PLEASE SCROLL DOWN FOR ARTICLE This article was downloaded by: [Queensland University of Technology] On: 18 February 2010 Access details: Access Details: [subscription number 908224431] Publisher Informa Healthcare Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37- 41 Mortimer Street, London W1T 3JH, UK Leukemia and Lymphoma Publication details, including instructions for authors and subscription information: http://www.informaworld.com/smpp/title~content=t713643806 Potential Adhesion Mechanisms for Localisation of Haemopoietic Progenitors to Bone Marrow Stroma Paul J. Simmons a ; Andrew Zannettino a ; Stan Gronthos a ; David Leavesley a a Matthew Roberts Laboratory, Leukaemia Research Unit, Division of Haematology, Hanson Centre for Cancer Research, Adelaide, South Australia To cite this Article Simmons, Paul J., Zannettino, Andrew, Gronthos, Stan and Leavesley, David(1994) 'Potential Adhesion Mechanisms for Localisation of Haemopoietic Progenitors to Bone Marrow Stroma', Leukemia and Lymphoma, 12: 5, 353 — 363 To link to this Article: DOI: 10.3109/10428199409073776 URL: http://dx.doi.org/10.3109/10428199409073776 Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf This article may be used for research, teaching and private study purposes. Any substantial or systematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The accuracy of any instructions, formulae and drug doses should be independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

PLEASE SCROLL DOWN FOR ARTICLE
This article was downloaded by: [Queensland University of Technology]On: 18 February 2010Access details: Access Details: [subscription number 908224431]Publisher Informa HealthcareInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Leukemia and LymphomaPublication details, including instructions for authors and subscription information:http://www.informaworld.com/smpp/title~content=t713643806
Potential Adhesion Mechanisms for Localisation of HaemopoieticProgenitors to Bone Marrow StromaPaul J. Simmons a; Andrew Zannettino a; Stan Gronthos a; David Leavesley a
a Matthew Roberts Laboratory, Leukaemia Research Unit, Division of Haematology, Hanson Centre forCancer Research, Adelaide, South Australia
To cite this Article Simmons, Paul J., Zannettino, Andrew, Gronthos, Stan and Leavesley, David(1994) 'Potential AdhesionMechanisms for Localisation of Haemopoietic Progenitors to Bone Marrow Stroma', Leukemia and Lymphoma, 12: 5, 353— 363To link to this Article: DOI: 10.3109/10428199409073776URL: http://dx.doi.org/10.3109/10428199409073776
Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf
This article may be used for research, teaching and private study purposes. Any substantial orsystematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply ordistribution in any form to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae and drug dosesshould be independently verified with primary sources. The publisher shall not be liable for any loss,actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directlyor indirectly in connection with or arising out of the use of this material.

Leukemia and Lymphoma, zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAVol. 12, pp. 353-363 Reprints available directly from the publisher Photocopying permitted by license only zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA0 1994 Harwood Academic Publishers GmbH
Printed in the United States zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAof America
Potential Adhesion Mechanisms for Localisation of Haemopoietic Progenitors to
Bone Marrow Stroma PAUL J. SIMMONS, ANDREW ZANNETTINO, STAN GRONTHOS and DAVID LEAVESLEY
Matthew Roberts Laboratory, Leukaemia Research Unit, Division of Haematology, Hanson Centre for Cancer Research, Adelaide, South Australia, 5000
(Received 30 May 1993)
Haemopoiesis occurs in intimate physical association with the stromal elements of the bone marrow. Current evidence supports the hypothesis that the restriction of primitive haemo- poietic progenitor cells (HPC) to the bone marrow involves developmentally regulated ad- hesive interactions between HPC and the stromal cell microenvironment. This review ex- amines the expression and function of cell adhesion molecules (CAM) on human HPC and marrow stromal cells. These data demonstrate that a broad range of CAMS representing at least three adhesion molecule superfamilies (integrins, selectins, immunoglobulin gene su- perfamily) participate in these adhesive interactions. We discuss the potential contribution of these various adhesion molecules to homing of HPC to the bone marrow, their retention within the extravascular haemopoietic compartment and their egress into the peripheral circulation. It is likely that each process is mediated not by a single binding event but requires the co- ordinated participation of multiple receptor-ligand pairs.
KEY WORDS: bone marrow stroma
STROMAL CELLS AND THE RESTRICTION OF HAEMOPOIESIS TO THE BONE MARROW
The importance of cellular interactions in haemo- poietic cell development is well established. '-* De- spite this, precise definition of the nature of many of these interactions at a molecular level is lacking and remains an objective which is of fundamental impor- tance to our understanding of the regulation of hae- mopoiesis. Under steady state conditions the majority of primitive stem cells appear to reside in the bone marrow where they and their progeny develop in as-
Address for correspondence: Paul J. Simmons, Matthew Rob- erts Laboratory, Leukaemia Research Unit, Division of Haema- tology, Hanson Centre for Cancer Research, Frome Road, P.O. Box 14, Rundle Mall, Adelaide, South Australia 5000.
adhesion haemopoietic progenitors
sociation with a phenotypically and probably func- tionally heterogeneous population of stromal cells .7s9-15
The various cellular elements of the stroma together with their associated biosynthetic products including extracellular matrix (ECM) components and haemo- poietic growth factors (HGF) constitute the haemo- poietic microenvironment (HM) of the bone mar- row. 1,2,4,7 Accumulating evidence supports the proposal that the localisation of haemopoiesis to the bone mar- row involves developmentally regulated adhesive in- teractions between primitive haemopoietic cells and this complex stromal cell-mediated HM. 16-19 In this review we ,will summarise recent data describing the expression and function of adhesion molecules on primitive human haemopoietic progenitor cells (HPC) and discuss the potential role of these molecules in the homing of HPC to the bone marrow, their reten- tion within the HM and their release into the periph- eral circulation.
353
Downloaded By: [Queensland University of Technology] At: 00:08 18 February 2010

P. J. SIMMONS zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAETAL. 354 zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBACELLULAR INTERACTIONS IN HAEMOPOIESIS: A MULTIPLICITY OF MECHANISMS
The cellular interactions that regulate haemopoiesis are many and varied. This is hardly surprising when one considers the hierarchical nature of the haemo- poietic system (with discrete stem cell, progenitor cell and maturing cell compartments), the number of cell lineages generated and the diversity of tissue envi- ronments associated with their development and mat- uration (bone marrow, thymus, secondary lymphoid organs). In all likelihood the molecules which me- diate these interactions may function in a stage and/ or lineage-specific manner as occurs, for example in the regulation of T-cell development.20,21 Thus, a zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAva- zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAriety of molecular species are probably involved in supporting a range of interactions throughout hae- mopoiesis. As this review will show, current data support this hypothesis which, in many respects par- allels the diversity of adhesive interactions involved in regulation of the immune system.20
In recent years there has been an exponential in- crease in our understanding of the mechanisms that regulate cell adhesion in multicellular organisms. A broad range of cell surface moieties have been iden- tified that function as cell adhesion molecules (CAMs) for diverse ligands including integral membrane gly- coproteins and ECM components zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA.20,22-24 Several dis- tinct superfamilies of CAMs have been identified in- cluding the immunoglobulin gene superfamily, integrins, selectins, cadherins and the CD44 family (Table l ) , many of which are expressed by haemo- poietic cells. Studies to investigate the function and contribution of particular CAMs to interactions of HPC with the stromal HM are complicated by the diversity of potential ligands for many of these CAMs which are exhibited by marrow stromal tissue (Table 2). This is further compounded by an additional category of interactions which involve HGF-receptor mediated adhesion of HPC to stromal cell-derived HGF pre- sented either as integral membrane protein^^^,^^ or bound to ECM molecules.27 This multiplicity of potential ad- hesive mechanisms presents a major technical and conceptual challenge to the identification of those CAM-ligand pairs which are essential for localisation of HPC to the BM. Nevertheless, some likely can- didates are beginning to emerge. These are reviewed below.
Table 1 Cell adhesion molecule superfamilies
Immunoglobulin superfamily of adhesion receptors
ICAM-2 LFA-3 (CD58) MHC
ICAM-3 CD3/T Cell Receptor N-CAM
PECAM- 1 (CD3 zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA1) CD8 c-kit
ICAM-1 (CD54) LFA-2 (CDZ) Thy- 1
Class I & I1
VCAM- 1 CD4 Ng-CAM
Cation-dependent CAMS N-Cadherin E-Cadherin P-Cadherin
Integrins Numerous: 8 p-chains & 13 a-chains in multiple
combinations
Selectins P-Selectin L-Selectin E-Selectin
Miscellaneous CD44 S y ndecan CD36
Table 2 ECM, Glycoproteins and CAMs expressed on bone marrow stromal cells
ECM components GAGs/Proteoglycans CAMS
Collagen I Chondroitin sulphate ICAM- 1 (CD54) Collagen 111 Heparan sulphate N-CAM (CD56) Collagen IV CD44 VCAM-1 Collagen V H yaluronate LFA-3 (CD58) Collagen VI Thy- 1 Fibronectin Vitronectin Laminin Thrombospondin Haemonectin
Baaed on data from Zuckerman and Wichd (1983). Long and Dixit (1990). Campbell er zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAal. (19901, Gordon zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA(1988), Gallagher (19891, Simmons zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAernl. (1992), Kincade er ul. (1989). Teixido er 01. (1992) [References 28-35],
INTEGRIN-MEDIATED ADHESION OF HPC
Integrins are a large family of CAMs with well doc- umented roles in a variety of cellular functions in- cluding cell migration and tissue organisation during embryonic development, cell differentiation, inflam- mation and m e t a ~ t a s i s . ~ ~ Integrins mediate both cell- cell and cell-ECM interactions and are so-named be- cause of their ability to integrate the intracellular cytoskeleton with the ECM. Structurally, they rep- resent a phylogenetically conserved family of integral membrane glycoproteins that consist of at least 20 distinct heterodimers formed by non-covalent asso- ciation between 13 a subunits and 8 different p sub-
Downloaded By: [Queensland University of Technology] At: 00:08 18 February 2010

MECHANISMS FOR LOCALISATION OF PROGENITORS IN MARROW 355
units, each with distinct ligand-binding properties. zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA37.38
Compounding the diversity of adhesive interactions mediated by integrins, most, if not all integrins can assume multiple functional states. 37 The integrins are subdivided on the basis of /I-chain composition al- though it is important to note that some a-chains (par- ticularly zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAa”) can associate with more than one zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAp sub- unit (Figure 1) . The zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAp1 (CD29) or Very Late Antigen (VLA) integrins comprise the largest subfamily and mediate cell-ECM and cell-cell adhesion. The p2 (CD18) or leukocyte integrins bind to cell surface counter-receptors of the immunoglobulin gene super- family and therefore mediate cell-cell adhesion. The /I3 (CD61) or cytoadhesion integrins predominantly bind to ECM proteins (Figure 1).
To date, the majority of published studies have ex- amined the expression of only the pl, zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA/ I 2 and zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAp3 in-
tegrins in human bone marrow. 33*35341-45 Of these, p3 (CD61) integrin is present on a minor proportion of CD34’ cells (approximately 10%) which are thought to include megakaryocyte precursors zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA.44,46 The expres- sion of p3 by more primitive HPC with the capacity to initiate and sustain haemopoiesis in long-term mar- row culture (so-called long-term culture initiating cells, LTCIC)47 has not been determined. Of the p2 (CD18) integrin family, CDl la (aL) is the only a-chain de- tectable on CD34+ cells (approximately 80%) .35943*48
LTCIC however, are restricted to the CDI la/CD18- subp~pulat ion.~~ There is good evidence for expres- sion of at least one member of the /Il (CD29) integrin family, a4P1 (CD49d/CD29), by human LTCIC33 and because of the brief nature of this overview we will focus on the role of PI integrins in mediating adhesion of HPC to marrow stromal elements.
Col, Fn, Lam Vn, Col, zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA(Lam) Fn,Fg Col, Lam vWF
Fn, Col, Vn vWF, Lam, Fg Tsp, osp, Bsp
Fg, iC3b
FactorX zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAaM ICAM-1
ICAM-1 \ ICAM-3 zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA““7 ICAM-2
ax Fg, iC3b
’ a7 a* zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBALam
?
Figure 1 Integrin Pairing and Ligand Specificity. Integrin heterodimers are formed by non-covalent association of unique a chains with one, or more, p chains. Composition of subunit pairs determines ligand specificity. The p4, p5, p6, p7 and p8 integrins have not yet been shown to be expressed by haemopoietic cells and have been omitted for simplicity. Ligands: Col, collagens; Lam, laminin; Fn, fibronectin; VCAM- 1, vascular cell adhesion molecule- 1; Fg, fibrinogen; vWF, von Willebrand Factor; Tsp, thrombospondin; Osp, osteopontin; Bsp, bone sialoprotein; ICAM- 1, intercellular adhesion molecule- 1 ; ICAM-2, intercellular adhesion molecule-2; ICAM- 3, intercellular adhesion molecule-3; iC3b, complement factor 3b.
Downloaded By: [Queensland University of Technology] At: 00:08 18 February 2010

356 P. J . SIMMONS zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAET AL..
INTEGRIN zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAa4/3,: DISTINCT ROLES IN CELL-CELL AND CELL-ECM ADHESION zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAIntegrin zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAa4P, is expressed by a wide variety of cir- culating lymphoid and myeloid ~ e l l s ~ ~ , ’ ~ and functions as a receptor for two distinct ligands. The first, vas- cular cell adhesion molecule-I (VCAM-l), a member of the immunoglobulin gene superfamily, is an inte- gral membrane glycoprotein which was first identified as an inducible CAM on human endothelial cells by Bevilacqua and c o l l e a g ~ e s . ~ ~ The second ligand for a4P1 is fibronectin (FN),” a major component of the ECM synthesised by bone marrow stromal cells in vivo and in vitro. Three distinct binding sites for zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAa4P1 have been identified on the FN m o l e c ~ l e . ~ ’ . ~ ~ The sites on a4P1 involved in interaction with FN are distinct from those which bind to VCAM-1.53 a4PI therefore me- diates both cell-cell and cell-ECM adhesion.
The pioneering work of Kincade and colleagues first demonstrated the importance of a4P, in regulating haemopoiesis in vitro. Addition of an anti-a, subunit monoclonal antibody (MAb) to murine long term bone marrow culture (LTBMC) was found to completely abrogate lymphopoiesis and to retard myelopoiesis .54
It is not clear from these data however, whether in- hibition of cellular adhesion is responsible for the ob- served suppression or whether some antibody-me- diated growth modulatory effect in LTBMC may also be involved. Recent data suggest an equally important role for a4Pl integrin in human haemopoietic cell in- teractions. Between 46-90% of CD34’ bone marrow cells are reported to co-express zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAa, PI 3333sz43-4s which include the majority of lineage restricted clonogenic progenitors, CFU-GM and BFU-E33,4s and B-cell pre- c u r s o r ~ . ~ ~ Our group has shown, in addition, that LTCIC express a4P1 and that anti-CD49d antibody partially inhibits their adhesion to marrow stromal cells in ~ i t r o . ~ ~ VCAM-1 is constitutively expressed by hu- man bone marrow stromal cells in vitro33*55 and in vivo (SG and PJS; unpublished observations) and its expression can be up-regulated by inflammatory me- diators such as IL- 1, IL-4 and TNFa resulting in con- cordant increases in CD34’ cell adhesion. ’’ Antibody to VCAM-1 partially inhibits the adhesion of LTCIC, myeloid and erythroid progenitors and normal and leukaemic B-lymphoid precursors to cytokine-in- duced marrow stromal cell^.'^.'^ Thus, in addition to roles in physiological processes involving migration and localisation of immune ~ e l l s , ~ ~ ~ ~ ~ the a4y4pI/VCAM- 1 counter receptors also mediate adhesion of primitive HPC to bone marrow stromal cells.
While there is accordance between different groups concerning the role of VCAM-1 in this setting, con- flicting observations have been made regarding the role of FN, the alternative ligand for a4P1. A number of groups have reported insignificant binding of HPC to
while others have demonstrated adhesion of murines8 and human HPC (including LTCIC) to intact zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAFN or proteolytic fragments there~f .~ ’ ”~ Recent work by Hemler and colleaguesm may, at least in part, re- solve these differing observations. The authors iden- tify 3 activation states of a4PI; the fully active recep- tor binds both ligands, the inactive form, neither. A third, partially active form of aqP1 binds only to VCAM-1. Thus, in order to bind FN, further acti- vation of a4PI is required. In accord with this, Kerst zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAet d4’ demonstrated the acquisition of a4PI and aspI- dependent adhesion of CD34’ cells to FN following protein kinase C activation. Given the high proportion of CD34’ cells which bind to VCAM- 1’’ collectively these data suggest that HPC constitutively express the partially active form of a4PI.
Antibodies to a4PI or VCAM-1 alone or in com- bination do not completely block the adhesion of HPC to marrow ~ t r ~ m a ~ ~ , ~ ~ , ~ ~ suggesting that more than one CAM interaction is involved in this process. Recent studies in our laboratory have identified several ad- ditional CAMS on primitive HPC. These are briefly summarised below.
~ ~ 4 5 . 4 8
PECAM-l/CD31: AN AMPLIFIER OF INTEGRIN-MEDIATED ADHESION
Platelet-endothelial cell adhesion molecule- 1 (PECAM- 1) or CD31 is an immunoglobulin gene superfamily member of 130-kD that is expressed at high density on endothelium, platelets, granulocytes, monocytes and by a subset of l y rnph~cy tes .~ ’ -~~ Although the precise function of CD31 is not known in all cell types that express it, a number of studies clearly document its ability to function as a Our own stud- ies [67 and PJS manuscript submitted for publication] demonstrate that in addition to the abovementioned mature leukocyte populations, essentially all CD34+ cells express CD31; BFU-E at low level, CFU-GM at high level and LTCIC at an intermediate level. Anti- CD31 Fab fragments partially inhibit adhesion of CD34’ cells to marrow ~ t r o m a . ~ ~ Although the ligand for CD31 is not yet defined, sulphated proteoglycans are a possible candidate.68 The latter are produced in abundant quantities by marrow stromal cells6* and have
Downloaded By: [Queensland University of Technology] At: 00:08 18 February 2010

MECHANISMS FOR LOCALISATION OF PROGENITORS IN MARROW zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA357 zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAbeen shown to participate in the adhesion of primitive HPC.70 In addition to its role as a CAM, recent work by Shaw and colleagues has demonstrated a role for CD31 in amplifying PI and P2 integrin-mediated adhesion of T cells.71 Whether CD31 has a similar role on HPC has not been investigated.
THE SELECTINS: ADHESION OF HPC
RECOGNITION MEDIATED BY LECTIN-CARBOHYDRATE
The selectins are three structurally related glycopro- teins that participate in leukocyte adhesion to vascular endothelium and platelet^.'^,^^-^^ Leukocyte (L), En- dothelial (E) and Platelet (P)-selectins function as Ca" dependent lectins, mediating rapid, shear-resistant adhesion by recognition of specific carbohydrate li- gands.24,16-78 A role for such protein-carbohydrate in- teractions has already been demonstrated in murine haemopoietic tissues by Tavassoli and colleagues. 1737y
However, few studies have investigated the potential involvement of the selectin family in the interaction of human HPC with the HM.
L-selectin is expressed on mature myeloid cells (neutrophils, eosinophils and monocytes) and most lymphocytes, mediating the adhesion of leukocytes to endothelium at sites of inflammation and homing of lymphocytes to peripheral lymph nodes.247s0781 Our own studies67 demonstrate expression of L-selectin by ap- proximately 75% of CD34' cells which include 290% of CFU-GM and 60% of BFU-E, confirming earlier work by Griffin and colleagues." Additionally, LTCIC express L-selectin at low level and adhere to a TNFa- inducible ligand on marrow stromal cells.67 The na- ture of the carbohydrate and/or protein components of this L-selectin ligand is not yet established. How- ever, based on studies using murine bone marrow stromal cells it appears to be distinct from the 50 kD sialomucin GlyCAM-1 identified as the ligand for L- selectin on murine peripheral lymph node high en- dothelial venules (HEV)83 (AZ and PJS; unpublished observations).
P-selectin (CD62) is a receptor for neutrophils and monocytes that is rapidly translocated from secretory granule membranes to the plasma membrane of ac- tivated platelets and endothelial cells zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA.24,74,77784 Pre- vious studies have documented expression of P-selec- tin by a proportion of bone marrow endothelial cells.85 We have recently demonstrated binding of bone mar- row CD34' cells, including LTCIC, to purified P-se-
lectin [PJS; manuscript submitted for publication]. Of interest, neither sialyl Le" nor sialyl Lea, both well documented ligands for P-selectin, are expressed by CD34' cells.24 Treatment of CD34' cells with neur- aminidase inhibits their adhesion to P-selectin imply- ing a prominent role for sialic acid in the structure and function of the ligand. Antibody HECA 452 which identifies the cutaneous lymphocyte-associated anti- gen (CLA), a ligand for E-selectin on skin-associated memory T-cells, 86 binds to a neuraminidase sensitive epitope on CD34' cells and partially inhibits their adhesion to P-selectin. We are currently investigating the nature of the glycoprotein identified by HECA- 452 on CD34' cells.
HAEMOPOIETIC PROGENITOR CELL- GROWTH FACTOR INTERACTIONS
Haemopoiesis is regulated by a wide variety of hae- mopoietic growth factors (HGF). In the context of ad- hesive interactions between HPC and the stromal HM, HGF are of central importance.
Growth factors which act on HPC have been shown to modulate the adhesive interactions of cells by reg- ulating the expression and/or activation state of CAMs33,73,82,83,87 or by influencing ECM biosynthesis and degradation.87x88 In addition, cell adhesion has been demonstrated to induce the production of HGF by a variety of cell Many HGF are bound in bi- ologically active form by ECM and in this form are able to mediate adhesion of HPC." In ad- dition, certain HGF can exist in both membrane bound and soluble forms. 25v263y2 The integral membrane iso- forms of macrophage colony-stimulating factor (M- CSF) and stem cell factor (SCF) are expressed by marrow stromal cells and have been demonstrated to promote the adhesion of cells bearing the appropriate HGF receptors, c-fms and c-kit, respectively.25326 A single receptor-ligand pair is thus able to stimulate HPC proliferation and support HPC-stromal cell adhesion. A number of cell types which express c-kit, including mast cells and megakaryocytes, adhere to marrow fi- broblasts via interaction with SCF.25,93 c-kit is ex- pressed by 75% of CD34' cells in the bone marrow, including the majority of CFU-GM, BRJ-E and LTCIC [Reference 94; PJS, G Aylett, LK Ashman; manu- script submitted for publication], Although HPC have not previously been reported to utilize this adhesion mechanism, preliminary data from this laboratory suggest they do and furthermore, that cell-cell adhe-
Downloaded By: [Queensland University of Technology] At: 00:08 18 February 2010

358 zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAP. zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAJ. SIMMONS zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAETAL.
sion mediated by the c-kit-SCF receptor/ligand pair is additive with respect to that supported by other CAMs such as integrin zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA(~4pI [AZ and PJS; unpub- lished observations]. At this time it is unclear (1) to what extent binding to immobilized HGFs is utilized by HPC as an adhesive interaction, (2) the range of HGF that mediate cell adhesion, and (3) whether ECM- bound HGF are as effective at promoting adhesion as integral membrane isoforms. This adhesion mecha- nism clearly warrants further exploration.
OTHER CANDIDATE CAMS
Because of the brief nature of this overview we have, of necessity, focussed on a limited number of CAM- ligand interactions with roles in supporting adhesion between HPC and the marrow stroma. However, other CAMs are clearly involved in this complex process. CD44, the major cell surface receptor for hyaluronic acid95 exhibits multiple activation states96 and exists in a multitude of i s o f o ~ m s . ~ ~ Anti-CD44 antibodies identify primitive human HPC43,98 and marrow stromal cells. Addition of anti-CD44 antibodies to murine LTBMC results in a profound inhibition of haemo- poiesis, in particular, B-lymphoid de~e lopmen t .~~ Several CAMs belonging to the Ig superfamily are expressed by human HPC including LFA-3 (CD58),43 ICAM- 1 (CD54) ,4391M) and Thy- 1. ICAM- 1 and Thy- 1 are also present on marrow stromal cells [References 35, 102; SG and PJS; unpublished observations], as is N-CAM (CD56)lo3 but, with the exception of ICAM- 1, have not been shown to participate in adhesive in- teractions with HPC. CD43, a counter-receptor for ICAM-1,'O4 is expressed at high density by CD34+ cells although its potential role in HPC-stromal cell adhesion has not been investigated. CD34, which shares several biochemical features with CD43 has been speculated to function as a CAM for HPC although such a role has not been documented. In fact, based on recent studies with vascular endothelial cells, CD34 is proposed to have a negative modulating role on cell- cell adhesion, perhaps by increasing the stringency re- quirements of ligand-receptor interactions that facili- tate adhesion. lo5
A MULTIPLICITY OF CAMS: REDUNDANCY OR COMBINATORIAL DIVERSITY?
There is now considerable data demonstrating that primitive human HPC exhibit a multitude of CAMs
each with specificity for distinct counter receptors on marrow stromal cells. Does this imply redundancy within this repertoire, or do particular CAMs perform separate functions associated with, for example, the homing of HPC to the bone marrow, or their lodge- ment and retention within the extravascular HM? (Figure 2). Given the potential complexity of both phenomena such an array of CAMs on HPC might be anticipated. Studies of the mechanism of entry of ma- ture leukocytes into tissues provide important insights into this question. From such studies has come the realisation that this is a multi-step process controlled by a dynamic interaction between CAMs expressed by both leukocytes and endothelial cells. Three fam- ilies of CAMs participate in this process. '06-'08 The initial attachment and "rolling" of leukocytes along the endothelium is mediated by selectins. The next stage involves firm attachment and is accomplished by a triggering event which causes activation of in- tegrins thereby facilitating binding to their counter- receptors, the various members of the immunoglob- ulin gene superfamily zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA. The final stage, extravasation, involves a change in the shape of the leukocyte and transmigration through the endothelium. Thus, rather than being dependent on a single binding event, the specificity and diversity of leukocyte-endothelial cell interactions relies on sequential and combinatorial ad- hesive interactions. Given that HPC express a sim- ilar cohort of CAMs to those found on circulating ~ymp~0cytes20.22.24.38.&1 a similar scenario of events may be involved in the homing of HPC to the bone marrow (Figure 2). Since the marrow circulation is closed, in order to enter the marrow sinus extravascular com- partment HPC must first recognise, or be recognised, by endothelial cells that separate the two compart- ments. Endothelial cells with this specialised function have yet to be identified. Once in the haemopoietic compartment, HPC interact with stromal elements that regulate their subsequent growth and development. It remains to be determined if the CAMs responsible for guiding HPC to the marrow endothelium are the same as those involved in binding to marrow stromal cells or whether distinct CAMs regulate the latter process of lodgement (Figure 2).
The mechanisms governing the release of primitive HPC from the bone marrow are unknown but may be distinct from the developmentally regulated changes in CAM expression that presumably contribute to the release of mature haemopoietic cells from the mar- row. A variety of perturbations result in the transient release of primitive HPC into the peripheral circula-
Downloaded By: [Queensland University of Technology] At: 00:08 18 February 2010
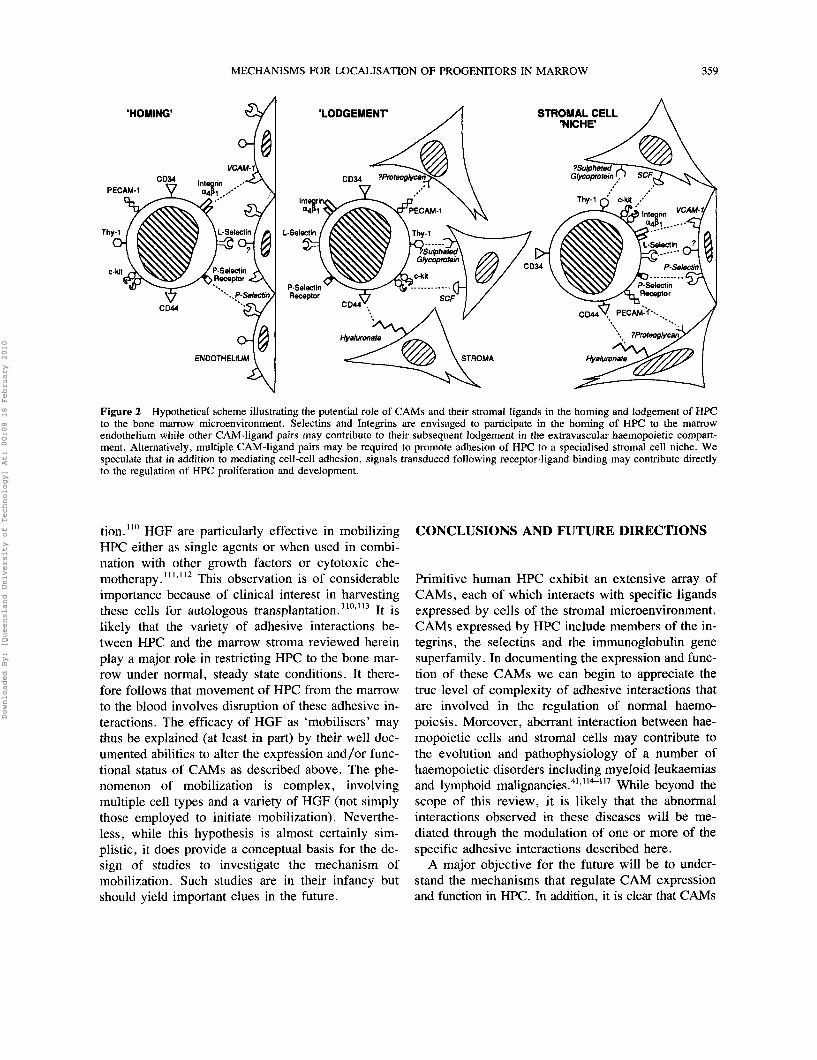
MECHANISMS FOR LOCALISATION OF PROGENITORS IN MARROW zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA359 zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA'LODGEMENT' zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAA STROMAL CELL
'NICHE
Figure 2 Hypothetical scheme illustrating the potential role of CAMs and their stromal ligands in the homing and lodgement of HPC to the bone marrow microenvironment. Selectins and Integrins are envisaged to participate in the homing of HPC to the marrow endothelium while other CAM-ligand pairs may contribute to their subsequent lodgement in the extravascular haemopoietic compart- ment. Alternatively, multiple CAM-ligand pairs may be required to promote adhesion of HPC to a specialised stromal cell niche. We speculate that in addition to mediating cell-cell adhesion, signals transduced following receptor-ligand binding may contribute directly to the regulation of HPC proliferation and development.
tion."' HGF are particularly effective in mobilizing HPC either as single agents or when used in combi- nation with other growth factors or cytotoxic che- motherapy. ' ' 1 *112 This observation is of considerable importance because of clinical interest in harvesting these cells for autologous transplantation. 110~113 It is likely that the variety of adhesive interactions be- tween HPC and the marrow stroma reviewed herein play a major role in restricting HPC to the bone mar- row under normal, steady state conditions. It there- fore follows that movement of HPC from the marrow to the blood involves disruption of these adhesive in- teractions. The efficacy of HGF as 'mobilisers' may thus be explained (at least in part) by their well doc- umented abilities to alter the expression and/or func- tional status of CAMs as described above. The phe- nomenon of mobilization is complex, involving multiple cell types and a variety of HGF (not simply those employed to initiate mobilization). Neverthe- less, while this hypothesis is almost certainly sim- plistic, it does provide a conceptual basis for the de- sign of studies to investigate the mechanism of mobilization. Such studies are in their infancy but should yield important clues in the future.
CONCLUSIONS AND FUTURE DIRECTIONS
Primitive human HPC exhibit an extensive array of CAMs, each of which interacts with specific ligands expressed by cells of the stromal microenvironment. CAMs expressed by HPC include members of the in- tegrins, the selectins and the immunoglobulin gene superfamily . In documenting the expression and func- tion of these CAMs we can begin to appreciate the true level of complexity of adhesive interactions that are involved in the regulation of normal haemo- poiesis. Moreover, aberrant interaction between hae- mopoietic cells and stromal cells may contribute to the evolution and pathophysiology of a number of haemopoietic disorders including myeloid leukaemias and lymphoid malignancies .417114-117 While beyond the scope of this review, it is likely that the abnormal interactions observed in these diseases will be me- diated through the modulation of one or more of the specific adhesive interactions described here.
A major objective for the future will be to under- stand the mechanisms that regulate CAM expression and function in HPC. In addition, it is clear that CAMs
Downloaded By: [Queensland University of Technology] At: 00:08 18 February 2010

360 P. J. SIMMONS zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAET zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAAL.
are signalling molecules. In the case of the integrins, for example, the signal transduction pathways share a number of features with those of polypeptide growth factors such as EGF, PDGF and insulin that bind to cell surface receptor tyrosine kinases. zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAL18-12" More- over, a number of cell types including haemopoietic cells, demonstrate adhesion induced changes in gene e x p r e s ~ i o n . ~ ~ ~ ' ~ ~ Thus the diverse CAM-ligand inter- actions reviewed here, rather than simply serving to initiate zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAand maintain contact between HPC and stromal cells, might also have an additional more direct role in controlling the growth and development of primi- tive haemopoietic cells as suggested by the 'niche' model (based on that of Schofield'22) depicted in Fig- ure 2. Such a model has obvious implications for the regulation of haemopoiesis, both normal and abnor- mal, and remains a provocative line of investigation for the future.
Acknowledgements The authors express their gratitude to Dr. Pri- tinder Kaur, Dr. Chris Juttner, and David Haylock for critically reviewing the manuscript. We also thank Silvanna Niutta and Sharon Paton for their excellent technical assistance in much of the ex- perimental work reported herein and to Mandy Huxtable for typing and preparation of the manuscript. This work was supported by grants from the National Health and Medical Research Council of Australia and the Anti-Cancer Foundation of the Universities of South Australia.
REFERENCES
1.
2.
3.
4. zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA5 .
6.
7. zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA8.
9.
10.
11.
Trentin, J. J . (1970) Influence of hematopoietic organ stroma (Hematopoietic Inductive Microenvironments) on stem cell differentiation. In Regulation offiematopoiesis, edited by A. S. Gordon, vol. zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA1, pp. 161-186. New York: Appleton-Cen- tury-Crofts. Tavassoli, M. (1975) Studies in hemopoietic microenviron- ments. Exp. Hematol., 3, 213-226. Price, G. B. and McCulloch, E. A. (1978) Cell surfaces and the regulation of hemopoiesis. Semin. Hematol., 15, 283- 300. Wolf, N. S. (1979) The hemopoietic microenvironment. Clin. Hematol., 8, 469-500. Cline, M. J. and Golde, D. W. (1979) Cellular interactions in haematopoiesis. Nature, 277, 177-181. Dexter, T. M. (1979) Cell Interaction in vitro. zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAClin. He- matol., 8, 453-468. Dexter, T. M. (1982) Stromal cell-associated haemopoiesis. J . Cell. Physiol., (Suppl. 1). 87-94. Torok-Storb, B. (1988) Cellular Interactions. Blood, 73, 373- 380. Weiss, L. (1976) The hematopoietic microenvironment of the bone marrow: An ultrastructural study of the stroma in rats. Anat. Rec., 186, 161-184. Lichtman, M. A. (1981) The ultrastructure of the hemopoi- etic environment of the marrow: A Review. Exp. Hematol., 9, 391-410. Bentley, S. A. (1982) Bone marrow connective tissue and
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
the haemopoietic microenvironment. Br. J . Haematol., 50, 1-6. Tavassoli, M. and Friedenstein, A. (1983) Hemopoietic stromal microenvironment. Ann. J . Hematol., 15, 195-203. Simmons, P. J., Przepiorka, D., Thomas, E. D. and Torok- Storb, B. (1987) Host origin of bone marrow stromal cells following allogeneic bone marrow transplantation. Nature,
Allen, T. D., Dexter, T. M. and Simmons, P. J. (1990) Marrow Biology and Stem Cells. In Colony Stimulating Fac- tors: Molecular and Cellular Biology, edited by T. M. Dex- ter, J. M. Garland, and N. G. Testa, pp. 1-38. New York: Marcel Dekker. Simmons, P. J. and Torok-Storb, B. (1991) Identification of stromal cell precursors in human bone marrow by a novel monoclonal antibody, STRO-1. Blood, 78, 55-58. Dexter, T. M., Allen, T. D. and Lajtha, L. G . (1977) Con- ditions controlling the proliferation of hemopoietic stem cells in vitro. J . Cell. Physiol., 91, 335-344. Tavassoli, M. and Hardy, C. L. (1990) Molecular basis of homing of intravenously transplanted stem cells. Blood, 76. 1059- 1070. Clark, B. R., Gallagher, J. T. and Dexter, T. M. (1992) Cell adhesion in the stromal regulation of adhesion. Bail- liere's Clinical Hematol., 5 , 21-53. Long, M. W. (1992) Blood cell cytoadhesion molecules. Exp. Hematol., 20, 288-301. Springer, T. A. (1990) Adhesion receptors of the immune system. Nature, 346, 425-434. Dunon, D. and Imhof, B. A. (1993) Mechanisms of thymus homing. Blood, 81, 1-8. Albelda, S. M. and Buck, C. A. (1990) Integrins and other cell adhesion molecules. FASEB J . , 4, 2868-2879. Haynes, B. F., Telen, M. J., Hale, L. P. and Denning, S. M. (1989) CD44-A molecule involved in leukocyte ad- herence and T-cell activation. Immunol. Today, 10, 423- 428. Bevilacqua, M. P. and Nelson, R. M. (1993) Selectins. J . Clin. Invest., 91, 379-387. Flanagan, J. G., Chan, D. C. and Leder, P. (1991) Trans- membrane form of the kit ligand growth factor is determined by alternate splining and is missing in the SLd mutant. Cell,
Rettenmier, C. W., Roussel, M. F. , Ashmun, R. A., Ralph, P., Price, K. and Sherr, C. J. (1987) Synthesis of membrane bound colony-stimulating factor-I (CSF-1) and down-mod- ulation of CSF-1 receptors in NIH-3T3 cells transformed by co-transfection of the human CSF-1 and c-fms (CSF-1 re- ceptor) genes. Mol. Cell. Biol., 7, 2378-2387. Roberts, R. A., Gallagher, J. T., Spooner, E., Allen, T. D. and Dexter, T. M. (1987) Heparan sulphate-bound growth factors: a mechanism for stromal cell mediated haemo- poiesis. Nature, 332, 376-378. Zuckerman, K. S. and Wicha, M. S. (1983) Extracellular matrix production by the adherent cells in long-term murine bone marrow cultures. Blood, 61, 540-547. Long, M. W. and Dixit, V. (1990) Thrombospondin func- tions as a cyto-adhesion molecule for human hematopoietic progenitors. Blood, 75, 231 1-2318. Campbell, A. D., Long, M. W. and Wicha, M. S . (1990) Developmental regulation of granulocytic cell binding to haemonectin. Blood, 75, 1758-1764. Gordon, M. Y. (1988) Extracellular matrix of the marrow microenvironment Br. J . Haematol., 70, 1-4. Gallagher, J. T. (1989) The extended family of proteogly- cans: social residents of the pericellular zone. Curr. Opin. Cell Biol., 1, 1201-1218. Simmons, P. J., Masinovsky, B., Longenecker, B. M., Ber- enson, R., Torok-Storb, B. and Gallatin, W. M. (1992) Vas-
328, 429-432.
64, 1025-1035.
Downloaded By: [Queensland University of Technology] At: 00:08 18 February 2010

MECHANISMS FOR LOCALISATION OF PROGENITORS IN MARROW 36 zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA1
cular Cell Adhesion Molecule-1 expressed by bone marrow stromal cells mediates the binding of hematopoietic progen- itor cells. zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBABlood, 80, 388-395.
34. Kincade, P. W., Lee, G., Pietrangeli, C. E., Hayashi, S. I. and Gimble, J. M. (1989) Cells and molecules that regulate B-lymphopoiesis in bone marrow. Ann. Rev. Immunol., zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA7,
35. Teixido, J., Hemler, M. E., Greenberger, J. S. and Ank- lesaria, P. (1992) Role of pl and p2 integrins in the adhesion of human CD34H' cells to bone marrow stroma. J . Clin. In- vest., 90, 358-367.
36. Hynes, R. 0. (1987) Integrins: A Family of cell surface re- ceptors. Cell, 48, 549-554.
37. Hynes, R. 0. (1992) Integrins: Versatility, modulation and signalling in cell adhesion. Cell, 69, 11-25.
38. Hemler, M. E. (1990) VLA proteins in the integrin family: structures, functions and their roles on leukocytes. Ann. Rev. Immunol., 8, 365-400.
1 1 1- 143.
39. Ruoslahti, E. (1991) Integrins. J. Clin. Invest., 87, 1-5. 40. Arnaout, A. M. (1990) Structure and function of the leu-
kocyte adhesion molecules CDll/CD18. Blood, 75, 1037- 1050.
41. Soligo, D., Schiro, R., Luksch, R., Manara, G., Quirici, N., Parravine, C. and Deliliers, G. L. (1990) Expression of integrins in human bone marrow. Br. J . Haematol., 76, 323- 330.
42. Rosemblatt, M., Vuillet Gaugler, M. H., Leroy, C. and Coulombel, L. (1991) Co-expression of two fibronectin re- ceptors, VLA-4 and VLA-5 by immature erythroblastic pre- cursor cells. J. zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAClin. Invest., 87, 6-12.
43. Saeland, S., Duvert, V., Caux, C., Pandrau, D., Favre, C., Valle, A,, Durand, I., Charboard, P., de Vries, J. and Banchereau, J. (1992) Distribution of surface membrane molecules on bone marrow and cord blood CD34' hema- topoietic cells. Exp. Hematol., 20, 24-33.
44. Liesveld, J. L., Winslow, J. M., Frediani, K. E., Ryan, D. H. and Abboud, C. N. (1993) Expression of integrins and examination of their adhesive function in normal and leu- kaemic hematopoietic cells. Blood, 81, 112-121.
45. Kerst, B. J. M., Sanders, J. B., Slaper-Cortenbach, I. C. M., Doorakkers, M. C., Hooibrink, B., van Ders, R. H. J., von dem Borne, A. E. G. K. and van der Schoot, C. E. (1993) a& and a& are differentially expressed during myelopoiesis and mediate the adherence of human CD34 cells to fibronectin in an activation-dependent way. Blood, 81,
46. Bender, J. G., Williams, S. F., Myers, S., Nottleman, D., Lee, W. J., Unverzagt, K. L., Walker, D., To, L. B. and van Epps, D. E. (1992) Characterisation of chemotherapy- mobilized peripheral blood progenitor cells for use in au- tologous stem cell transplantation. Bone Marrow Trans- plant., 10, 281-285.
47. Sutherland, H. J., Eaves, C. J., Eaves, A. C., Dragowska, W. and Lansdorp, zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAP. (1989) Characterisation and partial pu- rification of human marrow cells capable of initiating long- term hemopoiesis in vitro. Blood, 74, 1563-1570.
48. Liesveld, J. L., Winslow, J. M., Kempski, M. C., Ryan, D. H., Brennan, J. K. and Abboud, C. N. (1991) Adhesive interactions of normal and leukaemic human CD34' myeloid progenitors: role of marrow stromal, fibroblast and cyto- matrix components. Exp. Hematol., 19, 63-71.
49. Gunji, Y., Nakamura, M., Hagiwara, T., Hayakonwa, K., Matsushuta, H., Osawa, H., Nagagoshi, K. and Suda, T. (1992) Expression and function of adhesion molecules on human hematopietic stem cells: CD34'LFA- 1 ~ cells are more primitive than CD34'LFA-1' cells. Blood, 80, 429-436.
50. Rice, G. E., Munro, J. M. and Bevilacqua, M. P. (1990) Inducible cell adhesion molecule 110 (INCAM-I 10) is an
344-35 1.
endothelial receptor for lymphocytes. J. Exp. Med., 171,
51. Wayner, E. A., Garcia-Pardo, A,, Humphries, M. J., McDonald, J. A. and Carter, W. G. (1989) Identification and characterisation of the T-lymphocyte receptor for an al- ternative cell attachment domain (CS-l) in plasma fibronec- tin. J . Cell Biol., 109, 1321-1330.
52. Mould, A. P. and Humphries, M. J. (1991) Identification of a novel recognition sequence for the integrin zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAalp, in the COOH-terminal heparin-binding domain of fibronectin. EMBO
53. Elices, M. J., Oxborn, L., Takada, Y., Crouse, C., Lu- howskyj, S., Hemler, M. E. and Lobb, R. R. (1990) VCAM- 1 on activated endothelium interacts with the leukocyte in- tegrin VLA-4 at a site distinct from the VLA-4/fibronectin binding site. Cell, 60, 577-583.
54. Miyake, K., Weissman, K., Greenberger, J. S. and Kin- cade, P. W. (1991) Evidence for a role of the integrin VLA- 4 in lympho-hemopoiesis. J. Exp. Med., 173, 599-607.
55. Ryan, D. H., Nuccie, B. L., Abboud, C. N. and Winslow, J. M. (1991) Vascular cell adhesion molecule-1 and the in- tegrin VLA-4 mediate adhesion of human B cell precursors to cultured bone marrow adherent cells. J . Clin. Invest., zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA88,
56. Cybulsky, M. I. and Gimbrone, M. A. (1991) Endothelial expression of a leukocyte adhesion molecule during ather- ogeneis. Science, 251, 788-791.
57. Rice, G. E., Munro, J. M., Corless, C. and Bevilacqua, M. P. (1991) Vascular and non-vascular expression of INCAM- 110. A target for mononuclear leukocyte adhesion in normal and inflamed tissues. Am. J . Pathol., 138, 385-392.
58. Williams, D. A,, Rios, M., Stephens, C. and Patel, V. P. (1991) Fibronectin and VLA-4 in hematopoietic stem cell- microenvironment interactions. Nature, 352, 438-441.
59. Verfaille, C. M., McCarthy, J. B. and McGlave, P. B. (1991) Differentiation of primitive human multipotent hemato- poietic progenitors into single lineage clonogenic progenitors is accompanied by alterations in the interaction with fibro- nectin. J . Exp. Med., 174, 693-703.
60. Masumoto, A. and Hemler, M. E. (1993) Multiple activa- tion states of VLA-4. Mechanistic differences between adhe- sion to CS-l/fibronectin and to vascular cell adhesion mol- ecule-l. J . Biol. Chem., 268, 228-234.
61. Albelda, S. M., Muller, W. A,, Buck, C. A. and Newman, P. J. (1991) Molecular and cellular properties of PECAM-1 (endoCAM/CD3 1): a novel vascular cell-cell adhesion mol- ecule. J. Cell Biol., 114, 1059-1068.
62. Newman, P. J., Berndt, M. C., Gorski, J., White, C., Ly- man, S., Paddock, C. and Muller, W. A. (1990) PECAM- 1 (CD31) cloning and relationship to adhesion molecules of the immunoglobulin gene superfamily. Science, 247, 1219- 1222.
63. Stockinger, H., Gadd, S. J., Eher, R., Majdic, O., Schrei- ber, W., Kasinrerk, W., Strass, B., Schanbl, E. and Knapp, W. (1990) Molecular characterisation and functional analy- sis of the leukocyte surface protein CD31. J . Immunol., 14, 3889-3897,
64. Torimoto, Y., Rothstein, D. M., Dang, N. H., Schlossman, S. F. and Morimoto, C. (1992) CD31, a novel cell surface marker for CD4 cells of suppressor lineage, unaltered by state of activation. J . Immunol., 148, 388-396.
65. Muller, W. A,, Berman, M. E., Newman, P. J. , DeLisser, H. M. and Albelda, S. M. (1992) A heterophilic adhesion mechanism for platelet/endothelial cell adhesion molecule- 1 (CD31). J. Exp. Med., 175, 1401-1404.
66. Schirnmerti, L. A,, Yan, H.-C., Madri, J. A. and Albelda, S. M. (1992) Platelet endothelial cell adhesion molecule, PECAM-1, modulates cell migration. J. Cell Physiol., 153, 417-428.
1369-1374.
J.. 10, 4039-4095.
995- 1004.
Downloaded By: [Queensland University of Technology] At: 00:08 18 February 2010

362 P. J. SIMMONS zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAE T A L .
67. Simmons, P. J., Niutta, zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAS. , Ashman, L. K., Berndt, M. C. and Juttner, C. A. (1992) Primitive human bone marrow cells express L-selectin and PECAM-I. Exp. Hematol., zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA20, 329 (Abstr.).
68. Delisser, H. M., Muller, W. A,, Newman, P. J. and Al- belda, S. M. (1991) PECAM-l (CD31) mediates hetero- philic cell-cell adhesion. J . Cell Biol., 115, 2317.
69. Spooncer, E., Gallagher, J. T. , Krizsa, F. and Dexter, T. M. (1983) Regulation of hematopoiesis in long-term bone marrow cultures IV. Glycosaminoglycan synthesis and the stimulation of haematopoiesis by beta-D-xylosides. J . Cell Biol., 96, 510-520.
70. Siczkowski, M., Clark, D. and Gordon, M. Y. (1992) Bind- ing of primitive haemopoietic progenitor cells to marrow stromal cells involves heparan sulphate. Blood, 80, 912-919.
71. Tanaka, Y., Albelda, S. M., Horgan, K. J., van Seventer, G. A, , Shimizu, Y., Newman, W., Hallam, J., Newman, P. J., Buck, C. A. and Shaw, S. (1992) CD31 expressed on distinctive T cell subsets is a preferential amplifier of inte- grin-mediated adhesion. J . Exp. Med., 176, 245-253.
72. Lasky, L. A,, Singer, M. S . , Yednock, T. A,, Dowbenko, D., Fennic, C., Rodriguez, H. , Nguyen, T., Stachel, S. and Rosen, S. D. (1989) Cloning of a lymphocyte homing re- ceptor reveals a lectin domain. Cell, 56, 1045-1055.
73. Bevilacqua, M. P., Stengelin, S . , Gimbrone, M. A. and Seed, B. (1989) Endothelial leukocyte adhesion molecule 1: an in- ducible receptor for neutrophils related to complement reg- ulatory proteins and lectins. Science, 243, 1160-1 165.
74. Johnston, G. I., Cook, R. G. and McEver, R. P. (1989) Cloning of GMP-140, a granule membrane protein of plate- lets and endothelium: sequence similarity to proteins in- volved in cell adhesion and inflammation. zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBACell, 56, 1033- 1055.
75. Bevilacqua, M. P., Butcher, E., Furie, B., Gallatin, M., Gimbrone, M. A., Harlan, J., Kishimoto, K . , Lasky, L . , McEver, R. and Paulson, J. (1991) Selectins: a family of adhesion receptors. CelI, 67, 233.
76. Imai, Y. , Singer, M. S . , Fennic, C. , Lasky, L. A. and Ro- sen, S . D. (1991) Identification of a carbohydrate based en- dothelial ligand for a lymphocyte homing receptor. J . Cell Biol., 113, 1213-1221.
77. Moore, K . L., Stults, N. L. , Diaz, S. , Smith, D. F., Cum- mings, R. D. , Varki, A. and McEver, R. P. (1992) Iden- tification of a specific glycoprotein ligand for P-selectin (CD62) on myeloid cells. J . Cell Biol., 118, 445-456.
78. Polley, M. J., Phillips, M. L., Wayner, E., Nudelman, E . , Singhal, A. K., Hakomori, S. 1. and Paulson, J. C. (1991) CD62 and endothelial cell-leukocyte adhesion molecule- 1 (ELAM-1) recognize the same carbohydrate ligand, sialyl- lewis x. Proc. Natl. Acad. Sci. USA, 88, 6224-6228.
79. Aizawa, S. and Tavassoli, M. (1987) In vitro homing of hemopoietic stem cells is mediated by a recognition mech- anism with galactosyl and mannosyl specificity. Proc. Natl. Acud. Sci. USA, 84, 4485-4489.
80. Gallatin, W. M., Weissman, I. L. and Butcher, E. C. (1983) A cell-surface molecule involved in organ-specific homing of lymphocytes. Nature. 304, 30-34.
81. Tedder, T. F., Penta, A. C., Levine, H. B. and Friedman, A. S. (1990) Expression of the human leukocyte adhesion molecule, LAMI: Identity with the TQI and Leu8 differ- entiation antigens. J . Immunol., 144, 532-540.
82. Griffin, J. D., Spertini, O., Ernst, T. J., Belvin, M. P., Levine, H. B., Kanakura, Y. and Tedder, T. F. (1990) Granulocyte-macrophage colony-stimulating factor and other cytokines regulate surface expression of the leukocyte adhe- sion molecule LAM-I zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAon human neutrophils, monocytes and their precursors. J . Immunol., 145, 576-584.
83. Lasky, L. A., Singer, M. S . , Dowbenko, D., Imai, Y., Henzel, W. J., Grimley, C., Fennic, C., Gillet, N., Watson,
S. R. and Rosen, S. D. (1992) An endothelial ligand for L- selectin is a novel mucin-like molecule. Cell, 69, 927-938.
84. Hamburger, S. A. and McEver, R. P. (1990) GMP-140 me- diates adhesion of stimulated platelets to neutrophils. Blood,
85. Beckstead, J. H., Stenberg, P. E., McEver, R. P., Shuman, M. A. and Bainton, D. F. (1986) Immunohistochemical lo- calization of membrane and a-granule protein in human megakaryocytes: Application to plastic-embedded bone mar- row biopsy specimens. Blood, 67, 285-293.
86. Berg, E. L., Yoshino, T., Rott, L. S., Robinson, M. K., Warnock, A, , Kishimoto, T. K., Picker, L . J . andButcher, E. C. (1991) The cutaneous lymphocyte antigen is a skin lymphocyte homing receptor for the vascular lectin endothe- lial cell-leukocyte adhesion molecule 1. J . Exp. Med., 174,
87. Nathan, C. and Sporn, M. (1991) Cytokines in context. J . Cell Biol . , 113, 981-986.
88. Nugent, M. A. and Neuman, M. J. (1989) Inhibition of nor- mal rat kidney cell growth by transforming growth factor /3 is mediated by collagen. J . Biol. Chem., 264, 18060-18067.
89. Thorens, B., Mermod, J.-J. and Vassalli, P. (1987) Phago- cytosis and inflammatory stimuli induce GM-CSF mRNA in macrophages through post-transcriptional regulation. Cell, zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
90. Rathjen, P. D., Toth, S., Willis, A., Heath, J. K. and Smith, A. G . (1990) Differentiation-inhibiting activity is produced in matrix-associated and diffusible forms that are generated by alternate promotor usage. Cell, 62, 1105-1 114.
91. Long, M. W., Briddell, R., Walter, A. W., Bruno, E. and Hoffman, R. (1992). Human hernatopoietic stem cell ad- herence to cytokine and matrix molecules. J . Clin. Invesr.,
92. Massague, J. (1990) Transforming growth factor-a: a model for membrane-anchored growth factors. J . Biol. Chem., 265,
93. Avraham, H., Scadden, D. T., Chi, S., Broudy, V. S. , Zsebo, K. M. and Groopman, J. E. (1992) Interaction of human bone marrow fibroblasts with megakaryocytes: Role of the c-kit ligand. Blood, 80, 1679-1684.
94. Papayannopoulou, T., Brice, M., Broudy, V. C. and Zsebo, K. M. (1991) Isolation of c-kit receptor-expressing cells from bone marrow, peripheral blood and foetal liver: Functional properties and composite antigenic profile. Blood, 78, 1403- 1410.
95. Armfo, A., Stamenkovic, I., Melnick, M., Underhill, C . B. and Seed, B. (1990) CD44 is the principal cell surface re- ceptor for hyaluronate. Cell, 61, 1303-13 13.
96. Lesley, J. and Hyman, R. (1992) CD44 can be activated to function as an hyaluronic acid receptor in normal murine T cells. Eur. J . Immunol., 22, 2719-2723.
97. Screaton, G. R., Bell, M. V., Jackson, D. G . , Cornelis, F. B., Gerth, U. and Bell, J. I. (1992) Genomic structure of CD44 reveals at least 12 alternatively spliced exons. Proc. Natl. Acad. Sci. USA, 89, 12160-12164.
98. Lewinsohn, D. M., Nagler, A., Ginzton, N., Greenberg, P. and Butcher, E. C. (1990) Hematopoietic progenitor cell expression of the H-CAM (CD44) homing-associated adhe- sion molecule. Blood, 75, 589-595.
99. Miyake, K., Medina, K. L . , Hayashi, S.-I. , Ono, S . , Ha- maoka, T. and Kincade, P. w. (1990) Monoclonal antibod- ies to Pgp-l/CD44 block lymphopoiesis in long-term bone marrow cultures. J . Exp. Med., 171, 477-488.
100. Arkin, S., Naprstek, B., Guarini, L . , Ferrone, S. and Lip- ton, J. M. (1991) Expression of intercellular adhesion mol- ecule- l (CD54) on hernatopoietic progenitors. Blood, 77,948- 953.
101. Baum, C . M., Weissman, I. L . , Tsukamoto, A. S . , Buckle, A. M. and Peault, B. (1992) Isolation of a candidate human
75, 550-554.
I46 1 - 1466.
48, 671-679.
90, 251-255.
21393-31396.
Downloaded By: [Queensland University of Technology] At: 00:08 18 February 2010

MECHANISMS FOR LOCALISATION OF PROGENITORS IN MARROW 363
haemopoietic stem-cell population. zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAProc. Natl. Acad. zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBASci. USA, zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA89, 2804-2808.
102. Caligaris-Cappio, F., Berguli, L. and Gregoretti, M. G. (1991) Role of bone marrow stromal cells in the growth of human multiple myeloma. Blood, zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA77, 2688-2693.
103. Kincade, P. W., Lee, G., Pietrangeli, C. E., Hayashi, S. I. and Gimble, J. M. (1989) Cells and molecules that regulate B lymphopoiesis in bone marrow. Annu. Rev. Immunol., 7, zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA11 1-143.
104. Rosenstein, Y., Park, J. K., Hahn, W. C., Rosen, F. S., Bierer, B. E. and Burakoff, S. zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBAJ . , (1991) CD43, a molecule defective in Wiscott-Aldrich syndrome, binds ICAM-I zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA. Na- ture, 354, 233-235.
105. Delia, D., Lampugnani, M. G., Resnati, M., Dejana, E., Aiello, A., Fontanella, E., Soligo, D., Pierotti, M. A. and Greaves, M. F. (1993) CD34 expression is regulated recip- rocally with adhesion molecules in vascular endothelial cells in vitro. Blood, 81, 1001-1008.
106. Lawrence, M. B. and Springer, T. A. (1991) Leukocytes roll on selectin at physiologic flow rates: distinction from the prerequisite for adhesion through integrins. Cell, 65, 859- 873.
107. Mackay, C. R. and Imhof, B. (1993) Cell adhesion in the immune system. Immunol. Today, 14, 99-102.
108. Hogg, N. (1992) Roll, roll, your leukocyte gently down the vein. Immunol. Today, 13, 113-115.
109. Butcher, E. C. (1991) Leukocyte-endothelial cell recogni- tion: three (or more) steps to specificity and diversity. Cell,
110. To, L. B., Roberts, M. M., Haylock, D. N., Dyson, P. G., Branford, A. L., Thorp, D., Ho, J. Q. K. H., Dart, G. W. D., Horvath, N., Davy, M. L. J., Olweny, C. L. M., Abdi, E. and Juttner, C. A. (1992) Comparison of haematology recovery times and supportive care requirements of autolo- gous recovery phase peripheral blood stem cell transplants, autologous bone marrow transplants and allogeneic bone marrow transplants. Bone Marrow Transplant, 4, 277- 284.
111. Sheridan, W. P., Begley, C. G., Juttner, C. A., Szer, J., To, L. B., Maher, D., McCrath, K. M., Morstyn, G. and Fox, R. M. (1992) Effect of peripheral-blood progenitor cells mobilized by filgrastim (G-CSF) on platelet recovery after high dose chemotherapy. Lancer, 339, 640-644.
67, 1033-1036.
112. Siena, S., Bregni, M., Brando, B., Ravagnani, F., Bona- donna, G. and Gianni, A. M. (1989) Circulation of CD34' hematopoietic stem cells in the peripheral blood of high-dose cyclophosphamide treated patients: enhancement by intra- venous recombinant human granulocyte-macrophage colony- stimulating factor. Blood, 74, 1905-1914.
113. Kessinger, A. and Armitage, J. 0. (1991) The evolving role of autologous peripheral blood stem cell transplantation fol- lowing high dose therapy for malignancies. Blood, 77, 212- 213.
114. Gordon, M. Y., Dowding, C. R., Riley, G. P., Clark, D., Goldman, J. M. and Greaves, M. F. (1987) Altered adhesive interactions with marrow stroma of hematopoietic progenitor cells in chronic myeloid leukaemia. Nature, 328, 342-344.
115. Emerson, S. G., Guba, S. C., Upadhyaya, G. H., Deisser- oth, A. B. and Thomas, S. (1989) Chronic myelogenous leu- kaemia progenitor cells are deficient in cell surface LFA-3 and are not recognised by autoregulatory T lymphocytes. Clin. Res., 37, 901A.
116. Denkers, I. A., de Jong-de Boer, J. J., Beelen, R. H., Os- senkoppele, G. J. and Langenhuijsen, M. M. (1992) VLA molecule expression may be involved in the release of acute myeloid leukaemia cells from the bone marrow. Leuk. Res., 16, 469-474.
117. Inghirami, G., Wiecmrek, R., Zhu, B.-Y., Silber, R., Dalla- Favera, R. and Knowles, D. M. (1988) Differential expres- sion of LFA-1 molecules in non-Hodgkin's lymphoma and lymphoid leukaemia. Blood, 72, 1431-1434.
1 18. Kornberg, L., Earp, H. S., Turner, C., Prokop, C. and Juliano, R. L. (1991) Signal transduction by integrins: in- creased protein tyrosine phosphorylation caused by cluster- ing of beta 1 integrins. Proc. Natl. Acad. Sci. USA, 88, 8392- 8396.
119. Schwartz, M. A. (1993) Transmembrane signalling by in- tegrins. Trends Cell Biol., 2 , 304-308.
120. Juliano, R. L. and Haskill, S. (1993) Signal transduction from the extracellular matrix. J. Cell Biol., 120, 577-585.
121. Menko, A. S. and Boettiger, D. (1987) Occupation of the extracellular matrix receptor, integrin, is a control point for myogenic differentiation. Cell, 51, 51-57.
122. Schofield, R. (1978) The relationship between the spleen colony-forming cell and the haemopoietic stem cell. Blood Cells, 25, 195-202.
Downloaded By: [Queensland University of Technology] At: 00:08 18 February 2010
Related Documents











