Theriogenology41:1333-1346,1994 PRESERVATION OF OOCYTE AND GRANULOSA CELL MORPHOLOGY IN BOVINE PREANTRAL FOLLICLES CULTURED IN VITRO J.R. Figueiredo,Iya S.C.J. Hulshof,3 R. Van den Hurk,3 B. Nusgens,2 M.M. BeversP F.J. Ectorsl and J.F. Beckers’ 1 Department of Animal Endocrinology and Reproduction Faculty of Vetermary Medicine 2 Laboratory of Experimental Dermatology University of Liege, B-4000 Liege, Belgium 3 Department of Functional Morphology 4 Department of Herd Health and Reproduction Faculty of Veterinary Medicine, University of Utrecht Yalelaan 8,350s TD, Utrecht, The Netherlands Received for publication: &ne 30, 1993 Accepted:February 12, 1994 ABSTRACT Described in the present paper is a culture system that preserves oocyte and granulosa cell morphology in bovine preantral follicles during 5 d in vitro. The effects of additional hypoxantbine and energy substrata (i.e., pyruvate and glutamine) on the morphology of cultured preantml follicles were investigated. It was shown that addition of a mixture of pyruvate, glutamine and hypoxantine to the culture medium increased the percentage of follicles with an intact oocyte from 29.4 to 78.6%. Morphological criteria are described to discriminate between normal and degenerated preantral follicles during culture by inverted microscopy. In addition, the importance of histological evaluation to judge the quality of oocyte and granulosa cells is demonstrated. Key words: culture, preantral follicles, substrate, bovine Acknowledgments This work was supported by an IRSIA grant from the Faculty of Veterinary Medicine, The State University of Liege; by FRSM; and by les Actions de Recherches Concert&s de la Communaute Francaise de Belgique. We thank G. Dijkstra for histology and H.H. Otter, 0. van der Veen and H. Post for assistance in photography. aJ.R. Figueiredo is a recipient of a grant from CNPq-BIOAGRO (UFV) of Brazil. Copyright 0 1994 Butterwotth-Heinemann

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Theriogenology41:1333-1346,1994
PRESERVATION OF OOCYTE AND GRANULOSA CELL MORPHOLOGY IN BOVINE PREANTRAL FOLLICLES CULTURED IN VITRO
J.R. Figueiredo,Iya S.C.J. Hulshof,3 R. Van den Hurk,3 B. Nusgens,2 M.M. BeversP F.J. Ectorsl and J.F. Beckers’
1 Department of Animal Endocrinology and Reproduction Faculty of Vetermary Medicine
2 Laboratory of Experimental Dermatology University of Liege, B-4000 Liege, Belgium
3 Department of Functional Morphology 4 Department of Herd Health and Reproduction
Faculty of Veterinary Medicine, University of Utrecht Yalelaan 8,350s TD, Utrecht, The Netherlands
Received for publication: &ne 30, 1993
Accepted: February 12, 1994
ABSTRACT
Described in the present paper is a culture system that preserves oocyte and granulosa cell morphology in bovine preantral follicles during 5 d in vitro. The effects of additional hypoxantbine and energy substrata (i.e., pyruvate and glutamine) on the morphology of cultured preantml follicles were investigated. It was shown that addition of a mixture of pyruvate, glutamine and hypoxantine to the culture medium increased the percentage of follicles with an intact oocyte from 29.4 to 78.6%.
Morphological criteria are described to discriminate between normal and degenerated preantral follicles during culture by inverted microscopy. In addition, the importance of histological evaluation to judge the quality of oocyte and granulosa cells is demonstrated.
Key words: culture, preantral follicles, substrate, bovine
Acknowledgments
This work was supported by an IRSIA grant from the Faculty of Veterinary Medicine,
The State University of Liege; by FRSM; and by les Actions de Recherches Concert&s de
la Communaute Francaise de Belgique. We thank G. Dijkstra for histology and H.H. Otter, 0. van der Veen and H. Post for
assistance in photography.
aJ.R. Figueiredo is a recipient of a grant from CNPq-BIOAGRO (UFV) of Brazil.
Copyright 0 1994 Butterwotth-Heinemann

1334 Theriogenology
INTRODUCTION
The ability to culture ovarian follicles from the primary stage to the preovulatory stage,
followed by fertilization of their oocytes would have important applications in animal breeding
and human in vitro fertilization programs (4, 40). Exploitation of this resource, however,
requires the establishment of protocols for their growth and development that would promote
and maintain competence for fertilization and embryogenesis.
In mice (8, 9, 17), pigs (27) rats (12) and cats (30) isolated preantral follicles were
cultured and growth of the follicles was observed. Antral follicle development from preantral
follicles in culture has been reported in mice (37,40). Complete development of murine primary
follicles to mature stages was obtained and development to term achieved when oocytes derived
from in vitro-grown follicles were fertilized in vitro (20). Several culture media were used in the
different experiments (8, 12, 17,34,35,40,45). Pyruvate (8, 12, 17, 20, 45) and glutamine
(8, 37) have been used as energy substrate for the culture of ovarian preantral follicles in rats
and mice. Hypoxantbine has been added to the culture medium to prevent meiotic resumption
of the oocyte in several species (15, 19, 47). In addition, Eppig et al. (19) showed that
hypoxanthine reduces the percentage of denuded oocytes in mu&e preantral follicles cultured in
vitro.
Maintaining oocyte viability in preantral follicles has been shown to be difficult. Maresh et
al. (35) reported that in rabbit preantral follicles all oocytes were degenerated after a 15-d culture
period. Hirao et al. (27) reported that in pig pmantral follicles the survival rate of oocytes
cultured in medium with or without 10 mg/ml FSH was 36 and 15%, respectively. In the cow,
Jewgenow et al. (29) showed that after 48 h of culture, most of the oocytes in preantral follicles
were degenerated. Recently, Nuttinck et al. (38) found that 85% of the preantral bovine follicles
remained viable during a 7-d culture period on irradiated fibroblasts. It is, however, unclear
whether oocytes were present in those follicles.
The lack of efficient methods for the isolation of preantral follicles from the ovaries as
well as of defined culture systems has restricted studies in follicular viability and growth.
Recently, we reported on mechanical and enzymatic methods for the dissociation of bovine
ovaries to yield large numbers of intact, small preantral follicles (22). The present paper deals
with the culture of isolated bovine preantral follicles over a 5-d period, with emphasis on the
(micro)morphological condition of granulosa cells and oocytes, and on the effects of pyruvate,
glutamine and hypoxanthine.

Theriogenology 1335
MATERIALS AND METHODS
Isolation of Preantral Follicle
Ovaries from adult cows were collected at a local slaughterhouse. The ovaries were
aseptically removed, stripped of surrounding fat tissue and ligaments, and transferred into 10 ml
PBS supplemented with 200 IU/ml of penicillin and 200 pg/ml of streptomycin. Following 3
successive washings in this solution, each ovary was transferred into 10 ml of Ml99 HEPES
containing 200 IU/ml of penicillin, 200 @ml of streptomycin, and 5% fetal calf serum (KS,
Boehringer Mannheim, Germany) and transported to the laboratory within 1 h in a thermos
flask tilled with water at 39°C.
Preantral follicles were isolated from the ovaries by applying the simple mechanical
procedure described by Figueiredo et al. (22) without the use of enzymes. Briefly, the ovaries
were cut into small fragments using a tissue chopper. The ovarian fragments were put in Ml99
HEPES supplemented with 5% FCS, antibiotics (200 III/ml of penicillin, 200 pglml of
streptomycin), and 0.23mM pyruvate and then suspended 40 times with a pasteur pipette and filtered successively through 500- and lOO-ym nylon mesh filters. Isolated preantral follicles
(30 to 70 ym in diameter) were selected using a mouth-operated micropipette. Groups of 3
follicles were placed in 5@-pl droplets of isolation medium to facilitate the onset of culture. The
total procedure (isolation, selection and manipulation of follicles before culture) took approximately 3 h.
General culture procedure
Isolated preantral follicles were placed in 350 pl of culture medium in groups of 3 follicles
per well (plastic petri dishes, Nunc) and cultured for 5 d at 39’ C in 5% CO, in air. Culture in
groups of 3 follicles per well and their localization after attachment at Day 1 of culture allows
each follicle to be studied individually during the culture period.
The control medium was alpha Minimal Essential Medium (MEM; Gibco-BRL, Geithersburg, MD, USA) supplemented with antibiotics (200 III/ml of penicillin, 200 pglml of
streptomycin, 10% FCS and ITS (Insulin 6.25 pglml, transferin 6.25 pg/ml and selenium 6.25
nglml). Fresh medium was prepared immediately before use in culture.
Culture Experiments
A total of 341 follicles was cultured for 5 d in 8 different culture media. The media were compared using preantral follicles from the same ovaries. Medium was changed 24 h after the
start of culture, and then every other day.

1336 Theriogenology
The media were:
Glu: Hyp: PyrGlu:
PyrHyP GluHyp: PyrGluHyp:
Control medium Control medium plus 0.23 mM pyruvate (Sigma); Control medium plus 2 mM glutamine (Sigma); Control medium plus 2 mM hypoxanthine (Sigma); Control medium plus 0.23 mM pyruvate and 2 mM glutamine; Control medium plus 0.23 mM pyruvate and 2 mM hypoxanthine; Control medium plus 2 mM glutamine and 2 mM hypoxanthine; Control medium plus 0.23 mM pyruvate, 2 mM glutamine and 2 mM hypoxanthine.
Fohicular Morphology and Histological Evaluation
The morphology of each follicle during culture was observed using an inverted microscope. Follicular morphology, particularly the clarity of the oocyte and surrounding granulosa cell layer, was recorded at Days 1 and 5 of culture. Simultaneously, follicular diameters were measured with an ocular micrometer.
After 5 d of culture, the follicles were fmed in Bouins fluid for 24 h, dehydrated in a graded series of ethanol, and embedded in paraffin wax. Then 5ym tissue sections were stained with PAS haematoxylin. Freshly isolated preantral follicles were also fixed and sectioned to compare the in vivo appearance of preantral follicles with the in vitro picture.
The number of cells per mm2 in the granulosa layer was measured in 20 sections of uncultured preantral follicles, and of follicles cultured for 5 d in medium with or without the addition of hypoxanthine. In the same 20 sections, the area of 50 granulosa cell nuclei was
measured. The number of cells per mm2 in the granulosa layer and the area of the nuclei were determined by means of image analysis using the TIM image processing program (Version 3.20, Difa Measuring systems, Breda, The Netherlands) and a black and white CCD video camera (MO- Type High Technology Holland, Eindhoven, The Netherlands).
Statistical Analysis of Data
The percentages of morphologically normal follicles after 24 h (Day 1) of culture and after an additional 4 d of culture (Day 5) were analyzed by Chi-square. Since follicles were monitored individually during culture, the increase in diameter of morphologically normal follicles from Day 1 to Day 5 was analyzed by t-test (analysis of variance) and were considered paired data. The number of cells per mm2 in the granulosa cell layer and the area of the granulosa cell nuclei were analyzed by the nonparametric Mann Whitney U test. Values were considered statistically significant when PcO.05.
Theriogenology
RESULTS
1337
Selection of Bovine Preatmal Follicles Before Culture
After the isolation procedure, the remaining suspension was studied under an inverted
microscope and appeared to contain numerous morphologically normal follicles, dark follicles
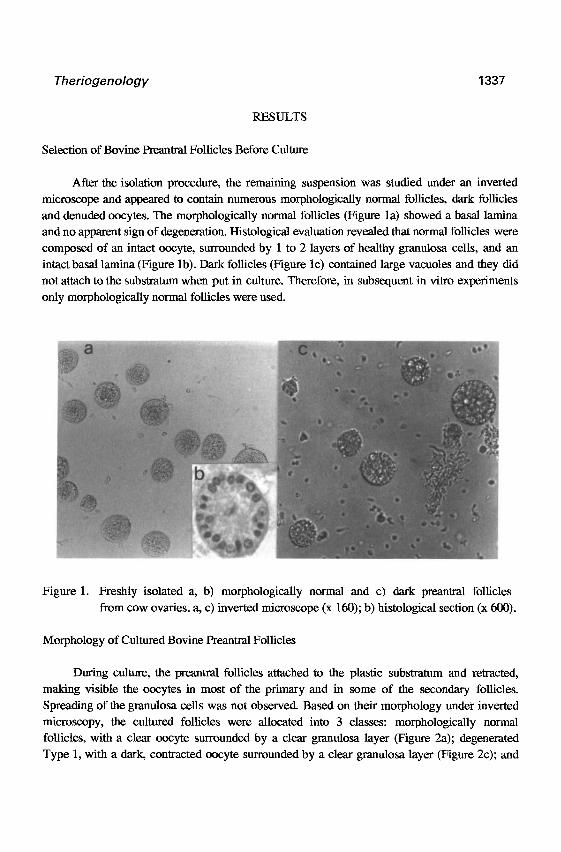
and denuded oocytes. The morphologically normal follicles (Figure la) showed a basal lamina
and no apparent sign of degeneration. Histological evaluation revealed that normal follicles were
composed of an intact oocyte, surrounded by 1 to 2 layers of healthy granulosa cells, and an
intact basal lamina (Figure lb). Dark follicles (Figure lc) contained large vacuoles and they did
not attach to the substratum when put in culture. Therefore, in subsequent in vitro experiments
only morphologically normal follicles were used.
Figure 1. Freshly isolated a, b) morphologically normal and c) dark preantral follicles
from cow ovaries. a, c) inverted microscope (x 160); b) histological section (x 600).
Morphology of Cultured Bovine Freantral Follicles
During culture, the preantral follicles attached to the plastic substratum and retracted,
making visible the oocytes in most of the primary and in some of the secondary follicles.
Spreading of the gmnulosa cells was not observed. Based on their morphology under inverted
microscopy, the cultured follicles were allocated into 3 classes: morphologically normal follicles, with a clear oocyte surrounded by a clear granulosa layer (Figure 2a); degenerated
Type 1, with a dark, contracted oocyte surrounded by a clear granulosa layer (Figure 2~); and
Theriogenology
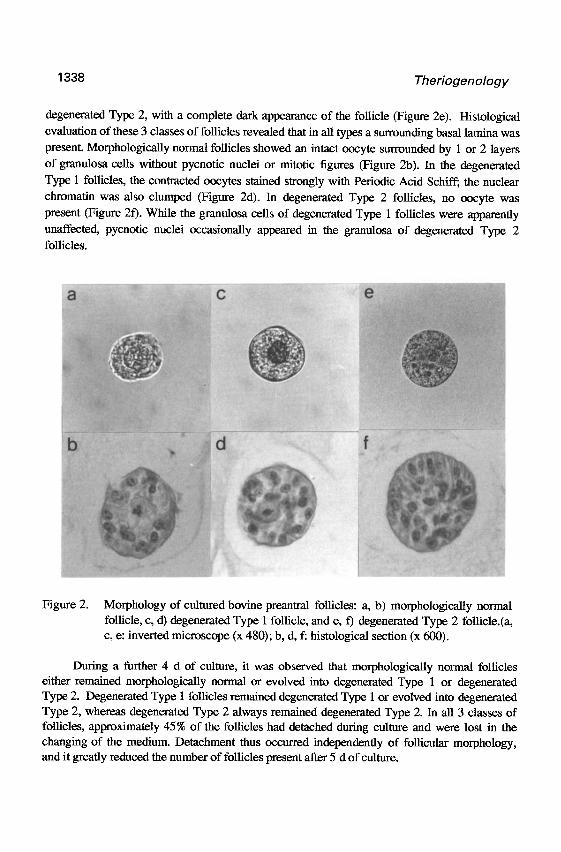
degenerated Type 2, with a complete dark appearance of the follicle (Figure 2e). Histological evaluation of these 3 classes of follicles revealed that in all types a surrounding basal lamina was
present. Morphologically normal follicles showed an intact oocyte surrounded by 1 or 2 layers
of granulosa cells without pycnotic nuclei or mitotic figures (Figure 2b). In the degenerated
Type 1 follicles, the contracted oocytes stained strongly with Periodic Acid Schiff; the nuclear
chromatin was also clumped (Figure 2d). In degenerated Type 2 follicles, no oocyte was
present (Figure 20. While the granulosa cells of degenerated Type 1 follicles were apparently
unaffected, pycnotic nuclei occasionally appeared in the granulosa of degenerated Type 2
follicles.
Figure 2. Morphology of cultured bovine preantral follicles: a, b) morphologically normal follicle, c, d) degenerated Type 1 follicle, and e, f) degenerated Type 2 follicle.(a, c, e: inverted microscope (x 480); b, d, f: histological section (x 600).
During a further 4 d of culture, it was observed that morphologically normal follicles either remained morphologically normal or evolved into degenerated Type 1 or degenerated Type 2. Degenerated Type 1 follicles remained degenerated Type 1 or evolved into degenerated Type 2, whereas degenerated Type 2 always remained degenerated Type 2. In all 3 classes of follicles, approximately 45% of the follicles had detached during culture and were lost in the changing of the medium. Detachment thus occurred independently of follicular morphology, and it greatly reduced the number of follicles present after 5 d of culture.

Theriogenology
Effect of Pyruvate, Glutamine and Hypoxanthine on the Percentage of Morphologically Normal
Follicles During Culture
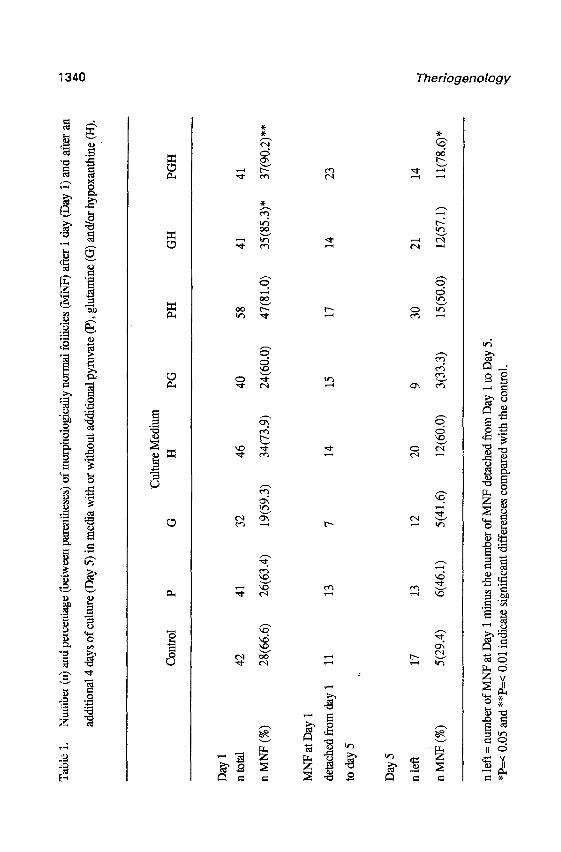
Table 1 shows the numbers and percentages of morphologically normal follicles at Day 1
and Day 5 of culture in 8 different media. At Day 1, a higher percentage of morphologically
normal follicles was observed in medium supplemented with a mixture of pyruvate, glutamine
and hypoxanthine (PcO.01) or a mixture of hypoxanthine and glutamine (PcO.05) compared
with the control. Other mixtures (pyruvate plus glutamine and pyruvate plus hypoxanthine) or
supplements alone (pyruvate, glutamine and hypoxanthine) did not significantly increase the
percentage of morphologically normal follicles compared with the control.
To evaluate the capacity of different culture media in preserving normal folhcular
morphology during an additional 4 d of culture, morphologically normal follicles presented at
Day 1 were individually reevaluated at Day 5 (Table 1). During this period, 36.2 to 62.5% of
the morphologically normal follicles had detached from the plastic substrate and were lost
during changing of the medium. This explains the difference between the number of follicles
classified as morphologically normal follicles at Day 1 of culture and the number available to
calculate the percentages of morphologically normal follicles at Day 5 (e.g., for the control
group; of the 28 follicles classified as morphologically normal follicles at Day 1 of culture, 11
had detached and were lost during the changing of medium, whereas 17 follicles were still
attached. of these 17 follicles, 5 (29.4%) remained morphologically normal follicles). During
an additional 4 d of culture (Day 5), the highest percentage of morphologically normal follicles
was observed when control medium was supplemented with a mixture of pyruvate, glutamine
and hypoxanthine. Addition of pyruvate, glutamine and hypoxanthine alone and mixtures of
pyruvate plus glutamine, pyruvate plus hypoxanthine, and glutamine plus hypoxanthine
increased the percentage of morphologically normal follicles compared with that of the wntrol
group, but the differences were not statistically significant_
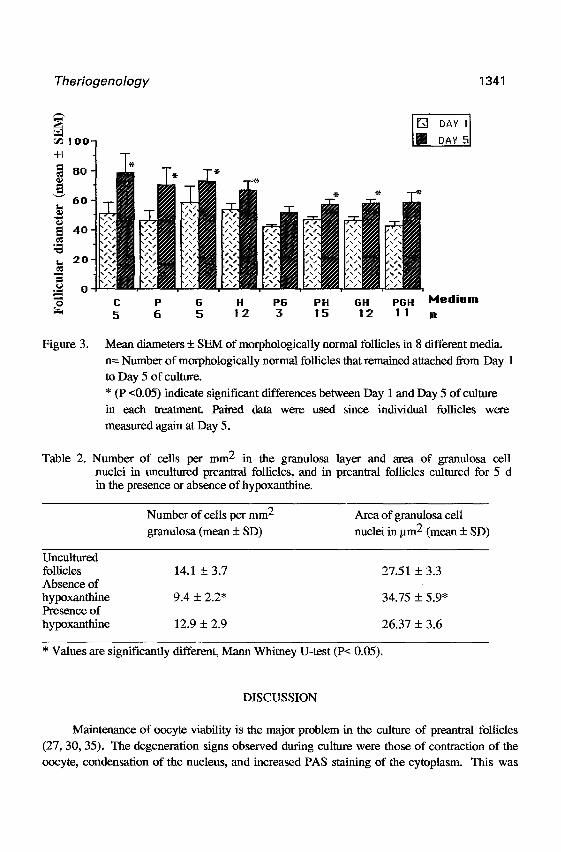
Figure 3 shows the mean diameter f SEM of morphologically normal follicles at Day 1
and Day 5 of culture. Mean diameters of morphologically normal follicles significantly increased
during culture in all the media tested (PcO.05) excepted for the medium suplemented with a
mixture of pyruvate and glutamine (PsO.05). The mean diameters among the different media
were not statistically different, either at Day 1 or at Day 5 (PrO.05). Larger variations in the
increase of follicular diameter were observed among follicles cultured in the absence of
hypoxanthine. Furthermore, the area of the granulosa cell nuclei was larger and the number of
cells per mm2 in the granulosa layer was lower in follicles cultured for 5 d in the absence of
hypoxanthine than in the uncultured follicles or in follicles cultured in the presence of hypoxanthine (Table 2).
Tab
le 1
. N
umbe
r (n)
and
per
cent
age (
betw
een p
aren
thes
es) o
f mor
phol
ogic
ally
norm
al fo
llicl
es (M
NP)
afte
r 1
day
(Day
1) a
nd a
fter
an
z
addi
tiona
l 4 d
ays o
f cul
ture
(Day
5) i
n m
edia
with
or w
ithou
t add
ition
al py
ruva
te (P
), g
luta
min
e (G
) and
/or h
ypox
anth
ine (
H).
$
Con
trol
P
%ul
ture
Med
ium
G
H
PG
PH
G
H
PGH
Day
1
ntot
al
n M
NF
(%)
MN
PatD
ay 1
deta
ched
from
day
1
toda
y5
Day
5
n le
ft
n M
NF
(%)
42
41
32
46
40
58
41
41
28(6
6.6)
26
(63.
4)
19(5
9.3)
34
(73.
9)
24(6
0.0)
47
(81.
0)
35(8
5.3)
* 37
(90.
2)**
11
13
7 14
15
17
14
17
13
12
20
9 30
21
5(29
.4)
6(46
.1)
5(41
.6)
12(6
0.0)
3(
33.3
) 15
(50.
0)
12(5
7.1)
n le
ft =
num
ber o
f MN
P at
Day
1 m
inus
the
num
ber o
f MN
F d
etac
hed
from
Day
1 to
Day
5.
*P=
< 0
.05
and
**P=
< 0.
01 in
dica
te si
gnif
ican
t dif
fere
nces
com
pare
d w
ith th
e co
ntro
l. 9 0 0‘
23
14
5r
1 l(7
8.6)
” 5.
3
_ __
__
___.
__
-.
.-
. ._
-
_I
-.
__
_~
-.
- -
- -
. -
Theriogenology 1341
ij 40 .C( a Li 2f9 m ‘3 .Y 0 = g
13 P G PG PH GH ~GH Medium 5 6 5 lH2 3 15 12 11 u
Figure 3. Mean diameters f SEW of morphologically normal follicles in 8 different media.
n= Number of morphologically normal follicles that remained attached from Day 1 to Day 5 of culture.
* (P ~0.05) indicate significant differences between Day 1 and Day 5 of culture
in each treatment. Paired data were used since individual follicles were
measured again at Day 5.
Table 2. Number of cells per mm2 in the granulosa layer and area of granulosa cell nuclei in uncultured prea.ntral follicles, and in preantral follicles cultured for 5 d in the presence or absence of hypoxanthine.
Number of cells per mm2
granulosa (mean + SD)
Area of granulosa cell
nuclei in pm2 (mean f SD)
Uncultured follicles Absence of hypoxanthine Presence of hypoxanthine
14.1 f 3.7 27.51 f 3.3
9.4 f 2.2* 34.75 f. 5.9*
12.9 rt 2.9 26.37 + 3.6
* Values are significantly different, Mann Whitney U-test (PC 0.05).
DISCUSSION
Maintenance of oocyte viability is the major problem in the culture of pmantral follicles (27,30,35). The degeneration signs observed during culture were those of contraction of the oocyte, condensation of the nucleus, and increased PAS staining of the cytoplasm. This was

1342 Theriogenology
followed by complete disappearance of the oocyte. These degenerative changes correspond with those described for atretic preantral follicles in vivo (6). In preliminary unpublished experiments using bovine follicles, we observed that follicle viability appeared to be negatively
affected by excessive manipulation of preantral follicles before culture, by the duration of time
(more than 4 h) between the beginning of the isolation procedure and the onset of culture, and
by the composition of the isolation medium. When PBS was used as the isolation medium,
most follicles were degenerated Type 1 follicles after 1 d of culture.
The present study showed that addition of pyruvate, glutamine or hypoxanthine to the
culture medium increased the percentage of follicles with an intact oocyte from 29.4 to 78.6 %.
Maintenance of the oocyte in almost 30 % of the follicles cultured in control medium may be due
to the presence of pyruvate in MEM. Previously it was shown that pyruvate was the
predominant substrate used by immature and mature bovine oocytes and zygotes (32). In
addition, Eppig (16) reported that isolated growing mouse oocytes utilized exogeneously
administered pyruvate more efficiently than glucose, lactate or succinate. Several authors (2, 3,
11, 43) have shown that glutamine is an efficient energy substrate for bovine and mouse
embryos, and for hamster and rabbit oocytes. Eppig et al. (19) demonstrated that hypoxanthine
promoted the association of oocytes and accompanying granulosa cells during the culture of
mouse preantral follicles; approximately 70 % of the oocytes became spontaneously denuded of
their granulosa cells when cultured in medium without hypoxanthine. The maintenance of
oocyte-granulosa cell communication via gap junctions has been shown to promote oocyte
growth in vitro (5). More than 85% of the metabolites of the oocytes are transported from the
granulosa cells to the oocyte via gap junctions (26). We showed that micromorphological
appearance of follicles cultured in the presence of hypoxanthine was similar to that of uncultured
follicles. It is probable that under our conditions, hypoxanthine improves the utilization of additional energy substrata by maintaining interactions between the oocyte and the surrounding
granulosa cells. In addition, hypoxanthine has been used in vitro to prevent the resumption of
meiosis in oocytes of different species (14), although a limited inhibitory effect has been reported in bovine oocytes (41). Oocytes acquire meiotic competence when they reach 80% of
their !dnal size (44). In the bovine species, this occurs when the follicles reach the antral stage
(i.e., diameter larger than 3 mm; 42). Consequently, the oocytes enclosed in our preantral
follicles used in our present study were far from acquiring meiotic competence.
Morphological examination using the inverted microscope has been shown to be very useful for the evaluation of preantral follicles in vitro. However, histological evaluation has been shown indispensable for the identification of an intact oocyte and to judge the quality of the
gmnulosa cells. Enlargement of preantral follicles after culture in all tested media can only be understood after histological examination. Increase in follicle diameter may be caused by
increase in oocyte diameter and by proliferation of the granulosa cells. Oocyte diameter is difficult to follow during culture, primarily in secondary follicles, due to the limited visibility of

Theriogenology 1343
the oocytes under an inverted microscope. proliferation of the granulosa cells should be tested
in future experiments by adding 3H-thymidine (28) or by bromo-deoxy-uridine (31) to the
culture medium followed by autoradiography or immunohistochemistry. Increase in diameter
may be also be due to enlargement of the granulosa cells, as indicated by the lower number of
cells per mm2 in the granulosa layer, and observed in follicles cultured in medium without
hypoxanthine. The increase in size of the granulosa cells and their nuclei resembles the
differentiation of granulosa cells in response to LH/hCG (1, 10, 13), and thus probably reflects
abnormal follicular development. In follicles cultured with a mixture of pyruvate, glutamine and
hypoxanthine, the appearance of the granulosa cells corresponds with that of uncultured
follicles. The increase in follicular diameter in these latter follicles may, therefore, be due to
proliferation of the granulosa cells.
We observed that 36.2 to 62.5 % of the morphologically normal follicles present at Day 1
of culture later detached from the plastic substratum and were lost during changing of the
medium. Loss of follicles can be attributed to the culture method used and not to the
degeneration of follicles since morphologically normal, degenerated Type 1, and degenerated
Type 2 follicles were lost to the same extent. It is known that attachment of cells to the
substratum may be affected by several factors such as temperature (24, 46) and pH, the
presence of C!a+2 and Mg+2, the concentration of serum (33), the type of cell (21, 36) and
adhesive proteins such as fibronectin (23,25). In culture, cells attach to the substratum either by
producing their own attachment factor or by binding to the attachment factors absorbed on the
substratum or those present in the culture medium (39). Fetal calf serum promotes the
attachment of many cell types in vitro by providing attachment factors (33), although in the
present study, addition of 10% FCS to the culture medium did not prevent follicular
detachment. Follicles probably attach to the substratum by synthesizing attachment factors.
Carnegie (7) reported that the secretion of fibronectin by rat granulosa cells occurs during early follicular development. When dark follicles were placed in culture, they did not attach to the
substratum and are apparently not able to produce attachment factors.
In our present study, granulosa cells did not spread away from the oocytes during culture.
In other studies (18, 35), spreading of granulosa cells from the oocytes was observed when preantral follicles were cultured. These preantral follicles were obtained after enzymatic
digestion with collagenase or trypsin, and it was reported that they were not surrounded by a
basal lamina. Using a mild mechanical isolation procedure, the basal lamina was preserved, and this may have prevented the spreading of granulosa cells in culture and consequently maintained
follicle structure in vitro.
In summary, we conclude that small bovine preantral follicles can be cultured in vitro for
5 d in a medium containing pyruvate, glutamine and hypoxanthine. Furthermore, we have
presented morphological criteria for cultured normal and degenerated follicles and showed that histological examination is indispensable for evaluating the quality of cultured follicles.

1344 Theriogenology
REFERENCES
1. Amsterdam A, Rothemensch S. Structure-function relationships during granulosa cell differentiation.
Endocr Rev 1987;8: 309337. 2. Bae M, Foote RH. Carbohydrate and aminoacids requirements and ammonia production of rabbit
follicularoocytes in vitro. Expl Cell Res 1991;91: 113-l 18. 3. Bavister BD, Leibfried ML, Lieberman G. Development of preimplantation embryos of golden hamster
in a defined culture medium. Biol Reprod 1983;28: 235-247. 4. Betteridge RI, Smith C, Stubbings RB, Xu XP, Ring WA. Potential genetic improvement of cattle by
fertilization of fetal oocytes in vitro. J Reprod Fertil 1989;38 (Suppl.): 87-98. 5. Buccione R, Schroeder AC, Eppig JJ. Interactions between somatic cells and germ cells throughout
mammalian oogenesis. Biol Reprod 199@,43: 543-547. 6. Byskov AG. Follicular atresia. In: Jones, R.E. (ed) The vertebrate ovary, plena in press, New York,
1978,533-562. 7. Carnegie JA. Secretion of fibronectin by rat granulosa cells occurs primarily during early follicular
development. J Reprod Fertil 1991;89: 579-589. 8. Carroll J, Whittingham DG, Wood MJ. Effect of dibutyryl cyclic adenosme monophosphate on granulosa
cell proliferation, oocyte growth and meiotic maturation in isolated mouse primary ovarian follicles cultured in collagen gels. J Reprod fertil 199 1;92: 197-207.
9. Carroll J, Whittingham DG, Wood MJ. Effect of gonadotrophin environment on growth and development of isolated mouse primary ovarian follicles. J Reprod Fertii 1991;93: 71-79.
10. Cavender JL, Murdoch WJ. Morphological studies of microcirculatory system of periovulatory ovine follicles. Biol Reprod 1989;41: 309-316.
11. Chatot CL, Tasca RJ, Ziomek CA. Glutamine uptake and utilization by preimplantation mouse embryos in CZB medium. J Reprod Fertil1990;89: 335-346.
12. Daniel AJ, Armstrong DT, Gore-langton RE. Growth and development of rat oocyte in vitro. Gam Res 1989;24: 109-121.
13. Dieleman SJ, Kruip TAM, Fontijne P, De Jong WHR, van der Weyden GC. Changes in oestradiol, progesterone and testosterone concentration in foll@rlar fluid and in the micromorphology of preovulatory bovine follicles relative to the peak of luteinizing hormone. J Endocrinol1983;97: 3 l-42.
14. Downs SM. Factors affecting the resumption of meiotic maturation in mammalian oocytes. Theriogenology 1993;39: 65-69.
15. Downs SM, Coleman DL, Ward-Bailey PF, Eppig JJ. Hypoxanthine is the principal inhibitor of murine oocyte maturation in a low molecular weight traction of porcine follicular fluid. Proc Nat1 Acad Sci USA 1985;82: 454-458.
16. EppigJJ. Analysis of mouse oogenesis in vitro. Oocyte isolation and the utilization of exogenous energy sources by growing oocytes. J Bxp Zoo1 1976;198: 375-382.
17. Eppig JJ. Mouse oozyte development in vitro with various culture systems. Dev Biol 1977;60: 371-378. 18. Eppig JJ. Growth and development of mammalian oocytes in vitro. Arch Path01 Lab Med 1992; 116: 379-
382. 19. Eppig J, Downs SM. The effect of hypoxanthine on mouse oocyte growth and development in vitro:
maintenance of meiotic arrest and gonadotropin-induced oocyte maturation. Dev Biol 1987;119: 313-321. 20. Eppig JJ, Schoeder AC. Capacity of mouse oocyte t?om preanttal follicles undergoes embryogenesis and
development to live young after growth, maturation, and fertilization in vitro. Biol Reprod 1989;41: 268- 276.
2 1. Elsale T, Bard J. Collagen substrata for studies on cell behavior. J Cell Biol 1972;54: 626-637.

Theriogenology 1345
22. Figueiredo JR, Hulshof SCJ, Van den Hurk R, Ectors FJ, Fontes RS, Nusgens B, Bevers MM, Beckers JF. Development of a new mechanical and enzymatic method for the isolation of intact preanlral follicles from fetal, calf and adult bovine ovaries. Theriogenology 1993;40: 789-799.
23. Gold LI, Pearlstein E. Fibronectin-Collagen binding and requirement during cellular adhesion. B&hem J 1980;186: 551-559.
24. Goldberg B. Binding of soluble type I collagen molecules to the fibroblast plasma membrane. Cell 1979;16: 265-275.
25. Grinnell F, Feld MK. Initial adhesion of human fibroblasts in serum free-medium: Possible role of secreted fibronectin. Cell 1979;17: 117-129.
26. Heller D, Cahill D, Schultz RM. Biochemical studies of mammalian oogenesis: metabolic cooperativity between gmnulosa and growing oocytes. Dev Biol 1981;84: 455-464.
27. Hirao Y, Miyano T, Kato S. In vitro growth of porcine oocytes. 12th Int. Cong Anim Reprod 1992;l: 333-335.
28. Hirshifield AN. Granulosa cell proliferation in very small follicles of cycling rats studied by long-term continuous tritiated-thymidme infusion. Biol Reprod 1989;41: 309-316.
29. Jewgenow K, Pitra C. Die Isolierung van Praamralfolikeln aus EierstGcken des Rindes. Reprod Dom Anim 1991;26: 281-289.
30. Jewgenow K, Pitra C. Hormone-contiolled culture of secondary follicles of domestic cats. Theriogenology 1993;39: 527-535.
3 1. Kant HJG. Periodic acid incubation can be relate hydrochloric acid hydrolysis and trypsin digestion in immunogold-silver staing of bromo-deoxy-uridine incorporation in plastic sections and allows the PAS reaction. Histochem J 1992;24: 170-175.
32. Khurana N, Niemann H. Energy metabolism of bovine oocytes matured and fertilized in vitro. 12th Int. Cong Anim Reprod 1992;l: 348-350.
33. Klebe RJ. Isolation of a collagen-dependent cell attachment factor. Nature 1974;250: 248-251. 34. Leichthammer F, Baunack E, Brem G. Behavior of living primordial germ cells of livestock in vitro.
Theriogenology 1990;33: 1221-1230. 35. Maresh G A, Timmons TT, Dunbar BD. Effect of matrix on the expression of specific ovarian proteins.
Biol Reprod 1990;43: 965-976. 36. Murray JC, Liotta L, Rennard SI, Martin GR. Adhesion characteristics of murine me&static and
nonmetastatic tumor cells in vitro. Cancer Res 1979;40: 347-35 1. 37. Nayudu PL, Osbom SM. Factors influencing the rate of preantml and antral growth of mouse ovarian
follicles in vitro. J Reprod Fertil 1992;95: 349-362. 38. Nuttinck F, Mermillod P, Massip A, Dessy F. Characterization of in vitro growth of bovine preantral
follicles: A preliminary study. Theriogenology 1993;39: 81 l- 821.
39. Pearlsten E. Substrata activation of the cell adhesion factor as prerequisite for cell attachment. Int J Cancer 1978;22: 32-35.
40. Qvist R, Blackwell LF, Boume H, Brown JB. Development of mouse ovarian follicles from primary to ovulatory stage in vitro. J Reprod Fertil 1990: 89: 169-180.
4 1. Sirard MA. ~Temporary inhibition of in vitro meiotic resumption by adenylate cyclase stimulation in immanture bovine oocytes. Theriogenology 1990;33: 757-767.
42. Szijllijsi D. Maturation de l’ovocyte. In: Thibault, C. et Ievasseur, M.C. (ed) Ia reproduction chez les mammif&s et l’homme, INRA-Ellipses, 1991,299-314.
43. Takahashi Y, First NL. In vitro development of bovine one cell embryos: influence of glucose, lactate, pyruvate, amino acids and vitamins. Theriogenology 1992;37: 963-968.

Theriogenology
44. Thibault C, Sziilliisi D, G&ard M. Mammalian oocyte maturation. Reprod Nutr D&elop 198727: 865 896
45. Torrance C, Telfer E?, Gosden RG. Quantitative study of development of isolated preantral follicles in collagen gel culture. _I Reprod Fertil 1990;87: 367-374.
46. UedaMJy Ito T, Oka& TS, Ohnishi iA. Correlation between membrane fluidity and the critical temperature for cell adhesion. J Cell BioI 197671: 67@674.
47. Warikoo PK, Bavister BD. Hypoxanthine and cyclic adenosine 5’-monophosphate maintain meiotic arrest of rhesus monkey oocytes in vitro. Fertil Steril1989;51: 886-889.
Related Documents