Presymbiotic growth and sporal morphology are affected in the arbuscular mycorrhizal fungus Gigaspora margarita cured of its endobacteria Erica Lumini, 1† Valeria Bianciotto, 1† Patricia Jargeat, 2,3† Mara Novero, 1 Alessandra Salvioli, 1 Antonella Faccio, 1 Guillaume Bécard 3 and Paola Bonfante 1 * 1 Dipartimento di Biologia Vegetale dell’Università and Istituto per la Protezione delle Piante – CNR, Viale Mattioli 25, 10125-I, Torino, Italy. 2 UMR 5174 University of Toulouse 3/CNRS/ENFA, 118, route de Narbonne, 31062 Toulouse, France. 3 UMR 5546 University of Toulouse 3/CNRS, 24 chemin de Borde-Rouge, 31326 Castanet-Tolosan, Toulouse, France. Summary Some arbuscular mycorrhizal fungi contain endocel- lular bacteria. In Gigaspora margarita BEG 34, a homogenous population of b-Proteobacteria is hosted inside the fungal spore. The bacteria, named Candidatus Glomeribacter gigasporarum, are verti- cally transmitted through fungal spore generations. Here we report how a protocol based on repeated passages through single-spore inocula caused dilu- tion of the initial bacterial population eventually leading to cured spores. Spores of this line had a distinct phenotype regarding cytoplasm organization, vacuole morphology, cell wall organization, lipid bodies and pigment granules. The absence of bacteria severely affected presymbiotic fungal growth such as hyphal elongation and branching after root exudate treatment, suggesting that Ca. Glomeribacter gigasporarum is important for optimal development of its fungal host. Under laboratory conditions, the cured fungus could be propagated, i.e. could form mycorrhizae and sporulate, and can therefore be considered as a stable variant of the wild type. The results demonstrated that – at least for the G. margarita BEG 34 isolate – the absence of endo- bacteria affects the spore phenotype of the fungal host, and causes delays in the growth of germinating mycelium, possibly affecting its ecological fitness. This cured line is the first manipulated and stable isolate of an arbuscular mycorrhizal fungus. Introduction Arbuscular mycorrhizal (AM) fungi colonize the root cortex of the majority of land plants, supplying them with nutri- ents, including phosphate, and conferring resistance against a variety of biotic and abiotic stresses (Smith and Read, 1997). In addition to the strong emphasis which is currently placed on the ecological roles played by AM fungi in nature (Van der Heijden and Sanders, 2002), there are other aspects of their biology which make them unique microbes: they are obligate biotrophs, meaning they get carbon exclusively through a symbiotic relation- ship with a host plant, lack host specificity (being present in approximately 80% of land plants), and are multinucle- ate, opening the question whether they are heterokaryotic (Hijri and Sanders, 2005) or homocaryotic polyploid organisms (Pawlowska and Taylor, 2004). In addition and as a further element of their genomic complexity, AM fungi offer the best-known example of an association between fungi and bacteria (Bonfante, 2003). Many bacteria com- plete their life cycle within eukaryotic cells (Moran, 2001), but the Fungal Kingdom offers a limited number of examples of endobacteria (Partida-Martinez and Hertweck, 2005; Lumini et al., 2006). Glomeromycota rep- resent a niche for a largely undescribed population of bacteria (Scannerini and Bonfante, 1991; Schüßler, 2002). Until a few years ago, the biology of these bacterial populations – with only their morphology described – was a black box. Recently we have shown that an isolate of Gigaspora margarita hosts a homogenous population of bacteria (at least 20 000 per fungal spore) (Bianciotto et al., 2003; Jargeat et al., 2004; Lumini et al., 2006). The spores of G. margarita seem to be a specialized niche for rod-shaped bacteria which have also been consistently found in many other Gigasporaceae in all stages of their life cycle (Bianciotto et al., 1996; 2000). On the basis of their 16S RNA sequence, the bacteria have been grouped into a new taxon, which – due to their current unculturable status – has been named as Candidatus Glomeribacter gigasporarum (Bianciotto et al., 2003), clustering within Received 10 July, 2006; revised and accepted 23 January, 2007. *For correspondence. E-mail [email protected]; Tel. (+39) 0116502927; Fax (+39) 0116705962. † These authors contri- buted equally to the work. Cellular Microbiology (2007) 9(7), 1716–1729 doi:10.1111/j.1462-5822.2007.00907.x First published online 28 February 2007 © 2007 The Authors Journal compilation © 2007 Blackwell Publishing Ltd

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Presymbiotic growth and sporal morphology areaffected in the arbuscular mycorrhizal fungusGigaspora margarita cured of its endobacteria
Erica Lumini,1† Valeria Bianciotto,1†
Patricia Jargeat,2,3† Mara Novero,1
Alessandra Salvioli,1 Antonella Faccio,1
Guillaume Bécard3 and Paola Bonfante1*1Dipartimento di Biologia Vegetale dell’Università andIstituto per la Protezione delle Piante – CNR, VialeMattioli 25, 10125-I, Torino, Italy.2UMR 5174 University of Toulouse 3/CNRS/ENFA, 118,route de Narbonne, 31062 Toulouse, France.3UMR 5546 University of Toulouse 3/CNRS, 24 cheminde Borde-Rouge, 31326 Castanet-Tolosan, Toulouse,France.
Summary
Some arbuscular mycorrhizal fungi contain endocel-lular bacteria. In Gigaspora margarita BEG 34, ahomogenous population of b-Proteobacteria ishosted inside the fungal spore. The bacteria, namedCandidatus Glomeribacter gigasporarum, are verti-cally transmitted through fungal spore generations.Here we report how a protocol based on repeatedpassages through single-spore inocula caused dilu-tion of the initial bacterial population eventuallyleading to cured spores. Spores of this line had adistinct phenotype regarding cytoplasm organization,vacuole morphology, cell wall organization, lipidbodies and pigment granules. The absence ofbacteria severely affected presymbiotic fungal growthsuch as hyphal elongation and branching after rootexudate treatment, suggesting that Ca. Glomeribactergigasporarum is important for optimal developmentof its fungal host. Under laboratory conditions, thecured fungus could be propagated, i.e. could formmycorrhizae and sporulate, and can therefore beconsidered as a stable variant of the wild type. Theresults demonstrated that – at least for theG. margarita BEG 34 isolate – the absence of endo-bacteria affects the spore phenotype of the fungalhost, and causes delays in the growth of germinating
mycelium, possibly affecting its ecological fitness.This cured line is the first manipulated and stableisolate of an arbuscular mycorrhizal fungus.
Introduction
Arbuscular mycorrhizal (AM) fungi colonize the root cortexof the majority of land plants, supplying them with nutri-ents, including phosphate, and conferring resistanceagainst a variety of biotic and abiotic stresses (Smith andRead, 1997). In addition to the strong emphasis which iscurrently placed on the ecological roles played by AMfungi in nature (Van der Heijden and Sanders, 2002),there are other aspects of their biology which make themunique microbes: they are obligate biotrophs, meaningthey get carbon exclusively through a symbiotic relation-ship with a host plant, lack host specificity (being presentin approximately 80% of land plants), and are multinucle-ate, opening the question whether they are heterokaryotic(Hijri and Sanders, 2005) or homocaryotic polyploidorganisms (Pawlowska and Taylor, 2004). In addition andas a further element of their genomic complexity, AM fungioffer the best-known example of an association betweenfungi and bacteria (Bonfante, 2003). Many bacteria com-plete their life cycle within eukaryotic cells (Moran, 2001),but the Fungal Kingdom offers a limited number ofexamples of endobacteria (Partida-Martinez andHertweck, 2005; Lumini et al., 2006). Glomeromycota rep-resent a niche for a largely undescribed population ofbacteria (Scannerini and Bonfante, 1991; Schüßler,2002). Until a few years ago, the biology of these bacterialpopulations – with only their morphology described – wasa black box. Recently we have shown that an isolate ofGigaspora margarita hosts a homogenous population ofbacteria (at least 20 000 per fungal spore) (Bianciottoet al., 2003; Jargeat et al., 2004; Lumini et al., 2006). Thespores of G. margarita seem to be a specialized niche forrod-shaped bacteria which have also been consistentlyfound in many other Gigasporaceae in all stages of theirlife cycle (Bianciotto et al., 1996; 2000). On the basis oftheir 16S RNA sequence, the bacteria have been groupedinto a new taxon, which – due to their current unculturablestatus – has been named as Candidatus Glomeribactergigasporarum (Bianciotto et al., 2003), clustering within
Received 10 July, 2006; revised and accepted 23 January, 2007.*For correspondence. E-mail [email protected]; Tel.(+39) 0116502927; Fax (+39) 0116705962. †These authors contri-buted equally to the work.
Cellular Microbiology (2007) 9(7), 1716–1729 doi:10.1111/j.1462-5822.2007.00907.xFirst published online 28 February 2007
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd

the large group of b-Proteobacteria related to theBurkholderia, Pandorea and Ralstonia genera. Theirgenome is highly reduced: 1.4 Mb consisting of a chro-mosome and a plasmid (Jargeat et al., 2004). The statusof uncultivable microbe and this small genome stronglysuggest that Ca. Glomeribacter gigasporarum is an obli-gate endosymbiont, like other comparable endosymbiontsliving in aphids (Moran, 2003), and justifies fundamentalquestions regarding the exact nature of a potentially inti-mate fungus–bacterium symbiotic relationship.
As antibiotic treatments to cure G. margarita from itsendobacteria were unsuccessful (P. Jargeat and G.Bécard, unpubl. results), biological material appropriate tounderstand the role of Ca. Glomeribacter gigasporarumwas not available. Here, we report how a protocol basedon the use of successive in vitro single-spore inocula(Bianciotto et al., 2004) caused a dilution of the microbialpopulation eventually leading to bacteria-cured spores.Morphological analyses as well as mycorrhizal experi-ments showed that the cured spore line (B–) was a stablevariant of the original wild type (WT) (B+), showing clear-cut morphological differences and important changes inthe growth of the germinating mycelium.
Results
Production of G. margarita cured spores
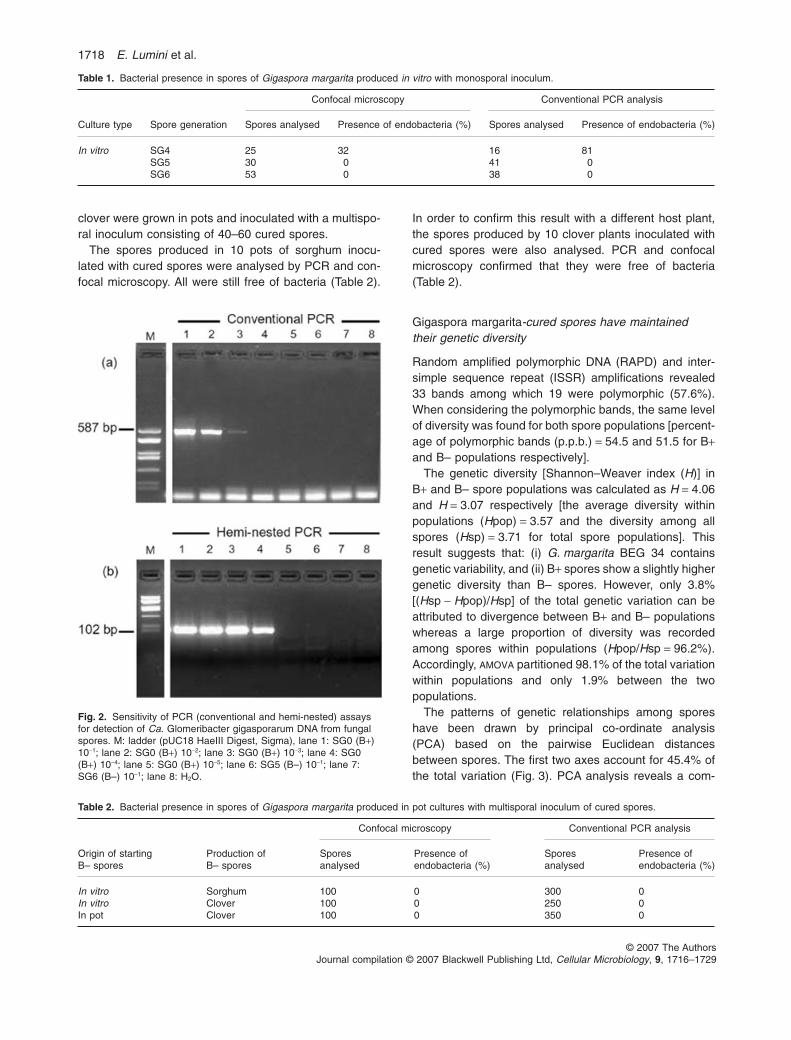
Successive spore generations (SGs) were obtained afterinoculating transformed carrot roots with single-sporeinocula originating from two separate batch pots. Sporesoriginated from a single SG0 spore were identified as a lot(Fig. 1). Six lots were considered during the experiments.As previously described (Bianciotto et al., 2004), theendobacterial population in successive spore generationswas found to rapidly decrease from SG0 to SG4. Here(irrespectively of the starting lot), 68% of the SG4 sporeswere found free of bacteria when observed by confocalmicroscopy (Table 1), while conventional PCR showedthat only 19% of SG4 spores were indeed free of bacteria(Table 1). The discrepancy between the two methods sug-gests a different level of sensitivity. Single SG4 sporeswere used to inoculate new root cultures in order to obtainSG5 and SG6 spores. No endobacteria were detected byconfocal microscopy and by conventional PCR in SG5and SG6 spores (Table 1).
To assess the sensitivity of the PCR technique to detectthe endobacteria in our biological system serial dilutionsof DNA template from the WT spores (B+) were amplifiedwith the specific 23S rDNA bacterial primers in conven-tional and hemi-nested PCR (Fig. 2). With conventionalPCR the amplification product was visible until 10-3 sporalDNA dilution, while the hemi-nested technique detectedthe presence of the endobacteria until the 10-4 dilution.
Real-time quantitative PCR assays confirmed the dataobtained by hemi-nested technique, allowing us to detecta minimum of 10 bacterial genomes for each PCR mixturein the same 10-4 dilution (A. Salvioli, E. Lumini and V.Bianciotto, unpubl. results). DNA extracts from randomlyselected SG5 and SG6 spores did not give any amplifica-tion, even in undiluted samples (Fig. 2). The fungal DNAwas detected in all the dilutions tested, demonstrating thegood quality of the DNA extract and the absence ofinhibition. The results show that the hemi-nested PCRtechnique is a suitable tool to detect endobacteria,increasing the level of sensitivity of conventional PCR atleast 10-fold.
Cured spores of G. margarita can still complete theirlife cycle
To examine the stability of the bacteria-free status ofcured spores, and to obtain them in quantity, sorghum and
Fig. 1. Schematic representation of the conditions for theproduction and maintaining of Gigaspora margarita-cured spores.The production starts from a single batch (one pot) and the singleSG0 spore originates a lot of cured spore. The figure describes theproduction of one lot. The spores (coloured spots) change from red(WT) to yellow (Cured).
Endocellular bacteria affect AMF presymbiotic growth and morphology 1717
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Cellular Microbiology, 9, 1716–1729
clover were grown in pots and inoculated with a multispo-ral inoculum consisting of 40–60 cured spores.
The spores produced in 10 pots of sorghum inocu-lated with cured spores were analysed by PCR and con-focal microscopy. All were still free of bacteria (Table 2).
In order to confirm this result with a different host plant,the spores produced by 10 clover plants inoculated withcured spores were also analysed. PCR and confocalmicroscopy confirmed that they were free of bacteria(Table 2).
Gigaspora margarita-cured spores have maintainedtheir genetic diversity
Random amplified polymorphic DNA (RAPD) and inter-simple sequence repeat (ISSR) amplifications revealed33 bands among which 19 were polymorphic (57.6%).When considering the polymorphic bands, the same levelof diversity was found for both spore populations [percent-age of polymorphic bands (p.p.b.) = 54.5 and 51.5 for B+and B– populations respectively].
The genetic diversity [Shannon–Weaver index (H)] inB+ and B– spore populations was calculated as H = 4.06and H = 3.07 respectively [the average diversity withinpopulations (Hpop) = 3.57 and the diversity among allspores (Hsp) = 3.71 for total spore populations]. Thisresult suggests that: (i) G. margarita BEG 34 containsgenetic variability, and (ii) B+ spores show a slightly highergenetic diversity than B– spores. However, only 3.8%[(Hsp - Hpop)/Hsp] of the total genetic variation can beattributed to divergence between B+ and B– populationswhereas a large proportion of diversity was recordedamong spores within populations (Hpop/Hsp = 96.2%).Accordingly, AMOVA partitioned 98.1% of the total variationwithin populations and only 1.9% between the twopopulations.
The patterns of genetic relationships among sporeshave been drawn by principal co-ordinate analysis(PCA) based on the pairwise Euclidean distancesbetween spores. The first two axes account for 45.4% ofthe total variation (Fig. 3). PCA analysis reveals a com-
Table 1. Bacterial presence in spores of Gigaspora margarita produced in vitro with monosporal inoculum.
Culture type Spore generation
Confocal microscopy Conventional PCR analysis
Spores analysed Presence of endobacteria (%) Spores analysed Presence of endobacteria (%)
In vitro SG4 25 32 16 81SG5 30 0 41 0SG6 53 0 38 0
Fig. 2. Sensitivity of PCR (conventional and hemi-nested) assaysfor detection of Ca. Glomeribacter gigasporarum DNA from fungalspores. M: ladder (pUC18 HaeIII Digest, Sigma), lane 1: SG0 (B+)10-1; lane 2: SG0 (B+) 10-2; lane 3: SG0 (B+) 10-3; lane 4: SG0(B+) 10-4; lane 5: SG0 (B+) 10-5; lane 6: SG5 (B–) 10-1; lane 7:SG6 (B–) 10-1; lane 8: H2O.
Table 2. Bacterial presence in spores of Gigaspora margarita produced in pot cultures with multisporal inoculum of cured spores.
Origin of startingB– spores
Production ofB– spores
Confocal microscopy Conventional PCR analysis
Sporesanalysed
Presence ofendobacteria (%)
Sporesanalysed
Presence ofendobacteria (%)
In vitro Sorghum 100 0 300 0In vitro Clover 100 0 250 0In pot Clover 100 0 350 0
1718 E. Lumini et al.
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Cellular Microbiology, 9, 1716–1729

plete overlap among B+ and B– spores, without anyclustering.
Gigaspora margarita-cured spores do not appeardifferent in mycorrhizal phenotype
To assess more carefully the infectivity of cured spores,mycorrhization experiments were set up in pots ofsorghum and clover. Sorghum pots were inoculated withB+ spores and four lots of B– spores, identified as B-1,B-2, B-3 and B-4 (i.e. groups of cured spores originatedfrom independent SG0 spores). Clover pots were inocu-lated with B+ spores and one additional lot of B– spores(lot 5). The frequency of mycorrhization (F), the intensityof mycorrhization (M), the percentage of arbuscules per
infected areas (a) and per root apparatus (A) were notdifferent between plants inoculated with B+ spores or B–spores (Table 3). Similar results showing no differencebetween B+ and B– mycorrhization were obtained withclover as well as with the in vitro system using trans-formed carrot roots (Table 3).
No cytological difference was noted, using light micros-copy and electron microscopy, in the respective mycor-rhizal structures observed in clover roots. The integrity ofimportant biotrophic structures such as the plant mem-brane surrounding the arbuscules looked normal in B–mycorrhizae (not shown).
The experiment with sorghum allowed us to evaluatesporulation of the cured line versus that of the original line.No significant difference was observed between the lines.After 3 months the mean number of spores produced was980.2 and 875.4 for WT and cured spores respectively(Table 3). Similarly, no significant difference was foundafter 3 months in the spore production of the in vitrosystem using carrot hairy roots (Table 3).
Gigaspora margarita-cured spores showedmorphological changes
Spores from both B+ and B– lines had the same meansize, with a diameter ranging from 333 to 325 mm(Table 4). In agreement with previous descriptions(Bonfante et al., 1994), the B+ spores of G. margaritaBEG 34 had a laminated wall and complex protoplasm(Fig. 4A–C), containing many nuclei, membranous secre-
Table 3. Mycorrhizal colonization intensity.
Host plant F% M% a% A% Spores number
Sorghum WT 49.50 (a) 25.98 (b) 70.40 (c,d) 21.93 (e) 980.2 (f)Sorghum-cured B-1 40.56 (a) 21.13 (b) 82.66 (c) 17.93 (e) 576.8 (f)Sorghum-cured B-2 45.17 (a) 23.41 (b) 87.85 (c) 24.31 (e) 679.8 (f)Sorghum-cured B-3 61.61 (a) 30.76 (b) 86.89 (c) 26.86 (e) 1111.5 (f)Sorghum-cured B-4 61.11 (a) 43.49 (b) 61.30 (d) 27.03 (e) 1133.5 (f)Clover WT 58.43 (a) 20.57 (b) 88.60 (c) 19.00 (d)Clover cured 61.00 (a) 21.40 (b) 80.30 (c) 21.78 (d)Transformed carrot root WT 46.25 (a) 8.65 (b) 87.72 (c) 7.59 (d) 31 (e)Transformed carrot root cured 47.50 (a) 11.18 (b) 82.89 (c) 9.26 (d) 31 (e)
Different letters indicate significantly different values according to the Krukall–Wallis test of variance (P < 0.05). Statistical analyses of clover andsorghum data were treated separately.
Fig. 3. PCA of 33 RAPD and ISSR markers in Gigaspora margaritaspores containing (B+) or not (B–) the endobacterium Ca.Glomeribacter gigasporarum.
Table 4. Main features of WT and cured spores of Gigasporamargarita.
Cured spore WT spore
Spore diameter (mm) 333 (a) 324.75 (a)Wall thickness (mm) 16.1 (b) 12.15 (c)Nuclear diameter (mm) 6.5 (d) 6.3 (d)
Different letters indicate significantly different values according to theKrukall–Wallis test of variance (P < 0.01).
Endocellular bacteria affect AMF presymbiotic growth and morphology 1719
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Cellular Microbiology, 9, 1716–1729
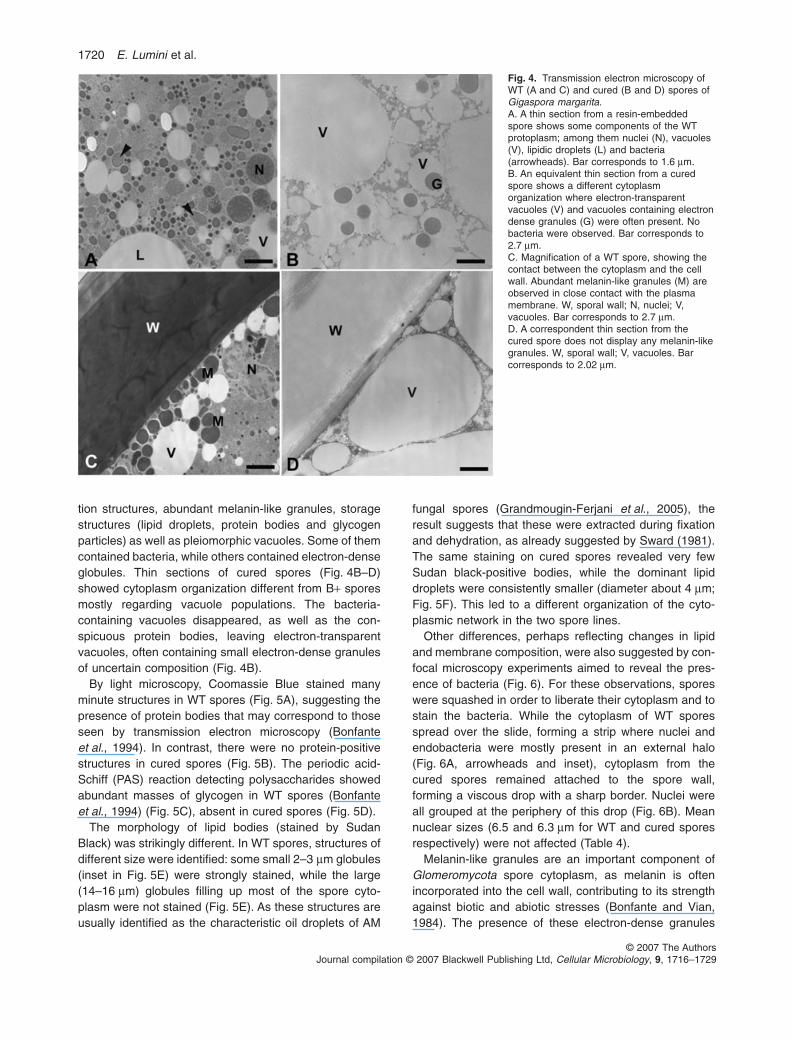
tion structures, abundant melanin-like granules, storagestructures (lipid droplets, protein bodies and glycogenparticles) as well as pleiomorphic vacuoles. Some of themcontained bacteria, while others contained electron-denseglobules. Thin sections of cured spores (Fig. 4B–D)showed cytoplasm organization different from B+ sporesmostly regarding vacuole populations. The bacteria-containing vacuoles disappeared, as well as the con-spicuous protein bodies, leaving electron-transparentvacuoles, often containing small electron-dense granulesof uncertain composition (Fig. 4B).
By light microscopy, Coomassie Blue stained manyminute structures in WT spores (Fig. 5A), suggesting thepresence of protein bodies that may correspond to thoseseen by transmission electron microscopy (Bonfanteet al., 1994). In contrast, there were no protein-positivestructures in cured spores (Fig. 5B). The periodic acid-Schiff (PAS) reaction detecting polysaccharides showedabundant masses of glycogen in WT spores (Bonfanteet al., 1994) (Fig. 5C), absent in cured spores (Fig. 5D).
The morphology of lipid bodies (stained by SudanBlack) was strikingly different. In WT spores, structures ofdifferent size were identified: some small 2–3 mm globules(inset in Fig. 5E) were strongly stained, while the large(14–16 mm) globules filling up most of the spore cyto-plasm were not stained (Fig. 5E). As these structures areusually identified as the characteristic oil droplets of AM
fungal spores (Grandmougin-Ferjani et al., 2005), theresult suggests that these were extracted during fixationand dehydration, as already suggested by Sward (1981).The same staining on cured spores revealed very fewSudan black-positive bodies, while the dominant lipiddroplets were consistently smaller (diameter about 4 mm;Fig. 5F). This led to a different organization of the cyto-plasmic network in the two spore lines.
Other differences, perhaps reflecting changes in lipidand membrane composition, were also suggested by con-focal microscopy experiments aimed to reveal the pres-ence of bacteria (Fig. 6). For these observations, sporeswere squashed in order to liberate their cytoplasm and tostain the bacteria. While the cytoplasm of WT sporesspread over the slide, forming a strip where nuclei andendobacteria were mostly present in an external halo(Fig. 6A, arrowheads and inset), cytoplasm from thecured spores remained attached to the spore wall,forming a viscous drop with a sharp border. Nuclei wereall grouped at the periphery of this drop (Fig. 6B). Meannuclear sizes (6.5 and 6.3 mm for WT and cured sporesrespectively) were not affected (Table 4).
Melanin-like granules are an important component ofGlomeromycota spore cytoplasm, as melanin is oftenincorporated into the cell wall, contributing to its strengthagainst biotic and abiotic stresses (Bonfante and Vian,1984). The presence of these electron-dense granules
Fig. 4. Transmission electron microscopy ofWT (A and C) and cured (B and D) spores ofGigaspora margarita.A. A thin section from a resin-embeddedspore shows some components of the WTprotoplasm; among them nuclei (N), vacuoles(V), lipidic droplets (L) and bacteria(arrowheads). Bar corresponds to 1.6 mm.B. An equivalent thin section from a curedspore shows a different cytoplasmorganization where electron-transparentvacuoles (V) and vacuoles containing electrondense granules (G) were often present. Nobacteria were observed. Bar corresponds to2.7 mm.C. Magnification of a WT spore, showing thecontact between the cytoplasm and the cellwall. Abundant melanin-like granules (M) areobserved in close contact with the plasmamembrane. W, sporal wall; N, nuclei; V,vacuoles. Bar corresponds to 2.7 mm.D. A correspondent thin section from thecured spore does not display any melanin-likegranules. W, sporal wall; V, vacuoles. Barcorresponds to 2.02 mm.
1720 E. Lumini et al.
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Cellular Microbiology, 9, 1716–1729

was decreased in the cured spores. In contrast with theWT spores (Fig. 4C), there was no evidence of their incor-poration into the fungal wall (Fig. 4D).
The lamination of the spore wall appeared the same inboth fungal populations, as also confirmed by PAS stain-ing for polysaccharides, but walls of cured spores weresignificantly thicker than those of WT spores (16.1 mmversus 12.15) (Fig. 7 and Table 4).
More detailed differences in the cell wall of the twolines could be seen by electron microscopy. The cell wallinner layers (Iw) were comparable in organization butnot in thickness. The middle layer (Mw) always showedthe architecture described as helicoidal (Bonfante andVian, 1984), and contained many chitin-rich sheets(Fig. 7C and D). Surprisingly, additional sheets werepresent in the middle layer of cured spores, and also in
Fig. 5. Light microscopy of WT (left) andcured spores (right) of Gigaspora margarita.A and B. Blue Coomassie protein staining. Inthe WT spores (A) there are many positiveminute structures (arrowheads) suggestingthe presence of proteinaceous bodies. In thecured spores (B) the staining did not revealany protein-positive structure.C and D. Polysaccharides PAS detection.Positive masses corresponding to glycogenparticles (arrowheads) are present in WTspores (C) while comparable staining is notdetectable in the cured spores (D).E and F. Sudan black lipid staining. The WTspores (E) display an intense black staining ofa population of globular bodies (arrowheads).Inset: detail of the black stained globularminute bodies. Some minute Sudanblack-positive bodies are revealed on thecured spores among the lipid droplets (F).Bar corresponds to 20 mm in (A)–(E), to10 mm in the inset and to 15 mm in (F). W,sporal wall; L, lipid droplet.
Fig. 6. Manually crushed WT (A) and curedspores (B) of Gigaspora margarita (S) stainedwith the bacteria counting kit and observed byusing an Olympus FluoView confocalmicroscope. In the WT spores (A) thecytoplasm spreads over the slide forming astripe where endobacteria were mostlypresent in an external halo (arrowheads). Inthe inset fungal nuclei (N) and endobacteria(arrowheads) fluoresce bright green underblue light. The cytoplasm from the curedspores (B) is dense and forms a drop with aclear cut edge. No bacterial endosymbiontswere detected in the cytoplasm, only fungalnuclei (N) are visible (inset). Bars correspondto 72 mm in (A), 66 mm in (B) and 32 mm inthe insets.
Endocellular bacteria affect AMF presymbiotic growth and morphology 1721
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Cellular Microbiology, 9, 1716–1729
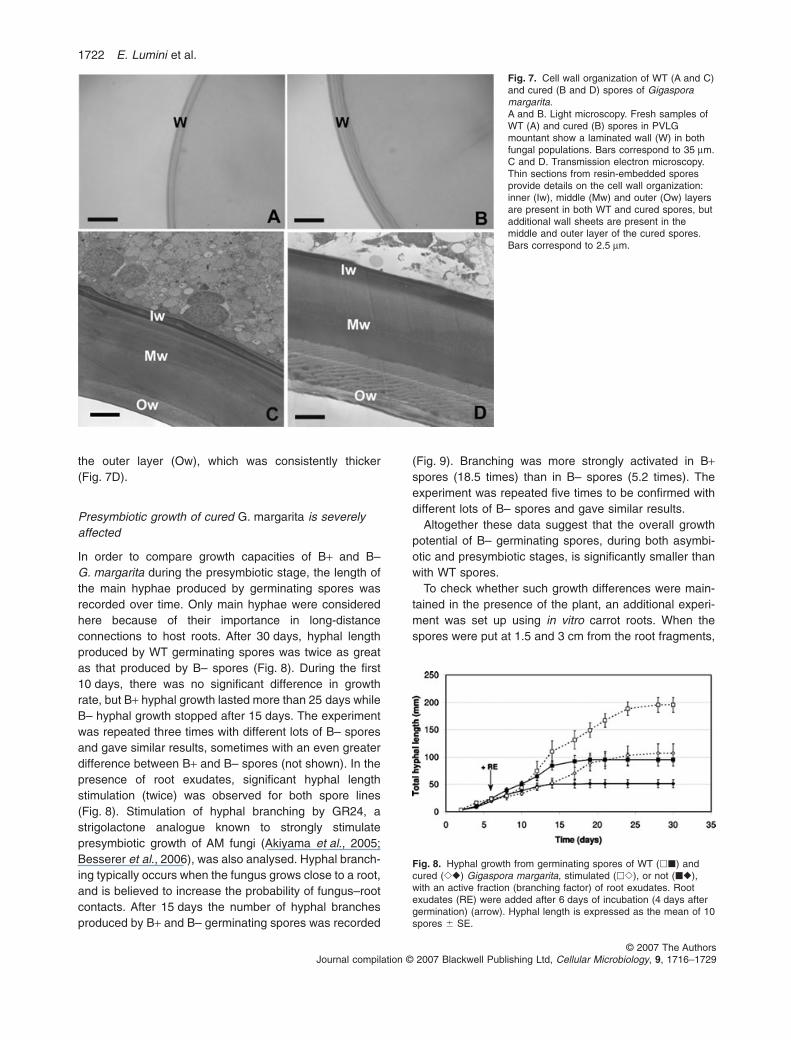
the outer layer (Ow), which was consistently thicker(Fig. 7D).
Presymbiotic growth of cured G. margarita is severelyaffected
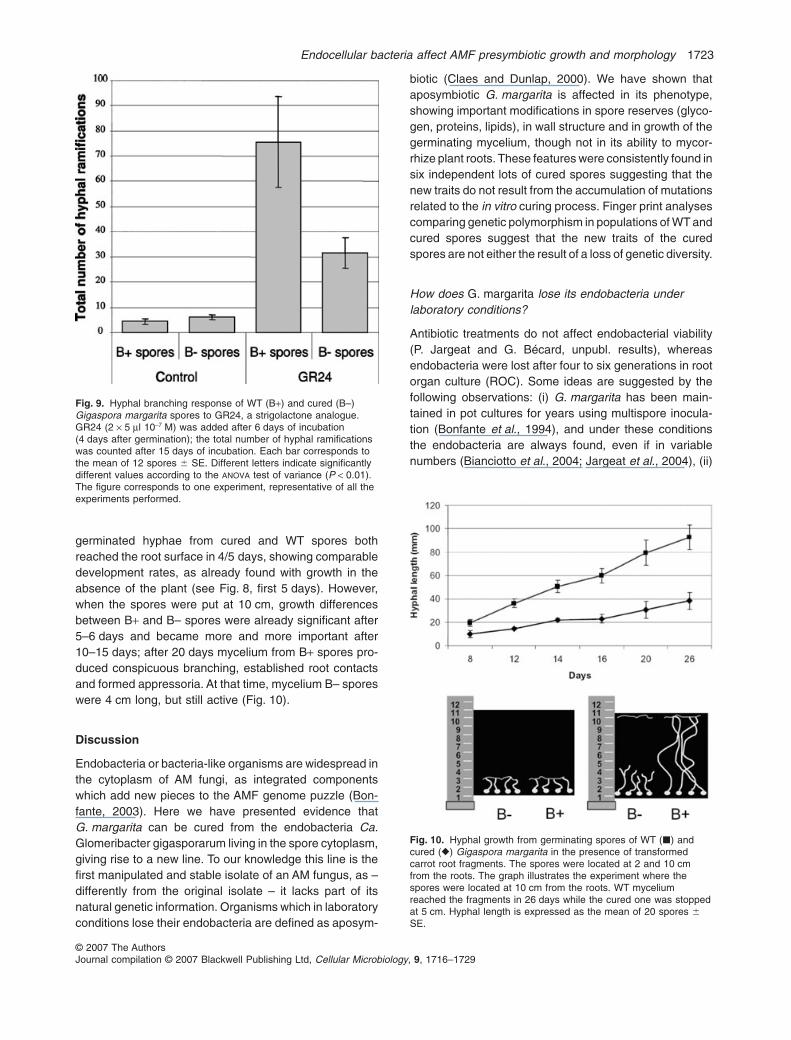
In order to compare growth capacities of B+ and B–G. margarita during the presymbiotic stage, the length ofthe main hyphae produced by germinating spores wasrecorded over time. Only main hyphae were consideredhere because of their importance in long-distanceconnections to host roots. After 30 days, hyphal lengthproduced by WT germinating spores was twice as greatas that produced by B– spores (Fig. 8). During the first10 days, there was no significant difference in growthrate, but B+ hyphal growth lasted more than 25 days whileB– hyphal growth stopped after 15 days. The experimentwas repeated three times with different lots of B– sporesand gave similar results, sometimes with an even greaterdifference between B+ and B– spores (not shown). In thepresence of root exudates, significant hyphal lengthstimulation (twice) was observed for both spore lines(Fig. 8). Stimulation of hyphal branching by GR24, astrigolactone analogue known to strongly stimulatepresymbiotic growth of AM fungi (Akiyama et al., 2005;Besserer et al., 2006), was also analysed. Hyphal branch-ing typically occurs when the fungus grows close to a root,and is believed to increase the probability of fungus–rootcontacts. After 15 days the number of hyphal branchesproduced by B+ and B– germinating spores was recorded
(Fig. 9). Branching was more strongly activated in B+spores (18.5 times) than in B– spores (5.2 times). Theexperiment was repeated five times to be confirmed withdifferent lots of B– spores and gave similar results.
Altogether these data suggest that the overall growthpotential of B– germinating spores, during both asymbi-otic and presymbiotic stages, is significantly smaller thanwith WT spores.
To check whether such growth differences were main-tained in the presence of the plant, an additional experi-ment was set up using in vitro carrot roots. When thespores were put at 1.5 and 3 cm from the root fragments,
Fig. 7. Cell wall organization of WT (A and C)and cured (B and D) spores of Gigasporamargarita.A and B. Light microscopy. Fresh samples ofWT (A) and cured (B) spores in PVLGmountant show a laminated wall (W) in bothfungal populations. Bars correspond to 35 mm.C and D. Transmission electron microscopy.Thin sections from resin-embedded sporesprovide details on the cell wall organization:inner (Iw), middle (Mw) and outer (Ow) layersare present in both WT and cured spores, butadditional wall sheets are present in themiddle and outer layer of the cured spores.Bars correspond to 2.5 mm.
Fig. 8. Hyphal growth from germinating spores of WT (��) andcured (��) Gigaspora margarita, stimulated (��), or not (��),with an active fraction (branching factor) of root exudates. Rootexudates (RE) were added after 6 days of incubation (4 days aftergermination) (arrow). Hyphal length is expressed as the mean of 10spores � SE.
1722 E. Lumini et al.
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Cellular Microbiology, 9, 1716–1729
germinated hyphae from cured and WT spores bothreached the root surface in 4/5 days, showing comparabledevelopment rates, as already found with growth in theabsence of the plant (see Fig. 8, first 5 days). However,when the spores were put at 10 cm, growth differencesbetween B+ and B– spores were already significant after5–6 days and became more and more important after10–15 days; after 20 days mycelium from B+ spores pro-duced conspicuous branching, established root contactsand formed appressoria. At that time, mycelium B– sporeswere 4 cm long, but still active (Fig. 10).
Discussion
Endobacteria or bacteria-like organisms are widespread inthe cytoplasm of AM fungi, as integrated componentswhich add new pieces to the AMF genome puzzle (Bon-fante, 2003). Here we have presented evidence thatG. margarita can be cured from the endobacteria Ca.Glomeribacter gigasporarum living in the spore cytoplasm,giving rise to a new line. To our knowledge this line is thefirst manipulated and stable isolate of an AM fungus, as –differently from the original isolate – it lacks part of itsnatural genetic information. Organisms which in laboratoryconditions lose their endobacteria are defined as aposym-
biotic (Claes and Dunlap, 2000). We have shown thataposymbiotic G. margarita is affected in its phenotype,showing important modifications in spore reserves (glyco-gen, proteins, lipids), in wall structure and in growth of thegerminating mycelium, though not in its ability to mycor-rhize plant roots. These features were consistently found insix independent lots of cured spores suggesting that thenew traits do not result from the accumulation of mutationsrelated to the in vitro curing process. Finger print analysescomparing genetic polymorphism in populations of WT andcured spores suggest that the new traits of the curedspores are not either the result of a loss of genetic diversity.
How does G. margarita lose its endobacteria underlaboratory conditions?
Antibiotic treatments do not affect endobacterial viability(P. Jargeat and G. Bécard, unpubl. results), whereasendobacteria were lost after four to six generations in rootorgan culture (ROC). Some ideas are suggested by thefollowing observations: (i) G. margarita has been main-tained in pot cultures for years using multispore inocula-tion (Bonfante et al., 1994), and under these conditionsthe endobacteria are always found, even if in variablenumbers (Bianciotto et al., 2004; Jargeat et al., 2004), (ii)
Fig. 9. Hyphal branching response of WT (B+) and cured (B–)Gigaspora margarita spores to GR24, a strigolactone analogue.GR24 (2 ¥ 5 ml 10-7 M) was added after 6 days of incubation(4 days after germination); the total number of hyphal ramificationswas counted after 15 days of incubation. Each bar corresponds tothe mean of 12 spores � SE. Different letters indicate significantlydifferent values according to the ANOVA test of variance (P < 0.01).The figure corresponds to one experiment, representative of all theexperiments performed.
Fig. 10. Hyphal growth from germinating spores of WT (�) andcured (�) Gigaspora margarita in the presence of transformedcarrot root fragments. The spores were located at 2 and 10 cmfrom the roots. The graph illustrates the experiment where thespores were located at 10 cm from the roots. WT myceliumreached the fragments in 26 days while the cured one was stoppedat 5 cm. Hyphal length is expressed as the mean of 20 spores �SE.
Endocellular bacteria affect AMF presymbiotic growth and morphology 1723
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Cellular Microbiology, 9, 1716–1729

bacteria are present in the spores produced in ROCsfollowing multispore inoculation, and (iii) spores withoutbacteria can easily be found after several single-spore-inoculated ROCs. Thus single-spore inoculation can beidentified as the condition affecting maintenance of theendobacterium. If we speculate that spores containing thehighest population of endobacteria are more infective(better fitness, see below) and therefore more competi-tive, they will be positively selected within a multisporeinoculum and will transmit large endobacterial populationsto the next generation. With single-spore inoculation,there is no selection, and even spores with low endobac-terial content may establish mycorrhizae, and producenew spores with a similar low bacterial content, increasingthe chance of losing bacteria along the generations.
The progressive decrease in ROC conditions (bacterialnumbers fall by one-fifth at each ROC generation accord-ing to Bianciotto et al., 2004) could also be the result of areduced capacity of the bacteria to multiply in this system.Identification of a bacterial cell division marker for Ca.Glomeribacter, i.e. the FtsZ gene belonging to the dcwcluster and differentially expressed during the fungal lifecycle (I. Anca et al., unpubl. results) should provide amolecular tool to test the hypothesis.
The endobacteria affect the spore phenotype
A detailed ultrastructural study of the B+ and B– spores ofG. margarita revealed that the B– spores had a thickercell wall. Not only did the bacteria disappear in curedspores, but the protein bodies also became less abundant(or less detectable with Coomassie blue). The smallelectron-dense granules, identified as melanin-likepigment granules (Grippiolo and Bonfante, 1984), areusually incorporated in the cell wall of WT spores but wererarely detectable in cured spores. The glycogen particlesalso seemed to be affected in the cured spores, but themost striking morphological difference involved lipids, thelipid bodies appearing smaller in cured spores. Sporemorphology thus shows that the presence of bacteriamodifies some metabolic aspects involving protein, lipidsand melanin; cell wall changes can be a consequence ofthe nutritional balance between the fungus and itsendobacteria.
All these important differences, likely reflecting signifi-cant metabolic modifications, suggest that the bacterialand fungal partners strongly and intimately interact at thelevel of their C metabolism. To address the questionwhether the bacterial presence represents a carbon costfor the fungus, we compared the multiple germinationcapacities of B– and B+ spores. Both types of sporesexhibited the same capacity as they could germinate suc-cessively up to 35 times (not shown data). Simple sugarsmust be sequestered by Ca. Glomeribacter but without
causing any serious quantitative carbon deprivation forthe fungal host. We hypothesize that both the partnersexploit common resources, such as sugars, in a coopera-tive manner for fuelling their own metabolism. Examplesof such functional integration are available from the well-known insect endosymbionts. Studies of endobacteriasuch as Buchnera and Wigglesworthia show that notwith-standing their reduced genome these bacteria provideimportant metabolites to their host, like essential aminoacids (Wernegreen, 2004).
To unambiguously confirm that the loss of bacteria isresponsible for these phenotypic changes, a complemen-tation experiment should be performed showing that rein-troduction of the bacteria into a cured line restores the WTphenotype. As Ca. Glomeribacter gigasporarum cannotbe grown in pure culture (Jargeat et al., 2004), such anexperiment is not yet feasible. In this context, our analy-ses of B+ and B– spores were made by using several linesof B– spores, coming from independent curing (motherspores) runs. Thus, when the different B– lines showsimilar phenotypic traits, we can be more confident thatthe differences are the result of the loss of bacteria, andnot the outcome of variability between spores.
Cured spores still keep their biotrophic capacities
The mycorrhization experiments using clover andsorghum showed that cured lines keep their biotrophiccapacities. This trait, already suggested by our first in vitroexperiments (Fig. 1, spore generations SG4, SG5 andSG6), allows us to conclude that production of curedspores is possible because they can still colonize rootsand complete their life cycle; without mycorrhization, anew generation of spores is not possible.
Symbiotic status depends on many other physiologicalfeatures; first the fungus has to perceive the plant in therhizosphere in order to successfully interact (Harrison,2005). Fungal branching after root exudate treatment isone phenotypic marker of this event (Giovannetti et al.,1996; Buée et al., 2000). Strigolactones have recentlybeen identified to stimulate branching (Akiyama et al.,2005) and to activate respiration and mitochondrial activi-ties in the fungus (Tamasloukht et al., 2003; Bessereret al., 2006). Interestingly, the growth rate of B+ and B–mycelium was comparable for the first 10 days, but afterthat the WT mycelium developed faster and was moreabundantly branched. The B– mycelium was also lessstimulated by the root exudates and the strigolactoneanalogue. The reduced lipid and protein reserves of thecured spores could offer a first explanation for this impor-tant physiological difference. The colonization experimentperformed on using transformed roots showed that bothcured and WT spores germinated and reached the rootsurface more or less at the same time (5 days), when
1724 E. Lumini et al.
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Cellular Microbiology, 9, 1716–1729

distances were short (not longer than 3 cm). At least inthis time and space frames, B– spores are vigorousenough to establish mycorrhizae. However, with sporesplaced 10 cm away from the root during long-time experi-ments (20 days), mycorrhization with B– spores wasseverely affected.
Taken as a whole, our results suggest that cured sporesare disadvantaged at a specific moment of their life cycle(the presymbiotic phase), from the sixth day aftergermination. The germinating mycelium is less fitted inreaching its host, and this may cause the positive selec-tion towards the B+ condition.
Conclusion
Symbiosis covers a wide range of situations, in which oneof the partners may be more or less dispensable. Forexample, plants, which naturally live in association withAM fungi, can grow without AM fungi under controlledconditions. The reverse is not true: AM fungi cannotaccomplish their life cycle without a plant host. In a similarway, AM fungi directly isolated from the field often containbacteria or bacteria–like organisms (BLOs) (Scanneriniand Bonfante, 1991), while AM fungi maintained under invitro or laboratory conditions often do not contain bacteria.Here also if the endobacteria are, to some extent (thisstudy), dispensable for the fungus, the reverse seems notto be true, at least if we recall the obligate biotrophicstatus of Ca. Glomeribacter gigasporarum (Jargeat et al.,2004). AM fungi are therefore dispensable for plants, andendobacteria for AM fungi. However, in ecological condi-tions natural selection has favoured mycorrhizal fungi asindispensable for plant success (Read, 1991). The samemay be true for endobacteria.
Experimental procedures
Fungal material
Spores of G. margarita Becker and Hall (BEG 34; deposited atthe European Bank of Glomeromycota) containing the Ca. Glom-eribacter gigasporarum endobacteria and produced in pot cultureby BIORIZE (Dijon, France) were used for all experiments. Theyare referred as WT or B+ (with bacteria) spores, while the curedspores are identified as B– (without bacteria). WT or cured sporepopulations are indicated as lines. A voucher of the cured sporesis deposited with the number E32 HC/F at Herbarium Crypto-gamicum Fungi, Department of Plant Biology, Torino, Italy.
Conditions for obtaining and cultivatingG. margarita-cured spore
Figure 1 summarizes the main steps of the procedure.
Production. A clone of root-inducing T-DNA-transformed carrotroots, established by Bécard and Fortin (1988), was propagated
on minimal (M) medium in 120 ¥ 120 mm square Petri dishes.For use as the plant partner in interaction with the fungus, rootexplants were standardized and prepared as described byBécard and Piché (1992). Spores of G. margarita were collectedfrom pot cultures (each pot culture representing a batch), surfacesterilized, stored at 4°C in sterile water and used as fungalinoculum (Bianciotto et al., 2004). A single starting spore (SG0)was used to inoculate the transformed root and to initiate myc-orrhizal cultures in Petri dishes. The dishes were carefully sealedwith Parafilm to confine the internal atmosphere and incubated inthe dark at 26°C. They were held vertically so that the spore germtubes elongated upward (as a result of negative geotropism) andcontacted the transformed root. In each Petri dish, new spores(SG1) were produced within 2 months as a result of mycorrhizalcolonization. These spores were used individually to inoculatenew root cultures in order to obtain SG2 spores. Similar succes-sive cycles were reproduced to obtain SG3, SG4, SG5 and SG6spores. A decreasing number of bacteria were found in spores ofeach new generation (Bianciotto et al., 2004). These successivesubcultures were therefore used to eventually produce curedspores. All the cured spores originated from an independentsingle SG0 spore were referred as belonging to the same lot.
Cultivation. In pot culture. To maintain and propagate the curedspores, leek, sorghum and clover plants were inoculated with amultisporal inoculum of cured spores obtained as describedabove.
Plants were grown in pots in order to become colonized and toproduce a new generation of cured spores within 3 months. Thesame protocol was used to maintain WT spores. The cycles ofspore production continued from 2002 to 2006.
In vitro culture. To maintain and propagate the cured spores,T-DNA-transformed carrot roots were propagated on minimalmedium in Petri dishes and inoculated with a multisporal inocu-lum of cured spores. The same protocol was used to maintain theWT spores.
Identification of batches and lots of cured spores. Six lots ofcured spores were originated by two batches (two starting pots):batch 1 (lot 1, 2, 3, 4) and batch 2 (lot 5 and 6). In particular thedifferent lots of cured spores, originating from independent invitro curing treatments, were submitted to different culture cycles.Cured spores of lot 1 were propagated through three cycles ofleek pot culture, lot 2 through one cycle of leek pot culture, lot 4through one cycle of clover pot culture, lot 5 through four cyclesof clover pot cultures, lot 6 through one cycle (of 2 years) ofsorghum pot culture, while spores of lot 3 were maintained underin vitro conditions throughout.
Checking the presence of bacteria
Microscopical detection of bacteria. After each mycorrhizalcycle, the presence or absence of bacteria was monitored inrandomly selected spores. These were placed on microscopeslides in 20 ml of Bacteria Counting Kit component A (B-7277;Molecular Probes) diluted 1:1.000 according to the manufactur-er’s directions. The spores were then crushed with a coverslip,incubated in the dark for at least 5 min, and observed with anOlympus FluoView confocal microscope. Bacteria wereobserved directly using a mercury arc fluorescence lamp, anexcitation band-pass (BP) filter at 490 nm (blue) and an emis-sion BP filter at 520 nm (green). For confocal imaging, the
Endocellular bacteria affect AMF presymbiotic growth and morphology 1725
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Cellular Microbiology, 9, 1716–1729

488 nm band of a Kr-Ar laser and a BP 510–540 nm emissionfilter were used.
Detection of Ca. Glomeribacter gigasporarum by conventionaland hemi-nested PCR amplification. For the molecular analyses,DNA extracts either from SG0 (B+) or from SG6 (B–) spores(single or pooled in 10) were used in conventional and hemi-nested PCR assays. Miniprep DNA extraction was performed bycrushing the spores in a volume of 30 ml (single spore) or 50 ml(pool of 10 spores) of 1:1 H2O:10X RedTaq PCR Buffer (100 mMTris-HCl, pH 8.3, 500 mM KCl, 11 mM MgCl2 and 0.1% gelatine).After incubation at 95°C for 15 min, the crude extract was cen-trifuged at 10 000 g for 5 min, and the supernatant was used for10-fold serial dilutions. Extreme care was taken to avoid externaland cross-bacterial contamination, and all steps were carried outunder a laminar flow hood. For the conventional PCR, DNAextracted from B+ and B– spores was amplified by specific bac-terial primers BLOf-BLOr and GlomGIGf-GlomGIGr. PrimersNS31-AM1 (Helgason et al., 1998) that amplify a portion of thefungal 18S ribosomal gene were used as positive controls toassess the quality of the DNA extraction and the absence ofinhibition of the PCR assays. Conventional PCR was carried outas described in Bianciotto et al. (2004).
To confirm the results obtained by conventional PCR we per-formed a hemi-nested PCR assay.
For such reaction, 1 ml of the conventional PCR productsobtained with GlomGIGf-GlomGIGr on DNA extracted from B+and B– was added as DNA template to 24 ml of PCR mix. Thereaction mixture contained 10 mM Tris-HCl, 50 mM KCl, 1.5 mMMgCl2, 0.1% gelatine, 0.2 mM of each dNTP, 0.5 mM primersGlomGIGf and GIGrA(5′-GTTGTTGCCCTCTTGACACC-3′), 1 Uof Taq polymerase. Amplifications were carried out in 0.2 ml PCRtubes using a GeneAmp PCR System 9700 thermocycler. Theprogramme consisted of 25 cycles of 45 s at 94°C, 1 min at 60°Cand 45 s at 72°C and a final extension of 7 min at 72°C. Theresulting 106 bp PCR products were visualized in a 2.5% high-resolution agarose gel using SYBR Safe™ DNA gel stain (Invit-rogen) for visualization under UV light.
Estimation of genetic diversity among WT and curedspores by RAPD and ISSR amplification
PCR and electrophoretic conditions. To estimate and comparelevels of genetic diversity among B+ and B– G. margarita spores,finger printing experiments were carried out. Forty B+ spores and36 B– spores (belonging to lots 5 and 6) were analysed by RAPDand ISSR amplification. These two lots were also used for growthtest (see Growth capacities of WT versus cured germinatingspores).
DNA was extracted from individual spores by using theWIZARD Genomic DNA Purification kit (Promega – Lyon,France). The final pellet was re-suspended in 20 ml of sterilewater. To avoid any contamination, DNA extraction and PCRpreparation were always performed under sterile conditions.
For RAPD analysis, primers 152 (Sélosse et al., 1998), cTB6and cTB9 (http://plantbio.berkeley.edu/~bruns/primers.html)were used. Reactions were carried out in a 25 ml volume con-taining 5 ml of 5¥ GoTaq buffer (Promega – Lyon, France), 1 mMof primer, 250 mM of each dNTP, 1 unit of GoTaq Polymerase(Promega – Lyon, France) and 1 ml of DNA solution. PCR cyclingconditions of the Mastercycler thermocycler (Eppendorf) were asfollows: initial denaturation at 94°C for 3 min; 40 cycles at 94°C
for 1 min, 40°C for 1 min, 72°C for 2 min and a final extension at72°C for 10 min.
For ISSR analysis, reactions were carried out using primersR1 (DHB CGA CGA CGA CGA CGA; Hantula et al., 1996), R2(DDB CCA CCA CCA CCA CCA; Hantula et al., 1996) and R3(BDB ACA ACA ACA ACA ACA; Hantula et al., 1996) asdescribed for RAPD procedure except amplifications were per-formed with an initial denaturation of 3 min at 94°C, followed by37 repetitions of 45 s at 94°C, 45 s at 58°C (R1 and R2) or 50°C(R3) and 3 min at 72°C, and ended by a final extension of 10 minat 72°C.
Amplification products were separated by electrophoresis in1¥ TAE buffer using a 1.5% agarose gel. Gels were ethidiumbromide stained and numerical pictures of banding patterns werecaptured under UV light with a CDD camera supported byVISION-CAPT software (Vilber Lourmat, France) for laterscoring.
Reproducibility of the patterns given by each primer waschecked by comparing results of two independent PCR carriedout with 10 subsamples. Once reproducibility was assessed foreach primer, the 76 samples were amplified in one run of PCR. Anegative control without template DNA was included in each runto test the absence of DNA contamination.
Scoring of RAPD and ISSR fragments and data analysis. Onlyunambiguously clear fragments that were present in at least twosamples were used in the analysis of RAPD and ISSR patterns.Each fragment was scored as present (1) or absent (0) in a binarymatrix.
Genetic diversity within each spore populations (B+ and B–)was estimated by calculating the percentage of polymorphicbands (p.p.b.) and the Shannon-Weaver diversity index (H)(Shannon and Weaver, 1949). This index is based on numberand frequency of markers in fingerprints and calculated as:H = –Spi·log2pi, where pi is the frequency of the ith band. H wasalso calculated at two levels: the diversity among all spores (Hsp)and the average diversity within populations (Hpop). Then, theproportion of genetic variation between B+ and B– spore popu-lations could be estimated by calculating (Hsp - Hpop)/Hsp.
In order to estimate the genetic divergence between B+ andB– populations by another way, an AMOVA analysis (Excoffieret al., 1992) was made using ARLEQUIN 2.000 software(Schneider et al., 2000). AMOVA allowed (i) to apportion the totalmolecular variance in among- and in within-population compo-nents and (ii) to test the significance of the partitioning.
Pairwise genetic distances were estimated as Euclidean dis-tance (Excoffier et al., 1992). PCA based on genetic distanceswas performed using GENALEX software (Peakall and Smouse,2002) to provide a graphical representation of the genetic rela-tionships between all the spores.
Evaluation of mycorrhizal capacities of the cured spores
To analyse the mycorrhizal phenotype of the cured spores, dif-ferent host plants were used: a dicot, Trifolium repens grown ingrowth chamber (16 clover plants mycorrhized with curedspores and four plants mycorrhized with WT spores), amonocot, Sorghum bicolor grown in greenhouse (24 sorghumplants mycorrhized with the cured spores and six plants inocu-lated with WT spores) and transformed carrot roots grown invitro. In the sorghum experiment, the cured spores we used
1726 E. Lumini et al.
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Cellular Microbiology, 9, 1716–1729

belong to the lots 1, 2, 3 and 4. In the clover and in the trans-formed carrot roots experiments, an additional lot (lot 5) ofcured spores was used.
Clover and sorghum plants were inoculated with 40–60 sporesand grown under greenhouse conditions for 12 weeks. The entireroot was then stained overnight at room temperature in 0.1%Cotton blue (w:v) in lactic acid, and washed several times in lacticacid. Root segments (1 cm long) were randomly selected andobserved under an optical microscope (Nikon Eclipse E400) forscoring mycorrhizal colonization. Four mycorrhizal variableswere considered as described by Trouvelot et al. (1986). Rootsegments were classified according to the percentage of theirlength occupied by mycelium and/or arbuscules. In total, 72 mand 18 m of sorghum roots inoculated, respectively, with curedand WT spores, 157 cm and 100 cm of clover roots inoculated,respectively, with cured and WT spores, and 90 cm and 90 cm oftransformed carrot roots inoculated, respectively, with cured andWT spores were carefully examined. In order to quantify thesporulation capacity, the number of new spores generated duringthe sorghum and transformed carrot roots experiments was alsocounted.
Morphological characterization of the cured spores
Light microscopy on fresh material. To check the morphologyand wall organization of spores, 69 B+ spores and 144 B– sporescoming from pot cultures inoculated with the five lots (1, 2, 3, 4and 5) described in Conditions for obtaining and cultivatingG. margarita-cured spore were collected, sterilized with 3% (w:v)chloramine T, 0.03% (w:v) streptomycin and mounted in a solu-tion of Polyvinyl-Lacto-Glycerol (PVLG) prepared with 100 ml ofdistilled water, 100 ml of lactic acid and 16.6 g of Polyvinylalcohol (PVA).
Light and transmission electron microscopy on embeddedmaterial. Twenty WT and 20 cured spores (15 of them originatingfrom pot cultures inoculated with the five lots previouslydescribed and five of them directly picked from a multisporal invitro culture) (Fig. 1, Production step) were cryo- or chemicallyfixed and then resin-embedded. For cryo-fixation, spores werevacuum-infiltrated in 12% Dextran water solution; each sporewas then placed in an aluminium holder and frozen at highpressure using the Balzers HPM 010 apparatus. Frozen sampleswere stored in liquid nitrogen until further processing. Freezesubstitution (FS) was performed in two FS solutions; the sporeswere incubated in 1% glutaraldehyde, 1% tannic acid in dryacetone at -85°C for 48–72 h, rinsed in dry, cold acetone threetimes, then transferred to 1% osmium tetroxide in acetone at-85°C for 1 h. To warm it up slowly, the osmium solution was heldfor 2 h at -20°C, 2 h at 4°C and 1 h at room temperature. Afterthree rinses in fresh acetone, samples were infiltrated with anEpon/Araldite-acetone mixture using 10% increase of resin eachstep for 1 h and then polymerized at 60°C for 10 h.
For chemical fixation, spores were perforated with a sterileneedle and fixed under vacuum with 2.5% glutaraldehyde in50 mM sodium cacodylate buffer pH 7.2 for 2 h at room tempera-ture and overnight at 4°C. Samples were then post-fixed in 1%osmium tetroxide in cacodylate buffer for 2 h, dehydrated in anacetone series and embedded in Epon/Araldite.
Thin sections (70 nm thick) were stained with uranyl acetateand lead citrate and examined with a Philips CM10 transmission
electron microscope, while semi-thin sections (1 mm thick) werestained for general observation with 1% toluidine blue in 1%aqueous sodium tetraborate and observed with a Nikon EclipseE400 optical microscope. Some sections were stained for 10 minat 50°C in 0.25% Coomassie brilliant blue in 7% acetic acid(O’Brien and McCully, 1981) to visualize proteins. Others weretreated with 70% ethanol for 2 min, stained with 0.3% SudanBlack B in 70% ethanol for 1 h at 60°C and rinsed with 70%ethanol for 1 min, for lipid detection. A third group of sections wastreated with 1% periodic acid for 10 min, rinsed briefly with dis-tilled water, stained with Schiff’s reagent for 30 min and rinsedtwice with distilled water to evaluate the presence ofpolysaccharides.
Growth capacities of WT versus cured germinatingspores
Spores were germinated on solid medium (0.1% w:v MgSO4;0.4% w:v Phytagel) in vertically incubated Petri dishes (90 mmdiameter or 120 ¥ 120 mm square plates), in a 2% CO2 incubatorat 28°C. Eight to 10 Petri dishes with one spore per dish wereused per treatment. The experiments were performed by usingcured spores belonging to four lots (lots 1, 3, 5 and 6 as previ-ously described).
Wild-type and cured germinating spores were treated 4 daysafter germination with an active fraction (branching factor) ofcarrot root exudates prepared as described by Buée et al. (2000).The branching factor was extracted with ethyl acetate from carrotroot exudate in water, dried and re-suspended in (50:50) metha-nol:water (Buée et al., 2000). Ten microlitres (2 ¥ 5 microlitres) ofthis solution, or of methanol:water for control, were injected nearthe growing germ tubes. Hyphal length from germinating sporeswas measured under a binocular microscope by using a 2 mmgrid every 2–3 days from day 4 to 30.
Under the same conditions, WT and cured germinating sporeswere also treated with the strigolactone analogue GR 24 at10-7 M. After 15 days of growth, the number of hyphal branchesformed by each germinating spore was recorded. Differenceswere tested for significance by analysis of variance and Tukey’spairwise comparison.
To further check whether the growth capacities of WT andcured germinating spores were affected by the presence of thehost plant, an additional small scale in vitro experiment was setup to follow the timing of infection. Carrot roots, as describedinProduction, were inoculated with B+ and B– spores and infec-tion was followed daily. In each plate, four segments of carrotroots were placed and two spores were put under each segment.The distance between spore and root was 1.5 cm, 3 cm and10 cm, for each experiment. Five plates with cured spores andfive plates with WT spores were prepared.
Acknowledgements
The authors thanks Bachar Blal (BIORIZE, Dijon, France) for themycorrhization experiments on Sorghum, Sara Torrielli for sporecollection, Santina Buonomo and Maria Teresa Della Beffa formaintaining ROC cultures and mycorrhizal plants respectively.The authors are particularly indebted to Dr Robert W. Roberson(University of Arizona, USA) for the freeze substitution experi-ments performed in his laboratory and to Dr Hervé Gryta (Tou-
Endocellular bacteria affect AMF presymbiotic growth and morphology 1727
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Cellular Microbiology, 9, 1716–1729

louse 3 University) for genetic variability analyses. The facilitiesof the Laboratory of Advanced Microscopy (University of Torino)are acknowledged, in particular Andrea Genre for confocal obser-vations and Silvano Panero for assistance with transmissionelectron microscopy. This research was funded by the EuropeanUnion GENOMYCA project (Project No. QLK5-CT-2000-01319),CEBIOVEM programme (D.M. 193/2003) and IPP-CNR (Biodi-versity Project AGP02 n. 371–Torino). E.L. was founded byLagrange Project – CRT Foundation with a Post-Doc LagrangeFellow 2004.
References
Akiyama, K., Matsuzaki, K., and Hayashi, H. (2005) Plantsesquiterpenes induce hyphal branching in arbuscularmycorrhizal fungi. Nature 435: 824–827.
Bécard, G., and Fortin, J.A. (1988) Early events of vesicular-arbuscular mycorrhiza formation on Ri T-DNA transformedroots. New Phytol 108: 211–218.
Bécard, G., and Piché, Y. (1992) Establishment of VA myc-orrhizae in root organ culture: review and proposedmethodology. In Methods in Microbiology, Vol. 24. Norris,J.R., Read, D.J., and Varma, A.K. (eds). London: AcademicPress, pp. 89–108.
Besserer, A., Puech-Pagès, V., Kiefer, P., Gomez-Roldan,V., Jauneau, A., Roy, S., et al. (2006) Strigolactonesstimulate arbuscular mycorrhizal fungi by activatingmitochondria. PLoS Biol 4: 1239–1247.
Bianciotto, V., Bandi, C., Minerdi, D., Sironi, M., Tichy, H.V.,and Bonfante, P. (1996) An obligately endosymbioticfungus itself harbors obligately intracellular bacteria. ApplEnviron Microbiol 62: 3005–3010.
Bianciotto, V., Lumini, E., Lanfranco, L., Minerdi, D., Bon-fante, P., and Perotto, S. (2000) Detection and identifica-tion of bacterial endosymbionts in arbuscular mycorrhizalfungi belonging to Gigasporaceae. App Environ Microbiol66: 4503–4509.
Bianciotto, V., Lumini, E., Bonfante, P., and Vandamme, P.(2003) ‘Candidatus Glomeribacter gigasporarum’ gen.nov., sp nov., an endosymbiont of arbuscular mycorrhizalfungi. Int J Syst Evol Microb 53: 121–124.
Bianciotto, V., Genre, A., Jargeat, P., Lumini, E., Bécard, G.,and Bonfante, P. (2004) Vertical transmission of endobac-teria in the arbuscular mycorrhizal fungus Gigasporamargarita through generation of vegetative spores. ApplEnviron Microbiol 70: 3600–3608.
Bonfante, P. (2003) Plants, mycorrhizal fungi and endobac-teria: a dialog among cells and genomes. Biol Bull 204:215–220.
Bonfante, P., and Vian, B. (1984) Wall texture in the spore ofa vesicular-arbuscular mycorrhizal fungus. Protoplasma120: 51–60.
Bonfante, P., Balestrini, R., and Mendgen, K. (1994)Storage and secretion processes in the spore ofGigasporamargarita Becker & Hall as revealed by high-pressure freezing and freeze substitution. New Phytol 128:93–101.
Buée, M., Rossignol, M., Jauneau, A., Ranjeva, R., andBécard, G. (2000) The pre-symbiotic growth of arbuscularmycorrhizal fungi is induced by a branching factor partially
purified from plant root exudates. Mol Plant Microbe Inter-act 13: 693–698.
Claes, P.M., and Dunlap, F.V. (2000) Aposymbiotic culture ofthe sepiolid squid Euprymna scolopes: role of the symbioticbacterium Vibrio fischeri in hostanimal growth, develop-ment, and light organ morphogenesis. J Exp Zool 286:280–296.
Excoffier, L., Smouse, P.E., and Quattro, J.M. (1992) Analy-sis of molecular variance inferred from metric distancesamong DNA haplotypes: application to human mitochon-drial DNA restriction data. Genetics 131: 479–491.
Giovannetti, M., Sbrana, C., Silvia, A., and Avio, L. (1996)Analysis of factors involved in fungal recognition responseto host-derived signals by arbuscular mycorrhizal fungi.New Phytol 133: 65–71.
Grandmougin-Ferjani, A., Fontaine, J., and Durand, R.(2005) Carbon metabolism, lipid composition and metabo-lism in arbuscular mycorrhizal fungi. In In Vitro Culture ofMycorrhizas. Declerck, S., Strullu, D.G., and Fortin, J.(eds). Berlin: Springer-Verlag, pp. 159–176.
Grippiolo, R., and Bonfante, P. (1984) Sporopollenin andmelanin-like pigments in the wall of a Glomus spore. GiornBot Ital 118: 88–90.
Hantula, J., Dusabenygasani, M., and Hamelin, R.C. (1996)Random amplified microsatellites (RAMS) – a novelmethod for characterizing genetic variation within fungi.Eur J Forest Pathol 26: 159–166.
Harrison, M.J. (2005) Signaling in the arbuscular mycorrhizalsymbiosis. Annu Rev Microbiol 59: 19–42.
Helgason, T., Daniell, T.J., Husband, R., Fitter, A.H., andYoung, J.P.W. (1998) Ploughing up the wood-wide web?Nature 394: 431.
Hijri, M., and Sanders, I.R. (2005) Low gene copy numbershows that arbuscular mycorrhizal fungi inherit geneticallydifferent nuclei. Nature 433: 160–163.
Jargeat, P., Cosseau, C., Ola’h, B., Jauneau, A., Bonfante,P., Batut, J., and Bécard, G. (2004) Isolation, free-livingcapacities, and genome structure of ‘Candidatus Glomeri-bacter gigasporarum’, the endocellular bacterium of themycorrhizal fungus Gigaspora margarita. J Bacteriol 186:6876–6884.
Lumini, E., Ghignone, S., Bianciotto, V., and Bonfante, P.(2006) Endobacteria or bacterial endosymbionts? To be ornot to be. New Phytol 170: 205–208.
Moran, N.A. (2001) Bacterial menageries inside insects. ProcNatl Acad Sci USA 98: 1338–1340.
Moran, N.A. (2003) Tracing the evolution of gene loss inobligate bacterial symbionts. Curr Opin Microbiol 6: 512–518.
O’Brien, T.P., and McCully, M.E. (1981) The Study of PlantStructure Principle and Selected Methods. Melbourne: Ter-macarphi Pty.
Partida-Martinez, L.P., and Hertweck, C. (2005) Pathogenicfungus harbours endosymbiotic bacteria for toxinproduction. Nature 437: 884–888.
Pawlowska, T.E., and Taylor, J.W. (2004) Organization ofgenetic variation in individuals of arbuscularmycorrhizalfungi. Nature 427: 733–737.
Peakall, R., and Smouse, P.E. (2002) GENALEXV5.04:Genetic Analysis in Excel. Population Genetic Software for
1728 E. Lumini et al.
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Cellular Microbiology, 9, 1716–1729

Teaching and Research. Canberra: Australian NationalUniversity. [WWW document]. URL http://www.anu.edu.au/BoZo/GenAlEx/.
Read, D.J. (1991) Mycorrhizas in ecosystems. Experientia47: 376–390.
Scannerini, S., and Bonfante, P. (1991) Bacteria and bac-teria like objects in endomycorrhizal fungi (Glomaceae).In Symbiosis as a Source of Evolutionary Innovation:Speciation and Morphogenesis. Margulis, L., andFester, R. (eds). Cambridge: MIT Press, pp. 273–287.
Schneider, S., Roessli, D., and Excoffier, L. (2000) ARLE-QUIN Version 2.000: a Software for Population GeneticsData Analysis. Geneva: Genetics and Biometry Laboratory,University of Geneva. [WWW document]. URL http://lgb.unige.ch/arlequin/.
Schüßler, A. (2002) Molecular phylogeny, taxonomy, andevolution of Geosiphon pyriformis and arbuscular mycor-rhizal fungi. Plant Soil 244: 75–83.
Sélosse, M.A., Jacquot, D., Bouchard, D., Martin, F., and LeTacon, F. (1998) Temporal persistence and spatial distri-bution of an American inoculant strain of the ectomycor-rhizal basidiomycete Laccaria bicolor in a French forestplantation. Mol Ecol 7: 561–573.
Shannon, C.E., and Weaver, W. (1949) The MathematicalTheory of Communication. Urbana, IL: University of IllinoisPress.
Smith, S.E., and Read, D.J. (1997) Mycorrhizal Symbiosis,2nd edn. San Diego, CA: Academic Press.
Sward, R.J. (1981) The structure of the spores of Gigasporamargarita. I. The dormant spore. New Phytol 87: 761–768.
Tamasloukht, M.B., Sejalon-Delmas, N., Kluever, A.,Jauneau, A., Roux, C., Bècard, G., and Franken, P. (2003)Root factors induce mitochondrial-related gene expressionand fungal respiration during the developmental switchfrom asymbiosis to presymbiosis in the arbuscular mycor-rhizal fungus Gigaspora rosea. Plant Physiol 131: 1468–1478.
Trouvelot, A., Kough, J.L., and Gianinazzi-Pearson, V. (1986)Mesure du taux de mycorhization VA d’un systèmeradiculaire. Recherche de méthodes d’estimation ayantune signification functionnelle. In Les Mycorhizes: Physi-ologie et Génétique. Gianinazzi-Pearson, V. and Giani-nazzi, S. (eds). Paris: INRA Press, pp. 217–221.
Van der Heijden, M.G.A., and Sanders, J.R. (2002) Mycor-rhizal Ecology. Berlin: Springer-Verlag.
Wernegreen, J.J. (2004) Endosymbiosis: lessons in conflictresolution. PLoS Biol 2: 307–311.
Endocellular bacteria affect AMF presymbiotic growth and morphology 1729
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Cellular Microbiology, 9, 1716–1729
Related Documents



![Diet of Arbuscular Mycorrhizal Fungi: Bread and Butter?...Gigaspora rosea), while FA elongation and desaturation can occur independently of the plant [13]. ... in plastids, these findings](https://static.cupdf.com/doc/110x72/5f7f050c12485b013f6da226/diet-of-arbuscular-mycorrhizal-fungi-bread-and-butter-gigaspora-rosea-while.jpg)







