JOURNAL OF BACTERIOLOGY, Sept. 1968, p. 652-659 Copyright ( 1968 American Society for Microbiology Vol. 96, No. 3 Printed in U.S.A Spontaneous Lethal Sectoring, a Further Feature of Escherichia coli Strains Deficient in the Function of rec and uvr Genes K. HAEFNER1 Division of Biology, Southwest Center for Advanced Studies, Dallas, Texas 75230 Received for publication 25 March 1968 Eight recombination-deficient (Rec-) mutants of Escherichia coli were studied. Progeny lines were obtained on solid media, by means of micromanipulation, and the colony-forming ability of individual cells was analyzed. Cells of all eight strains gave rise to colony-forming as well as non-colony-forming descendants ("lethal sectoring"). Lethal sectors, i.e., groups of non-colony-forming cells which originate from a common ancestor, appeared with frequencies per generation ranging between 4 and 20 % in Rec- strains, whereas lethal sectors were rare in Rec+ strains (less than 1 %). A strain carrying a mutation (uvrA6) in one of the genes involved in pyrimidine dimer excision from deoxyribonucleic acid (DNA) showed twice as many lethal sectors per generation as a strain with the genotype uvrA+. Similarly, a double mutant (AB2480, uvrA6, recA13) showed twice as much spontaneous lethal sectoring as the corresponding Rec- strain (uvrA+, recA13). The kinetics of growth curves obtained in nutrient broth and the frequency of non-colony-forming units in stationary-phase broth cultures indicate clearly that lethal sectors occur in liquid cultures too. The causes for spontaneous lethal sectoring are unknown at present. It seems reasonable to assume that gene uvrA and the rec genes are somehow involved in the repair of spontaneously occurring DNA lesions, since a deficiency in this type of repair may cause lethal sectors. The extent to which spontaneous lethal sectoring (observed in all Rec- strains of E. coli studied) may contribute indirectly to the failure to form recombinants is discussed. Several ultraviolet light (UV)-sensitive mutants of Escherichia coli show lower frequencies of recombination than the corresponding strains with normal UV-sensitivity (3, 4, 11, 17). The observation that recombination-deficient mutants are also UV-sensitive suggests that these mutants are blocked in the repair of deoxyribonucleic acid (DNA) containing UV lesions (9, 11). However, there is some evidence (2) that Rec- strains are able to excise pyrimidine dimers, the first step in DNA repair. Furthermore, a strain defective in pyrimidine dimer excision as well as in recombination shows a considerably higher UV sensitivity than other UV-sensitive strains (9). UV-sensitive mutants of E. coli that are excision-defective are able to form recombinants at the normal frequencies (10); thus, there appear to be at least two (more or less independent) systems that promote cell survival following UV irradiation: the recombination system and the 1 Present address: Institut fur Genetik, Universi- tat Freiburg, 78 Freiburg/Br., Germany. excision system. If these also are involved in the repair of spontaneous DNA lesions, which might occur in normally growing cells, a deficiency in one or both of them should result in higher rates of spontaneous mutations or in other cellular defects. Such effects have not yet been reported. In this paper, results obtained by micromanipu- latory pedigree analysis of individual cells are presented. They show that normally growing Rec- cells and cells which lack the ability to excise pyrimidine dimers give rise spontaneously to many non-colony-forming descendants. We have called this phenomenon spontaneous lethal sector- ing (SLS) in analogy to radiation-induced lethal sectoring (see, e.g., 8). The finding of SLS shows clearly that a deficiency in the recombination system or in the excision repair system can result in cellular defects. Some models explaining SLS will be discussed. MATERIALS AND METHODS E. coli strains. AB1157 ("wild type"), AB2463 (recA13, "reckless"), AB2470 (recB21, "cautious"), 652 on May 15, 2018 by guest http://jb.asm.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

JOURNAL OF BACTERIOLOGY, Sept. 1968, p. 652-659Copyright ( 1968 American Society for Microbiology
Vol. 96, No. 3Printed in U.S.A
Spontaneous Lethal Sectoring, a Further Feature ofEscherichia coli Strains Deficient in the
Function of rec and uvr GenesK. HAEFNER1
Division of Biology, Southwest Center for Advanced Studies, Dallas, Texas 75230
Received for publication 25 March 1968
Eight recombination-deficient (Rec-) mutants of Escherichia coli were studied.Progeny lines were obtained on solid media, by means of micromanipulation, andthe colony-forming ability of individual cells was analyzed. Cells of all eight strainsgave rise to colony-forming as well as non-colony-forming descendants ("lethalsectoring"). Lethal sectors, i.e., groups of non-colony-forming cells which originatefrom a common ancestor, appeared with frequencies per generation ranging between4 and 20% in Rec- strains, whereas lethal sectors were rare in Rec+ strains (lessthan 1 %). A strain carrying a mutation (uvrA6) in one of the genes involved inpyrimidine dimer excision from deoxyribonucleic acid (DNA) showed twice asmany lethal sectors per generation as a strain with the genotype uvrA+. Similarly,a double mutant (AB2480, uvrA6, recA13) showed twice as much spontaneouslethal sectoring as the corresponding Rec- strain (uvrA+, recA13). The kinetics ofgrowth curves obtained in nutrient broth and the frequency of non-colony-formingunits in stationary-phase broth cultures indicate clearly that lethal sectors occur inliquid cultures too. The causes for spontaneous lethal sectoring are unknown atpresent. It seems reasonable to assume that gene uvrA and the rec genes are somehowinvolved in the repair of spontaneously occurring DNA lesions, since a deficiencyin this type of repair may cause lethal sectors. The extent to which spontaneouslethal sectoring (observed in all Rec- strains of E. coli studied) may contributeindirectly to the failure to form recombinants is discussed.
Several ultraviolet light (UV)-sensitive mutantsof Escherichia coli show lower frequencies ofrecombination than the corresponding strainswith normal UV-sensitivity (3, 4, 11, 17). Theobservation that recombination-deficient mutantsare also UV-sensitive suggests that these mutantsare blocked in the repair of deoxyribonucleicacid (DNA) containing UV lesions (9, 11).However, there is some evidence (2) that Rec-strains are able to excise pyrimidine dimers, thefirst step in DNA repair. Furthermore, a straindefective in pyrimidine dimer excision as well asin recombination shows a considerably higherUV sensitivity than other UV-sensitive strains(9). UV-sensitive mutants of E. coli that areexcision-defective are able to form recombinantsat the normal frequencies (10); thus, there appearto be at least two (more or less independent)systems that promote cell survival following UVirradiation: the recombination system and the
1 Present address: Institut fur Genetik, Universi-tat Freiburg, 78 Freiburg/Br., Germany.
excision system. If these also are involved in therepair of spontaneous DNA lesions, which mightoccur in normally growing cells, a deficiency inone or both of them should result in higher ratesof spontaneous mutations or in other cellulardefects. Such effects have not yet been reported.
In this paper, results obtained by micromanipu-latory pedigree analysis of individual cells arepresented. They show that normally growingRec- cells and cells which lack the ability to excisepyrimidine dimers give rise spontaneously tomany non-colony-forming descendants. We havecalled this phenomenon spontaneous lethal sector-ing (SLS) in analogy to radiation-induced lethalsectoring (see, e.g., 8). The finding of SLS showsclearly that a deficiency in the recombinationsystem or in the excision repair system can resultin cellular defects. Some models explaining SLSwill be discussed.
MATERIALS AND METHODSE. coli strains. AB1157 ("wild type"), AB2463
(recA13, "reckless"), AB2470 (recB21, "cautious"),652
on May 15, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

SPONTANEOUS LETHAL SECTORING
AB2437 (uvrA6), and AB2480 (uvrA6, recA13) wereobtained from P. Howard-Flanders (some of themthrough W. Harm; for further characteristics of thesestrains, see references 10 and 11). KMBL146 ("wildtype"), KMBL239 (rec-34), KMBL240 (rec-35),KMBL241 (rec-36), and KMBL243 (rec-38) wereobtained from A. Rorsch (these strains are furthercharacterized in reference 17). Strain KMBL243seems to be "cautious" Rorsch, personal communica-tion. JC411 ("wildtype"), JC1553 (recAl, "reckless"),and JC4535 (recB60) were obtained from A. J. Clark(for further characteristics of strains JC411 andJC1553, see reference 3).
Media. Nutrient broth (NB) contained 8 g ofNutrient Broth (Difco) and 5 g of NaCl in 1,000 mlof water. Minimal medium always contained 5 g ofNH4Cl, 1 g ofNH4NO3, 2 g ofNa2SO4, 3 g ofK2HPO4,1 g of KH2PO4, 0.1 g of MgSO4-7H20, and 20 g ofglucose, in 1,000 ml of water. To allow growth of theAB strains, it was supplemented with 20 mg of threo-nine, 20 mg of proline, 20 mg of histidine, 20 mg ofarginine, 20 mg of leucine, and 1 mg of thiamine perliter. This supplemented medium is called SMH. Asecond supplemented medium called SMR was usedfor the culture of KMBL strains; SMR containedminimal medium supplemented with 20 mg of threo-nine, 20 mg of leucine, 20 mg of arginine, 20 mg ofhistidine, 20 mg of isoleucine, 20 mg of thymine, 20mg of uracil, 20 mg of cytosine, and 1 mg of thiamine.Agar (2%) was added for solid media.
Pedigree analysis. The same procedure was fol-lowed in all experiments on individual cells. Theywere grown at 37 C until they reached the conditionsdesired in the experiment. Then they were washedonce in 0.05 M KH2PO4 (pH 6.5) and streaked atthe edge of an agar medium slab (14 X 36 mm; uni-form thickness, 1.5 mm) attached to a cover glass.(The production of such agar layers is described inreference 7.) The slab was placed upside down in achamber which was open on one side. The chamberwith the slab was resting on the stage of a Zeiss phase-contrast microscope equipped with long working con-densor and objective (40X). Transfer of cells on theslab surface was accomplished by means of a deFonbrune micromanipulator.Three single cells per slab were picked with the
micromanipulator needle from the streak of cells at
the edge, and placed individually in chosen positions.Cells were incubated at 37 C and inspected frequentlyunder the microscope at room temperature. (Nor-mally cells were kept at 37 C for about 60% of thetime of pedigree analysis.) After each division, theprogeny cells were separated and placed within thesame field of vision of the microscope in a patternwhich allowed the arrangement of 16 descendants.Pedigrees normally were followed through the fourthgeneration. The 16 fourth-generation descendantsand their small progeny, were later relocated withsufficient space between them to enable the growth ofindividual macroscopic colonies. At the time ofrelocation, transferred cell groups were counted andregistered to obtain the data for the clone-size distri-bution of lethal sectors. After the micromanipulationperiod, the slabs were incubated at 37 C for 18 hr;then colonies were counted. Normally, 30 four-generation-pedigrees were handled simultaneously inone experiment.
Growth curves. A Brinkman biophotometer wasused to determine the rate of growth of the variousE. coli strains. This instrument plots the opticaltransmission values for six cultures simultaneously.Growth curves were obtained in NB at approximately37 C. The inoculum was 0.02 ml from an overnightshaker culture into 8 ml. The slopes of the growthcurves were expressed as the percentage decrease inoptical transmission per hour, taking only the expo-nential growth phase into consideration.
RESULTS
Pattern in which non-colony-forming cells ap-pear. Eight genetically different Rec- mutantsand two strains deficient in the excision repairsystem were studied by means of micromanipula-tion pedigree analysis of individual cells. In allstrains non-colony-forming descendants occurredfrequently in the progeny of cells growing nor-mally for four generations on solid media. Twoexamples of the pattern in which such non-colony-formers appear in pedigrees derived from strainAB2480 are given in Fig. 1. It can be seen that
Frequency
I?O of LS
I: 1/4 =0.25If: 2/6 =0.33
4 M: 1/8=0.13
{SE6 LBSi SJJ lLETAL SECTORJ i LETHA CTOR LETHAL SECTOR JFIG. 1. Pattern in which inactive cells occur in pedigrees from strain AB2480. An open circle in the fourth
generation refers to a cell which forms a colony. A closed circle indicates a cell which is unable to form a colony.Such a cell, however, may undergo reisdual divisions. A lethal sector (LS) is defined as a group of non-colony-forming cells which descend from a common ancestor. The clone size ofa lethal sector is defined by the number ofcells forming this lethal sector, including all descendants of inactive cells which have been observed within 6 hrafter placing the parental cell. The Roman numbers refer to the generations. In each generation, the frequency oflethal sectors per generation (W/l + c) was computed for the two pedigrees. They are shown on the right side. Thevalues can never exceed 0.50.
653VOL. 96, 1968
on May 15, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

J. BACrERIOL.
groups of non-colony-forming cells occur whichoriginate from a common ancester. Each of thesegroups of non-colony-formers will be called alethal sector.We will present four kinds of information
which can be derived from pedigree experiments.(i) The fraction of cells of the original populationthat gives rise to at least one colony-formingdescendant (survival) was determined. If we as-sume, for example, that studying a population oftwo cells has led to the pedigrees shown in Fig. 1,this value would be 1.0, i.e., two out of two.(ii) The fraction of the colony-forming cellswhich give rise to at least one non-colony-forming descendant within the four generationsstudied is presented. For the population shownin the figure, this value would be 1.0 i.e. two outof two. (iii) The frequency of lethal sectors pergeneration (I/l + c) was computed by dividingthe number of lethal sectors (1) originating in agiven generation by the sum of lethal sectors pluscolony-forming cells (c) in this generation (see thecalculations in Fig. 1). (iv) The clone-size distribu-tion of the lethal sectors, expressed in terms ofthe frequency of lethal sectors which are formedby a given number of inactive cells, is given.Assuming that none of the non-colony-formingcells in Fig. 1 had undergone residual divisions,44% (i.e., three of the total of seven lethal sectors)would have the clone size of one cell; 14% (i.e.,one of seven) would have the clone size of twocells; 14% (i.e., one of seven) would have theclone size of three cells; and 28% (i.e., two ofseven) would have the clone size of four cells.The time at which cell division took place was
noted in these experiments, but will not be con-sidered in this paper.Experiments with Rec- cells growing on NB
agar. In the first set of experiments on NBagar, pedigrees were derived from small cells(length: width > 1.3) of an overnight culture inNB grown on a shaker. For eight Rec- strains andthree related "wild-type" strains, survival and thefraction of colony-forming cells which give riseto at least one inactive descendant are shown inTable 1. In comparison with the "wild-type"strains, survival of Rec- strains was lower andthe frequency of colony-forming cells which gaverise to a non-colony-forming descendant was con-siderably higher. For example, in the Rec- strainAB2470, only 39% of the cells formed a colony,and of those 82% gave rise to lethal sectors; incontrast, 89% of the cell from strain AB1157(Rec+) formed a colony and only 6% showedlethal sectors.The frequency of lethal sectors per generation
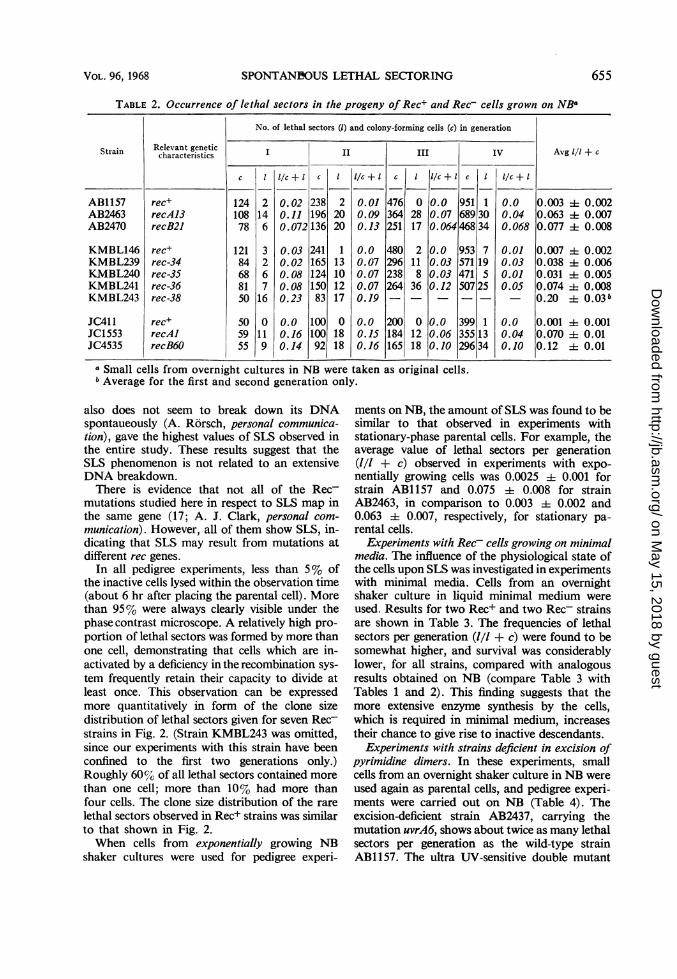
(1/1 + c) is 0.007 or less in Rec+ strains, whereasit is considerably higher in Rec- strains (Table 2).
TABLE 1. Survival and the frequency of colony-form-ing cells with inactive descendants for eight Recrstrains
and the appropriate Rec+ parental strainsa
Strain
AB 1157AB 2463
AB 2470
KMBL 146KMBL 239KMBL 240KMBL 241KMBL-243
JC 411JC 1553
JC 4535
Relevant geneticcharacteristics
rec+recA13 ("reck-
less")recB21 ("cau-
tious")
rec+rec-34rec-35rec-36rec-38 ("cau-
tious")
rec+recAl ("reck-
less")recB60
No. ofcellsana-lyzed
6361
42
6243374433
2535
32
Survival(frequen-cy ofcolony-formingcells)
0.890.74
0.39
0.710.710.330.500.18
0.970.63
0.53
Fractionof colony-formingcells withinactivedescen-dants
0.060.85
0.82
0.180.540.660.910.79
0.040.81
0.94
a Progeny lines were followed for four generations on NB,starting with small cells from a stationary phase overnightshaker culture in NB.
(Since there seems to be a clear correlation be-tween the Rec- characteristics and SLS in E. coli,it was asked whether every strain showing SLSis recombination-deficient. We studied this inSchizosaccharomyces pombe. However, two mu-tants which gave about 10% lethal sectors pergeneration did not show altered recombinationfrequencies in intragenic crosses of two ad6 alleles.)For all Rec- strains, there is a slight decrease ofthe frequency of lethal sectors per generation ifone compares the first and second generationswith later generations. In experiments in whichup to seven generations were taken into considera-tion, the frequency of lethal sectors per generationdid not decrease below the values obtained forthe third and fourth generations. Therefore, itseems justified to average the frequencies over thefirst four generations. This allows the derivationof a simple quantitative term for SLS, which isgiven in the last column of Table 2. This averagevalue of lethal sectors per generation varies con-siderably for various Rec- strains. The highestvalues (0.12 and '-..0.20) were observed for strainsJC4535 and KMBL243, respectively.
Strains AB2463 and JC1553, which are knownto break down their DNA spontaneously ["reck-less" strains, (2, 9)] gave the same frequencies oflethal sectors per generation, or even less, relativeto strain AB2470, which does not break down itsDNA spontaneously (9). Strain KMBL243, which
654 HAEFNER
on May 15, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
SPONTANIBOUS LETHAL SECTORING
TABLE 2. Occurrence of lethal sectors in the progeny of Rec+ and Rec- cells grown on NB
No. of lethal sectors (I) and colony-forming cells (c) in generation
I II III IV
12410878
12184688150
505955
2146
326716
0
119
I/c+l c
0.020.110.072
0.030.020.080.080.23
0.00.160.14
238196136
24116512415083
10010092
22020
113101217
0
1818
I/c+ l
0.010.090.13
0.00.070.070.070.19
0.00.Js0.16
c
476364251
480296238264
200184165
I I/c +I
0 0.028 0.0717 0.064
2118
36
0
1218
0.00.030.030.12
0.00.060.10
c
951689468
953571471507
399355296
a Small cells from overnight cultures in NB were taken as original cells.bAverage for the first and second generation only.
1
' 3034
7195
25
1
13
34
I/c + I
0.00.040.068
0.010.030.010.05
0.00.040.10
Avg 1/1 + c
0.003 A 0.0020.063 4 0.0070.077 i 0.008
0.007 4 0.0020.038 1 0.0060.031 4 0.0050.074 4 0.0080.20 4 0.03b
0.001 h 0.0010.070 =1: 0.010.12 + 0.01
also does not seem to break down its DNAspontaueously (A. Rorsch, personal communica-tion), gave the highest values of SLS observed inthe entire study. These results suggest that theSLS phenomenon is not related to an extensiveDNA breakdown.There is evidence that not all of the Rec-
mutations studied here in respect to SLS map inthe same gene (17; A. J. Clark, personal com-
munication). However, all of them show SLS, in-dicating that SLS may result from mutations atdifferent rec genes.
In all pedigree experiments, less than 5% ofthe inactive cells lysed within the observation time(about 6 hr after placing the parental cell). Morethan 95% were always clearly visible under thephase contrast microscope. A relatively high pro-portion of lethal sectors was formed by more thanone cell, demonstrating that cells which are in-activated by a deficiency in the recombination sys-tem frequently retain their capacity to divide atleast once. This observation can be expressedmore quantitatively in form of the clone sizedistribution of lethal sectors given for seven Rec-strains in Fig. 2. (Strain KMBL243 was omitted,since our experiments with this strain have beenconfined to the first two generations only.)Roughly 60% of all lethal sectors contained morethan one cell; more than 10% had more thanfour cells. The clone size distribution of the rarelethal sectors observed in Rec+ strains was similarto that shown in Fig. 2.When cells from exponentially growing NB
shaker cultures were used for pedigree experi-
ments on NB, the amount of SLS was found to besimilar to that observed in experiments withstationary-phase parental cells. For example, theaverage value of lethal sectors per generation(1/1 + c) observed in experiments with expo-nentially growing cells was 0.0025 i 0.001 forstrain AB1157 and 0.075 i 0.008 for strainAB2463, in comparison to 0.003 + 0.002 and0.063 i4 0.007, respectively, for stationary pa-rental cells.
Experiments with Rec- cells growing on minimalmedia. The influence of the physiological state ofthe cells upon SLS was investigated in experimentswith minimal media. Cells from an overnightshaker culture in liquid minimal medium wereused. Results for two Rec+ and two Rec- strainsare shown in Table 3. The frequencies of lethalsectors per generation (1/1 + c) were found to besomewhat higher, and survival was considerablylower, for all strains, compared with analogousresults obtained on NB (compare Table 3 withTables 1 and 2). This finding suggests that themore extensive enzyme synthesis by the cells,which is required in minimal medium, increasestheir chance to give rise to inactive descendants.
Experiments with strains deficient in excision ofpyrimidine dimers. In these experiments, smallcells from an overnight shaker culture in NB wereused again as parental cells, and pedigree experi-ments were carried out on NB (Table 4). Theexcision-deficient strain AB2437, carrying themutation uvrA6, shows about twice as many lethalsectors per generation as the wild-type strainABI 157. The ultra UV-sensitive double mutant
Strain
AB1157AB2463AB2470
KMBL146KMBL239KMBL240KMBL241KMBL243
JC411JC1553JC4535
Relevant geneticcharacteristics
rec+recA13recB21
rec+rec-34rec-35rec-36rec-38
rec+recAlrecB60
VOL. 96, 1968 655
I
on May 15, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

656
w-1L1
wz0-J
C-)
0
L)zwawLL
JC15532 AB24631
JC45351 AB2470J
KMBL239H
KMBL22041
KMBL241]
5 -8
CLONE SIZE OF LETHAL SECTORS (CELLS)FIG. 2. Clone size distributionz of lethal sectors. In this distribution all lethal sectors observed were taken i/ito
account, irrespective of the generationi in which they occurred. Several LS have beenl observed which were formedby more than 32 cells.
TABLE 3. Occurrence of lethal sectors in the progeny of Rec+ and Rec- cells grown on minimal mediaa
No of colony-forming cells (c) and lethal sectors (1) in generationRelevant
Strain character- Survival I II III IV Avg 1/l + £istics
c 1 f+c c 1 I+c c 1+c c I +c
AB 1157 rec+ 0.59 80 2i 0.02 158 2 0.01 - _- |_ - 0.016 4± 0.01lAB 2463 recA13 0.10 25 9 0.26 44 6 0.12 - - - - 0.18 4t 0.05b
KMBL 146 rec+ 0.67 85 5s 0.06 157 11 0.07 303 7 0.02 554 16 0.03 0.035 i 0.06KMBL 239 rec-34 0.29 98 121 0.11 176 18 0.09 332 28 0.09 637 29 0.04 0.065 S 0.007
a Cells from an overnight shaker culture in minimal medium (SMH and SMR, respectively) weretaken as parental cells. Medium SMH was used for the AB strains; medium SMR, for the KMBL strains.
b Average values for two generations only.
AB2480 (uvrA6, recA 13) gives twice as manylethal sectors per generation as strain AB2463carrying mutation recA13 alone. Apparently, thepresence of mutation uvrA6 increases the fre-quency of lethal sectors per generation by roughlya factor of two, independently of the level of SLSexpressed in the original strain.
E. coli B8_1, which is known not to excisepyrimidine dimers (15), shows a significantlyhigher rate of lethal sectors per generation thanB/r, the corresponding wild-type strain (Table 4).Since B.-, carries an hcr and an exr mutation,beside the lon mutation (14), it is not clear whichof those causes the observed increase in SLS.
Spontaneous lethal sectoring in liquid NB. Non-colony-forming descendants occurred when Rec-cells or cells deficient in the excision repair systemgrow on solid media. It seemed of interest toknow to what extent this phenomenon exists inliquid cultures. Micromanipulation, however, isimpossible, under these conditions; therefore,evidence for SLS has to be derived indirectly.Assuming that the frequency of lethal sectors per
generation is similar in liquid broth and on solidmedia, one should find: (i) a large fraction ofinactive cells in stationary-phase cultures ofstrains showing high rates of SLS on solid media,and (ii) smaller slopes of growth curves obtained
HAEFNER J. BACTERIOL.
70
60
50
40
20
10
2 3 -4
on May 15, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

SPONTANEOUS LETHAL SECTORING
TABLE 4. Spontaneous lethal sectoring in strains deficient in various systems which are involved in repairof radiation lesionsa
No. of colony-forming cells (c) and lethal sectors (1) ingeneration
Strain Rlcharacteristics Siural II III IV Avg 1/1 + c
c _ +C c -+c 'c I+Cl c I c+AB 1157 rec+uvrA+ 0.85 124 2 0.02 238 2 0.01 476 00.0 951 1 0.0 0.00340.002AB 2437 rec+ uvrA6 0.95 152 4 0.02 301 3 0.01 599 3 0.005 1192 6 0.005 0.007±0.002AB 2463 recA13 uvrA+ 0.74 108 14 0.11 196 20 0.09 364 28 0.07 689 30 0.04 0.063±0.007AB 2480 recA13 uvrA6 0.73 96 260.20 152 42 0.22 271 33 0.11 500400.074 0.122±4-0.01
B/r exr+ hcr+ 1.00 100 0 0.0 200 0 0.0 400 00.0 800 0 0.0 0.000±0.002BS-1 exr- hcr-b 0.96 100 2 0.02 196 4 0.02 387 5 0.01 771 3 0.005 0.0104±0.003
Small cells from an overnight NB shaker culture were taken as original cells. Pedigree experimentswere carried out on NB. Data for strains B/r and B.-, were taken from Haefner and Striebeck (8).
b See reference 13.
from strains with high SLS in comparison to"wild-type" strains.
Figure 3a shows that the frequency of non-colony-forming cells (1-fraction of colony-form-ing cells) and the average frequency of lethalsectors per generation show a positive correla-tion. Strains with high SLS values give highpercentages of non-colony-formers. Strain AB-2480 gave less non-survival than most of the othermutant strains, suggesting that this strain pro-duces fewer lethal sectors in liquid NB. The sta-tionary-phase culture of KMBL240 containedmany non-colony-forming units, which may indi-cate that this strain shows more lethal sectors inliquid NB than on NB agar.The interpretation of the growth cutves is more
complex. Within closely related strains, the ob-served slope values corresponded, as expected, tothe amount of SLS (Fig. 3b). In each group,strains with low average frequencies of lethalsectors per generation had higher slope valuesthan strains which showed extensive SLS.The observation of non-colony-forming cells
and the characteristics of the growth curvesindicate qualitatively that SLS also occurs inliquid NB at 37 C. A quantitative estimate of thefrequency of lethal sectors per generation underliquid conditions will be published elsewhere.
DISCUSSIONEight Rec- strains of E. coli, probably resulting
from eight different rec mutations, showed spon-taneous lethal sectoring (SLS) to a considerableextent (Table 1 and 2). This suggests that the recgenes, which appear to be involved in DNA inte-gration and in repair of UV-inducedDNA lesions,have a function also in normally growing cells.
A similar conclusion can be drawn for geneuvrA, which is involved in the excision repair ofUV photoproducts from DNA. A mutation inthis gene (uvrA6) increases the (1/1 + c) valuesby a factor of two (Table 4), thus indicating thatthis gene functions also in cells which have notbeen UV-irradiated.The SLS phenomenon seems to be the only
common feature of all rec mutants which isdisplayed in untreated normally growing cells[recA mutants exhibit spontaneous DNA degra-dation also (2, 11)]. Other properties, especiallypolymerase, nuclease, and "sealase" activities,have been reported to be unaffected in Rec- cells(6, 9).
Causes for SLS. There is no direct support forany molecular mechanisms causing SLS; there-fore, explanations for this phenomenon have tobe highly speculative. Our findings that "reckless"Rec- strains, which break down their DNAspontaneously, show lower, or at most equal,rates of SLS compared with "cautious" Rec-strains (Table 2) make it unlikely that SLS resultsfrom extensive DNA breakdown.
Tlhere is no way of ruling out the idea that SLSin Rec- strains may result from attempts ofrecombination between sister DNA moleculeswhich might be lethal in such strains. This pos-sibility of lethal recombination was considered byClark et al. (2) for Rec- strains, since there isevidence for recombination of sister chromosomesin Rec+ strains (5). The high level of SLS in thedouble mutant (uvrA6, recA13) relative to theuvr+ recA13 strain, however, is difficult to under-stand in these terms, since the uvrA6 mutation isknown not to affect recombination (10).
Since it is well established that the rec and uvr
VOL. 96, 1968 657
on May 15, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

HAEFNER
a
(X) KMBL243
AB2480 JC4535x x
JC 1553 X KMBL 241 X AB 2470AB2463X KM/L 2
AB 11571XKMBL239
AB 2437X KMBL 146%(-7.I X?~ 1 1
240
l
O 10 20 30 40 50 60 70 80 90 100PERCENT OF NON-COLONY-FORMING CELLS IN A STATIONARYPHASED NUTRIENT BROTH CULTURE
b
(X) KMBL 243
JC4553\- K"~AB27
NtAB2473 JC1553 6KMBL241AB 2463
KMBL 240 XX KMBL 239
AB\17 [KMBL146 CA II I
10 20 30SLOPE OF GROWTH CURVE (%/hr)
40
FIG. 3. Correlation between SLS observed on NB agar anid growth characteristics of cells grown in liquid NB.Itn Fig. 3a, the average frequency oflethal sectors per generation (from Tables 2 and 4) is plotted as a functioni ofthe fraction ofnon-colony-forming cells, using the survival values from Tables I and 4. In Fig. 3b, the averagefrequency oflethal sectors per generation (from Tables 2 and 4) is plotted as a function of the slope of the cor-
responding growth curve. The slope is given as percentage decrease in optical transmission per hour, taking intoconsideration the exponential growth period only. For strain KMBL 243, the average frequency oflethal sectorsper genterationi was for two generations only.
genes are involved in repair of UV-induced DNAlesions, it seems reasonable to assume that theyrepair spontaneously occurring DNA lesions too.The observed SLS might then be caused by a lackof repair of such DNA lesions. If this hypothesisis valid, then both the excision repair system andthe recombination system have to be consideredas natural "DNA guard systems," which keep thelevel of DNA lesions low.
Howard-Flanders and Boyce (9) have discusseda function of this type for the excision repairsystem. However, this idea was not supported by
their finding that the spontaneous mutation rateto valine resistance is the same in strains with thegenotypes uvr- and uvr+. [In Proteus mirabilis,Bohme (la) found increased frequencies of spon-taneous mutations in UV-sensitive strains.] Sinceno quantitative data were given in their paper, wecarried out similar determinations with the sameresult. However, this does not necessarily excludea more general function of the excision repairsystem, since it might well be that this system actspredominantly on lethal lesions.
Influence of SLS on recombination frequencies.
658 J. BACTERIOL.
i -
4 25M-_
w
i o 20U.4
o 15
LWW*
W cr,
L.aJ
10
n
o0 S
cn
25F-J4
zw
-r
U-
w
w
4t
e-
Z lS
20
bi 15zwCD
z 10
w
cn
0
on May 15, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

SPONTANEOUS LETHAL SECTORING
The classical experiments of Lederberg (12) andAnderson (1) and more recent investigations (16,18, 19); indicate that, in the course of bacterialrecombination in Rec+ strains of E. coli, DNAintegration takes place soon after DNA transferas well as in the exconjugant progeny carryingthe transferred and unreplicated Hfr genome.Integration in later generations, however, seemsto be of greater importance only in crosses withHfr Hayes donor strains (19). Low (13) presentedevidence that in crosses with Rec- "cautious"strains regular recombinants are formed, whereasRec- "reckless" strains tend to form no realrecombinants but do form merodiploids withpossibly integrated Hfr DNA pieces. In bothcases, it is, however, unknown at which stageafter cell conjugation the DNA integration stepmight occur.Presuming that any type of probably rare DNA
integration does occur, not only in the zygoteitself but also in the progeny ofRec- exconjugantscarrying the unintegrated Hfr DNA, and assum-ing further that these progeny lines show the SLSphenomenon, one has to expect that a givenfraction of such lines is inactivated before DNAintegration can occur. This would lead to de-creased numbers of recombinants and mero-diploids, respectively. Quantitative predictionsabout the extent to which the recombinationfrequencies are influenced by such a process,however, cannot be made, since nothing is knownin respect to SLS of Rec- cells carrying unin-tegrated Hfr DNA.
ACKNOWLEDGMENTSThis investigation was supported by AEC contract
AT-(40-1)-3601 and by grant GM 1323401 Al fromthe National Public Health Service. The excellentwork of Ute Striebeck, who did most of the difficultpedigree experiments, is greatly appreciated. I thankA. J. Clark, P. Howard-Flanders, A. Rorsch, andW. Harm for providing bacterial strains and A. J.Clark, W. Harm, and R. Hausmann for reading themanuscript critically.
LITERATURE CITED1. Anderson, T. K. 1958. Recombination and segre-
gation in Escherichia coli. Cold Spring HarborSymp. Quant. Biol. 23:47-58.
la. Bohme, H. 1967. Ultraviolet-sensitive mutantsof Proteus mirabilis with increased spontaneousmutability. Microbiol. Gen. Bull. 27:4.
2. Clark, A. J., M. Chamberlin, R. P. Boyce, andP. Howard-Flanders. 1966. Abnormal meta-bolic response to ultraviolet light of a recom-bination deficient mutant of Escherichia coli.J. Mol. Biol. 19:442-454.
3. Clark A. J., and A. D. Margulies. 1965. Isolation
and characterization of recombination-de-ficient mutants of Escherichia coli K 12. Proc.Natl. Acad. Sci. U.S. 53:451-459.
4. Clowes, R. C., and E. E. M. Moody. 1966.Chromosomal transfer from "recombination-deficient" strains of Escherichia coli K-12.Genetics 53:717-726.
5. Forro, F. 1965. Autoradiographic studies of bac-terial chromosome replication in amino-aciddeficient Escherichia coli 15T-. Biophys. J.5:629-649.
6. Gefter, M. L., A. Becker, and J. Hurwitz. 1967.The enzymatic repair of DNA. I. Formationof circular DNA. Proc. Natl. Acad. Sci. U.S.58:240-247.
7. Haefner, K. 1967. A simple apparatus for pro-ducing agar layers of uniform thickness formicrobiological micromanipulator work. Z.Allgem. Microbiol. 7:229-231.
8. Haefner, K., and U. Striebeck. 1966. Radiation-induced lethal sectoring in Escherichia coli B/rand B.-,. Mutation Res. 4:399-407.
9. Howard-Flanders, P., and R. P. Boyce. 1966.DNA repair and genetic recombination:Studies on mutants of Escherichia coli defectivein these processes. Radiation Res. Suppl.6:156-184.
10. Howard-Flanders, P., R. P. Boyce, and L.Theriot. 1966. Three loci in Escherichia coliK-12 that control the excision of pyrimidinedimers and certain other mutagen productsfrom DNA. Genetics 53:1119-1136.
11. Howard-Flanders, P., and L. Theriot. 1966.Mutants of Escherichia coli K-12 defective inDNA repair and in genetic recombination.Genetics 53:1137-1150.
12. Lederberg, J. 1957. Sibling recombinants inzygote pedigrees of Escherichia coli. Proc.Natl. Acad. Sci. U.S. 43:1060-1065.
13. Low, K. B. 1967. Discussion. J. Cell. Physiol.70(Suppl. 1) :165-180.
14. Mattern, I. E., H. Zwenk, and A. Rorsch. 1966.The genetic constitution of the radiation-sensi-tive mutant Escherichia coli B,,_. MutationRes. 3:374-380.
15. Setlow, R. B., and W. L. Carrier. 1964. The dis-appearance of thymine dimers from DNA: Anerror-correction mechanism. Proc. Natl. Acad.Sci. U.S. 51:226-231.
16. Tomizawa, J. 1960. Genetic structure of recom-binant chromosomes formed after mating inEscherichia coli K-12. Proc. Natl. Acad. Sci.U.S. 46:91-101.
17. van de Putte, P., H. Zwenk, and A. Rorsch. 1966.Properties of four mutants of Escherichia colidefective in genetic recombination. MutationRes. 3:381-392.
18. Wann, M., and T. H. Wood. 1967. Radiationmodification and kinetics of genetic integrationin E. coli. (Abstract) Radiation Res. 31:628.
19. Wood, T. H. 1967. Genetic recombination inEscherichia coli: Clone heterogeneity and thekinetics of segregation. Science 157:319-321.
VOL. 96, 1968 659
on May 15, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
Related Documents











