Cancer Therapeutics Insights Preclinical Evaluation of Transcriptional Targeting Strategy for Human Hepatocellular Carcinoma in an Orthotopic Xenograft Mouse Model Kian Chuan Sia 1 , Hung Huynh 1 , Alexander Yaw Fui Chung 2 , London Lucien Peng Jin Ooi 2 , Kiat Hon Lim 3 , Kam Man Hui 1,4,6,7 , and Paula Yeng Po Lam 1,5,7 Abstract Gene regulation of many key cell-cycle players in S-, G 2 phase, and mitosis results from transcriptional repression in their respective promoter regions during the G 0 and G 1 phases of cell cycle. Within these promoter regions are phylogenetically conserved sequences known as the cell-cycle–dependent element (CDE) and cell- cycle genes homology regions (CHR) sites. Thus, we hypothesize that transcriptional regulation of cell-cycle regulation via the CDE/CHR region together with liver-specific apolipoprotein E (apoE)-hAAT promoter could bring about a selective transgene expression in proliferating human hepatocellular carcinoma. We show that the newly generated vector AH-6CC-L2C could mediate hepatocyte-targeted luciferase gene expression in tumor cells and freshly isolated short-term hepatocellular carcinoma cultures from patient biopsy. In contrast, normal murine and human hepatocytes infected with AH-6CC-L2C expressed minimal or low luciferase activities. In the presence of prodrug 5-fluorocytosine (5-FC), AH-6CC-L2C effectively suppressed the growth of orthotopic hepatocellular carcinoma patient-derived xenograft mouse model via the expression of yeast cytosine deaminase (yCD) that converts 5-FC to anticancer metabolite 5-fluoruracil. More importantly, we show that combination treatment of AH-6CC-L2C with an EZH2 inhibitor, DZNep, that targets EpCAM- positive hepatocellular carcinoma, can bring about a greater therapeutic efficacy compared with a single treatment of virus or inhibitor. Our study showed that targeting proliferating human hepatocellular carcinoma cells through the transcriptional control of therapeutic gene could represent a feasible approach against hepatocellular carcinoma. Mol Cancer Ther; 12(8); 1651–64. Ó2013 AACR. Introduction Hepatocellular carcinoma (HCC) is the third leading cause of death from cancer in the world (1). Although curative treatments are available for patients with hepato- cellular carcinoma such as liver transplantation, surgical resection, and radiofrequency ablation (2), these treatment regimens are not always possible due to the size and location of the tumors. Even after liver resection, tumor recurrence remains high due to intrahepatic metastases via the portal veins or metachronous multicentric hepatocar- cinogenesis (3). Currently, the most effective medical treat- ment for advanced hepatocellular carcinoma is with the antiangiogenic drug sorafenib. Despite the success of sor- afenib to serve as the only approved medical drug for treat- ing advanced hepatocellular carcinoma, only 2.3 months improvement in median overall survival was observed in Asian hepatocellular carcinoma patients (4). Therefore, treatment options for hepatocellular carcinoma remained universally dismal and require a concerted effort from research and innovative medical interventions. Viral gene therapy has emerged as a potential alterna- tive treatment modality for cancer treatment as evidenced by the significant number of reported clinical trials in recent years (5). Diverse strategies and a plethora of cell type–specific promoters have been used to achieve tar- geted cancer gene therapy. Recent advances in molecular biologic techniques such as cDNA microarray have unra- veled differentially expressed genes in human hepatocel- lular carcinoma versus normal controls (6, 7). Among the differentially expressed gene is a prominent overlap of cell-cycle genes including the Aurora kinase family members A and B (AURKA and AURKB, respectively), cyclin A2 (CCNA2), cyclin B1 (CCNB1), proliferating cell nuclear antigen, and polo-like kinase 1. Interestingly, all of these genes are regulated by phylogenetically conserved Authors' Affiliations: 1 Division of Cellular and Molecular Research, National Cancer Center; Departments of 2 General Surgery and 3 Pathology, Singapore General Hospital; Departments of 4 Biochemistry and 5 Physiol- ogy, Yong Loo Lin School of Medicine, National University of Singapore; 6 Institute of Molecular and Cell Biology, A STAR; and 7 Division of Cancer and Stem Cell Biology, Duke-National University of Singapore (NUS) Graduate Medical School, Singapore, Singapore Note: Supplementary data for this article are available at Molecular Cancer Therapeutics Online (http://mct.aacrjournals.org/). Corresponding Author: Paula Yeng Po Lam, Laboratory of Cancer Gene Therapy, Cellular and Molecular Research Division, Humphrey Oei Institute of Cancer Research, National Cancer Centre, 11, Hospital Drive, Singapore 169610. Phone: 65-6436-8357; Fax: 65-6226-5694; E-mail: [email protected] doi: 10.1158/1535-7163.MCT-13-0056 Ó2013 American Association for Cancer Research. Molecular Cancer Therapeutics www.aacrjournals.org 1651 on May 25, 2016. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from Published OnlineFirst May 29, 2013; DOI: 10.1158/1535-7163.MCT-13-0056

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Cancer Therapeutics Insights
Preclinical Evaluation of Transcriptional Targeting Strategyfor Human Hepatocellular Carcinoma in an OrthotopicXenograft Mouse Model
Kian Chuan Sia1, Hung Huynh1, Alexander Yaw Fui Chung2, London Lucien Peng Jin Ooi2, Kiat Hon Lim3,Kam Man Hui1,4,6,7, and Paula Yeng Po Lam1,5,7
AbstractGene regulation of many key cell-cycle players in S-, G2 phase, and mitosis results from transcriptional
repression in their respective promoter regionsduring theG0 andG1phases of cell cycle.Within thesepromoter
regions are phylogenetically conserved sequences known as the cell-cycle–dependent element (CDE) and cell-
cycle genes homology regions (CHR) sites. Thus, we hypothesize that transcriptional regulation of cell-cycle
regulation via the CDE/CHR region together with liver-specific apolipoprotein E (apoE)-hAAT promoter
could bring about a selective transgene expression in proliferating human hepatocellular carcinoma.We show
that the newly generated vector AH-6CC-L2C could mediate hepatocyte-targeted luciferase gene expression
in tumor cells and freshly isolated short-term hepatocellular carcinoma cultures from patient biopsy. In
contrast, normal murine and human hepatocytes infected with AH-6CC-L2C expressed minimal or low
luciferase activities. In the presence of prodrug 5-fluorocytosine (5-FC), AH-6CC-L2C effectively suppressed
the growth of orthotopic hepatocellular carcinoma patient-derived xenograft mouse model via the expression
of yeast cytosine deaminase (yCD) that converts 5-FC to anticancermetabolite 5-fluoruracil. More importantly,
we show that combination treatment of AH-6CC-L2C with an EZH2 inhibitor, DZNep, that targets EpCAM-
positive hepatocellular carcinoma, can bring about a greater therapeutic efficacy compared with a single
treatment of virus or inhibitor. Our study showed that targeting proliferating humanhepatocellular carcinoma
cells through the transcriptional control of therapeutic gene could represent a feasible approach against
hepatocellular carcinoma. Mol Cancer Ther; 12(8); 1651–64. �2013 AACR.
IntroductionHepatocellular carcinoma (HCC) is the third leading
cause of death from cancer in the world (1). Althoughcurative treatments are available for patients with hepato-cellular carcinoma such as liver transplantation, surgicalresection, and radiofrequency ablation (2), these treatmentregimens are not always possible due to the size andlocation of the tumors. Even after liver resection, tumorrecurrence remains high due to intrahepaticmetastases viathe portal veins or metachronous multicentric hepatocar-
cinogenesis (3). Currently, themost effectivemedical treat-ment for advanced hepatocellular carcinoma is with theantiangiogenic drug sorafenib. Despite the success of sor-afenib to serve as the only approvedmedical drug for treat-ing advanced hepatocellular carcinoma, only 2.3 monthsimprovement in median overall survival was observed inAsian hepatocellular carcinoma patients (4). Therefore,treatment options for hepatocellular carcinoma remaineduniversally dismal and require a concerted effort fromresearch and innovative medical interventions.
Viral gene therapy has emerged as a potential alterna-tive treatmentmodality for cancer treatment as evidencedby the significant number of reported clinical trials inrecent years (5). Diverse strategies and a plethora of celltype–specific promoters have been used to achieve tar-geted cancer gene therapy. Recent advances in molecularbiologic techniques such as cDNAmicroarray have unra-veled differentially expressed genes in human hepatocel-lular carcinoma versus normal controls (6, 7). Amongthe differentially expressed gene is a prominent overlapof cell-cycle genes including the Aurora kinase familymembers A and B (AURKA and AURKB, respectively),cyclin A2 (CCNA2), cyclin B1 (CCNB1), proliferating cellnuclear antigen, andpolo-like kinase 1. Interestingly, all ofthese genes are regulated by phylogenetically conserved
Authors' Affiliations: 1Division of Cellular and Molecular Research,National Cancer Center; Departments of 2General Surgery and 3Pathology,Singapore General Hospital; Departments of 4Biochemistry and 5Physiol-ogy, Yong Loo Lin School of Medicine, National University of Singapore;6Institute of Molecular and Cell Biology, A�STAR; and 7Division of Cancerand Stem Cell Biology, Duke-National University of Singapore (NUS)Graduate Medical School, Singapore, Singapore
Note: Supplementary data for this article are available at Molecular CancerTherapeutics Online (http://mct.aacrjournals.org/).
Corresponding Author: Paula Yeng Po Lam, Laboratory of Cancer GeneTherapy, Cellular andMolecular Research Division, Humphrey Oei Instituteof Cancer Research, National Cancer Centre, 11, Hospital Drive, Singapore169610. Phone: 65-6436-8357; Fax: 65-6226-5694; E-mail:[email protected]
doi: 10.1158/1535-7163.MCT-13-0056
�2013 American Association for Cancer Research.
MolecularCancer
Therapeutics
www.aacrjournals.org 1651
on May 25, 2016. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst May 29, 2013; DOI: 10.1158/1535-7163.MCT-13-0056

sequences known as the cell-cycle–dependent element(CDE) and cell cycle genes homology regions (CHR) sitesin their promoters (8). Thus, cell-cycle-dependent trans-gene regulation is achieved through the binding of tran-scriptional repressors, suchas cell-cycle–dependent factor1 (CDF-1) and cyclin A CHR-binding factor (CHF), to theCDEand/orCHRelementswithin the cyclinA2promoter(9, 10). Depending on the phase of the cell cycle, theMMBand DREAM complexes could regulate cell-cycle–depen-dent gene transcription through the CHR element ofmouse and human cyclin B2 promoter (11). Furthermore,the transitional switch from MMB to DREAM bound toCHR sites is dependent on the activation status of p53,which subsequently regulates downstream transcription-al targets (12). Mutation of either CDE or CHR or bothelements leads to a substantial deregulation of cell-cyclecontrol (8, 13). Multimerization of CDE/CHR elementswithin the cyclin A2 promoter could also improve thekinetics of gene regulation (13). On the basis of thisconcept, we have previously generated a herpes simplexvirus type I (HSV-1) amplicon viral vector whereby thechimeric gene consisting of nuclear transcription factor Ysubunit A (NF-YA) fuses with the DNA-binding domainof Gal4 and is expressed under the liver-specific hybrid4�ApoE/hAAT promoter (14). This strategy is indepen-dent of exogenous stimulation for gene expression regu-lation; activation of the transgene expression is triggeredby the proliferation status of the host cells. Thus, inproliferating liver cells, the binding of Gal4/NF-YAfusion proteins to Gal4-binding sites cloned upstream ofthe minimal cyclin A2 promoter will transactivate thepromoter, as measured by the luciferase reporter geneactivity. In nondividing or quiescent cells, the binding ofthe Gal4/NF-YA proteins will be prevented by repressorproteins occupying the CDE/CHR site, thus suppressingtranscriptional activation of the reporter gene.
As aproof-of-concept that these vectors holdpromise asnovel viral vectors against hepatocellular carcinoma, thereporter gene is substituted with a bicistronic cassetteencoding the yeast cytosine deaminase gene (yCD) fusedin-framewith the luciferase reporter gene. TheCDE/CHRregulatory motifs within the minimal cyclin A2 promoterare alsomultimerized to improve the kinetics of cell-cycleregulation. The yCD is a well-characterized suicide genethat converts nontoxic antifungal agent 5-fluorocytosine(5-FC) to cytotoxic 5-fluorouracil (5-FU; ref. 15). The toxicmetabolites could freely diffuse to surrounding uninfect-ed cells to exert additional bystander killing effect (16). Inview of future clinical application, the vectors were stud-ied in hepatocellular carcinomapatient-derived xenograftmouse models and freshly isolated human hepatocellularcarcinoma samples with matched controls. Recently,Chiba and colleagues have also shown that 5-FU–resistanthepatocellular carcinoma cells as a result of continuous 5-FU treatment can be efficiently abolished by an EZH2inhibitor called 3-Deazaneplanocin A (DZNep; ref. 17).DZNep is a pharmacologic inhibitor of the enhancer ofzeste homolog 2 (EZH2; ref. 18). EZH2 is a histonemethyl-
transferase that plays an important regulatory role in theself-renewal and differentiation of murine hepatic stemcells (19) and cancer stem cells (20). In a recent study,DZNepwas shown todiminish the self-renewal capabilityof hepatocellular carcinoma, as indicated by the reducedEpCAM-positive hepatocellular carcinoma cells (17).Interestingly, the biologic effects of DZNep on hepatocel-lular carcinoma are distinct from those of 5-FU and thuswe postulate that the combination treatment of our vec-tors with DZNepwould bring about a greater therapeuticefficacy than a single treatment.
Materials and MethodsCell lines and tissue specimens
A human hepatocellular carcinoma cell line (PLC/PRF/5) and a human lung adenocarcinoma cell line(A549) were obtained from the American Type CultureCollection (ATCC). A human hepatocellular carcinomacell line (HuH-7) was obtained from the Japanese Collec-tion of Research Bioresources (JCRB cell bank). HumanDGli36 glioma cells (also known as Gli36qEGFR) werekindly provided by M. Sena-Esteves (University of Mas-sachusettsMedical School,Worcester,MA).Africangreenmonkey kidney (2–2) cells, used for vector packaging,werederived fromVero cells that constitutively expressedthe HSV-1 ICP27 proteins (kindly provided by R.M. San-dri-Goldin, University of California, Irvine, CA). All cellsweremaintained at 37�C in a 5%CO2–95% air atmosphereand cultured in Dulbecco’s modified Eagle medium sup-plemented with 10% FBS (Hyclone Laboratories), peni-cillin (100 U/mL; Life Technologies), streptomycin (100mg/mL; Life Technologies), and 2 mmol/L L-glutamine(Life Technologies). The DGli36 cells were further supple-mented with 1 mg/mL puromycin (Invivogen), whereasthe 2–2 and PLC/PRF/5-DsRed2 (PLC/PRF/5 cell stablyexpressed DsRed2 protein) cells were cultured in thepresence of 500 mg/mL of Geneticin (Life Technologies).The immortalized untransformed human neonatal livercell line (NeHepLxHT) was kindly provided by Lee CG(National Cancer Centre, Singapore). Human primaryhepatocellular carcinoma and its adjacent matched nor-mal liver tissues were obtained from the SingHealthTissue Repository following approval from the Sin-gHealthCentralized Institutional ReviewBoard, andwithpatient informed consent. Preparation of short-term cul-ture of hepatocellular carcinoma tumor or matched nor-mal liver specimens can be found in SupplementaryMaterials and Methods. All cell lines except 2–2 used inthis study were verified by using short tandem repeatanalysis by Genetica DNA laboratories, Inc. NeHepLxHTcell line authentication was conducted by ATCC.
HSV-1 amplicon plasmid DNA constructsThe generation of pAH-6CC-L2C plasmid DNA con-
struct was constructed from pApoE/hAAT-cc-Luc (14),pC8-6CC-Luc (13), and pHGCX-L2C (21) that were previ-ously constructed in our laboratory. 6CC promoter wasfirst PCR from pC8-6CC-Luc using the forward primer
Sia et al.
Mol Cancer Ther; 12(8) August 2013 Molecular Cancer Therapeutics1652
on May 25, 2016. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst May 29, 2013; DOI: 10.1158/1535-7163.MCT-13-0056

50 –CTGTCCTCCGTCTAGAATAGTCGCGGGATACandreverse primer 50-GATATAGAATTCAGACCACGCAG-GGCCGAGG with the introduction of XbaI and EcoRIsites. The PCR product of 6CC was then used to replacethe minimal cyclin A2 promoter of pApoE/hAAT-cc-Lucusing the same restriction sites to generate pApoE/hAAT-6CC-Luc. Finally, luciferase foot and mouth disease virus2A-yeast cytosine deaminase (Luc2ACD) cassette wasreleased from the pHGCX-L2C construct and substitutedwith Luc gene in pApoE/hAAT-6CC-Luc using the NotIand PacI restriction site to generate pApoE/hAAT-6CC-L2C. This construct is later renamed as pAH-6CC-L2C.Subsequent helper virus-free packaging methods aredescribed in Supplementary Materials and Methods.
Luciferase reporter gene assayHepatocellular carcinoma cells (HuH-7 and PLC/PRF/
5) or non–hepatocellular carcinoma cells (DGli36 andA549) were seeded at 3 � 105 cells in 6-well plate beforebeing transfected with 3 mg of pAH-6CC-L2C plasmidusing 8 mL of Lipofectamine (Life Technologies) and 5 mLof Plus reagent (Life Technologies) according to manu-facturer’s instructions. Completed culture medium with10% FBS was added to cells after transfection to maintainits proliferation status, whereas fresh culture mediumsupplementedwith 0.2% FBS and 70 mmol/L of lovastatin(Merck)was added to cells to induceG1 arrest for 48 hoursbefore subjecting to luciferase assay. For single-cell sus-pensions isolated from tumors or matched normal liver,the cells were seeded at 1 � 105 cells in 24-well plate andinfected with AH-6CC-L2C at the appropriate multiplic-ity of infection (MOI) for 24 hours before subjecting toluciferase assays.All cellswere harvested and lysed in 120mL lysis buffer (50mmol/LTris-Cl, 150mmol/LNaCl, 1%Triton X-100). Luciferase activity and protein concentra-tion were measured as described previously (14).
Determination of transfection and infectionefficiencyIn vitro transfection efficiency of pAH-6CC-L2C and in
vivo tumor infectivity of AH-6CC-L2C were determinedon the basis of the quantification of the percentage ofenhanced green fluorescence proteins (eGFP)-positivecells determined using FACS Calibur (Becton Dickinson).All generated data were analyzed using FlowJo software(version 7.6.4; TreeStar Inc.). In vitro AH-6CC-L2C infec-tivity in 24-well plate was determined by counting thetotal eGFP-positive cells per well using Eclipse TE300inverted fluorescence microscope (Nikon).
Cell viability assayCell viabilitywas assayed using the Cell CountingKit-8
(CCK-8; Dojindo Laboratories). A total of 5 � 103 cellswere seeded in 96-well plate with 100 mL complete medi-um per well. After incubation for 24 hours at 37�C, 5%CO2, these cells were infected with AH-6CC-L2C at MOIof 1 for 6 hours and replenished with fresh completeculture medium or fresh complete culture medium con-taining2mmol/L5-FC (Sigma-Aldrich) and/or10mmol/L
DZNep (Sigma-Aldrich). These cells were then incubatedat 37�C, 5% CO2 for another 72 hours before cell viabilityassays were carried out. To conduct an assay for cellviability, 10 mL of CCK-8 solution was added to each welland the plate was incubated for another 1 to 2 hours at37�C, 5%CO2 depending on the cells. The absorbancewasfinally read at 450 nm with a reference at 650 nm using aVictor 3V plate reader (PerkinElmer Life Sciences).
Establishment of PLC/PRF/5-DsRed2 tumorxenografts andprimary patient-derived tumormodelin SCID mice
PLC/PRF/5-DsRed2 tumor xenografts were estab-lished in the left lateral liver lobe of severe combinedimmunodeficient (SCID) mice as previously described(21). HCC 26-1004 tumors were derived, with informedconsent, from patients undergoing liver resection andhistology analysis with confirmed diagnosis of hepato-cellular carcinoma (22). Samples for xenografting weretaken from morphologically viable and non-necrotictumor regions. hepatocellular carcinoma diagnoses weresubsequently verified in all cases by our pathologists.Detailed procedure for the establishment of subcutane-ous HCC 26-1004 tumor xenograft in SCID mice wasdescribed previously (21). Cell-cycle analysis is describ-ed in Supplementary Materials and Methods. OrthotopicHCC 26-1004 tumor was established using the subcuta-neous HCC 26-1004 tumor. When a subcutaneous HCC26-1004 tumor size reached about 1,000 mm3, the tumorwas harvested and cut into 1 to 2 mm3 tumor cubes.Using 8-0 absorbable suture (Johnson & Johnson), thetumor cubes were orthotopically implanted into the leftlateral liver lobe of 6-week-old female immunodeficientSCID mice (Animal Resource Centre).
Animal worksAll animal experiments were carried out according
to the guidelines and protocols approved by theSingHealth Institutional Animal Care andUseCommittee(Singapore). After 4 to 6 days of HCC 26-1004 or RCC 09-1210C tumor implantation, 3–6 � 106 transduction units(TU) of AH-6CC-L2C in 100 ml Hank’s balanced saltsolution (HBSS) were intratumorally injected. This wasfollowedby intraperitoneal injectionof 5-FCat 500 to 1,000mg/kg doses, twice a day at 6-hour interval. At variousdesignated time points, tumor growth was monitoredusing calipers, and the volumes were calculated usingthe formula V (volume)¼ p/6� [L� (W)2], where L is thelength and W is the width. After acquiring the measure-ments, mice were subjected to noninvasive biolumines-cence imaging using the Xenogen IVIS Lumina (CaliperLife Sciences) system as previously described (21). Allimage analyses to quantitate the bioluminescence signalswere conducted using Living Image software version 3.0(Caliper Life Sciences) where regions of interest (ROI)were drawn at sites of vector injection. Background signalcorrections were conducted with an ROI drawn over theregions on mice where no vector had been administered.
Transcriptional Targeting Strategy for Human HCC
www.aacrjournals.org Mol Cancer Ther; 12(8) August 2013 1653
on May 25, 2016. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst May 29, 2013; DOI: 10.1158/1535-7163.MCT-13-0056

Statistical analysisData are presented throughout this study as means �
SEM. Student unpaired t test was used for comparing thestatistical significance for study involving two groups.One-way ANOVA followed by the Bonferroni multiplecomparisons test were used for comparing the statisticalsignificance formore than twogroups. Two-wayANOVAwas used for comparing the statistical significance formore than two factors.P< 0.05was considered statisticallysignificant.
ResultsAH-6CC-L2C mediates preferential transgeneexpression in proliferating orthotopic hepatocellularcarcinoma tumors compared with normal livers
Previous studies from our laboratory have shown that abasic HSV-1 amplicon vector engineered with cell-cycle–regulatory elements could confer luciferase activities in aproliferation-dependent manner (14). In this study, addi-tional modifications were made to this vector whichincluded the use of (i) multimerized CDE/CHR regula-tory motifs within the minimal cyclin A2 promoter toimprove the kinetics of cell-cycle regulation and (ii) abicistronic 2A-mediated therapeutic and reporter geneexpression cassette for assessing the preclinical therapeu-tic efficacy while providing real-time monitoring of thevector activities. The newly generated vector is denoted aspAH-6CC-L2C in its amplicon plasmid form (Fig. 1A) andAH-6CC-L2C when it has been packaged into ampliconviral vectors.
To evaluate the cell-cycle–dependent gene expressionproperty of the newly generated vector, PLC/PRF/5hepatocellular carcinoma cells were either transfectedwith pAH-6CC-L2C plasmids or infected with AH-6CC-L2C amplicon viral vectors at a MOI equivalent to1. In both cases, luciferase activities were significantlyenhanced in proliferating cells versus G1-arrested cellpopulation (5.9-foldunder transfectionand4.2-foldunderinfection conditions; Supplementary Fig. S1A and S1B,respectively). Flow cytometric analysis confirmed theproliferation and G1 arrest status of the cells (Supplemen-tary Fig. S1C).
We next generated an orthotopic PLC/PRF/5-DsRed2hepatocellular carcinoma tumor model to study the func-tionality of viruses inmalignant hepatocellular carcinomatumors versus normal liver. Before viral vector injection,the proliferating status of the implanted hepatocellularcarcinoma tumors was confirmed by analyzing the cell-cycle profiles of cells prepared from the tumor andnormalliver in representative animal. The results showed thatcells originated fromanorthotopic tumor region consistedof 9.7-fold higher fractions of S-phase compared withnormal liver tissues that consisted of about 95% of qui-escent cells in the G1 state (Supplementary Fig. S2A).Equal viral doses were subsequently intrahepaticallyinjected into normal mice or mice bearing orthotopichepatocellular carcinoma tumors. Luciferase activitieswere measured at 24 hours after viral injection. As shown
in Supplementary Fig. S2B, luciferase activities were sig-nificantly enhanced in liver regions that corresponded tothe site of implanted tumors, as indicated by the redcoloredfluorescent proteins from tumors (SupplementaryFig. S2C; left). This was further confirmed using theluciferase assays on lysates prepared from the corre-sponding PLC/PRF/5-DsRed2 tumors and normal livers(Supplementary Fig. S2C; right).
The activities of AH-6CC-L2C in proliferating versuspostmitotic hepatocytes were further confirmed in prima-ry patient-derivedHCC26-1004 xenograftmousemodels.The tumor was derived from a patient diagnosed withhepatocellular carcinoma and should be a better repre-sentation of the human hepatocellular carcinoma archi-tecture because the tumors, stromal fibroblast cells, andinflammatory cells are retained (22).Rapidproliferationofthe tumor cells in these xenografts was further confirmedby the enhanced endogenous cyclin A expression (Fig.1B). It is therefore not surprising that high levels ofluciferase activitiesweredetected in the tumors comparedwith normal liver tissueswhen both regions were injectedwith the same amount ofAH-6CC-L2C (Fig. 1C andD). Toexclude tissue difference, AH-6CC-L2C-transduced pri-mary cells derived from HCC 26-1004 tumors were com-pared with those in NeHepLxHT, a human telomerasereverse transcriptase immortalized human liver cell line.At 24 hours after infection, the number of eGFP-positivecells was similar in HCC 26-1004 versus NeHepLxHT(Fig. 1E). Luciferase activities mediated by AH-6CC-L2C were significantly higher in HCC 26-1004 comparedwith NeHepLxHT (Fig. 1F) as well as after normalizingagainst the number of infected eGFP-positive cells (Fig.1G). Taken together, our results showed that AH-6CC-L2C could mediate elevated transgene expression in pro-liferating hepatocellular carcinoma in preference to nor-mal mouse liver and immortalized normal humanhepatocytes.
AH-6CC-L2C confers hepatocellular carcinomaspecificity using hepatocellular carcinoma patient-derived xenograft mouse models
Given thatmost proliferating tumor cellswoulddisplayelevated Ki-67 and cell-cycle proteins, we next determinewhether AH-6CC-L2C could function preferentially inpatient-derived hepatocellular carcinoma versus non-hepatocellular carcinoma xenograft mouse models. Thiswas first conducted in vitrowith cell line–derived cells bycomparing the pAH-6CC-L2C–transfected hepatocellularcarcinoma cells to non–hepatocellular carcinoma cellswhere all of these cells consisted of high percentage ofproliferating cells in S-phase (Supplementary Fig. S3).Using A549 as a more stringent representative of non-hepatocellular carcinoma lines, HuH-7 and PLC/PRF/5cells exhibited a corresponding 6.8- to 12.3-fold increase intransgene expression (Fig. 2A). One-way ANOVA anal-ysis indicated that the differences between all cell lineswere statistically significant (P < 0.0001) where the Bon-ferroni multiple comparisons test also confirmed that
Sia et al.
Mol Cancer Ther; 12(8) August 2013 Molecular Cancer Therapeutics1654
on May 25, 2016. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst May 29, 2013; DOI: 10.1158/1535-7163.MCT-13-0056
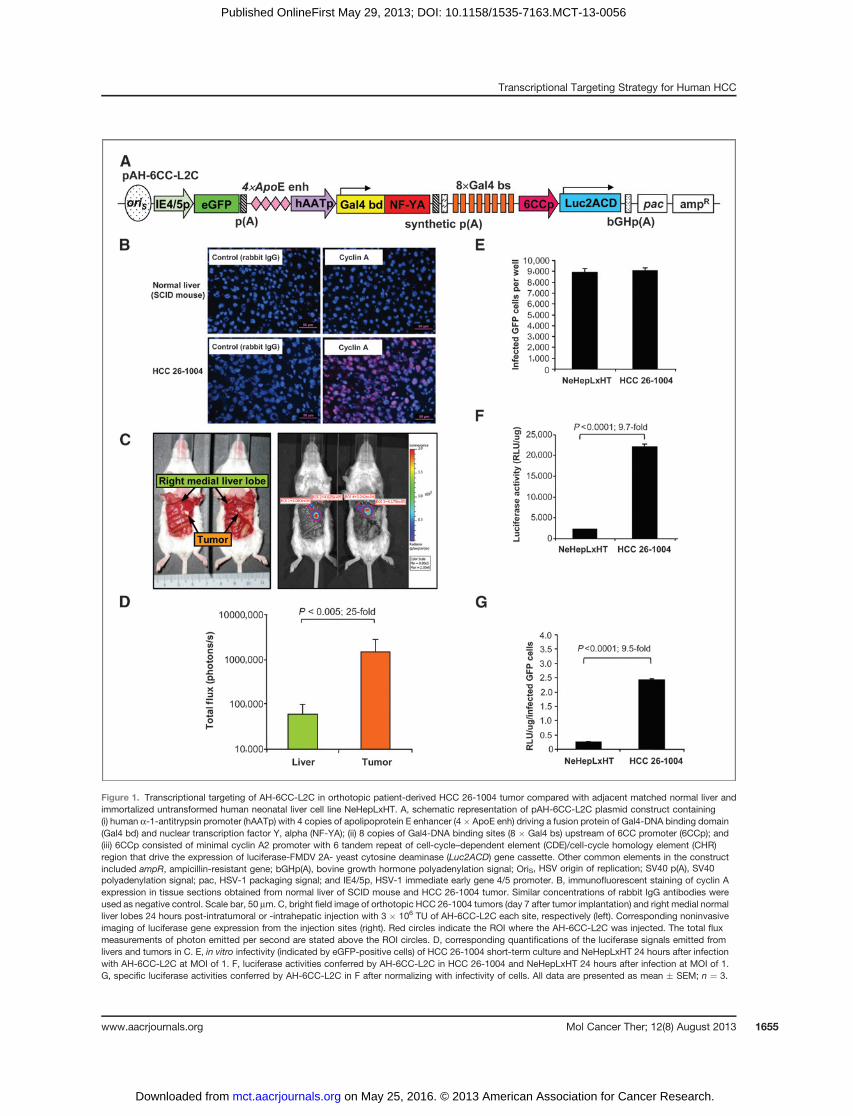
Figure 1. Transcriptional targeting of AH-6CC-L2C in orthotopic patient-derived HCC 26-1004 tumor compared with adjacent matched normal liver andimmortalized untransformed human neonatal liver cell line NeHepLxHT. A, schematic representation of pAH-6CC-L2C plasmid construct containing(i) human a-1-antitrypsin promoter (hAATp) with 4 copies of apolipoprotein E enhancer (4� ApoE enh) driving a fusion protein of Gal4-DNA binding domain(Gal4 bd) and nuclear transcription factor Y, alpha (NF-YA); (ii) 8 copies of Gal4-DNA binding sites (8 � Gal4 bs) upstream of 6CC promoter (6CCp); and(iii) 6CCp consisted of minimal cyclin A2 promoter with 6 tandem repeat of cell-cycle–dependent element (CDE)/cell-cycle homology element (CHR)region that drive the expression of luciferase-FMDV 2A- yeast cytosine deaminase (Luc2ACD) gene cassette. Other common elements in the constructincluded ampR, ampicillin-resistant gene; bGHp(A), bovine growth hormone polyadenylation signal; OriS, HSV origin of replication; SV40 p(A), SV40polyadenylation signal; pac, HSV-1 packaging signal; and IE4/5p, HSV-1 immediate early gene 4/5 promoter. B, immunofluorescent staining of cyclin Aexpression in tissue sections obtained from normal liver of SCID mouse and HCC 26-1004 tumor. Similar concentrations of rabbit IgG antibodies wereused as negative control. Scale bar, 50 mm. C, bright field image of orthotopic HCC 26-1004 tumors (day 7 after tumor implantation) and right medial normalliver lobes 24 hours post-intratumoral or -intrahepatic injection with 3 � 106 TU of AH-6CC-L2C each site, respectively (left). Corresponding noninvasiveimaging of luciferase gene expression from the injection sites (right). Red circles indicate the ROI where the AH-6CC-L2C was injected. The total fluxmeasurements of photon emitted per second are stated above the ROI circles. D, corresponding quantifications of the luciferase signals emitted fromlivers and tumors in C. E, in vitro infectivity (indicated by eGFP-positive cells) of HCC 26-1004 short-term culture and NeHepLxHT 24 hours after infectionwith AH-6CC-L2C at MOI of 1. F, luciferase activities conferred by AH-6CC-L2C in HCC 26-1004 and NeHepLxHT 24 hours after infection at MOI of 1.G, specific luciferase activities conferred by AH-6CC-L2C in F after normalizing with infectivity of cells. All data are presented as mean � SEM; n ¼ 3.
Transcriptional Targeting Strategy for Human HCC
www.aacrjournals.org Mol Cancer Ther; 12(8) August 2013 1655
on May 25, 2016. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst May 29, 2013; DOI: 10.1158/1535-7163.MCT-13-0056

Figure 2. Hepatocellular carcinoma specificity of AH-6CC-L2C in cell line-derived or patient-derived human hepatocellular carcinoma and non–hepatocellularcarcinoma cells. A, luciferase activities conferred by pAH-6CC-L2C in cell line–derived hepatocellular carcinoma (HuH-7 and PLC/PRF/5) and non–hepatocellular carcinoma (DGli36 andA549) cells at 48 hours after transfection. Total relative luciferase units (RLU)were normalizedwith total protein contentsand transfection efficiency (indicated by eGFP-positive cells). B, luciferase gene expression in in vitro cell cultures of patient-derived HCC 26-1004and RCC 09-1210C (non–hepatocellular carcinoma) 24 hours after infection with AH-6CC-L2C atMOI of 0.5, 1.0, 1.5, and 2.0. Total RLUwas normalizedwithtotal protein contents. C, comparison of in vitro transduction efficiency of HCC 26-1004 (infected with AH-6CC-L2C at MOI of 0.5) with RCC 09-1210C(infectedwith AH-6CC-L2C atMOI of 0.5, 1.0, and 1.5) as indicated by eGFP-positive cells. D, specific luciferase activities conferred by AH-6CC-L2C in HCC26-1004 and RCC 09-1210C after normalizing with total protein content and transduction efficiency. All in vitro data are presented as mean � SEM;n ¼ 3. E, noninvasive imaging of luciferase gene expression conferred by AH-6CC-L2C in HCC 26-1004 and RCC 09-1210C subcutaneous primarytumor xenograft 24hours after injectionwith 3�106TUand6�106TUofAH-6CC-L2C, respectively. Redcircles indicate theROIwhere theAH-6CC-L2Cwasinjected intratumorally. The total flux measurements of photon emitted per second are stated above the ROI circles. F, corresponding quantifications of theluciferase signal emitted from tumors in E. All in vivo data are presented as mean � SEM; n ¼ 5. G, immunofluorescent staining for hAAT andcyclin A expression in HCC 26-1004 and RCC 09-1210C tumor sections. Similar concentrations of rabbit IgG antibodies were used as negative control.Scale bar, 50 mm.
Sia et al.
Mol Cancer Ther; 12(8) August 2013 Molecular Cancer Therapeutics1656
on May 25, 2016. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst May 29, 2013; DOI: 10.1158/1535-7163.MCT-13-0056

luciferase expression in non–hepatocellular carcinomacells is significantly different than in hepatocellular car-cinoma cells.The cell-type specificity of AH-6CC-L2C was further
confirmed in primary patient-derived HCC 26-1004 andRCC 09-1210Cmice xenografts. Single-cell suspensions ofHCC 26-1004 and RCC 09-1210C tumors were prepared.Given that these two tumor types may express differentreceptors that resulted in differential HSV-1 infectivity,we tested a range of MOI of vectors on primary cellsderived from RCC 09-1210C against HCC 26-1004cells infected at a constantMOI of 0.5 (Fig. 2B). The resultsindicated that twice as much of the vector was requiredto infect RCC 09-1210C cells to achieve similar infectivitywith HCC 26-1004 (Fig. 2C). However, after normalizingthe luciferase activities against infected cells, the func-tional expression of luciferase reporter activities remain-ed constant regardless of MOIs, and AH-6CC-L2C stillmediate significantly increased transgene expression forabout 4.7-fold higher in hepatocellular carcinoma 26-1004compared with RCC 09-1210C (one-way ANOVA: P <0.0001; Fig. 2D).On the basis of the in vitro virus optimization study,
twice the amount of AH-6CC-L2C viral vectors was sub-sequently injected subcutaneously into RCC 09-1210Ctumor-bearing mice. The human origin and proliferationstatus of bothHCC26-1004 (Supplementary Fig. S4A) andRCC 09-1210C (Supplementary Fig. S4B) primary patient-derived xenografts were validated through positive stain-ing against human nuclei and a corresponding high Ki-67–labeling index.As represented in Fig. 2E, the luciferaseactivitieswere 4.6-foldhigher inHCC26-1004 than inRCC09-1210C (Fig. 2F) despite the fact that mice bearing renalcell carcinoma tumors were injected with twice theamount of viruses. Primary cell suspensions were pre-pared from tumors in representative animal and thesimilar infectivity of AH-6CC-L2C in the two tumor typeswas further confirmed by flow cytometric analysis (datanot shown). In an attempt to address the differentialluciferase expression, the tumor sections were stainedagainst human a-1-antitrypsin (hAAT) and cyclin A (seeSupplementary Materials and Methods for details). Bothtumor types expressed high levels of endogenous cyclinAprotein but only hepatocellular carcinoma tumorsexpressed hAAT proteins (Fig. 2G). Taken together, theseresults consistently showed that AH-6CC-L2C mediateenhanced transgene expression in proliferating tumorsexpressing elevated levels of hAAT proteins.
Intratumoral administration of AH-6CC-L2Cinhibits growth of patient-derived humanhepatocellular carcinoma xenograft mouse modelsNext, we evaluate the therapeutic efficacy of AH-6CC-
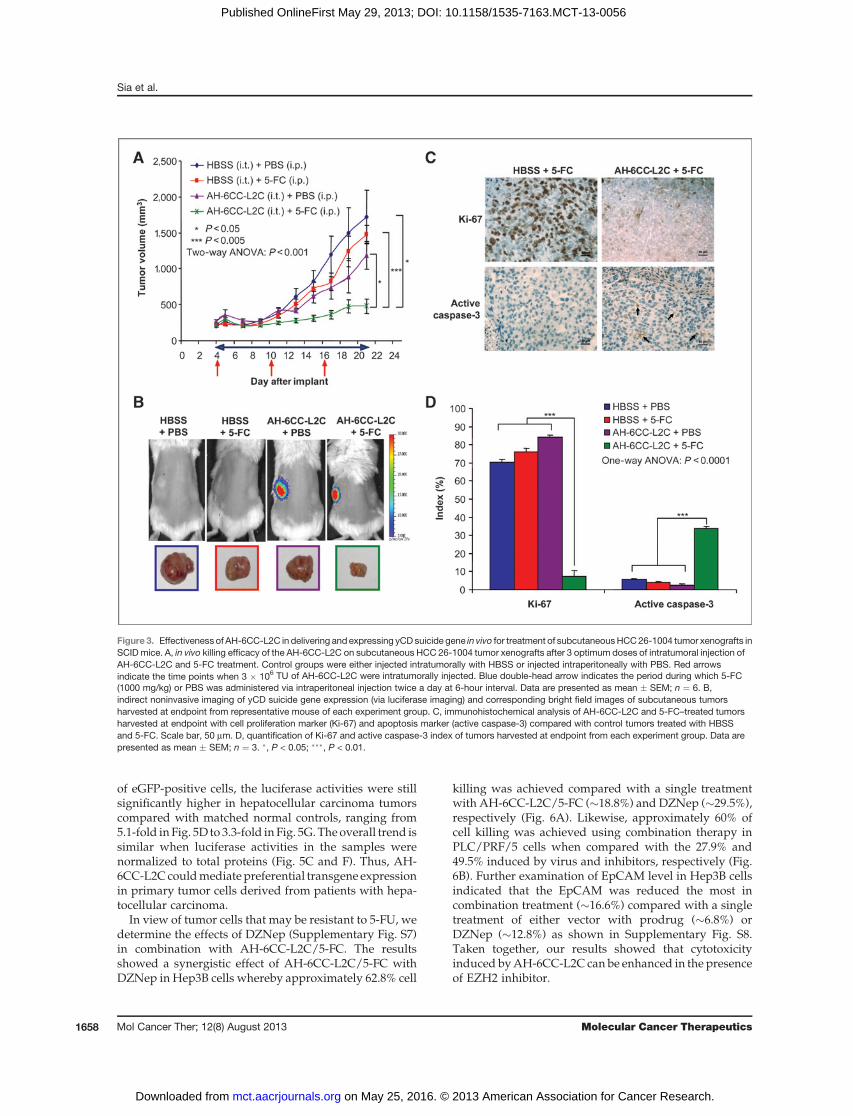
L2C viral vectors in patient-derived subcutaneous andorthotopic hepatocellular carcinoma tumor xenograftmodels. The functionality of the yCD suicide gene in thepresence of its prodrug (5-FC) was confirmed in vitro inPLC/PRF/5 cells (Supplementary Fig. S5). Because AH-
6CC-L2C vector comprised of yCD gene fused in-framewith the luciferase reporter, we were able to monitor theestimated levels of therapeutic proteins produced in vivo,and the mice were injected with a booster shot of vectorswhen minimal luciferase activities were detected (Sup-plementary Fig. S6A; red arrows). Similar volumeofHBSSto those of injected vectors was used as control. Asshown in Fig. 3A, significant tumor growth inhibitionwas observed in AH-6CC-L2C/5-FC–treated hepatocel-lular carcinoma patient–derived subcutaneous xenograftcomparedwithHBSS/5-FC control group (3.1-fold).Mea-surement of tumor volumes from each group of animalscorresponded to the size of the tumors in the representa-tive animal sacrificed at the endpoint of study (Fig. 3B).Furthermore, animals treated with AH-6CC-L2C/5-FCexhibited marked reduction in the levels of Ki-67, whichwas accompanied by elevated caspase-3 expression (Fig.3C andD). TheKi-67 index ofAH-6CC-L2C/5-FC–treatedtumors were about 10-fold lowerwhen comparedwith allother control groups (Fig. 3D).
To confirm the functionality of these viruses in thelysosome-enriched liver microenvironment, similarexperiment was carried out in orthotopically implantedHCC 26-1004 animals injected with AH-6CC-L2C viralvectors as outlined in Fig. 4A. The results showed thatthere was a significant decrease in tumor volume andweight in animals treated with AH-6CC-L2C/5-FC whencompared with HBSS/5-FC group (Fig. 4B) at the end-point of the study. Expression of yCD was monitoredindirectly through the reporter activities with boostershots of vectors administered at the indicated times (Sup-plementary Fig. S6B). Corresponding luciferase activitiesand tumors in the representative animals were showninFig. 4C. Similar topreviousfindings, high levels ofKi-67were observed in control animals but not in AH-6CC-L2C/5-FC group (Fig. 4D and E). In contrast, activecaspase-3 was detected only in animals treated withAH-6CC-L2C/5-FC (Fig. 4D and E). Taken together, wehave clearly shown that AH-6CC-L2C could effectivelysuppress the growth of patient-derived human hepato-cellular carcinoma in subcutaneous and orthotopic xeno-graft mouse models.
Combination treatment of AH-6CC-L2C and DZNepexerts synergistic cytotoxic effects againsthepatocellular carcinoma
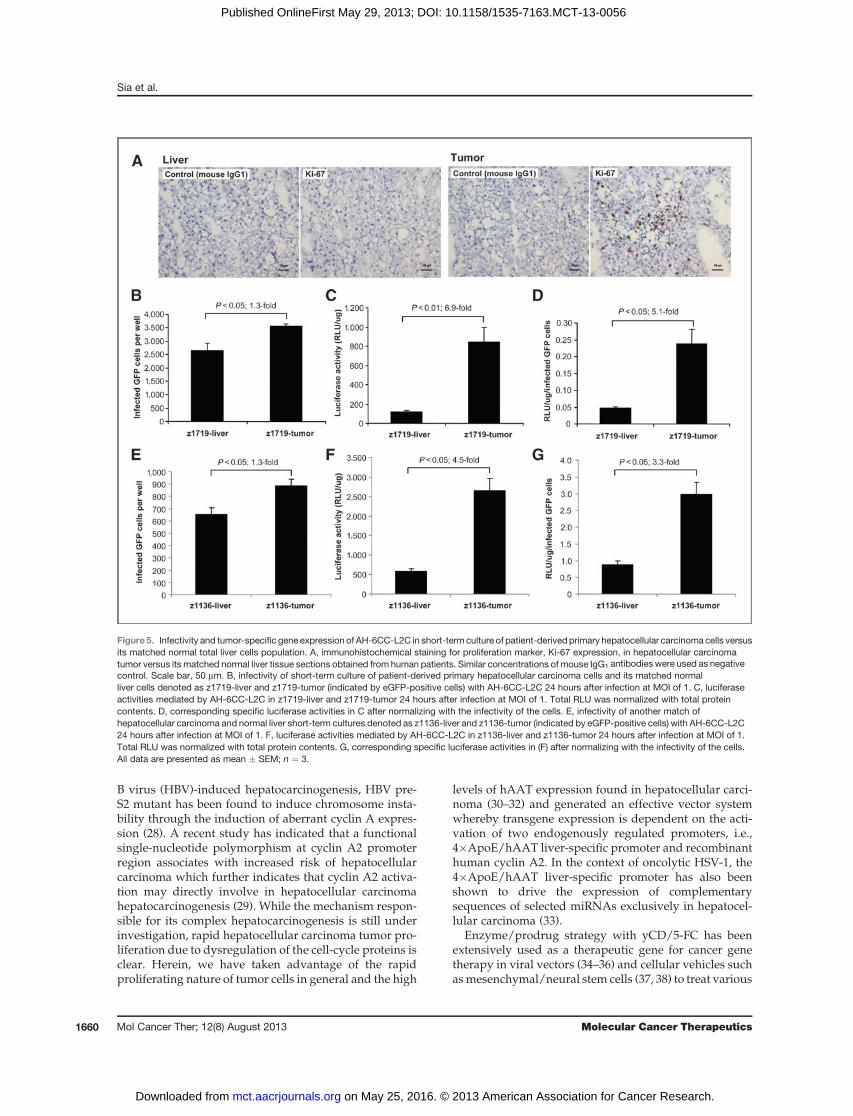
For clinical relevance, we asked whether AH-6CC-L2Cwould enhance transgene expression in short-term cul-tures consisting of patient-derived hepatocellular carci-noma compared with the correspondingmatched normaladjacent liver cells. Representative liver and tumor sec-tions showed increased proliferation as characterized bythe Ki-67 labeling index (Fig. 5A), which may be used asprognostic marker for hepatocellular carcinoma associat-ed with poor survival (23). In two representative short-term cultures, AH-6CC-L2C infected hepatocellularcarcinoma slightly better than matched normal control(Fig. 5B andE).However, after normalizing to the number
Transcriptional Targeting Strategy for Human HCC
www.aacrjournals.org Mol Cancer Ther; 12(8) August 2013 1657
on May 25, 2016. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst May 29, 2013; DOI: 10.1158/1535-7163.MCT-13-0056
of eGFP-positive cells, the luciferase activities were stillsignificantly higher in hepatocellular carcinoma tumorscompared with matched normal controls, ranging from5.1-fold in Fig. 5D to 3.3-fold in Fig. 5G.Theoverall trend issimilar when luciferase activities in the samples werenormalized to total proteins (Fig. 5C and F). Thus, AH-6CC-L2Ccouldmediatepreferential transgene expressionin primary tumor cells derived from patients with hepa-tocellular carcinoma.
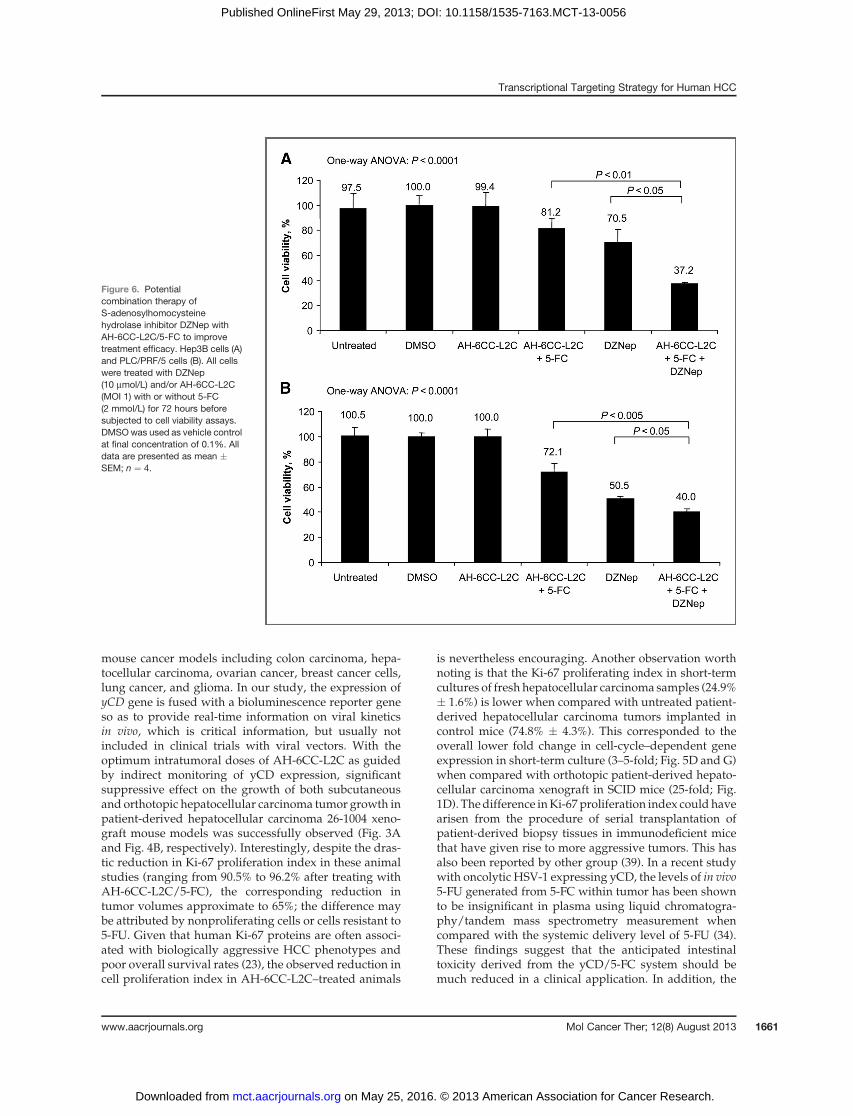
In view of tumor cells that may be resistant to 5-FU, wedetermine the effects of DZNep (Supplementary Fig. S7)in combination with AH-6CC-L2C/5-FC. The resultsshowed a synergistic effect of AH-6CC-L2C/5-FC withDZNep in Hep3B cells whereby approximately 62.8% cell
killing was achieved compared with a single treatmentwith AH-6CC-L2C/5-FC (�18.8%) and DZNep (�29.5%),respectively (Fig. 6A). Likewise, approximately 60% ofcell killing was achieved using combination therapy inPLC/PRF/5 cells when compared with the 27.9% and49.5% induced by virus and inhibitors, respectively (Fig.6B). Further examination of EpCAM level in Hep3B cellsindicated that the EpCAM was reduced the most incombination treatment (�16.6%) compared with a singletreatment of either vector with prodrug (�6.8%) orDZNep (�12.8%) as shown in Supplementary Fig. S8.Taken together, our results showed that cytotoxicityinduced byAH-6CC-L2C can be enhanced in the presenceof EZH2 inhibitor.
Figure 3. Effectiveness of AH-6CC-L2C in delivering and expressing yCD suicide gene in vivo for treatment of subcutaneousHCC26-1004 tumor xenografts inSCID mice. A, in vivo killing efficacy of the AH-6CC-L2C on subcutaneous HCC 26-1004 tumor xenografts after 3 optimum doses of intratumoral injection ofAH-6CC-L2C and 5-FC treatment. Control groups were either injected intratumorally with HBSS or injected intraperitoneally with PBS. Red arrowsindicate the time points when 3 � 106 TU of AH-6CC-L2C were intratumorally injected. Blue double-head arrow indicates the period during which 5-FC(1000 mg/kg) or PBS was administered via intraperitoneal injection twice a day at 6-hour interval. Data are presented as mean � SEM; n ¼ 6. B,indirect noninvasive imaging of yCD suicide gene expression (via luciferase imaging) and corresponding bright field images of subcutaneous tumorsharvested at endpoint from representative mouse of each experiment group. C, immunohistochemical analysis of AH-6CC-L2C and 5-FC–treated tumorsharvested at endpoint with cell proliferation marker (Ki-67) and apoptosis marker (active caspase-3) compared with control tumors treated with HBSSand 5-FC. Scale bar, 50 mm. D, quantification of Ki-67 and active caspase-3 index of tumors harvested at endpoint from each experiment group. Data arepresented as mean � SEM; n ¼ 3. �, P < 0.05; ���, P < 0.01.
Sia et al.
Mol Cancer Ther; 12(8) August 2013 Molecular Cancer Therapeutics1658
on May 25, 2016. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst May 29, 2013; DOI: 10.1158/1535-7163.MCT-13-0056

DiscussionOver the last decade, advanced technologies such as
DNA microarrays have enabled us to characterize theaberrant expression profiles with respect to tumor typesand/or stages in comparison with the adjacent normaltissues. For example, comparison of differentially regu-lated genes in hepatocellular carcinoma compared withnormal liver using microarray analysis has enabled theidentification of several hepatocellular carcinoma-specificpromoterswhichmaybe a valuable tool for the generationof hepatocellular carcinoma-specific viral vectors (24).Herein, we have incorporated multimerized CDE/CHRmodules within theminimal cyclin A2 promoter of a HSVamplicon vector as ameanof regulating transgene expres-sion in response to the proliferation status of the host cells.When coupled with 4�ApoE/hAAT liver-specific pro-moter, the results obtained are encouraging in that these
AH-6CC-L2C viral vectors could mediate specific trans-gene expression in hepatocellular carcinoma but not renalcell carcinoma (RCC) patient-derived xenografts despiteboth tumor types originated from patients with advancedgrade 3 tumor and expressed high Ki-67 proliferatingindex. Taken together, these data suggest that AH-6CC-L2C vectors could represent a novel therapeutic vectorplatform for hepatocellular carcinoma.
In the clinical setting, overexpression of cyclin A andcyclin E proteins have been associatedwith tumor relapseof human hepatocellular carcinoma and are independentpredictivemarkers for their recurrence and prognosis (25,26). Silencing of b-catenin gene in hepatocellular carcino-ma induces changes in the expression of cyclin A andcyclin E, suggesting that the Wnt/b-catenin signalingpathwaymaybe involved in the hepatocellular carcinomapathogenesis through cell-cycle proteins (27). In hepatitis
Figure 4. Effective tumor growth suppression mediated by AH-6CC-L2C in orthotopic HCC 26-1004 tumor xenografts in SCID mice. A, treatment timelineof orthotopic HCC 26-1004 tumors with 5-FC and 4 intratumoral doses of 3 � 106 TU of AH-6CC-L2C. Control tumors were intratumorally injectedwith HBSS. B, tumor volume (left) and tumor weight (right) measurements for AH-6CC-L2C/5-FC- or HBSS/5-FC–treated orthotopic HCC 26-1004tumors harvested at endpoint. Data are presented as mean � SEM; n ¼ 5. C, indirect noninvasive imaging of yCD suicide gene expression (via luciferaseimaging) and corresponding bright field images of orthotopic tumors harvested at endpoint from representative mouse of each experiment group. D,immunohistochemical analysis of AH-6CC-L2C- and 5-FC–treated tumors harvested at endpoint with cell proliferation marker (Ki-67) and apoptosis marker(active caspase-3) compared with control tumors treated with HBSS and 5-FC. Scale bar, 50 mm. E, quantification of Ki-67 and active caspase-3 index oftumors harvested at endpoint from each experiment group. Data are presented as mean � SEM; n ¼ 3.
Transcriptional Targeting Strategy for Human HCC
www.aacrjournals.org Mol Cancer Ther; 12(8) August 2013 1659
on May 25, 2016. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst May 29, 2013; DOI: 10.1158/1535-7163.MCT-13-0056
B virus (HBV)-induced hepatocarcinogenesis, HBV pre-S2 mutant has been found to induce chromosome insta-bility through the induction of aberrant cyclin A expres-sion (28). A recent study has indicated that a functionalsingle-nucleotide polymorphism at cyclin A2 promoterregion associates with increased risk of hepatocellularcarcinoma which further indicates that cyclin A2 activa-tion may directly involve in hepatocellular carcinomahepatocarcinogenesis (29). While the mechanism respon-sible for its complex hepatocarcinogenesis is still underinvestigation, rapid hepatocellular carcinoma tumor pro-liferation due to dysregulation of the cell-cycle proteins isclear. Herein, we have taken advantage of the rapidproliferating nature of tumor cells in general and the high
levels of hAAT expression found in hepatocellular carci-noma (30–32) and generated an effective vector systemwhereby transgene expression is dependent on the acti-vation of two endogenously regulated promoters, i.e.,4�ApoE/hAAT liver-specific promoter and recombinanthuman cyclin A2. In the context of oncolytic HSV-1, the4�ApoE/hAAT liver-specific promoter has also beenshown to drive the expression of complementarysequences of selected miRNAs exclusively in hepatocel-lular carcinoma (33).
Enzyme/prodrug strategy with yCD/5-FC has beenextensively used as a therapeutic gene for cancer genetherapy in viral vectors (34–36) and cellular vehicles suchasmesenchymal/neural stem cells (37, 38) to treat various
Figure 5. Infectivity and tumor-specific gene expression of AH-6CC-L2C in short-term culture of patient-derived primary hepatocellular carcinoma cells versusits matched normal total liver cells population. A, immunohistochemical staining for proliferation marker, Ki-67 expression, in hepatocellular carcinomatumor versus its matched normal liver tissue sections obtained from human patients. Similar concentrations of mouse IgG1 antibodies were used as negativecontrol. Scale bar, 50 mm. B, infectivity of short-term culture of patient-derived primary hepatocellular carcinoma cells and its matched normalliver cells denoted as z1719-liver and z1719-tumor (indicated by eGFP-positive cells) with AH-6CC-L2C 24 hours after infection at MOI of 1. C, luciferaseactivities mediated by AH-6CC-L2C in z1719-liver and z1719-tumor 24 hours after infection at MOI of 1. Total RLU was normalized with total proteincontents. D, corresponding specific luciferase activities in C after normalizing with the infectivity of the cells. E, infectivity of another match ofhepatocellular carcinoma and normal liver short-term cultures denoted as z1136-liver and z1136-tumor (indicated by eGFP-positive cells) with AH-6CC-L2C24 hours after infection at MOI of 1. F, luciferase activities mediated by AH-6CC-L2C in z1136-liver and z1136-tumor 24 hours after infection at MOI of 1.Total RLU was normalized with total protein contents. G, corresponding specific luciferase activities in (F) after normalizing with the infectivity of the cells.All data are presented as mean � SEM; n ¼ 3.
Sia et al.
Mol Cancer Ther; 12(8) August 2013 Molecular Cancer Therapeutics1660
on May 25, 2016. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst May 29, 2013; DOI: 10.1158/1535-7163.MCT-13-0056
mouse cancer models including colon carcinoma, hepa-tocellular carcinoma, ovarian cancer, breast cancer cells,lung cancer, and glioma. In our study, the expression ofyCD gene is fused with a bioluminescence reporter geneso as to provide real-time information on viral kineticsin vivo, which is critical information, but usually notincluded in clinical trials with viral vectors. With theoptimum intratumoral doses of AH-6CC-L2C as guidedby indirect monitoring of yCD expression, significantsuppressive effect on the growth of both subcutaneousand orthotopic hepatocellular carcinoma tumor growth inpatient-derived hepatocellular carcinoma 26-1004 xeno-graft mouse models was successfully observed (Fig. 3Aand Fig. 4B, respectively). Interestingly, despite the dras-tic reduction in Ki-67 proliferation index in these animalstudies (ranging from 90.5% to 96.2% after treating withAH-6CC-L2C/5-FC), the corresponding reduction intumor volumes approximate to 65%; the difference maybe attributed by nonproliferating cells or cells resistant to5-FU. Given that human Ki-67 proteins are often associ-ated with biologically aggressive HCC phenotypes andpoor overall survival rates (23), the observed reduction incell proliferation index in AH-6CC-L2C–treated animals
is nevertheless encouraging. Another observation worthnoting is that the Ki-67 proliferating index in short-termcultures of fresh hepatocellular carcinoma samples (24.9%� 1.6%) is lower when compared with untreated patient-derived hepatocellular carcinoma tumors implanted incontrol mice (74.8% � 4.3%). This corresponded to theoverall lower fold change in cell-cycle–dependent geneexpression in short-term culture (3–5-fold; Fig. 5D and G)when compared with orthotopic patient-derived hepato-cellular carcinoma xenograft in SCID mice (25-fold; Fig.1D). Thedifference inKi-67proliferation index couldhavearisen from the procedure of serial transplantation ofpatient-derived biopsy tissues in immunodeficient micethat have given rise to more aggressive tumors. This hasalso been reported by other group (39). In a recent studywith oncolytic HSV-1 expressing yCD, the levels of in vivo5-FU generated from 5-FC within tumor has been shownto be insignificant in plasma using liquid chromatogra-phy/tandem mass spectrometry measurement whencompared with the systemic delivery level of 5-FU (34).These findings suggest that the anticipated intestinaltoxicity derived from the yCD/5-FC system should bemuch reduced in a clinical application. In addition, the
Figure 6. Potentialcombination therapy ofS-adenosylhomocysteinehydrolase inhibitor DZNep withAH-6CC-L2C/5-FC to improvetreatment efficacy. Hep3B cells (A)and PLC/PRF/5 cells (B). All cellswere treated with DZNep(10 mmol/L) and/or AH-6CC-L2C(MOI 1) with or without 5-FC(2 mmol/L) for 72 hours beforesubjected to cell viability assays.DMSOwas used as vehicle controlat final concentration of 0.1%. Alldata are presented as mean �SEM; n ¼ 4.
Transcriptional Targeting Strategy for Human HCC
www.aacrjournals.org Mol Cancer Ther; 12(8) August 2013 1661
on May 25, 2016. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst May 29, 2013; DOI: 10.1158/1535-7163.MCT-13-0056

use of yCD/5-FC system can be further improved in thepresence of radiation as 5-FU is a potent radiosensitizer(40, 41). Alternatively, a humanized yCD has shown 3-fold increase in specific activities compared with theoriginal prototype due to an improvement in thermosta-bility (42).
For clinical relevance, we have also tested the function-ality of AH-6CC-L2C in short-term culture of fresh hepa-tocellular carcinoma samples versus its matched normaltotal liver cell population. Primary hepatocellular carci-noma and normal cells were isolated immediately aftersurgery resection. These short-term cultures did not stainagainst human fibroblast markers but in hepatocellularcarcinoma cultures were immunopositive against cyto-keratin-18, cyclin A, and hAAT (data not shown). It maybeworth noting that hepatocellular carcinoma tumors are1.3-fold more susceptible to AH-6CC-L2C infection whencompared with its matched normal liver cells (Fig. 5B andE). This differential in infectivity may be attributed to theoverexpression of glypican-3 (GPC3) which is a cell sur-face heparin sulfate proteoglycan required by the HSV-1viruses for adsorption (43). Increasing studies havereported on the use of replication-competent HSV-1against hepatocellular carcinoma (44, 45) and hepaticmetastases (46). We and others (47, 48) have shown thatreplication-defective HSV-1 amplicon viral vectors areequally efficient in delivering genes to the cells of hepaticorigin. For clinical application of AH-6CC-L2C, it may beworthwhile to study whether these vectors could prefer-entially infect GPC3-positive hepatocellular carcinomatumors when delivered intratumorally with the aid of acatheter and ultrasound imaging. Unfortunately, the clin-ical applications of these amplicon vectors are restrictedby the current technology platform for clinical scale-upmanufacturing and production.
Although our results show that AH-6CC-L2C couldinhibit hepatocellular carcinoma tumor growth efficient-ly, most treatments would be faced with problems ofchemotherapeutic drug resistance including those of 5-FU (49). Prolonged 5-FU treatment has been shown toincrease in EpCAMhigh and CD133high 5-FU–resistanthepatocellular carcinoma cells (HuH-7; ref17). Hepatocel-lular carcinoma cells expressing EpCAM and CD133proteins have been implicated to possess features char-acteristics of tumor-initiating cells (50). Interestingly,EpCAMhigh and CD133high 5-FU–resistant hepatocellularcarcinoma cells could be selectively killed byDZNep (17).Our results supported the finding in that combinedtreatment of viral vectors and DZNep significantlyenhances cell kill in Hep3B (Fig. 6A) and PLC/PRF/5(Fig. 6B) when compared with a single treatment. Weanticipated that after treatment with AH-6CC-L2C/5-FC,the residual 5-FU–resistant hepatocellular carcinomacells could increase the fractions of EpCAM expressionin hepatocellular carcinoma cells. Instead, our resultsshowed that EpCAM level was slightly reduced in hepa-tocellular carcinoma treated with AH-6CC-L2C in 5-FC(Supplementary Fig. S8) which could imply that the time
point chosen for FACS analysis was suboptimal, thus 5-FU–resistant cells hadnot been enriched.DZNephas beenreported to selectively inhibit the trimethylation of lysine27 on histoneH3 (H3K27me3) and lysine 20 on histoneH4(H4K20me3) and could induce demethylation of CpGsites on genes such as FBXO32, which subsequently leadto efficient apoptosis (51). In acute myeloid leukemia, thedecrease inH3K27me3 after DZNep treatment resulted inanupregulation of thioredoxin-binding protein 2 (TXNIP)which resulted in an overexpression of reaction oxygenspecies and led to apoptosis (52). A recent study showedthat combination treatment of DZNep and gemcitabine inpancreatic cancer cells significantly reduce EZH2 andH3K27me3 expression as well as depleted the intracellu-lar adenosine content (53). This synergist interactionresulted in the reduction of another cancer stemcell (CSC) marker–expressing cells, CD133, suggestingthat dual targeting of CSC and non-CSC could representan effective treatmentmodality in human tumors. Havingsaid this, the specificity and possible toxicity of DZNep orsimilar compound in vivo require further investigation.
In conclusion, we have shown that AH-6CC-L2Cvectors could mediate elevated transgene expression inproliferating hepatocellular carcinoma tumors in pref-erence to normal mouse or human hepatocytes. Selec-tive transgene expression in hepatocellular carcinomamay be attributed to the combined transcriptional reg-ulation of cell-cycle regulation and liver-specific pro-moters. In the presence of prodrug 5-FC, AH-6CC-L2Ceffectively suppressed the growth of orthotopic hepa-tocellular carcinoma patient-derived xenograft mousemodel. More importantly, we show that combinationtreatment of AH-6CC-L2C with an EZH2 inhibitor,DZNep, can bring about a greater therapeutic efficacycompared with a single treatment. Taken together, weshowed that targeting proliferating human hepatocel-lular carcinoma cells through transcriptional control oftherapeutic gene could represent a feasible approachagainst hepatocellular carcinoma.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: K.C. Sia, P.Y.P. LamDevelopment of methodology: K.C. Sia, K.H. LimAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.): K.C. Sia, H. Huynh, A.Y.F. Chung, L.L.P.J. Ooi,K.H. Lim, P.Y.P. LamAnalysis and interpretation of data (e.g., statistical analysis, biostatis-tics, computational analysis): K.C. Sia, P.Y.P. LamWriting, review, and/or revision of the manuscript: K.C. Sia, A.Y.F.Chung, L.L.P.J. Ooi, P.Y.P. LamAdministrative, technical, or material support (i.e., reporting or orga-nizing data, constructing databases): K.C. Sia, A.Y.F. Chung, K.M. Hui,P.Y.P. LamStudy supervision: P.Y.P. Lam
AcknowledgmentsThe authors thank Caroline Lee (National Cancer Center), Way Champ
Mah (National Cancer Center), Ivy Ho (National Cancer Center), Xin YiToh (National Cancer Center), and Jennifer P. Newman (National CancerCenter) for providing technical assistances.
Sia et al.
Mol Cancer Ther; 12(8) August 2013 Molecular Cancer Therapeutics1662
on May 25, 2016. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst May 29, 2013; DOI: 10.1158/1535-7163.MCT-13-0056

Grant SupportThis study is supported by research grant from the Singapore Ministry
of Health’s National Medical Research Council (NMRC/CG/NCC/2010;to P.Y.P. Lam).
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby marked
advertisement in accordance with 18 U.S.C. Section 1734 solely to indicatethis fact.
Received January 22, 2013; revised May 6, 2013; accepted May 20, 2013;published OnlineFirst May 29, 2013.
References1. El-Serag HB, Rudolph KL. Hepatocellular carcinoma: epidemio-
logy and molecular carcinogenesis. Gastroenterology 2007;132:2557–76.
2. Bruix J, Sherman M. Management of hepatocellular carcinoma: anupdate. Hepatology 2011;53:1020–2.
3. Poon RT, Fan ST, Ng IO, Lo CM, Liu CL, Wong J. Different risk factorsand prognosis for early and late intrahepatic recurrence after resectionof hepatocellular carcinoma. Cancer 2000;89:500–7.
4. ChengAL, KangYK,ChenZ, TsaoCJ,Qin S, KimJS, et al. Efficacy andsafety of sorafenib in patients in the Asia-Pacific region with advancedhepatocellular carcinoma: a phase III randomised, double-blind, pla-cebo-controlled trial. Lancet Oncol 2009;10:25–34.
5. PatelMR, KratzkeRA.Oncolytic virus therapy for cancer: the first waveof translational clinical trials. Transl Res 2013;161:355–64..
6. Chen X, Cheung ST, So S, Fan ST, Barry C, Higgins J, et al. Geneexpression patterns in human liver cancers. Mol Biol Cell 2002;13:1929–39.
7. Lee JS, Chu IS, Heo J, Calvisi DF, Sun Z, Roskams T, et al. Classi-fication and prediction of survival in hepatocellular carcinoma bygene expression profiling. Hepatology 2004;40:667–76.
8. Muller GA, Engeland K. The central role of CDE/CHR promoter ele-ments in the regulation of cell cycle-dependent gene transcription.FEBS J 2010;277:877–93.
9. Liu N, Lucibello FC, Korner K,Wolfraim LA, Zwicker J,Muller R CDF-1,a novel E2F-unrelated factor, interacts with cell cycle-regulatedrepressor elements in multiple promoters. Nucleic Acids Res 1997;25:4915–20.
10. Philips A, Chambeyron S, Lamb N, Vie A, Blanchard JM. CHF: a novelfactor binding to cyclin A CHR corepressor element. Oncogene1999;18:6222–32.
11. Muller GA, Quaas M, SchumannM, Krause E, Padi M, Fischer M, et al.The CHR promoter element controls cell cycle-dependent gene tran-scription and binds the DREAM and MMB complexes. Nucleic AcidsRes 2012;40:1561–78.
12. Quaas M, Muller GA, Engeland K. p53 can repress transcription of cellcycle genes through a p21(WAF1/CIP1)-dependent switch from MMBto DREAM protein complex binding at CHR promoter elements. CellCycle 2012;11:4661–72.
13. Wang GY, Ho IA, Sia KC, Miao L, Hui KM, Lam PY. Engineering animproved cell cycle-regulatable herpes simplex virus type 1 ampli-con vector with enhanced transgene expression in proliferating cellsyet attenuated activities in resting cells. Hum Gene Ther 2007;18:222–31.
14. Lam PY, Sia KC, Khong JH, De Geest B, Lim KS, Ho IA, et al. Anefficient and safe herpes simplex virus type 1 amplicon vector fortranscriptionally targeted therapy of human hepatocellular carcino-mas. Mol Ther 2007;15:1129–36.
15. Kievit E, Bershad E, Ng E, Sethna P, Dev I, Lawrence TS, et al.Superiority of yeast over bacterial cytosine deaminase for enzyme/prodrug gene therapy in colon cancer xenografts. Cancer Res 1999;59:1417–21.
16. Dachs GU, Hunt MA, Syddall S, Singleton DC, Patterson AV. Bystand-er or no bystander for gene directed enzyme prodrug therapy. Mole-cules 2009;14:4517–45.
17. Chiba T, Suzuki E, Negishi M, Saraya A, Miyagi S, Konuma T, et al. 3-Deazaneplanocin A is apromising therapeutic agent for the eradicationof tumor-initiating hepatocellular carcinoma cells. Int J Cancer2012;130:2557–67.
18. Miranda TB, Cortez CC, Yoo CB, Liang G, Abe M, Kelly TK, et al.DZNep is a global histone methylation inhibitor that reactivates devel-
opmental genes not silenced by DNA methylation. Mol Cancer Ther2009;8:1579–88.
19. Aoki R,Chiba T,Miyagi S, NegishiM, KonumaT, Taniguchi H, et al. Thepolycomb group gene product Ezh2 regulates proliferation and differ-entiation of murine hepatic stem/progenitor cells. J Hepatol 2010;52:854–63.
20. Chang CJ, Yang JY, Xia W, Chen CT, Xie X, Chao CH, et al. EZH2promotes expansion of breast tumor initiating cells through acti-vation of RAF1-beta-catenin signaling. Cancer Cell 2011;19:86–100.
21. Sia KC, Huynh H, Chinnasamy N, Hui KM, Lam PY. Suicidal genetherapy in the effective control of primary human hepatocellular car-cinoma as monitored by noninvasive bioimaging. Gene Ther 2012;19:532–42.
22. Huynh H, Soo KC, Chow PK, Panasci L, Tran E. Xenografts of humanhepatocellular carcinoma: a usefulmodel for testing drugs.ClinCancerRes 2006;12:4306–14.
23. Stroescu C, Dragnea A, Ivanov B, Pechianu C, Herlea V, Sgarbura O,et al. Expression of p53, Bcl-2, VEGF, Ki67 and PCNA and prognosticsignificance in hepatocellular carcinoma. J Gastrointestin Liver Dis2008;17:411–7.
24. Foka P, Pourchet A, Hernandez-Alcoceba R, Doumba PP, Pissas G,Kouvatsis V, et al. Novel tumour-specific promoters for transcriptionaltargeting of hepatocellular carcinoma by herpes simplex virus vectors.J Gene Med 2010;12:956–67.
25. Chao Y, Shih YL, Chiu JH, Chau GY, Lui WY, Yang WK, et al.Overexpression of cyclin A but not Skp 2 correlates with the tumorrelapse of human hepatocellular carcinoma. Cancer Res 1998;58:985–90.
26. Zhou Q, He Q, Liang LJ. Expression of p27, cyclin E and cyclin A inhepatocellular carcinoma and its clinical significance. World J Gastro-enterol 2003;9:2450–4.
27. Wang X, Meng X, Sun X, Liu M, Gao S, Zhao J, et al. Wnt/beta-cateninsignaling pathway may regulate cell cycle and expression of cyclin Aand cyclin E protein in hepatocellular carcinoma cells. Cell Cycle2009;8:1567–70.
28. Wang LH, Huang W, Lai MD, Su IJ. Aberrant cyclin A expression andcentrosome overduplication induced by hepatitis B virus pre-S2mutants and its implication in hepatocarcinogenesis. Carcinogenesis2012;33:466–72.
29. Kim DH, Park SE, Kim M, Ji YI, Kang MY, Jung EH, et al. A functionalsingle nucleotide polymorphism at the promoter region of cyclin A2 isassociatedwith increased risk of colon, liver, and lung cancers. Cancer2011;117:4080–91.
30. Chen XL, Zhou L, Yang J, Shen FK, Zhao SP,Wang YL. Hepatocellularcarcinoma-associated protein markers investigated by MALDI-TOFMS. Mol Med Rep 2010;3:589–96.
31. Topic A, LjujicM, Radojkovic D. Alpha-1-antitrypsin in pathogenesis ofhepatocellular carcinoma. Hepat Mon 2012;12:e7042.
32. Qin X, Chen Q, Sun C, Wang C, Peng Q, Xie L, et al. High-throughputscreening of tumor metastatic-related differential glycoprotein inhepatocellular carcinoma by iTRAQ combines lectin-related techni-ques. Med Oncol 2013;30:420.
33. Fu X, Rivera A, Tao L, De Geest B, Zhang X. Construction of anoncolytic herpes simplex virus that precisely targets hepatocellularcarcinoma cells. Mol Ther 2012;20:339–46.
34. Yamada S, Kuroda T, Fuchs BC, He X, Supko JG, Schmitt A, et al.Oncolytic herpes simplex virus expressing yeast cytosine deaminase:relationship between viral replication, transgene expression, prodrugbioactivation. Cancer Gene Ther 2012;19:160–70.
Transcriptional Targeting Strategy for Human HCC
www.aacrjournals.org Mol Cancer Ther; 12(8) August 2013 1663
on May 25, 2016. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst May 29, 2013; DOI: 10.1158/1535-7163.MCT-13-0056

35. Lu YC, Chen YJ, Yu YR, Lai YH, Cheng JC, Li YF, et al. Replicatingretroviral vectors for oncolytic virotherapy of experimental hepatocel-lular carcinoma. Oncol Rep 2012;28:21–6.
36. Warmann SW, Armeanu S, Heigoldt H, Ruck P, Vonthein R, HeitmannH, et al. Adenovirus-mediated cytosine deaminase/5-fluorocytosinesuicide gene therapy of human hepatoblastoma in vitro. Pediatr BloodCancer 2009;53:145–51.
37. Altanerova V, Cihova M, Babic M, Rychly B, Ondicova K, Mravec B,et al. Human adipose tissue-derived mesenchymal stem cells expres-sing yeast cytosinedeaminase:uracil phosphoribosyltransferase inhib-it intracerebral rat glioblastoma. Int J Cancer 2012;130:2455–63.
38. Yi BR, KimSU, KimYB, LeeHJ, ChoMH,Choi KC. Antitumor effects ofgenetically engineered stem cells expressing yeast cytosine deami-nase in lung cancer brain metastases via their tumor-tropic properties.Oncol Rep 2012;27:1823–8.
39. Chiba T, Kita K, Zheng YW, YokosukaO, SaishoH, IwamaA, et al. Sidepopulationpurified fromhepatocellular carcinomacells harbors cancerstem cell-like properties. Hepatology 2006;44:240–51.
40. Lawrence TS, Davis MA, Maybaum J. Dependence of 5-fluorouracil-mediated radiosensitization on DNA-directed effects. Int J RadiatOncol Biol Phys 1994;29:519–23.
41. Kievit E, Nyati MK, Ng E, Stegman LD, Parsels J, Ross BD, et al. Yeastcytosine deaminase improves radiosensitization and bystander effectby 5-fluorocytosine of human colorectal cancer xenografts. CancerRes 2000;60:6649–55.
42. Perez OD, Logg CR, Hiraoka K, Diago O, Burnett R, Inagaki A, et al.Design and selection of toca 511 for clinical use: modified retroviralreplicating vector with improved stability and gene expression. MolTher 2012;20:1689–98.
43. Spear PG. Herpes simplex virus: receptors and ligands for cell entry.Cell Microbiol 2004;6:401–10.
44. Pawlik TM, Nakamura H, Yoon SS, Mullen JT, Chandrasekhar S,Chiocca EA, et al. Oncolysis of diffuse hepatocellular carcinoma by
intravascular administration of a replication-competent, geneticallyengineered herpesvirus. Cancer Res 2000;60:2790–5.
45. Song TJ, Eisenberg DP, Adusumilli PS, Hezel M, Fong Y. Oncolyticherpes viral therapy is effective in the treatment of hepatocellularcarcinoma cell lines. J Gastrointest Surg 2006;10:532–42.
46. Geevarghese SK, Geller DA, de Haan HA, Horer M, Knoll AE, Mesche-der A, et al. Phase I/II studyof oncolytic herpes simplex virusNV1020 inpatients with extensively pretreated refractory colorectal cancer met-astatic to the liver. Hum Gene Ther 2010;21:1119–28.
47. LuB,GuptaS, Federoff H. Ex vivo hepatic gene transfer inmouse usinga defective herpes simplex virus-1 vector. Hepatology 1995;21:752–9.
48. Muller L, Saydam O, Saeki Y, Heid I, Fraefel C. Gene transfer intohepatocytes mediated by herpes simplex virus-Epstein-Barr virushybrid amplicons. J Virol Methods 2005;123:65–72.
49. Ma S, Lee TK, Zheng BJ, Chan KW, Guan XY. CD133þ HCC cancerstem cells confer chemoresistance by preferential expression of theAkt/PKB survival pathway. Oncogene 2008;27:1749–58.
50. Chen Y, Yu D, Zhang H, He H, Zhang C, Zhao W, et al. CD133(þ)EpCAM(þ) phenotype possesses more characteristics of tumor initi-ating cells in hepatocellular carcinoma Huh7 cells. Int J Biol Sci2012;8:992–1004.
51. TanJ,YangX,ZhuangL, JiangX,ChenW,LeePL, et al. Pharmacologicdisruption of Polycomb-repressive complex 2-mediated gene repres-sion selectively induces apoptosis in cancer cells. Genes Dev 2007;21:1050–63.
52. Zhou J, Bi C, Cheong LL, Mahara S, Liu SC, Tay KG, et al. The histonemethyltransferase inhibitor, DZNep, up-regulates TXNIP, increasesROS production, and targets leukemia cells in AML. Blood 2011;118:2830–9.
53. Avan A, Crea F, Paolicchi E, Funel N, Galvani E, Marquez VE, et al.Molecular mechanisms involved in the synergistic interaction of theEZH2 inhibitor 3-deazaneplanocin A with gemcitabine in pancreaticcancer cells. Mol Cancer Ther 2012;11:1735–46.
Sia et al.
Mol Cancer Ther; 12(8) August 2013 Molecular Cancer Therapeutics1664
on May 25, 2016. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst May 29, 2013; DOI: 10.1158/1535-7163.MCT-13-0056

2013;12:1651-1664. Published OnlineFirst May 29, 2013.Mol Cancer Ther Kian Chuan Sia, Hung Huynh, Alexander Yaw Fui Chung, et al. Mouse ModelHuman Hepatocellular Carcinoma in an Orthotopic Xenograft Preclinical Evaluation of Transcriptional Targeting Strategy for
Updated version
10.1158/1535-7163.MCT-13-0056doi:
Access the most recent version of this article at:
Material
Supplementary
http://mct.aacrjournals.org/content/suppl/2013/05/24/1535-7163.MCT-13-0056.DC1.html
Access the most recent supplemental material at:
Cited articles
http://mct.aacrjournals.org/content/12/8/1651.full.html#ref-list-1
This article cites 53 articles, 13 of which you can access for free at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications Department at
on May 25, 2016. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst May 29, 2013; DOI: 10.1158/1535-7163.MCT-13-0056
Related Documents









![Current status and perspectives of patient-derived xenograft models in cancer research · 2017. 8. 26. · pancreas [131, 132], kidney [26],and ovary [11], which is called orthotopic](https://static.cupdf.com/doc/110x72/6129a53441008e1a43776d58/current-status-and-perspectives-of-patient-derived-xenograft-models-in-cancer-research.jpg)

