+ B: Structure of ad-ducts used in this work: 1, azidophenacyl bromide (ABP); 2, phena-cyl bromide (PB); 3, iodoacetamide (IAA)+ C: Reaction of RNA-SHwith APB generating APA-RNA+ APA places the reactive nitrene ap-proximately 9 Å from the attachment site on phosphorothioate in theRNA+ D: Analysis of the derivatization products in a 20% polyacryl-amide/8 M urea gel+ Thio-modified G15 59SS RNA (lane 1) wasmock treated (lane 2) or derivatized with APB (lane 3), PB (lane 4),or IAA (lane 5)+ Unmodified, wt 59SS RNA (lane 6) was derivatizedwith APB (lane 7)+
1070 M. Sha et al.
Cold Spring Harbor Laboratory Press on July 13, 2011 - Published by rnajournal.cshlp.orgDownloaded from

study additionally contain a 29-O-methyl (29-O-Me) or a29-H adjacent to the thiol residue (Fig+ 1A, RNA-SH)+We have confirmed that both types of modifications atthe 29 position yield similar results (data not shown)+Typically, more than 80–90% of the input RNA yieldsthe derivatized product (Fig+ 1D)+ To test if the APAadduct is coupled specifically at the site of thio-modification, an unmodified, wild-type (wt) 59SS RNAwas subjected to derivatization with APB and the reac-tion products were resolved in a polyacrylamide gel+Although a slowly migrating APA-derivatized adductformed with thio-modified RNA (Fig+ 1D, lane 3), a prod-uct of intermediate gel mobility was observed when theunmodified 59SS RNA was used as substrate (Fig+ 1D,lane 7)+ Trace amounts of this product are also gener-ated with thio-RNA (Fig+ 1D, lane 3)+ Because forma-tion of this product does not require a thiol group, itmust reflect addition of the APA to other, less reactivegroups in the RNA that might be located on the bases+Depending on conditions of derivatization, this type ofthio-nonspecific product represents from 5 to 25% ofinput RNA, as monitored by phosphorimager scanningof denaturing gels (data not shown)+ Upon irradiation
with UV light (lmax at 302 nm) the azido group of APAis converted to a reactive nitrene, generating covalentcrosslinks to neighboring molecules+ The relatively smallsize of APA (;9 Å in length) makes it a useful tool forprobing a variety of nucleic acid complexes+ In thisstudy, APA-RNAs were used to analyze spliceosomalcomponents present in close proximity to the 59SS+
Crosslinking of the APA-derivatized59SS RNA in splicing reactions
For the initial series of experiments, the 59SS RNAcontaining a thiol group and derivatized with APA atposition G15 (G15-APA 59SS RNA, see Fig+ 2A) wasused+ The 59 end-labeled G15-APA 59SS RNA wasincubated in the presence of HeLa cell nuclear extractsunder splicing conditions (Konforti & Konarska, 1995),the reactions were irradiated with 302-nm UV light andthe crosslinked products were analyzed by SDS gelelectrophoresis+ Several abundant crosslinks were gen-erated under these conditions (Fig+ 2B, lanes 2–4)+Two of these products were identified as RNA–RNAcrosslinks, as judged by their resistance to protein-
FIGURE 2. APA-derivatized 59SS RNA crosslinks to protein and RNA via the APA group when incubated under splicingconditions and irradiated with 302-nm UV light+ A: Sequence of the 59SS RNA substrate used+ Numbering of positions isindicated+ The “/” indicates the exon/intron junction+ B: Splicing reactions containing the unmodified 59SS RNA (wt, lane 1),the 59SS RNA modified with thiol at position 15 (G15, lanes 5–7), or the APA-derivatized G15 thio-59SS RNA (lanes 2–4)were UV-irradiated as indicated+ C: Splicing reactions containing the thiol-G15 59SS RNA with (lanes 1–3) or without(lanes 4–6) APA derivatization were UV-irradiated as indicated+ D: Crosslinking profiles of the wt 59SS RNA (lane 1), theAPA-derivatized wt 59SS RNA (lane 2), and the APA-derivatized A16 thio-59SS RNA (lane 3)+ All 59SS RNAs were 59end-labeled and crosslinked products were resolved in a 12+5% (B, C) or 12% (D) SDS gel+ Positions of the 59SS RNAcrosslinks as well as protein size markers are indicated+
Spliceosome components in proximity of the 59 splice site 1071
Cold Spring Harbor Laboratory Press on July 13, 2011 - Published by rnajournal.cshlp.orgDownloaded from

ase K and sensitivity to RNase A digestion using asubstrate that lacks RNase A cleavage sites within the59SS RNA (data not shown)+ These RNA–RNA cross-links were further analyzed by RNase H digestion usingDNAs complementary to selected snRNA sequencesand identified as the 59SS RNA:U1 and 59SS RNA:U6 snRNA crosslinks (data not shown)+ In addition, sev-eral abundant RNA–protein crosslinks (p114, p70, andp54) and a number of less prominent products, includ-ing the hPrp8p crosslink, were also detected in thesereactions (Fig+ 2B, lanes 2–4)+ Although these cross-links are sensitive to digestion with either proteinase K,nuclease P1, or RNase T2, they do not contain anyRNA components other than the 59SS RNA, as indi-cated by their resistance to RNase A (using the appro-priately modified RNase A-resistant 59SS RNA, datanot shown)+ As expected, electrophoretic mobilities ofthese crosslinks change with the length of the attached59SS RNA+ As a control, a sample containing unmodi-fied, wt 59SS RNA was irradiated with 254-nm UV lightto generate two spliceosome-specific crosslinks, hPrp8pand U6 snRNA (Fig+ 2B, lane 1)+ No detectable cross-links were observed in the absence of UV irradiation(Fig+ 2C, lanes 1, 4)+ Only after 15 min irradiation at302-nm UV were traces of hPrp8p and U6 snRNA cross-links,which reflect APA-independent photoreactivity, de-tected in reactions with the underivatized G15 59SSRNA (Fig+ 2B, lane 7)+ Because irradiation at 302 nmfor 2 min and 15 min yielded similar patterns, the shorter2-min irradiation was selected as a standard treatmentfor subsequent experiments+
Crosslinking specificity in these reactions can be as-sessed by comparison of patterns obtained using theAPA-derivatized and underivatized (i+e+, containing thethiol and 29-O-methyl groups, but not the APA group)59SS RNAs (Fig+ 2C, lane 2 and 5)+ However, becausea detectable level of APA derivatization was observedeven with unmodified, wt 59SS RNA (Fig+ 1D, lane 7), itwas necessary to compare the crosslinking profiles ofRNAs derivatized in the thio-dependent and thio-independent fashions (Fig+ 2D)+As expected, reactionscontaining the wt 59SS RNA yielded no detectable cross-links upon irradiation with 302-nm light (Fig+ 2D,lane 1)+ However, a low level of crosslinks was ob-served with the wt 59SS RNA derivatized with APA(Fig+ 2D, lane 2)+ The crosslinking pattern representeda subset of products generated with the thio-59SS RNAAPA-derivatized at position A16 (A16-APA 59SS RNA,Fig+ 2D, lane 3) and included hPrp8p, p54, and U6snRNA crosslinks+ Thus, although a fraction of cross-links generated with thio-RNAs may originate from APAgroups attached through the bases, the majority of thesignal is derived from thio-specific APA derivatization+
A number of crosslinks detected in these reactionsmay represent RNA–protein interactions not related tosplicing+ To enrich for spliceosome-specific crosslinks,UV-irradiated reactions were centrifuged through glyc-
erol cushions (Fig+ 3A, see Materials and Methods)+Under these conditions, the majority of crosslinks notstably associated with spliceosomes remain in the su-pernatant (Fig+ 3A, lane 2), whereas those bound tosplicing complexes or snRNP particles are recoveredin the pellet (Fig+ 3A, lane 3)+ To further characterizecrosslinks generated upon UV irradiation of the G15-APA 59SS RNA, we analyzed the crosslinking profileunder various incubation conditions (Fig+ 3C)+ In par-allel, formation of complex B was monitored in nativegels (Fig+ 3B)+ As expected, both complex B formationand 59SS RNA crosslinking require the presence ofnuclear extract (Fig+ 3B,C, lane 3)+ The appearanceof the U6 snRNA crosslink correlates with spliceosomeformation because it can be clearly detected under stan-dard splicing conditions in the presence of two different39SS RNA sequences (adenovirus or rabbit b-globin39SS RNAs, Fig+ 3C, lanes 2, 6), but not in the absenceof ATP (Fig+ 3C, lane 4)+ Similarly, an abundant p54crosslink can be detected in these reactions (Fig+ 3C,lanes 2, 6), whereas its level decreases in the absenceof ATP (Fig+ 3C, lane 4)+ Interestingly, the p54 crosslinkis also detectable in reactions performed in the ab-sence of the 39SS RNA (Fig+ 3C, lane 5)+ Although theintensity of the p54 crosslink is significantly reducedcompared with analogous reactions containing the39SS RNA (Fig+ 3C, lane 2 or 6), this result correlateswell with the reduced stability of the 59SS RNA:U2/U4/U5/U6 snRNP complex that forms under these condi-tions (Konforti & Konarska, 1994)+ Furthermore, thep54 crosslink is not detectable in reactions in which U2,U4, or U6 snRNA is cleaved by RNase H in the pres-ence of complementary DNAs (Fig+ 3D, lane 2–4)+ In-stead, the p54 crosslink is easily detectable when the59 end of U1 snRNA is removed by RNase H, a treat-ment known to stimulate spliceosome formation andsplicing in this system (Fig+ 3D, lane 1)+ Thus, formationof the p54 crosslink correlates with requirements forassociation of the 59SS RNA with U2/U4/U5/U6 snRNPsin complex B+
Modification-interference analysisof splicing complex B
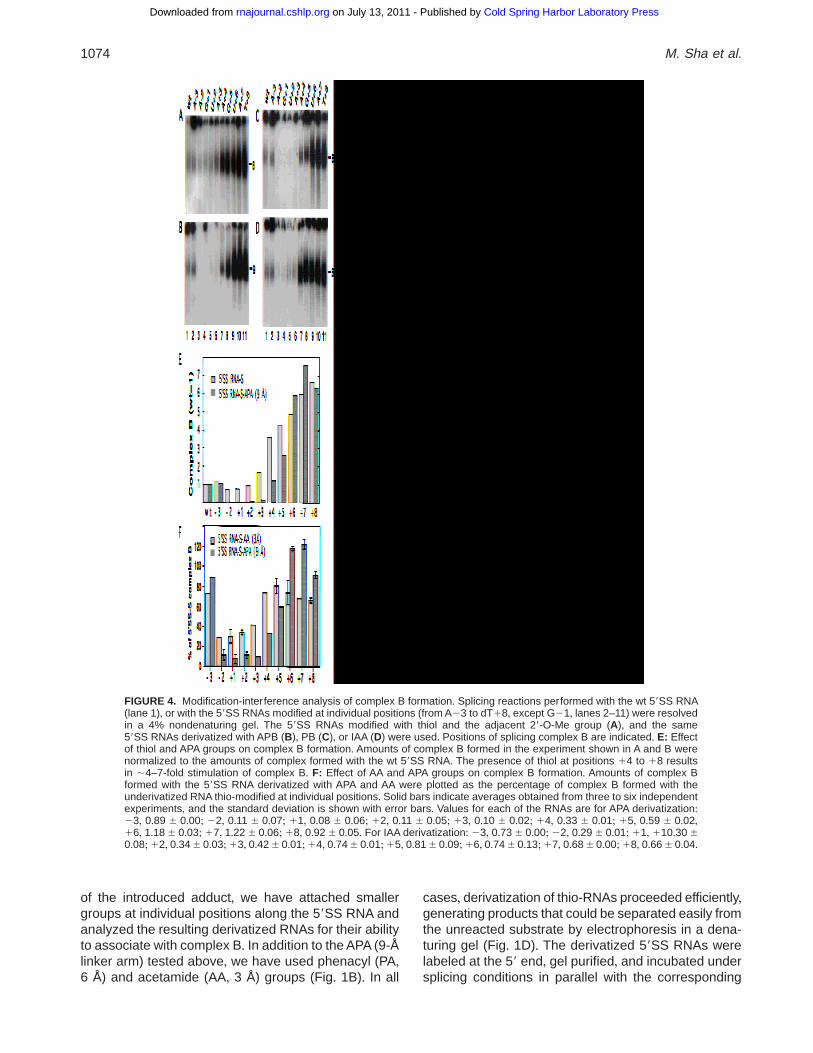
To study the possible interactions of spliceosome com-ponents with the 59SS, thio-modifications were intro-duced at individual positions along the entire 59SS RNA(Fig+ 4)+ As with the G15 59SS RNA, the 29 positionadjacent to the thiol in each of these RNAs was blockedby a 29-O-Me or a 29-H to ensure integrity of the APA-derivatized product+ The individual 59SS RNAs werelabeled at the 59 end with 32P and derivatized with APA+To test the APA-59SS RNAs for the ability to form splic-ing complexes, aliquots of each reaction were resolvedin native gels (Fig+ 4B)+ Although the 59SS RNAs de-rivatized at positions 22 to 13 appear to have lost theability to form complex B, analogous RNAs derivatized
1072 M. Sha et al.
Cold Spring Harbor Laboratory Press on July 13, 2011 - Published by rnajournal.cshlp.orgDownloaded from

at positions 14 to 18 associate with complex B withsignificantly increased efficiency (Fig+ 4B)+ Thus, to ad-dress the effect of the APA on spliceosome assembly,we have compared profiles of complex B formation usinga battery of thio-modified, but underivatized 59SS RNAswith the analogous substrates subjected to APA deriv-atization (Fig+ 4E)+
Interestingly, thio-modification alone, in the absenceof APA, strongly stimulated complex B formation whenintroduced at positions from A14 to T18 (Fig+ 4A,lanes 7–11)+ The stability of all thio-modified 59SS RNAstested (except for G-1 59SS RNA, see below), uponincubation in nuclear extracts under splicing condi-tions,was comparable to that of the unmodified wt 59SSRNA (data not shown)+ Furthermore, thio-modificationalone, regardless of the identity of the 29 position, gen-erated a similar stimulation of complex B formation,indicating that the thiol group itself is responsible forthe observed effect (Fig+ 5A)+ As expected, the strong
enhancement of complex B formed with thio-59SS RNAscorrelated with increased levels of hPrp8p (Fig+ 5B)and U6 snRNA (data not shown) crosslinks generatedby 254-nm UV irradiation+
Thus, to monitor the effect of the APA separately fromthe positive effect of thiol, the efficiency of spliceosomeassembly using the APA-59SS RNAs was normalizedto the level of complex B formed using the correspond-ing thio-modified, underivatized RNA (Fig+ 4F)+ Strongreduction in the levels of complex B formed with theAPA-59SS RNAs derivatized at positions near the 59splice junction (from A22 to A13) suggested that thebulky APA group may interfere with spliceosome as-sembly+ This interference appears to be site-specificbecause the same APA placed further away from the59SS, at positions 23 and 24 or from A14 to T18,does not inhibit complex B formation (Fig+ 4B, lanes 1,2, 7–11)+ To test the possibility that the observed inhi-bition of complex B assembly is correlated with the size
FIGURE 3. 54-kDa polypeptide crosslinks to the 59SS RNA under splicing conditions+ A: Splicing reactions containing the59 end-labeled G15-APA 59SS RNA were irradiated with 302-nm UV and spun through a 20% glycerol cushion+ Crosslinkproducts present in a complete reaction (lane 1), recovered from the supernatant (lane 2), or pellet (lane 3), were resolvedin a 12% SDS gel+ B: Reactions with the wt 59SS RNA (lane 1) were UV-irradiated at 254 nm for 15 min and those containingthe G15-APA 59SS RNA (lanes 2–6) were UV-irradiated at 302 nm for 2 min, and resolved in a 4% nondenaturing gel+Reactions were performed under standard splicing conditions (lanes 1, 2), in the absence of nuclear extract (lane 3), ATP(lane 4), 39SS RNA (lane 5), or in the presence of globin 39SS RNA (lane 6)+ Position of splicing complex B is indicated+ C:Aliquots of reactions shown in B were spun through a glycerol cushion and resolved in a 12+5% SDS gel+ D: Nuclear extractssupplemented with RNase H were pre-incubated in the absence (lane 5) or presence (KO: U1, U2, U4, U6, lanes 1–4,respectively) of DNAs complementary to the following regions of snRNAs: U1, 28–41; U2g, 29–42; U4b, 58–76; and U6f,33–48+ KO extracts and a standard extract without preincubation (NE, lane 6) were incubated in the presence of the59 end-labeled G15-APA 59SS RNA+ Reactions were UV-irradiated at 302 nm, spun through a glycerol cushion, andresolved in a 12+5% SDS gel+ Positions of the 59SS RNA crosslinks and of size markers are indicated+
Spliceosome components in proximity of the 59 splice site 1073
Cold Spring Harbor Laboratory Press on July 13, 2011 - Published by rnajournal.cshlp.orgDownloaded from
of the introduced adduct, we have attached smallergroups at individual positions along the 59SS RNA andanalyzed the resulting derivatized RNAs for their abilityto associate with complex B+ In addition to the APA (9-Ålinker arm) tested above, we have used phenacyl (PA,6 Å) and acetamide (AA, 3 Å) groups (Fig+ 1B)+ In all
cases, derivatization of thio-RNAs proceeded efficiently,generating products that could be separated easily fromthe unreacted substrate by electrophoresis in a dena-turing gel (Fig+ 1D)+ The derivatized 59SS RNAs werelabeled at the 59 end, gel purified, and incubated undersplicing conditions in parallel with the corresponding
FIGURE 4. Modification-interference analysis of complex B formation+ Splicing reactions performed with the wt 59SS RNA(lane 1), or with the 59SS RNAs modified at individual positions (from A23 to dT18, except G21, lanes 2–11) were resolvedin a 4% nondenaturing gel+ The 59SS RNAs modified with thiol and the adjacent 29-O-Me group (A), and the same59SS RNAs derivatized with APB (B), PB (C), or IAA (D) were used+ Positions of splicing complex B are indicated+ E: Effectof thiol and APA groups on complex B formation+ Amounts of complex B formed in the experiment shown in A and B werenormalized to the amounts of complex formed with the wt 59SS RNA+ The presence of thiol at positions 14 to 18 resultsin ;4–7-fold stimulation of complex B+ F: Effect of AA and APA groups on complex B formation+ Amounts of complex Bformed with the 59SS RNA derivatized with APA and AA were plotted as the percentage of complex B formed with theunderivatized RNA thio-modified at individual positions+ Solid bars indicate averages obtained from three to six independentexperiments, and the standard deviation is shown with error bars+ Values for each of the RNAs are for APA derivatization:23, 0+89 6 0+00; 22, 0+11 6 0+07; 11, 0+08 6 0+06; 12, 0+11 6 0+05; 13, 0+10 6 0+02; 14, 0+33 6 0+01; 15, 0+59 6 0+02,16, 1+18 6 0+03; 17, 1+22 6 0+06; 18, 0+92 6 0+05+ For IAA derivatization: 23, 0+73 6 0+00; 22, 0+29 6 0+01; 11, 110+30 60+08;12, 0+34 6 0+03;13, 0+42 6 0+01;14, 0+74 6 0+01;15, 0+81 6 0+09;16, 0+74 6 0+13;17, 0+68 6 0+00;18, 0+66 6 0+04+
1074 M. Sha et al.
Cold Spring Harbor Laboratory Press on July 13, 2011 - Published by rnajournal.cshlp.orgDownloaded from

thio-modified, underivatized 59SS RNAs, and the re-sulting complexes were resolved in a native gel+ Thepattern of complex B formation is similar for the APA-(9 Å) and PA- (6 Å) derivatized 59SS RNAs (Fig+ 4B,C)+Specifically, derivatization of phosphodiester bonds nearthe splice junction, between positions 22 and 13 ofthe 59SS RNA, resulted in a significant reduction ofspliceosome formation ($ 10-fold) compared with un-derivatized 59SS RNAs modified only with thiol groups+Furthermore, the PA group introduced at positions14 and 15 partially reduced spliceosome formation(;40–60% of the control thio-59SS RNA, respectively)+However, a smaller, 3-Å AA group resulted in a com-paratively smaller degree of inhibition of complex Bformation (30–40% of the thio-59SS RNA at positionsfrom 22 to 13)+ Thus, with the decreasing size of ad-duct, the inhibitory effect was less severe and ex-tended over a shorter region of the RNA, centeringaround the 59SS junction+
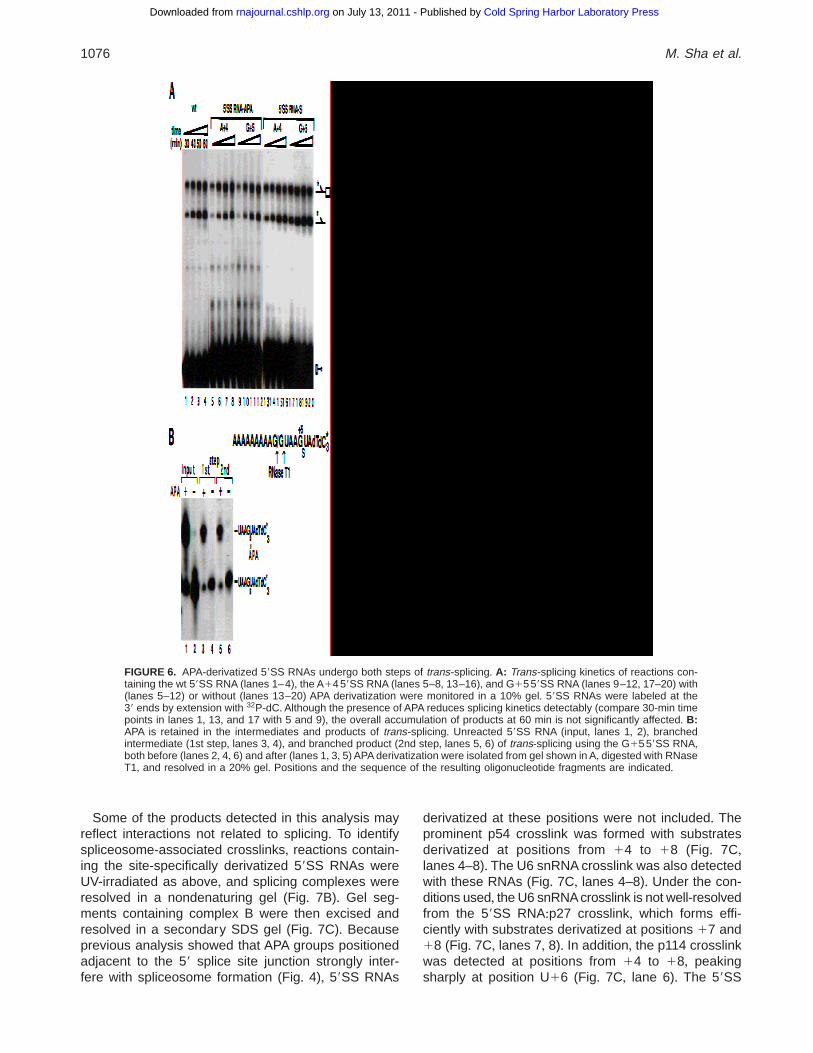
Independently, we have confirmed that derivatized59SS RNAs capable of complex B formation are alsospliced efficiently+ For these experiments, the 59SSRNAs containing a longer exon (A9G, resulting in ahigher efficiency of splicing) were labeled at the 39 endby the addition of three 32P-dC residues, allowing fordetection of branched intermediates and products oftrans-splicing (Fig+ 6A)+ Splicing kinetics using the wt(Fig+ 6A, lanes 1–4), thio-modified (lanes 13–20), and
APA-derivatized (lanes 5–12) 59SS RNAs are similar,indicating that splicing is not significantly affected bythe presence of APA at the positions tested+ To confirmthat APA is not removed from the RNA prior to splicing,branched intermediates and products generated withthe G15 59SS RNA were isolated along with the un-reacted 59SS RNA substrate and subjected to diges-tion with RNase T1 (Fig+ 6B)+ The labeled 11-nt intronfragment produced in this reaction spans the site ofmodification (the thio-G15 position is not cleaved withRNase T1 because of the presence of a 29OMe) andthus allows for separation of derivatized and underiv-atized products+ The APA group was detected in bothbranched intermediates and products, confirming thatits presence does not interfere with splicing and thusdoes not significantly alter the geometry of the assem-bled splicing complexes+
Modification interference at the splice junction (posi-tion G21) was analyzed in greater detail+ Complex Bformation was reduced (;50% of wt) in the presence of29H at this position, whereas the analogous modifica-tion at neighboring positions (23, 22, or from 11 to14) had no detectable effect (data not shown)+ A com-bination of a thiol together with the adjacent 29H groupcaused an additional inhibition of complex B formation(to ;35–40% of wt, data not shown)+ The thio-effect atposition G21 in the context of all-RNA 59SS could notbe measured reliably because of the unusual instabilityof such RNAs upon incubation in nuclear extracts (datanot shown)+ Thus, complex B formation appears to beparticularly sensitive to ribose modifications at positionG21, complicating the analysis of derivatization ef-fects+
Probing of the spliceosome withsite-specifically derivatized 5 9SS RNAs
To detect spliceosomal components positioned in prox-imity to the 59SS, the crosslinking patterns of splicingreactions containing site-specifically derivatized 59SSRNAs were examined (Fig+ 7A)+ The crosslinking pat-terns obtained with individual 59SS RNAs differed de-pending on the position of the APA in the substrate+ The59SS RNA:U6 snRNA crosslink was formed when theAPA was present at positions U16, A17, and T18 ofthe 59SS RNA, and could be detected with lower effi-ciency at positions G15 and A14 (Fig+ 7A, lanes 7–11,and data not shown)+Similarly, the 59SS RNA:p54 cross-link was formed efficiently using the 59SS RNAs deriv-atized at positions U16, A17, and T18, and was alsodetectable at positions G15 and A14 (Fig+ 7A, lanes 7–11)+However, these crosslinks were not detectable whenthe APA was placed closer to the splice junction, atpositions from A24 to A13 (Fig+ 7A, lanes 1–6)+ In-stead, 59SS RNA containing the APA within the exonsegment at position 23 or 24 produced an abundanthPrp8p crosslink (Fig+ 7A, lanes 1, 2)+
FIGURE 5. Phosphorothioate modification of the 59SS RNA in-creases accumulation of complex B+ Reactions were performed un-der splicing conditions using the 59SS RNA containing ribo (lanes 1,2), 29O-Me (lanes 3, 4), or deoxy (lanes 5, 6) residues at positionG15+ RNAs in lanes 2, 4, and 6 contained in addition a thiol atposition G15+ A: Complex B formation was monitored in a 4% non-denaturing gel+ B: Aliquots of reactions shown in A were UV-irradiated(254 nm), spun through a glycerol cushion, and crosslinks resolvedin a 10% SDS gel+ Positions of the hPrp8p:59SS RNA crosslink andsize markers are indicated+
Spliceosome components in proximity of the 59 splice site 1075
Cold Spring Harbor Laboratory Press on July 13, 2011 - Published by rnajournal.cshlp.orgDownloaded from
Some of the products detected in this analysis mayreflect interactions not related to splicing+ To identifyspliceosome-associated crosslinks, reactions contain-ing the site-specifically derivatized 59SS RNAs wereUV-irradiated as above, and splicing complexes wereresolved in a nondenaturing gel (Fig+ 7B)+ Gel seg-ments containing complex B were then excised andresolved in a secondary SDS gel (Fig+ 7C)+ Becauseprevious analysis showed that APA groups positionedadjacent to the 59 splice site junction strongly inter-fere with spliceosome formation (Fig+ 4), 59SS RNAs
derivatized at these positions were not included+ Theprominent p54 crosslink was formed with substratesderivatized at positions from 14 to 18 (Fig+ 7C,lanes 4–8)+ The U6 snRNA crosslink was also detectedwith these RNAs (Fig+ 7C, lanes 4–8)+ Under the con-ditions used, the U6 snRNA crosslink is not well-resolvedfrom the 59SS RNA:p27 crosslink, which forms effi-ciently with substrates derivatized at positions 17 and18 (Fig+ 7C, lanes 7, 8)+ In addition, the p114 crosslinkwas detected at positions from 14 to 18, peakingsharply at position U16 (Fig+ 7C, lane 6)+ The 59SS
FIGURE 6. APA-derivatized 59SS RNAs undergo both steps of trans-splicing+ A: Trans-splicing kinetics of reactions con-taining the wt 59SS RNA (lanes 1–4), the A14 59SS RNA (lanes 5–8, 13–16), and G15 59SS RNA (lanes 9–12, 17–20) with(lanes 5–12) or without (lanes 13–20) APA derivatization were monitored in a 10% gel+ 59SS RNAs were labeled at the39 ends by extension with 32P-dC+ Although the presence of APA reduces splicing kinetics detectably (compare 30-min timepoints in lanes 1, 13, and 17 with 5 and 9), the overall accumulation of products at 60 min is not significantly affected+ B:APA is retained in the intermediates and products of trans-splicing+ Unreacted 59SS RNA (input, lanes 1, 2), branchedintermediate (1st step, lanes 3, 4), and branched product (2nd step, lanes 5, 6) of trans-splicing using the G15 59SS RNA,both before (lanes 2, 4, 6) and after (lanes 1, 3, 5) APA derivatization were isolated from gel shown in A, digested with RNaseT1, and resolved in a 20% gel+ Positions and the sequence of the resulting oligonucleotide fragments are indicated+
1076 M. Sha et al.
Cold Spring Harbor Laboratory Press on July 13, 2011 - Published by rnajournal.cshlp.orgDownloaded from

RNA:p90 crosslink could be detected with substratesderivatized at positions 16 or 17 (Fig+ 7C, lanes 6, 7),however, formation of this product was not reproduc-ible+Complex B assembled with the A-3-APA 59SS RNAyielded a prominent 59SS RNA:hPrp8p crosslink andlower amounts of the 59SS:p70 crosslink (Fig+ 7C,lane 1)+ Significant levels of hPrp8p and p70 crosslinkswere also detected with 59SS RNAs derivatized at allother positions+ This could indicate that hPrp8p andp70 are positioned in the proximity of the 59SS RNAalong its entire length+ However, because a detectablelevel of thio-independent APA derivatization can alsooccur (Fig+ 1D, lane 7), a subset of the generated prod-ucts might originate from such thio-independent cross-linking+ To distinguish between these possibilities, weprepared two 59SS RNAs thio-modified at position 16,32P-labeled at either the 59 or the 39 end, and deriva-tized with APB+ Upon RNase A digestion, the 59 end-labeled RNA yields a 32P-fragment that consists of the59 exon and the intron GU dinucleotide, whereas the39 end-labeled RNA generates a 32P-labeled fragmentcontaining the remaining portion of the intron (positions13 to 18, in this substrate dT residues are present atpositions 16 and 18)+ Analysis of products generatedwith the 59 end-labeled RNA revealed that the p114and p54 crosslinks are eliminated by RNase A diges-tion (Fig+ 7D, lane 2), as expected for crosslinks at-tached to the RNA at position 16+ In addition, the U6
snRNA crosslink is also degraded by RNase A+ How-ever, a subset of crosslinks, including hPrp8p and p70,retain the 32P-label after RNase A treatment, suggest-ing that these proteins are attached to the 59SS RNAthrough its 59-terminal segment+ In contrast, among theproducts detected using the 39 end-labeled RNA, onlyp114, p54, and low levels of p27 were detected afterRNase A digestion (Fig+ 7D, lane 6), consistent with thenotion that these crosslinks are formed through theAPA at position 16+ Instead, hPrp8p and p70 cross-links present in the undigested reaction (Fig+ 7D,lane 5) are eliminated by RNase A treatment+ Thus,hPrp8p and p70 are crosslinked only to the 59-terminalportion of the 59SS RNA, whereas p114, p54, and p27crosslinks are formed at the site of thiol modification(position 16 in this case)+
Together, a number of proteins can be crosslinked tothe 59SS RNA within the splicing complex B+ Amongthese, we have been able to confirm the identity of the59SS RNA:hPrp8p crosslink+ hPrp8p crosslinks weregenerated using the A-3-APA and wt 59SS RNAs irradi-ated with 302-nm and 254-nm UV light, respectively,and gel purified+ Both crosslinks were specifically im-munoprecipitated using anti-hPrp8 antiserum (a gift fromDr+ M+ Moore, Fig+ 8, lanes 3, 5) but not pre-immuneserum (Fig+ 8, lanes 2, 4)+ The identity of other splice-osomal proteins crosslinked to the 59SS RNA via APAgroups remains to be determined+
FIGURE 7. Crosslinking profiles of the 59SS RNAs derivatized with APA at individual positions along the sequence incomplete splicing reactions (A) or in the isolated complex B (C)+ A: Splicing reaction containing 59 end-labeled wt 59SS RNAwas UV-irradiated (254 nm for 15 min, lane 12)+ In all other cases, splicing reactions using 59 end-labeled APA-derivatized59SS RNAs were UV-irradiated (302 nm for 2 min), spun through a glycerol cushion, and crosslinked products were resolvedin a 12+5% SDS gel+ B,C: Splicing reactions with 59 end-labeled 59SS RNAs derivatized with APA at the indicated positionswere resolved in a native gel (B)+ Gel segments containing complex B were excised and resolved in a second dimensionin a 15% SDS gel (C)+ D: A16 59SS RNA underivatized (lanes 3, 4) or APA-derivatized (lanes 1, 2 and 5, 6), labeled at the59 end (lanes 1, 2) or the 39 end (lanes 3–6) was incubated under splicing conditions and UV-irradiated as indicated+Resulting crosslinks were digested with RNase A (lanes 2, 4, 6) or mock treated (lanes 1, 3, 5) and resolved in a 12% SDSgel+ Positions of the 59SS RNA crosslinks as well as protein size markers are indicated+
Spliceosome components in proximity of the 59 splice site 1077
Cold Spring Harbor Laboratory Press on July 13, 2011 - Published by rnajournal.cshlp.orgDownloaded from

DISCUSSION
Because of the remarkable complexity of the splice-osome, a detailed structural analysis of this ribonucleo-protein complex is very difficult+ To identify spliceosomalcomponents present in close proximity to the splicesites, we have developed an efficient and versatilemethod for the introduction of photoreactive groups intothe RNA and have identified protein and snRNA com-ponents that interact with the 59 splice site+
Photocrosslinking technique
A large number of RNA–protein and RNA–RNA inter-actions have been studied using UV crosslinking, in-cluding systems as complex as the ribosome (Ofenganget al+, 1977) or the spliceosome (Sawa & Abelson, 1992;Sawa & Shimura, 1992; Wassarman & Steitz, 1992)+Specifically, crosslinking techniques identified a num-ber of proteins interacting with pre-mRNA sequences
important for splicing, including the polypyrimidine tract,branch site, and 59 or 39SS signals (e+g+, Sontheimer &Steitz, 1993; MacMillan et al+, 1994; Gaur et al+, 1995;Singh et al+, 1995; Chiara et al+, 1996; Query et al+,1996; Reyes et al+, 1996)+ To date, most crosslinkingapproaches concentrated on the analysis of interactionsinvolving the bases+ These approaches utilize the nat-ural photoreactivity of bases at 254 nm, photoreactivebase analogues (e+g+, 5-BrU, 5-IU, 4-thioU, 2-azidoA,8-azidoA), or photoreactive groups attached to basesthrough a longer linker (Sontheimer & Steitz, 1993;Wil-lis et al+, 1993; MacMillan et al+, 1994; Query et al+,1996)+ However, because of the high degree of con-servation of the pre-mRNA consensus sequences, onlya few selected positions in the RNA can be modifiedwithout affecting spliceosome assembly+ Thus, to bet-ter characterize the spliceosomal components involvedin recognition of the 59SS, we chose to introduce aphotoreactive group through the phosphodiester back-bone in the RNA+ Derivatization of DNA through phos-phorothioate groups has been used previously in anumber of systems (Cassetti & Moss, 1996; Lagrangeet al+, 1996)+ However, due to the high degree of insta-bility of the resulting phosphorotriester in the presenceof the adjacent 29 OH group, most applications of thistechnique in the context of RNA were restricted to de-rivatization of 59-terminal positions (Burgin & Pace, 1990;Wang et al+, 1993; Harris et al+, 1994)+ This problemwas circumvented by introducing a single deoxyribose59 to the benzophenone-derivatized phosphorothioate(Musier-Forsyth & Schimmel, 1994)+ In this work, thephotoreactive APA group was covalently attached atinternal positions of the 59SS RNA through a thiol groupintroduced during the synthesis of RNA+ To stabilize thephosphorotriester, the 29-OH adjacent to the deriva-tized thiol was replaced by a 29-O-Me or a 29-H in asite-specific fashion+ The APA positions the photoreac-tive nitrene ;9 Å from the thiol, a distance comparablein length to three nucleotides in the RNA chain+
We have applied this method to mapping the splice-osomal components present in close proximity to the59SS and thus expected to participate in formation ofthe catalytic site+ To facilitate the generation of largeamounts of splicing complexes, we have used a sim-plified in vitro system developed previously in our lab-oratory (Konforti & Konarska, 1995)+ In this system, ashort 59SS RNA can induce spliceosome assembly inthe presence of a longer 39SS RNA and undergo bothsteps of splicing+ By increasing the concentrations ofthe 59 and 39SS RNAs, it is possible to achieve veryefficient spliceosome assembly, such that the vast ma-jority of endogenous U4/U5/U6 snRNP can be en-gaged in complex B formation+ Furthermore, becausesubsequent rearrangements of this complex leading tocomplex C formation and splicing catalysis are slow,complex B accumulates in these reactions and resultsin a highly homogenous population of spliceosomes+
FIGURE 8. Immunoprecipitation of the 59SS RNA:hPrp8p crosslink+59SS RNA:hPrp8p crosslink formed either with the wt 59SS RNA inthe splicing reaction irradiated with 254 nm UV (lanes 1–3) or withthe A-3-APA 59SS UV-irradiated with 302 nm UV (lanes 4–5) was gelpurified and subjected to immunoprecipitation with anti-hPrp8p anti-body 70R (lanes 3, 5) or with pre-immune serum (lanes 2, 4)+ Mate-rial bound to the Protein A-Trisacryl resin was resolved in a 10%SDS gel+ Positions of the 59SS RNA:hPrp8p crosslink and of themolecular weight size markers are indicated+
1078 M. Sha et al.
Cold Spring Harbor Laboratory Press on July 13, 2011 - Published by rnajournal.cshlp.orgDownloaded from

The structure of complex B does not appear to be af-fected significantly by the presence of the bulky APAgroup in the 59SS RNA (at positions that allow for com-plex B formation) because derivatized 59SS RNAs ef-ficiently undergo both steps of splicing (Fig+ 6)+
Mapping of the 5 9SS RNA contactswithin splicing complex B
Several spliceosomal components were detected in theproximity of the 59SS using this system, includinghPrp8p, U6 snRNA, and a set of previously unidentifiedproteins, p114, p70, p54, and p27 (Fig+ 9)+ The mostabundant of these products, p54, becomes crosslinkedto the 59SS RNA when the APA is introduced at intronpositions 14 to 18+ The crosslinking signal dependson the site of APA attachment, suggesting that p54 islocated near the intron segment involved in the pairinginteraction with U6 snRNA+ In addition, several lessabundant crosslinks are detected in these reactions+The p27 crosslink is detected through the APA locatedat the 39 end of the 59SS RNA (positions 17 and 18),whereas the p114 crosslink is detected predominantlyat position 16+ Furthermore, the p90 crosslink can bedetected at position 16 or 17, however, its appear-ance is somewhat variable+ Finally, two crosslinks,hPrp8p and p70, are detected with the 59SS RNAsderivatized in the exon segment+ Interestingly, althoughboth these crosslinks can also be detected with RNAsthio-modified at various intron positions (e+g+, Fig+ 7A,lanes 7–11, Fig+ 7C, lanes 3–8), they represent prod-ucts linked to the 59SS RNA through APA groups located
only in the 59-exon segment (Fig+ 7D)+ Short-wavelengthUV irradiation of splicing reactions containing wt, un-modified 59SS RNA induces formation of a specific,direct crosslink between hPrp8p and the GU dinucle-otide at the 59 end of the intron (Reyes et al+, 1996)+The APA groups positioned near the 59SS strongly in-terfere with spliceosome assembly (Fig+ 4), whereastheir presence in the exon just a few nucleotides up-stream allow a crosslink to hPrp8+ Together, these re-sults suggest that although the direct, close interactionbetween hPrp8p and the 59SS RNA is restricted to afew nucleotides around the 59SS junction itself, the prox-imity of hPrp8 extends over the exon segment and theexon/intron junction+
Formation of the described crosslinks with the 59SSRNA takes place in complex B and requires conditionsthat are necessary for splicing, e+g+, the presence ofATP, intact U2, U4, and U6 snRNAs+ However, closecontact between these proteins and the 59SS may betransient+ Chase experiments indicate that the pre-formed 59SS RNA:p54 crosslink does not participate insplicing (data not shown) and suggest that the 59SSRNA:p54 interaction is not maintained at later stages inthe reaction+ Further analysis will be necessary to de-termine the identity of p114, p70, p54, and p27 andtheir role in spliceosome assembly+ There are severalcandidate factors among the known spliceosome com-ponents in a size range comparable to the most abun-dant 59SS RNA:p54 crosslink+ These include 62-, 61-,57-, 55-, and 49-kDa SAPs (spliceosome-associatedproteins), which are implicated in complex A formationbut are also present at later stages of spliceosomeassembly (Chiara et al+, 1996)+ Furthermore, a 54-kDasplicing factor (Chaudhary et al+, 1991) has been foundto interact with U2AF65, and thus to influence complexA formation (Zhang & Wu, 1996)+ In addition, anotherrecently identified protein associated with U2AF65,UAP56, is required for the branch site–U2 snRNP in-teraction (Fleckner et al+, 1997)+ Finally, several U4/U6and U4/U5/U6 snRNP-specific proteins of this sizerange have been identified+ These include a 60-kDaU4/U6 snRNP component recently identified as a ho-mologue of yeast Prp4 (Lauber et al+, 1997), and threeother proteins associated with U4/U5/U6 snRNP (Beh-rens & Lührmann, 1991)+ Thus far, immunoprecipitationanalysis using antibodies against a variety of the fac-tors listed above failed to identify the p54 crosslinkdetected in this work (data not shown)+ Similarly, anti-bodies against the 116-kDa protein of U5 snRNP (Fa-brizio et al+, 1997) failed to recognize the U16-APA59SS RNA:p114 crosslink (data not shown)+ It is pos-sible that proteins detected in this study correspond toother known components of snRNP particles or otherknown splicing factors for which antibodies are not yetavailable+ Alternatively, they may represent factors thathave not been identified previously in the mammaliansystem+
FIGURE 9. Schematic representation of the spliceosomal proteinscrosslinked to the 59SS RNA in the context of snRNA–snRNA inter-actions present at the stage of complex B+ Sizes and relative posi-tions of the crosslinked proteins are arbitrary+
Spliceosome components in proximity of the 59 splice site 1079
Cold Spring Harbor Laboratory Press on July 13, 2011 - Published by rnajournal.cshlp.orgDownloaded from

59SS RNA modifications within the59SS:U6 snRNA interaction domain
The 59SS RNA intron segment implicated in contactingp54 and p114 (positions 14 to 18) has also been shownto interact with U6 snRNA+ This 59SS RNA:U6 snRNAinteraction is formed already upon the joining of U4/U5/U6 snRNP into the spliceosome (Konforti & Konar-ska, 1994)+ The 59SS:U6 snRNA pairing appears to becritical for proper positioning of the 59SS for catalysisand, in fact, stability of this interaction can be rate-limiting for splicing (Crispino & Sharp, 1995)+ Similarly,stability of the 59SS:U6 snRNA duplex affects forma-tion of complex B in our system (data not shown)+ In-terestingly, formation of complex B is also stronglyaffected by the presence of thiol in the 59SS RNA+ Thispositive thio-effect is regio-specific and coincides withthe positions of 59SS:U6 snRNA base pairing+ Becausethio-modification introduces a number of changes inthe RNA, a definitive interpretation of its effect is diffi-cult+ The sulfur atom is slightly bigger than oxygen andpossibly differently solvated+Phosphorothioates are alsoslightly more acidic, with a negative charge localizedon sulfur (Frey & Sammons, 1985)+ Finally, the coordi-nation of metal ions changes with sulfur substitution+Although Mg21 coordinates preferentially to oxygen,Mn21 has a higher affinity for sulfur in a phosphoro-thioate+ Both positive and negative thio-effects havebeen observed in several other protein–nucleic acidsinteractions (e+g+, Milligan & Uhlenbeck, 1989; Jeltschet al+, 1995)+ The most detailed analysis of thio-effectson RNA–protein interaction was reported for phage MS2(R17) coat protein binding to its operator RNA (Milligan& Uhlenbeck, 1989; LeCuyer et al+, 1996)+ Further stud-ies will be necessary to understand the mechanism ofthe thio-effect in our system+ It is unlikely that this effectreflects a direct, specific interaction between the splice-osome and the 59SS ribose backbone at a unique phos-phodiester bond, because the observed effect is notstereospecifc (data not shown)+ Rather, the presenceof sulfur in this segment may permit more favorablehydrogen bonding interactions with one of the splice-osome components than does the oxygen in the wt59SS RNA+
Effect of modifications near the 5 9SS junction
Analysis of derivatized 59SS RNAs clearly indicatesthat the introduction of a large APA group in close prox-imity to the 59SS strongly interferes with complex Bformation+ Furthermore, the size of the adduct intro-duced into the 59SS correlates with the distance of itsposition from the 59SS junction at which it has an effect+For the APA group (;9 Å), strong interference of com-plex B formation (to ;10% of wt) is observed fromposition 22 to 13, whereas a lesser effect is detect-able even at positions 14 and 15 (Fig+ 4)+ The smaller
AA group (;3 Å) interferes with complex B formation atpositions from 22 to 12 and the observed inhibition isless severe (to ;40% of wt)+ The observed interfer-ence is apparently associated with complex B forma-tion and does not affect the interaction between the59SS RNA and U1 or U4/U5/U6 snRNP (data notshown)+ Thus, the site-specific derivatization of 59SSRNA allows for a precise modification-interference anal-ysis of the spliceosome+ This type of approach hasbeen used extensively in studies of protein–RNA inter-actions in general (Conway & Wickens, 1989; Krol &Carbon, 1989) and has also been applied to analysis ofthe spliceosome (Rymond & Rosbash, 1988)+ The de-rivatization technique presented here introduces site-specific modifications in the phosphodiester backbonerather than selected bases in RNA+ In addition to theability to direct the site of modification to a specificnucleotide position, this method allows for control ofthe size and chemical identity of the modifying group,significantly improving the flexibility of this approach+This technique should be applicable to a wide variety ofother projects involving RNA structure–function studies+
Determination of precise contact sites for several ofthe identified pre-mRNA–spliceosome interactions, com-bined with known distances on the pre-mRNA, will pro-vide parameters enabling the construction of schematic,three-dimensional models of the spliceosome+Althoughsuch models will be, by necessity, very simplistic, theywill provide a useful framework for further experimentsand a new perspective for interpretation of the existingdata+
MATERIALS AND METHODS
Oligonucleotides
RNA and DNA oligonucleotides were synthesized using anApplied Biosystems 390 synthesizer using phosphoramiditesfrom Glen Research (Sterling, Virginia)+ Phosphorothioatemodification was introduced during the chemical synthesis ofRNA using tetraethylthiuram disulfide (TETD) in acetonitrile,according to manufacturer’s recommendations (Applied Bio-systems)+ The sequence of the 11-nt wt 59SS RNA used inmost experiments was 59-AAG/GUAAGUAdT-39+ A24 59SSRNA used in Figure 7A is one nucleotide longer than othersubstrates because it contains an additional A residue in the59 exon segment+ For trans-splicing experiments (Fig+ 6), 59-A9G/GUAAGUAT 18-nt 59SS RNA labeled at the 39 end byextension with three 32P-dC residues was used+ In addition,the 59SS DNA, CAGGTAAGTAT, and oligos complementaryto the following regions of snRNAs were used: U1, 1–11;U2g, 29–42; U4b, 58–76; and U6f, 33–48+
Most of the 59SS RNAs used in this study were 59 end-labeled using T4 polynucleotide kinase (NEB) and g-32P-ATP(NEN)+ To accurately measure the RNA concentration, at leasta twofold molar excess of g-32P-ATP was used to ensurequantitative labeling+ The RNAs were purified in a 20% poly-acrylamide/8 M urea gel+ For the 39 end-labeling, 59SS RNAs
1080 M. Sha et al.
Cold Spring Harbor Laboratory Press on July 13, 2011 - Published by rnajournal.cshlp.orgDownloaded from

were annealed to a complementary DNA oligo containingthree 59-terminal dGs+ The extension reaction contained theannealed RNA–DNA duplex, 32P-adCTP, and Klenow frag-ment of DNA polymerase (Boehringer), and was incubatedfor 4 h at room temperature, generating RNA with three ad-ditional 32P-dCs at the 39 end+ The RNA used in Figure 7D,lanes 3–6,A5G/GUAAGdT(s)dA32P-dT, was labeled in a sim-ilar reaction containing thio-dATP and 32P-adTTP, extendingthe product by two nucleotides at the 39 end+
To allow for RNase A digestion of crosslinks generated withAPA-RNAs without the loss of label, substrates containing29-H groups at both U12 and U16 positions were used+Alternatively, 39 end-labeled 59SS RNAs were used in whichonly one position (U16 for RNase A or G15 for RNase T1digestions) was modified with 29-H or 29-O-Me groups+
Derivatization of the 5 9SS RNA
The thio-59SS RNAs were stored in the presence of 1 mMDTT+ Derivatization was performed in a 10-mL reaction con-taining 10 mM 4-azidophenacyl bromide (Fluka) in 80% DMSOat 37 8C for 2 h+ Derivatizations with PB and IAA (Sigma)were performed under similar conditions+ Excess of the re-agent was removed by phenol, followed by chloroform ex-traction, and the RNA was recovered by ethanol precipitation+The extent of derivatization was monitored in a 20% gel fol-lowed by autoradiography and/or phosphorimager scanning+For experiments shown in Figure 4, all derivatized 59SS RNAswere gel purified to ensure homogeneity of products+
Crosslinking experiments
Splicing reactions (10 mL) were performed as described (Kon-forti & Konarska, 1995)+ In most experiments, the 39SS RNAcontained 83 nt of intron and 45 nt exon and was transcribedfrom pBSAd13 plasmid (Konarska, 1989)+ In Figure 3A andB, lane 6, the 39SS RNA contained 112 nt of the second intronand 54 nt of the third exon of the rabbit b-globin (Lamondet al+, 1987)+ For crosslinking experiments, reactions weretransferred to a microtiter plate embedded in ice and UV-irradiated 3 3 5 min (15 min total) at 254 nm for the wt RNAor for 2 min at 302 nm for APA-RNAs+ A 302-nm UV lampfrom UVP Inc+ (model UVM-57) or a 254-nm UV lamp fromSpectroline (model EF 280C) were used+ To minimize shortUV irradiation, the lid of a tissue culture dish (Becton Dick-inson, Lincoln Park, New Jersey) was used as a filter with the302-nm UV lamp+ Samples were resolved in a 12+5% SDSgel+ In most experiments, crosslinks are not treated withRNases prior to gel analysis and thus the entire RNA oligo iscovalently attached to the crosslink+ Thus, the estimated mo-lecular weights of products were corrected for the knownmass of the 59SS RNA+
In order to reduce the background of crosslinks not asso-ciated with the spliceosomes, the crosslinked reactions(10 mL) were spun through a 100-mL cushion of buffer D(20 mM Hepes, pH 7+6, 20% glycerol, 100 mM KCl, and0+2 mM EDTA) for 30 min at 40K rpm in a Beckman TL-100ultracentrifuge in a TLA 100+3 rotor+ Under these conditions,splicing complexes and individual snRNPs are recovered inthe pellet, except for U1 snRNP, which is pelleted only par-tially+ Pelleted complexes were resolved by SDS-PAGE+
For the experiments shown in Figure 7B, splicing com-plexes were UV-irradiated and resolved in a 4% nondenatur-ing gel+ Gel sections containing complexes were cut out andresolved in a 15% SDS gel+ For quantitative analysis of splice-osome formation (Fig+ 4), electrophoresis was performed suchthat the unbound/unreacted 59SS RNA remained in the gel+Quantitation was performed using a Molecular DynamicsPhosphorImager+
To ensure that the signals shown in Figure 4 are not af-fected by differential stabilities of the RNA oligos, degradationprofiles of individual 59SS RNAs were determined+ Kinetics ofdegradation was obtained by incubating the 59SS RNA understandard splicing reactions as a function of time, and moni-toring the extent of degradation in a 20% gel+ Except for the59SS RNAs containing modifications at position G21, otherRNAs did not show major differences in their stability+
Immunoprecipitation experiments shown in Figure 8 wereperformed as described previously (Reyes et al+, 1996), usingthe rabbit polyclonal antiserum against human Prp8p, (70R,provided by Dr+ M+ Moore, Brandeis)+
ACKNOWLEDGMENTS
We are particularly grateful to M+ Moore, R+ Lührmann, B+Laggerbauer, A+ Krämer, R+ Reed, M+ Green, and N+Chaudhary for antibody reagents and to C+ Query and mem-bers of the Konarska laboratory for helpful discussions andcritical reading of the manuscript+ This work was supported inpart by the Norman and Rosita Winston Foundation Fellow-ship Award to M+S+ and by the Monique Weill-Caulier Awardand an NIH grant to M+M+K+
Received April 30, 1998; returned for revision May 20,1998; revised manuscript received June 5, 1998
REFERENCES
Anderson GJ, Bach M, Lührmann R, Beggs JD+ 1989+ Conservationbetween yeast and man of a protein associated with U5 snRNP+Nature 342:819–821+
Behrens SE, Lührmann R+ 1991+ Immunoaffinity purification of a [U4/U6+U5] tri-snRNP from human cells+ Genes & Dev 5:1439–1452+
Burgin AB, Pace NR+ 1990+ Mapping the active site of ribonucleaseP RNA using a substrate containing a photoaffinity agent+ EMBOJ 9:4111–4118+
Cassetti MC, Moss B+ 1996+ Interaction of the 82-kDa subunit of thevaccinia virus early transcription factor heterodimer with the pro-moter core sequence directs downstream DNA binding of the70-kDa subunit+ Proc Natl Acad Sci USA 93:7540–7545+
Chaudhary N, McMahon C, Blobel G+ 1991+ Primary structure of ahuman arginine-rich nuclear protein that colocalizes with splice-osome components+ Proc Natl Acad Sci USA 88:8189–8193+
Chiara MD, Gozani O, Bennett M, Champion-Arnaud P, Palandjian L,Reed R+ 1996+ Identification of proteins that interact with exonsequences, splice sites, and the branchpoint sequence duringeach stage of spliceosome assembly+ Mol Cell Biol 16:3317–3326+
Conway L, Wickens M+ 1989+ Modification interference analysis ofreactions using RNA substrates+ Methods Enzymol 180:369–379+
Crispino JD, Sharp PA+ 1995+A U6 snRNA:pre-mRNA interaction canbe rate-limiting for U1-independent splicing+Genes & Dev 9:2314–2323+
Fabrizio P, Esser S, Kastner B, Luhrmann R+ 1994+ Isolation of S.cerevisiae snRNPs: Comparison of U1 and U4/U6+U5 to theirhuman counterparts+ Science 264:261–265+
Spliceosome components in proximity of the 59 splice site 1081
Cold Spring Harbor Laboratory Press on July 13, 2011 - Published by rnajournal.cshlp.orgDownloaded from

Fabrizio P, Laggerbauer B, Lauber J, Lane WS, Lührmann R+ 1997+An evolutionarily conserved U5 snRNP-specific protein is a GTP-binding factor closely related to the ribosomal translocase EF-2+EMBO J 16:4092–4106+
Fleckner J, Zhang M, Valcarcel J, Green MR+ 1997+ U2AF65 recruitsa novel human DEAD box protein required for the U2 snRNP-branchpoint interaction+ Genes & Dev 11:1864–1872+
Frey PA, Sammons RD+ 1985+ Bond order and charge localization innucleoside phosphorothioates+ Science 228:541–545+
Fu XD+ 1995+ The superfamily of arginine/serine-rich splicing factors+RNA 1:663–680+
Gaur RK, Valcarcel J, Green MR+ 1995+ Sequential recognition of thepre-mRNA branch point by U2AF65 and a novel spliceosome-associated 28-kDa protein+ RNA 1:407–417+
Gish G, Eckstein F+ 1988+ DNA and RNA sequence determinationbased on phosphorothioate chemistry+ Science 240:1520–1522+
Gozani O, Patton JG, Reed R+ 1994+ A novel set of spliceosome-associated proteins and the essential splicing factor PSF bindstably to pre-mRNA prior to catalytic step II of the splicing reac-tion+ EMBO J 13:3356–3367+
Harris ME, Nolan JM, Malhotra A, Brown JW, Harvey SC, Pace NR+1994+ Use of photoaffinity crosslinking and molecular modeling toanalyze the global architecture of ribonuclease P RNA+ EMBO J13:3953–3963+
Hixson SH, Hixson SS+ 1975+ P-Azidophenacyl bromide, a versatilephotolabile bifunctional reagent+ Reaction with glyceraldehyde-3-phosphate dehydrogenase+ Biochemistry 14:4251–4254+
Jeltsch A, Maschke H, Selent U, Wenz C, Köhler E, Connolly BA,Thorogood H, Pingoud A+ 1995+ DNA binding specificity of theEcoRV restriction endonuclease is increased by Mg21 binding toa metal ion binding site distinct from the catalytic center of theenzyme+ Biochemistry 34:6239–6246+
Konarska MM+ 1989+ Analysis of splicing complexes and small nu-clear ribonucleoprotein particles by native gel electrophoresis+Methods Enzymol 180(Part A):442–453+
Konforti BB, Konarska MM+ 1995+A short 59 splice site RNA oligo canparticipate in both steps of splicing in mammalian extracts+ RNA1:815–827+
Konforti BB, Konarska MM+ 1994+ U4/U5/U6 snRNP recognizes the59 splice site in the absence of U2 snRNP+ Genes & Dev 8:1962–1973+
Konforti BB, Koziolkiewicz MJ, Konarska MM+ 1993+ Disruption ofbase pairing between the 59 splice site and the 59 end of U1snRNA is required for spliceosome assembly+ Cell 75:863–873+
Krämer A+ 1996+ The structure and function of proteins involved inmammalian pre-mRNA splicing+ Annu Rev Biochem 65:367–409+
Krol A, Carbon P+ 1989+A guide for probing native small nuclear RNAand ribonucleoprotein structures+ Methods Enzymol 180:212–227+
Lagrange T, Kim TK, Orphanides G, Ebright YW, Ebright RH, Rein-berg D+ 1996+High-resolution mapping of nucleoprotein complexesby site-specific protein–DNAphotocrosslinking:Organization of thehuman TBP-TFIIA-TFIIB-DNAquaternary complex+Proc Natl AcadSci USA 93:10620–10625+
Lamond AI, Konarska MM, Sharp PA+ 1987+ A mutational analysis ofspliceosome assembly: Evidence for splice site collaboration dur-ing spliceosome formation+ Genes & Dev 1:532–543+
Lauber J, Fabrizio P, Teigelkamp S, Lane WS, Hartmann E, Luhr-mann R+ 1996+ The HeLa 200 kDa U5 snRNP-specific proteinand its homologue in Saccharomyces cerevisiae are members ofthe DEXH-box protein family of putative RNA helicases+ EMBO J15:4001–4015+
Lauber J, Plessel G, Prehn S, Will CL, Fabrizio P, Gröning K, LaneWS, Lührmann R+ 1997+ The human U4/U6 snRNP contains 60and 90kD proteins that are structurally homologous to the yeastsplicing factors Prp4p and Prp3p+ RNA 3:926–941+
LeCuyer KA, Behlen LS, Uhlenbeck OC+ 1996+ Mutagenesis of astacking contact in the MS2 coat protein–RNA complex+ EMBO J15:6847–6853+
MacMillan AM, Query CC, Allerson CR, Chen S, Verdine GL, SharpPA+ 1994+ Dynamic association of proteins with the pre-mRNAbranch region+ Genes & Dev 8:3008–3020+
Milligan JF, Uhlenbeck OC+ 1989+ Determination of RNA–protein con-tacts using thiophosphate substitutions+ Biochemistry 28:2849–2855+
Moore MJ, Query CC, Sharp PA+ 1993+ Splicing of precursors tomRNA by the spliceosome+ In: Gesteland RF, Atkins JF, eds+ TheRNA world+ Cold Spring Harbor, New York: Cold Spring HarborLaboratory Press+ pp 303–357+
Musier-Forsyth K, Schimmel P+ 1994+ Acceptor helix interactions in aclass II tRNA synthetase: Photoaffinity cross-linking of an RNAminiduplex substrate+ Biochemistry 33:773–779+
Newman AJ, Norman C+ 1992+ U5 snRNA interacts with exon se-quences at 59 and 39 splice sites+ Cell 68:743–754+
Newman AJ, Teigelkamp S, Beggs JD+ 1995+ snRNA interactions at59 and 39 splice sites monitored by photoactivated crosslinking inyeast spliceosomes+ RNA 1:968–980+
Nilsen TW+ 1994+ RNA–RNA interactions in the spliceosome: Unrav-eling the ties that bind+ Cell 78:1–4+
O’Keefe RT, Norman C, Newman AJ+ 1996+ The invariant U5 snRNAloop 1 sequence is dispensable for the first catalytic step of pre-mRNA splicing in yeast+ Cell 86:679–689+
Ofengang J, Schwartz I, Chinali G, Hixson SS, Hixson SH+ 1977+Photoaffinity-probe-modified tRNA for the analysis of ribosomalbinding sites+ Methods Enzymol XLVI:683–702+
Pinto AL, Steitz JA+ 1989+ The mammalian analogue of the yeastPRP8 splicing protein is present in the U4/5/6 small nuclear ri-bonucleoprotein particle and the spliceosome+ Proc Natl Acad SciUSA 86:8742–8746+
Query CC, Strobel SA, Sharp PA+ 1996+ Three recognition events atthe branch-site adenine+ EMBO J 15:1392–1402+
Reyes JL, Kois P, Konforti BB, Konarska MM+ 1996+ The canonicalGU dinucleotide at the 59 splice site is recognized by p220 of theU5 snRNP within the spliceosome+ RNA 2:213–225+
Rymond BC, Rosbash M+ 1988+A chemical modification/interferencestudy of yeast pre-mRNA spliceosome assembly and splicing+Genes & Dev 2:428–439+
Sawa H,Abelson J+ 1992+ Evidence for a base-pairing interaction be-tween U6 small nuclear RNA and 59 splice site during the splicingreaction in yeast+ Proc Natl Acad Sci USA 89:11269–11273+
Sawa H, Shimura Y+ 1992+ Association of U6 snRNA with the 59-splice site region of pre-mRNA in the spliceosome+ Genes & Dev6:244–254+
Singh R, Valcarcel J, Green MR+ 1995+ Distinct binding specificitiesand functions of higher eukaryotic polypyrimidine tract-bindingproteins+ Science 268:1173–1176+
Sontheimer EJ, Steitz JA+ 1993+ The U5 and U6 snRNAs as activesite components of the spliceosome+ Science 262:1989–1996+
Staley JP, Guthrie C+ 1998+ Mechanical devices of the spliceosome:Motors, clocks, springs, and things+ Cell 92:315–326+
Teigelkamp S, Newman AJ, Beggs JD+ 1995+ Extensive interactionsof PRP8 protein with the 59 and 39 splice sites during splicingsuggest a role in stabilization of exon alignment by U5 snRNA+EMBO J 14:2602–2612+
Umen JG, Guthrie C+ 1995+ A novel role for a U5 snRNP protein in 39splice site selection+ Genes & Dev 9:855–868+
Umen JG, Guthrie C+ 1996+ Mutagenesis of the yeast gene PRP8reveals domains governing the specificity and fidelity of 39 splicesite selection+ Genetics 143:723–739+
Wang JF, Downs WD, Cech TR+ 1993+ Movement of the guide se-quence during RNA catalysis by a group I ribozyme+ Science 260:504–508+
Wassarman DA, Steitz JA+ 1992+ Interactions of small nuclear RNAswith precursor mRNA during in vitro splicing+ Science 257:1918–1925+
Willis MC, Hicke BJ, Uhlenbeck OC, Cech TR, Koch TH+ 1993+ Photo-crosslinking of 5-iodouracil-substituted RNA and DNA to proteins+Science 262:1255–1257+
Wu S, Green MR+ 1997+ Identification of a human protein that rec-ognizes the 39 splice site during the second step of pre-mRNAsplicing+ EMBO J 16:4421–4432+
Zhang WJ, Wu JY+ 1996+ Functional properties of p54, a novel SRprotein active in constitutive and alternative splicing+Mol Cell Biol16:5400–5408+
1082 M. Sha et al.
Cold Spring Harbor Laboratory Press on July 13, 2011 - Published by rnajournal.cshlp.orgDownloaded from
Related Documents











