Primary succession trajectories on a barren plain, Mount St. Helens, Washington Roger del Moral, Jason M. Saura & Jennifer N. Emenegger Abstract Questions: Have predictable relationships between environmental variables and vegetation developed in primary succession following a volcanic eruption? Has the rate of succession changed? Have vegetation trajectories converged or diverged? Location: The Abraham Plain of Mount St. Helens, Washington, USA (46112 0 42 00 N, 122108 0 27 00 W, ele- vation 1360 m), was sterilized in 1980 by a blast, scoured by lahars and buried by pumice. Method: We monitored 400 100 m 2 contiguous per- manent plots annually (1988–2008), and classified each plot from every year into ten community types (CTs). We characterized the terrain by topography and surface features. Redundancy analysis assessed relationships between vegetation and possible ex- planatory variables, which included sample location. We used detrended correspondence analy- sis (DCA) to assess successional rates and trends. Results: Relationships between species composition and explanatory variables were only significant after 1996, when position and presence of rills became significant. By 2006, explained variation remained low (13%) but significant. Species accumulated slowly, restricted by stress and isolation. Changes in mean DCA position slowed. Composition shifted from pioneer to persistent species and vegetation became more stable with time. Species accumulated for two decades and then stabilized, while cover has continued to increase. Diversity increased and then declined slightly as dominance developed and pio- neer species became less common. Conclusions: We demonstrate weak but increasingly predictable trends in species composition using en- vironmental variables. The rate of succession slowed and trajectories formed a reticulate network of transitions dominated by divergence. Convergence was not evident because vegetation responded dis- tinctively to minor topographic features that allowed alternative stable communities to develop. Keywords: Community assembly; Convergence; Di- vergence; Permanent plots; Rate of succession; Redundancy analysis; Restoration; Stochastic as- sembly; Succession network. Nomenclature: U.S.D.A. NRCS (2009; Appendix S1). Introduction One goal of the study of succession is to assess how vegetation develops in response to explanatory environmental variables. Studies of succession in severe habitats suggest that the control of succession shifts from stochastic events to predictable causes (Baasch et al. 2009), but this shift is poorly under- stood and rarely documented directly. Several succession trajectories are probable during early primary succession when competitive interactions are weak. Early in primary succession, landscape factors that influence dispersal may be the only predictors of plant patterns (Prach & R ˇ ehounkova´ 2006).This can produce variable patterns in composition with respect to topography (Felinks & Wiegand 2008). Rarity of seedling establishment and extreme weather events can obscure predictable pattern. Therefore, variability of early primary succession vegetation is expected (Robbins & Matthews 2009) because similar sites that receive different colonists often follow multiple trajectories. While the response of a species to one environ- mental factor might be predictable, vegetation patterns are less tractable due to complex, inter- dependent species responses to environmental factors. Variables that are often related to species patterns include soil properties that favour species differentially, favourable microsites and biotic in- teractions (del Moral 2009a). The effects of such factors should increase with time because, as dis- persal limitations ease, more biomass can produce stronger interactions and relationships between to- pography, or nurse plants can become more evident (but see Walker et al. 2006). A successional trajectory describes vegetation change; these were once thought inevitably to be linear, convergent and predictable (Pickett et al. del Moral, R. (corresponding author, [email protected]), Saura, J. M. ([email protected]) & Emenegger, J. N. (jne2 @u.washington.edu): Department of Biology, Box 351800, University of Washington, Seattle, Washington, USA. Journal of Vegetation Science 21: 857–867, 2010 DOI: 10.1111/j.1654-1103.2010.01189.x & 2010 International Association for Vegetation Science

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Primary succession trajectories on a barren plain Mount St Helens
Washington
Roger del Moral Jason M Saura amp Jennifer N Emenegger
AbstractQuestions Have predictable relationships betweenenvironmental variables and vegetation developedin primary succession following a volcanic eruptionHas the rate of succession changed Have vegetationtrajectories converged or divergedLocation The Abraham Plain of Mount St HelensWashington USA (4611204200N 12210802700W ele-vation 1360m) was sterilized in 1980 by a blastscoured by lahars and buried by pumiceMethod We monitored 400 100m2 contiguous per-manent plots annually (1988ndash2008) and classifiedeach plot from every year into ten community types(CTs) We characterized the terrain by topographyand surface features Redundancy analysis assessedrelationships between vegetation and possible ex-planatory variables which included samplelocation We used detrended correspondence analy-sis (DCA) to assess successional rates and trendsResults Relationships between species compositionand explanatory variables were only significant after1996 when position and presence of rills becamesignificant By 2006 explained variation remainedlow (13) but significant Species accumulatedslowly restricted by stress and isolation Changesin mean DCA position slowed Composition shiftedfrom pioneer to persistent species and vegetationbecame more stable with time Species accumulatedfor two decades and then stabilized while cover hascontinued to increase Diversity increased and thendeclined slightly as dominance developed and pio-neer species became less commonConclusions We demonstrate weak but increasinglypredictable trends in species composition using en-vironmental variables The rate of succession slowedand trajectories formed a reticulate network oftransitions dominated by divergence Convergencewas not evident because vegetation responded dis-tinctively to minor topographic features thatallowed alternative stable communities to develop
Keywords Community assembly Convergence Di-vergence Permanent plots Rate of successionRedundancy analysis Restoration Stochastic as-sembly Succession network
NomenclatureUSDA NRCS (2009 Appendix S1)
Introduction
One goal of the study of succession is to assesshow vegetation develops in response to explanatoryenvironmental variables Studies of succession insevere habitats suggest that the control of successionshifts from stochastic events to predictable causes(Baasch et al 2009) but this shift is poorly under-stood and rarely documented directly Severalsuccession trajectories are probable during earlyprimary succession when competitive interactionsare weak
Early in primary succession landscape factorsthat influence dispersal may be the only predictorsof plant patterns (Prach amp Rehounkova 2006) Thiscan produce variable patterns in composition withrespect to topography (Felinks amp Wiegand 2008)Rarity of seedling establishment and extremeweather events can obscure predictable patternTherefore variability of early primary successionvegetation is expected (Robbins amp Matthews 2009)because similar sites that receive different colonistsoften follow multiple trajectories
While the response of a species to one environ-mental factor might be predictable vegetationpatterns are less tractable due to complex inter-dependent species responses to environmentalfactors Variables that are often related to speciespatterns include soil properties that favour speciesdifferentially favourable microsites and biotic in-teractions (del Moral 2009a) The effects of suchfactors should increase with time because as dis-persal limitations ease more biomass can producestronger interactions and relationships between to-pography or nurse plants can become more evident(but see Walker et al 2006)
A successional trajectory describes vegetationchange these were once thought inevitably to belinear convergent and predictable (Pickett et al
del Moral R (corresponding author moraluwedu)
Saura J M (saurauwedu) amp Emenegger J N (jne2
uwashingtonedu) Department of Biology Box 351800
University of Washington Seattle Washington USA
Journal of Vegetation Science 21 857ndash867 2010DOI 101111j1654-1103201001189xamp 2010 International Association for Vegetation Science
2009) Now ecologists recognize that vegetation candiverge or follow intricate temporal braids in re-sponse to stochastic processes contingencies andlandscape factors
The vegetation of most new surfaces on MountSt Helens has developed markedly since the massiveeruption of 1980 but on the east slope vegetationremains sparse Slow primary succession on thisbarren plain offers a superb chance to evaluaterelationships between environmental factors andvegetation at different stages of development Weask has a more predictable relationship betweenenvironmental variables and vegetation developedover time has the rate of succession changed asspecies accumulate and have trajectories convergedor diverged
Methods
Study site
Abraham Plain a barren nearly level site 4 kmeast of the cone of Mount St Helens is centered at4611204200N12210802700Wmean elevation 1360m Itreceived three catastrophic volcanic impacts in rapidsuccession on 18051980 The lateral blast removedall soil and melted ice fields to produce massive la-hars that scoured the site (Swanson amp Major 2005)Pumice then smothered the landscape Pumice rockshave decomposed to gravel and erosion has carvedgullies and rills to create protected microsites Iso-lation from surviving vegetation few animaldispersal vectors and winds that direct seeds awayfrom barren sites all restricted colonization Estab-lishment was constrained by drought stress andinfertility
Vegetation sampling
Errors using chronosequence approaches canaccrue when multiple trajectories remain un-recognized Using permanent plots mitigates thepotential for such errors (Johnson amp Miyanishi2008) In 1988 we established 400 contiguous10m10m plots (a 1040 grid) We estimated spe-cies cover with this index 15o6 individuals 25 6to 20 individuals 35420 individuals or cover of025m2 to 05m2 45 cover 405m2 to 1m25541m2 to 2m2 6542m2 to 4m2 7544m2 to8m2 48m2 recorded directly (Wood amp del Moral1988) R del Moral determined cover from the ver-tical projection of the canopy of each speciesannually between 1988 and 2008
Explanatory variables
We estimated percentage cover of topographic(smooth rill and gully) and surface (rock pumiceand sand) features in each plot Rills are narrowwith gentle slopes and have not exposed the originalsurface Gullies are more than 1-m wide with steepslopes and have reached the original surface Cate-gories were as follows For smooth 05o20cover 15 21ndash50 cover 25 51ndash90 cover35490 cover For rills 05 none 15 1ndash5cover 25 6ndash10 cover 35410 cover For gul-ly 05 none 15 1ndash10 25 11ndash20 cover35420 cover For rocks 05 none 15 1ndash2cover 25 3ndash5 cover 3545 cover For pu-mice 05o80 cover 15 81ndash90 cover 25 91ndash97 cover 35497 cover For sand 05 0ndash3cover 15 4ndash6 cover 25 7ndash10 cover35410 cover Position was defined by x (northndashsouth) and y (eastndashwest) grid coordinates
Analyses
We calculated species richness plot cover indexSimpsonrsquos dominance [D5 1Spi
2] and Shannondiversity (H05 [Spi log pi]) where pi is the pro-portion of the cover index represented by the ithspecies from the cover index of each species (MjMSoftware Design Gleneden Beach OR US) Weclassified all plots in each year into community types(CTs) with flexible sorting (an effective space-con-serving group-linkage method that employsb5 025 to limit chaining) using the Euclideandistance between samples (MjM Software Design)The 6464 samples formed a dendrogram that we di-vided into 10 CTs recognized when members sharedat least 50 of the information Large differences inspecies richness distort dendrograms (Clifford1976) so divisions occurred at different fusion le-vels We used multiple response permutationprocedures (MRPP MjM Software Design) to as-sess validity of the CTs MRPP calculates aweighted mean within-group distance and a T-valuethat describes the distinctiveness of the groups A(0 to 10) by describing group homogeneitycompared to a random classification We formed sixhabitat types (HTs) from the topographic and sur-face variables using the same classification approachas for species
Recognizing that succession has encompassedonly early primary succession we divided the CTsinto four stages For convenience these are pioneerearly mid- and late seral The categories were based
858 del Moral Roger et al
on the dispersal growth form and longevity ofdominant species (Appendix S1)
After 1990 annual species turnover was low solinear methods could be applied (Legendre amp An-derson 1999) We used redundancy analysis (RDALeps amp Smilauer 2003 CANOCO DLO-Agri-cultural Mathematics Group Wageningen NL) toexplore the relationship between species composi-tion (index values) and explanatory variables in1990 1993 1996 1999 2003 2006 and 2008 Speciescomposition in each of these years was regressedwith explanatory variables to create fitted speciesscores We applied principal components analysis tothese values to produce canonical plot scores RDAthen formed linear equations to predict the positionof plots We assessed variation by comparing theresult to a null model (2000 random trials) The re-gression coefficient of each variable with each RDAaxis estimated how well variables predicted plot po-sition determined by a t-value (df5 400 samples ndash 8variables ndash 15 391) The surplus species variationsuggested the importance of unmeasured variablessampling error and stochastic effects
Spatial pattern of CTs was determined by com-paring the inter-plot distances of a given CT to thedistances between N random plots and the nearestplot of that CT Adjacent plots were 0 distance lar-ger spacings were calculated by Euclidean distancein grid units The observed mean between-plot dis-tance was compared by t-test to that determinedfrom the hypothesis of random distribution
We used Statistix 9 (Analytical Software Talla-hassee FL US) to conduct statistical analysesComparisons of cover index values among species inthe CTs and among DCA scores of CTs were madewith one-way ANOVA or repeat measures ANO-VA as appropriate each followed by a Bonferronitest of differences We used w2 association tests torelate community patterns to surface patterns Weused Axum 7 (Mathsoft Insightful CorporationSeattle WA US) to produce graphs and generaterandom numbers
Results
Vegetation
Succession on Abraham Plain has been leisurelyBy 2009 vegetation remained sparse (Fig S1) andpercentage cover was one-quarter of that observedon a similar grid on the north side of Mount StHelens with three-quarters as many species (delMoral 2009b)
The composition of each of the 10 CTs was dis-tinctive (MRPP A5 0278 T5 1772 Po00001Table 1) Each pair-wise comparison by MRPPshowed that CTs differed significantly Pioneer CTsvanished by 2001 (Fig 1a) they had lower richness(Fig 1b) and cover indices (Fig 1c) than did persis-tent CTs H0 (Fig 1d) increased with time and washigher in persistent CTs Each plot supported amean of 495 106 CTs between 1988 and 2008We characterize CT composition below The num-ber of plots within which the CT ever occurred is inparentheses
There were two pioneer community types CT-A (Anaphalis-Chamerion-Hypochaeris n5 397 of400 plots) had sparse cover of a few pioneer specieseach with long-distance wind dispersal herbaceouslife form and short life span (Appendix S1) As coverincreased plots developed into other CTs CT-B(Lupinus lepidus n5 11) established first in sevenplots near surviving vegetation and was character-ized by L lepidus with sparse Anaphalis and Carexmertensii We recognized three early seral CTs thatalways developed from other CTs and had morespecies and cover than did pioneer CTs These in-cluded a few species that are more persistent Plotscharacterized as CT-C (Anaphalis n5 107) usuallydeveloped from CT-A plots CT-D (Anaphalis-Cis-tanthe-Agrostis) n5 319) was the main nexusthrough which pioneer CTs developed It had lowerrichness and cover than did persistent CTs CT-E(Anaphalis-Chamerion n5 208) had more Hy-pochaeris Luetkea andRacomitrium than did CT-D
Transitions to persistent CTs involved increas-ing species richness and cover and greaterdominance by longer-lived often woody speciesWe characterized two persistent CTs as mid-seralMost common species have less effective dispersalthan those in pioneer CTs The richness of CT-F(Penstemon-Agrostis-Cistanthe n5 338) peakedwhen it contained both pioneer and seral speciesH0
declined as the vegetation matured CT-G (Pen-stemon-Agrostis-Juncus n5 75) retained Anaphalisand developed more of the persistent species (egLuetkea and Racomitrium) than did CT-F Threelate-seral CTs may represent divergent trajectoriestowards mature vegetation CT-H (Penstemon-Agrostis-Salix n5 266) also had Juncus Cistantheand Luetkea Common species are long-lived andseveral are shrubs CT-H had more species highercover and greater diversity than other CTs It hadchanged little since 2004 CT-I (Penstemon-Agrostis-Cistanthe n5 125) also supported Juncus and mos-ses Richness cover and H0 were intermediateamong persistent CTs CT-J (Penstemon-Agrostis-
Primary succession trajectories 859
Cistanthe-Juncus n5 145) also had substantialLuetkea and Racomitrium Pioneer species were un-common and taller woody species (Abies spp andSalix) absent Richness cover and diversity were allleast of the persistent CTs
Species patterns
Many species increased in cover (Fig 2 Fig S2)during the study but cover of pioneer species de-clined after the mid-1990s In 2008 1093 speciesplot were wind-dispersed while only 011 speciesplot were animal-dispersed (Appendix S1) Pioneerspecies have effective long-distance dispersal me-chanisms and while seral species are also wind-
dispersed their dispersal abilities are more limited(Fuller amp del Moral 2003) Penstemon and Agrostispallens expanded to occupy all plots while Juncusparryi and Cistanthe umbellata occurred in mostLuetkea increased steadily but by 2008 it appearedto be stable Salix increased gradually and spor-adicallyRacomitrium occurred in four plots in 1988while Polytrichum did not occur until 1995 Carexmertensii and C microptera were widely distributedand occasionally abundant both occur on halfthe grid Saxifraga once common declined in2008 but this may be associated with dry summerconditions
Relative cover clearly shows the contrast be-tween pioneer and persistent species (Fig 3 Fig S3)
Table 1 Structure and mean cover index in 10 community types (CTs) Numbers in parentheses after species are occurrences(plotsyears) Values in bold indicate dominant species (high cover index) in that CT Each structural variable differedsignificantly among CTs (one-way ANOVA Po0001 values with same superscript fall within the same group as determinedby Bonferroni comparisons) Species cover index means also differed significantly (ANOVA Po0001) but patterns weretoo complex to display grouping patterns CTs and species were each arranged in order of their DCA scores
Structure CT-A CT-B CT-C CT-D CT-E CT-F CT-G CT-H CT-I CT-J
Richness 37a 31a 53b 95d 89c 109e 125f 127f 111e 91cd
Cover (sum indexscores)
52a 38a 70a 159c 136b 242d 321g 298f 281f 238e
Dominance 0516a 0575a 0734b 0851cde 0840c 0871def 0890ef 0890ef 0877def 0843cd
H0 1001a 0999a 1512b 2078d 2008cd 2204e 2347f 2361f 2227e 2001c
Number of plotsyears 1558 21 108 877 444 1289 219 1334 421 495First year observed 1988 1988 1988 1990 1989 1992 1996 1994 1997 1997Last year observed 2000 1991 1998 2001 2000 2008 2008 2008 2008 2008Hypochaeris radicata
(3011)0444 0000 0491 1197 1380 0497 0365 0412 0214 0040
Anaphalis margaritacea(5784)
1557 0810 1713 3304 3459 2370 2717 2147 1337 0802
Chamerionangustifolium (4712)
0514 0191 0593 1911 1803 1442 0991 1077 0691 0513
Hieracium albiflorum(3907)
0336 0143 0398 0904 0930 0929 0753 0813 0411 0293
Lupinus lepidus (248) 0007 1667 0028 0051 0220 0076 0383 0021 0088 0036Lupinus latifolius (122) o0001 0095 0000 o0001 0047 0020 0932 0037 0000 0000Abies sp (736) 0106 0000 0120 0123 0124 0087 0032 0352 1062 0020Luetkea pectinata
(4075)0182 0095 0157 0691 0916 1389 2078 2524 1981 1432
Carex mertensii (2691) 0128 0428 0046 0379 0373 0680 0827 1249 1057 0649Saxifraga ferruginea
(1596)0045 0095 0083 0276 0253 0632 1082 1079 0456 0247
Racomitrium canescens(2498)
0072 0000 0019 0243 0943 0421 2183 1080 1064 0990
Cistanthe umbellata(4867)
0123 0048 1370 2222 0523 3199 3603 3469 4033 4853
Agrostis pallens (5060) 0294 0000 0528 1331 0738 2855 3457 3271 3924 4301
Eriogonum pyrolifolium(653)
0016 0000 0000 0071 0057 0120 0338 0281 0591 0059
Juncus parryi (4421) 0116 0191 0250 1008 0550 2066 4064 2633 3565 3465
Penstemon cardwellii(4489)
0195 0000 0278 0520 0484 3165 4489 3591 4197 4216
Salix sitchensis (1858) 0037 0000 0000 0140 0188 0216 1237 3144 0271 0073Carex microptera
(1723)0035 0095 0065 0161 0091 0412 0763 0752 0644 1372
Polytrichumjuniperinum (1133)
o0001 0000 0000 0047 0093 0182 0182 1601 2183 0168
860 del Moral Roger et al
Species such as Penstemon Agrostis Juncus parryiCarex spp and Luetkea increased across the CTswhen listed by their first appearance These speciesare long-lived with strong vegetative growth
Fig 1 Structure of CTs from 1988 to 2008 (a) Number of plots (b) Mean species richness (c) Mean cover index (d) Meandiversity index (H0)
Fig 2 Changes in mean cover index for abundant species
Fig 3 Changes in relative cover (proportion of total cov-er in the sample) of common species in the four stages ofearly primary succession The first four species are persis-tent the last two are pioneer species (defined in AppendixS1)
Primary succession trajectories 861
In contrast pioneer species soon peaked and thendeclined
Habitat relationships
We could use only position topography andsurface conditions as explanatory variables in RDAExplained variation was low but increased four-fold from 1990 to 2006 (Table 2) In 1996 we ob-tained the first significant result when the y-axis(possibly related to distance from survivors) and rillfraction became significant By 2006 smooth sur-face also contributed but topography did notinfluence the vegetation
We recognized six habitat types (Table 3) A fivepersistent CT (Table S1) by six HT w2 test was sig-nificant (w2 5 935 Po00001 df 5 20) CT-F(Penstemon-Agrostis-Cistanthe) was more commonin rills CT-G (Penstemon-Agrostis-Juncus) wasmore common in gullies CT-H (Penstemon-Agros-tis-Salix) tended to avoid smooth plots and wasmore common in plots with rills or gullies and CT-J(Penstemon-Agrostis-Cistanthe-Juncus) tended tooccur in smooth plots but avoided rills or gullies
The spatial pattern displayed by the CTs re-flected some environmental sorting (Fig 4) Nearestneighbour distances compared to the randommodel suggested that CT-F formed scattered clus-ters (mean observed distance5 142 mean randomdistance5 224 t5 225 P5 003) Some plots ofCT-G were confined to the eastern edge while oth-ers were in the southwest corner but overall it wasnot clustered (observed5 190 random5 243t5 190 P5 006) CT-H was clustered (ob-served5 015 random5 058 t5 483 Po00001)as was CT-I (observed5 052 random5 145t5 448 Po00001) concentrated on the east halfof the grid CT-J was clustered (observed5 119random5 264 t5 383 P5 0002) and con-centrated on the eastern part of the grid
Trajectories
We explored vegetation trajectories in severalways In 1988 richness the mean cover index(Fig 5) H0 and dominance (Fig S4) were leastRichness increased until 1999 after which time itvaried around 11 species per plot The cover indexcontinued to increase while H0 and dominance sta-bilized by the mid-1990s
Succession began with colonization by highlyvagile pioneers such as Anaphalis Chamerion Hier-acium and Hypochaeris Plots changed into otherCTs at variable rates While we found transitionsamong all CTs some trajectories were more typicalthan were others (Fig 6) The most likely trajec-
Table 2 Summary of redundancy analysis results usingposition topography and surface variables on the gridThe alternate plots sampled in 2007 and 2008 were ana-lysed separately Percentage variation5 how much ofspecies variation is explained by variables F-values5 sig-nificance level of Axis 1 trace5 percentage variance onAxis 1 Correlations 405
Years Percentagevariation
F-value
Trace Correlatedvariables
1990 34 707 157 ndash1993 65 128 340 ndash1996 79 178 420 y rill1999 109 331 598 y rill2003 112 338 616 y rill2006 125 390 698 y rill flat2008 122 335 654 y rill flat
Table 3 Mean value of characteristics of six habitat types (HT) HTs differed significantly for each terrain characteristic(Po00001 one-way ANOVA followed by Bonferroni tests for differences among means Values with same superscript fallwithin the same group)
Terrain HT-1 HT-2 HT-3 HT-4 HT-5 HT-6 F-value(n5 128) (n5 28) (n5 37) (n5 64) (n5 61) (n5 82)
Smooth 300a 286a 289a 192bc 198b 180c 1911Gully 005b 036b 030b 203a 026b 222a 2085Rill 015d 029cd 057c 061c 238a 117b 1141Rock 000d 100bc 132b 080c 062c 216a 1466Sand 000b 000b 019b 002b 010b 090a 306Pumice 300a 300a 200c 261b 257b 128d 1470
Fig 4 Distribution of CTs on the grid in 2008
862 del Moral Roger et al
tories proceeded from pioneer plots dominated byAnaphalis to early seral types that had accumulatedCistanthe Agrostis and Luetkea These developedfurther into mid-seral communities when Penstemonbecame dominant and Juncus became widespreador they developed directly into late seral commu-nities dominated by Penstemon and Agrostis In2008 some mid-seral CTs persisted but most plotswere in a late-seral community that had changedlittle since 2005 For each annual interval 1988 to2008 a plot could persist in a CT progress to a moredeveloped one or regress to an earlier stage Therewere 301 retrogressions and 1014 progressions The
five extant CTs persisted significantly longer thandid the five vanished CTs (Table 4)
We clarified temporal vegetation changes usingDCA (Fig 7) Total variance (l) was 159 (DCA-15 155 DCA-25 74) Turnover on DCA-1was 53 half-changes on DCA-2 it was 39 Pioneerspecies had high values on DCA-1 species with lowDCA-1 values appeared later and were persistentMean DCA-1 position of the plots changed sig-nificantly between years (r2 5 0637 Po00001Spearman nonparametric correlation r5 079Po00001) DCA-1 scores by year changed rapidlythrough 2001 (ANOVA Po00001) then slowedDCA-2 changes ceased after 1997 but ANOVA was
Fig 5 Annual changes of mean species richness and meancover index Differences (Po005) among measures de-termined by a repeat measures ANOVA followed by theBonferroni test Values with the same associated letter fallwithin the same group of values
Fig 6 Trajectories showing the number of transitionsbetween CTs Numbers in parentheses after persistent CTsare the number of plots in 2008 Many arrows transitionnear the mid-point to differentiate between transitions toand from a given CT
Table 4 Number of continuous years (standard devia-tion) for which each CT persisted Differences amongmeans are significant (one-way ANOVA Po00001 fol-lowed by Bonferroni tests for differences among meansSuperscripts indicate membership in same group) Mini-mum run is one in each case
CT Years SD Max
A 329b 135 7B 200ab 115 4C 107a 026 2D 238b 142 7E 181ab 117 7F 398c 274 10G 391c 172 6H 558d 256 10I 419cd 167 8J 378c 185 7
Fig 7 Detrended correspondence analysis of plots by CTand by year Differences (Po005) among measures de-termined by ANOVA followed by the Bonferroni testFor each set values with the same associated letter fallwithin the same group of values for DCA-1 (no data for1998 and 2002 alternate plots were sampled in 2007 and2008) The inset is a separate DCA of plots in 2008
Primary succession trajectories 863
significant (r2 5 0206 Po00001 Spearman non-parametric correlation r5 025 Po00001) Thevariation in DCA-1 scores within years declined (thelinear correlation of years vs standard deviationwas Po00001 t5 533 r2 5 0637)
The mean DCA position of the 10 CTs did notreveal a single trajectory CT-C (Anaphalis) wasmore closely associated with pioneer CTs but wasnever the first CT on a plot CT-G (Penstemon-Agrostis-Cistanthe) was aligned with late-seral CTsWe also conducted a DCA of 2008 plots (Fig 7inset) b-diversity was low with l5 097 (DCA-15
133 DCA-25 84 ca 18 HC along DCA-1)No trajectory was evident from this analysis
Discussion
There are so few long-term longitudinal studiesof early primary succession on stressful sites be-cause initially so little happens The vegetation ofthe Abraham Plain remains sparse 30 years after theeruption It has developed by interwoven succes-sional trajectories that include plots that divergeconverge regress and progress to form a braidedsuccession Neither habitat factors nor competitionappears to have provided strong filtering effects onspecies composition although unmeasured soilvariables (eg moisture and nitrogen content) andfactors related to stochastic colonization and estab-lishment (cf del Moral amp Ellis 2004) may explainsome of the remaining unexplained variation Theimmigration seed density was low (Wood amp delMoral 2000) many suitable sites appeared vacant inearly years (del Moral amp Bliss 1993) and severalspecies occupied similar sites Chance may governsuch patterns We propose that when environmentalvariables have only slight effects on species compo-sition consistent successional trajectories areunlikely to emerge Below we discuss questionsabout predictability rate and pattern of successionon Abraham Plain that are the focus of this study
Environmentndashvegetation connection
Predictable relationships between environ-mental variables and vegetation had developed by2008 but they remained low Priority effects sec-ondary disturbances elk grazing and seasonalfluctuations may affect these trajectories in the fu-ture rendering it unlikely that explained variationwould become the dominant factor before forestsdevelop
We did not use soil factors because plants es-tablish at a smaller scale than the plot size and it isimpractical to measure such factors as moisture ineven a few plots Although soils were young andhomogeneous it is likely that inclusion of soil fac-tors (eg pH and moisture) would have improvedexplanatory power based on similar studies (delMoral 2009a) explained variance might doubleTopographic surface and position factors alone diddemonstrate increasing explained variance of spe-cies pattern over time but this trend was lesspronounced than in other studies On the northslope of Mount St Helens in 1992 only spatial fac-tors explained any variation (15) but after 10 yrenvironmental factors dominated (28 del Moralamp Lacher 2005) Baasch et al (2009) produced oneof the very few primary succession studies to showthat relationships between vegetation and ex-planatory variables increased over time They toosuggested that the impact of stochastic processesdeclined over time
The spatial distribution of CTs suggested someenvironmental influence Rills provide havens forpioneers (eg Anaphalis Hieracium and Hy-pochaeris) Gullies offer protection from wind andaccumulate snow which allowed species such asAnaphalis and Salix to colonize and persist Pen-stemon and Juncus were common in more exposedsites where cover and diversity were least In suchsites species that are less stress tolerant (eg SalixPolytrichum Saxifraga and Juncus mertensianus)were uncommon while stress-tolerant species suchas Cistanthe Juncus parryi Agrostis pallens andCarex microptera reached their maximum values
Rates
Habitats with limited fertility or moisture typi-cally have low succession rates (Donnegan ampRebertus 1999 Anderson 2007) and succession onAbraham Plain has been no exception Mean annualDCA scores were in a single group of values Be-tween 1988 and 1999 these scores formed sevengroups Surrounding habitats on Mount St Helensdeveloped more rapidly On an exposed lahar on thesouth slope richness averaged 16 species and thecover index averaged 73 (del Moral 2009b) prior to2008 In contrast mean richness on the AbrahamPlain in 2008 was 108 and the mean cover index was314 The proximity to forest more fertile soils andabundance of the N-fixing Lupinus may all con-tribute to accelerated primary succession at suchsites relative to Abraham Plain
864 del Moral Roger et al
Trajectories
Succession trajectories have been discussedthoroughly (cf Leps amp Rejmanek 1991) When thevegetation of initially homogeneous sites developslocally distinctive characteristics divergence occurs(Bossuyt et al 2003) Divergent trajectories occur onmany surfaces including landslides glacier fore-lands wet slack dune vegetation floodplains andmine tailings (Walker amp del Moral 2003) Lanta ampLeps (2009) showed experimentally that differentialdispersal could promote divergence when there arepriority effects Divergence may be more likely instressful habitats when different species arrive firstand where climatic factors are highly variable(MacDougall et al 2008) Convergence describestrajectories that become more similar over timeConvergence has been assumed to result when cli-matic factors dominate and there is evidence thatwhen dispersal effects are small and competitive ef-fects are strong convergence to a single communitydoes occur (van Oijen et al 2005 Anthelme et al2007) Convergence is also more likely when thereare few alternative stages as on glacial forelands(Hodkinson et al 2003 but see Robbins ampMathews2009)
Here we show directly that early succession un-der stressful conditions can include trajectories thatdiverge converge regress and progress to form areticulate succession Our study joins a growingbody of work that demonstrates the frequency ofmultiple trajectories in early primary succession (egdel Moral 2007 Prach amp Hobbs 2008 Walker amp delMoral 2009) Forecasting trajectories and identify-ing factors that limit their development is crucial tounderstanding community assembly mechanismsOn the Abraham Plain persistent species graduallyreplaced readily dispersed pioneer species Subtlehabitat variation annual weather fluctuation andlandscape effects combined to produce communitiesrelated through a network of development and re-sponsible to topographic variation
The first colonists were species with excellentdispersal but isolation allowed for only a meagerseed rain they established pioneer communities thatsoon developed into other CTs characterized byspecies with greater persistence (cf Ejrnaes et al2006) Several alternative transitions were possiblefor most CTs suggesting that stochastic processesinitially dominated transitions Gradually persis-tent species invaded and attained dominance Minortopographic variation affected composition some-what resulting in some successional transitions thatare more common than others Of annual transi-
tions between successional stages 51 wereregressions and 170 were progressions DCA re-vealed no single trend among the CTs The two mid-seral community types were central to the transitionpatterns while the three late-seral CTs occurred indifferent directions from this centre
It is unlikely that complete convergence will oc-cur Even when trees dominate understoreyvegetation is likely to retain variation initiated dur-ing early succession and maintained by topographicvariation and priority effects The understorey offorests on the south side of Mount St Helens de-monstrates such variation (del Moral amp Ellis 2004)There is no evidence yet for a single target commu-nity
Strong assembly rules are more likely to pro-duce convergent trajectories than weak ones Whilestrong assembly rules may exist in mature vegeta-tion (see Navas amp Violle 2009) they appear weak onAbraham Plain and are more effective for functionaltypes than species One rule may be that long-livedwind-dispersed species accumulate at the expense ofshort-lived pioneers A second may be that grami-noids with rhizomatous growth (eg Agrostispallens) and prostrate shrubs (eg Penstemon) arecomplementary and can coexist indefinitely A thirdmay be that fleshy fruit-producing shrubs (eg Ru-bus spp Vaccinium spp) cannot establish undercurrent conditions Finally although seeds of con-ifers and Populus balsamifera are commonconditions do not yet favour their developmentThese rules are weak and exceptions are commonThe spontaneously developing vegetation of theAbraham Plain is constrained by isolation from po-tential colonists as well as a habitat that challengesplant growth
Acknowledgements We thank the US National Science
Foundation for funding (BSR8906544 DEB9406987
DEB0087040 and DEB0541972) and the Mount St He-
lens National Volcanic Monument for permissions
Valuable comments by J G Bishop M P Fleming Jan-
neke Hille Ris Lambers L R Walker J H Titus and two
anonymous reviewers improved the paper Field assis-
tance was provided through the years by S Anderson W
Arnesen S Bard A Coogan S Crawford K Dlugosch
A Eckert E Ellis C Fairbourn C Muerdter M Flem-
ing T Fletcher R Fuller A Grant E Jenkins M
Johns C Jones L McMullan P Moody R Robham L
Rozzell J Sandler R Sewell-Nesteruk M Spasojevic
M Tweiten S Wilson B Witte and C Wolfe This is
contribution No 60 to the Mount St Helens Succession
Project
Primary succession trajectories 865
References
Anderson KJ 2007 Temporal patterns in rates of
community change during succession American
Naturalist 169 780ndash793
Anthelme F Villaret JC amp Brun JJ 2007 Shrub
encroachment in the Alps gives rise to the
convergence of sub-alpine communities on a regional
scale Journal of Vegetation Science 18 355ndash362
Baasch A Tischew S amp Bruelheide H 2009 Insights
into succession processes using temporally repeated
habitat models results from a long-term study in a
post-mining landscape Journal of Vegetation Science
20 629ndash638
Bossuyt B Honnay O amp Hermy M 2003 An island
biogeographical view of the successional pathway in
wet dune slacks Journal of Vegetation Science 14 781ndash
788
Clifford HT 1976 Dendrograms and their
interpretation In Williams WT (ed) Pattern
analysis in agricultural science pp 96ndash109 Elsevier
Science Amsterdam NL
del Moral R 2007 Vegetation dynamics in space and
time an example from Mount St Helens Journal of
Vegetation Science 18 479ndash488
del Moral R 2009a Increasing deterministic control of
primary succession onMount St Helens Washington
Journal of Vegetation Science 20 1145ndash1154
del Moral R 2009b Mount St Helens permanent plots
and grids 1980 to present Available at http
protistbiologywashingtonedudelmoral Accessed 8
December 2009
del Moral R amp Bliss LC 1993 Mechanism of primary
succession insights resulting from the eruption of
Mount St Helens Advances in Ecological Research
24 1ndash66
del Moral R amp Ellis EE 2004 Gradients in hetero-
geneity and structure on lahars Mount St Helens
Washington USA Plant Ecology 175 273ndash286
del Moral R amp Lacher IL 2005 Vegetation patterns 25
years after the eruption of Mount St Helens
WashingtonAmerican Journal of Botany 92 1948ndash1956
Donnegan JA amp Rebertus AJ 1999 Rates and
mechanisms of subalpine forest succession along an
environmental gradient Ecology 80 1370ndash1384
Ejrnaes R Bruun HH amp Graae BJ 2006 Community
assembly in experimental grasslands suitable
environment or timely arrival Ecology 87 1225ndash1233
Felinks B ampWiegand T 2008 Exploring spatiotemporal
patterns in early stages of primary succession on
former lignite mining sites Journal of Vegetation
Science 19 267ndash276
Fuller RN amp del Moral R 2003 The role of refugia and
dispersal in primary succession on Mount St Helens
Washington Journal of Vegetation Science 14 637ndash
644
Hodkinson IA Coulson SJ amp Webb NR 2003
Community assembly along proglacial chrono-
sequences in the high Arctic vegetation and soil
development in north-west Svalbard Journal of
Ecology 91 651ndash663
Johnson EA amp Miyanishi K 2008 Testing the
assumptions of chronosequences in succession
Ecology Letters 11 419ndash431
Lanta V amp Leps J 2009 How does surrounding
vegetation affect the course of succession a five-year
container experiment Journal of Vegetation Science
20 686ndash694
Legendre P amp Anderson MJ 1999 Distance-based
redundancy analysis testing multispecies responses in
multi-factorial ecological experiments Ecological
Monographs 69 1ndash24
Leps J amp Rejmanek M 1991 Convergence or
divergence what should we expect from vegetation
succession Oikos 62 261ndash264
Leps J amp Smilauer P 2003 Multivariate analysis of
ecological data using CANOCO Cambridge
University Press Cambridge UK
MacDougall AS Wilson SD amp Bakker JD 2008
Climatic variability alters the outcome of longterm
community assembly Journal of Ecology 96 346ndash354
Navas ML amp Violle C 2009 Plant traits related to
competition how do they shape the functional
diversity of communities Community Ecology 10
131ndash137
Pickett STA Cadenasso ML amp Meiners SJ 2009
Ever since Clements from succession to vegetation
dynamics and understanding to intervention Applied
Vegetation Science 12 9ndash21
Prach K amp Hobbs RJ 2008 Spontaneous succession
versus technical reclamation in the restoration of
disturbed sites Restoration Ecology 16 363ndash366
Prach K amp Rehounkova K 2006 Vegetation succession
over broad geographical scales which factors
determine the patterns Preslia 78 469ndash480
Robbins JA amp Matthews JA 2009 Pioneer vegetation
on glacier forelands in southern Norway emerging
communities Journal of Vegetation Science 20 889ndash
902
Swanson FJ amp Major JJ 2005 Physical events
environments and geologicalndashecological interactions
at Mount St Helens March 1980ndash2004 In Dale
VH Swanson FJ amp Crisafulli CM (eds)
Ecological recovery after the 1980 eruption of Mount
St Helens pp 27ndash44 Springer New York NY US
USDA National Resource Conservation Service 2009
The Plants Data Base National Plants Data Center
Baton Rouge LA US Available at http
plantsusdagov Accessed October 15 2009
van Oijen D Hommel P den Ouden J amp de Waal R
2005 Effects of tree species composition on within-
forest distribution of understorey species Applied
Vegetation Science 8 155ndash166
Walker LR amp del Moral R 2003 Primary succession
and ecosystem rehabilitation Cambridge University
Press Cambridge UK
866 del Moral Roger et al
Walker LR amp del Moral R 2009 Transition dynamics
in succession implications for rates trajectories and
restoration In Hobbs RJ amp Suding KN (eds)
New models for ecosystem dynamics and restoration
pp 33ndash49 Island Press Washington DC US
Walker LR Bellingham PJ amp Peltzer DA 2006
Plant characteristics are poor predictors of microsite
colonization during the first two years of pri-
mary succession Journal of Vegetation Science 17
397ndash406
Wood DM amp del Moral R 1988 Colonizing plants on
the Pumice Plains Mount St Helens Washington
American Journal of Botany 75 1228ndash1237
Wood DM amp del Moral R 2000 Seed rain during early
primary succession onMount St Helens Washington
Madrono 47 1ndash9
Supporting Information
Additional supporting information may befound in the online version of this article
Fig S1 Abraham Plain Sept 1980 pumice de-posits over scoured bedrock looking north towardsthe study site
Fig S2 Abraham Plain July 2009 lookingnorthwest across the study site
Fig S3 Changes in mean cover index for lesscommon species
Fig S4 Relative cover (proportion of totalcover in the sample) of less common species in thefour stages of early primary succession The firstfour species are persistent the last two are pioneerspecies (defined in Appendix S1)
Fig S5 Annual changes of mean H0 and meanSimpsonrsquos dominance index Differences (Po005)among measures determined by a repeat measuresANOVA followed by the Bonferroni test Valueswith same associated letter fall within the samegroup of values
Table S1 Structure and species composition ofCTs found in 2008
Appendix S1 Characteristics of species en-countered on Abraham Plain grid
Please note Wiley-Blackwell is not responsiblefor the content or functionality of any supportingmaterials supplied by the authors Any queries(other than missing material) should be directed tothe corresponding author for the article
Received 9 December 2009
Accepted 14 March 2010
Co-ordinating Editor Dr Kerry Woods
Primary succession trajectories 867

2009) Now ecologists recognize that vegetation candiverge or follow intricate temporal braids in re-sponse to stochastic processes contingencies andlandscape factors
The vegetation of most new surfaces on MountSt Helens has developed markedly since the massiveeruption of 1980 but on the east slope vegetationremains sparse Slow primary succession on thisbarren plain offers a superb chance to evaluaterelationships between environmental factors andvegetation at different stages of development Weask has a more predictable relationship betweenenvironmental variables and vegetation developedover time has the rate of succession changed asspecies accumulate and have trajectories convergedor diverged
Methods
Study site
Abraham Plain a barren nearly level site 4 kmeast of the cone of Mount St Helens is centered at4611204200N12210802700Wmean elevation 1360m Itreceived three catastrophic volcanic impacts in rapidsuccession on 18051980 The lateral blast removedall soil and melted ice fields to produce massive la-hars that scoured the site (Swanson amp Major 2005)Pumice then smothered the landscape Pumice rockshave decomposed to gravel and erosion has carvedgullies and rills to create protected microsites Iso-lation from surviving vegetation few animaldispersal vectors and winds that direct seeds awayfrom barren sites all restricted colonization Estab-lishment was constrained by drought stress andinfertility
Vegetation sampling
Errors using chronosequence approaches canaccrue when multiple trajectories remain un-recognized Using permanent plots mitigates thepotential for such errors (Johnson amp Miyanishi2008) In 1988 we established 400 contiguous10m10m plots (a 1040 grid) We estimated spe-cies cover with this index 15o6 individuals 25 6to 20 individuals 35420 individuals or cover of025m2 to 05m2 45 cover 405m2 to 1m25541m2 to 2m2 6542m2 to 4m2 7544m2 to8m2 48m2 recorded directly (Wood amp del Moral1988) R del Moral determined cover from the ver-tical projection of the canopy of each speciesannually between 1988 and 2008
Explanatory variables
We estimated percentage cover of topographic(smooth rill and gully) and surface (rock pumiceand sand) features in each plot Rills are narrowwith gentle slopes and have not exposed the originalsurface Gullies are more than 1-m wide with steepslopes and have reached the original surface Cate-gories were as follows For smooth 05o20cover 15 21ndash50 cover 25 51ndash90 cover35490 cover For rills 05 none 15 1ndash5cover 25 6ndash10 cover 35410 cover For gul-ly 05 none 15 1ndash10 25 11ndash20 cover35420 cover For rocks 05 none 15 1ndash2cover 25 3ndash5 cover 3545 cover For pu-mice 05o80 cover 15 81ndash90 cover 25 91ndash97 cover 35497 cover For sand 05 0ndash3cover 15 4ndash6 cover 25 7ndash10 cover35410 cover Position was defined by x (northndashsouth) and y (eastndashwest) grid coordinates
Analyses
We calculated species richness plot cover indexSimpsonrsquos dominance [D5 1Spi
2] and Shannondiversity (H05 [Spi log pi]) where pi is the pro-portion of the cover index represented by the ithspecies from the cover index of each species (MjMSoftware Design Gleneden Beach OR US) Weclassified all plots in each year into community types(CTs) with flexible sorting (an effective space-con-serving group-linkage method that employsb5 025 to limit chaining) using the Euclideandistance between samples (MjM Software Design)The 6464 samples formed a dendrogram that we di-vided into 10 CTs recognized when members sharedat least 50 of the information Large differences inspecies richness distort dendrograms (Clifford1976) so divisions occurred at different fusion le-vels We used multiple response permutationprocedures (MRPP MjM Software Design) to as-sess validity of the CTs MRPP calculates aweighted mean within-group distance and a T-valuethat describes the distinctiveness of the groups A(0 to 10) by describing group homogeneitycompared to a random classification We formed sixhabitat types (HTs) from the topographic and sur-face variables using the same classification approachas for species
Recognizing that succession has encompassedonly early primary succession we divided the CTsinto four stages For convenience these are pioneerearly mid- and late seral The categories were based
858 del Moral Roger et al
on the dispersal growth form and longevity ofdominant species (Appendix S1)
After 1990 annual species turnover was low solinear methods could be applied (Legendre amp An-derson 1999) We used redundancy analysis (RDALeps amp Smilauer 2003 CANOCO DLO-Agri-cultural Mathematics Group Wageningen NL) toexplore the relationship between species composi-tion (index values) and explanatory variables in1990 1993 1996 1999 2003 2006 and 2008 Speciescomposition in each of these years was regressedwith explanatory variables to create fitted speciesscores We applied principal components analysis tothese values to produce canonical plot scores RDAthen formed linear equations to predict the positionof plots We assessed variation by comparing theresult to a null model (2000 random trials) The re-gression coefficient of each variable with each RDAaxis estimated how well variables predicted plot po-sition determined by a t-value (df5 400 samples ndash 8variables ndash 15 391) The surplus species variationsuggested the importance of unmeasured variablessampling error and stochastic effects
Spatial pattern of CTs was determined by com-paring the inter-plot distances of a given CT to thedistances between N random plots and the nearestplot of that CT Adjacent plots were 0 distance lar-ger spacings were calculated by Euclidean distancein grid units The observed mean between-plot dis-tance was compared by t-test to that determinedfrom the hypothesis of random distribution
We used Statistix 9 (Analytical Software Talla-hassee FL US) to conduct statistical analysesComparisons of cover index values among species inthe CTs and among DCA scores of CTs were madewith one-way ANOVA or repeat measures ANO-VA as appropriate each followed by a Bonferronitest of differences We used w2 association tests torelate community patterns to surface patterns Weused Axum 7 (Mathsoft Insightful CorporationSeattle WA US) to produce graphs and generaterandom numbers
Results
Vegetation
Succession on Abraham Plain has been leisurelyBy 2009 vegetation remained sparse (Fig S1) andpercentage cover was one-quarter of that observedon a similar grid on the north side of Mount StHelens with three-quarters as many species (delMoral 2009b)
The composition of each of the 10 CTs was dis-tinctive (MRPP A5 0278 T5 1772 Po00001Table 1) Each pair-wise comparison by MRPPshowed that CTs differed significantly Pioneer CTsvanished by 2001 (Fig 1a) they had lower richness(Fig 1b) and cover indices (Fig 1c) than did persis-tent CTs H0 (Fig 1d) increased with time and washigher in persistent CTs Each plot supported amean of 495 106 CTs between 1988 and 2008We characterize CT composition below The num-ber of plots within which the CT ever occurred is inparentheses
There were two pioneer community types CT-A (Anaphalis-Chamerion-Hypochaeris n5 397 of400 plots) had sparse cover of a few pioneer specieseach with long-distance wind dispersal herbaceouslife form and short life span (Appendix S1) As coverincreased plots developed into other CTs CT-B(Lupinus lepidus n5 11) established first in sevenplots near surviving vegetation and was character-ized by L lepidus with sparse Anaphalis and Carexmertensii We recognized three early seral CTs thatalways developed from other CTs and had morespecies and cover than did pioneer CTs These in-cluded a few species that are more persistent Plotscharacterized as CT-C (Anaphalis n5 107) usuallydeveloped from CT-A plots CT-D (Anaphalis-Cis-tanthe-Agrostis) n5 319) was the main nexusthrough which pioneer CTs developed It had lowerrichness and cover than did persistent CTs CT-E(Anaphalis-Chamerion n5 208) had more Hy-pochaeris Luetkea andRacomitrium than did CT-D
Transitions to persistent CTs involved increas-ing species richness and cover and greaterdominance by longer-lived often woody speciesWe characterized two persistent CTs as mid-seralMost common species have less effective dispersalthan those in pioneer CTs The richness of CT-F(Penstemon-Agrostis-Cistanthe n5 338) peakedwhen it contained both pioneer and seral speciesH0
declined as the vegetation matured CT-G (Pen-stemon-Agrostis-Juncus n5 75) retained Anaphalisand developed more of the persistent species (egLuetkea and Racomitrium) than did CT-F Threelate-seral CTs may represent divergent trajectoriestowards mature vegetation CT-H (Penstemon-Agrostis-Salix n5 266) also had Juncus Cistantheand Luetkea Common species are long-lived andseveral are shrubs CT-H had more species highercover and greater diversity than other CTs It hadchanged little since 2004 CT-I (Penstemon-Agrostis-Cistanthe n5 125) also supported Juncus and mos-ses Richness cover and H0 were intermediateamong persistent CTs CT-J (Penstemon-Agrostis-
Primary succession trajectories 859
Cistanthe-Juncus n5 145) also had substantialLuetkea and Racomitrium Pioneer species were un-common and taller woody species (Abies spp andSalix) absent Richness cover and diversity were allleast of the persistent CTs
Species patterns
Many species increased in cover (Fig 2 Fig S2)during the study but cover of pioneer species de-clined after the mid-1990s In 2008 1093 speciesplot were wind-dispersed while only 011 speciesplot were animal-dispersed (Appendix S1) Pioneerspecies have effective long-distance dispersal me-chanisms and while seral species are also wind-
dispersed their dispersal abilities are more limited(Fuller amp del Moral 2003) Penstemon and Agrostispallens expanded to occupy all plots while Juncusparryi and Cistanthe umbellata occurred in mostLuetkea increased steadily but by 2008 it appearedto be stable Salix increased gradually and spor-adicallyRacomitrium occurred in four plots in 1988while Polytrichum did not occur until 1995 Carexmertensii and C microptera were widely distributedand occasionally abundant both occur on halfthe grid Saxifraga once common declined in2008 but this may be associated with dry summerconditions
Relative cover clearly shows the contrast be-tween pioneer and persistent species (Fig 3 Fig S3)
Table 1 Structure and mean cover index in 10 community types (CTs) Numbers in parentheses after species are occurrences(plotsyears) Values in bold indicate dominant species (high cover index) in that CT Each structural variable differedsignificantly among CTs (one-way ANOVA Po0001 values with same superscript fall within the same group as determinedby Bonferroni comparisons) Species cover index means also differed significantly (ANOVA Po0001) but patterns weretoo complex to display grouping patterns CTs and species were each arranged in order of their DCA scores
Structure CT-A CT-B CT-C CT-D CT-E CT-F CT-G CT-H CT-I CT-J
Richness 37a 31a 53b 95d 89c 109e 125f 127f 111e 91cd
Cover (sum indexscores)
52a 38a 70a 159c 136b 242d 321g 298f 281f 238e
Dominance 0516a 0575a 0734b 0851cde 0840c 0871def 0890ef 0890ef 0877def 0843cd
H0 1001a 0999a 1512b 2078d 2008cd 2204e 2347f 2361f 2227e 2001c
Number of plotsyears 1558 21 108 877 444 1289 219 1334 421 495First year observed 1988 1988 1988 1990 1989 1992 1996 1994 1997 1997Last year observed 2000 1991 1998 2001 2000 2008 2008 2008 2008 2008Hypochaeris radicata
(3011)0444 0000 0491 1197 1380 0497 0365 0412 0214 0040
Anaphalis margaritacea(5784)
1557 0810 1713 3304 3459 2370 2717 2147 1337 0802
Chamerionangustifolium (4712)
0514 0191 0593 1911 1803 1442 0991 1077 0691 0513
Hieracium albiflorum(3907)
0336 0143 0398 0904 0930 0929 0753 0813 0411 0293
Lupinus lepidus (248) 0007 1667 0028 0051 0220 0076 0383 0021 0088 0036Lupinus latifolius (122) o0001 0095 0000 o0001 0047 0020 0932 0037 0000 0000Abies sp (736) 0106 0000 0120 0123 0124 0087 0032 0352 1062 0020Luetkea pectinata
(4075)0182 0095 0157 0691 0916 1389 2078 2524 1981 1432
Carex mertensii (2691) 0128 0428 0046 0379 0373 0680 0827 1249 1057 0649Saxifraga ferruginea
(1596)0045 0095 0083 0276 0253 0632 1082 1079 0456 0247
Racomitrium canescens(2498)
0072 0000 0019 0243 0943 0421 2183 1080 1064 0990
Cistanthe umbellata(4867)
0123 0048 1370 2222 0523 3199 3603 3469 4033 4853
Agrostis pallens (5060) 0294 0000 0528 1331 0738 2855 3457 3271 3924 4301
Eriogonum pyrolifolium(653)
0016 0000 0000 0071 0057 0120 0338 0281 0591 0059
Juncus parryi (4421) 0116 0191 0250 1008 0550 2066 4064 2633 3565 3465
Penstemon cardwellii(4489)
0195 0000 0278 0520 0484 3165 4489 3591 4197 4216
Salix sitchensis (1858) 0037 0000 0000 0140 0188 0216 1237 3144 0271 0073Carex microptera
(1723)0035 0095 0065 0161 0091 0412 0763 0752 0644 1372
Polytrichumjuniperinum (1133)
o0001 0000 0000 0047 0093 0182 0182 1601 2183 0168
860 del Moral Roger et al
Species such as Penstemon Agrostis Juncus parryiCarex spp and Luetkea increased across the CTswhen listed by their first appearance These speciesare long-lived with strong vegetative growth
Fig 1 Structure of CTs from 1988 to 2008 (a) Number of plots (b) Mean species richness (c) Mean cover index (d) Meandiversity index (H0)
Fig 2 Changes in mean cover index for abundant species
Fig 3 Changes in relative cover (proportion of total cov-er in the sample) of common species in the four stages ofearly primary succession The first four species are persis-tent the last two are pioneer species (defined in AppendixS1)
Primary succession trajectories 861
In contrast pioneer species soon peaked and thendeclined
Habitat relationships
We could use only position topography andsurface conditions as explanatory variables in RDAExplained variation was low but increased four-fold from 1990 to 2006 (Table 2) In 1996 we ob-tained the first significant result when the y-axis(possibly related to distance from survivors) and rillfraction became significant By 2006 smooth sur-face also contributed but topography did notinfluence the vegetation
We recognized six habitat types (Table 3) A fivepersistent CT (Table S1) by six HT w2 test was sig-nificant (w2 5 935 Po00001 df 5 20) CT-F(Penstemon-Agrostis-Cistanthe) was more commonin rills CT-G (Penstemon-Agrostis-Juncus) wasmore common in gullies CT-H (Penstemon-Agros-tis-Salix) tended to avoid smooth plots and wasmore common in plots with rills or gullies and CT-J(Penstemon-Agrostis-Cistanthe-Juncus) tended tooccur in smooth plots but avoided rills or gullies
The spatial pattern displayed by the CTs re-flected some environmental sorting (Fig 4) Nearestneighbour distances compared to the randommodel suggested that CT-F formed scattered clus-ters (mean observed distance5 142 mean randomdistance5 224 t5 225 P5 003) Some plots ofCT-G were confined to the eastern edge while oth-ers were in the southwest corner but overall it wasnot clustered (observed5 190 random5 243t5 190 P5 006) CT-H was clustered (ob-served5 015 random5 058 t5 483 Po00001)as was CT-I (observed5 052 random5 145t5 448 Po00001) concentrated on the east halfof the grid CT-J was clustered (observed5 119random5 264 t5 383 P5 0002) and con-centrated on the eastern part of the grid
Trajectories
We explored vegetation trajectories in severalways In 1988 richness the mean cover index(Fig 5) H0 and dominance (Fig S4) were leastRichness increased until 1999 after which time itvaried around 11 species per plot The cover indexcontinued to increase while H0 and dominance sta-bilized by the mid-1990s
Succession began with colonization by highlyvagile pioneers such as Anaphalis Chamerion Hier-acium and Hypochaeris Plots changed into otherCTs at variable rates While we found transitionsamong all CTs some trajectories were more typicalthan were others (Fig 6) The most likely trajec-
Table 2 Summary of redundancy analysis results usingposition topography and surface variables on the gridThe alternate plots sampled in 2007 and 2008 were ana-lysed separately Percentage variation5 how much ofspecies variation is explained by variables F-values5 sig-nificance level of Axis 1 trace5 percentage variance onAxis 1 Correlations 405
Years Percentagevariation
F-value
Trace Correlatedvariables
1990 34 707 157 ndash1993 65 128 340 ndash1996 79 178 420 y rill1999 109 331 598 y rill2003 112 338 616 y rill2006 125 390 698 y rill flat2008 122 335 654 y rill flat
Table 3 Mean value of characteristics of six habitat types (HT) HTs differed significantly for each terrain characteristic(Po00001 one-way ANOVA followed by Bonferroni tests for differences among means Values with same superscript fallwithin the same group)
Terrain HT-1 HT-2 HT-3 HT-4 HT-5 HT-6 F-value(n5 128) (n5 28) (n5 37) (n5 64) (n5 61) (n5 82)
Smooth 300a 286a 289a 192bc 198b 180c 1911Gully 005b 036b 030b 203a 026b 222a 2085Rill 015d 029cd 057c 061c 238a 117b 1141Rock 000d 100bc 132b 080c 062c 216a 1466Sand 000b 000b 019b 002b 010b 090a 306Pumice 300a 300a 200c 261b 257b 128d 1470
Fig 4 Distribution of CTs on the grid in 2008
862 del Moral Roger et al
tories proceeded from pioneer plots dominated byAnaphalis to early seral types that had accumulatedCistanthe Agrostis and Luetkea These developedfurther into mid-seral communities when Penstemonbecame dominant and Juncus became widespreador they developed directly into late seral commu-nities dominated by Penstemon and Agrostis In2008 some mid-seral CTs persisted but most plotswere in a late-seral community that had changedlittle since 2005 For each annual interval 1988 to2008 a plot could persist in a CT progress to a moredeveloped one or regress to an earlier stage Therewere 301 retrogressions and 1014 progressions The
five extant CTs persisted significantly longer thandid the five vanished CTs (Table 4)
We clarified temporal vegetation changes usingDCA (Fig 7) Total variance (l) was 159 (DCA-15 155 DCA-25 74) Turnover on DCA-1was 53 half-changes on DCA-2 it was 39 Pioneerspecies had high values on DCA-1 species with lowDCA-1 values appeared later and were persistentMean DCA-1 position of the plots changed sig-nificantly between years (r2 5 0637 Po00001Spearman nonparametric correlation r5 079Po00001) DCA-1 scores by year changed rapidlythrough 2001 (ANOVA Po00001) then slowedDCA-2 changes ceased after 1997 but ANOVA was
Fig 5 Annual changes of mean species richness and meancover index Differences (Po005) among measures de-termined by a repeat measures ANOVA followed by theBonferroni test Values with the same associated letter fallwithin the same group of values
Fig 6 Trajectories showing the number of transitionsbetween CTs Numbers in parentheses after persistent CTsare the number of plots in 2008 Many arrows transitionnear the mid-point to differentiate between transitions toand from a given CT
Table 4 Number of continuous years (standard devia-tion) for which each CT persisted Differences amongmeans are significant (one-way ANOVA Po00001 fol-lowed by Bonferroni tests for differences among meansSuperscripts indicate membership in same group) Mini-mum run is one in each case
CT Years SD Max
A 329b 135 7B 200ab 115 4C 107a 026 2D 238b 142 7E 181ab 117 7F 398c 274 10G 391c 172 6H 558d 256 10I 419cd 167 8J 378c 185 7
Fig 7 Detrended correspondence analysis of plots by CTand by year Differences (Po005) among measures de-termined by ANOVA followed by the Bonferroni testFor each set values with the same associated letter fallwithin the same group of values for DCA-1 (no data for1998 and 2002 alternate plots were sampled in 2007 and2008) The inset is a separate DCA of plots in 2008
Primary succession trajectories 863
significant (r2 5 0206 Po00001 Spearman non-parametric correlation r5 025 Po00001) Thevariation in DCA-1 scores within years declined (thelinear correlation of years vs standard deviationwas Po00001 t5 533 r2 5 0637)
The mean DCA position of the 10 CTs did notreveal a single trajectory CT-C (Anaphalis) wasmore closely associated with pioneer CTs but wasnever the first CT on a plot CT-G (Penstemon-Agrostis-Cistanthe) was aligned with late-seral CTsWe also conducted a DCA of 2008 plots (Fig 7inset) b-diversity was low with l5 097 (DCA-15
133 DCA-25 84 ca 18 HC along DCA-1)No trajectory was evident from this analysis
Discussion
There are so few long-term longitudinal studiesof early primary succession on stressful sites be-cause initially so little happens The vegetation ofthe Abraham Plain remains sparse 30 years after theeruption It has developed by interwoven succes-sional trajectories that include plots that divergeconverge regress and progress to form a braidedsuccession Neither habitat factors nor competitionappears to have provided strong filtering effects onspecies composition although unmeasured soilvariables (eg moisture and nitrogen content) andfactors related to stochastic colonization and estab-lishment (cf del Moral amp Ellis 2004) may explainsome of the remaining unexplained variation Theimmigration seed density was low (Wood amp delMoral 2000) many suitable sites appeared vacant inearly years (del Moral amp Bliss 1993) and severalspecies occupied similar sites Chance may governsuch patterns We propose that when environmentalvariables have only slight effects on species compo-sition consistent successional trajectories areunlikely to emerge Below we discuss questionsabout predictability rate and pattern of successionon Abraham Plain that are the focus of this study
Environmentndashvegetation connection
Predictable relationships between environ-mental variables and vegetation had developed by2008 but they remained low Priority effects sec-ondary disturbances elk grazing and seasonalfluctuations may affect these trajectories in the fu-ture rendering it unlikely that explained variationwould become the dominant factor before forestsdevelop
We did not use soil factors because plants es-tablish at a smaller scale than the plot size and it isimpractical to measure such factors as moisture ineven a few plots Although soils were young andhomogeneous it is likely that inclusion of soil fac-tors (eg pH and moisture) would have improvedexplanatory power based on similar studies (delMoral 2009a) explained variance might doubleTopographic surface and position factors alone diddemonstrate increasing explained variance of spe-cies pattern over time but this trend was lesspronounced than in other studies On the northslope of Mount St Helens in 1992 only spatial fac-tors explained any variation (15) but after 10 yrenvironmental factors dominated (28 del Moralamp Lacher 2005) Baasch et al (2009) produced oneof the very few primary succession studies to showthat relationships between vegetation and ex-planatory variables increased over time They toosuggested that the impact of stochastic processesdeclined over time
The spatial distribution of CTs suggested someenvironmental influence Rills provide havens forpioneers (eg Anaphalis Hieracium and Hy-pochaeris) Gullies offer protection from wind andaccumulate snow which allowed species such asAnaphalis and Salix to colonize and persist Pen-stemon and Juncus were common in more exposedsites where cover and diversity were least In suchsites species that are less stress tolerant (eg SalixPolytrichum Saxifraga and Juncus mertensianus)were uncommon while stress-tolerant species suchas Cistanthe Juncus parryi Agrostis pallens andCarex microptera reached their maximum values
Rates
Habitats with limited fertility or moisture typi-cally have low succession rates (Donnegan ampRebertus 1999 Anderson 2007) and succession onAbraham Plain has been no exception Mean annualDCA scores were in a single group of values Be-tween 1988 and 1999 these scores formed sevengroups Surrounding habitats on Mount St Helensdeveloped more rapidly On an exposed lahar on thesouth slope richness averaged 16 species and thecover index averaged 73 (del Moral 2009b) prior to2008 In contrast mean richness on the AbrahamPlain in 2008 was 108 and the mean cover index was314 The proximity to forest more fertile soils andabundance of the N-fixing Lupinus may all con-tribute to accelerated primary succession at suchsites relative to Abraham Plain
864 del Moral Roger et al
Trajectories
Succession trajectories have been discussedthoroughly (cf Leps amp Rejmanek 1991) When thevegetation of initially homogeneous sites developslocally distinctive characteristics divergence occurs(Bossuyt et al 2003) Divergent trajectories occur onmany surfaces including landslides glacier fore-lands wet slack dune vegetation floodplains andmine tailings (Walker amp del Moral 2003) Lanta ampLeps (2009) showed experimentally that differentialdispersal could promote divergence when there arepriority effects Divergence may be more likely instressful habitats when different species arrive firstand where climatic factors are highly variable(MacDougall et al 2008) Convergence describestrajectories that become more similar over timeConvergence has been assumed to result when cli-matic factors dominate and there is evidence thatwhen dispersal effects are small and competitive ef-fects are strong convergence to a single communitydoes occur (van Oijen et al 2005 Anthelme et al2007) Convergence is also more likely when thereare few alternative stages as on glacial forelands(Hodkinson et al 2003 but see Robbins ampMathews2009)
Here we show directly that early succession un-der stressful conditions can include trajectories thatdiverge converge regress and progress to form areticulate succession Our study joins a growingbody of work that demonstrates the frequency ofmultiple trajectories in early primary succession (egdel Moral 2007 Prach amp Hobbs 2008 Walker amp delMoral 2009) Forecasting trajectories and identify-ing factors that limit their development is crucial tounderstanding community assembly mechanismsOn the Abraham Plain persistent species graduallyreplaced readily dispersed pioneer species Subtlehabitat variation annual weather fluctuation andlandscape effects combined to produce communitiesrelated through a network of development and re-sponsible to topographic variation
The first colonists were species with excellentdispersal but isolation allowed for only a meagerseed rain they established pioneer communities thatsoon developed into other CTs characterized byspecies with greater persistence (cf Ejrnaes et al2006) Several alternative transitions were possiblefor most CTs suggesting that stochastic processesinitially dominated transitions Gradually persis-tent species invaded and attained dominance Minortopographic variation affected composition some-what resulting in some successional transitions thatare more common than others Of annual transi-
tions between successional stages 51 wereregressions and 170 were progressions DCA re-vealed no single trend among the CTs The two mid-seral community types were central to the transitionpatterns while the three late-seral CTs occurred indifferent directions from this centre
It is unlikely that complete convergence will oc-cur Even when trees dominate understoreyvegetation is likely to retain variation initiated dur-ing early succession and maintained by topographicvariation and priority effects The understorey offorests on the south side of Mount St Helens de-monstrates such variation (del Moral amp Ellis 2004)There is no evidence yet for a single target commu-nity
Strong assembly rules are more likely to pro-duce convergent trajectories than weak ones Whilestrong assembly rules may exist in mature vegeta-tion (see Navas amp Violle 2009) they appear weak onAbraham Plain and are more effective for functionaltypes than species One rule may be that long-livedwind-dispersed species accumulate at the expense ofshort-lived pioneers A second may be that grami-noids with rhizomatous growth (eg Agrostispallens) and prostrate shrubs (eg Penstemon) arecomplementary and can coexist indefinitely A thirdmay be that fleshy fruit-producing shrubs (eg Ru-bus spp Vaccinium spp) cannot establish undercurrent conditions Finally although seeds of con-ifers and Populus balsamifera are commonconditions do not yet favour their developmentThese rules are weak and exceptions are commonThe spontaneously developing vegetation of theAbraham Plain is constrained by isolation from po-tential colonists as well as a habitat that challengesplant growth
Acknowledgements We thank the US National Science
Foundation for funding (BSR8906544 DEB9406987
DEB0087040 and DEB0541972) and the Mount St He-
lens National Volcanic Monument for permissions
Valuable comments by J G Bishop M P Fleming Jan-
neke Hille Ris Lambers L R Walker J H Titus and two
anonymous reviewers improved the paper Field assis-
tance was provided through the years by S Anderson W
Arnesen S Bard A Coogan S Crawford K Dlugosch
A Eckert E Ellis C Fairbourn C Muerdter M Flem-
ing T Fletcher R Fuller A Grant E Jenkins M
Johns C Jones L McMullan P Moody R Robham L
Rozzell J Sandler R Sewell-Nesteruk M Spasojevic
M Tweiten S Wilson B Witte and C Wolfe This is
contribution No 60 to the Mount St Helens Succession
Project
Primary succession trajectories 865
References
Anderson KJ 2007 Temporal patterns in rates of
community change during succession American
Naturalist 169 780ndash793
Anthelme F Villaret JC amp Brun JJ 2007 Shrub
encroachment in the Alps gives rise to the
convergence of sub-alpine communities on a regional
scale Journal of Vegetation Science 18 355ndash362
Baasch A Tischew S amp Bruelheide H 2009 Insights
into succession processes using temporally repeated
habitat models results from a long-term study in a
post-mining landscape Journal of Vegetation Science
20 629ndash638
Bossuyt B Honnay O amp Hermy M 2003 An island
biogeographical view of the successional pathway in
wet dune slacks Journal of Vegetation Science 14 781ndash
788
Clifford HT 1976 Dendrograms and their
interpretation In Williams WT (ed) Pattern
analysis in agricultural science pp 96ndash109 Elsevier
Science Amsterdam NL
del Moral R 2007 Vegetation dynamics in space and
time an example from Mount St Helens Journal of
Vegetation Science 18 479ndash488
del Moral R 2009a Increasing deterministic control of
primary succession onMount St Helens Washington
Journal of Vegetation Science 20 1145ndash1154
del Moral R 2009b Mount St Helens permanent plots
and grids 1980 to present Available at http
protistbiologywashingtonedudelmoral Accessed 8
December 2009
del Moral R amp Bliss LC 1993 Mechanism of primary
succession insights resulting from the eruption of
Mount St Helens Advances in Ecological Research
24 1ndash66
del Moral R amp Ellis EE 2004 Gradients in hetero-
geneity and structure on lahars Mount St Helens
Washington USA Plant Ecology 175 273ndash286
del Moral R amp Lacher IL 2005 Vegetation patterns 25
years after the eruption of Mount St Helens
WashingtonAmerican Journal of Botany 92 1948ndash1956
Donnegan JA amp Rebertus AJ 1999 Rates and
mechanisms of subalpine forest succession along an
environmental gradient Ecology 80 1370ndash1384
Ejrnaes R Bruun HH amp Graae BJ 2006 Community
assembly in experimental grasslands suitable
environment or timely arrival Ecology 87 1225ndash1233
Felinks B ampWiegand T 2008 Exploring spatiotemporal
patterns in early stages of primary succession on
former lignite mining sites Journal of Vegetation
Science 19 267ndash276
Fuller RN amp del Moral R 2003 The role of refugia and
dispersal in primary succession on Mount St Helens
Washington Journal of Vegetation Science 14 637ndash
644
Hodkinson IA Coulson SJ amp Webb NR 2003
Community assembly along proglacial chrono-
sequences in the high Arctic vegetation and soil
development in north-west Svalbard Journal of
Ecology 91 651ndash663
Johnson EA amp Miyanishi K 2008 Testing the
assumptions of chronosequences in succession
Ecology Letters 11 419ndash431
Lanta V amp Leps J 2009 How does surrounding
vegetation affect the course of succession a five-year
container experiment Journal of Vegetation Science
20 686ndash694
Legendre P amp Anderson MJ 1999 Distance-based
redundancy analysis testing multispecies responses in
multi-factorial ecological experiments Ecological
Monographs 69 1ndash24
Leps J amp Rejmanek M 1991 Convergence or
divergence what should we expect from vegetation
succession Oikos 62 261ndash264
Leps J amp Smilauer P 2003 Multivariate analysis of
ecological data using CANOCO Cambridge
University Press Cambridge UK
MacDougall AS Wilson SD amp Bakker JD 2008
Climatic variability alters the outcome of longterm
community assembly Journal of Ecology 96 346ndash354
Navas ML amp Violle C 2009 Plant traits related to
competition how do they shape the functional
diversity of communities Community Ecology 10
131ndash137
Pickett STA Cadenasso ML amp Meiners SJ 2009
Ever since Clements from succession to vegetation
dynamics and understanding to intervention Applied
Vegetation Science 12 9ndash21
Prach K amp Hobbs RJ 2008 Spontaneous succession
versus technical reclamation in the restoration of
disturbed sites Restoration Ecology 16 363ndash366
Prach K amp Rehounkova K 2006 Vegetation succession
over broad geographical scales which factors
determine the patterns Preslia 78 469ndash480
Robbins JA amp Matthews JA 2009 Pioneer vegetation
on glacier forelands in southern Norway emerging
communities Journal of Vegetation Science 20 889ndash
902
Swanson FJ amp Major JJ 2005 Physical events
environments and geologicalndashecological interactions
at Mount St Helens March 1980ndash2004 In Dale
VH Swanson FJ amp Crisafulli CM (eds)
Ecological recovery after the 1980 eruption of Mount
St Helens pp 27ndash44 Springer New York NY US
USDA National Resource Conservation Service 2009
The Plants Data Base National Plants Data Center
Baton Rouge LA US Available at http
plantsusdagov Accessed October 15 2009
van Oijen D Hommel P den Ouden J amp de Waal R
2005 Effects of tree species composition on within-
forest distribution of understorey species Applied
Vegetation Science 8 155ndash166
Walker LR amp del Moral R 2003 Primary succession
and ecosystem rehabilitation Cambridge University
Press Cambridge UK
866 del Moral Roger et al
Walker LR amp del Moral R 2009 Transition dynamics
in succession implications for rates trajectories and
restoration In Hobbs RJ amp Suding KN (eds)
New models for ecosystem dynamics and restoration
pp 33ndash49 Island Press Washington DC US
Walker LR Bellingham PJ amp Peltzer DA 2006
Plant characteristics are poor predictors of microsite
colonization during the first two years of pri-
mary succession Journal of Vegetation Science 17
397ndash406
Wood DM amp del Moral R 1988 Colonizing plants on
the Pumice Plains Mount St Helens Washington
American Journal of Botany 75 1228ndash1237
Wood DM amp del Moral R 2000 Seed rain during early
primary succession onMount St Helens Washington
Madrono 47 1ndash9
Supporting Information
Additional supporting information may befound in the online version of this article
Fig S1 Abraham Plain Sept 1980 pumice de-posits over scoured bedrock looking north towardsthe study site
Fig S2 Abraham Plain July 2009 lookingnorthwest across the study site
Fig S3 Changes in mean cover index for lesscommon species
Fig S4 Relative cover (proportion of totalcover in the sample) of less common species in thefour stages of early primary succession The firstfour species are persistent the last two are pioneerspecies (defined in Appendix S1)
Fig S5 Annual changes of mean H0 and meanSimpsonrsquos dominance index Differences (Po005)among measures determined by a repeat measuresANOVA followed by the Bonferroni test Valueswith same associated letter fall within the samegroup of values
Table S1 Structure and species composition ofCTs found in 2008
Appendix S1 Characteristics of species en-countered on Abraham Plain grid
Please note Wiley-Blackwell is not responsiblefor the content or functionality of any supportingmaterials supplied by the authors Any queries(other than missing material) should be directed tothe corresponding author for the article
Received 9 December 2009
Accepted 14 March 2010
Co-ordinating Editor Dr Kerry Woods
Primary succession trajectories 867

on the dispersal growth form and longevity ofdominant species (Appendix S1)
After 1990 annual species turnover was low solinear methods could be applied (Legendre amp An-derson 1999) We used redundancy analysis (RDALeps amp Smilauer 2003 CANOCO DLO-Agri-cultural Mathematics Group Wageningen NL) toexplore the relationship between species composi-tion (index values) and explanatory variables in1990 1993 1996 1999 2003 2006 and 2008 Speciescomposition in each of these years was regressedwith explanatory variables to create fitted speciesscores We applied principal components analysis tothese values to produce canonical plot scores RDAthen formed linear equations to predict the positionof plots We assessed variation by comparing theresult to a null model (2000 random trials) The re-gression coefficient of each variable with each RDAaxis estimated how well variables predicted plot po-sition determined by a t-value (df5 400 samples ndash 8variables ndash 15 391) The surplus species variationsuggested the importance of unmeasured variablessampling error and stochastic effects
Spatial pattern of CTs was determined by com-paring the inter-plot distances of a given CT to thedistances between N random plots and the nearestplot of that CT Adjacent plots were 0 distance lar-ger spacings were calculated by Euclidean distancein grid units The observed mean between-plot dis-tance was compared by t-test to that determinedfrom the hypothesis of random distribution
We used Statistix 9 (Analytical Software Talla-hassee FL US) to conduct statistical analysesComparisons of cover index values among species inthe CTs and among DCA scores of CTs were madewith one-way ANOVA or repeat measures ANO-VA as appropriate each followed by a Bonferronitest of differences We used w2 association tests torelate community patterns to surface patterns Weused Axum 7 (Mathsoft Insightful CorporationSeattle WA US) to produce graphs and generaterandom numbers
Results
Vegetation
Succession on Abraham Plain has been leisurelyBy 2009 vegetation remained sparse (Fig S1) andpercentage cover was one-quarter of that observedon a similar grid on the north side of Mount StHelens with three-quarters as many species (delMoral 2009b)
The composition of each of the 10 CTs was dis-tinctive (MRPP A5 0278 T5 1772 Po00001Table 1) Each pair-wise comparison by MRPPshowed that CTs differed significantly Pioneer CTsvanished by 2001 (Fig 1a) they had lower richness(Fig 1b) and cover indices (Fig 1c) than did persis-tent CTs H0 (Fig 1d) increased with time and washigher in persistent CTs Each plot supported amean of 495 106 CTs between 1988 and 2008We characterize CT composition below The num-ber of plots within which the CT ever occurred is inparentheses
There were two pioneer community types CT-A (Anaphalis-Chamerion-Hypochaeris n5 397 of400 plots) had sparse cover of a few pioneer specieseach with long-distance wind dispersal herbaceouslife form and short life span (Appendix S1) As coverincreased plots developed into other CTs CT-B(Lupinus lepidus n5 11) established first in sevenplots near surviving vegetation and was character-ized by L lepidus with sparse Anaphalis and Carexmertensii We recognized three early seral CTs thatalways developed from other CTs and had morespecies and cover than did pioneer CTs These in-cluded a few species that are more persistent Plotscharacterized as CT-C (Anaphalis n5 107) usuallydeveloped from CT-A plots CT-D (Anaphalis-Cis-tanthe-Agrostis) n5 319) was the main nexusthrough which pioneer CTs developed It had lowerrichness and cover than did persistent CTs CT-E(Anaphalis-Chamerion n5 208) had more Hy-pochaeris Luetkea andRacomitrium than did CT-D
Transitions to persistent CTs involved increas-ing species richness and cover and greaterdominance by longer-lived often woody speciesWe characterized two persistent CTs as mid-seralMost common species have less effective dispersalthan those in pioneer CTs The richness of CT-F(Penstemon-Agrostis-Cistanthe n5 338) peakedwhen it contained both pioneer and seral speciesH0
declined as the vegetation matured CT-G (Pen-stemon-Agrostis-Juncus n5 75) retained Anaphalisand developed more of the persistent species (egLuetkea and Racomitrium) than did CT-F Threelate-seral CTs may represent divergent trajectoriestowards mature vegetation CT-H (Penstemon-Agrostis-Salix n5 266) also had Juncus Cistantheand Luetkea Common species are long-lived andseveral are shrubs CT-H had more species highercover and greater diversity than other CTs It hadchanged little since 2004 CT-I (Penstemon-Agrostis-Cistanthe n5 125) also supported Juncus and mos-ses Richness cover and H0 were intermediateamong persistent CTs CT-J (Penstemon-Agrostis-
Primary succession trajectories 859
Cistanthe-Juncus n5 145) also had substantialLuetkea and Racomitrium Pioneer species were un-common and taller woody species (Abies spp andSalix) absent Richness cover and diversity were allleast of the persistent CTs
Species patterns
Many species increased in cover (Fig 2 Fig S2)during the study but cover of pioneer species de-clined after the mid-1990s In 2008 1093 speciesplot were wind-dispersed while only 011 speciesplot were animal-dispersed (Appendix S1) Pioneerspecies have effective long-distance dispersal me-chanisms and while seral species are also wind-
dispersed their dispersal abilities are more limited(Fuller amp del Moral 2003) Penstemon and Agrostispallens expanded to occupy all plots while Juncusparryi and Cistanthe umbellata occurred in mostLuetkea increased steadily but by 2008 it appearedto be stable Salix increased gradually and spor-adicallyRacomitrium occurred in four plots in 1988while Polytrichum did not occur until 1995 Carexmertensii and C microptera were widely distributedand occasionally abundant both occur on halfthe grid Saxifraga once common declined in2008 but this may be associated with dry summerconditions
Relative cover clearly shows the contrast be-tween pioneer and persistent species (Fig 3 Fig S3)
Table 1 Structure and mean cover index in 10 community types (CTs) Numbers in parentheses after species are occurrences(plotsyears) Values in bold indicate dominant species (high cover index) in that CT Each structural variable differedsignificantly among CTs (one-way ANOVA Po0001 values with same superscript fall within the same group as determinedby Bonferroni comparisons) Species cover index means also differed significantly (ANOVA Po0001) but patterns weretoo complex to display grouping patterns CTs and species were each arranged in order of their DCA scores
Structure CT-A CT-B CT-C CT-D CT-E CT-F CT-G CT-H CT-I CT-J
Richness 37a 31a 53b 95d 89c 109e 125f 127f 111e 91cd
Cover (sum indexscores)
52a 38a 70a 159c 136b 242d 321g 298f 281f 238e
Dominance 0516a 0575a 0734b 0851cde 0840c 0871def 0890ef 0890ef 0877def 0843cd
H0 1001a 0999a 1512b 2078d 2008cd 2204e 2347f 2361f 2227e 2001c
Number of plotsyears 1558 21 108 877 444 1289 219 1334 421 495First year observed 1988 1988 1988 1990 1989 1992 1996 1994 1997 1997Last year observed 2000 1991 1998 2001 2000 2008 2008 2008 2008 2008Hypochaeris radicata
(3011)0444 0000 0491 1197 1380 0497 0365 0412 0214 0040
Anaphalis margaritacea(5784)
1557 0810 1713 3304 3459 2370 2717 2147 1337 0802
Chamerionangustifolium (4712)
0514 0191 0593 1911 1803 1442 0991 1077 0691 0513
Hieracium albiflorum(3907)
0336 0143 0398 0904 0930 0929 0753 0813 0411 0293
Lupinus lepidus (248) 0007 1667 0028 0051 0220 0076 0383 0021 0088 0036Lupinus latifolius (122) o0001 0095 0000 o0001 0047 0020 0932 0037 0000 0000Abies sp (736) 0106 0000 0120 0123 0124 0087 0032 0352 1062 0020Luetkea pectinata
(4075)0182 0095 0157 0691 0916 1389 2078 2524 1981 1432
Carex mertensii (2691) 0128 0428 0046 0379 0373 0680 0827 1249 1057 0649Saxifraga ferruginea
(1596)0045 0095 0083 0276 0253 0632 1082 1079 0456 0247
Racomitrium canescens(2498)
0072 0000 0019 0243 0943 0421 2183 1080 1064 0990
Cistanthe umbellata(4867)
0123 0048 1370 2222 0523 3199 3603 3469 4033 4853
Agrostis pallens (5060) 0294 0000 0528 1331 0738 2855 3457 3271 3924 4301
Eriogonum pyrolifolium(653)
0016 0000 0000 0071 0057 0120 0338 0281 0591 0059
Juncus parryi (4421) 0116 0191 0250 1008 0550 2066 4064 2633 3565 3465
Penstemon cardwellii(4489)
0195 0000 0278 0520 0484 3165 4489 3591 4197 4216
Salix sitchensis (1858) 0037 0000 0000 0140 0188 0216 1237 3144 0271 0073Carex microptera
(1723)0035 0095 0065 0161 0091 0412 0763 0752 0644 1372
Polytrichumjuniperinum (1133)
o0001 0000 0000 0047 0093 0182 0182 1601 2183 0168
860 del Moral Roger et al
Species such as Penstemon Agrostis Juncus parryiCarex spp and Luetkea increased across the CTswhen listed by their first appearance These speciesare long-lived with strong vegetative growth
Fig 1 Structure of CTs from 1988 to 2008 (a) Number of plots (b) Mean species richness (c) Mean cover index (d) Meandiversity index (H0)
Fig 2 Changes in mean cover index for abundant species
Fig 3 Changes in relative cover (proportion of total cov-er in the sample) of common species in the four stages ofearly primary succession The first four species are persis-tent the last two are pioneer species (defined in AppendixS1)
Primary succession trajectories 861
In contrast pioneer species soon peaked and thendeclined
Habitat relationships
We could use only position topography andsurface conditions as explanatory variables in RDAExplained variation was low but increased four-fold from 1990 to 2006 (Table 2) In 1996 we ob-tained the first significant result when the y-axis(possibly related to distance from survivors) and rillfraction became significant By 2006 smooth sur-face also contributed but topography did notinfluence the vegetation
We recognized six habitat types (Table 3) A fivepersistent CT (Table S1) by six HT w2 test was sig-nificant (w2 5 935 Po00001 df 5 20) CT-F(Penstemon-Agrostis-Cistanthe) was more commonin rills CT-G (Penstemon-Agrostis-Juncus) wasmore common in gullies CT-H (Penstemon-Agros-tis-Salix) tended to avoid smooth plots and wasmore common in plots with rills or gullies and CT-J(Penstemon-Agrostis-Cistanthe-Juncus) tended tooccur in smooth plots but avoided rills or gullies
The spatial pattern displayed by the CTs re-flected some environmental sorting (Fig 4) Nearestneighbour distances compared to the randommodel suggested that CT-F formed scattered clus-ters (mean observed distance5 142 mean randomdistance5 224 t5 225 P5 003) Some plots ofCT-G were confined to the eastern edge while oth-ers were in the southwest corner but overall it wasnot clustered (observed5 190 random5 243t5 190 P5 006) CT-H was clustered (ob-served5 015 random5 058 t5 483 Po00001)as was CT-I (observed5 052 random5 145t5 448 Po00001) concentrated on the east halfof the grid CT-J was clustered (observed5 119random5 264 t5 383 P5 0002) and con-centrated on the eastern part of the grid
Trajectories
We explored vegetation trajectories in severalways In 1988 richness the mean cover index(Fig 5) H0 and dominance (Fig S4) were leastRichness increased until 1999 after which time itvaried around 11 species per plot The cover indexcontinued to increase while H0 and dominance sta-bilized by the mid-1990s
Succession began with colonization by highlyvagile pioneers such as Anaphalis Chamerion Hier-acium and Hypochaeris Plots changed into otherCTs at variable rates While we found transitionsamong all CTs some trajectories were more typicalthan were others (Fig 6) The most likely trajec-
Table 2 Summary of redundancy analysis results usingposition topography and surface variables on the gridThe alternate plots sampled in 2007 and 2008 were ana-lysed separately Percentage variation5 how much ofspecies variation is explained by variables F-values5 sig-nificance level of Axis 1 trace5 percentage variance onAxis 1 Correlations 405
Years Percentagevariation
F-value
Trace Correlatedvariables
1990 34 707 157 ndash1993 65 128 340 ndash1996 79 178 420 y rill1999 109 331 598 y rill2003 112 338 616 y rill2006 125 390 698 y rill flat2008 122 335 654 y rill flat
Table 3 Mean value of characteristics of six habitat types (HT) HTs differed significantly for each terrain characteristic(Po00001 one-way ANOVA followed by Bonferroni tests for differences among means Values with same superscript fallwithin the same group)
Terrain HT-1 HT-2 HT-3 HT-4 HT-5 HT-6 F-value(n5 128) (n5 28) (n5 37) (n5 64) (n5 61) (n5 82)
Smooth 300a 286a 289a 192bc 198b 180c 1911Gully 005b 036b 030b 203a 026b 222a 2085Rill 015d 029cd 057c 061c 238a 117b 1141Rock 000d 100bc 132b 080c 062c 216a 1466Sand 000b 000b 019b 002b 010b 090a 306Pumice 300a 300a 200c 261b 257b 128d 1470
Fig 4 Distribution of CTs on the grid in 2008
862 del Moral Roger et al
tories proceeded from pioneer plots dominated byAnaphalis to early seral types that had accumulatedCistanthe Agrostis and Luetkea These developedfurther into mid-seral communities when Penstemonbecame dominant and Juncus became widespreador they developed directly into late seral commu-nities dominated by Penstemon and Agrostis In2008 some mid-seral CTs persisted but most plotswere in a late-seral community that had changedlittle since 2005 For each annual interval 1988 to2008 a plot could persist in a CT progress to a moredeveloped one or regress to an earlier stage Therewere 301 retrogressions and 1014 progressions The
five extant CTs persisted significantly longer thandid the five vanished CTs (Table 4)
We clarified temporal vegetation changes usingDCA (Fig 7) Total variance (l) was 159 (DCA-15 155 DCA-25 74) Turnover on DCA-1was 53 half-changes on DCA-2 it was 39 Pioneerspecies had high values on DCA-1 species with lowDCA-1 values appeared later and were persistentMean DCA-1 position of the plots changed sig-nificantly between years (r2 5 0637 Po00001Spearman nonparametric correlation r5 079Po00001) DCA-1 scores by year changed rapidlythrough 2001 (ANOVA Po00001) then slowedDCA-2 changes ceased after 1997 but ANOVA was
Fig 5 Annual changes of mean species richness and meancover index Differences (Po005) among measures de-termined by a repeat measures ANOVA followed by theBonferroni test Values with the same associated letter fallwithin the same group of values
Fig 6 Trajectories showing the number of transitionsbetween CTs Numbers in parentheses after persistent CTsare the number of plots in 2008 Many arrows transitionnear the mid-point to differentiate between transitions toand from a given CT
Table 4 Number of continuous years (standard devia-tion) for which each CT persisted Differences amongmeans are significant (one-way ANOVA Po00001 fol-lowed by Bonferroni tests for differences among meansSuperscripts indicate membership in same group) Mini-mum run is one in each case
CT Years SD Max
A 329b 135 7B 200ab 115 4C 107a 026 2D 238b 142 7E 181ab 117 7F 398c 274 10G 391c 172 6H 558d 256 10I 419cd 167 8J 378c 185 7
Fig 7 Detrended correspondence analysis of plots by CTand by year Differences (Po005) among measures de-termined by ANOVA followed by the Bonferroni testFor each set values with the same associated letter fallwithin the same group of values for DCA-1 (no data for1998 and 2002 alternate plots were sampled in 2007 and2008) The inset is a separate DCA of plots in 2008
Primary succession trajectories 863
significant (r2 5 0206 Po00001 Spearman non-parametric correlation r5 025 Po00001) Thevariation in DCA-1 scores within years declined (thelinear correlation of years vs standard deviationwas Po00001 t5 533 r2 5 0637)
The mean DCA position of the 10 CTs did notreveal a single trajectory CT-C (Anaphalis) wasmore closely associated with pioneer CTs but wasnever the first CT on a plot CT-G (Penstemon-Agrostis-Cistanthe) was aligned with late-seral CTsWe also conducted a DCA of 2008 plots (Fig 7inset) b-diversity was low with l5 097 (DCA-15
133 DCA-25 84 ca 18 HC along DCA-1)No trajectory was evident from this analysis
Discussion
There are so few long-term longitudinal studiesof early primary succession on stressful sites be-cause initially so little happens The vegetation ofthe Abraham Plain remains sparse 30 years after theeruption It has developed by interwoven succes-sional trajectories that include plots that divergeconverge regress and progress to form a braidedsuccession Neither habitat factors nor competitionappears to have provided strong filtering effects onspecies composition although unmeasured soilvariables (eg moisture and nitrogen content) andfactors related to stochastic colonization and estab-lishment (cf del Moral amp Ellis 2004) may explainsome of the remaining unexplained variation Theimmigration seed density was low (Wood amp delMoral 2000) many suitable sites appeared vacant inearly years (del Moral amp Bliss 1993) and severalspecies occupied similar sites Chance may governsuch patterns We propose that when environmentalvariables have only slight effects on species compo-sition consistent successional trajectories areunlikely to emerge Below we discuss questionsabout predictability rate and pattern of successionon Abraham Plain that are the focus of this study
Environmentndashvegetation connection
Predictable relationships between environ-mental variables and vegetation had developed by2008 but they remained low Priority effects sec-ondary disturbances elk grazing and seasonalfluctuations may affect these trajectories in the fu-ture rendering it unlikely that explained variationwould become the dominant factor before forestsdevelop
We did not use soil factors because plants es-tablish at a smaller scale than the plot size and it isimpractical to measure such factors as moisture ineven a few plots Although soils were young andhomogeneous it is likely that inclusion of soil fac-tors (eg pH and moisture) would have improvedexplanatory power based on similar studies (delMoral 2009a) explained variance might doubleTopographic surface and position factors alone diddemonstrate increasing explained variance of spe-cies pattern over time but this trend was lesspronounced than in other studies On the northslope of Mount St Helens in 1992 only spatial fac-tors explained any variation (15) but after 10 yrenvironmental factors dominated (28 del Moralamp Lacher 2005) Baasch et al (2009) produced oneof the very few primary succession studies to showthat relationships between vegetation and ex-planatory variables increased over time They toosuggested that the impact of stochastic processesdeclined over time
The spatial distribution of CTs suggested someenvironmental influence Rills provide havens forpioneers (eg Anaphalis Hieracium and Hy-pochaeris) Gullies offer protection from wind andaccumulate snow which allowed species such asAnaphalis and Salix to colonize and persist Pen-stemon and Juncus were common in more exposedsites where cover and diversity were least In suchsites species that are less stress tolerant (eg SalixPolytrichum Saxifraga and Juncus mertensianus)were uncommon while stress-tolerant species suchas Cistanthe Juncus parryi Agrostis pallens andCarex microptera reached their maximum values
Rates
Habitats with limited fertility or moisture typi-cally have low succession rates (Donnegan ampRebertus 1999 Anderson 2007) and succession onAbraham Plain has been no exception Mean annualDCA scores were in a single group of values Be-tween 1988 and 1999 these scores formed sevengroups Surrounding habitats on Mount St Helensdeveloped more rapidly On an exposed lahar on thesouth slope richness averaged 16 species and thecover index averaged 73 (del Moral 2009b) prior to2008 In contrast mean richness on the AbrahamPlain in 2008 was 108 and the mean cover index was314 The proximity to forest more fertile soils andabundance of the N-fixing Lupinus may all con-tribute to accelerated primary succession at suchsites relative to Abraham Plain
864 del Moral Roger et al
Trajectories
Succession trajectories have been discussedthoroughly (cf Leps amp Rejmanek 1991) When thevegetation of initially homogeneous sites developslocally distinctive characteristics divergence occurs(Bossuyt et al 2003) Divergent trajectories occur onmany surfaces including landslides glacier fore-lands wet slack dune vegetation floodplains andmine tailings (Walker amp del Moral 2003) Lanta ampLeps (2009) showed experimentally that differentialdispersal could promote divergence when there arepriority effects Divergence may be more likely instressful habitats when different species arrive firstand where climatic factors are highly variable(MacDougall et al 2008) Convergence describestrajectories that become more similar over timeConvergence has been assumed to result when cli-matic factors dominate and there is evidence thatwhen dispersal effects are small and competitive ef-fects are strong convergence to a single communitydoes occur (van Oijen et al 2005 Anthelme et al2007) Convergence is also more likely when thereare few alternative stages as on glacial forelands(Hodkinson et al 2003 but see Robbins ampMathews2009)
Here we show directly that early succession un-der stressful conditions can include trajectories thatdiverge converge regress and progress to form areticulate succession Our study joins a growingbody of work that demonstrates the frequency ofmultiple trajectories in early primary succession (egdel Moral 2007 Prach amp Hobbs 2008 Walker amp delMoral 2009) Forecasting trajectories and identify-ing factors that limit their development is crucial tounderstanding community assembly mechanismsOn the Abraham Plain persistent species graduallyreplaced readily dispersed pioneer species Subtlehabitat variation annual weather fluctuation andlandscape effects combined to produce communitiesrelated through a network of development and re-sponsible to topographic variation
The first colonists were species with excellentdispersal but isolation allowed for only a meagerseed rain they established pioneer communities thatsoon developed into other CTs characterized byspecies with greater persistence (cf Ejrnaes et al2006) Several alternative transitions were possiblefor most CTs suggesting that stochastic processesinitially dominated transitions Gradually persis-tent species invaded and attained dominance Minortopographic variation affected composition some-what resulting in some successional transitions thatare more common than others Of annual transi-
tions between successional stages 51 wereregressions and 170 were progressions DCA re-vealed no single trend among the CTs The two mid-seral community types were central to the transitionpatterns while the three late-seral CTs occurred indifferent directions from this centre
It is unlikely that complete convergence will oc-cur Even when trees dominate understoreyvegetation is likely to retain variation initiated dur-ing early succession and maintained by topographicvariation and priority effects The understorey offorests on the south side of Mount St Helens de-monstrates such variation (del Moral amp Ellis 2004)There is no evidence yet for a single target commu-nity
Strong assembly rules are more likely to pro-duce convergent trajectories than weak ones Whilestrong assembly rules may exist in mature vegeta-tion (see Navas amp Violle 2009) they appear weak onAbraham Plain and are more effective for functionaltypes than species One rule may be that long-livedwind-dispersed species accumulate at the expense ofshort-lived pioneers A second may be that grami-noids with rhizomatous growth (eg Agrostispallens) and prostrate shrubs (eg Penstemon) arecomplementary and can coexist indefinitely A thirdmay be that fleshy fruit-producing shrubs (eg Ru-bus spp Vaccinium spp) cannot establish undercurrent conditions Finally although seeds of con-ifers and Populus balsamifera are commonconditions do not yet favour their developmentThese rules are weak and exceptions are commonThe spontaneously developing vegetation of theAbraham Plain is constrained by isolation from po-tential colonists as well as a habitat that challengesplant growth
Acknowledgements We thank the US National Science
Foundation for funding (BSR8906544 DEB9406987
DEB0087040 and DEB0541972) and the Mount St He-
lens National Volcanic Monument for permissions
Valuable comments by J G Bishop M P Fleming Jan-
neke Hille Ris Lambers L R Walker J H Titus and two
anonymous reviewers improved the paper Field assis-
tance was provided through the years by S Anderson W
Arnesen S Bard A Coogan S Crawford K Dlugosch
A Eckert E Ellis C Fairbourn C Muerdter M Flem-
ing T Fletcher R Fuller A Grant E Jenkins M
Johns C Jones L McMullan P Moody R Robham L
Rozzell J Sandler R Sewell-Nesteruk M Spasojevic
M Tweiten S Wilson B Witte and C Wolfe This is
contribution No 60 to the Mount St Helens Succession
Project
Primary succession trajectories 865
References
Anderson KJ 2007 Temporal patterns in rates of
community change during succession American
Naturalist 169 780ndash793
Anthelme F Villaret JC amp Brun JJ 2007 Shrub
encroachment in the Alps gives rise to the
convergence of sub-alpine communities on a regional
scale Journal of Vegetation Science 18 355ndash362
Baasch A Tischew S amp Bruelheide H 2009 Insights
into succession processes using temporally repeated
habitat models results from a long-term study in a
post-mining landscape Journal of Vegetation Science
20 629ndash638
Bossuyt B Honnay O amp Hermy M 2003 An island
biogeographical view of the successional pathway in
wet dune slacks Journal of Vegetation Science 14 781ndash
788
Clifford HT 1976 Dendrograms and their
interpretation In Williams WT (ed) Pattern
analysis in agricultural science pp 96ndash109 Elsevier
Science Amsterdam NL
del Moral R 2007 Vegetation dynamics in space and
time an example from Mount St Helens Journal of
Vegetation Science 18 479ndash488
del Moral R 2009a Increasing deterministic control of
primary succession onMount St Helens Washington
Journal of Vegetation Science 20 1145ndash1154
del Moral R 2009b Mount St Helens permanent plots
and grids 1980 to present Available at http
protistbiologywashingtonedudelmoral Accessed 8
December 2009
del Moral R amp Bliss LC 1993 Mechanism of primary
succession insights resulting from the eruption of
Mount St Helens Advances in Ecological Research
24 1ndash66
del Moral R amp Ellis EE 2004 Gradients in hetero-
geneity and structure on lahars Mount St Helens
Washington USA Plant Ecology 175 273ndash286
del Moral R amp Lacher IL 2005 Vegetation patterns 25
years after the eruption of Mount St Helens
WashingtonAmerican Journal of Botany 92 1948ndash1956
Donnegan JA amp Rebertus AJ 1999 Rates and
mechanisms of subalpine forest succession along an
environmental gradient Ecology 80 1370ndash1384
Ejrnaes R Bruun HH amp Graae BJ 2006 Community
assembly in experimental grasslands suitable
environment or timely arrival Ecology 87 1225ndash1233
Felinks B ampWiegand T 2008 Exploring spatiotemporal
patterns in early stages of primary succession on
former lignite mining sites Journal of Vegetation
Science 19 267ndash276
Fuller RN amp del Moral R 2003 The role of refugia and
dispersal in primary succession on Mount St Helens
Washington Journal of Vegetation Science 14 637ndash
644
Hodkinson IA Coulson SJ amp Webb NR 2003
Community assembly along proglacial chrono-
sequences in the high Arctic vegetation and soil
development in north-west Svalbard Journal of
Ecology 91 651ndash663
Johnson EA amp Miyanishi K 2008 Testing the
assumptions of chronosequences in succession
Ecology Letters 11 419ndash431
Lanta V amp Leps J 2009 How does surrounding
vegetation affect the course of succession a five-year
container experiment Journal of Vegetation Science
20 686ndash694
Legendre P amp Anderson MJ 1999 Distance-based
redundancy analysis testing multispecies responses in
multi-factorial ecological experiments Ecological
Monographs 69 1ndash24
Leps J amp Rejmanek M 1991 Convergence or
divergence what should we expect from vegetation
succession Oikos 62 261ndash264
Leps J amp Smilauer P 2003 Multivariate analysis of
ecological data using CANOCO Cambridge
University Press Cambridge UK
MacDougall AS Wilson SD amp Bakker JD 2008
Climatic variability alters the outcome of longterm
community assembly Journal of Ecology 96 346ndash354
Navas ML amp Violle C 2009 Plant traits related to
competition how do they shape the functional
diversity of communities Community Ecology 10
131ndash137
Pickett STA Cadenasso ML amp Meiners SJ 2009
Ever since Clements from succession to vegetation
dynamics and understanding to intervention Applied
Vegetation Science 12 9ndash21
Prach K amp Hobbs RJ 2008 Spontaneous succession
versus technical reclamation in the restoration of
disturbed sites Restoration Ecology 16 363ndash366
Prach K amp Rehounkova K 2006 Vegetation succession
over broad geographical scales which factors
determine the patterns Preslia 78 469ndash480
Robbins JA amp Matthews JA 2009 Pioneer vegetation
on glacier forelands in southern Norway emerging
communities Journal of Vegetation Science 20 889ndash
902
Swanson FJ amp Major JJ 2005 Physical events
environments and geologicalndashecological interactions
at Mount St Helens March 1980ndash2004 In Dale
VH Swanson FJ amp Crisafulli CM (eds)
Ecological recovery after the 1980 eruption of Mount
St Helens pp 27ndash44 Springer New York NY US
USDA National Resource Conservation Service 2009
The Plants Data Base National Plants Data Center
Baton Rouge LA US Available at http
plantsusdagov Accessed October 15 2009
van Oijen D Hommel P den Ouden J amp de Waal R
2005 Effects of tree species composition on within-
forest distribution of understorey species Applied
Vegetation Science 8 155ndash166
Walker LR amp del Moral R 2003 Primary succession
and ecosystem rehabilitation Cambridge University
Press Cambridge UK
866 del Moral Roger et al
Walker LR amp del Moral R 2009 Transition dynamics
in succession implications for rates trajectories and
restoration In Hobbs RJ amp Suding KN (eds)
New models for ecosystem dynamics and restoration
pp 33ndash49 Island Press Washington DC US
Walker LR Bellingham PJ amp Peltzer DA 2006
Plant characteristics are poor predictors of microsite
colonization during the first two years of pri-
mary succession Journal of Vegetation Science 17
397ndash406
Wood DM amp del Moral R 1988 Colonizing plants on
the Pumice Plains Mount St Helens Washington
American Journal of Botany 75 1228ndash1237
Wood DM amp del Moral R 2000 Seed rain during early
primary succession onMount St Helens Washington
Madrono 47 1ndash9
Supporting Information
Additional supporting information may befound in the online version of this article
Fig S1 Abraham Plain Sept 1980 pumice de-posits over scoured bedrock looking north towardsthe study site
Fig S2 Abraham Plain July 2009 lookingnorthwest across the study site
Fig S3 Changes in mean cover index for lesscommon species
Fig S4 Relative cover (proportion of totalcover in the sample) of less common species in thefour stages of early primary succession The firstfour species are persistent the last two are pioneerspecies (defined in Appendix S1)
Fig S5 Annual changes of mean H0 and meanSimpsonrsquos dominance index Differences (Po005)among measures determined by a repeat measuresANOVA followed by the Bonferroni test Valueswith same associated letter fall within the samegroup of values
Table S1 Structure and species composition ofCTs found in 2008
Appendix S1 Characteristics of species en-countered on Abraham Plain grid
Please note Wiley-Blackwell is not responsiblefor the content or functionality of any supportingmaterials supplied by the authors Any queries(other than missing material) should be directed tothe corresponding author for the article
Received 9 December 2009
Accepted 14 March 2010
Co-ordinating Editor Dr Kerry Woods
Primary succession trajectories 867

Cistanthe-Juncus n5 145) also had substantialLuetkea and Racomitrium Pioneer species were un-common and taller woody species (Abies spp andSalix) absent Richness cover and diversity were allleast of the persistent CTs
Species patterns
Many species increased in cover (Fig 2 Fig S2)during the study but cover of pioneer species de-clined after the mid-1990s In 2008 1093 speciesplot were wind-dispersed while only 011 speciesplot were animal-dispersed (Appendix S1) Pioneerspecies have effective long-distance dispersal me-chanisms and while seral species are also wind-
dispersed their dispersal abilities are more limited(Fuller amp del Moral 2003) Penstemon and Agrostispallens expanded to occupy all plots while Juncusparryi and Cistanthe umbellata occurred in mostLuetkea increased steadily but by 2008 it appearedto be stable Salix increased gradually and spor-adicallyRacomitrium occurred in four plots in 1988while Polytrichum did not occur until 1995 Carexmertensii and C microptera were widely distributedand occasionally abundant both occur on halfthe grid Saxifraga once common declined in2008 but this may be associated with dry summerconditions
Relative cover clearly shows the contrast be-tween pioneer and persistent species (Fig 3 Fig S3)
Table 1 Structure and mean cover index in 10 community types (CTs) Numbers in parentheses after species are occurrences(plotsyears) Values in bold indicate dominant species (high cover index) in that CT Each structural variable differedsignificantly among CTs (one-way ANOVA Po0001 values with same superscript fall within the same group as determinedby Bonferroni comparisons) Species cover index means also differed significantly (ANOVA Po0001) but patterns weretoo complex to display grouping patterns CTs and species were each arranged in order of their DCA scores
Structure CT-A CT-B CT-C CT-D CT-E CT-F CT-G CT-H CT-I CT-J
Richness 37a 31a 53b 95d 89c 109e 125f 127f 111e 91cd
Cover (sum indexscores)
52a 38a 70a 159c 136b 242d 321g 298f 281f 238e
Dominance 0516a 0575a 0734b 0851cde 0840c 0871def 0890ef 0890ef 0877def 0843cd
H0 1001a 0999a 1512b 2078d 2008cd 2204e 2347f 2361f 2227e 2001c
Number of plotsyears 1558 21 108 877 444 1289 219 1334 421 495First year observed 1988 1988 1988 1990 1989 1992 1996 1994 1997 1997Last year observed 2000 1991 1998 2001 2000 2008 2008 2008 2008 2008Hypochaeris radicata
(3011)0444 0000 0491 1197 1380 0497 0365 0412 0214 0040
Anaphalis margaritacea(5784)
1557 0810 1713 3304 3459 2370 2717 2147 1337 0802
Chamerionangustifolium (4712)
0514 0191 0593 1911 1803 1442 0991 1077 0691 0513
Hieracium albiflorum(3907)
0336 0143 0398 0904 0930 0929 0753 0813 0411 0293
Lupinus lepidus (248) 0007 1667 0028 0051 0220 0076 0383 0021 0088 0036Lupinus latifolius (122) o0001 0095 0000 o0001 0047 0020 0932 0037 0000 0000Abies sp (736) 0106 0000 0120 0123 0124 0087 0032 0352 1062 0020Luetkea pectinata
(4075)0182 0095 0157 0691 0916 1389 2078 2524 1981 1432
Carex mertensii (2691) 0128 0428 0046 0379 0373 0680 0827 1249 1057 0649Saxifraga ferruginea
(1596)0045 0095 0083 0276 0253 0632 1082 1079 0456 0247
Racomitrium canescens(2498)
0072 0000 0019 0243 0943 0421 2183 1080 1064 0990
Cistanthe umbellata(4867)
0123 0048 1370 2222 0523 3199 3603 3469 4033 4853
Agrostis pallens (5060) 0294 0000 0528 1331 0738 2855 3457 3271 3924 4301
Eriogonum pyrolifolium(653)
0016 0000 0000 0071 0057 0120 0338 0281 0591 0059
Juncus parryi (4421) 0116 0191 0250 1008 0550 2066 4064 2633 3565 3465
Penstemon cardwellii(4489)
0195 0000 0278 0520 0484 3165 4489 3591 4197 4216
Salix sitchensis (1858) 0037 0000 0000 0140 0188 0216 1237 3144 0271 0073Carex microptera
(1723)0035 0095 0065 0161 0091 0412 0763 0752 0644 1372
Polytrichumjuniperinum (1133)
o0001 0000 0000 0047 0093 0182 0182 1601 2183 0168
860 del Moral Roger et al
Species such as Penstemon Agrostis Juncus parryiCarex spp and Luetkea increased across the CTswhen listed by their first appearance These speciesare long-lived with strong vegetative growth
Fig 1 Structure of CTs from 1988 to 2008 (a) Number of plots (b) Mean species richness (c) Mean cover index (d) Meandiversity index (H0)
Fig 2 Changes in mean cover index for abundant species
Fig 3 Changes in relative cover (proportion of total cov-er in the sample) of common species in the four stages ofearly primary succession The first four species are persis-tent the last two are pioneer species (defined in AppendixS1)
Primary succession trajectories 861
In contrast pioneer species soon peaked and thendeclined
Habitat relationships
We could use only position topography andsurface conditions as explanatory variables in RDAExplained variation was low but increased four-fold from 1990 to 2006 (Table 2) In 1996 we ob-tained the first significant result when the y-axis(possibly related to distance from survivors) and rillfraction became significant By 2006 smooth sur-face also contributed but topography did notinfluence the vegetation
We recognized six habitat types (Table 3) A fivepersistent CT (Table S1) by six HT w2 test was sig-nificant (w2 5 935 Po00001 df 5 20) CT-F(Penstemon-Agrostis-Cistanthe) was more commonin rills CT-G (Penstemon-Agrostis-Juncus) wasmore common in gullies CT-H (Penstemon-Agros-tis-Salix) tended to avoid smooth plots and wasmore common in plots with rills or gullies and CT-J(Penstemon-Agrostis-Cistanthe-Juncus) tended tooccur in smooth plots but avoided rills or gullies
The spatial pattern displayed by the CTs re-flected some environmental sorting (Fig 4) Nearestneighbour distances compared to the randommodel suggested that CT-F formed scattered clus-ters (mean observed distance5 142 mean randomdistance5 224 t5 225 P5 003) Some plots ofCT-G were confined to the eastern edge while oth-ers were in the southwest corner but overall it wasnot clustered (observed5 190 random5 243t5 190 P5 006) CT-H was clustered (ob-served5 015 random5 058 t5 483 Po00001)as was CT-I (observed5 052 random5 145t5 448 Po00001) concentrated on the east halfof the grid CT-J was clustered (observed5 119random5 264 t5 383 P5 0002) and con-centrated on the eastern part of the grid
Trajectories
We explored vegetation trajectories in severalways In 1988 richness the mean cover index(Fig 5) H0 and dominance (Fig S4) were leastRichness increased until 1999 after which time itvaried around 11 species per plot The cover indexcontinued to increase while H0 and dominance sta-bilized by the mid-1990s
Succession began with colonization by highlyvagile pioneers such as Anaphalis Chamerion Hier-acium and Hypochaeris Plots changed into otherCTs at variable rates While we found transitionsamong all CTs some trajectories were more typicalthan were others (Fig 6) The most likely trajec-
Table 2 Summary of redundancy analysis results usingposition topography and surface variables on the gridThe alternate plots sampled in 2007 and 2008 were ana-lysed separately Percentage variation5 how much ofspecies variation is explained by variables F-values5 sig-nificance level of Axis 1 trace5 percentage variance onAxis 1 Correlations 405
Years Percentagevariation
F-value
Trace Correlatedvariables
1990 34 707 157 ndash1993 65 128 340 ndash1996 79 178 420 y rill1999 109 331 598 y rill2003 112 338 616 y rill2006 125 390 698 y rill flat2008 122 335 654 y rill flat
Table 3 Mean value of characteristics of six habitat types (HT) HTs differed significantly for each terrain characteristic(Po00001 one-way ANOVA followed by Bonferroni tests for differences among means Values with same superscript fallwithin the same group)
Terrain HT-1 HT-2 HT-3 HT-4 HT-5 HT-6 F-value(n5 128) (n5 28) (n5 37) (n5 64) (n5 61) (n5 82)
Smooth 300a 286a 289a 192bc 198b 180c 1911Gully 005b 036b 030b 203a 026b 222a 2085Rill 015d 029cd 057c 061c 238a 117b 1141Rock 000d 100bc 132b 080c 062c 216a 1466Sand 000b 000b 019b 002b 010b 090a 306Pumice 300a 300a 200c 261b 257b 128d 1470
Fig 4 Distribution of CTs on the grid in 2008
862 del Moral Roger et al
tories proceeded from pioneer plots dominated byAnaphalis to early seral types that had accumulatedCistanthe Agrostis and Luetkea These developedfurther into mid-seral communities when Penstemonbecame dominant and Juncus became widespreador they developed directly into late seral commu-nities dominated by Penstemon and Agrostis In2008 some mid-seral CTs persisted but most plotswere in a late-seral community that had changedlittle since 2005 For each annual interval 1988 to2008 a plot could persist in a CT progress to a moredeveloped one or regress to an earlier stage Therewere 301 retrogressions and 1014 progressions The
five extant CTs persisted significantly longer thandid the five vanished CTs (Table 4)
We clarified temporal vegetation changes usingDCA (Fig 7) Total variance (l) was 159 (DCA-15 155 DCA-25 74) Turnover on DCA-1was 53 half-changes on DCA-2 it was 39 Pioneerspecies had high values on DCA-1 species with lowDCA-1 values appeared later and were persistentMean DCA-1 position of the plots changed sig-nificantly between years (r2 5 0637 Po00001Spearman nonparametric correlation r5 079Po00001) DCA-1 scores by year changed rapidlythrough 2001 (ANOVA Po00001) then slowedDCA-2 changes ceased after 1997 but ANOVA was
Fig 5 Annual changes of mean species richness and meancover index Differences (Po005) among measures de-termined by a repeat measures ANOVA followed by theBonferroni test Values with the same associated letter fallwithin the same group of values
Fig 6 Trajectories showing the number of transitionsbetween CTs Numbers in parentheses after persistent CTsare the number of plots in 2008 Many arrows transitionnear the mid-point to differentiate between transitions toand from a given CT
Table 4 Number of continuous years (standard devia-tion) for which each CT persisted Differences amongmeans are significant (one-way ANOVA Po00001 fol-lowed by Bonferroni tests for differences among meansSuperscripts indicate membership in same group) Mini-mum run is one in each case
CT Years SD Max
A 329b 135 7B 200ab 115 4C 107a 026 2D 238b 142 7E 181ab 117 7F 398c 274 10G 391c 172 6H 558d 256 10I 419cd 167 8J 378c 185 7
Fig 7 Detrended correspondence analysis of plots by CTand by year Differences (Po005) among measures de-termined by ANOVA followed by the Bonferroni testFor each set values with the same associated letter fallwithin the same group of values for DCA-1 (no data for1998 and 2002 alternate plots were sampled in 2007 and2008) The inset is a separate DCA of plots in 2008
Primary succession trajectories 863
significant (r2 5 0206 Po00001 Spearman non-parametric correlation r5 025 Po00001) Thevariation in DCA-1 scores within years declined (thelinear correlation of years vs standard deviationwas Po00001 t5 533 r2 5 0637)
The mean DCA position of the 10 CTs did notreveal a single trajectory CT-C (Anaphalis) wasmore closely associated with pioneer CTs but wasnever the first CT on a plot CT-G (Penstemon-Agrostis-Cistanthe) was aligned with late-seral CTsWe also conducted a DCA of 2008 plots (Fig 7inset) b-diversity was low with l5 097 (DCA-15
133 DCA-25 84 ca 18 HC along DCA-1)No trajectory was evident from this analysis
Discussion
There are so few long-term longitudinal studiesof early primary succession on stressful sites be-cause initially so little happens The vegetation ofthe Abraham Plain remains sparse 30 years after theeruption It has developed by interwoven succes-sional trajectories that include plots that divergeconverge regress and progress to form a braidedsuccession Neither habitat factors nor competitionappears to have provided strong filtering effects onspecies composition although unmeasured soilvariables (eg moisture and nitrogen content) andfactors related to stochastic colonization and estab-lishment (cf del Moral amp Ellis 2004) may explainsome of the remaining unexplained variation Theimmigration seed density was low (Wood amp delMoral 2000) many suitable sites appeared vacant inearly years (del Moral amp Bliss 1993) and severalspecies occupied similar sites Chance may governsuch patterns We propose that when environmentalvariables have only slight effects on species compo-sition consistent successional trajectories areunlikely to emerge Below we discuss questionsabout predictability rate and pattern of successionon Abraham Plain that are the focus of this study
Environmentndashvegetation connection
Predictable relationships between environ-mental variables and vegetation had developed by2008 but they remained low Priority effects sec-ondary disturbances elk grazing and seasonalfluctuations may affect these trajectories in the fu-ture rendering it unlikely that explained variationwould become the dominant factor before forestsdevelop
We did not use soil factors because plants es-tablish at a smaller scale than the plot size and it isimpractical to measure such factors as moisture ineven a few plots Although soils were young andhomogeneous it is likely that inclusion of soil fac-tors (eg pH and moisture) would have improvedexplanatory power based on similar studies (delMoral 2009a) explained variance might doubleTopographic surface and position factors alone diddemonstrate increasing explained variance of spe-cies pattern over time but this trend was lesspronounced than in other studies On the northslope of Mount St Helens in 1992 only spatial fac-tors explained any variation (15) but after 10 yrenvironmental factors dominated (28 del Moralamp Lacher 2005) Baasch et al (2009) produced oneof the very few primary succession studies to showthat relationships between vegetation and ex-planatory variables increased over time They toosuggested that the impact of stochastic processesdeclined over time
The spatial distribution of CTs suggested someenvironmental influence Rills provide havens forpioneers (eg Anaphalis Hieracium and Hy-pochaeris) Gullies offer protection from wind andaccumulate snow which allowed species such asAnaphalis and Salix to colonize and persist Pen-stemon and Juncus were common in more exposedsites where cover and diversity were least In suchsites species that are less stress tolerant (eg SalixPolytrichum Saxifraga and Juncus mertensianus)were uncommon while stress-tolerant species suchas Cistanthe Juncus parryi Agrostis pallens andCarex microptera reached their maximum values
Rates
Habitats with limited fertility or moisture typi-cally have low succession rates (Donnegan ampRebertus 1999 Anderson 2007) and succession onAbraham Plain has been no exception Mean annualDCA scores were in a single group of values Be-tween 1988 and 1999 these scores formed sevengroups Surrounding habitats on Mount St Helensdeveloped more rapidly On an exposed lahar on thesouth slope richness averaged 16 species and thecover index averaged 73 (del Moral 2009b) prior to2008 In contrast mean richness on the AbrahamPlain in 2008 was 108 and the mean cover index was314 The proximity to forest more fertile soils andabundance of the N-fixing Lupinus may all con-tribute to accelerated primary succession at suchsites relative to Abraham Plain
864 del Moral Roger et al
Trajectories
Succession trajectories have been discussedthoroughly (cf Leps amp Rejmanek 1991) When thevegetation of initially homogeneous sites developslocally distinctive characteristics divergence occurs(Bossuyt et al 2003) Divergent trajectories occur onmany surfaces including landslides glacier fore-lands wet slack dune vegetation floodplains andmine tailings (Walker amp del Moral 2003) Lanta ampLeps (2009) showed experimentally that differentialdispersal could promote divergence when there arepriority effects Divergence may be more likely instressful habitats when different species arrive firstand where climatic factors are highly variable(MacDougall et al 2008) Convergence describestrajectories that become more similar over timeConvergence has been assumed to result when cli-matic factors dominate and there is evidence thatwhen dispersal effects are small and competitive ef-fects are strong convergence to a single communitydoes occur (van Oijen et al 2005 Anthelme et al2007) Convergence is also more likely when thereare few alternative stages as on glacial forelands(Hodkinson et al 2003 but see Robbins ampMathews2009)
Here we show directly that early succession un-der stressful conditions can include trajectories thatdiverge converge regress and progress to form areticulate succession Our study joins a growingbody of work that demonstrates the frequency ofmultiple trajectories in early primary succession (egdel Moral 2007 Prach amp Hobbs 2008 Walker amp delMoral 2009) Forecasting trajectories and identify-ing factors that limit their development is crucial tounderstanding community assembly mechanismsOn the Abraham Plain persistent species graduallyreplaced readily dispersed pioneer species Subtlehabitat variation annual weather fluctuation andlandscape effects combined to produce communitiesrelated through a network of development and re-sponsible to topographic variation
The first colonists were species with excellentdispersal but isolation allowed for only a meagerseed rain they established pioneer communities thatsoon developed into other CTs characterized byspecies with greater persistence (cf Ejrnaes et al2006) Several alternative transitions were possiblefor most CTs suggesting that stochastic processesinitially dominated transitions Gradually persis-tent species invaded and attained dominance Minortopographic variation affected composition some-what resulting in some successional transitions thatare more common than others Of annual transi-
tions between successional stages 51 wereregressions and 170 were progressions DCA re-vealed no single trend among the CTs The two mid-seral community types were central to the transitionpatterns while the three late-seral CTs occurred indifferent directions from this centre
It is unlikely that complete convergence will oc-cur Even when trees dominate understoreyvegetation is likely to retain variation initiated dur-ing early succession and maintained by topographicvariation and priority effects The understorey offorests on the south side of Mount St Helens de-monstrates such variation (del Moral amp Ellis 2004)There is no evidence yet for a single target commu-nity
Strong assembly rules are more likely to pro-duce convergent trajectories than weak ones Whilestrong assembly rules may exist in mature vegeta-tion (see Navas amp Violle 2009) they appear weak onAbraham Plain and are more effective for functionaltypes than species One rule may be that long-livedwind-dispersed species accumulate at the expense ofshort-lived pioneers A second may be that grami-noids with rhizomatous growth (eg Agrostispallens) and prostrate shrubs (eg Penstemon) arecomplementary and can coexist indefinitely A thirdmay be that fleshy fruit-producing shrubs (eg Ru-bus spp Vaccinium spp) cannot establish undercurrent conditions Finally although seeds of con-ifers and Populus balsamifera are commonconditions do not yet favour their developmentThese rules are weak and exceptions are commonThe spontaneously developing vegetation of theAbraham Plain is constrained by isolation from po-tential colonists as well as a habitat that challengesplant growth
Acknowledgements We thank the US National Science
Foundation for funding (BSR8906544 DEB9406987
DEB0087040 and DEB0541972) and the Mount St He-
lens National Volcanic Monument for permissions
Valuable comments by J G Bishop M P Fleming Jan-
neke Hille Ris Lambers L R Walker J H Titus and two
anonymous reviewers improved the paper Field assis-
tance was provided through the years by S Anderson W
Arnesen S Bard A Coogan S Crawford K Dlugosch
A Eckert E Ellis C Fairbourn C Muerdter M Flem-
ing T Fletcher R Fuller A Grant E Jenkins M
Johns C Jones L McMullan P Moody R Robham L
Rozzell J Sandler R Sewell-Nesteruk M Spasojevic
M Tweiten S Wilson B Witte and C Wolfe This is
contribution No 60 to the Mount St Helens Succession
Project
Primary succession trajectories 865
References
Anderson KJ 2007 Temporal patterns in rates of
community change during succession American
Naturalist 169 780ndash793
Anthelme F Villaret JC amp Brun JJ 2007 Shrub
encroachment in the Alps gives rise to the
convergence of sub-alpine communities on a regional
scale Journal of Vegetation Science 18 355ndash362
Baasch A Tischew S amp Bruelheide H 2009 Insights
into succession processes using temporally repeated
habitat models results from a long-term study in a
post-mining landscape Journal of Vegetation Science
20 629ndash638
Bossuyt B Honnay O amp Hermy M 2003 An island
biogeographical view of the successional pathway in
wet dune slacks Journal of Vegetation Science 14 781ndash
788
Clifford HT 1976 Dendrograms and their
interpretation In Williams WT (ed) Pattern
analysis in agricultural science pp 96ndash109 Elsevier
Science Amsterdam NL
del Moral R 2007 Vegetation dynamics in space and
time an example from Mount St Helens Journal of
Vegetation Science 18 479ndash488
del Moral R 2009a Increasing deterministic control of
primary succession onMount St Helens Washington
Journal of Vegetation Science 20 1145ndash1154
del Moral R 2009b Mount St Helens permanent plots
and grids 1980 to present Available at http
protistbiologywashingtonedudelmoral Accessed 8
December 2009
del Moral R amp Bliss LC 1993 Mechanism of primary
succession insights resulting from the eruption of
Mount St Helens Advances in Ecological Research
24 1ndash66
del Moral R amp Ellis EE 2004 Gradients in hetero-
geneity and structure on lahars Mount St Helens
Washington USA Plant Ecology 175 273ndash286
del Moral R amp Lacher IL 2005 Vegetation patterns 25
years after the eruption of Mount St Helens
WashingtonAmerican Journal of Botany 92 1948ndash1956
Donnegan JA amp Rebertus AJ 1999 Rates and
mechanisms of subalpine forest succession along an
environmental gradient Ecology 80 1370ndash1384
Ejrnaes R Bruun HH amp Graae BJ 2006 Community
assembly in experimental grasslands suitable
environment or timely arrival Ecology 87 1225ndash1233
Felinks B ampWiegand T 2008 Exploring spatiotemporal
patterns in early stages of primary succession on
former lignite mining sites Journal of Vegetation
Science 19 267ndash276
Fuller RN amp del Moral R 2003 The role of refugia and
dispersal in primary succession on Mount St Helens
Washington Journal of Vegetation Science 14 637ndash
644
Hodkinson IA Coulson SJ amp Webb NR 2003
Community assembly along proglacial chrono-
sequences in the high Arctic vegetation and soil
development in north-west Svalbard Journal of
Ecology 91 651ndash663
Johnson EA amp Miyanishi K 2008 Testing the
assumptions of chronosequences in succession
Ecology Letters 11 419ndash431
Lanta V amp Leps J 2009 How does surrounding
vegetation affect the course of succession a five-year
container experiment Journal of Vegetation Science
20 686ndash694
Legendre P amp Anderson MJ 1999 Distance-based
redundancy analysis testing multispecies responses in
multi-factorial ecological experiments Ecological
Monographs 69 1ndash24
Leps J amp Rejmanek M 1991 Convergence or
divergence what should we expect from vegetation
succession Oikos 62 261ndash264
Leps J amp Smilauer P 2003 Multivariate analysis of
ecological data using CANOCO Cambridge
University Press Cambridge UK
MacDougall AS Wilson SD amp Bakker JD 2008
Climatic variability alters the outcome of longterm
community assembly Journal of Ecology 96 346ndash354
Navas ML amp Violle C 2009 Plant traits related to
competition how do they shape the functional
diversity of communities Community Ecology 10
131ndash137
Pickett STA Cadenasso ML amp Meiners SJ 2009
Ever since Clements from succession to vegetation
dynamics and understanding to intervention Applied
Vegetation Science 12 9ndash21
Prach K amp Hobbs RJ 2008 Spontaneous succession
versus technical reclamation in the restoration of
disturbed sites Restoration Ecology 16 363ndash366
Prach K amp Rehounkova K 2006 Vegetation succession
over broad geographical scales which factors
determine the patterns Preslia 78 469ndash480
Robbins JA amp Matthews JA 2009 Pioneer vegetation
on glacier forelands in southern Norway emerging
communities Journal of Vegetation Science 20 889ndash
902
Swanson FJ amp Major JJ 2005 Physical events
environments and geologicalndashecological interactions
at Mount St Helens March 1980ndash2004 In Dale
VH Swanson FJ amp Crisafulli CM (eds)
Ecological recovery after the 1980 eruption of Mount
St Helens pp 27ndash44 Springer New York NY US
USDA National Resource Conservation Service 2009
The Plants Data Base National Plants Data Center
Baton Rouge LA US Available at http
plantsusdagov Accessed October 15 2009
van Oijen D Hommel P den Ouden J amp de Waal R
2005 Effects of tree species composition on within-
forest distribution of understorey species Applied
Vegetation Science 8 155ndash166
Walker LR amp del Moral R 2003 Primary succession
and ecosystem rehabilitation Cambridge University
Press Cambridge UK
866 del Moral Roger et al
Walker LR amp del Moral R 2009 Transition dynamics
in succession implications for rates trajectories and
restoration In Hobbs RJ amp Suding KN (eds)
New models for ecosystem dynamics and restoration
pp 33ndash49 Island Press Washington DC US
Walker LR Bellingham PJ amp Peltzer DA 2006
Plant characteristics are poor predictors of microsite
colonization during the first two years of pri-
mary succession Journal of Vegetation Science 17
397ndash406
Wood DM amp del Moral R 1988 Colonizing plants on
the Pumice Plains Mount St Helens Washington
American Journal of Botany 75 1228ndash1237
Wood DM amp del Moral R 2000 Seed rain during early
primary succession onMount St Helens Washington
Madrono 47 1ndash9
Supporting Information
Additional supporting information may befound in the online version of this article
Fig S1 Abraham Plain Sept 1980 pumice de-posits over scoured bedrock looking north towardsthe study site
Fig S2 Abraham Plain July 2009 lookingnorthwest across the study site
Fig S3 Changes in mean cover index for lesscommon species
Fig S4 Relative cover (proportion of totalcover in the sample) of less common species in thefour stages of early primary succession The firstfour species are persistent the last two are pioneerspecies (defined in Appendix S1)
Fig S5 Annual changes of mean H0 and meanSimpsonrsquos dominance index Differences (Po005)among measures determined by a repeat measuresANOVA followed by the Bonferroni test Valueswith same associated letter fall within the samegroup of values
Table S1 Structure and species composition ofCTs found in 2008
Appendix S1 Characteristics of species en-countered on Abraham Plain grid
Please note Wiley-Blackwell is not responsiblefor the content or functionality of any supportingmaterials supplied by the authors Any queries(other than missing material) should be directed tothe corresponding author for the article
Received 9 December 2009
Accepted 14 March 2010
Co-ordinating Editor Dr Kerry Woods
Primary succession trajectories 867
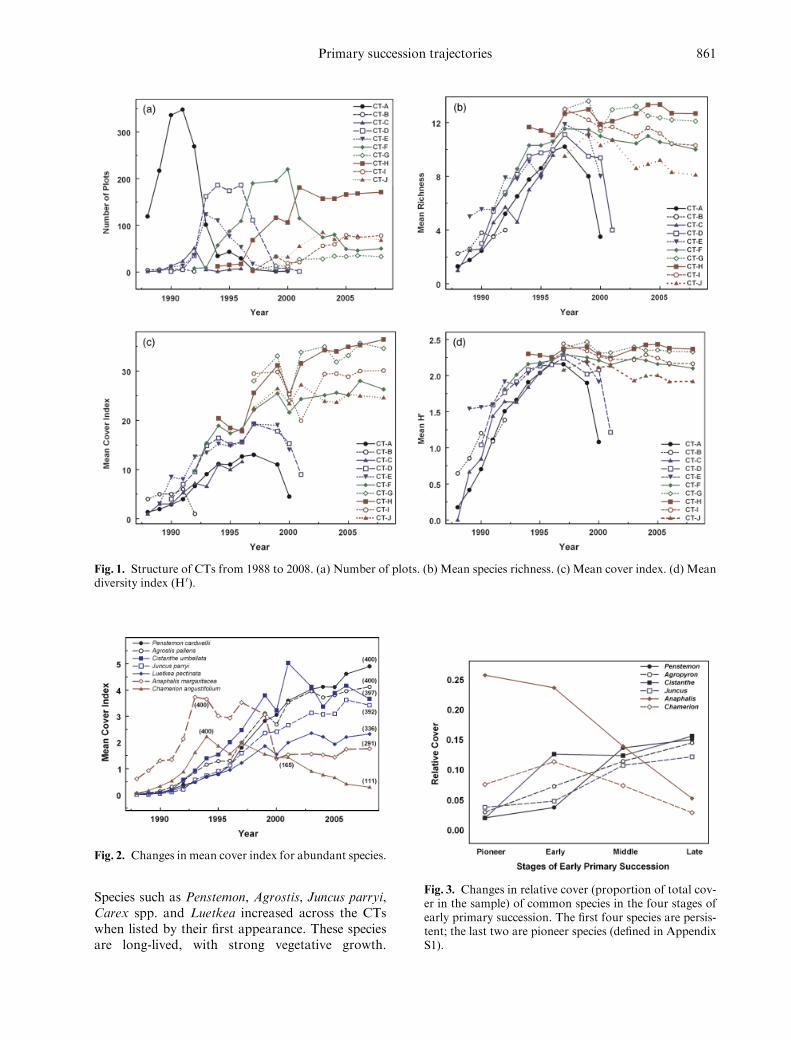
Species such as Penstemon Agrostis Juncus parryiCarex spp and Luetkea increased across the CTswhen listed by their first appearance These speciesare long-lived with strong vegetative growth
Fig 1 Structure of CTs from 1988 to 2008 (a) Number of plots (b) Mean species richness (c) Mean cover index (d) Meandiversity index (H0)
Fig 2 Changes in mean cover index for abundant species
Fig 3 Changes in relative cover (proportion of total cov-er in the sample) of common species in the four stages ofearly primary succession The first four species are persis-tent the last two are pioneer species (defined in AppendixS1)
Primary succession trajectories 861
In contrast pioneer species soon peaked and thendeclined
Habitat relationships
We could use only position topography andsurface conditions as explanatory variables in RDAExplained variation was low but increased four-fold from 1990 to 2006 (Table 2) In 1996 we ob-tained the first significant result when the y-axis(possibly related to distance from survivors) and rillfraction became significant By 2006 smooth sur-face also contributed but topography did notinfluence the vegetation
We recognized six habitat types (Table 3) A fivepersistent CT (Table S1) by six HT w2 test was sig-nificant (w2 5 935 Po00001 df 5 20) CT-F(Penstemon-Agrostis-Cistanthe) was more commonin rills CT-G (Penstemon-Agrostis-Juncus) wasmore common in gullies CT-H (Penstemon-Agros-tis-Salix) tended to avoid smooth plots and wasmore common in plots with rills or gullies and CT-J(Penstemon-Agrostis-Cistanthe-Juncus) tended tooccur in smooth plots but avoided rills or gullies
The spatial pattern displayed by the CTs re-flected some environmental sorting (Fig 4) Nearestneighbour distances compared to the randommodel suggested that CT-F formed scattered clus-ters (mean observed distance5 142 mean randomdistance5 224 t5 225 P5 003) Some plots ofCT-G were confined to the eastern edge while oth-ers were in the southwest corner but overall it wasnot clustered (observed5 190 random5 243t5 190 P5 006) CT-H was clustered (ob-served5 015 random5 058 t5 483 Po00001)as was CT-I (observed5 052 random5 145t5 448 Po00001) concentrated on the east halfof the grid CT-J was clustered (observed5 119random5 264 t5 383 P5 0002) and con-centrated on the eastern part of the grid
Trajectories
We explored vegetation trajectories in severalways In 1988 richness the mean cover index(Fig 5) H0 and dominance (Fig S4) were leastRichness increased until 1999 after which time itvaried around 11 species per plot The cover indexcontinued to increase while H0 and dominance sta-bilized by the mid-1990s
Succession began with colonization by highlyvagile pioneers such as Anaphalis Chamerion Hier-acium and Hypochaeris Plots changed into otherCTs at variable rates While we found transitionsamong all CTs some trajectories were more typicalthan were others (Fig 6) The most likely trajec-
Table 2 Summary of redundancy analysis results usingposition topography and surface variables on the gridThe alternate plots sampled in 2007 and 2008 were ana-lysed separately Percentage variation5 how much ofspecies variation is explained by variables F-values5 sig-nificance level of Axis 1 trace5 percentage variance onAxis 1 Correlations 405
Years Percentagevariation
F-value
Trace Correlatedvariables
1990 34 707 157 ndash1993 65 128 340 ndash1996 79 178 420 y rill1999 109 331 598 y rill2003 112 338 616 y rill2006 125 390 698 y rill flat2008 122 335 654 y rill flat
Table 3 Mean value of characteristics of six habitat types (HT) HTs differed significantly for each terrain characteristic(Po00001 one-way ANOVA followed by Bonferroni tests for differences among means Values with same superscript fallwithin the same group)
Terrain HT-1 HT-2 HT-3 HT-4 HT-5 HT-6 F-value(n5 128) (n5 28) (n5 37) (n5 64) (n5 61) (n5 82)
Smooth 300a 286a 289a 192bc 198b 180c 1911Gully 005b 036b 030b 203a 026b 222a 2085Rill 015d 029cd 057c 061c 238a 117b 1141Rock 000d 100bc 132b 080c 062c 216a 1466Sand 000b 000b 019b 002b 010b 090a 306Pumice 300a 300a 200c 261b 257b 128d 1470
Fig 4 Distribution of CTs on the grid in 2008
862 del Moral Roger et al
tories proceeded from pioneer plots dominated byAnaphalis to early seral types that had accumulatedCistanthe Agrostis and Luetkea These developedfurther into mid-seral communities when Penstemonbecame dominant and Juncus became widespreador they developed directly into late seral commu-nities dominated by Penstemon and Agrostis In2008 some mid-seral CTs persisted but most plotswere in a late-seral community that had changedlittle since 2005 For each annual interval 1988 to2008 a plot could persist in a CT progress to a moredeveloped one or regress to an earlier stage Therewere 301 retrogressions and 1014 progressions The
five extant CTs persisted significantly longer thandid the five vanished CTs (Table 4)
We clarified temporal vegetation changes usingDCA (Fig 7) Total variance (l) was 159 (DCA-15 155 DCA-25 74) Turnover on DCA-1was 53 half-changes on DCA-2 it was 39 Pioneerspecies had high values on DCA-1 species with lowDCA-1 values appeared later and were persistentMean DCA-1 position of the plots changed sig-nificantly between years (r2 5 0637 Po00001Spearman nonparametric correlation r5 079Po00001) DCA-1 scores by year changed rapidlythrough 2001 (ANOVA Po00001) then slowedDCA-2 changes ceased after 1997 but ANOVA was
Fig 5 Annual changes of mean species richness and meancover index Differences (Po005) among measures de-termined by a repeat measures ANOVA followed by theBonferroni test Values with the same associated letter fallwithin the same group of values
Fig 6 Trajectories showing the number of transitionsbetween CTs Numbers in parentheses after persistent CTsare the number of plots in 2008 Many arrows transitionnear the mid-point to differentiate between transitions toand from a given CT
Table 4 Number of continuous years (standard devia-tion) for which each CT persisted Differences amongmeans are significant (one-way ANOVA Po00001 fol-lowed by Bonferroni tests for differences among meansSuperscripts indicate membership in same group) Mini-mum run is one in each case
CT Years SD Max
A 329b 135 7B 200ab 115 4C 107a 026 2D 238b 142 7E 181ab 117 7F 398c 274 10G 391c 172 6H 558d 256 10I 419cd 167 8J 378c 185 7
Fig 7 Detrended correspondence analysis of plots by CTand by year Differences (Po005) among measures de-termined by ANOVA followed by the Bonferroni testFor each set values with the same associated letter fallwithin the same group of values for DCA-1 (no data for1998 and 2002 alternate plots were sampled in 2007 and2008) The inset is a separate DCA of plots in 2008
Primary succession trajectories 863
significant (r2 5 0206 Po00001 Spearman non-parametric correlation r5 025 Po00001) Thevariation in DCA-1 scores within years declined (thelinear correlation of years vs standard deviationwas Po00001 t5 533 r2 5 0637)
The mean DCA position of the 10 CTs did notreveal a single trajectory CT-C (Anaphalis) wasmore closely associated with pioneer CTs but wasnever the first CT on a plot CT-G (Penstemon-Agrostis-Cistanthe) was aligned with late-seral CTsWe also conducted a DCA of 2008 plots (Fig 7inset) b-diversity was low with l5 097 (DCA-15
133 DCA-25 84 ca 18 HC along DCA-1)No trajectory was evident from this analysis
Discussion
There are so few long-term longitudinal studiesof early primary succession on stressful sites be-cause initially so little happens The vegetation ofthe Abraham Plain remains sparse 30 years after theeruption It has developed by interwoven succes-sional trajectories that include plots that divergeconverge regress and progress to form a braidedsuccession Neither habitat factors nor competitionappears to have provided strong filtering effects onspecies composition although unmeasured soilvariables (eg moisture and nitrogen content) andfactors related to stochastic colonization and estab-lishment (cf del Moral amp Ellis 2004) may explainsome of the remaining unexplained variation Theimmigration seed density was low (Wood amp delMoral 2000) many suitable sites appeared vacant inearly years (del Moral amp Bliss 1993) and severalspecies occupied similar sites Chance may governsuch patterns We propose that when environmentalvariables have only slight effects on species compo-sition consistent successional trajectories areunlikely to emerge Below we discuss questionsabout predictability rate and pattern of successionon Abraham Plain that are the focus of this study
Environmentndashvegetation connection
Predictable relationships between environ-mental variables and vegetation had developed by2008 but they remained low Priority effects sec-ondary disturbances elk grazing and seasonalfluctuations may affect these trajectories in the fu-ture rendering it unlikely that explained variationwould become the dominant factor before forestsdevelop
We did not use soil factors because plants es-tablish at a smaller scale than the plot size and it isimpractical to measure such factors as moisture ineven a few plots Although soils were young andhomogeneous it is likely that inclusion of soil fac-tors (eg pH and moisture) would have improvedexplanatory power based on similar studies (delMoral 2009a) explained variance might doubleTopographic surface and position factors alone diddemonstrate increasing explained variance of spe-cies pattern over time but this trend was lesspronounced than in other studies On the northslope of Mount St Helens in 1992 only spatial fac-tors explained any variation (15) but after 10 yrenvironmental factors dominated (28 del Moralamp Lacher 2005) Baasch et al (2009) produced oneof the very few primary succession studies to showthat relationships between vegetation and ex-planatory variables increased over time They toosuggested that the impact of stochastic processesdeclined over time
The spatial distribution of CTs suggested someenvironmental influence Rills provide havens forpioneers (eg Anaphalis Hieracium and Hy-pochaeris) Gullies offer protection from wind andaccumulate snow which allowed species such asAnaphalis and Salix to colonize and persist Pen-stemon and Juncus were common in more exposedsites where cover and diversity were least In suchsites species that are less stress tolerant (eg SalixPolytrichum Saxifraga and Juncus mertensianus)were uncommon while stress-tolerant species suchas Cistanthe Juncus parryi Agrostis pallens andCarex microptera reached their maximum values
Rates
Habitats with limited fertility or moisture typi-cally have low succession rates (Donnegan ampRebertus 1999 Anderson 2007) and succession onAbraham Plain has been no exception Mean annualDCA scores were in a single group of values Be-tween 1988 and 1999 these scores formed sevengroups Surrounding habitats on Mount St Helensdeveloped more rapidly On an exposed lahar on thesouth slope richness averaged 16 species and thecover index averaged 73 (del Moral 2009b) prior to2008 In contrast mean richness on the AbrahamPlain in 2008 was 108 and the mean cover index was314 The proximity to forest more fertile soils andabundance of the N-fixing Lupinus may all con-tribute to accelerated primary succession at suchsites relative to Abraham Plain
864 del Moral Roger et al
Trajectories
Succession trajectories have been discussedthoroughly (cf Leps amp Rejmanek 1991) When thevegetation of initially homogeneous sites developslocally distinctive characteristics divergence occurs(Bossuyt et al 2003) Divergent trajectories occur onmany surfaces including landslides glacier fore-lands wet slack dune vegetation floodplains andmine tailings (Walker amp del Moral 2003) Lanta ampLeps (2009) showed experimentally that differentialdispersal could promote divergence when there arepriority effects Divergence may be more likely instressful habitats when different species arrive firstand where climatic factors are highly variable(MacDougall et al 2008) Convergence describestrajectories that become more similar over timeConvergence has been assumed to result when cli-matic factors dominate and there is evidence thatwhen dispersal effects are small and competitive ef-fects are strong convergence to a single communitydoes occur (van Oijen et al 2005 Anthelme et al2007) Convergence is also more likely when thereare few alternative stages as on glacial forelands(Hodkinson et al 2003 but see Robbins ampMathews2009)
Here we show directly that early succession un-der stressful conditions can include trajectories thatdiverge converge regress and progress to form areticulate succession Our study joins a growingbody of work that demonstrates the frequency ofmultiple trajectories in early primary succession (egdel Moral 2007 Prach amp Hobbs 2008 Walker amp delMoral 2009) Forecasting trajectories and identify-ing factors that limit their development is crucial tounderstanding community assembly mechanismsOn the Abraham Plain persistent species graduallyreplaced readily dispersed pioneer species Subtlehabitat variation annual weather fluctuation andlandscape effects combined to produce communitiesrelated through a network of development and re-sponsible to topographic variation
The first colonists were species with excellentdispersal but isolation allowed for only a meagerseed rain they established pioneer communities thatsoon developed into other CTs characterized byspecies with greater persistence (cf Ejrnaes et al2006) Several alternative transitions were possiblefor most CTs suggesting that stochastic processesinitially dominated transitions Gradually persis-tent species invaded and attained dominance Minortopographic variation affected composition some-what resulting in some successional transitions thatare more common than others Of annual transi-
tions between successional stages 51 wereregressions and 170 were progressions DCA re-vealed no single trend among the CTs The two mid-seral community types were central to the transitionpatterns while the three late-seral CTs occurred indifferent directions from this centre
It is unlikely that complete convergence will oc-cur Even when trees dominate understoreyvegetation is likely to retain variation initiated dur-ing early succession and maintained by topographicvariation and priority effects The understorey offorests on the south side of Mount St Helens de-monstrates such variation (del Moral amp Ellis 2004)There is no evidence yet for a single target commu-nity
Strong assembly rules are more likely to pro-duce convergent trajectories than weak ones Whilestrong assembly rules may exist in mature vegeta-tion (see Navas amp Violle 2009) they appear weak onAbraham Plain and are more effective for functionaltypes than species One rule may be that long-livedwind-dispersed species accumulate at the expense ofshort-lived pioneers A second may be that grami-noids with rhizomatous growth (eg Agrostispallens) and prostrate shrubs (eg Penstemon) arecomplementary and can coexist indefinitely A thirdmay be that fleshy fruit-producing shrubs (eg Ru-bus spp Vaccinium spp) cannot establish undercurrent conditions Finally although seeds of con-ifers and Populus balsamifera are commonconditions do not yet favour their developmentThese rules are weak and exceptions are commonThe spontaneously developing vegetation of theAbraham Plain is constrained by isolation from po-tential colonists as well as a habitat that challengesplant growth
Acknowledgements We thank the US National Science
Foundation for funding (BSR8906544 DEB9406987
DEB0087040 and DEB0541972) and the Mount St He-
lens National Volcanic Monument for permissions
Valuable comments by J G Bishop M P Fleming Jan-
neke Hille Ris Lambers L R Walker J H Titus and two
anonymous reviewers improved the paper Field assis-
tance was provided through the years by S Anderson W
Arnesen S Bard A Coogan S Crawford K Dlugosch
A Eckert E Ellis C Fairbourn C Muerdter M Flem-
ing T Fletcher R Fuller A Grant E Jenkins M
Johns C Jones L McMullan P Moody R Robham L
Rozzell J Sandler R Sewell-Nesteruk M Spasojevic
M Tweiten S Wilson B Witte and C Wolfe This is
contribution No 60 to the Mount St Helens Succession
Project
Primary succession trajectories 865
References
Anderson KJ 2007 Temporal patterns in rates of
community change during succession American
Naturalist 169 780ndash793
Anthelme F Villaret JC amp Brun JJ 2007 Shrub
encroachment in the Alps gives rise to the
convergence of sub-alpine communities on a regional
scale Journal of Vegetation Science 18 355ndash362
Baasch A Tischew S amp Bruelheide H 2009 Insights
into succession processes using temporally repeated
habitat models results from a long-term study in a
post-mining landscape Journal of Vegetation Science
20 629ndash638
Bossuyt B Honnay O amp Hermy M 2003 An island
biogeographical view of the successional pathway in
wet dune slacks Journal of Vegetation Science 14 781ndash
788
Clifford HT 1976 Dendrograms and their
interpretation In Williams WT (ed) Pattern
analysis in agricultural science pp 96ndash109 Elsevier
Science Amsterdam NL
del Moral R 2007 Vegetation dynamics in space and
time an example from Mount St Helens Journal of
Vegetation Science 18 479ndash488
del Moral R 2009a Increasing deterministic control of
primary succession onMount St Helens Washington
Journal of Vegetation Science 20 1145ndash1154
del Moral R 2009b Mount St Helens permanent plots
and grids 1980 to present Available at http
protistbiologywashingtonedudelmoral Accessed 8
December 2009
del Moral R amp Bliss LC 1993 Mechanism of primary
succession insights resulting from the eruption of
Mount St Helens Advances in Ecological Research
24 1ndash66
del Moral R amp Ellis EE 2004 Gradients in hetero-
geneity and structure on lahars Mount St Helens
Washington USA Plant Ecology 175 273ndash286
del Moral R amp Lacher IL 2005 Vegetation patterns 25
years after the eruption of Mount St Helens
WashingtonAmerican Journal of Botany 92 1948ndash1956
Donnegan JA amp Rebertus AJ 1999 Rates and
mechanisms of subalpine forest succession along an
environmental gradient Ecology 80 1370ndash1384
Ejrnaes R Bruun HH amp Graae BJ 2006 Community
assembly in experimental grasslands suitable
environment or timely arrival Ecology 87 1225ndash1233
Felinks B ampWiegand T 2008 Exploring spatiotemporal
patterns in early stages of primary succession on
former lignite mining sites Journal of Vegetation
Science 19 267ndash276
Fuller RN amp del Moral R 2003 The role of refugia and
dispersal in primary succession on Mount St Helens
Washington Journal of Vegetation Science 14 637ndash
644
Hodkinson IA Coulson SJ amp Webb NR 2003
Community assembly along proglacial chrono-
sequences in the high Arctic vegetation and soil
development in north-west Svalbard Journal of
Ecology 91 651ndash663
Johnson EA amp Miyanishi K 2008 Testing the
assumptions of chronosequences in succession
Ecology Letters 11 419ndash431
Lanta V amp Leps J 2009 How does surrounding
vegetation affect the course of succession a five-year
container experiment Journal of Vegetation Science
20 686ndash694
Legendre P amp Anderson MJ 1999 Distance-based
redundancy analysis testing multispecies responses in
multi-factorial ecological experiments Ecological
Monographs 69 1ndash24
Leps J amp Rejmanek M 1991 Convergence or
divergence what should we expect from vegetation
succession Oikos 62 261ndash264
Leps J amp Smilauer P 2003 Multivariate analysis of
ecological data using CANOCO Cambridge
University Press Cambridge UK
MacDougall AS Wilson SD amp Bakker JD 2008
Climatic variability alters the outcome of longterm
community assembly Journal of Ecology 96 346ndash354
Navas ML amp Violle C 2009 Plant traits related to
competition how do they shape the functional
diversity of communities Community Ecology 10
131ndash137
Pickett STA Cadenasso ML amp Meiners SJ 2009
Ever since Clements from succession to vegetation
dynamics and understanding to intervention Applied
Vegetation Science 12 9ndash21
Prach K amp Hobbs RJ 2008 Spontaneous succession
versus technical reclamation in the restoration of
disturbed sites Restoration Ecology 16 363ndash366
Prach K amp Rehounkova K 2006 Vegetation succession
over broad geographical scales which factors
determine the patterns Preslia 78 469ndash480
Robbins JA amp Matthews JA 2009 Pioneer vegetation
on glacier forelands in southern Norway emerging
communities Journal of Vegetation Science 20 889ndash
902
Swanson FJ amp Major JJ 2005 Physical events
environments and geologicalndashecological interactions
at Mount St Helens March 1980ndash2004 In Dale
VH Swanson FJ amp Crisafulli CM (eds)
Ecological recovery after the 1980 eruption of Mount
St Helens pp 27ndash44 Springer New York NY US
USDA National Resource Conservation Service 2009
The Plants Data Base National Plants Data Center
Baton Rouge LA US Available at http
plantsusdagov Accessed October 15 2009
van Oijen D Hommel P den Ouden J amp de Waal R
2005 Effects of tree species composition on within-
forest distribution of understorey species Applied
Vegetation Science 8 155ndash166
Walker LR amp del Moral R 2003 Primary succession
and ecosystem rehabilitation Cambridge University
Press Cambridge UK
866 del Moral Roger et al
Walker LR amp del Moral R 2009 Transition dynamics
in succession implications for rates trajectories and
restoration In Hobbs RJ amp Suding KN (eds)
New models for ecosystem dynamics and restoration
pp 33ndash49 Island Press Washington DC US
Walker LR Bellingham PJ amp Peltzer DA 2006
Plant characteristics are poor predictors of microsite
colonization during the first two years of pri-
mary succession Journal of Vegetation Science 17
397ndash406
Wood DM amp del Moral R 1988 Colonizing plants on
the Pumice Plains Mount St Helens Washington
American Journal of Botany 75 1228ndash1237
Wood DM amp del Moral R 2000 Seed rain during early
primary succession onMount St Helens Washington
Madrono 47 1ndash9
Supporting Information
Additional supporting information may befound in the online version of this article
Fig S1 Abraham Plain Sept 1980 pumice de-posits over scoured bedrock looking north towardsthe study site
Fig S2 Abraham Plain July 2009 lookingnorthwest across the study site
Fig S3 Changes in mean cover index for lesscommon species
Fig S4 Relative cover (proportion of totalcover in the sample) of less common species in thefour stages of early primary succession The firstfour species are persistent the last two are pioneerspecies (defined in Appendix S1)
Fig S5 Annual changes of mean H0 and meanSimpsonrsquos dominance index Differences (Po005)among measures determined by a repeat measuresANOVA followed by the Bonferroni test Valueswith same associated letter fall within the samegroup of values
Table S1 Structure and species composition ofCTs found in 2008
Appendix S1 Characteristics of species en-countered on Abraham Plain grid
Please note Wiley-Blackwell is not responsiblefor the content or functionality of any supportingmaterials supplied by the authors Any queries(other than missing material) should be directed tothe corresponding author for the article
Received 9 December 2009
Accepted 14 March 2010
Co-ordinating Editor Dr Kerry Woods
Primary succession trajectories 867

In contrast pioneer species soon peaked and thendeclined
Habitat relationships
We could use only position topography andsurface conditions as explanatory variables in RDAExplained variation was low but increased four-fold from 1990 to 2006 (Table 2) In 1996 we ob-tained the first significant result when the y-axis(possibly related to distance from survivors) and rillfraction became significant By 2006 smooth sur-face also contributed but topography did notinfluence the vegetation
We recognized six habitat types (Table 3) A fivepersistent CT (Table S1) by six HT w2 test was sig-nificant (w2 5 935 Po00001 df 5 20) CT-F(Penstemon-Agrostis-Cistanthe) was more commonin rills CT-G (Penstemon-Agrostis-Juncus) wasmore common in gullies CT-H (Penstemon-Agros-tis-Salix) tended to avoid smooth plots and wasmore common in plots with rills or gullies and CT-J(Penstemon-Agrostis-Cistanthe-Juncus) tended tooccur in smooth plots but avoided rills or gullies
The spatial pattern displayed by the CTs re-flected some environmental sorting (Fig 4) Nearestneighbour distances compared to the randommodel suggested that CT-F formed scattered clus-ters (mean observed distance5 142 mean randomdistance5 224 t5 225 P5 003) Some plots ofCT-G were confined to the eastern edge while oth-ers were in the southwest corner but overall it wasnot clustered (observed5 190 random5 243t5 190 P5 006) CT-H was clustered (ob-served5 015 random5 058 t5 483 Po00001)as was CT-I (observed5 052 random5 145t5 448 Po00001) concentrated on the east halfof the grid CT-J was clustered (observed5 119random5 264 t5 383 P5 0002) and con-centrated on the eastern part of the grid
Trajectories
We explored vegetation trajectories in severalways In 1988 richness the mean cover index(Fig 5) H0 and dominance (Fig S4) were leastRichness increased until 1999 after which time itvaried around 11 species per plot The cover indexcontinued to increase while H0 and dominance sta-bilized by the mid-1990s
Succession began with colonization by highlyvagile pioneers such as Anaphalis Chamerion Hier-acium and Hypochaeris Plots changed into otherCTs at variable rates While we found transitionsamong all CTs some trajectories were more typicalthan were others (Fig 6) The most likely trajec-
Table 2 Summary of redundancy analysis results usingposition topography and surface variables on the gridThe alternate plots sampled in 2007 and 2008 were ana-lysed separately Percentage variation5 how much ofspecies variation is explained by variables F-values5 sig-nificance level of Axis 1 trace5 percentage variance onAxis 1 Correlations 405
Years Percentagevariation
F-value
Trace Correlatedvariables
1990 34 707 157 ndash1993 65 128 340 ndash1996 79 178 420 y rill1999 109 331 598 y rill2003 112 338 616 y rill2006 125 390 698 y rill flat2008 122 335 654 y rill flat
Table 3 Mean value of characteristics of six habitat types (HT) HTs differed significantly for each terrain characteristic(Po00001 one-way ANOVA followed by Bonferroni tests for differences among means Values with same superscript fallwithin the same group)
Terrain HT-1 HT-2 HT-3 HT-4 HT-5 HT-6 F-value(n5 128) (n5 28) (n5 37) (n5 64) (n5 61) (n5 82)
Smooth 300a 286a 289a 192bc 198b 180c 1911Gully 005b 036b 030b 203a 026b 222a 2085Rill 015d 029cd 057c 061c 238a 117b 1141Rock 000d 100bc 132b 080c 062c 216a 1466Sand 000b 000b 019b 002b 010b 090a 306Pumice 300a 300a 200c 261b 257b 128d 1470
Fig 4 Distribution of CTs on the grid in 2008
862 del Moral Roger et al
tories proceeded from pioneer plots dominated byAnaphalis to early seral types that had accumulatedCistanthe Agrostis and Luetkea These developedfurther into mid-seral communities when Penstemonbecame dominant and Juncus became widespreador they developed directly into late seral commu-nities dominated by Penstemon and Agrostis In2008 some mid-seral CTs persisted but most plotswere in a late-seral community that had changedlittle since 2005 For each annual interval 1988 to2008 a plot could persist in a CT progress to a moredeveloped one or regress to an earlier stage Therewere 301 retrogressions and 1014 progressions The
five extant CTs persisted significantly longer thandid the five vanished CTs (Table 4)
We clarified temporal vegetation changes usingDCA (Fig 7) Total variance (l) was 159 (DCA-15 155 DCA-25 74) Turnover on DCA-1was 53 half-changes on DCA-2 it was 39 Pioneerspecies had high values on DCA-1 species with lowDCA-1 values appeared later and were persistentMean DCA-1 position of the plots changed sig-nificantly between years (r2 5 0637 Po00001Spearman nonparametric correlation r5 079Po00001) DCA-1 scores by year changed rapidlythrough 2001 (ANOVA Po00001) then slowedDCA-2 changes ceased after 1997 but ANOVA was
Fig 5 Annual changes of mean species richness and meancover index Differences (Po005) among measures de-termined by a repeat measures ANOVA followed by theBonferroni test Values with the same associated letter fallwithin the same group of values
Fig 6 Trajectories showing the number of transitionsbetween CTs Numbers in parentheses after persistent CTsare the number of plots in 2008 Many arrows transitionnear the mid-point to differentiate between transitions toand from a given CT
Table 4 Number of continuous years (standard devia-tion) for which each CT persisted Differences amongmeans are significant (one-way ANOVA Po00001 fol-lowed by Bonferroni tests for differences among meansSuperscripts indicate membership in same group) Mini-mum run is one in each case
CT Years SD Max
A 329b 135 7B 200ab 115 4C 107a 026 2D 238b 142 7E 181ab 117 7F 398c 274 10G 391c 172 6H 558d 256 10I 419cd 167 8J 378c 185 7
Fig 7 Detrended correspondence analysis of plots by CTand by year Differences (Po005) among measures de-termined by ANOVA followed by the Bonferroni testFor each set values with the same associated letter fallwithin the same group of values for DCA-1 (no data for1998 and 2002 alternate plots were sampled in 2007 and2008) The inset is a separate DCA of plots in 2008
Primary succession trajectories 863
significant (r2 5 0206 Po00001 Spearman non-parametric correlation r5 025 Po00001) Thevariation in DCA-1 scores within years declined (thelinear correlation of years vs standard deviationwas Po00001 t5 533 r2 5 0637)
The mean DCA position of the 10 CTs did notreveal a single trajectory CT-C (Anaphalis) wasmore closely associated with pioneer CTs but wasnever the first CT on a plot CT-G (Penstemon-Agrostis-Cistanthe) was aligned with late-seral CTsWe also conducted a DCA of 2008 plots (Fig 7inset) b-diversity was low with l5 097 (DCA-15
133 DCA-25 84 ca 18 HC along DCA-1)No trajectory was evident from this analysis
Discussion
There are so few long-term longitudinal studiesof early primary succession on stressful sites be-cause initially so little happens The vegetation ofthe Abraham Plain remains sparse 30 years after theeruption It has developed by interwoven succes-sional trajectories that include plots that divergeconverge regress and progress to form a braidedsuccession Neither habitat factors nor competitionappears to have provided strong filtering effects onspecies composition although unmeasured soilvariables (eg moisture and nitrogen content) andfactors related to stochastic colonization and estab-lishment (cf del Moral amp Ellis 2004) may explainsome of the remaining unexplained variation Theimmigration seed density was low (Wood amp delMoral 2000) many suitable sites appeared vacant inearly years (del Moral amp Bliss 1993) and severalspecies occupied similar sites Chance may governsuch patterns We propose that when environmentalvariables have only slight effects on species compo-sition consistent successional trajectories areunlikely to emerge Below we discuss questionsabout predictability rate and pattern of successionon Abraham Plain that are the focus of this study
Environmentndashvegetation connection
Predictable relationships between environ-mental variables and vegetation had developed by2008 but they remained low Priority effects sec-ondary disturbances elk grazing and seasonalfluctuations may affect these trajectories in the fu-ture rendering it unlikely that explained variationwould become the dominant factor before forestsdevelop
We did not use soil factors because plants es-tablish at a smaller scale than the plot size and it isimpractical to measure such factors as moisture ineven a few plots Although soils were young andhomogeneous it is likely that inclusion of soil fac-tors (eg pH and moisture) would have improvedexplanatory power based on similar studies (delMoral 2009a) explained variance might doubleTopographic surface and position factors alone diddemonstrate increasing explained variance of spe-cies pattern over time but this trend was lesspronounced than in other studies On the northslope of Mount St Helens in 1992 only spatial fac-tors explained any variation (15) but after 10 yrenvironmental factors dominated (28 del Moralamp Lacher 2005) Baasch et al (2009) produced oneof the very few primary succession studies to showthat relationships between vegetation and ex-planatory variables increased over time They toosuggested that the impact of stochastic processesdeclined over time
The spatial distribution of CTs suggested someenvironmental influence Rills provide havens forpioneers (eg Anaphalis Hieracium and Hy-pochaeris) Gullies offer protection from wind andaccumulate snow which allowed species such asAnaphalis and Salix to colonize and persist Pen-stemon and Juncus were common in more exposedsites where cover and diversity were least In suchsites species that are less stress tolerant (eg SalixPolytrichum Saxifraga and Juncus mertensianus)were uncommon while stress-tolerant species suchas Cistanthe Juncus parryi Agrostis pallens andCarex microptera reached their maximum values
Rates
Habitats with limited fertility or moisture typi-cally have low succession rates (Donnegan ampRebertus 1999 Anderson 2007) and succession onAbraham Plain has been no exception Mean annualDCA scores were in a single group of values Be-tween 1988 and 1999 these scores formed sevengroups Surrounding habitats on Mount St Helensdeveloped more rapidly On an exposed lahar on thesouth slope richness averaged 16 species and thecover index averaged 73 (del Moral 2009b) prior to2008 In contrast mean richness on the AbrahamPlain in 2008 was 108 and the mean cover index was314 The proximity to forest more fertile soils andabundance of the N-fixing Lupinus may all con-tribute to accelerated primary succession at suchsites relative to Abraham Plain
864 del Moral Roger et al
Trajectories
Succession trajectories have been discussedthoroughly (cf Leps amp Rejmanek 1991) When thevegetation of initially homogeneous sites developslocally distinctive characteristics divergence occurs(Bossuyt et al 2003) Divergent trajectories occur onmany surfaces including landslides glacier fore-lands wet slack dune vegetation floodplains andmine tailings (Walker amp del Moral 2003) Lanta ampLeps (2009) showed experimentally that differentialdispersal could promote divergence when there arepriority effects Divergence may be more likely instressful habitats when different species arrive firstand where climatic factors are highly variable(MacDougall et al 2008) Convergence describestrajectories that become more similar over timeConvergence has been assumed to result when cli-matic factors dominate and there is evidence thatwhen dispersal effects are small and competitive ef-fects are strong convergence to a single communitydoes occur (van Oijen et al 2005 Anthelme et al2007) Convergence is also more likely when thereare few alternative stages as on glacial forelands(Hodkinson et al 2003 but see Robbins ampMathews2009)
Here we show directly that early succession un-der stressful conditions can include trajectories thatdiverge converge regress and progress to form areticulate succession Our study joins a growingbody of work that demonstrates the frequency ofmultiple trajectories in early primary succession (egdel Moral 2007 Prach amp Hobbs 2008 Walker amp delMoral 2009) Forecasting trajectories and identify-ing factors that limit their development is crucial tounderstanding community assembly mechanismsOn the Abraham Plain persistent species graduallyreplaced readily dispersed pioneer species Subtlehabitat variation annual weather fluctuation andlandscape effects combined to produce communitiesrelated through a network of development and re-sponsible to topographic variation
The first colonists were species with excellentdispersal but isolation allowed for only a meagerseed rain they established pioneer communities thatsoon developed into other CTs characterized byspecies with greater persistence (cf Ejrnaes et al2006) Several alternative transitions were possiblefor most CTs suggesting that stochastic processesinitially dominated transitions Gradually persis-tent species invaded and attained dominance Minortopographic variation affected composition some-what resulting in some successional transitions thatare more common than others Of annual transi-
tions between successional stages 51 wereregressions and 170 were progressions DCA re-vealed no single trend among the CTs The two mid-seral community types were central to the transitionpatterns while the three late-seral CTs occurred indifferent directions from this centre
It is unlikely that complete convergence will oc-cur Even when trees dominate understoreyvegetation is likely to retain variation initiated dur-ing early succession and maintained by topographicvariation and priority effects The understorey offorests on the south side of Mount St Helens de-monstrates such variation (del Moral amp Ellis 2004)There is no evidence yet for a single target commu-nity
Strong assembly rules are more likely to pro-duce convergent trajectories than weak ones Whilestrong assembly rules may exist in mature vegeta-tion (see Navas amp Violle 2009) they appear weak onAbraham Plain and are more effective for functionaltypes than species One rule may be that long-livedwind-dispersed species accumulate at the expense ofshort-lived pioneers A second may be that grami-noids with rhizomatous growth (eg Agrostispallens) and prostrate shrubs (eg Penstemon) arecomplementary and can coexist indefinitely A thirdmay be that fleshy fruit-producing shrubs (eg Ru-bus spp Vaccinium spp) cannot establish undercurrent conditions Finally although seeds of con-ifers and Populus balsamifera are commonconditions do not yet favour their developmentThese rules are weak and exceptions are commonThe spontaneously developing vegetation of theAbraham Plain is constrained by isolation from po-tential colonists as well as a habitat that challengesplant growth
Acknowledgements We thank the US National Science
Foundation for funding (BSR8906544 DEB9406987
DEB0087040 and DEB0541972) and the Mount St He-
lens National Volcanic Monument for permissions
Valuable comments by J G Bishop M P Fleming Jan-
neke Hille Ris Lambers L R Walker J H Titus and two
anonymous reviewers improved the paper Field assis-
tance was provided through the years by S Anderson W
Arnesen S Bard A Coogan S Crawford K Dlugosch
A Eckert E Ellis C Fairbourn C Muerdter M Flem-
ing T Fletcher R Fuller A Grant E Jenkins M
Johns C Jones L McMullan P Moody R Robham L
Rozzell J Sandler R Sewell-Nesteruk M Spasojevic
M Tweiten S Wilson B Witte and C Wolfe This is
contribution No 60 to the Mount St Helens Succession
Project
Primary succession trajectories 865
References
Anderson KJ 2007 Temporal patterns in rates of
community change during succession American
Naturalist 169 780ndash793
Anthelme F Villaret JC amp Brun JJ 2007 Shrub
encroachment in the Alps gives rise to the
convergence of sub-alpine communities on a regional
scale Journal of Vegetation Science 18 355ndash362
Baasch A Tischew S amp Bruelheide H 2009 Insights
into succession processes using temporally repeated
habitat models results from a long-term study in a
post-mining landscape Journal of Vegetation Science
20 629ndash638
Bossuyt B Honnay O amp Hermy M 2003 An island
biogeographical view of the successional pathway in
wet dune slacks Journal of Vegetation Science 14 781ndash
788
Clifford HT 1976 Dendrograms and their
interpretation In Williams WT (ed) Pattern
analysis in agricultural science pp 96ndash109 Elsevier
Science Amsterdam NL
del Moral R 2007 Vegetation dynamics in space and
time an example from Mount St Helens Journal of
Vegetation Science 18 479ndash488
del Moral R 2009a Increasing deterministic control of
primary succession onMount St Helens Washington
Journal of Vegetation Science 20 1145ndash1154
del Moral R 2009b Mount St Helens permanent plots
and grids 1980 to present Available at http
protistbiologywashingtonedudelmoral Accessed 8
December 2009
del Moral R amp Bliss LC 1993 Mechanism of primary
succession insights resulting from the eruption of
Mount St Helens Advances in Ecological Research
24 1ndash66
del Moral R amp Ellis EE 2004 Gradients in hetero-
geneity and structure on lahars Mount St Helens
Washington USA Plant Ecology 175 273ndash286
del Moral R amp Lacher IL 2005 Vegetation patterns 25
years after the eruption of Mount St Helens
WashingtonAmerican Journal of Botany 92 1948ndash1956
Donnegan JA amp Rebertus AJ 1999 Rates and
mechanisms of subalpine forest succession along an
environmental gradient Ecology 80 1370ndash1384
Ejrnaes R Bruun HH amp Graae BJ 2006 Community
assembly in experimental grasslands suitable
environment or timely arrival Ecology 87 1225ndash1233
Felinks B ampWiegand T 2008 Exploring spatiotemporal
patterns in early stages of primary succession on
former lignite mining sites Journal of Vegetation
Science 19 267ndash276
Fuller RN amp del Moral R 2003 The role of refugia and
dispersal in primary succession on Mount St Helens
Washington Journal of Vegetation Science 14 637ndash
644
Hodkinson IA Coulson SJ amp Webb NR 2003
Community assembly along proglacial chrono-
sequences in the high Arctic vegetation and soil
development in north-west Svalbard Journal of
Ecology 91 651ndash663
Johnson EA amp Miyanishi K 2008 Testing the
assumptions of chronosequences in succession
Ecology Letters 11 419ndash431
Lanta V amp Leps J 2009 How does surrounding
vegetation affect the course of succession a five-year
container experiment Journal of Vegetation Science
20 686ndash694
Legendre P amp Anderson MJ 1999 Distance-based
redundancy analysis testing multispecies responses in
multi-factorial ecological experiments Ecological
Monographs 69 1ndash24
Leps J amp Rejmanek M 1991 Convergence or
divergence what should we expect from vegetation
succession Oikos 62 261ndash264
Leps J amp Smilauer P 2003 Multivariate analysis of
ecological data using CANOCO Cambridge
University Press Cambridge UK
MacDougall AS Wilson SD amp Bakker JD 2008
Climatic variability alters the outcome of longterm
community assembly Journal of Ecology 96 346ndash354
Navas ML amp Violle C 2009 Plant traits related to
competition how do they shape the functional
diversity of communities Community Ecology 10
131ndash137
Pickett STA Cadenasso ML amp Meiners SJ 2009
Ever since Clements from succession to vegetation
dynamics and understanding to intervention Applied
Vegetation Science 12 9ndash21
Prach K amp Hobbs RJ 2008 Spontaneous succession
versus technical reclamation in the restoration of
disturbed sites Restoration Ecology 16 363ndash366
Prach K amp Rehounkova K 2006 Vegetation succession
over broad geographical scales which factors
determine the patterns Preslia 78 469ndash480
Robbins JA amp Matthews JA 2009 Pioneer vegetation
on glacier forelands in southern Norway emerging
communities Journal of Vegetation Science 20 889ndash
902
Swanson FJ amp Major JJ 2005 Physical events
environments and geologicalndashecological interactions
at Mount St Helens March 1980ndash2004 In Dale
VH Swanson FJ amp Crisafulli CM (eds)
Ecological recovery after the 1980 eruption of Mount
St Helens pp 27ndash44 Springer New York NY US
USDA National Resource Conservation Service 2009
The Plants Data Base National Plants Data Center
Baton Rouge LA US Available at http
plantsusdagov Accessed October 15 2009
van Oijen D Hommel P den Ouden J amp de Waal R
2005 Effects of tree species composition on within-
forest distribution of understorey species Applied
Vegetation Science 8 155ndash166
Walker LR amp del Moral R 2003 Primary succession
and ecosystem rehabilitation Cambridge University
Press Cambridge UK
866 del Moral Roger et al
Walker LR amp del Moral R 2009 Transition dynamics
in succession implications for rates trajectories and
restoration In Hobbs RJ amp Suding KN (eds)
New models for ecosystem dynamics and restoration
pp 33ndash49 Island Press Washington DC US
Walker LR Bellingham PJ amp Peltzer DA 2006
Plant characteristics are poor predictors of microsite
colonization during the first two years of pri-
mary succession Journal of Vegetation Science 17
397ndash406
Wood DM amp del Moral R 1988 Colonizing plants on
the Pumice Plains Mount St Helens Washington
American Journal of Botany 75 1228ndash1237
Wood DM amp del Moral R 2000 Seed rain during early
primary succession onMount St Helens Washington
Madrono 47 1ndash9
Supporting Information
Additional supporting information may befound in the online version of this article
Fig S1 Abraham Plain Sept 1980 pumice de-posits over scoured bedrock looking north towardsthe study site
Fig S2 Abraham Plain July 2009 lookingnorthwest across the study site
Fig S3 Changes in mean cover index for lesscommon species
Fig S4 Relative cover (proportion of totalcover in the sample) of less common species in thefour stages of early primary succession The firstfour species are persistent the last two are pioneerspecies (defined in Appendix S1)
Fig S5 Annual changes of mean H0 and meanSimpsonrsquos dominance index Differences (Po005)among measures determined by a repeat measuresANOVA followed by the Bonferroni test Valueswith same associated letter fall within the samegroup of values
Table S1 Structure and species composition ofCTs found in 2008
Appendix S1 Characteristics of species en-countered on Abraham Plain grid
Please note Wiley-Blackwell is not responsiblefor the content or functionality of any supportingmaterials supplied by the authors Any queries(other than missing material) should be directed tothe corresponding author for the article
Received 9 December 2009
Accepted 14 March 2010
Co-ordinating Editor Dr Kerry Woods
Primary succession trajectories 867
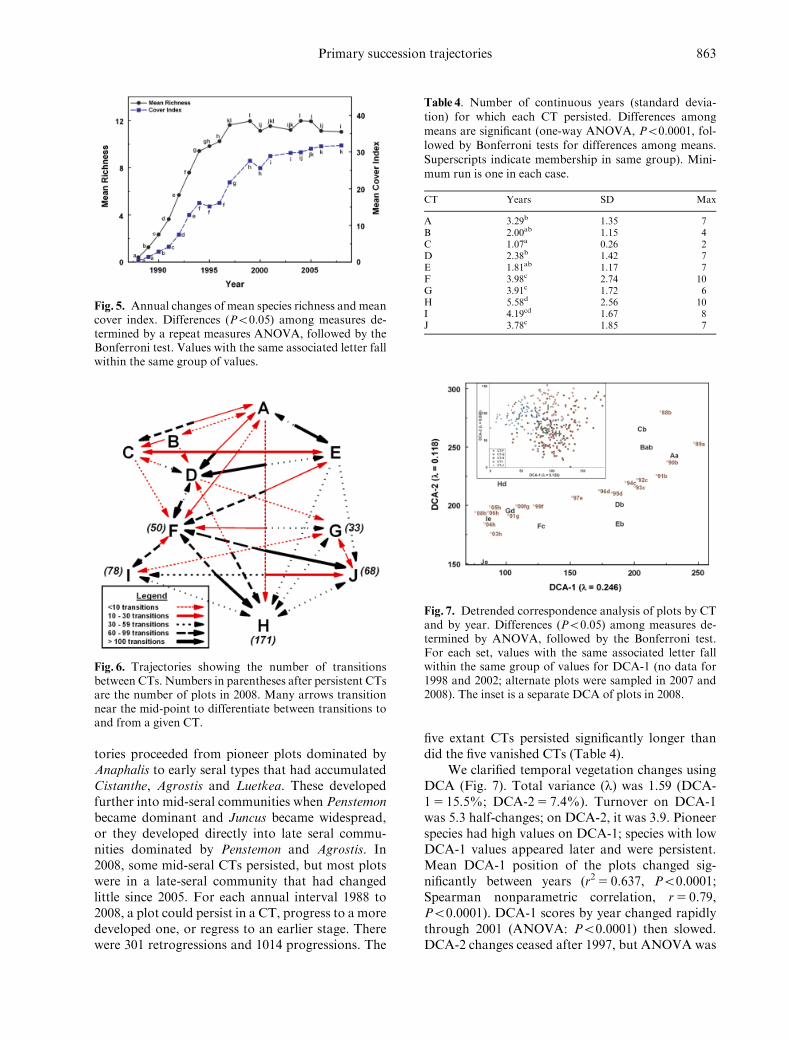
tories proceeded from pioneer plots dominated byAnaphalis to early seral types that had accumulatedCistanthe Agrostis and Luetkea These developedfurther into mid-seral communities when Penstemonbecame dominant and Juncus became widespreador they developed directly into late seral commu-nities dominated by Penstemon and Agrostis In2008 some mid-seral CTs persisted but most plotswere in a late-seral community that had changedlittle since 2005 For each annual interval 1988 to2008 a plot could persist in a CT progress to a moredeveloped one or regress to an earlier stage Therewere 301 retrogressions and 1014 progressions The
five extant CTs persisted significantly longer thandid the five vanished CTs (Table 4)
We clarified temporal vegetation changes usingDCA (Fig 7) Total variance (l) was 159 (DCA-15 155 DCA-25 74) Turnover on DCA-1was 53 half-changes on DCA-2 it was 39 Pioneerspecies had high values on DCA-1 species with lowDCA-1 values appeared later and were persistentMean DCA-1 position of the plots changed sig-nificantly between years (r2 5 0637 Po00001Spearman nonparametric correlation r5 079Po00001) DCA-1 scores by year changed rapidlythrough 2001 (ANOVA Po00001) then slowedDCA-2 changes ceased after 1997 but ANOVA was
Fig 5 Annual changes of mean species richness and meancover index Differences (Po005) among measures de-termined by a repeat measures ANOVA followed by theBonferroni test Values with the same associated letter fallwithin the same group of values
Fig 6 Trajectories showing the number of transitionsbetween CTs Numbers in parentheses after persistent CTsare the number of plots in 2008 Many arrows transitionnear the mid-point to differentiate between transitions toand from a given CT
Table 4 Number of continuous years (standard devia-tion) for which each CT persisted Differences amongmeans are significant (one-way ANOVA Po00001 fol-lowed by Bonferroni tests for differences among meansSuperscripts indicate membership in same group) Mini-mum run is one in each case
CT Years SD Max
A 329b 135 7B 200ab 115 4C 107a 026 2D 238b 142 7E 181ab 117 7F 398c 274 10G 391c 172 6H 558d 256 10I 419cd 167 8J 378c 185 7
Fig 7 Detrended correspondence analysis of plots by CTand by year Differences (Po005) among measures de-termined by ANOVA followed by the Bonferroni testFor each set values with the same associated letter fallwithin the same group of values for DCA-1 (no data for1998 and 2002 alternate plots were sampled in 2007 and2008) The inset is a separate DCA of plots in 2008
Primary succession trajectories 863
significant (r2 5 0206 Po00001 Spearman non-parametric correlation r5 025 Po00001) Thevariation in DCA-1 scores within years declined (thelinear correlation of years vs standard deviationwas Po00001 t5 533 r2 5 0637)
The mean DCA position of the 10 CTs did notreveal a single trajectory CT-C (Anaphalis) wasmore closely associated with pioneer CTs but wasnever the first CT on a plot CT-G (Penstemon-Agrostis-Cistanthe) was aligned with late-seral CTsWe also conducted a DCA of 2008 plots (Fig 7inset) b-diversity was low with l5 097 (DCA-15
133 DCA-25 84 ca 18 HC along DCA-1)No trajectory was evident from this analysis
Discussion
There are so few long-term longitudinal studiesof early primary succession on stressful sites be-cause initially so little happens The vegetation ofthe Abraham Plain remains sparse 30 years after theeruption It has developed by interwoven succes-sional trajectories that include plots that divergeconverge regress and progress to form a braidedsuccession Neither habitat factors nor competitionappears to have provided strong filtering effects onspecies composition although unmeasured soilvariables (eg moisture and nitrogen content) andfactors related to stochastic colonization and estab-lishment (cf del Moral amp Ellis 2004) may explainsome of the remaining unexplained variation Theimmigration seed density was low (Wood amp delMoral 2000) many suitable sites appeared vacant inearly years (del Moral amp Bliss 1993) and severalspecies occupied similar sites Chance may governsuch patterns We propose that when environmentalvariables have only slight effects on species compo-sition consistent successional trajectories areunlikely to emerge Below we discuss questionsabout predictability rate and pattern of successionon Abraham Plain that are the focus of this study
Environmentndashvegetation connection
Predictable relationships between environ-mental variables and vegetation had developed by2008 but they remained low Priority effects sec-ondary disturbances elk grazing and seasonalfluctuations may affect these trajectories in the fu-ture rendering it unlikely that explained variationwould become the dominant factor before forestsdevelop
We did not use soil factors because plants es-tablish at a smaller scale than the plot size and it isimpractical to measure such factors as moisture ineven a few plots Although soils were young andhomogeneous it is likely that inclusion of soil fac-tors (eg pH and moisture) would have improvedexplanatory power based on similar studies (delMoral 2009a) explained variance might doubleTopographic surface and position factors alone diddemonstrate increasing explained variance of spe-cies pattern over time but this trend was lesspronounced than in other studies On the northslope of Mount St Helens in 1992 only spatial fac-tors explained any variation (15) but after 10 yrenvironmental factors dominated (28 del Moralamp Lacher 2005) Baasch et al (2009) produced oneof the very few primary succession studies to showthat relationships between vegetation and ex-planatory variables increased over time They toosuggested that the impact of stochastic processesdeclined over time
The spatial distribution of CTs suggested someenvironmental influence Rills provide havens forpioneers (eg Anaphalis Hieracium and Hy-pochaeris) Gullies offer protection from wind andaccumulate snow which allowed species such asAnaphalis and Salix to colonize and persist Pen-stemon and Juncus were common in more exposedsites where cover and diversity were least In suchsites species that are less stress tolerant (eg SalixPolytrichum Saxifraga and Juncus mertensianus)were uncommon while stress-tolerant species suchas Cistanthe Juncus parryi Agrostis pallens andCarex microptera reached their maximum values
Rates
Habitats with limited fertility or moisture typi-cally have low succession rates (Donnegan ampRebertus 1999 Anderson 2007) and succession onAbraham Plain has been no exception Mean annualDCA scores were in a single group of values Be-tween 1988 and 1999 these scores formed sevengroups Surrounding habitats on Mount St Helensdeveloped more rapidly On an exposed lahar on thesouth slope richness averaged 16 species and thecover index averaged 73 (del Moral 2009b) prior to2008 In contrast mean richness on the AbrahamPlain in 2008 was 108 and the mean cover index was314 The proximity to forest more fertile soils andabundance of the N-fixing Lupinus may all con-tribute to accelerated primary succession at suchsites relative to Abraham Plain
864 del Moral Roger et al
Trajectories
Succession trajectories have been discussedthoroughly (cf Leps amp Rejmanek 1991) When thevegetation of initially homogeneous sites developslocally distinctive characteristics divergence occurs(Bossuyt et al 2003) Divergent trajectories occur onmany surfaces including landslides glacier fore-lands wet slack dune vegetation floodplains andmine tailings (Walker amp del Moral 2003) Lanta ampLeps (2009) showed experimentally that differentialdispersal could promote divergence when there arepriority effects Divergence may be more likely instressful habitats when different species arrive firstand where climatic factors are highly variable(MacDougall et al 2008) Convergence describestrajectories that become more similar over timeConvergence has been assumed to result when cli-matic factors dominate and there is evidence thatwhen dispersal effects are small and competitive ef-fects are strong convergence to a single communitydoes occur (van Oijen et al 2005 Anthelme et al2007) Convergence is also more likely when thereare few alternative stages as on glacial forelands(Hodkinson et al 2003 but see Robbins ampMathews2009)
Here we show directly that early succession un-der stressful conditions can include trajectories thatdiverge converge regress and progress to form areticulate succession Our study joins a growingbody of work that demonstrates the frequency ofmultiple trajectories in early primary succession (egdel Moral 2007 Prach amp Hobbs 2008 Walker amp delMoral 2009) Forecasting trajectories and identify-ing factors that limit their development is crucial tounderstanding community assembly mechanismsOn the Abraham Plain persistent species graduallyreplaced readily dispersed pioneer species Subtlehabitat variation annual weather fluctuation andlandscape effects combined to produce communitiesrelated through a network of development and re-sponsible to topographic variation
The first colonists were species with excellentdispersal but isolation allowed for only a meagerseed rain they established pioneer communities thatsoon developed into other CTs characterized byspecies with greater persistence (cf Ejrnaes et al2006) Several alternative transitions were possiblefor most CTs suggesting that stochastic processesinitially dominated transitions Gradually persis-tent species invaded and attained dominance Minortopographic variation affected composition some-what resulting in some successional transitions thatare more common than others Of annual transi-
tions between successional stages 51 wereregressions and 170 were progressions DCA re-vealed no single trend among the CTs The two mid-seral community types were central to the transitionpatterns while the three late-seral CTs occurred indifferent directions from this centre
It is unlikely that complete convergence will oc-cur Even when trees dominate understoreyvegetation is likely to retain variation initiated dur-ing early succession and maintained by topographicvariation and priority effects The understorey offorests on the south side of Mount St Helens de-monstrates such variation (del Moral amp Ellis 2004)There is no evidence yet for a single target commu-nity
Strong assembly rules are more likely to pro-duce convergent trajectories than weak ones Whilestrong assembly rules may exist in mature vegeta-tion (see Navas amp Violle 2009) they appear weak onAbraham Plain and are more effective for functionaltypes than species One rule may be that long-livedwind-dispersed species accumulate at the expense ofshort-lived pioneers A second may be that grami-noids with rhizomatous growth (eg Agrostispallens) and prostrate shrubs (eg Penstemon) arecomplementary and can coexist indefinitely A thirdmay be that fleshy fruit-producing shrubs (eg Ru-bus spp Vaccinium spp) cannot establish undercurrent conditions Finally although seeds of con-ifers and Populus balsamifera are commonconditions do not yet favour their developmentThese rules are weak and exceptions are commonThe spontaneously developing vegetation of theAbraham Plain is constrained by isolation from po-tential colonists as well as a habitat that challengesplant growth
Acknowledgements We thank the US National Science
Foundation for funding (BSR8906544 DEB9406987
DEB0087040 and DEB0541972) and the Mount St He-
lens National Volcanic Monument for permissions
Valuable comments by J G Bishop M P Fleming Jan-
neke Hille Ris Lambers L R Walker J H Titus and two
anonymous reviewers improved the paper Field assis-
tance was provided through the years by S Anderson W
Arnesen S Bard A Coogan S Crawford K Dlugosch
A Eckert E Ellis C Fairbourn C Muerdter M Flem-
ing T Fletcher R Fuller A Grant E Jenkins M
Johns C Jones L McMullan P Moody R Robham L
Rozzell J Sandler R Sewell-Nesteruk M Spasojevic
M Tweiten S Wilson B Witte and C Wolfe This is
contribution No 60 to the Mount St Helens Succession
Project
Primary succession trajectories 865
References
Anderson KJ 2007 Temporal patterns in rates of
community change during succession American
Naturalist 169 780ndash793
Anthelme F Villaret JC amp Brun JJ 2007 Shrub
encroachment in the Alps gives rise to the
convergence of sub-alpine communities on a regional
scale Journal of Vegetation Science 18 355ndash362
Baasch A Tischew S amp Bruelheide H 2009 Insights
into succession processes using temporally repeated
habitat models results from a long-term study in a
post-mining landscape Journal of Vegetation Science
20 629ndash638
Bossuyt B Honnay O amp Hermy M 2003 An island
biogeographical view of the successional pathway in
wet dune slacks Journal of Vegetation Science 14 781ndash
788
Clifford HT 1976 Dendrograms and their
interpretation In Williams WT (ed) Pattern
analysis in agricultural science pp 96ndash109 Elsevier
Science Amsterdam NL
del Moral R 2007 Vegetation dynamics in space and
time an example from Mount St Helens Journal of
Vegetation Science 18 479ndash488
del Moral R 2009a Increasing deterministic control of
primary succession onMount St Helens Washington
Journal of Vegetation Science 20 1145ndash1154
del Moral R 2009b Mount St Helens permanent plots
and grids 1980 to present Available at http
protistbiologywashingtonedudelmoral Accessed 8
December 2009
del Moral R amp Bliss LC 1993 Mechanism of primary
succession insights resulting from the eruption of
Mount St Helens Advances in Ecological Research
24 1ndash66
del Moral R amp Ellis EE 2004 Gradients in hetero-
geneity and structure on lahars Mount St Helens
Washington USA Plant Ecology 175 273ndash286
del Moral R amp Lacher IL 2005 Vegetation patterns 25
years after the eruption of Mount St Helens
WashingtonAmerican Journal of Botany 92 1948ndash1956
Donnegan JA amp Rebertus AJ 1999 Rates and
mechanisms of subalpine forest succession along an
environmental gradient Ecology 80 1370ndash1384
Ejrnaes R Bruun HH amp Graae BJ 2006 Community
assembly in experimental grasslands suitable
environment or timely arrival Ecology 87 1225ndash1233
Felinks B ampWiegand T 2008 Exploring spatiotemporal
patterns in early stages of primary succession on
former lignite mining sites Journal of Vegetation
Science 19 267ndash276
Fuller RN amp del Moral R 2003 The role of refugia and
dispersal in primary succession on Mount St Helens
Washington Journal of Vegetation Science 14 637ndash
644
Hodkinson IA Coulson SJ amp Webb NR 2003
Community assembly along proglacial chrono-
sequences in the high Arctic vegetation and soil
development in north-west Svalbard Journal of
Ecology 91 651ndash663
Johnson EA amp Miyanishi K 2008 Testing the
assumptions of chronosequences in succession
Ecology Letters 11 419ndash431
Lanta V amp Leps J 2009 How does surrounding
vegetation affect the course of succession a five-year
container experiment Journal of Vegetation Science
20 686ndash694
Legendre P amp Anderson MJ 1999 Distance-based
redundancy analysis testing multispecies responses in
multi-factorial ecological experiments Ecological
Monographs 69 1ndash24
Leps J amp Rejmanek M 1991 Convergence or
divergence what should we expect from vegetation
succession Oikos 62 261ndash264
Leps J amp Smilauer P 2003 Multivariate analysis of
ecological data using CANOCO Cambridge
University Press Cambridge UK
MacDougall AS Wilson SD amp Bakker JD 2008
Climatic variability alters the outcome of longterm
community assembly Journal of Ecology 96 346ndash354
Navas ML amp Violle C 2009 Plant traits related to
competition how do they shape the functional
diversity of communities Community Ecology 10
131ndash137
Pickett STA Cadenasso ML amp Meiners SJ 2009
Ever since Clements from succession to vegetation
dynamics and understanding to intervention Applied
Vegetation Science 12 9ndash21
Prach K amp Hobbs RJ 2008 Spontaneous succession
versus technical reclamation in the restoration of
disturbed sites Restoration Ecology 16 363ndash366
Prach K amp Rehounkova K 2006 Vegetation succession
over broad geographical scales which factors
determine the patterns Preslia 78 469ndash480
Robbins JA amp Matthews JA 2009 Pioneer vegetation
on glacier forelands in southern Norway emerging
communities Journal of Vegetation Science 20 889ndash
902
Swanson FJ amp Major JJ 2005 Physical events
environments and geologicalndashecological interactions
at Mount St Helens March 1980ndash2004 In Dale
VH Swanson FJ amp Crisafulli CM (eds)
Ecological recovery after the 1980 eruption of Mount
St Helens pp 27ndash44 Springer New York NY US
USDA National Resource Conservation Service 2009
The Plants Data Base National Plants Data Center
Baton Rouge LA US Available at http
plantsusdagov Accessed October 15 2009
van Oijen D Hommel P den Ouden J amp de Waal R
2005 Effects of tree species composition on within-
forest distribution of understorey species Applied
Vegetation Science 8 155ndash166
Walker LR amp del Moral R 2003 Primary succession
and ecosystem rehabilitation Cambridge University
Press Cambridge UK
866 del Moral Roger et al
Walker LR amp del Moral R 2009 Transition dynamics
in succession implications for rates trajectories and
restoration In Hobbs RJ amp Suding KN (eds)
New models for ecosystem dynamics and restoration
pp 33ndash49 Island Press Washington DC US
Walker LR Bellingham PJ amp Peltzer DA 2006
Plant characteristics are poor predictors of microsite
colonization during the first two years of pri-
mary succession Journal of Vegetation Science 17
397ndash406
Wood DM amp del Moral R 1988 Colonizing plants on
the Pumice Plains Mount St Helens Washington
American Journal of Botany 75 1228ndash1237
Wood DM amp del Moral R 2000 Seed rain during early
primary succession onMount St Helens Washington
Madrono 47 1ndash9
Supporting Information
Additional supporting information may befound in the online version of this article
Fig S1 Abraham Plain Sept 1980 pumice de-posits over scoured bedrock looking north towardsthe study site
Fig S2 Abraham Plain July 2009 lookingnorthwest across the study site
Fig S3 Changes in mean cover index for lesscommon species
Fig S4 Relative cover (proportion of totalcover in the sample) of less common species in thefour stages of early primary succession The firstfour species are persistent the last two are pioneerspecies (defined in Appendix S1)
Fig S5 Annual changes of mean H0 and meanSimpsonrsquos dominance index Differences (Po005)among measures determined by a repeat measuresANOVA followed by the Bonferroni test Valueswith same associated letter fall within the samegroup of values
Table S1 Structure and species composition ofCTs found in 2008
Appendix S1 Characteristics of species en-countered on Abraham Plain grid
Please note Wiley-Blackwell is not responsiblefor the content or functionality of any supportingmaterials supplied by the authors Any queries(other than missing material) should be directed tothe corresponding author for the article
Received 9 December 2009
Accepted 14 March 2010
Co-ordinating Editor Dr Kerry Woods
Primary succession trajectories 867

significant (r2 5 0206 Po00001 Spearman non-parametric correlation r5 025 Po00001) Thevariation in DCA-1 scores within years declined (thelinear correlation of years vs standard deviationwas Po00001 t5 533 r2 5 0637)
The mean DCA position of the 10 CTs did notreveal a single trajectory CT-C (Anaphalis) wasmore closely associated with pioneer CTs but wasnever the first CT on a plot CT-G (Penstemon-Agrostis-Cistanthe) was aligned with late-seral CTsWe also conducted a DCA of 2008 plots (Fig 7inset) b-diversity was low with l5 097 (DCA-15
133 DCA-25 84 ca 18 HC along DCA-1)No trajectory was evident from this analysis
Discussion
There are so few long-term longitudinal studiesof early primary succession on stressful sites be-cause initially so little happens The vegetation ofthe Abraham Plain remains sparse 30 years after theeruption It has developed by interwoven succes-sional trajectories that include plots that divergeconverge regress and progress to form a braidedsuccession Neither habitat factors nor competitionappears to have provided strong filtering effects onspecies composition although unmeasured soilvariables (eg moisture and nitrogen content) andfactors related to stochastic colonization and estab-lishment (cf del Moral amp Ellis 2004) may explainsome of the remaining unexplained variation Theimmigration seed density was low (Wood amp delMoral 2000) many suitable sites appeared vacant inearly years (del Moral amp Bliss 1993) and severalspecies occupied similar sites Chance may governsuch patterns We propose that when environmentalvariables have only slight effects on species compo-sition consistent successional trajectories areunlikely to emerge Below we discuss questionsabout predictability rate and pattern of successionon Abraham Plain that are the focus of this study
Environmentndashvegetation connection
Predictable relationships between environ-mental variables and vegetation had developed by2008 but they remained low Priority effects sec-ondary disturbances elk grazing and seasonalfluctuations may affect these trajectories in the fu-ture rendering it unlikely that explained variationwould become the dominant factor before forestsdevelop
We did not use soil factors because plants es-tablish at a smaller scale than the plot size and it isimpractical to measure such factors as moisture ineven a few plots Although soils were young andhomogeneous it is likely that inclusion of soil fac-tors (eg pH and moisture) would have improvedexplanatory power based on similar studies (delMoral 2009a) explained variance might doubleTopographic surface and position factors alone diddemonstrate increasing explained variance of spe-cies pattern over time but this trend was lesspronounced than in other studies On the northslope of Mount St Helens in 1992 only spatial fac-tors explained any variation (15) but after 10 yrenvironmental factors dominated (28 del Moralamp Lacher 2005) Baasch et al (2009) produced oneof the very few primary succession studies to showthat relationships between vegetation and ex-planatory variables increased over time They toosuggested that the impact of stochastic processesdeclined over time
The spatial distribution of CTs suggested someenvironmental influence Rills provide havens forpioneers (eg Anaphalis Hieracium and Hy-pochaeris) Gullies offer protection from wind andaccumulate snow which allowed species such asAnaphalis and Salix to colonize and persist Pen-stemon and Juncus were common in more exposedsites where cover and diversity were least In suchsites species that are less stress tolerant (eg SalixPolytrichum Saxifraga and Juncus mertensianus)were uncommon while stress-tolerant species suchas Cistanthe Juncus parryi Agrostis pallens andCarex microptera reached their maximum values
Rates
Habitats with limited fertility or moisture typi-cally have low succession rates (Donnegan ampRebertus 1999 Anderson 2007) and succession onAbraham Plain has been no exception Mean annualDCA scores were in a single group of values Be-tween 1988 and 1999 these scores formed sevengroups Surrounding habitats on Mount St Helensdeveloped more rapidly On an exposed lahar on thesouth slope richness averaged 16 species and thecover index averaged 73 (del Moral 2009b) prior to2008 In contrast mean richness on the AbrahamPlain in 2008 was 108 and the mean cover index was314 The proximity to forest more fertile soils andabundance of the N-fixing Lupinus may all con-tribute to accelerated primary succession at suchsites relative to Abraham Plain
864 del Moral Roger et al
Trajectories
Succession trajectories have been discussedthoroughly (cf Leps amp Rejmanek 1991) When thevegetation of initially homogeneous sites developslocally distinctive characteristics divergence occurs(Bossuyt et al 2003) Divergent trajectories occur onmany surfaces including landslides glacier fore-lands wet slack dune vegetation floodplains andmine tailings (Walker amp del Moral 2003) Lanta ampLeps (2009) showed experimentally that differentialdispersal could promote divergence when there arepriority effects Divergence may be more likely instressful habitats when different species arrive firstand where climatic factors are highly variable(MacDougall et al 2008) Convergence describestrajectories that become more similar over timeConvergence has been assumed to result when cli-matic factors dominate and there is evidence thatwhen dispersal effects are small and competitive ef-fects are strong convergence to a single communitydoes occur (van Oijen et al 2005 Anthelme et al2007) Convergence is also more likely when thereare few alternative stages as on glacial forelands(Hodkinson et al 2003 but see Robbins ampMathews2009)
Here we show directly that early succession un-der stressful conditions can include trajectories thatdiverge converge regress and progress to form areticulate succession Our study joins a growingbody of work that demonstrates the frequency ofmultiple trajectories in early primary succession (egdel Moral 2007 Prach amp Hobbs 2008 Walker amp delMoral 2009) Forecasting trajectories and identify-ing factors that limit their development is crucial tounderstanding community assembly mechanismsOn the Abraham Plain persistent species graduallyreplaced readily dispersed pioneer species Subtlehabitat variation annual weather fluctuation andlandscape effects combined to produce communitiesrelated through a network of development and re-sponsible to topographic variation
The first colonists were species with excellentdispersal but isolation allowed for only a meagerseed rain they established pioneer communities thatsoon developed into other CTs characterized byspecies with greater persistence (cf Ejrnaes et al2006) Several alternative transitions were possiblefor most CTs suggesting that stochastic processesinitially dominated transitions Gradually persis-tent species invaded and attained dominance Minortopographic variation affected composition some-what resulting in some successional transitions thatare more common than others Of annual transi-
tions between successional stages 51 wereregressions and 170 were progressions DCA re-vealed no single trend among the CTs The two mid-seral community types were central to the transitionpatterns while the three late-seral CTs occurred indifferent directions from this centre
It is unlikely that complete convergence will oc-cur Even when trees dominate understoreyvegetation is likely to retain variation initiated dur-ing early succession and maintained by topographicvariation and priority effects The understorey offorests on the south side of Mount St Helens de-monstrates such variation (del Moral amp Ellis 2004)There is no evidence yet for a single target commu-nity
Strong assembly rules are more likely to pro-duce convergent trajectories than weak ones Whilestrong assembly rules may exist in mature vegeta-tion (see Navas amp Violle 2009) they appear weak onAbraham Plain and are more effective for functionaltypes than species One rule may be that long-livedwind-dispersed species accumulate at the expense ofshort-lived pioneers A second may be that grami-noids with rhizomatous growth (eg Agrostispallens) and prostrate shrubs (eg Penstemon) arecomplementary and can coexist indefinitely A thirdmay be that fleshy fruit-producing shrubs (eg Ru-bus spp Vaccinium spp) cannot establish undercurrent conditions Finally although seeds of con-ifers and Populus balsamifera are commonconditions do not yet favour their developmentThese rules are weak and exceptions are commonThe spontaneously developing vegetation of theAbraham Plain is constrained by isolation from po-tential colonists as well as a habitat that challengesplant growth
Acknowledgements We thank the US National Science
Foundation for funding (BSR8906544 DEB9406987
DEB0087040 and DEB0541972) and the Mount St He-
lens National Volcanic Monument for permissions
Valuable comments by J G Bishop M P Fleming Jan-
neke Hille Ris Lambers L R Walker J H Titus and two
anonymous reviewers improved the paper Field assis-
tance was provided through the years by S Anderson W
Arnesen S Bard A Coogan S Crawford K Dlugosch
A Eckert E Ellis C Fairbourn C Muerdter M Flem-
ing T Fletcher R Fuller A Grant E Jenkins M
Johns C Jones L McMullan P Moody R Robham L
Rozzell J Sandler R Sewell-Nesteruk M Spasojevic
M Tweiten S Wilson B Witte and C Wolfe This is
contribution No 60 to the Mount St Helens Succession
Project
Primary succession trajectories 865
References
Anderson KJ 2007 Temporal patterns in rates of
community change during succession American
Naturalist 169 780ndash793
Anthelme F Villaret JC amp Brun JJ 2007 Shrub
encroachment in the Alps gives rise to the
convergence of sub-alpine communities on a regional
scale Journal of Vegetation Science 18 355ndash362
Baasch A Tischew S amp Bruelheide H 2009 Insights
into succession processes using temporally repeated
habitat models results from a long-term study in a
post-mining landscape Journal of Vegetation Science
20 629ndash638
Bossuyt B Honnay O amp Hermy M 2003 An island
biogeographical view of the successional pathway in
wet dune slacks Journal of Vegetation Science 14 781ndash
788
Clifford HT 1976 Dendrograms and their
interpretation In Williams WT (ed) Pattern
analysis in agricultural science pp 96ndash109 Elsevier
Science Amsterdam NL
del Moral R 2007 Vegetation dynamics in space and
time an example from Mount St Helens Journal of
Vegetation Science 18 479ndash488
del Moral R 2009a Increasing deterministic control of
primary succession onMount St Helens Washington
Journal of Vegetation Science 20 1145ndash1154
del Moral R 2009b Mount St Helens permanent plots
and grids 1980 to present Available at http
protistbiologywashingtonedudelmoral Accessed 8
December 2009
del Moral R amp Bliss LC 1993 Mechanism of primary
succession insights resulting from the eruption of
Mount St Helens Advances in Ecological Research
24 1ndash66
del Moral R amp Ellis EE 2004 Gradients in hetero-
geneity and structure on lahars Mount St Helens
Washington USA Plant Ecology 175 273ndash286
del Moral R amp Lacher IL 2005 Vegetation patterns 25
years after the eruption of Mount St Helens
WashingtonAmerican Journal of Botany 92 1948ndash1956
Donnegan JA amp Rebertus AJ 1999 Rates and
mechanisms of subalpine forest succession along an
environmental gradient Ecology 80 1370ndash1384
Ejrnaes R Bruun HH amp Graae BJ 2006 Community
assembly in experimental grasslands suitable
environment or timely arrival Ecology 87 1225ndash1233
Felinks B ampWiegand T 2008 Exploring spatiotemporal
patterns in early stages of primary succession on
former lignite mining sites Journal of Vegetation
Science 19 267ndash276
Fuller RN amp del Moral R 2003 The role of refugia and
dispersal in primary succession on Mount St Helens
Washington Journal of Vegetation Science 14 637ndash
644
Hodkinson IA Coulson SJ amp Webb NR 2003
Community assembly along proglacial chrono-
sequences in the high Arctic vegetation and soil
development in north-west Svalbard Journal of
Ecology 91 651ndash663
Johnson EA amp Miyanishi K 2008 Testing the
assumptions of chronosequences in succession
Ecology Letters 11 419ndash431
Lanta V amp Leps J 2009 How does surrounding
vegetation affect the course of succession a five-year
container experiment Journal of Vegetation Science
20 686ndash694
Legendre P amp Anderson MJ 1999 Distance-based
redundancy analysis testing multispecies responses in
multi-factorial ecological experiments Ecological
Monographs 69 1ndash24
Leps J amp Rejmanek M 1991 Convergence or
divergence what should we expect from vegetation
succession Oikos 62 261ndash264
Leps J amp Smilauer P 2003 Multivariate analysis of
ecological data using CANOCO Cambridge
University Press Cambridge UK
MacDougall AS Wilson SD amp Bakker JD 2008
Climatic variability alters the outcome of longterm
community assembly Journal of Ecology 96 346ndash354
Navas ML amp Violle C 2009 Plant traits related to
competition how do they shape the functional
diversity of communities Community Ecology 10
131ndash137
Pickett STA Cadenasso ML amp Meiners SJ 2009
Ever since Clements from succession to vegetation
dynamics and understanding to intervention Applied
Vegetation Science 12 9ndash21
Prach K amp Hobbs RJ 2008 Spontaneous succession
versus technical reclamation in the restoration of
disturbed sites Restoration Ecology 16 363ndash366
Prach K amp Rehounkova K 2006 Vegetation succession
over broad geographical scales which factors
determine the patterns Preslia 78 469ndash480
Robbins JA amp Matthews JA 2009 Pioneer vegetation
on glacier forelands in southern Norway emerging
communities Journal of Vegetation Science 20 889ndash
902
Swanson FJ amp Major JJ 2005 Physical events
environments and geologicalndashecological interactions
at Mount St Helens March 1980ndash2004 In Dale
VH Swanson FJ amp Crisafulli CM (eds)
Ecological recovery after the 1980 eruption of Mount
St Helens pp 27ndash44 Springer New York NY US
USDA National Resource Conservation Service 2009
The Plants Data Base National Plants Data Center
Baton Rouge LA US Available at http
plantsusdagov Accessed October 15 2009
van Oijen D Hommel P den Ouden J amp de Waal R
2005 Effects of tree species composition on within-
forest distribution of understorey species Applied
Vegetation Science 8 155ndash166
Walker LR amp del Moral R 2003 Primary succession
and ecosystem rehabilitation Cambridge University
Press Cambridge UK
866 del Moral Roger et al
Walker LR amp del Moral R 2009 Transition dynamics
in succession implications for rates trajectories and
restoration In Hobbs RJ amp Suding KN (eds)
New models for ecosystem dynamics and restoration
pp 33ndash49 Island Press Washington DC US
Walker LR Bellingham PJ amp Peltzer DA 2006
Plant characteristics are poor predictors of microsite
colonization during the first two years of pri-
mary succession Journal of Vegetation Science 17
397ndash406
Wood DM amp del Moral R 1988 Colonizing plants on
the Pumice Plains Mount St Helens Washington
American Journal of Botany 75 1228ndash1237
Wood DM amp del Moral R 2000 Seed rain during early
primary succession onMount St Helens Washington
Madrono 47 1ndash9
Supporting Information
Additional supporting information may befound in the online version of this article
Fig S1 Abraham Plain Sept 1980 pumice de-posits over scoured bedrock looking north towardsthe study site
Fig S2 Abraham Plain July 2009 lookingnorthwest across the study site
Fig S3 Changes in mean cover index for lesscommon species
Fig S4 Relative cover (proportion of totalcover in the sample) of less common species in thefour stages of early primary succession The firstfour species are persistent the last two are pioneerspecies (defined in Appendix S1)
Fig S5 Annual changes of mean H0 and meanSimpsonrsquos dominance index Differences (Po005)among measures determined by a repeat measuresANOVA followed by the Bonferroni test Valueswith same associated letter fall within the samegroup of values
Table S1 Structure and species composition ofCTs found in 2008
Appendix S1 Characteristics of species en-countered on Abraham Plain grid
Please note Wiley-Blackwell is not responsiblefor the content or functionality of any supportingmaterials supplied by the authors Any queries(other than missing material) should be directed tothe corresponding author for the article
Received 9 December 2009
Accepted 14 March 2010
Co-ordinating Editor Dr Kerry Woods
Primary succession trajectories 867

Trajectories
Succession trajectories have been discussedthoroughly (cf Leps amp Rejmanek 1991) When thevegetation of initially homogeneous sites developslocally distinctive characteristics divergence occurs(Bossuyt et al 2003) Divergent trajectories occur onmany surfaces including landslides glacier fore-lands wet slack dune vegetation floodplains andmine tailings (Walker amp del Moral 2003) Lanta ampLeps (2009) showed experimentally that differentialdispersal could promote divergence when there arepriority effects Divergence may be more likely instressful habitats when different species arrive firstand where climatic factors are highly variable(MacDougall et al 2008) Convergence describestrajectories that become more similar over timeConvergence has been assumed to result when cli-matic factors dominate and there is evidence thatwhen dispersal effects are small and competitive ef-fects are strong convergence to a single communitydoes occur (van Oijen et al 2005 Anthelme et al2007) Convergence is also more likely when thereare few alternative stages as on glacial forelands(Hodkinson et al 2003 but see Robbins ampMathews2009)
Here we show directly that early succession un-der stressful conditions can include trajectories thatdiverge converge regress and progress to form areticulate succession Our study joins a growingbody of work that demonstrates the frequency ofmultiple trajectories in early primary succession (egdel Moral 2007 Prach amp Hobbs 2008 Walker amp delMoral 2009) Forecasting trajectories and identify-ing factors that limit their development is crucial tounderstanding community assembly mechanismsOn the Abraham Plain persistent species graduallyreplaced readily dispersed pioneer species Subtlehabitat variation annual weather fluctuation andlandscape effects combined to produce communitiesrelated through a network of development and re-sponsible to topographic variation
The first colonists were species with excellentdispersal but isolation allowed for only a meagerseed rain they established pioneer communities thatsoon developed into other CTs characterized byspecies with greater persistence (cf Ejrnaes et al2006) Several alternative transitions were possiblefor most CTs suggesting that stochastic processesinitially dominated transitions Gradually persis-tent species invaded and attained dominance Minortopographic variation affected composition some-what resulting in some successional transitions thatare more common than others Of annual transi-
tions between successional stages 51 wereregressions and 170 were progressions DCA re-vealed no single trend among the CTs The two mid-seral community types were central to the transitionpatterns while the three late-seral CTs occurred indifferent directions from this centre
It is unlikely that complete convergence will oc-cur Even when trees dominate understoreyvegetation is likely to retain variation initiated dur-ing early succession and maintained by topographicvariation and priority effects The understorey offorests on the south side of Mount St Helens de-monstrates such variation (del Moral amp Ellis 2004)There is no evidence yet for a single target commu-nity
Strong assembly rules are more likely to pro-duce convergent trajectories than weak ones Whilestrong assembly rules may exist in mature vegeta-tion (see Navas amp Violle 2009) they appear weak onAbraham Plain and are more effective for functionaltypes than species One rule may be that long-livedwind-dispersed species accumulate at the expense ofshort-lived pioneers A second may be that grami-noids with rhizomatous growth (eg Agrostispallens) and prostrate shrubs (eg Penstemon) arecomplementary and can coexist indefinitely A thirdmay be that fleshy fruit-producing shrubs (eg Ru-bus spp Vaccinium spp) cannot establish undercurrent conditions Finally although seeds of con-ifers and Populus balsamifera are commonconditions do not yet favour their developmentThese rules are weak and exceptions are commonThe spontaneously developing vegetation of theAbraham Plain is constrained by isolation from po-tential colonists as well as a habitat that challengesplant growth
Acknowledgements We thank the US National Science
Foundation for funding (BSR8906544 DEB9406987
DEB0087040 and DEB0541972) and the Mount St He-
lens National Volcanic Monument for permissions
Valuable comments by J G Bishop M P Fleming Jan-
neke Hille Ris Lambers L R Walker J H Titus and two
anonymous reviewers improved the paper Field assis-
tance was provided through the years by S Anderson W
Arnesen S Bard A Coogan S Crawford K Dlugosch
A Eckert E Ellis C Fairbourn C Muerdter M Flem-
ing T Fletcher R Fuller A Grant E Jenkins M
Johns C Jones L McMullan P Moody R Robham L
Rozzell J Sandler R Sewell-Nesteruk M Spasojevic
M Tweiten S Wilson B Witte and C Wolfe This is
contribution No 60 to the Mount St Helens Succession
Project
Primary succession trajectories 865
References
Anderson KJ 2007 Temporal patterns in rates of
community change during succession American
Naturalist 169 780ndash793
Anthelme F Villaret JC amp Brun JJ 2007 Shrub
encroachment in the Alps gives rise to the
convergence of sub-alpine communities on a regional
scale Journal of Vegetation Science 18 355ndash362
Baasch A Tischew S amp Bruelheide H 2009 Insights
into succession processes using temporally repeated
habitat models results from a long-term study in a
post-mining landscape Journal of Vegetation Science
20 629ndash638
Bossuyt B Honnay O amp Hermy M 2003 An island
biogeographical view of the successional pathway in
wet dune slacks Journal of Vegetation Science 14 781ndash
788
Clifford HT 1976 Dendrograms and their
interpretation In Williams WT (ed) Pattern
analysis in agricultural science pp 96ndash109 Elsevier
Science Amsterdam NL
del Moral R 2007 Vegetation dynamics in space and
time an example from Mount St Helens Journal of
Vegetation Science 18 479ndash488
del Moral R 2009a Increasing deterministic control of
primary succession onMount St Helens Washington
Journal of Vegetation Science 20 1145ndash1154
del Moral R 2009b Mount St Helens permanent plots
and grids 1980 to present Available at http
protistbiologywashingtonedudelmoral Accessed 8
December 2009
del Moral R amp Bliss LC 1993 Mechanism of primary
succession insights resulting from the eruption of
Mount St Helens Advances in Ecological Research
24 1ndash66
del Moral R amp Ellis EE 2004 Gradients in hetero-
geneity and structure on lahars Mount St Helens
Washington USA Plant Ecology 175 273ndash286
del Moral R amp Lacher IL 2005 Vegetation patterns 25
years after the eruption of Mount St Helens
WashingtonAmerican Journal of Botany 92 1948ndash1956
Donnegan JA amp Rebertus AJ 1999 Rates and
mechanisms of subalpine forest succession along an
environmental gradient Ecology 80 1370ndash1384
Ejrnaes R Bruun HH amp Graae BJ 2006 Community
assembly in experimental grasslands suitable
environment or timely arrival Ecology 87 1225ndash1233
Felinks B ampWiegand T 2008 Exploring spatiotemporal
patterns in early stages of primary succession on
former lignite mining sites Journal of Vegetation
Science 19 267ndash276
Fuller RN amp del Moral R 2003 The role of refugia and
dispersal in primary succession on Mount St Helens
Washington Journal of Vegetation Science 14 637ndash
644
Hodkinson IA Coulson SJ amp Webb NR 2003
Community assembly along proglacial chrono-
sequences in the high Arctic vegetation and soil
development in north-west Svalbard Journal of
Ecology 91 651ndash663
Johnson EA amp Miyanishi K 2008 Testing the
assumptions of chronosequences in succession
Ecology Letters 11 419ndash431
Lanta V amp Leps J 2009 How does surrounding
vegetation affect the course of succession a five-year
container experiment Journal of Vegetation Science
20 686ndash694
Legendre P amp Anderson MJ 1999 Distance-based
redundancy analysis testing multispecies responses in
multi-factorial ecological experiments Ecological
Monographs 69 1ndash24
Leps J amp Rejmanek M 1991 Convergence or
divergence what should we expect from vegetation
succession Oikos 62 261ndash264
Leps J amp Smilauer P 2003 Multivariate analysis of
ecological data using CANOCO Cambridge
University Press Cambridge UK
MacDougall AS Wilson SD amp Bakker JD 2008
Climatic variability alters the outcome of longterm
community assembly Journal of Ecology 96 346ndash354
Navas ML amp Violle C 2009 Plant traits related to
competition how do they shape the functional
diversity of communities Community Ecology 10
131ndash137
Pickett STA Cadenasso ML amp Meiners SJ 2009
Ever since Clements from succession to vegetation
dynamics and understanding to intervention Applied
Vegetation Science 12 9ndash21
Prach K amp Hobbs RJ 2008 Spontaneous succession
versus technical reclamation in the restoration of
disturbed sites Restoration Ecology 16 363ndash366
Prach K amp Rehounkova K 2006 Vegetation succession
over broad geographical scales which factors
determine the patterns Preslia 78 469ndash480
Robbins JA amp Matthews JA 2009 Pioneer vegetation
on glacier forelands in southern Norway emerging
communities Journal of Vegetation Science 20 889ndash
902
Swanson FJ amp Major JJ 2005 Physical events
environments and geologicalndashecological interactions
at Mount St Helens March 1980ndash2004 In Dale
VH Swanson FJ amp Crisafulli CM (eds)
Ecological recovery after the 1980 eruption of Mount
St Helens pp 27ndash44 Springer New York NY US
USDA National Resource Conservation Service 2009
The Plants Data Base National Plants Data Center
Baton Rouge LA US Available at http
plantsusdagov Accessed October 15 2009
van Oijen D Hommel P den Ouden J amp de Waal R
2005 Effects of tree species composition on within-
forest distribution of understorey species Applied
Vegetation Science 8 155ndash166
Walker LR amp del Moral R 2003 Primary succession
and ecosystem rehabilitation Cambridge University
Press Cambridge UK
866 del Moral Roger et al
Walker LR amp del Moral R 2009 Transition dynamics
in succession implications for rates trajectories and
restoration In Hobbs RJ amp Suding KN (eds)
New models for ecosystem dynamics and restoration
pp 33ndash49 Island Press Washington DC US
Walker LR Bellingham PJ amp Peltzer DA 2006
Plant characteristics are poor predictors of microsite
colonization during the first two years of pri-
mary succession Journal of Vegetation Science 17
397ndash406
Wood DM amp del Moral R 1988 Colonizing plants on
the Pumice Plains Mount St Helens Washington
American Journal of Botany 75 1228ndash1237
Wood DM amp del Moral R 2000 Seed rain during early
primary succession onMount St Helens Washington
Madrono 47 1ndash9
Supporting Information
Additional supporting information may befound in the online version of this article
Fig S1 Abraham Plain Sept 1980 pumice de-posits over scoured bedrock looking north towardsthe study site
Fig S2 Abraham Plain July 2009 lookingnorthwest across the study site
Fig S3 Changes in mean cover index for lesscommon species
Fig S4 Relative cover (proportion of totalcover in the sample) of less common species in thefour stages of early primary succession The firstfour species are persistent the last two are pioneerspecies (defined in Appendix S1)
Fig S5 Annual changes of mean H0 and meanSimpsonrsquos dominance index Differences (Po005)among measures determined by a repeat measuresANOVA followed by the Bonferroni test Valueswith same associated letter fall within the samegroup of values
Table S1 Structure and species composition ofCTs found in 2008
Appendix S1 Characteristics of species en-countered on Abraham Plain grid
Please note Wiley-Blackwell is not responsiblefor the content or functionality of any supportingmaterials supplied by the authors Any queries(other than missing material) should be directed tothe corresponding author for the article
Received 9 December 2009
Accepted 14 March 2010
Co-ordinating Editor Dr Kerry Woods
Primary succession trajectories 867

References
Anderson KJ 2007 Temporal patterns in rates of
community change during succession American
Naturalist 169 780ndash793
Anthelme F Villaret JC amp Brun JJ 2007 Shrub
encroachment in the Alps gives rise to the
convergence of sub-alpine communities on a regional
scale Journal of Vegetation Science 18 355ndash362
Baasch A Tischew S amp Bruelheide H 2009 Insights
into succession processes using temporally repeated
habitat models results from a long-term study in a
post-mining landscape Journal of Vegetation Science
20 629ndash638
Bossuyt B Honnay O amp Hermy M 2003 An island
biogeographical view of the successional pathway in
wet dune slacks Journal of Vegetation Science 14 781ndash
788
Clifford HT 1976 Dendrograms and their
interpretation In Williams WT (ed) Pattern
analysis in agricultural science pp 96ndash109 Elsevier
Science Amsterdam NL
del Moral R 2007 Vegetation dynamics in space and
time an example from Mount St Helens Journal of
Vegetation Science 18 479ndash488
del Moral R 2009a Increasing deterministic control of
primary succession onMount St Helens Washington
Journal of Vegetation Science 20 1145ndash1154
del Moral R 2009b Mount St Helens permanent plots
and grids 1980 to present Available at http
protistbiologywashingtonedudelmoral Accessed 8
December 2009
del Moral R amp Bliss LC 1993 Mechanism of primary
succession insights resulting from the eruption of
Mount St Helens Advances in Ecological Research
24 1ndash66
del Moral R amp Ellis EE 2004 Gradients in hetero-
geneity and structure on lahars Mount St Helens
Washington USA Plant Ecology 175 273ndash286
del Moral R amp Lacher IL 2005 Vegetation patterns 25
years after the eruption of Mount St Helens
WashingtonAmerican Journal of Botany 92 1948ndash1956
Donnegan JA amp Rebertus AJ 1999 Rates and
mechanisms of subalpine forest succession along an
environmental gradient Ecology 80 1370ndash1384
Ejrnaes R Bruun HH amp Graae BJ 2006 Community
assembly in experimental grasslands suitable
environment or timely arrival Ecology 87 1225ndash1233
Felinks B ampWiegand T 2008 Exploring spatiotemporal
patterns in early stages of primary succession on
former lignite mining sites Journal of Vegetation
Science 19 267ndash276
Fuller RN amp del Moral R 2003 The role of refugia and
dispersal in primary succession on Mount St Helens
Washington Journal of Vegetation Science 14 637ndash
644
Hodkinson IA Coulson SJ amp Webb NR 2003
Community assembly along proglacial chrono-
sequences in the high Arctic vegetation and soil
development in north-west Svalbard Journal of
Ecology 91 651ndash663
Johnson EA amp Miyanishi K 2008 Testing the
assumptions of chronosequences in succession
Ecology Letters 11 419ndash431
Lanta V amp Leps J 2009 How does surrounding
vegetation affect the course of succession a five-year
container experiment Journal of Vegetation Science
20 686ndash694
Legendre P amp Anderson MJ 1999 Distance-based
redundancy analysis testing multispecies responses in
multi-factorial ecological experiments Ecological
Monographs 69 1ndash24
Leps J amp Rejmanek M 1991 Convergence or
divergence what should we expect from vegetation
succession Oikos 62 261ndash264
Leps J amp Smilauer P 2003 Multivariate analysis of
ecological data using CANOCO Cambridge
University Press Cambridge UK
MacDougall AS Wilson SD amp Bakker JD 2008
Climatic variability alters the outcome of longterm
community assembly Journal of Ecology 96 346ndash354
Navas ML amp Violle C 2009 Plant traits related to
competition how do they shape the functional
diversity of communities Community Ecology 10
131ndash137
Pickett STA Cadenasso ML amp Meiners SJ 2009
Ever since Clements from succession to vegetation
dynamics and understanding to intervention Applied
Vegetation Science 12 9ndash21
Prach K amp Hobbs RJ 2008 Spontaneous succession
versus technical reclamation in the restoration of
disturbed sites Restoration Ecology 16 363ndash366
Prach K amp Rehounkova K 2006 Vegetation succession
over broad geographical scales which factors
determine the patterns Preslia 78 469ndash480
Robbins JA amp Matthews JA 2009 Pioneer vegetation
on glacier forelands in southern Norway emerging
communities Journal of Vegetation Science 20 889ndash
902
Swanson FJ amp Major JJ 2005 Physical events
environments and geologicalndashecological interactions
at Mount St Helens March 1980ndash2004 In Dale
VH Swanson FJ amp Crisafulli CM (eds)
Ecological recovery after the 1980 eruption of Mount
St Helens pp 27ndash44 Springer New York NY US
USDA National Resource Conservation Service 2009
The Plants Data Base National Plants Data Center
Baton Rouge LA US Available at http
plantsusdagov Accessed October 15 2009
van Oijen D Hommel P den Ouden J amp de Waal R
2005 Effects of tree species composition on within-
forest distribution of understorey species Applied
Vegetation Science 8 155ndash166
Walker LR amp del Moral R 2003 Primary succession
and ecosystem rehabilitation Cambridge University
Press Cambridge UK
866 del Moral Roger et al
Walker LR amp del Moral R 2009 Transition dynamics
in succession implications for rates trajectories and
restoration In Hobbs RJ amp Suding KN (eds)
New models for ecosystem dynamics and restoration
pp 33ndash49 Island Press Washington DC US
Walker LR Bellingham PJ amp Peltzer DA 2006
Plant characteristics are poor predictors of microsite
colonization during the first two years of pri-
mary succession Journal of Vegetation Science 17
397ndash406
Wood DM amp del Moral R 1988 Colonizing plants on
the Pumice Plains Mount St Helens Washington
American Journal of Botany 75 1228ndash1237
Wood DM amp del Moral R 2000 Seed rain during early
primary succession onMount St Helens Washington
Madrono 47 1ndash9
Supporting Information
Additional supporting information may befound in the online version of this article
Fig S1 Abraham Plain Sept 1980 pumice de-posits over scoured bedrock looking north towardsthe study site
Fig S2 Abraham Plain July 2009 lookingnorthwest across the study site
Fig S3 Changes in mean cover index for lesscommon species
Fig S4 Relative cover (proportion of totalcover in the sample) of less common species in thefour stages of early primary succession The firstfour species are persistent the last two are pioneerspecies (defined in Appendix S1)
Fig S5 Annual changes of mean H0 and meanSimpsonrsquos dominance index Differences (Po005)among measures determined by a repeat measuresANOVA followed by the Bonferroni test Valueswith same associated letter fall within the samegroup of values
Table S1 Structure and species composition ofCTs found in 2008
Appendix S1 Characteristics of species en-countered on Abraham Plain grid
Please note Wiley-Blackwell is not responsiblefor the content or functionality of any supportingmaterials supplied by the authors Any queries(other than missing material) should be directed tothe corresponding author for the article
Received 9 December 2009
Accepted 14 March 2010
Co-ordinating Editor Dr Kerry Woods
Primary succession trajectories 867

Walker LR amp del Moral R 2009 Transition dynamics
in succession implications for rates trajectories and
restoration In Hobbs RJ amp Suding KN (eds)
New models for ecosystem dynamics and restoration
pp 33ndash49 Island Press Washington DC US
Walker LR Bellingham PJ amp Peltzer DA 2006
Plant characteristics are poor predictors of microsite
colonization during the first two years of pri-
mary succession Journal of Vegetation Science 17
397ndash406
Wood DM amp del Moral R 1988 Colonizing plants on
the Pumice Plains Mount St Helens Washington
American Journal of Botany 75 1228ndash1237
Wood DM amp del Moral R 2000 Seed rain during early
primary succession onMount St Helens Washington
Madrono 47 1ndash9
Supporting Information
Additional supporting information may befound in the online version of this article
Fig S1 Abraham Plain Sept 1980 pumice de-posits over scoured bedrock looking north towardsthe study site
Fig S2 Abraham Plain July 2009 lookingnorthwest across the study site
Fig S3 Changes in mean cover index for lesscommon species
Fig S4 Relative cover (proportion of totalcover in the sample) of less common species in thefour stages of early primary succession The firstfour species are persistent the last two are pioneerspecies (defined in Appendix S1)
Fig S5 Annual changes of mean H0 and meanSimpsonrsquos dominance index Differences (Po005)among measures determined by a repeat measuresANOVA followed by the Bonferroni test Valueswith same associated letter fall within the samegroup of values
Table S1 Structure and species composition ofCTs found in 2008
Appendix S1 Characteristics of species en-countered on Abraham Plain grid
Please note Wiley-Blackwell is not responsiblefor the content or functionality of any supportingmaterials supplied by the authors Any queries(other than missing material) should be directed tothe corresponding author for the article
Received 9 December 2009
Accepted 14 March 2010
Co-ordinating Editor Dr Kerry Woods
Primary succession trajectories 867
Related Documents