Palaeogeography, Palaeoclimatology, Palaeoecology 158 ( 2000) 175–195 www.elsevier.nl/ locate/palaeo Present-day coccolith fluxes recorded in central eastern Mediterranean sediment traps and surface sediments P. Ziveri a,b, * , A. Rutten c, G.J. de Lange c, J. Thomson d, C. Corselli a a Department of Earth Sciences, University of Milan, Via Mangiagalli 34, 20133 Milan, Italy b Faculty of Earth Sciences, Vrije Universiteit Amsterdam, de Boelelaan 1085, 1081 HV Amsterdam, The Netherlands c Institute of Earth Sciences, Department of Geochemistry, Utrecht University, Budapestlaan 4, 3584 CD Utrecht, The Netherlands d Southampton Oceanography Centre, European Way, Empress Dock, Southampton SO14 3ZH, UK Accepted for publication 15 June 1999 Abstract Two sediment traps were deployed in time series collection from November 1991 to August 1994 at 3000 and 3500 m, respectively, above and below the oxygenated sea water/anoxic brine interface in the Bannock Basin, central eastern Mediterranean. Here, the coccolithophore flux and its contribution to the carbonate particulate flux are presented for the 3000 m trap and compared with the record in eastern Mediterranean surface sediments. A marked seasonal variation is observed in the fluxes of total mass, total coccoliths and whole coccospheres, with flux maxima in late winter and spring. The annual coccolith flux of 1×1010 coccoliths m-2 year -1 measured in the deep waters of Bannock Basin is much lower than published data from most other oceanographic settings, even when corrected for the trap e ciency of ~23% calculated from the 230Th flux. The biogenic and lithogenic fluxes are primarily controlled by coccolithophore production and Saharan dust input, respectively. The calculated coccolith and coccosphere settling rates, estimated from the comparison of maximum pigment concentration in the surface ocean and arrival of maximum flux at 3000 m water depth, ranged from 17 to 25 m day-1 for coccoliths and 100 m day-1 for coccospheres. At the study site, carbonate dissolution is a minor process at both the trap depth and at the sea floor in both oxic and anoxic conditions, and there is a high preservation of coccolith CaCO 3 . Coccolithophores are the main contributor to the biogenic carbonate flux, followed by thoracosphaerids. Emiliania huxleyi and Florisphaera profunda followed by Syracosphaera, Helicosphaera carteri and Calcidiscus leptoporus are the dominant species in the sediment trap and surface sediments. © 2000 Elsevier Science B.V. All rights reserved. Keywords: calcareous composition; east Mediterranean; flux; nanoplankton; recent; sediment traps 1. Introduction production is tightly coupled with primary pro- duction. In particular, 85% of the primary pro- The eastern Mediterranean is an area with duction in the eastern Mediterranean flows to the extremely oligotrophic or ultra-oligotrophic condi- microbial food web ( Turley, 1997 ). Phytoplankton tions ( Berland et al., 1988; Dugdale and Wilkerson, populations are dominated by picoplankton (Li 1988; Yacobi et al., 1995). Heterotrophic bacterial et al., 1993) with the bacterial biomass constituting about 50% of total algal biomass (Robarts et al., 1996). The central eastern Mediterranean is net * Corresponding author. E-mail address: [email protected] (P. Ziveri) phototrophic, resulting in a sink for atmospheric 0031-0182/00/$ - see front matter © 2000 Elsevier Science B.V. All rights reserved. PII: S0031-0182(00)00049-3

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Palaeogeography, Palaeoclimatology, Palaeoecology 158 (2000) 175–195www.elsevier.nl/locate/palaeo
Present-day coccolith fluxes recorded in central easternMediterranean sediment traps and surface sediments
P. Ziveri a,b,*, A. Rutten c, G.J. de Lange c, J. Thomson d, C. Corselli aa Department of Earth Sciences, University of Milan, Via Mangiagalli 34, 20133 Milan, Italy
b Faculty of Earth Sciences, Vrije Universiteit Amsterdam, de Boelelaan 1085, 1081 HV Amsterdam, The Netherlandsc Institute of Earth Sciences, Department of Geochemistry, Utrecht University, Budapestlaan 4, 3584 CD Utrecht, The Netherlands
d Southampton Oceanography Centre, European Way, Empress Dock, Southampton SO14 3ZH, UK
Accepted for publication 15 June 1999
Abstract
Two sediment traps were deployed in time series collection from November 1991 to August 1994 at 3000 and3500 m, respectively, above and below the oxygenated sea water/anoxic brine interface in the Bannock Basin, centraleastern Mediterranean. Here, the coccolithophore flux and its contribution to the carbonate particulate flux arepresented for the 3000 m trap and compared with the record in eastern Mediterranean surface sediments. A markedseasonal variation is observed in the fluxes of total mass, total coccoliths and whole coccospheres, with flux maximain late winter and spring. The annual coccolith flux of 1×1010 coccoliths m−2 year−1 measured in the deep waters ofBannock Basin is much lower than published data from most other oceanographic settings, even when corrected forthe trap efficiency of ~23% calculated from the 230Th flux. The biogenic and lithogenic fluxes are primarily controlledby coccolithophore production and Saharan dust input, respectively. The calculated coccolith and coccosphere settlingrates, estimated from the comparison of maximum pigment concentration in the surface ocean and arrival of maximumflux at 3000 m water depth, ranged from 17 to 25 m day−1 for coccoliths and 100 m day−1 for coccospheres. At thestudy site, carbonate dissolution is a minor process at both the trap depth and at the sea floor in both oxic andanoxic conditions, and there is a high preservation of coccolith CaCO3. Coccolithophores are the main contributorto the biogenic carbonate flux, followed by thoracosphaerids. Emiliania huxleyi and Florisphaera profunda followedby Syracosphaera, Helicosphaera carteri and Calcidiscus leptoporus are the dominant species in the sediment trap andsurface sediments. © 2000 Elsevier Science B.V. All rights reserved.
Keywords: calcareous composition; east Mediterranean; flux; nanoplankton; recent; sediment traps
1. Introduction production is tightly coupled with primary pro-duction. In particular, 85% of the primary pro-
The eastern Mediterranean is an area with duction in the eastern Mediterranean flows to theextremely oligotrophic or ultra-oligotrophic condi- microbial food web (Turley, 1997). Phytoplanktontions (Berland et al., 1988; Dugdale and Wilkerson, populations are dominated by picoplankton (Li1988; Yacobi et al., 1995). Heterotrophic bacterial et al., 1993) with the bacterial biomass constituting
about 50% of total algal biomass (Robarts et al.,1996). The central eastern Mediterranean is net* Corresponding author.
E-mail address: [email protected] (P. Ziveri) phototrophic, resulting in a sink for atmospheric
0031-0182/00/$ - see front matter © 2000 Elsevier Science B.V. All rights reserved.PII: S0031-0182 ( 00 ) 00049-3

176 P. Ziveri et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 158 (2000) 175–195
CO2. Although eastern Mediterranean primary and to assess the quality and accuracy of theinformation preserved in the sediment record.production estimates are generally low (Dugdale
and Wilkerson, 1988), so far, no one has studied In the central eastern Mediterranean, coccoli-thophores are the dominant phytoplankton groupin detail how the productivity changes over the
annual cycle. that produces carbonate (Ziveri et al., 1995). Inthis study, we concentrate on the coccolithophoreCoccolithophores are pelagic unicellular algae
that secrete calcite plates. They are a major constit- flux (number m−2 day−1) and its seasonality. Inaddition, annual fluxes will be compared withuent of marine phytoplankton and make a major
contribution to the biogenic carbonate content in those estimated for nearby surface sediments.Present-day coccolithophore fluxes in the easterndeep-sea sediments (Bernard and Lecal, 1953;
Milliman and Muller, 1973; Milliman, 1993). With Mediterranean have been measured by two auto-mated sediment traps capable of collecting par-their fast turnover, coccolithophores may have an
important effect on the CO2 cycle in the global ticles for specified time intervals over a long period.ocean. They influence the global climate systemthrough the organic carbon and carbonate pumpsand by the emission of dimethyl sulfide, which 2. Material and methodsmay affect cloud albedo (Andreae, 1986; Charlsonet al., 1987; Matrai and Keller, 1993; Westbroek Two time-series sediment traps (Technicap
PPS5/2) were deployed in the southwesternet al., 1993). Satellite imagery by the Coastal ZoneColor Scanner (CZCS) revealed the formation of Bannock Basin (Libeccio subbasin; 34°18∞N;
20°01∞E), a central eastern Mediterranean anoxicextensive blooms of the coccolithophore Emilianiahuxleyi at higher latitudes in cooler, nutrient-rich basin (Scientific Staff of Cruise Bannock 1984-12,
1985) (Fig. 1). Collection was from Novemberwaters (Holligan et al., 1983; Brown and Yoder,1994; Balch et al., 1997). A better understanding 1991 until August 1994. During the 927 day
deployment, each sediment trap opened and closedof coccolithophore ecology is further required touse them as a biotic proxy of past climate change 96 times, providing a near-continuous time series
Fig. 1. Map of the central eastern Mediterranean showing the locations of the sediment trap mooring (ST) and box cores (MT6,UM15, UM35 and UM26) used in this study.
177P. Ziveri et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 158 (2000) 175–195
Table 1Sediment trap mooring phases, sampling period per phase, sample interval and deployment days per sampling phasea
Mooring phase Phase start date Phase end date Deployment (days) Sample interval (days)
ST2 Nov 10, 1991 May 10, 1992 182.5 5–10MT4O Jun 1, 1992 Feb 19, 1993 263 9–14MU-B Apr 1, 1993 Sep 16, 1993 168 7UM1STO Oct 20, 1993 Aug 29, 1994 313 10–20
a Sediment trap position: 34°17.84∞N 20°00.89∞E. Sediment trap depth: 3000 m. Water depth: 3530 m.
(Table 1). The upper trap was located at 3000 m, of the coccolithophores estimated from the traps(Fig. 1, Table 2).above the sea water/brine interface in oxygenated
conditions, whereas the lower trap, at 3500 m, waslocated in anoxic brine close to the bottom of 2.1. Coccolithophore sediment trap sample
preparationLibeccio Basin (Fig. 2 in Rutten et al., 2000).Samples 21, 22, 23 (from 20-04-1992 to 05-05-1992)collected by the trap at 3000 m were empty, All samples were split on recovery on board the
ship into equal fractions using a pneumatic splitterwhereas sample 24 was unusually large, but notechnical failure was apparent. In addition, a high (Tennant and Baker, 1992). For coccolith analysis,
the first three series of samples (1–72) were col-mass flux was recorded in the anoxic trap at thesame time in the synchronous sample 24. For this lected on board upon 0.45 mm cellulose acetate
(samples 1–48: 47 mm diameter; samples 49–72:reason, we include all samples in all calculations.A detailed description of the deployment and 25 mm diameter) using a gas pressure system. The
final set of samples (73–96) were maintained inrecovery of the sediment traps, sample additives,integrity and analytical methods is reported by solution and then split and filtered in the labora-
tory of the Department of Biology, University ofRutten et al. (2000) and Ziveri et al. (1995). Thesediment trap efficiency was evaluated using the Milan. After stirring the solution, a small portion
of the sample was collected using a precisionflux of the long-lived radionuclide 230Th (Ruttenet al., 2000). pipette and filtered on to a 0.45 mm cellulose
acetate filter (47 mm diameter).The surfice sediment of four easternMediterranean box cores, from 2160 to 3520 m For the coccolith analysis of samples 1–72, the
material on the filter had to be resuspended off thewater depth, were analyzed for comparison withthe accumulation rates and taxonomic composition filter. Each filter was put into a glass culture tube
Table 2Box core position, sampling interval, water depth, type of sediment and data used for the calculation of the coccolith accumulationrates (excluding coccolith concentration)a
Box Position ( latitude, Sampling Water Sediment type Sedimentation Dry bulk Mass Totalcore longitude) interval depth rate (cm ka−1) density accumulation CaCO3
(cm) (m) (g cm−3) rate (g cm2 ka−1) weight (%)
UM15 35° 17.39∞N 21° 24.82∞E 0.0–1.0 3307 Oxic, homogenized 4.7 0.8 4.2 51.3UM26 33° 23.58∞N 25° 00.93∞E 0.0–0.5 2160 Oxic, homogenized 2.7 0.8 2.6 53.7UM35 35° 11.04∞N 21° 12.54∞E 0.0–1.0 2670 Oxic, homogenized 3.3 0.9 3.1 54.5MT6 34° 17.09∞N 20° 01.16∞E 0.0–0.5 3520 Anoxic, laminated 14.1 0.2 6.1 37.7
a These data include the sedimentation rates (cm ka−1), dry bulk densities (g cm−3), percentage mass accumulation rates(mg cm−2 ka−1) and CaCO3 content (%) of the surface sediments analyzed in the present study. The sedimentation rates and the drybulk densities are from Van Santvoort et al. (1996), excluding the data for MT6 by using 210Pb (Rutten et al., 2000).

178 P. Ziveri et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 158 (2000) 175–195
containing a known volume of filtered (0.2 mm) filtration in order to improve the even distributionof the particles on the filter. Each membrane wasdistilled water buffered with sodium borate. Theoven-dried at 40°C and stored in an air-tightsolution was stirred for a few minutes to allow thecontainer. Coccolithophore quantification was car-detachment of the material from the fiber filter. Aried out on the <32 mm fraction; the >32 mm wasportion (1/8–1/1000 of the original sample) of thequalitatively analyzed to evaluate the sieving effi-solution was collected by precision pipette andciency. The weight percentage of the >32 andrefiltered on to a 0.45 mm cellulose acetate filter<32 mm size fractions for samples of UM15,(47 mm diameter). A 1–2 mm2 area of each filteredUM26 and UM35 was obtained by wet-sievingmembrane was subsequently analyzed using a polar-0.5 g sediment and subsequent oven drying.ized light optical microscope (LM) at 1250× mag-
The sedimentation rates (cm ka−1), dry bulknification to determine the total coccolith,densities (g cm−3), mass accumulation ratescoccosphere and calcareous dinocyst fluxes and the(mg cm−2 ka−1) and CaCO3 content (wt%) of thefluxes of the individual coccolithophore speciessurface sediments analyzed in this study are pre-(Ziveri and Thunell, in press). A larger area (aboutsented in Table 2. The sedimentation rates and dry30 mm2) was then used to quantify fluxes of coc-bulk densities are from Van Santvoort et al. (1996),cospheres and calcispheres of thoracosphaerids (cal-excluding the data for MT6 from Rutten et al. (incareous dinocysts). A scanning electron microscopepreparation). The carbonate content of the fine(SEM) was used to evaluate the sample prepara-fraction was determined by gasometric techniques,tion, coccolithophore taxonomical composition andwith a precision of ±2%coccolith preservation. The flux results on the other
Coccolith accumulation rates were calculatedmajor biogenic contributors will be given in a futureusing the following formula (Lototoskaya et al.,publication (Ziveri, P., Erba, E. Grandi, C.1998; Ziveri and Thunell, 2000):Stefanetti, A., Cita, M., in preparation).
The evenness of the distribution of coccoliths ARc=Dc · DBD · SR (1)across the filter surface was confirmed by quantify-
where ARc=coccolith accumulation rate (numbering their distribution on different areas across theof coccoliths cm−2 year−1), Dc=coccolith densityfilter on samples from different production epi-(number of coccoliths per gram of sediment),sodes. An average of 709 (405–1302) coccolithsDBD=dry bulk density (g cm−3), and SR=sedi-were counted per sample; the number of coccosph-mentation rate (cm year−1).eres counted per sample ranged from 0 to 340.
The coccolith taxonomic composition of thetop sediments was determined by LM in the same2.2. Coccolithophore surface sediment sampleway as the trap samples, and 500 specimens werepreparationcounted for each; SEM was used to evaluate thepreparation of the samples, coccolith carbonateThe total number of coccoliths per gram of drypreservation and species diversity.sediment (coccoliths/g) and the coccolith taxo-
Taxonomic identification for the determinationnomic composition were determined in theof the species diversity follows Ziveri et al. (1995).<32 mm fraction from the top sediment of fourFor quantification, all holococcoliths were classedbox cores (upper 0.5–1 cm) located close to theas holococcolith spp., and the Syracosphaera grouptrap mooring (UM15, UM35, MT6) and at aincludes all Syracosphaera species excluding S.more eastern site (UM26).pulchra and S. histrica.The <32 and >32 mm <1 mm fractions for
each sample were obtained by wet sieving 0.25–0.5 g of freeze-dried sediment following the filtra-
3. Resultstion method described in Lototoskaya et al.(1998). Each size fraction was collected on to 3.1. Sediment trap0.45 mm Millipore filters (47 mm diameter) usingprefiltered distilled water with a sodium borate The seasonal pattern of coccolithophore depos-
ition recorded in the 3000 m Bannock Basin sedi-buffer. A low-pressure vacuum pump was used for
179P. Ziveri et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 158 (2000) 175–195
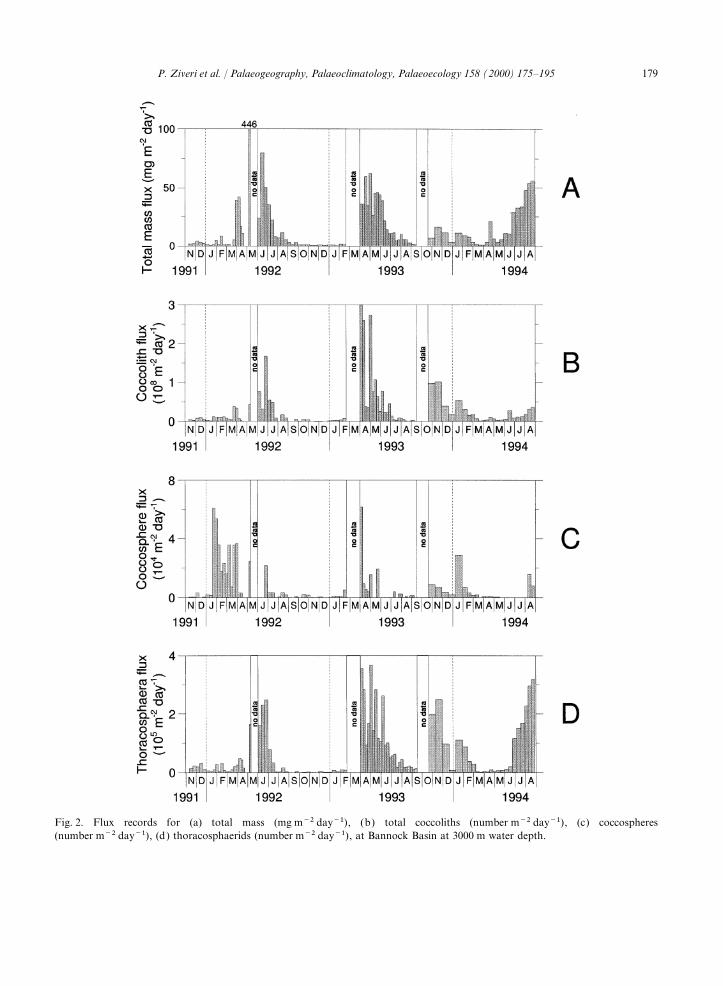
Fig. 2. Flux records for (a) total mass (mg m−2 day−1), (b) total coccoliths (number m−2 day−1), (c) coccospheres(number m−2 day−1), (d) thoracosphaerids (number m−2 day−1), at Bannock Basin at 3000 m water depth.
180 P. Ziveri et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 158 (2000) 175–195
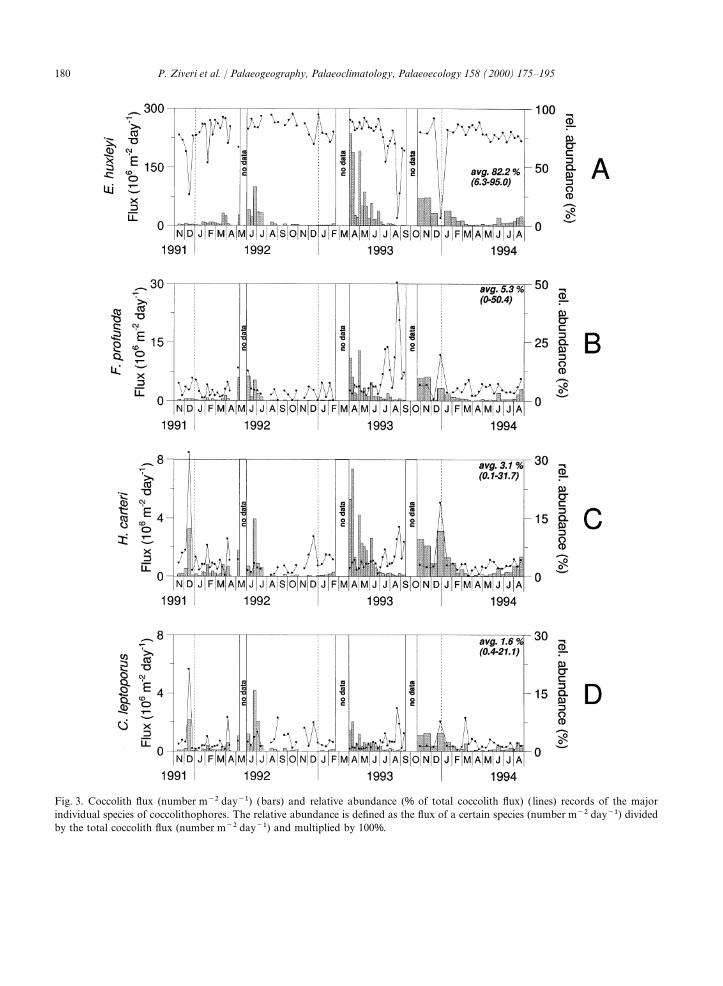
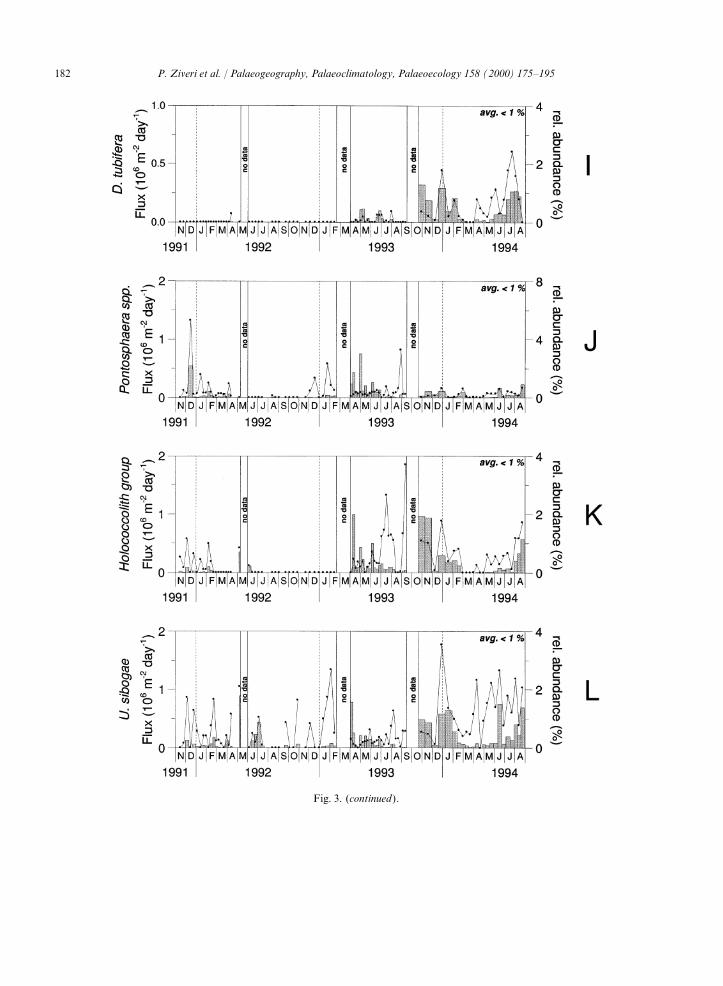
Fig. 3. Coccolith flux (number m−2 day−1) (bars) and relative abundance (% of total coccolith flux) ( lines) records of the majorindividual species of coccolithophores. The relative abundance is defined as the flux of a certain species (number m−2 day−1) dividedby the total coccolith flux (number m−2 day−1) and multiplied by 100%.
181P. Ziveri et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 158 (2000) 175–195
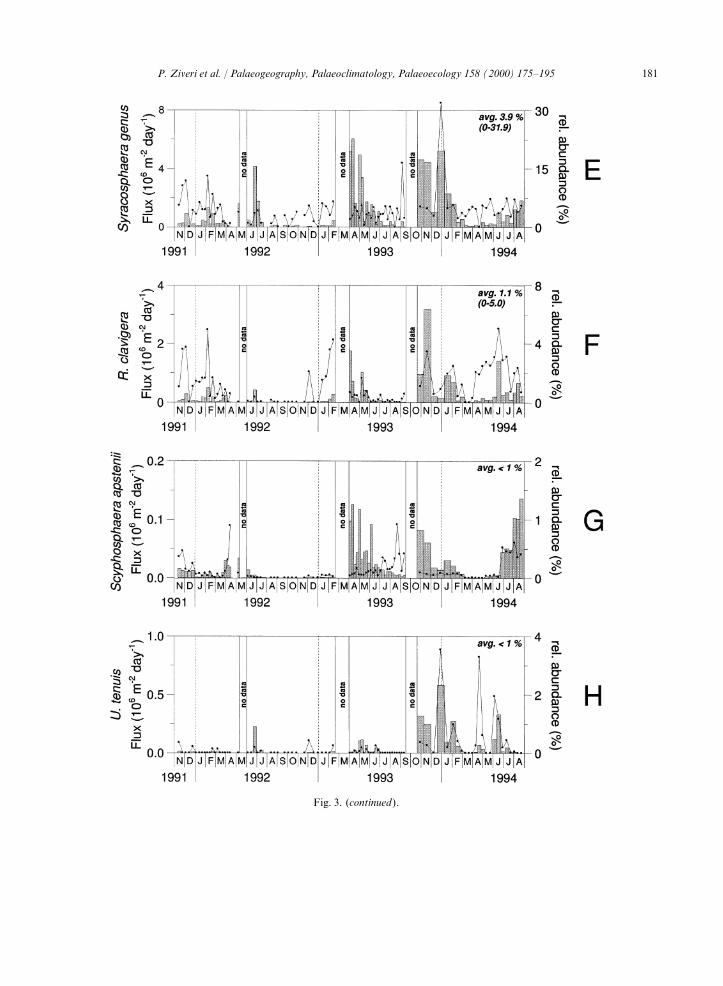
Fig. 3. (continued).
182 P. Ziveri et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 158 (2000) 175–195
Fig. 3. (continued).

183P. Ziveri et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 158 (2000) 175–195
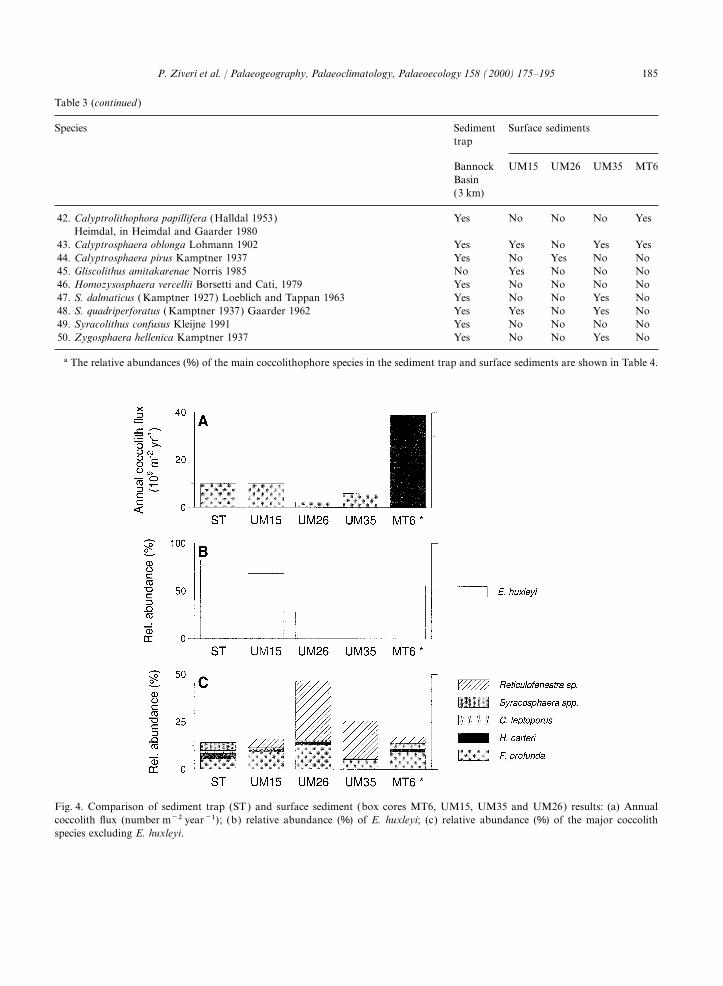
ment trap exhibits the highest coccosphere fluxes 24 wt% and is 15 wt% for box core MT6, locatedduring January–May 1992, April–May 1993 and in the anoxic brine basin. Higher annual coccolithJanuary 1994, and the highest individual cocco- accumulation rates were found in the anoxic sur-lith fluxes during late March–mid-July 1992, face sediments (core MT6) from Bannock BasinApril–June 1993 and late October–January 1994 (Fig. 4; Table 4). The coccolith accumulation(Fig. 2). The maximum coccolith and coccosphere rates range from 2×1012 to 3.9×1013fluxes occur in April 1993, reaching values of coccoliths m−2 ka−1. Emiliania huxleyi dominates3.0×108 coccoliths m−2 day−1 and 6.2×104 the sediment assemblage, followed by F. profunda,coccospheres m−2 day−1, respectively. The contri- H. carteri and C. leptoporus (Fig. 4; Table 4).bution of intact coccospheres to the total coccolithflux is relatively small in comparison to the settlingof individual coccoliths. 4. Discussion
The coccolithophore flux peaks are causedmainly by an increased flux of E. huxleyi (Fig. 3). 4.1. Seasonal trend of coccolithophore andThe taxonomic composition of the coccolithophore thoracosphaerid fluxesassemblages in the trap samples is dominated byE. huxleyi (on average 82.2% of the total coccolith During the 3 year time series, the mass fluxassemblage; range between 6.3 and 95%) and pattern was characterized by low-mass particlesecondarily by F. profunda (average 5.3%; range fluxes with an average of about 16 mg m−2 day–1.0–50.4%), Syracosphaera (average 3.9%; range 0– This value is several times lower than the average31.9%) and H. carteri (average 3.1%; range 0– mass flux measured at 1000 m depth in the North31.2%) (Fig. 3). Balearic Northwestern Mediterranean (185–
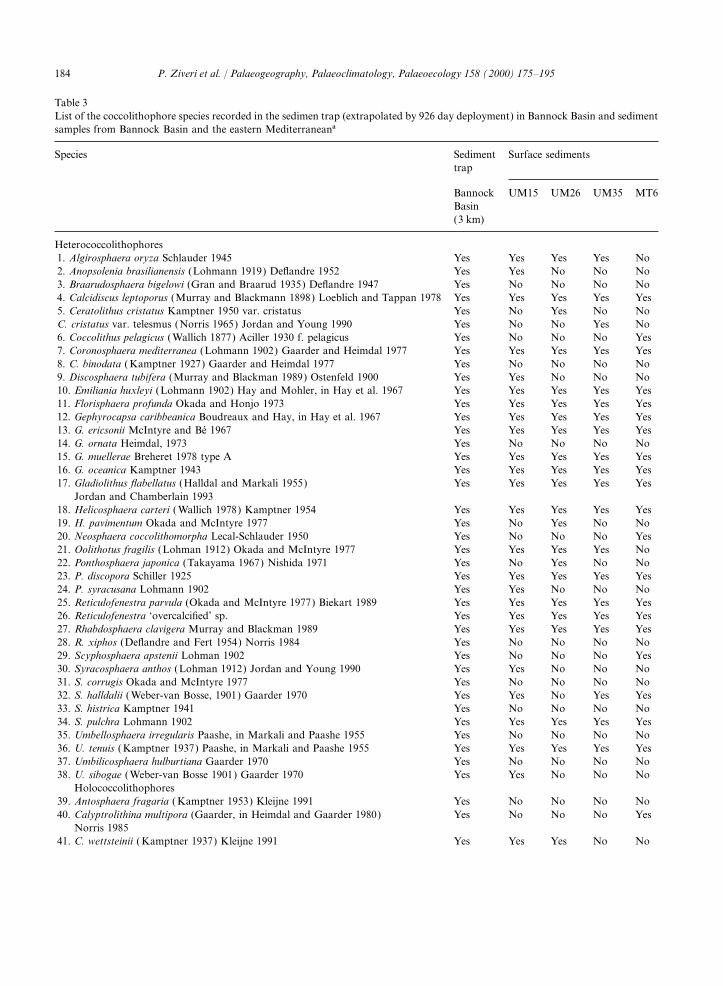
The 50 coccolithophore species identified in the 588 m−2 day–1), Adriatic sea (341 mg m−2 day–1)sediment trap samples are listed in Table 3. The and west of Crete (46–210 mg m−2 day–1)500 specimens counted for each sample lead to a
(Heussner and Monaco, 1996), and in the Alboran95% probability level of detecting a species presentSea (197 mg m−2 day–1; Dachs et al., 1986). It is,in the population at 1%.however, similar to the flux recorded in the westernMaximum fluxes of thoracosphaerids areMediterranean northwest of Corsica (22 mgrecorded during April–June 1992 and 1993, andm−2 day–1; Miquel et al., 1994).from November 1993 to January 1994 and July to
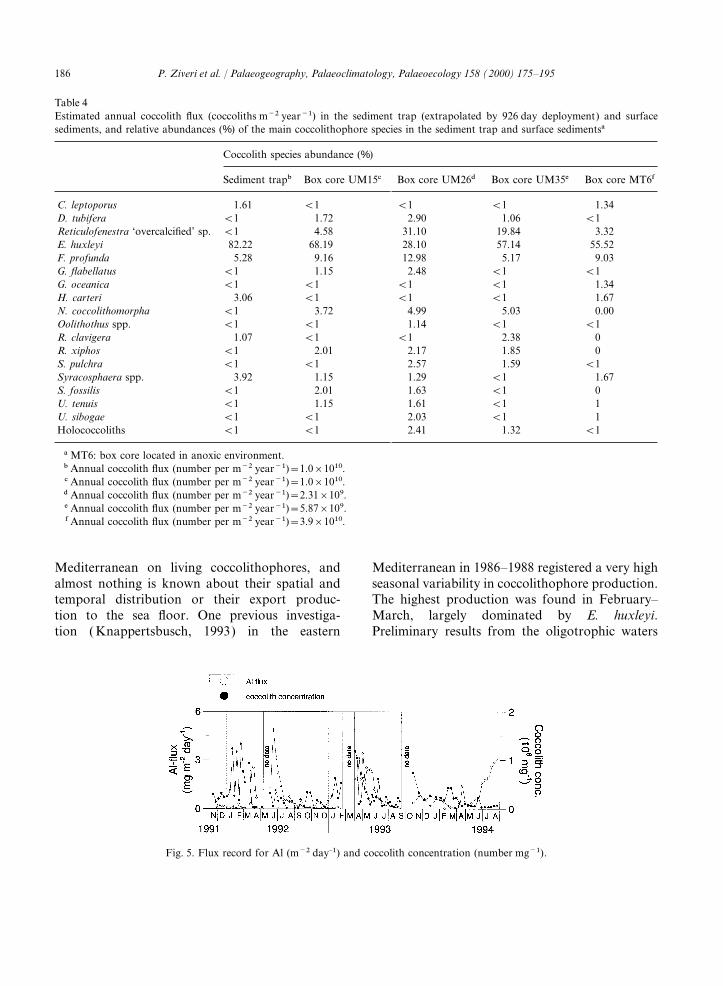
The total mass and coccolith fluxes recorded inAugust 1994 (Fig. 2). The mean flux isBannock Basin present a strong seasonal pattern6.5×104 calcispheres m−2 day−1.with a general increase in late winter–early spring.Coccolithophores are the major contributor toThe major component of the mass flux in thethe biogenic carbonate flux in Bannock Basin andsediment trap appears to be of terrestrial originare also the dominant phytoplankton group(60 wt%) (Rutten et al., 2000). The average totaldetected in the trap samples (Ziveri et al., 1995).carbonate content is relatively low (31.5 wt%)Calcareous dinoflagellates, mainly Thoracosphaeracompared to those in the top sediments of theheimii, are the only additional calcareous compo-eastern Mediterranean. In contrast, the biogenicnent that contributes considerably to the biogenicparticle fluxes in the sediment trap at the Bannockcarbonate fluxes (average of 17%). ForaminifersBasin site are largely dominated by carbonateare present mainly as juvenile forms and have low(70%). Dilution of the biogenic components is,fluxes. An increase in juvenile foraminifer flux hastherefore, an important process. The dilution effectbeen recorded in July–August 1994 (Ziveri et al.,by terrestrial input is demonstrated by the generalin preparation).inverse trends of the Al flux (an indicator ofterrestrial input) and the coccolith concentrations3.2. Surface sedimentsobtained by dividing the coccolith flux by the massflux (Fig. 5).The Ca content of the surface sediments of box
cores UM15, UM26 and UM35 varies from 18 to Little work has been performed in the eastern
184 P. Ziveri et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 158 (2000) 175–195
Table 3List of the coccolithophore species recorded in the sedimen trap (extrapolated by 926 day deployment) in Bannock Basin and sedimentsamples from Bannock Basin and the eastern Mediterraneana
Species Sediment Surface sedimentstrap
Bannock UM15 UM26 UM35 MT6Basin(3 km)
Heterococcolithophores1. Algirosphaera oryza Schlauder 1945 Yes Yes Yes Yes No2. Anopsolenia brasilianensis (Lohmann 1919) Deflandre 1952 Yes Yes No No No3. Braarudosphaera bigelowi (Gran and Braarud 1935) Deflandre 1947 Yes No No No No4. Calcidiscus leptoporus (Murray and Blackmann 1898) Loeblich and Tappan 1978 Yes Yes Yes Yes Yes5. Ceratolithus cristatus Kamptner 1950 var. cristatus Yes No Yes No NoC. cristatus var. telesmus (Norris 1965) Jordan and Young 1990 Yes No No Yes No6. Coccolithus pelagicus (Wallich 1877) Aciller 1930 f. pelagicus Yes No No No Yes7. Coronosphaera mediterranea (Lohmann 1902) Gaarder and Heimdal 1977 Yes Yes Yes Yes Yes8. C. binodata ( Kamptner 1927) Gaarder and Heimdal 1977 Yes No No No No9. Discosphaera tubifera (Murray and Blackman 1989) Ostenfeld 1900 Yes Yes No No No10. Emiliania huxleyi (Lohmann 1902) Hay and Mohler, in Hay et al. 1967 Yes Yes Yes Yes Yes11. Florisphaera profunda Okada and Honjo 1973 Yes Yes Yes Yes Yes12. Gephyrocapsa caribbeanica Boudreaux and Hay, in Hay et al. 1967 Yes Yes Yes Yes Yes13. G. ericsonii McIntyre and Be 1967 Yes Yes Yes Yes Yes14. G. ornata Heimdal, 1973 Yes No No No No15. G. muellerae Breheret 1978 type A Yes Yes Yes Yes Yes16. G. oceanica Kamptner 1943 Yes Yes Yes Yes Yes17. Gladiolithus flabellatus (Halldal and Markali 1955) Yes Yes Yes Yes Yes
Jordan and Chamberlain 199318. Helicosphaera carteri ( Wallich 1978) Kamptner 1954 Yes Yes Yes Yes Yes19. H. pavimentum Okada and McIntyre 1977 Yes No Yes No No20. Neosphaera coccolithomorpha Lecal-Schlauder 1950 Yes No No No Yes21. Oolithotus fragilis (Lohman 1912) Okada and McIntyre 1977 Yes Yes Yes Yes No22. Ponthosphaera japonica (Takayama 1967) Nishida 1971 Yes No Yes No No23. P. discopora Schiller 1925 Yes Yes Yes Yes Yes24. P. syracusana Lohmann 1902 Yes Yes No No No25. Reticulofenestra parvula (Okada and McIntyre 1977) Biekart 1989 Yes Yes Yes Yes Yes26. Reticulofenestra ‘overcalcified’ sp. Yes Yes Yes Yes Yes27. Rhabdosphaera clavigera Murray and Blackman 1989 Yes Yes Yes Yes Yes28. R. xiphos (Deflandre and Fert 1954) Norris 1984 Yes No No No No29. Scyphosphaera apstenii Lohman 1902 Yes No No No Yes30. Syracosphaera anthos (Lohman 1912) Jordan and Young 1990 Yes Yes No No No31. S. corrugis Okada and McIntyre 1977 Yes No No No No32. S. halldalii ( Weber-van Bosse, 1901) Gaarder 1970 Yes Yes No Yes Yes33. S. histrica Kamptner 1941 Yes No No No No34. S. pulchra Lohmann 1902 Yes Yes Yes Yes Yes35. Umbellosphaera irregularis Paashe, in Markali and Paashe 1955 Yes No No No No36. U. tenuis (Kamptner 1937) Paashe, in Markali and Paashe 1955 Yes Yes Yes Yes Yes37. Umbilicosphaera hulburtiana Gaarder 1970 Yes No No No No38. U. sibogae (Weber-van Bosse 1901) Gaarder 1970 Yes Yes No No No
Holococcolithophores39. Antosphaera fragaria ( Kamptner 1953) Kleijne 1991 Yes No No No No40. Calyptrolithina multipora (Gaarder, in Heimdal and Gaarder 1980) Yes No No No Yes
Norris 198541. C. wettsteinii ( Kamptner 1937) Kleijne 1991 Yes Yes Yes No No
185P. Ziveri et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 158 (2000) 175–195
Table 3 (continued )
Species Sediment Surface sedimentstrap
Bannock UM15 UM26 UM35 MT6Basin(3 km)
42. Calyptrolithophora papillifera (Halldal 1953) Yes No No No YesHeimdal, in Heimdal and Gaarder 1980
43. Calyptrosphaera oblonga Lohmann 1902 Yes Yes No Yes Yes44. Calyptrosphaera pirus Kamptner 1937 Yes No Yes No No45. Gliscolithus amitakarenae Norris 1985 No Yes No No No46. Homozysosphaera vercellii Borsetti and Cati, 1979 Yes No No No No47. S. dalmaticus ( Kamptner 1927) Loeblich and Tappan 1963 Yes No No Yes No48. S. quadriperforatus ( Kamptner 1937) Gaarder 1962 Yes Yes No Yes No49. Syracolithus confusus Kleijne 1991 Yes No No No No50. Zygosphaera hellenica Kamptner 1937 Yes No No Yes No
a The relative abundances (%) of the main coccolithophore species in the sediment trap and surface sediments are shown in Table 4.
Fig. 4. Comparison of sediment trap (ST) and surface sediment (box cores MT6, UM15, UM35 and UM26) results: (a) Annualcoccolith flux (number m−2 year−1); (b) relative abundance (%) of E. huxleyi; (c) relative abundance (%) of the major coccolithspecies excluding E. huxleyi.
186 P. Ziveri et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 158 (2000) 175–195
Table 4Estimated annual coccolith flux (coccoliths m−2 year−1) in the sediment trap (extrapolated by 926 day deployment) and surfacesediments, and relative abundances (%) of the main coccolithophore species in the sediment trap and surface sedimentsa
Coccolith species abundance (%)
Sediment trapb Box core UM15c Box core UM26d Box core UM35e Box core MT6f
C. leptoporus 1.61 <1 <1 <1 1.34D. tubifera <1 1.72 2.90 1.06 <1Reticulofenestra ‘overcalcified’ sp. <1 4.58 31.10 19.84 3.32E. huxleyi 82.22 68.19 28.10 57.14 55.52F. profunda 5.28 9.16 12.98 5.17 9.03G. flabellatus <1 1.15 2.48 <1 <1G. oceanica <1 <1 <1 <1 1.34H. carteri 3.06 <1 <1 <1 1.67N. coccolithomorpha <1 3.72 4.99 5.03 0.00Oolithothus spp. <1 <1 1.14 <1 <1R. clavigera 1.07 <1 <1 2.38 0R. xiphos <1 2.01 2.17 1.85 0S. pulchra <1 <1 2.57 1.59 <1Syracosphaera spp. 3.92 1.15 1.29 <1 1.67S. fossilis <1 2.01 1.63 <1 0U. tenuis <1 1.15 1.61 <1 1U. sibogae <1 <1 2.03 <1 1Holococcoliths <1 <1 2.41 1.32 <1
a MT6: box core located in anoxic environment.b Annual coccolith flux (number per m−2 year−1)=1.0×1010.c Annual coccolith flux (number per m−2 year−1)=1.0×1010.d Annual coccolith flux (number per m−2 year−1)=2.31×109.e Annual coccolith flux (number per m−2 year−1)=5.87×109.f Annual coccolith flux (number per m−2 year−1)=3.9×1010.
Mediterranean on living coccolithophores, and Mediterranean in 1986–1988 registered a very highseasonal variability in coccolithophore production.almost nothing is known about their spatial and
temporal distribution or their export produc- The highest production was found in February–March, largely dominated by E. huxleyi.tion to the sea floor. One previous investiga-
tion ( Knappertsbusch, 1993) in the eastern Preliminary results from the oligotrophic waters
Fig. 5. Flux record for Al (m−2 day–1) and coccolith concentration (number mg−1).
187P. Ziveri et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 158 (2000) 175–195
of the south Aegean Sea and the Straits of Cretan 4.2. Factors controlling the temporal and spatialvariability of coccolithophores, and the transferArc show that coccolithophores were the dominant
phytoplankton group in March and September processes to the deep Mediterranean waters(Balopoulos et al., 1996).
The annual coccolith flux of 1.0×1010 The particle flux in the ocean is mainly con-trolled by biological processes, occurring in thecoccoliths m−2 year−1 measured in the deep waters
of Bannock Basin is much lower than most pub- upper ocean, and by eolian and riverine input.Our work suggests that, in the central easternlished data recorded in other oceanographic set-
tings (1011–1012 coccoliths m−2 year−1; Steinmetz, Mediterranean, these two main particle fluxsources of biogenic and lithogenic origin are pri-1991; Broerse et al., 2000; Haidar et al., 2000;
Sprengel et al., 2000; Ziveri and Thunell, 2000). marily controlled by coccolithophore productionand Saharan dust input, respectively.This result remains low even after we correct the
value for the trap efficiency of 23% calculated from Satellite-derived, 7 year average (1979–1985),monthly surface pigment concentrations were esti-the 230Th flux (Rutten et al., in press).
Thoracosphaera fluxes parallel the mass flux (r= mated using data collected by the Coastal ZoneColor Scanner (CZCS) (courtesy of the Joint0.72). Data on thoracosphaerid fluxes in the ocean
still remain scarce. Their annual fluxes Research Centre, European Commission/European Space Agency) and show an annual varia-(2.4×107 calcispheres m−2 year−1) in Bannock
Basin are higher than those recorded in open ocean tion in primary production (Fig. 6). The pigmentconcentration record is used as a proxy for surfaceNortheast Atlantic stations at 48°N21°W, how-
ever, and are comparable to the data collected at primary productivity and is very similar from yearto year. In the eastern Mediterranean, the overall38°N21°W (Broerse et al., in press). Enhanced
calcareous cyst production can be observed in pigment concentrations are low when compared toother oceanic basins, even during the high-produc-regions and time intervals with stratified, oligotro-
phic conditions in the upper water masses (Holl tivity season, and most of the production occurs inthe period from December–March.et al., 1999; Zonneveld et al., 1999).
Fig. 6. Satellite observations: (a) Monthly averaged pigment concentrations observed for the period 1979–1985 derived from CZCSdata (solid line) at the trap location, with the gray area indicating the 1 standard deviation for each month, and the weekly compositeAVHRR sea surface temperature record at the trap location during the deployment period (dashed line). (b) Precipitation at thetrap location ( line) and desert dust mass over the Mediterranean (bars) during the deployment period. The dust data were redrawnafter Dulac et al. (1996).

188 P. Ziveri et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 158 (2000) 175–195
Sea surface temperatures (SST) were derived deposition of Saharan dust occur (compare alsoFigs. 2 and 6).from advanced very high resolution radiometer
(AVHRR) data for the study period (November The vertical stability of the water column mayplay a vital role in determining the productivity of1991–August 1994) and have been generated using
weekly composite AVHRR data for the Bannock Bannock surface waters. The breakdown of thisstability is most important in initiating the late fallBasin trap location (Fig. 6). The sea surface tem-
perature varies between 15 and 28°C at Bannock increase in plankton production. Coincident withthe changes in winds that occur in late fall, surfaceBasin, with the highest temperatures in summer
and fall (Fig. 6). The average SST in the summer waters begin to cool and evaporation is highestat this time. Nutrients are scarce in theand winter of 1994 appears to be 1°C higher than
those in 1992 and 1993. Mediterranean compared with the rest of the worldocean, because the sea’s main input comes fromMassive airborne plumes of desert dust from
Sahara are exported to the Mediterranean all year the surface waters of the Atlantic through theStrait of Gibraltar. As a consequence, the centrallong (Bergametti et al., 1989; Moulin et al.,
1997). Although dust concentrations in the eastern Mediterranean is characterized by anextremely oligotrophic regime (Berland et al., 1988;Mediterranean atmosphere and chlorophyll con-
centrations correlate negatively, there are differ- Dugdale and Wilkerson, 1988).The trap station in Bannock basin is located inences between the dry and wet dust deposition. In
the Mediterranean, wet dust deposition contributes the Ionian Sea province of Antoine et al. (1995)in which the winter bloom, as revealed by CZCS,65–80% to the total dust deposition (Molinaroli
et al., 1993). Because precipitation only occurs in coincides with the breakdown of stratification, andit is of moderate intensity because the nutrientwinter, most of the yearly dust deposition to the
sea water occurs in this period. A relationship levels are never high. Zohary and Robarts (1998)concluded that phosphorus was the primary limit-between dust input and primary production was
shown by the results of the Ironex II program, ing nutrient when other factors (such as light andgrazing) did not control microbial biomass orwhich established physiological limitation of phy-
toplankton by iron as the cause of the high- activity.The coccolith and coccosphere sinking rates werenutrient, low-chlorophyll phenomenon at eastern
equatorial Pacific Ocean (Martin et al., 1994; calculated from the time lag between associatedpigment maxima at the sea surface (Fig. 6) and theBehrenfeld et al., 1996). We are aware of the fact
that phosphorus, not iron, is the limiting nutrient coccolith/coccosphere flux maxima in the trapsamples and the vertical distance (3000 m). Thein the eastern Mediterranean ( Krom et al., 1992;
Zohary and Robarts, 1998), but the atmospheric coccolith sinking rates are very low, ranging from17 to 25 m day−1 (average time lag of 4–6 months).source can also contribute phosphorus to the
photic zone, as was shown for the western In contrast, maximum fluxes of coccospheres wererecorded mostly from January to April indicatingMediterranean (Bergametti et al., 1992). Because
E. huxleyi is an excellent competitor for phos- that the time lag between maximum primary pro-duction and maximum export flux at 3000 m waterphorus (Riegman et al., 1998), increases in the
production of this most abundant coccolithophore depth was about 1 month. Consequently, it appearsthat in the eastern Mediterranean, coccospheresspecies are expected in P-controlled systems such
as the eastern Mediterranean. sink individually with a rate of approximately100 m day−1. This settling speed concords with theOur work on particle fluxes in Bannock Basin
suggests that the productivity pattern could be the previous estimations of the sinking speed of particlesthrough the water column (Honjo et al., 1982;result of fertilization in the upper euphotic zone.
The increase in pigment concentration is observed Neuer et al., 1997). Therefore, a possible indicatorfor primary production in the sediment trap locatedwhen both wind-induced oceanographic changes
from a summer stratified upper water mass to a in the oligotrophic waters of the easternMediterranean might be the occurrence of highdeep winter mixing (Krom et al., 1992) and wet

189P. Ziveri et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 158 (2000) 175–195
coccosphere fluxes. However, the contribution of Mediterranean sapropelitic sediments has beenused to infer paleonutricline dynamicsintact coccospheres to the total coccolith flux is
relatively small in comparison to the settling flux (Castradori, 1993).Coccolith fluxes of Helicosphaera carteri andof individual coccoliths probably associated with
small macroaggregates and fecal pellets (considering Calcidiscus leptoporus show a similar seasonaltrend (r=0.79) and abundance (Fig. 3). This sim-also that grazing activity and organic matter pro-
duction is very low in the central eastern ilarity was previously observed in the Quaternarysediments from the North Atlantic and in sedimentMediterranean). The coccosphere–coccolith flux
ratio ranges from 0 to 5.8×10−3 (average trap samples from the Southern California Bightand Gulf of California (Gard, 1989; Ziveri et al.,5.1×10−4). Individual coccoliths may have been
resuspended and redeposited from the shelf into the 1995; Ziveri and Thunell, 2000). In the SouthernCalifornia Bight, the highest fluxes of the twodeep sea, through currents and winds. In contrast,
intact coccospheres disintegrate relatively rapidly species were associated with low to intermediatenutrient concentrations prior to upwelling whenafter the cell dies and are rarely found in sediments.
Consequently, the occurrence of coccospheres the total coccolithophore productivity was high.In the Gulf of California, both species tend todirectly refers to overlying surface water produc-
tivity, whereas coccoliths may also have a different decrease when the zooplankton grazing pressureincreases.origin (e.g. compare Fig. 2b and c for January–
March 1992). The Syracosphaera spp. fluxes represent all ofthe Syracosphaera species recorded in BannockBasin except S. pulchra and S. histrica, which have4.3. Flux patterns of Coccolithophore speciesa very low abundance in our samples. The fluxesof Syracosphaera spp. show a seasonal signal com-The most abundant and cosmopolitan coccoli-
thophorid species, Emiliania huxleyi, largely domi- parable to C. leptoporus and H. carteri (Fig. 3).The remaining coccolithophore species account fornates the observed assemblage of the 50
coccolithophore species recorded in the trap approximately 3.7% of the total assemblage.Umbellosphaera tenuis and U. irregularis have sea-samples (38 heterococcolith and 12 holococcolith
species; Fig. 3; Tables 3 and 4). The seasonal trend sonal trends similar to that of H. carteri (Fig. 3).Both species are typical of warm, tropical–sub-recorded in coccolith species fluxes is mainly driven
by changes in the flux of this species, a dominance tropical waters (McIntyre et al., 1970). TheRhabdospheraceae represent only a small part ofof this species already observed in the
Mediterranean surface waters by Knappertsbusch the species recorded in the present study and arepresent as D. tubifera and R. clavigera ad R.(1993) and Kleijne (1993).
The high correlation (r=0.90) between E. hux- xiphos. Okada and McIntyre (1979) have describedD. tubifera and R. clavigera as warm-water speciesleyi and the deep-dwelling species Florisphaera
profunda suggests coccolithophore production of the upper water layers. These two species havea subtropical temperature preference, of which R.throughout the water column. F. profunda is
restricted to the lower euphotic zone in low-middle clavigera (3–29°C) has a wider temperature rangethan D. tubifera (optimum 21–29°C; total rangelatitude regions (Okada and Honjo, 1973), and
changes in the F. profunda abundance through 14–30°C ). D. tubifera is more abundant in oligo-trophic water conditions, occurring outside thetime have been used as a proxy to model paleonut-
ricline dynamics (Molfino and McIntyre, 1990). upwelling areas, and has also been described inthe warmer areas of the eastern MediterraneanThe inverse relationship between the F. profunda
flux and the diatom export production shows that ( Kleijne, 1993). In our study, D. tubifera is alwaysless than 3% of the coccolith assemblage, but isF. profunda is inversely related to the intensity of
upwelling (Jordan et al., 1996; Ziveri and Thunell, remarkable in that the increase in both fluxes andrelative abundance during the last sampling year2000). The relative abundance of F. profunda
in the coccolith assemblages of eastern (1994) is associated with a low coccolith flux and

190 P. Ziveri et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 158 (2000) 175–195
in SST, being a few degrees higher than the previ- planktic foraminifers, the latter mainly as juvenileforms. The occurrence of mainly juvenile foramini-ous sampling years. Scyphosphaera apstenii,
Umbellosphaera tenuis and holococcoliths show the fers is probably due to the combination of twofactors. Firstly, extremely oligotrophic conditionssame increase during 1994.seriously hamper heterotrophic production in theeastern Mediterranean (Turley, 1997). Secondly,4.4. Comparison of sediment trap and surface
sediment results (Plates I and II) the small size of juvenile foraminifers (approxi-mately 20–60 mm) allows for easy lateral transportfrom higher production areas to the BannockThe importance of coccolith calcite in the east-
ern Mediterranean deep-sea sediments has been Basin site.The carbonate content of sediment trap samplesestablished previously (Bernard and Lecal, 1953;
Milliman and Muller, 1973). Coccolithophores are and surface sediment samples shows large regionalvariations. The carbonate content in the trapthe most abundant primary producers that are
being preserved in surface sediments in the eastern resembles that found in box core MT-6, but ismuch lower than in the other box cores. AluminumMediterranean. This is confirmed by our results
on particle fluxes in Bannock Basin, where coccol- correlates inversely with coccolith concentrations(coccoliths mg−1), suggesting the dilution of coc-iths constitute the major part of the biogenic
component. colith carbonate by terrestrial input (Fig. 5).The annual coccolith flux obtained by the sedi-The coarse-size fraction of the biogenic particle
flux, including foraminifers, pteropods, and radio- ment trap at 3000 m is comparable to the coccolithaccumulation rates of the aerobic surface sedimentlarians, has not been quantified in the trap samples.
However, during the analyses of the complete of box core UM15 and is about one-third of thecoccolith accumulation rate calculated in thefraction aliquot under both a binocular microscope
(400×) and LM, it was noted that this fraction anaerobic surface sediment of Bannock Basin(MT6) (Fig. 4). Differences with a similar orderwas consistently very low. In Bannock Basin,
coccolithophores are the dominant phytoplankton of magnitude have been obtained by the compari-son of the coccolith fluxes in the two traps in thegroup that manufactures a skeleton and, together
with calcareous dinoflagellates (mainly aerobic (this study) and anaerobic water condi-tions (Ziveri et al., 1996). The main differenceThoracosphaera heimii), are also the major contrib-
utors to the biogenic carbonate flux. In contrast, between the coccolith assemblages of sedimenttrap and surface sediment samples is the presencein the surface sediment, the >32 mm fraction con-
stitutes approximately 25% of the bulk sediment of Reticulofenestra ‘overcalcified’ sp. in the surfacesediment. Because this species has a very lowand contains about 75% biogenic carbonate
(mainly foraminifers and pteropods; Plate II ) abundance in the sediment trap, we conclude that,in combination with the same trend for the large-(Rutten et al., 2000).
There is no evidence of any carbonate dissolu- size fraction (high in surface sediments, low in thesediment trap), the long-term variability in faunaltion in the trap samples collected at 3000 m water
depth in Bannock Basin. The preservation of deli- and floral assemblages is not captured by thesediment trap, possibly due to irregular bloomscate coccoliths such as Oolithothus fragilis,
Discosphaera tubifera and Umbilicosphaera spp. (see also Rutten et al., 2000).(Roth and Berger, 1975; Schneidermann, 1977) isvery good. Perfectly preserved spines of foramini-fers were also detected, suggesting a good carbon-ate preservation. 5. Summary and conclusions
The rest of the flora consist of diatoms andsilicoflagellates. They show a flux pattern similar This work, concentrating on coccolithophores
and thoracosphaerid fluxes as the major contrib-to coccoliths (Ziveri et al., in preparation). Thefauna is poorly represented by radiolarians and utors to the biogenic carbonate flux recorded at
191P. Ziveri et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 158 (2000) 175–195
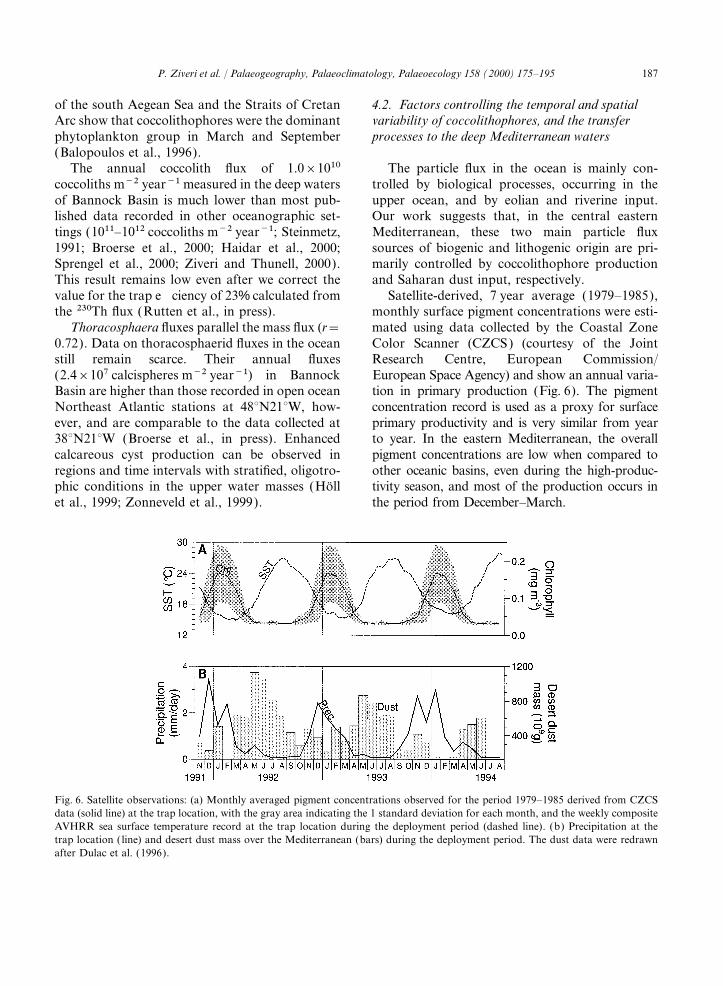
PLATE I
Sediment trap samples.
1. Assemblages of sample MT4O-1.2. Assemblages of sample MUB-1.3. Rhabdosphaera clavigera var. clavigera, rhabdolith (sample MT4O-18).4. Umbilicosphaera tenuis, umbelloliths, and Calcidiscus leptoporus, distal view of the distal shield (sample MT4O-16).5. Emiliania huxleyi, coccosphere, and Umbilicosphaera sibogae, distal view of the distal shield (sample ST2-20).6. Emiliania huxleyi and gephyrocapsid, distal view of the distal shields (sample MT40-1).7. Syracosphaera apstenii, lopadolith (sample MUB-10).8. Helicosphaera carteri, proximal view of coccolith, and Florisphaera profunda, coccolith (sample MUB-1).
192 P. Ziveri et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 158 (2000) 175–195
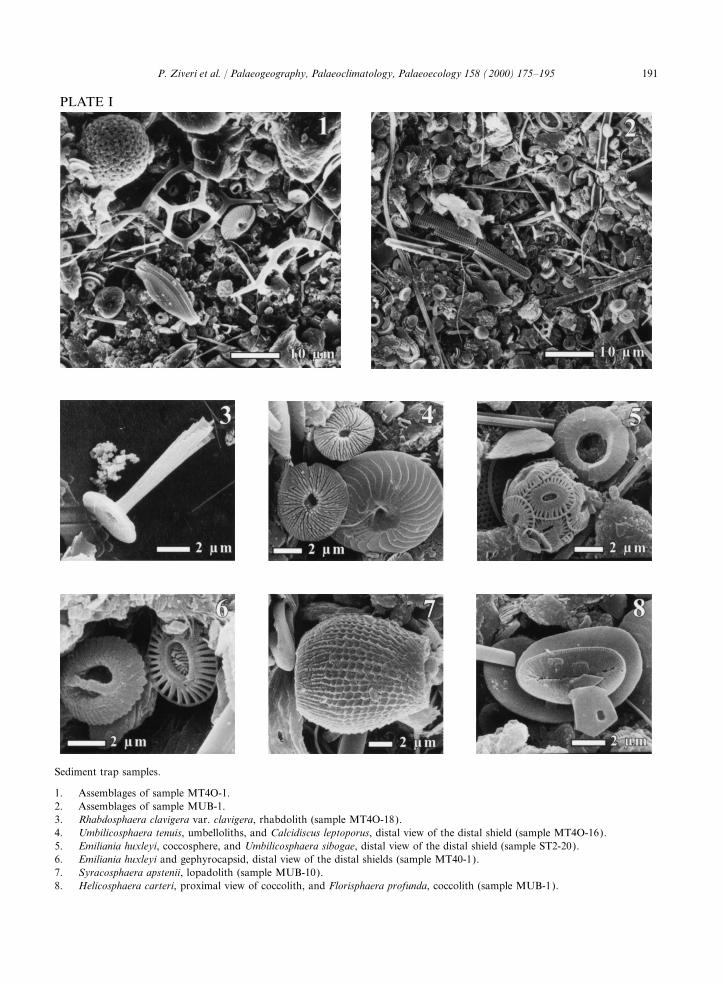
PLATE II
Surface sediment samples.
1. Calyptrosphaera pirus, areolith (sample UM15<32 mm).2. Helicosphaera carteri, distal view (sample UM35<32 mm).3. Emiliania huxleyi, distal view (sample UM26<32 mm).4. Florisphaera profunda, coccoliths (sample MT6).5. Discosphaera tubifera, rhabdolith (sample UM35).6. Syracosphaera pulchra, caneolith, distal view (sample UM35).7. Reticulofenestra, ‘overcalcified’ sp., distal view (sample UM26<32 mm).8. Reticulofenestra, ‘overcalcified’ sp., distal view (sample UM26<32 mm).9. Umbilicosphaera sibogae, distal view (sample UM15<32 mm).10. Assemblages of sample UM15<32 mm).11. Assemblages of sample UM15>32 mm).

193P. Ziveri et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 158 (2000) 175–195
3000 m in the oligotrophic waters of the central tation data from the GPCP Global CombinedPrecipitation Dataset (part of the Climatologyeastern Mediterranean Sea, has shown that:
1. A strong seasonal variation in the total mass Interdisciplinary Data Collection). This study wassupported by MARFLUX (MAST1-90022C),flux is recorded by the total coccolith and
coccosphere flux, with maximum fluxes occur- PALAEOFLUX (MAS2-CT93-0051) and SAP(MAS3-CT97-0137) European programs.ring in late winter and spring.
2. The annual coccolith flux of 1.0× This is Netherlands Sedimentary ResearchSchool (NSG) contribution 990309.1010 coccoliths m−2 year−1 measured in the
deep waters at 3000 m water depth is muchlower than most published data recorded inother oceanographic settings, even when cor- Referencesrected for the trap efficiency (23%).
3. The biogenic and lithogenic fluxes are primarily Andreae, M.O., 1986. The ocean as a source of atmosphericsulfur compounds. In: Buat-Menard, P. (Ed.), The Role ofcontrolled by coccoliths and Saharan dustAir–Sea Exchange in Geochemical Cycling. Reidel, Dor-input, respectively.drecht, pp. 331–362.
4. The calculated coccolith and coccosphere Antoine, D., Morel, A., Andre, J.-M., 1995. Algal pigment dis-settling speeds estimated from the comparison tribution and primary production in the eastern Mediterra-
nean as derived from coastal zone color scannerof maximum pigment concentration at the seaobservations. J. Geophys. Res. 100, 16193–16209.surface and maximum flux at 3000 m water
Balch, W.M., Kilpatrick, K.A., Tres, C.C., 1997. The 1991 coc-depth were 17–25 m day−1 for coccoliths andcolithophore bloom in the central North Atlantic. 1. Optical
100 m day−1 for coccospheres. properties and factors affecting their distribution. Limnol.5. Emiliania huxleyi and Florisphaera profunda, Oceanogr. 41 (8), 1669–1683.
Balopoulos, E., Theocharis, A., Price, B., Heussner, S., Souver-followed by H. carteri and C. leptoporus, aremezoglou, A., Kontoyiannis, H., Nakopoululou, C., Varna-the dominant coccolith species in sediment trapvas, S., Collins, M.B., Apostolopoulou, M., Ignatiades, L.,samples as well as surface sediments.Gotsi, O., Theodoru, A., 1996. A synthesis of preliminary
Reticulofenestra ‘overcalcified’ sp. is a major results from multidisciplinary oceanographic studies in thespecies in the surface sediments only. This South Aegean Sea and the straits of Cretan Arc. In: Second
Workshop of the Mediterranean Target Project, Crete 1996,anomaly is comparable to that observed for theExtended Abstract, 224–232.large-size (>32 mm) fraction, which is nearly
Behrenfeld, M.J., Bale, A.J., Kolber, Z.S., Aiken, J., Falkowski,absent in the sediment trap, but abundant inA., 1996. Confirmation of iron limitation of phytoplankton
surface sediments. Both might be related to photosynthesis in the equatorial Pacific Ocean. Naturelong-term differences in faunal and floral assem- 383, 508–511.
Bergametti, G., Gomes, L., Remoudaki, E., Desbois, M.,blages, possibly due to irregular blooms.Martin, D., Buat-Menard, M.P., 1989. Present transportand deposition patterns of African dusts to the north-west-ern Mediterranean. In: Leinen, M., Sarnthein, M. (Eds.),
Acknowledgements Paleoclimatology and Paleometeorology, Modern and PastPatterns of Global Atmospheric Transport, NATO ASISeries C 282., 227–252.P.J.M. van Santvoort, G. Nobbe and H. de
Bergametti, G., Remoudaki, E., Losno, R., Steiner, E., Cha-Waard are thanked for their analytical assistance.tenet, B., Buat-Menard, P., 1992. Source, transport andWe are grateful to Agostino Rizzi (University ofdeposition of atmospheric phosphorus over the Northwest-
Milan) and Saskia Kars (Vrije Universiteit ern Mediterranean. J. Atm. Chem. 14, 501–513.Amsterdam) for operating the SEM. The paper Berland, B.R., Benzhitski, A.G., Zensira, P., Burlakova, Z.P.,
Georgieva, L.V., Izmestieva, M.A., Kholodov, V.I., Maes-benefited greatly from comments by E. Erba andtrini, S.Y., 1988. Conditions hydrologiques estivales enR. Thunell. The pigment data are courtesy of theMediterranee, repartition du phytoplancton et de la matiereJoint Research Centre (European Commission/organique. Oceanol. Acta 9, 163–177.
European Space Agency). The SST data were Bernard, F., Lecal, J., 1953. Role des flagelles calcaires dans laextracted from the Weekly NCEP SST Database sedimentation actuelle en Mediterranee. Publ. 19th Geol.
Congres, Algiers 1952 (4), 11–23.at the NOAA/WRC Server Ferret and the precipi-

194 P. Ziveri et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 158 (2000) 175–195
Broerse, A.C., Ziveri, P., Van Hinte, J.E., Honjo, S., 2000. Coc- biogenic matter in the deep ocean. Deep-Sea Res. 29,colithophore export production, seasonal species composi- 609–625.tion, and coccolith-CaCO3 fluxes in the NE Atlantic (34°N Jordan, R., Meixhun, Z., Eglinton, G., Weaver, P.P.E., 1996.21°W and 48°N 21°W ). In: Ganssen, G., Wefer, G. (Eds.), Coccolith and alkenone stratigraphy and paleoceanographyDeep-Sea Research II, Topical Studies in Oceanograph. Par- at an upwelling site off NW Africa (ODP 658 C) during theticle Fluxes and its Preservation in Deep Sea Sediments last 130 000 years. In: Moguivelesky, A., Whatley, R. (Eds.),47., 9–11. Microfossil and Oceanic Environments. Aberystwyth Press,
Brown, C.W., Yoder, J.A., 1994. Distribution pattern of cocco- University of Wales, pp. 111–130.lithophorid blooms in the Western North Atlantic Ocean. Kleijne, A., 1993. Morphology, taxonomy and distribution ofContinent. Shelf Res. 14, 175–197. extant coccolithophorids (calcareous nannoplankton),
Castradori, D., 1993. Calcareous nannofossils and the origin of Ph.D. dissertation, Vrije Universiteit Amsterdam, 321 pp.eastern Mediterranean sapropels. Paleoceanography 8, Knappertsbusch, M., 1993. Geographic distribution of living459–471. and Holocene coccolithophores in the Mediterranean Sea.
Charlson, R.J., Lovelock, J.E., Andrea, M.O., Warren, S.G., Mar. Micropaleontol. 21, 219–247.1987. Oceanic phytoplankton, atmospheric sulfur, cloud Krom, M.D., Brenner, N.K., Neori, A., Gordon, L.I., 1992.albedo and climate. Nature 326, 655–661. Nutrient dynamics and new production in a warm-core eddy
Dachs, J., Bayona, J.M., Fowler, S.W., Miquel, J.C., Albaiges, from the Eastern Mediterranean Sea. Deep-Sea Res. 39,J., 1986. Vertical fluxes of polycyclic aromatic hydrocarbons 467–480.and organochlorine compounds in the western Alboran Sea Li, W.K.W., Yacobi, Y.Z., Wood, A.M., 1993. Ultraphy-(southwestern Mediterranean). Mar. Chem. 52, 75–86. toplankton in the eastern Mediterranean Sea: Towards
Dugdale, R.C., Wilkerson, F.P., 1988. Nutrient sources and deriving phytoplankton biomass from flow cytometric meas-primary production in the Eastern Mediterranean. Oceanol. urements of abundance, fluorescence and light scatter. Mar.Acta 9, 179–184. Ecol. Prog. Ser. 102, 79–97.
Dulac, F., Moulin, C., Lambert, C.E., Guillard, F., Poitou, J., Lototoskaya, A., Ziveri, P., Ganssen, G.M., Van Hinte, J.E.,Guelle, W., Quetel, C.R., Schneider, X., Ezat, U., 1996. 1998. Calcareous nanoflora response to Termination II atQuantitative remote sensing of African dust transport to the 45°N, 25°W (northeast Atlantic). Mar. Micropaleontol.Mediterranean. In: Guerzoni, S., Chester, R. (Eds.), The
34, 47–70.Impact of Desert Dust across the Mediterranean. Environm.
McIntyre, A., Be, A.W.H., Roche, M.B., 1970. Modern PacificSci. Technol. Libr. Vol. 11. Kluwer Academic, Dordrecht,
Coccolithophorida: A paleontological thermometer. Trans.pp. 25–49.
NY Acad. Sci. 32, 720–731.Gard, G., 1989. Variation in coccolith assemblages during the
Martin, J.H., Coale, K.H., Johnson, K.S., Fitzwater, S.E.,last glacial cycle in the high and mid-latitude Atlantic andGordon, R.M., Tanner, S.J., Hunter, C.N., Elrod, V.A.,Indian Oceans. In: Crux, J.A., van Heck, S.E. (Eds.), Nan-Nowicki, J.L., Coley, T.L., Barber, R.T., Lindley, S.,nofossils and their applications, proceedings of the Interna-Watson, A.J., Van Scoy, K., Law, C.S., Liddicoat, M.I.,tional Nannofossil Association conference, London, 1987.Ling, R., Stanton, T., Stockel, J., Collins, C., Anderson, A.,Br. Micropaleontol. Soc. Ser.. Ellis Horwood, pp. 108–121.Bidigare, R., Ondrusek, M., Latasa, M., Millero, F.J., Lee,Haidar, A.T., Thierstein, H.R., Deuser, W.G., 2000. CalcareousK., Yao, W., Zhang, J.Z., Friederich, G., Sakamoto, C.,phytoplankton standing stocks, fluxes and accumulation inChavez, F., Buck, K., Kolber, Z., Greene, R., Falkowski,Holocene sediments at Bermuda (N. Atlantic). In: Ganssen,P., Chisholm, S.W., Hoge, F., Swift, R., Yungel, J., Turner,G., Wefer, G. (Eds.), Deep-Sea Research II, Topical StudiesS., Nightingale, P., Hatton, A., Liss, P., Tindale, N.W.,in Oceanograph. Particle Fluxes and its Preservation in1994. Testing the iron hypothesis in ecosystems of the equa-Deep Sea Sediments 47, 9–11.torial Pacific Ocean. Nature 371, 123–129.Heussner, S., Monaco, A., 1996. The MTP sediment trap
Matrai, P.A., Keller, M.D., 1993. Dimethylsulfide in a large-experiments: highlights on particle fluxes through the eutro-scale coccolithophore bloom in the Gulf of Maine. Conti-phic and oligotrophic systems of the Mediterranean Sea. In:nent. Shelf Res. 13, 831–843.Second Workshop of the Mediterranean Target Project,
Milliman, J.D., Muller, J., 1973. Precipitation and lithificationCrete 1996, Extended Abstract, 233–239.of magnesian calcite in the deep-sea sediments of the easternHoll, C., Karwath, B., Ruhlemann, C., Zonneveld, K.A.F., Wil-Mediterranean sea. Sedimentology 20, 29–45.lems, H., 1999. Palaeoenvironmental information gained
Milliman, J.D., 1993. Production and accumulation of calciumfrom calcareous dinoflagellates: the late Quaternary easterncarbonate in the ocean: budget of a nonsteady state. Globaland western tropical Atlantic Ocean in comparison. Palaeo-Biogeochem. Cycles 7, 927–957.geogr. Palaeoclimatol. Palaeoecol. 146, 147–164.
Miquel, J.C., Fowler, S.W., La-Rosa, J., Buat-Menard, P.,Holligan, P.M., Viollier, M., Harbour, D.S., Camus, P., Cham-1994. Dynamics of the downward flux of particles andpagne-Philippe, M., 1983. Satellite and ship studies of coc-carbon in the open northwestern Mediterranean Sea. Deep-colithophore production along a continental shelf edge.Sea Res. I 41, 243–261.Nature 304, 339–342.
Honjo, S., Manganini, S.J., Cole, J.J., 1982. Sedimentation of Molfino, B., McIntyre, A., 1990. Precessional forcing of nutri-

195P. Ziveri et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 158 (2000) 175–195
cline dynamics in the equatorial Atlantic. Science 249, Steinmetz, J.C., 1991. Calcareous nannoplankton biocoenosis:Sediment trap studies in the Equatorial Atlantic Central766–769.Pacific and Panama Basin. Ocean Biocoenosis Series No. 1.Molinaroli, E., Guerzoni, S., Rampazzo, G., 1993. Contribu-Woods Hole Oceanographic Institution, 85 pp.tion of Saharan dust to the Central Mediterranean Basin.
Tennant, D.A., Baker, E.T., 1992. A fast, high-precision splitterGeol. Soc. Am. Spec. Pap. 284, 303–312.for particle suspensions. Mar. Geol. 108, 247–252.Moulin, C., Lambert, C.E., Dulac, F., Dayan, U., 1997. Control
Turley, C.M., 1997. In: The Changing Mediterranean Sea — Aof atmospheric export of dust from North Africa by theSensitive Ecosystem? Progress in Oceanography of theNorth Atlantic Oscillation. Nature 387, 691–694.Mediterranean Sea. European Commission Marine ScienceNeuer, S., Ratmeyer, V., Davenport, R., Fischer, G., Wefer,and Technology Programme, Rome, pp. 65–66.G., 1997. Deep water particle flux in the Canary Island
Van Santvoort, P.J.M., De Lange, G.J., Thomson, J., Cussen,region: seasonal trends in relation to long-term satelliteH., Wilson, T.R.S., Krom, M.D., Strohle, K., 1996. Activederived pigment data and lateral sources. Deep-Sea Res. 44,post-depositional oxidation of the most recent sapropel (S1)1451–1466.in sediments of the eastern Mediterranean Sea. Geochim.Okada, H., Honjo, S., 1973. The distribution of oceanic cocco-Cosmochim. Acta 60, 4007–4024.lithophorids in the Pacific. Deep-Sea Res. 20, 355–374.
Westbroek, P., Brown, C.W., Van Bleijswijk, J., Brownlee, C.,Okada, H., McIntyre, A., 1979. Seasonal distribution of modernBrummer, G.J., Conte, M., Egge, J., Fernandez, E., Jordan,coccolithophores in the Western North Atlantic Ocean.R., Knappertsbusch, M., Stefels, J., Veldhuis, M., Van derMar. Biol. 54, 319–328.Wal, P., Young, J., 1993. A model system approach to bio-Riegman, R., Stolte, W., Noordeloos, A.A.M., 1998. A modellogical climate forcing: the example of Emiliania huxleyi.System Approach to Biological Forcing: the Example ofGlobal Planet. Change 8, 27–46.Emiliania huxleyi. Final Report Subproject (b): Physiology
Yacobi, Y.Z., Zohary, T., Kress, N., Hecht, A., Robarts, R.D.,NIOZ-Rapport 1998-8. Netherlands Institute for SeaWaiser, M., Wood, A.M., Li, W.K.W., 1995. ChlorophyllResearch.distribution throughout the southeastern Mediterranean inRobarts, R.D., Waiser, M.J., Yacobi, Y.Z., 1996. Bacterialrelation to the physical structure of the water mass. J. Mar.
abundance, biomass, and production in relation to phyto-Syst. 6, 179–189.
plankton biomass in the Levantine Basin of the Southeast-Ziveri, P., Grandi, C., Stefanetti, A., 1995. Biogenic fluxes in
ern Mediterranean Sea. Mar. Ecol. Prog. Ser. 137, 273–281.Bannock Basin: first results from a sediment trap study
Roth, P.H., Berger, W.H., 1975. Distribution and dissolution (November 1991–May 1992). Rend. Fis. Acc. Lincei 9, 6,of coccoliths in the South and central Pacific. Cushman 131–145.Found. Foraminiferal Res. 13, 87–113. Ziveri, P., DeLange, G.J., Rossignol-Strick, M., Schrader, H.,
Rutten, A., De Lange, G.J., Ziveri, P., Thomson, J., Van Sant- Thomson, J., 1996. Biogechemical Cycles in the Easternvoort, P., Colley, S., Corselli, C., 2000. Recent terrestrial Mediterranean; Actuo (Seasonal/annual ) Variations. 2ndand carbonate fluxes in the pelagic eastern Mediterranean, Workshop of Mediterranean Target Project.a comparison between sediment trap and surface sediment. Ziveri, P, Thunell, R.C., 2000. Coccolithophore export pro-Palaeogeogr. Palaeoclimatol. Palaeoecol. 158, 197–213. duction in Guaymas Basin, Gulf of California: response to(this issue). climate forcing. In: Ganssen, G., Wefer, G. (Eds.), Deep-
Schneidermann, N., 1977. Selective dissolution of recent coccol- Sea Research II, Topical Studies in Oceanograph. Particleiths in the Atlantic Ocean. In: Ramsay, A.T.S. (Ed.), Oce- Fluxes and its Preservation in Deep Sea Sediments 47, 9–11.anic Micropaleontology Vol. 2. Academic Press, London, Zohary, T., Robarts, R.D., 1998. Experimental study of micro-pp. 1009–1053. bial P limitation in the eastern Mediterranean. Limnol.
Scientific Staff of Cruise Bannock 1984-12, 1985 1985. Gypsum Oceanogr. 43, 387–395.precipitation from cold brines in an anoxic basin in the Zonneveld, K.A.F., Holl, C., Janofske, D., Karwath, B., Kern-eastern Mediterranean. Nature 314, 152–154. topf, B., Ruhlemann, C., Willems, H., 1999. Calcareous
Sprengel, C., Baumann, K.-H., Neuer, S., 2000. Seasonal and dinoflagellate cysts as palaeo-environmental tools. In: Gans-interannual variations of coccolithophore fluxes and species sen, G., Wefer, G. (Eds.), Use of Proxies in Paleoceanog-compositions in sediment traps north of Gran Canaria. Mar. raphy: Examples from the South Atlantic. Springer, Berlin,
pp. 145–164.Micropaleontol. 39.
Related Documents










