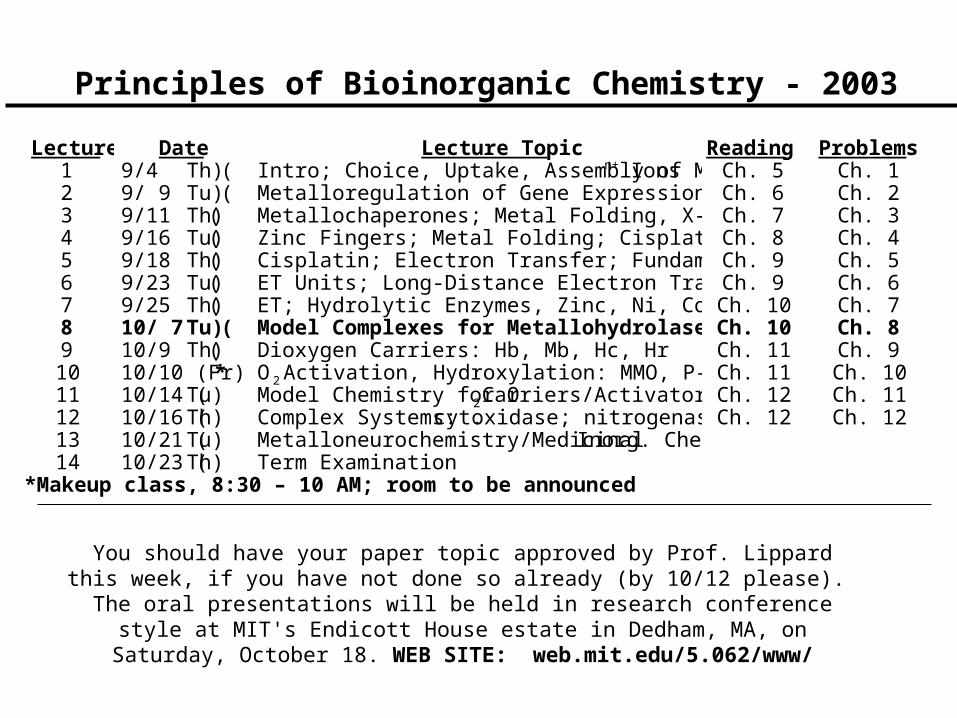
Principles of Bioinorganic Chemistry - 2003 Lecture Date Lecture Topic Reading Problems 1 9/4 ( Th) Intro; Choice, Uptake, Assembly of M n+ Ions Ch. 5 Ch. 1 2 9/ 9 ( Tu) Metalloregulation of Gene Expression Ch. 6 Ch. 2 3 9/11 ( Th) Metallochaperones; Metal Folding, X- Ch. 7 Ch. 3 4 9/16 ( Tu) Zinc Fingers; Metal Folding; Cisplat Ch. 8 Ch. 4 5 9/18 ( Th) Cisplatin; Electron Transfer; Fundam Ch. 9 Ch. 5 6 9/23 ( Tu) ET Units; Long-Distance Electron Tra Ch. 9 Ch. 6 7 9/25 ( Th) ET; Hydrolytic Enzymes, Zinc, Ni, Co Ch. 10 Ch. 7 8 10/ 7 ( Tu) Model Complexes for Metallohydrolase Ch. 10 Ch. 8 9 10/9 ( Th) Dioxygen Carriers: Hb, Mb, Hc, Hr Ch. 11 Ch. 9 10 10/10 (Fr) * O 2 Activation, Hydroxylation: MMO, P- Ch. 11 Ch. 10 11 10/14 ( Tu) Model Chemistry for O 2 Carriers/Activator Ch. 12 Ch. 11 12 10/16 ( Th) Complex Systems: cyt. oxidase; nitrogenas Ch. 12 Ch. 12 13 10/21 ( Tu) Metalloneurochemistry/Medicinal Inorg. Chem 14 10/23 ( Th) Term Examination *Makeup class, 8:30 – 10 AM; room to be announced You should have your paper topic approved by Prof. Lippard this week, if you have not done so already (by 10/12 please). The oral presentations will be held in research conference style at MIT's Endicott House estate in Dedham, MA, on Saturday, October 18. WEB SITE: web.mit.edu/5.062/www/

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript
Principles of Bioinorganic Chemistry - 2003
Lecture Date Lecture Topic Reading Problems1 9/4 (Th) Intro; Choice, Uptake, Assembly of Mn+ Ions Ch. 5 Ch. 12 9/ 9 (Tu) Metalloregulation of Gene Expression Ch. 6 Ch. 23 9/11 (Th) Metallochaperones; Metal Folding, X-linkingCh. 7 Ch. 34 9/16 (Tu) Zinc Fingers; Metal Folding; Cisplatin Ch. 8 Ch. 45 9/18 (Th) Cisplatin; Electron Transfer; Fundamentals Ch. 9 Ch. 56 9/23 (Tu) ET Units; Long-Distance Electron Transfer Ch. 9 Ch. 67 9/25 (Th) ET; Hydrolytic Enzymes, Zinc, Ni, Co Ch. 10 Ch. 78 10/ 7 (Tu) Model Complexes for MetallohydrolasesCh. 10 Ch. 89 10/9 (Th) Dioxygen Carriers: Hb, Mb, Hc, Hr Ch. 11 Ch. 910 10/10 (Fr)* O2 Activation, Hydroxylation: MMO, P-450, R2Ch. 11 Ch. 1011 10/14 (Tu) Model Chemistry for O2 Carriers/Activators Ch. 12 Ch. 1112 10/16 (Th) Complex Systems: cyt. oxidase; nitrogenase Ch. 12 Ch. 1213 10/21 (Tu) Metalloneurochemistry/Medicinal Inorg. Chem.14 10/23 (Th) Term Examination
*Makeup class, 8:30 – 10 AM; room to be announced
You should have your paper topic approved by Prof. Lippard this week, if you have not done so already (by 10/12 please). The oral presentations will be held in research conference style at MIT's Endicott House estate
in Dedham, MA, on Saturday, October 18. WEB SITE: web.mit.edu/5.062/www/

Hydrolytic Enzymes, Zinc and other Metal Ions
PRINCIPLES:
•M(OH)n+ centers supply OH- at pH 7 by lowering water pKa
•Mn+ serves as general Lewis acid, activating substrates•Rate acceleration occurs by internal attack within coord. sphere•Protein side chains greatly assist assembly of transition state•Carboxylate shifts can occur, especially at dimetallic centers•Electrostatic interactions predominate•Non-redox active metal ions often but not universally used
Illustrating the Principles:
•Carboxypeptidase, carbonic anhydrase - delivering hydroxide•Alcohol dehydrogenase: an oxidoreductase•Dimetallic metallohydrolases: are two metals better than one?

Carboxypeptidase A: A Hydrolytic Zinc Enzyme
Reaction catalyzed:
R–CH–C(O)–NH–R’
NH2R’’
R–CH–CO2- + +NH3–R’
NH2R’’Cleaves C-terminal peptide bonds; prefers aromatic residues.
Active site contains a single catalytic zinc, essential for activity. The glutamate can undergo a carboxylate shift. Thermolysin has a similar active site; it is an endopeptidase.

Carboxypeptidase A structure with the inhibitor glycyl-L-tyrosine bound at the active site. Note hydrogen bonds to key residues in the active site that position the substrate moiety for bond scission.

Catalytic Mechanism for Carboxypeptidase ASummary of events:1. Substrate binds; orients by the terminal carboxylate.2. Deprotonate bound H2O.3. Polarize scissile bond by Arg127.4. Bound OH- attacks peptide C(O).5. Form tetrahedral transition state.6.Lose 2 peptide fragments and recycle the enzyme.
Principles illustrated:1. Zinc serves as template.2.Metal supplies cleaving reagent, OH-, and organizes key groups.3. Chemistry achieved at neutral pH! Kcat ~ 100 s-1 .

Carbonic Anhydrase, the First Known Zn Enzyme
Reaction catalyzed:
CO2 + H2O H2CO3 ~ 106 s-1

Note: Rate 10-2 s-1 at pH 7; kf 106 s-1 in active site.Paradox: The reverse reaction is diffusion controlled, with kr ~ 1011 M-1 s-1
Thus kf ≤ 104 s-1. So how can the turnover be 106 s-1 ? Answer: Facilitated diffusion of protons by buffer components bound to the enzyme.
PZn(OH2)2+ PZn(OH)+ + H+Keq = 10-7 M = kf/kr
Carbonic Anhydrase

Possible Carbonic Anhydrase Mechanism
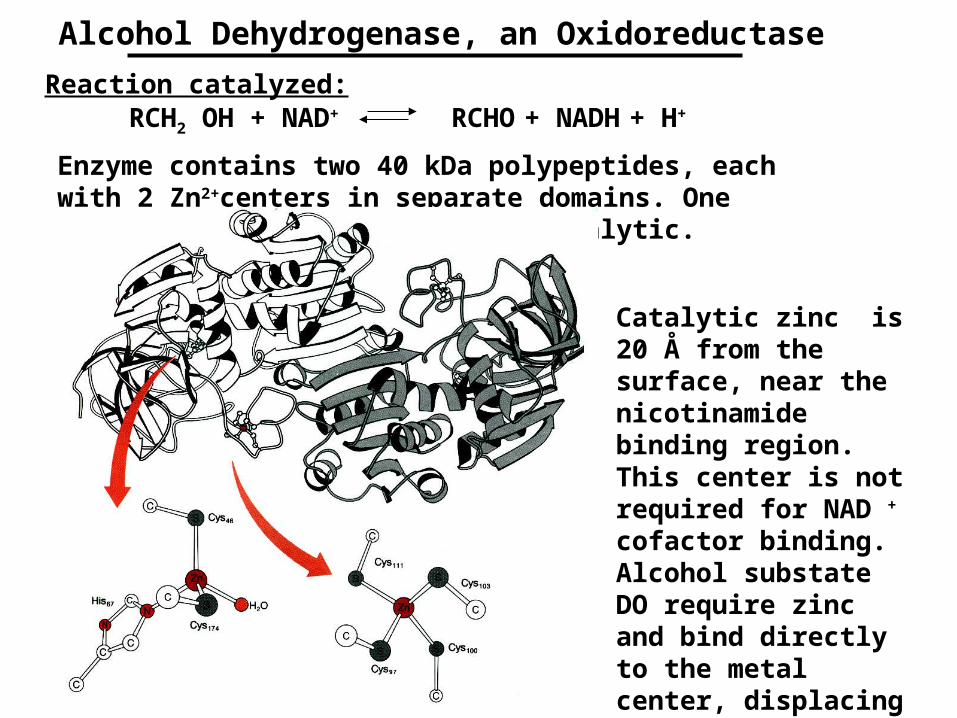
Alcohol Dehydrogenase, an Oxidoreductase
Reaction catalyzed:RCH2 OH + NAD+ RCHO + NADH + H+
Enzyme contains two 40 kDa polypeptides, each with 2 Zn2+centers in separate domains. One zinc is structural, the other catalytic.
Catalytic zinc is 20 Å from the surface, near the nicotinamide binding region. This center is not required for NAD + cofactor binding. Alcohol substate DO require zinc and bind directly to the metal center, displacing the coordinated water.

Schematic Diagram
NAD+ binding to the active site of LADH, with specific, well-positioned amino acid side chains holding it in place. Ethanol is shown bound to the zinc, displacing water. The system is set to undergo catalysis.

Hydride Transfer Mechanism
N
H2N O
R' +
OZn
H
RH
H2O
N
H2N O
R' ..
H
H
+ RCHO +
H2OZn
Note hydride transfers from -C of alcohol to nicotinamide ring.

Dinuclear Metalloenzymes
Peptide hydrolases: Methionine aminopeptidase (Zn2 or Co2)Leucine aminopeptidase (Zn2)
Phosphoester hydrolases:Ser/Thr phosphatases (Fe/Zn or Fe/Fe)Alkaline phosphatase (Zn2)Nuclease P1 (Zn2)Inositol Monophosphatase (Mg2)RNase (Mn2 and Mg2)DNA polymerase I (Mg2)
Other metallohydrolases:Arginase (Mn2, Co2)Urease (Ni2)β- (Lactamase Zn2)
(Xylose isomerase Mg2):Isomerase
- Redox active dinuclear:Metalloenzymes
(Methane monooxygenase Fe2) (Tyrosinase Cu2)
(Catalase Mn2)

Alkaline Phosphatase; a Dizinc(II) Center Activates the Substrate
1. The substrate binds to the dizinc center; a nearby Arg also helps activate it.2. A serine hydroxyl group attack the phosphoryl group, cleaving the ester. The phosphate is transferred to the enzyme, forming a phosphoryl-serine residue.3. Hydrolysis of this phosphate ester by a zinc-bound hydroxide com-pletes the catalytic cycle.
This mechanism is supported by studies with chiral phosphate esters (ROP18O17O16O)2-; there is no net change in chirality at phoshorus.
1.
2.
3.
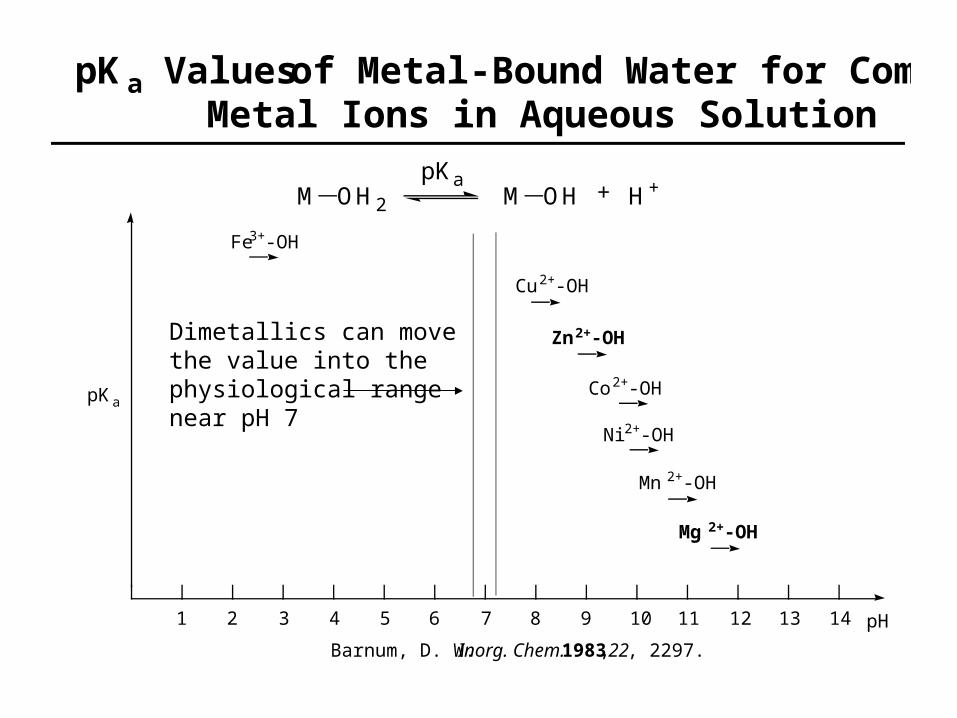
pKa Values of Metal-Bound Water for CommonMetal Ions in Aqueous Solution
pH1 2 3 4 5 6 7 8 9 10 11 12 13 14
Fe3+-OH
Cu2+-OH
Zn2+-OH
Co2+-OH
Ni2+-OH
Mn2+-OH
Mg2+-OH
M OH2 M OH H+pKa
pKa
Barnum, D. W. Inorg. Chem. 1983, 22, 2297.
+
Dimetallics can move the value into the physiological range near pH 7

MB
OC
O
MA
R
N S
MB
OC
O
MA
R
S
MB
OC
O
MA
R
N S
MB
OC
O
MA
R
N SMB
OC
O
MA
R
N S
Modes of Substrate (S) Attack by an Activated Nucleophile (N) at a Carboxylate-Bridged Dimetallic Center
A EB C D
N:: :: :
Advantages of Carboxylate-Bridged Dimetallic Centersin Chemistry and Biology

O
C
O-
H2C
C
H
H3N COO-
O
C
O
Carbonate
CH2H2C
C
H
H3N COO-
Biologically available carboxylates:
O
C
O
OH
O
C
O-
NHH2C
C
H
H3N COO-
Aspartate (Asp) D
Carboxylate Ligation in Metalloproteins and the Carboxylate Shift
Glutamate (Glu) E
CH2
H2C
-
Lys* Carbamate
-
CH2
Carbonate is encountered in transferrinLys* is found in urease and rubisco

C
O
O
R
R
CO
NM
OOC
O R
N
C
O
O
R
R
CO
N
MO O
COR
NM
O
OR
M M
O O
R
M M
M M
O
O
R
Rardin, Bino, Poganiuch, Tolman, Liu, LippardAngew. Chem. Int. Ed. Engl., 102, 812 (1990)
Chemistry Leading to the Definition of the Carboxylate Shift
[M3II(O2CCH3)6(BIPhMe)2]
M = Mn, FeM(O2CCH3)2 + BIPhMe methanol
"This movement, which we term 'the carboxylate shift,'may be of general importance in the active site structuresof metalloproteins."

Note syn/anti lonepair traversals
The Expanded Carboxylate Shift
M
O
M
OCR
M
OC
O
M
R
MO
M
CR O
O
M
M
OO
M
M
CR O
O
M
C
C
R
O R
M
OM
CR
O
MO
M
OOC
R
M
O
M
CR O
CR O
M MM

The Active Sites of Selected Dinuclear MetalloenzymesCatalyzing Hydrolysis of Biological Substrates
FeO
Zn
HO
H2O
O
H2O
NN
N
O
HN
NH
NH
His92
His281
His199
OAsp90
O
Asp118
H2N
Asn150
Calcineurin
Zn Zn
HO
ON
N
N
O
HN
NH
NH
His6
His60
His116
O
Asp45
OO
Asp122
NH2
Trp1
Nuclease P1
Zn Zn
HO
O
N
N
O
HN
NH
His97
His156
O
Glu152
OO
Asp117
O
Aminopeptidase
Asp179
Zn Zn
HO
N
N
N
O
HN
NH
NH
His230
His57
His55
O
Asp301
OO
N
Phosphotriesterase
NHN
His201
Lys169
H
+H3N NH
NH2
CO2-
NH2+
+H3N NH3+
CO2- H2N NH2
O+
L-Ornithine
H2O +
L-Arginine Urea
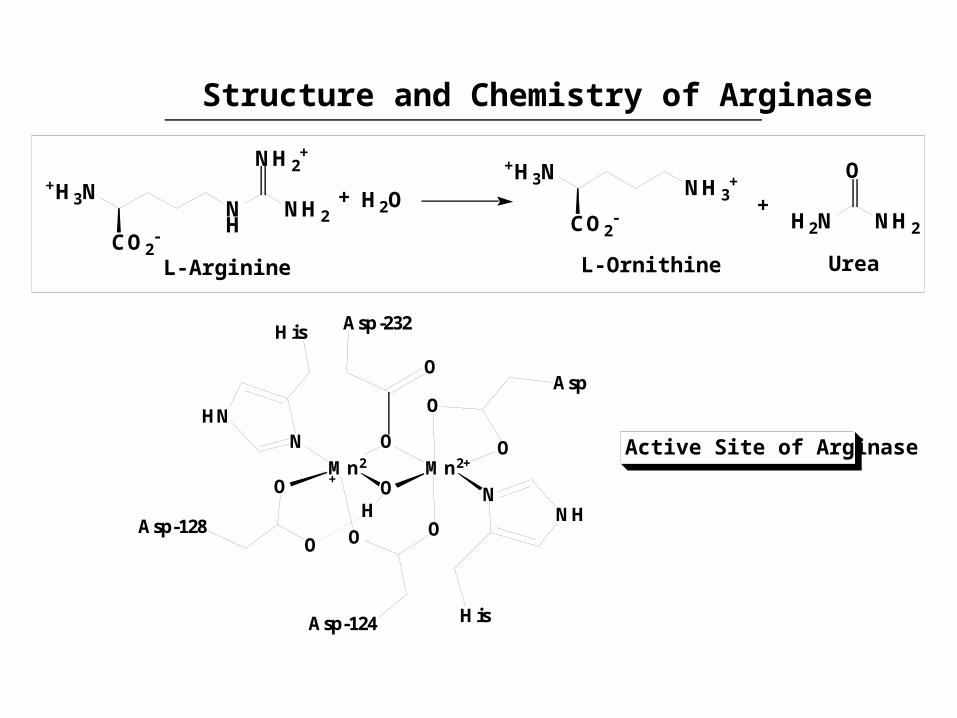
Structure and Chemistry of Arginase
Mn2+ O
Mn2+
O
O
O
Asp-232
O O
Asp-124
O
O
Asp
OAsp-128
NNH
His
NHN
His
H
Active Site of Arginase

NH
NH2
NH2
O
O
Mn2+ OMn2+
NH
NH2
NH2
O
O
OMn2+ Mn2+
H
NH2
O NH2
NH2
O
OMn2+
Mn2+
Mn2+
H2O
Mn2+
III
L-Ornithine
II
IV
H2O
Urea
H+
L-Arginine
Postulated Catalytic Mechanism for Arginase
I O OO
OH
OO
Asp 124
Asp 128
Christianson, 1996
Principles illustrated: the dimetallic affords hydroxide; the substrate is positioned by residues in the active site; the dimetallic stabilizes the urea leaving group; redox inactive metal; electrostatics

The Dinickel(II) Metalloenzyme Urease
History of Urease
1926, Sumner crystallizes urease
1975, Blakeley and Zerner discover that urease is a dinickel enzyme
1995, Hausinger and Karplus determine X-ray structure; unusual active site
Ni Ni
N
N
N
N
N
NN
N
N
OO
O
O
O
Urea Hydrolysis
H2N
O
NH2 H2N
O
OH
H2Ourease + NH3
NH3 + H2O

N
NH
N
HN
NHN
NHN
Ni1O O
Ni2
HN
Lys220*
OH(2)OH2 O
H2OO Asp363
His137
His139
Figure 1. Barrios & Lippard
NNH
N
HN
NHN
NHNNi1
O O
Ni2
HNLys220*
OO ONH2
O Asp363
His137
His139N
NH
N
HN
NHN
NHNNi1
O O
Ni2
HNLys220*
SHO O
O Asp363
His137
His139
P
NH2His249
His275His275
His249
His275
His249
Native and Inhibited Urease from B. Pasteurii
Benini et. al. Structure 1999, 7, 205-216.Benini et. al. JBIC 1998, 3, 268-273.
Native urease, 2.0 Å resolution
β- , Mercaptoethanol inhibited urease1.65 Å resolution
, 2.0 DAP inhibited urease Å resolution

NiNi
OH
NiNi
OH O
NH2H2N
NiNi
O O
NH2
O
NH2H2N
Proposed Mechanism of Urea Hydrolysis
O
NH2H
O
NH2
O
NHCH3H2N
O
NHOHH2N
O
NHOHHOHNOther urease substrates:
H2O
CO2 + (2)NH3

NiNi
OH
NiNi
OH O
NH2H2N
NiNi
OH2
O
NH2H2N
O
C
N
Alternative Mechanism of Urea Hydrolysis
H2O
CO2 + (2)NH3

PZn(OH2)2+ PZn(OH)+ + H+Keq = 10-7M = kf/kr
Metallo-β-lactamases, an Emerging Clinical Problem
Zn1
H162
H101
H99
HO
Zn2OH2
C181
D103
H223Zn1
H149
H86
H88
Zn2
H210
C168
OH
OO
D90
H2OZn1
H86
H84
H160
HO
Zn2H225
H2O
D88
H89
S185
N
S
C
OCOOH
R'
R" HN
SCO
COOH
R"
R'
OH
Bacillus cereus Stenotrophomonas maltophiliao
Zn Zn, 3.4 Å...Zn Zn, 3.5 and 4.4 Å... Zn Zn, 3.5 Å...
H2OReaction Catalyzed:
Active Sites:
Bacteroides fragilis
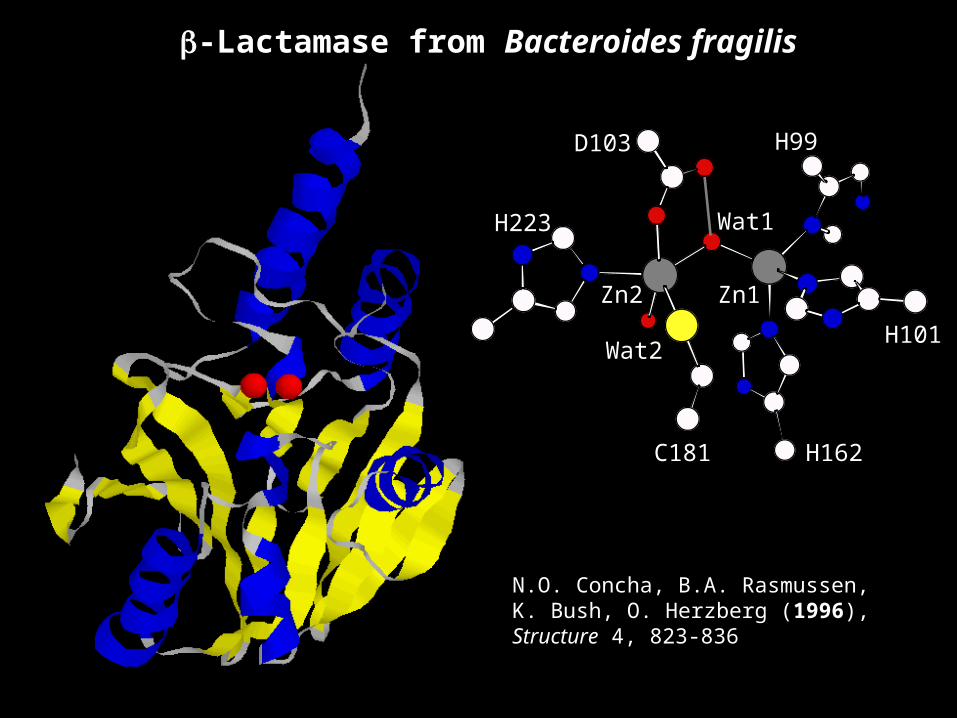
Zn2
Wat2
C181
Wat1
Zn1
H99
H101
H162
D103
H223
N.O. Concha, B.A. Rasmussen, K. Bush, O. Herzberg (1996), Structure 4, 823-836
β-Lactamase from Bacteroides fragilis
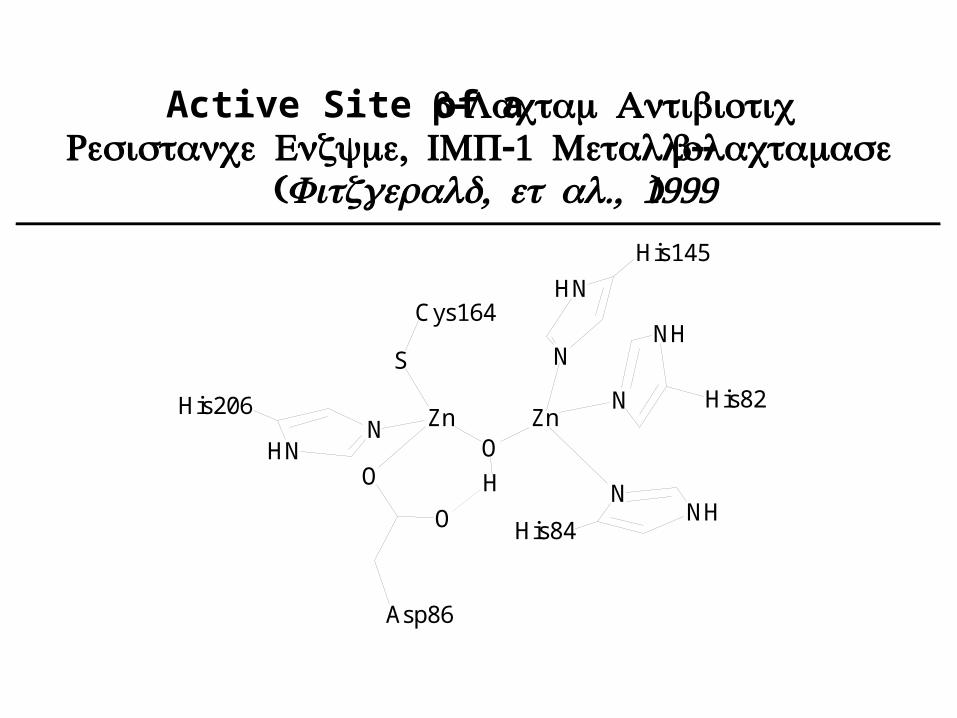
Active Site of a β- Lactam Antibiotic , -1 -Resistance Enzyme IMP Metalloβ-lactamase( , ., 1999Fitzgerald et al )
Zn ZnO
N
N
N
HN
HN
NH
His206
O
O
Asp86
His145
His84
N
NH
His82
H
S
Cys164

Zn Zn
N
S
O
O
O
R1
R2
NH
S
OH
O
O
R1
R2OH
HO
O O
Possible Mechanism for Metallo-β-lactamases
S
ONH
N
S
NO2
NO2
COO-O
SNH
O
O
OH HN
S
COO-
NO2
NO2
400 nm 496 nm
1
4
5 6
910
12
H2O
nitrocefin: a substrate for investigating the mechanism
Zn Zn
N-
S
O
O
O-
R1
R2
-O
O O
blue intermediate

•Both mono- and dimetallic centers lower the pKa value of bound water, allowing hydroxide to be delivered at pH 7.
•Coordination of the leaving group portion of the substrate to a metal ion activates the substrate for nucleophilic attack.
•Residues not coordinated but in the second coordination sphere can participate directly (serine in phophatases) or indirectly (arginine in alcohol dehydrogenase) in substrate attack, orientation, and/or activation.
•Carboxylate shifts facilitate substrate binding, activation.
•Redox inactive metal ions (Zn2+, Ni2+, Mn 2+, Co2+) preferred.
Summary - Points to Remember
Related Documents




![[Lucy Lippard and John Chandler] Dematerialization](https://static.cupdf.com/doc/110x72/563db86f550346aa9a93a929/lucy-lippard-and-john-chandler-dematerialization.jpg)






