Physiologia Plantarum 133: 651–669. 2007 Copyright ª Physiologia Plantarum 2007, ISSN 0031-9317 REVIEW Potassium transport and plant salt tolerance Sergey Shabala* and Tracey A. Cuin School of Agricultural Science, University of Tasmania, Private Bag 54, Hobart, Tasmania 7001, Australia Correspondence *Corresponding author, e-mail: [email protected] Received 8 October 2007 doi: 10.1111/j.1399-3054.2007.01008.x Salinity is a major abiotic stress affecting approximately 7% of the world’s total land area resulting in billion dollar losses in crop production around the globe. Recent progress in molecular genetics and plant electrophysiology suggests that the ability of a plant to maintain a high cytosolic K 1 /Na 1 ratio appears to be critical to plant salt tolerance. So far, the major efforts of plant breeders have been aimed at improving this ratio by minimizing Na 1 uptake and transport to shoot. In this paper, we discuss an alternative approach, reviewing the molecular and ionic mechanisms contributing to potassium homeostasis in salinized plant tissues and discussing prospects for breeding for salt tolerance by targeting this trait. Major K 1 transporters and their functional expression under saline conditions are reviewed and the multiple modes of their control are evaluated, including ameliorative effects of compatible solutes, poly- amines and supplemental calcium. Subsequently, the genetic aspects of inheritance of K 1 transport ‘markers’ are discussed in the general context of salt tolerance as a polygenic trait. The molecular identity of ‘salt tolerance’ genes is analysed, and prospects for future research and breeding are examined. Salinity as an issue The extent of the problem Global food production will need to increase by ap- proximately 50% by 2050 to match the projected population growth (Flowers 2004, Rengasamy 2006). At the same time, the most suitable land has already been cultivated, implying a need for either expansion into new areas to meet the above target or a dramatic increase in crop production on existing cultivated lands. Given the fact that about 15% of the total land area of the world has already been degraded by various factors, and the other half is ‘perennial desert’ or ‘drylands’ which can only be made more productive by irrigation (Flowers 2004), both tasks appear to be rather challenging. One of the major threats to this is soil salinity. Over 800 million hectares of land worldwide is affected by salinity (Munns 2005), comprising nearly 7% of the world’s total land area. Irrigation systems are particularly prone to salinization, with nearly one third of irrigated land being severely affected. Despite persistent questioning of the sustainability of irrigation as a method to increase food production (e.g. Flowers and Yeo 1995 and references within), it is highly unlikely that such practices will be stopped in the near future. Taking Australia as an example, the overall profit from irrigated crops is about $6 billion annually, which comprises 20– 25% of Australia’s gross value for agriculture, although irrigated land accounts for only 0.4% the country’s farmland (Haw et al. 2000). Thus, it can be predicted that such short-term economic benefits will prevail, despite the above concerns. Most crops are glycophytes, thus are not capable of growing in high concentrations of salt in the soil. Given the above trends, improving crop salt tolerance is fast becoming one of the key aspects of plant breeding in the future. However, all attempts so far towards improving crop salt tolerance through conventional breeding Abbreviations – CPA2, cation:proton antiporter family 2; CNGC, cyclic nucleotide-gated channels; iGluRs, glutamate receptors; NSCC, non-selective cation channels; KOR, outward-rectifying K 1 channel Physiol. Plant. 133, 2007 651

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Physiologia Plantarum 133: 651–669. 2007 Copyright ª Physiologia Plantarum 2007, ISSN 0031-9317
REVIEW
Potassium transport and plant salt toleranceSergey Shabala* and Tracey A. Cuin
School of Agricultural Science, University of Tasmania, Private Bag 54, Hobart, Tasmania 7001, Australia
Correspondence
*Corresponding author,
e-mail: [email protected]
Received 8 October 2007
doi: 10.1111/j.1399-3054.2007.01008.x
Salinity is a major abiotic stress affecting approximately 7% of the world’s total
land area resulting in billion dollar losses in crop production around the globe.
Recent progress in molecular genetics and plant electrophysiology suggests
that the ability of a plant to maintain a high cytosolic K1/Na1 ratio appears to
be critical to plant salt tolerance. So far, the major efforts of plant breeders havebeen aimed at improving this ratio by minimizing Na1 uptake and transport to
shoot. In this paper, we discuss an alternative approach, reviewing the
molecular and ionic mechanisms contributing to potassium homeostasis in
salinized plant tissues and discussing prospects for breeding for salt tolerance
by targeting this trait. Major K1 transporters and their functional expression
under saline conditions are reviewed and the multiple modes of their control
are evaluated, including ameliorative effects of compatible solutes, poly-
amines and supplemental calcium. Subsequently, the genetic aspects ofinheritance of K1 transport ‘markers’ are discussed in the general context of salt
tolerance as a polygenic trait. The molecular identity of ‘salt tolerance’ genes is
analysed, and prospects for future research and breeding are examined.
Salinity as an issue
The extent of the problem
Global food production will need to increase by ap-
proximately 50% by 2050 to match the projectedpopulation growth (Flowers 2004, Rengasamy 2006). At
the same time, the most suitable land has already been
cultivated, implying a need for either expansion into new
areas to meet the above target or a dramatic increase in
crop production on existing cultivated lands. Given the
fact that about 15% of the total land area of the world has
already been degraded by various factors, and the other
half is ‘perennial desert’ or ‘drylands’ which can only bemade more productive by irrigation (Flowers 2004), both
tasks appear to be rather challenging. One of the major
threats to this is soil salinity.
Over 800 million hectares of land worldwide is
affected by salinity (Munns 2005), comprising nearly
7% of the world’s total land area. Irrigation systems are
particularly prone to salinization, with nearly one third of
irrigated land being severely affected. Despite persistent
questioning of the sustainability of irrigation as a method
to increase food production (e.g. Flowers and Yeo 1995
and references within), it is highly unlikely that suchpractices will be stopped in the near future. Taking
Australia as an example, the overall profit from irrigated
crops is about $6 billion annually, which comprises 20–
25% of Australia’s gross value for agriculture, although
irrigated land accounts for only 0.4% the country’s
farmland (Haw et al. 2000). Thus, it can be predicted
that such short-term economic benefits will prevail,
despite the above concerns.Most crops are glycophytes, thus are not capable of
growing in high concentrations of salt in the soil. Given
the above trends, improving crop salt tolerance is fast
becoming one of the key aspects of plant breeding in the
future. However, all attempts so far towards improving
crop salt tolerance through conventional breeding
Abbreviations – CPA2, cation:proton antiporter family 2; CNGC, cyclic nucleotide-gated channels; iGluRs, glutamate receptors;
NSCC, non-selective cation channels; KOR, outward-rectifying K1 channel
Physiol. Plant. 133, 2007 651

programmes have met with very limited success, primar-
ily because of the physiological and genetic complexity of
this trait (Flowers 2004). In this review, we discuss some
of the reasons behind this, and suggest a new approach
of breeding plants for salt tolerance by targeting K1
homeostasis in plant tissues.
Physiological constraints and adaptation to salineenvironment
Salinity affects plant growth by imposing both ionic and
osmotic stresses. Because of the osmotic gradient
generated, elevated Na1 levels in the soil solution drives
water out of the cell (Fig. 1). To maintain the cytosolicwater activity (required for normal cell metabolism),
water moves from the vacuole into the cytosol to
compensate for such depletion. As a result, the cell turgor
is reduced almost instantaneously, and a substantial
reduction in leaf and root elongation rate is observed
within minutes after the onset of salt stress (Yeo et al.
1991). The growth is restored, albeit at a slower rate,after plant cells adjust to new osmotic conditions
(typically, approximately 1 h after stress onset; Munns
2002). Not only the root, but leaf tissues are also exposed
to this osmotic challenge as a result of the accumulation
of high Na1 concentrations in the leaf apoplast (Flowers
et al. 1991). Cell osmotic adjustment is, therefore, critical
to plant salt tolerance. Surprisingly, such osmotic
adjustment is usually taken for granted, and the essenti-ality of this trait is typically ignored by plant physiologists
who focus predominantly on Na1 exclusion from uptake
(Munns 2005). The specific mechanisms of cell osmotic
adjustment will be considered in detail in the section
Potassium and cell osmotic adjustment.
The second major constraint is the specific Na1
toxicity. Because of the similarity in physicochemical
properties between Na1 and K1 (i.e. ionic radius and ionhydration energy), the former competes with K1 for major
binding sites in key metabolic processes in the cytoplasm,
such as enzymatic reactions, protein synthesis and
ribosome functions (Marschner 1995). With over 50
cytoplasmic enzymes being activated by K1 (Marschner
1995), the disruption to metabolism is severe, both in root
and leaf tissues. Several mechanisms may reduce specific
Na1 toxicity. At the cellular level, restricted Na1 uptake,active Na1 exclusion back to the soil solution (via the
plasma membrane salt overly sensitive (SOS1) Na1/H1
antiporter; Zhu 2003) and compartmentation of excessive
Na1 in the vacuole (by the tonoplast Na1/H1 exchanger;
Zhang and Blumwald 2001) are considered to be central
to salt tolerance. There is also growing evidence that
a cell’s ability to repair damage, scavenge reactive oxygen
species (ROS) and protect sensitive cellular structures byincreased the synthesis of a wide variety of chaperone-
acting proteins, may be at least as important as the above
traits. At the whole-plant level, plant adaptation to salinity
may be achieved by traits such as reduced Na1 delivery to
the shoot, Na1 recirculation in the phloem, tissue- and
organ-specific compartmentation (such as preferential
Na1 loading into physiologically ‘unimportant’ struc-
tures, e.g. senescing leaves, leaf sheath or epidermis),and, in some species, by Na1 secretion from the leaf
through, e.g. specialized structures such as salt glands or
bladders (see Tester and Davenport 2003 for review).
Another important mechanism is the recirculation of Na1
from the photosynthetic organs back to the roots. There
are strong suggestions that this process is mediated
by high affinity K1 transporter (HKT) transporters
K+
K+,Na+
SKOR
NORC
KEK+
H+
H+
ATP
K+
H+
K+
HUK
NSCC
HKT
KIR
KOR
ATP
H+
Na+
K+, Na+
K+, Na+
HKT
KAT2
AKT2/3
GluR2
K+
K+, Na+
K+
Na+
K+, Na+
Na+
K+
Water
water
Na+
K+
STOP
STOP
Leaf
Root
Stele Epidermis
Fig. 1. Adverse effects of salinity on K1 homeostasis and water relations
in plants. Elevated Na1 levels in the soil solution drive water out of cell
following the osmotic gradient (1). Not only root, but also leaf tissues are
exposed to this osmotic challenge as a result of accumulation of high Na1
concentrations in the leaf apoplast (6). Salinity also substantially reduces
K1 availability to roots as a result of reduced K1 activity in the soil solution
(2). Na1 also competes with K1 for uptake sites at the plasmamembrane,
including both low-affinity (3) and high-affinity (4) transporters. Finally,
salinity dramatically increases K1 leak through depolarization-activated
outward-rectifying (KOR) K1 channels (5).
652 Physiol. Plant. 133, 2007

(Davenport et al. 2007, Rus et al. 2004). Given the fact
that these transporters are also involved in K1 uptake by
plant tissues (Rubio et al. 1995), it is not surprising that
HKT transporters have recently become a major target for
strategies in breeding for salt tolerance (Byrt et al. 2007).
Another major constraint is salinity-induced nutritionaldisorders and, specifically, K1 deficiency. First, high
concentrations of Na1 in the soil substantially reduce the
activity of many essential nutrients (including K1),
making them less available for plants (Fig. 1). Secondly,
Na1 competes with K1 for uptake sites at the plasma
membrane, including both low-affinity [e.g. non-selec-
tive cation channels (NSCC)] and high-affinity (e.g. HKT)
affinity transporters (Fig. 1; see also section Major targetsfor genetic improvement). Moreover, when positively
charged Na1 crosses the plasma membrane, a significant
membrane depolarization (by 60–80 mV for 100 mM
NaCl treatment; Shabala et al. 2003, 2005a) is observed.
Such depolarization makes passive K1 uptake through
inward-rectifying K1 channels thermodynamically im-
possible and, at the same time, dramatically increases K1
leak through depolarization-activated outward-rectifyingK1 channels (KOR) (Fig. 1). Finally, increased de novo
synthesis of various compatible solutes used for osmo-
protection under saline conditions (see section Potassium
and cell osmotic adjustment for details) severely reduces
the available ATP pool, making high-affinity K1 uptake
even more problematic.
Surprisingly, salinity-induced perturbations in K1
homeostasis are usually considered of ‘secondary impor-tance’ (Tester and Davenport 2003) and are often ignored
by most researchers. We believe that this is wrong and
argue here that overcoming this constraint may open up
new avenues for plant breeding for salt tolerance.
Improving salt tolerance: some principles anddogmas of modern breeding
Thus far, all attempts at improving salt tolerance, either
via traditional breeding or via marker-assisted selection or
genetic engineering, has focused along two major lines:
(1) breeding for better Na1 exclusion and (2) breeding for
better osmotic adjustment. Unfortunately, neither of these
approaches has, so far, resulted in salt-tolerant cultivars in
the farmers’ field (Flowers 2004).
The first approach was specifically advocated for wheatbreeding (Munns et al. 2006). According to Munns et al.
(1999), plants grown at 200 mM NaCl should exclude
over 97% of all Na1 presented at the root surface,
regardless of whether they are glycophytes or halophytes.
As Na1 is toxic to cell metabolism (see above), high
cytosolic Na1 is detrimental for plants. Unfortunately,
exclusion of Na1 from root uptake per se does not solve
the problem of the osmotic component of salt stress. The
existing osmotic gradient will keep driving water out of
the plant, reducing cell turgor, and slowing root and leaf
expansion growth. Moreover, such massive Na1 exclu-
sion may lead to a substantial and progressive build-up of
high Na1 concentrations in cell walls or near the rootsurface, further exacerbating the problem of the osmotic
component of salt stress. This problem may be especially
severe in perennial species. Thus, it is unlikely that such
an approach of breeding for Na1 exclusion only would
result in a significant improvement of plant performance
in saline environments. At the very least, a better Na1
excluding ability in plants should be complemented by
better osmotic adjustment. Literature analysis shows thatthe latter is rarely the case.
Moreover, the central dogma of a negative correlation
between Na1 accumulation and plant salt tolerance
(Munns 2002, 2005, Tester and Davenport 2003) does not
hold in many cases. The most obvious exception is
halophytes, capable of accumulating very large amounts
of Na1 (up to 50% of shoot dry weight) without dying.
One can argue that these species live longer thenglycophytes under saline conditions, hence, accumulate
more Na1 as a result. However, this argument is rebutted
by the fact that optimal halophyte growth is observed at
soil salinities ranging between 200 and 400 mM NaCl
(Khan et al. 2005), conditions at which most crops would
rapidly die. The Chenopodium family is another example
(Jacobsen et al. 2003). In glycophytes, no correlation
between Na1 accumulation and salinity tolerance wasfound in Arabidopsis, with salt overly sensitive mutants
(such as sos1) showing lower shoot Na1 content than the
wild-type (WT) (Ding and Zhu 1997). Maize and rice are
also two notable exceptions (reviewed in Tester and
Davenport 2003). In tomato, salt-tolerant Lycopersicon
peruvianum accumulates a much higher concentration of
Na1 than salt-sensitive Lycopersicon esculentum (Santa-
Cruz et al. 1999). Finally, recent screening of over 40varieties of Australian bread wheat has also found no
relationship between Na1 exclusion and salt tolerance in
this species (Genc et al. 2007).
The second approach to improve salt tolerance in
plants is to overexpress genes responsible for biosynthesis
of the so-called ‘compatible solutes’. These are organic
osmolytes that can accumulate in the cytosol in sub-
stantial quantities without interfering with cell metabo-lism. They are involved in both cell osmotic adjustment,
hence maintenance of normal turgor pressure, and in
osmoprotection of various membrane structures and
proteins. Osmolyte accumulation has long been empha-
sized as a selection criterion in traditional crop breeding
programmes (Ludlow and Muchow 1990, Morgan 1983).
Indeed, recent progress in molecular biology has made
Physiol. Plant. 133, 2007 653

this approach central to molecular breeding programmes,
largely because of the fact that osmolyte accumulation is
often controlled by only one gene (Serraj and Sinclair
2002), making it easy to manipulate. Approximately 13
species were transformed with nearly 40 genes between
1993 and 2003 (Flowers 2004), with most of the genesrelated to the biosynthesis of compatible solutes.
However, while a large body of data indicate a positive
correlation between proline accumulation and adapta-
tion to salt or drought stress (Delauney and Verma 1993),
this is not corroborated by other studies, particularly from
transgenic plants (Chandler and Thorpe 1987, Hassan
and Wilkins 1988). Differences between WT and trans-
genic plants are usually either small, restricted to a narrowdevelopmental stage, or achieved in non-physiological
(e.g. in the absence of transpiration) conditions. Several
more drawbacks such as the high energy cost of
biosynthesis of compatible solutes (between 40 and
50 mol ATP per 1 mol of compatible solute synthesized;
Raven 1985), and increased susceptibility to fungal dis-
eases and lodging (Bohnert and Shen 1999), negate the
beneficial effect of high osmolyte content on salt toler-ance. As a result, practical outcomes of this approach are
only marginal (Bajaj et al. 1999) and do not result in an
improved salt tolerance of plants under field conditions
(Flowers 2004).
Potassium homeostasis and plant salttolerance
Intracellular K1/Na1 ratio as a key determinantof salt tolerance
With Na1 toxicity occurring as a result of its competitionwith K1 for enzyme activation and protein biosynthesis, it
is obvious that it is not the absolute quantity of Na1 per se,
but rather the cytosolic K1/Na1 ratio that determines cell
metabolic competence and ultimately, the ability of
a plant to survive in saline environments. Indeed, the
cytosolic K1/Na1 ratio has been repeatedly named as
a key determinant of plant salt tolerance (Colmer et al.
2006, Cuin et al. 2003, Dvorak et al. 1994, Gaxiola et al.1992, Gorham et al. 1991, Maathuis and Amtmann
1999). Surprisingly though, not much direct experimental
evidence has been presented supporting this. Further-
more, there is, unfortunately, a tendency in the literature
to use the ratio between shoot (or leaf) tissue K1 and Na1
content as an equivalent of cytosolic K1/Na1 ratio
(Botella et al. 1997, Cramer et al. 1991). Such ‘surrogate’
estimates are rarely justified as they fail to take intoaccount the intracellular compartmentation of each of
these two ions. Indeed, because the vacuole occupies the
majority of the intracellular volume in most mature plant
cells, changes in tissue K1 concentration are largely
a reflection of the behaviour of K1 within that compart-
ment (Walker et al. 1996). It is, after all, the cytosolic K1
homeostasis rather than the vacuolar content that is
essential for plant metabolic processes (Maathuis and
Amtmann 1999, Marschner 1995, Walker et al. 1996).The major hurdle is a lack of appropriate and convenient
techniques to enable such compartmentational analysis.
So far, several methods including NMR, energy-dispersive
X-ray microanalysis and multi-barrelled microelectrodes
have been employed (reviewed by Shabala 2006), all
supporting the concept of the cytosolic K1 homeostasis.
The optimal cytosolic K1/Na1 ratio can be maintained
by either restricting Na1 accumulation in plant tissues orby preventing K1 loss from the cell. Plant breeders (Ashraf
and Khanum 1997, Garthwaite et al. 2005) and cell
physiologists (Tester and Davenport 2003) have focussed
almost exclusively on the former mechanism. This is
surprising considering the growing bulk of evidence
demonstrating that the ability of plant ability to retain K1
in the cytosol may be crucial in achieving increased salt
tolerance. Indeed, it has been shown that the difference insalt sensitivity between bread wheat (hexaploid, ABD
genomes) and durum wheat (tetraploid, AB genomes) may
be in enhanced K1/Na1 discrimination, a feature con-
trolled by a Kna1 locus on chromosome 4D (Dubcovsky
et al. 1996, Dvorak et al. 1994, Gorham et al. 1991).
Moreover, comparisons between Arabidopsis and its
salt-tolerant relative Thellungiella halophila have shown
that the latter is capable of actually increasing mesophyllK1 content under saline conditions, while Arabidopsis
shows a ‘classical’ decline (Volkov et al. 2003). This
suggests that mechanisms are in place within mesophyll
cells to ameliorate ionic changes, protecting and main-
taining their photosynthetic and metabolic activity under
saline conditions. This is consistent with recent studies in
our laboratory showing that improving tissue K1/Na1
ratios by externally applied divalent cations enablenormal leaf photochemistry in plants, even under high
(100 mM) salinity conditions (Shabala et al. 2005b).
Potassium and cell osmotic adjustment
As mentioned above, osmotic adjustment is an essential
attribute of plant salt tolerance. Under saline conditions,
Na1 and Cl2 are cheap osmotica that can be used tomaintain normal cell turgor, assuming they are efficiently
compartmentalized in the cell vacuole. However, as salt
levels increase in the vacuole, a substantial osmotic
potential gradient is established between the vacuole and
the cytosol, reducing water activity in the latter. This issue
needs to be managed by a concurrent increase of
osmolality in the cytosol to match the accumulation of
654 Physiol. Plant. 133, 2007
vacuolar Na1. There are two major avenues by which this
can be achieved. One is de novo synthesis of compatible
solutes, and the other, an increased uptake of inorganic
ions. The first avenue was discussed briefly in the section
Improving salt tolerance: some principles and dogmas of
the modern breeding. It should be added though, that theconventional view of increased concentrations of com-
patible solutes in the cytosol being responsible for water
retention, so providing the turgor necessary for cell
expansion (Delauney and Verma 1993, Zhang et al.
1999), has been strongly challenged over the last decade
for a number of reasons. First, the absolute osmolyte
concentrations in stressed plants are too low for an overall
osmotic adjustment (Hare et al. 1998). Secondly, theenergetic cost of osmolyte production is far too high
(Raven 1985). Thirdly, synthesis of compatible solutes is
a rather slow process, operating over a timescale of hours
and days, so cannot be responsible for the observed rapid
recovery of plant growth. Thus, a direct contribution of
compatible solutes to osmotic adjustment is highly un-
likely. Instead, other roles have been proposed for these
solutes such as osmoprotection of functional componentsthrough their stabilizing or chaperoning properties
(Bohnert and Shen 1999, Hare et al. 1998).
Given the relatively minor direct contribution of
organic osmolytes to cytosolic osmotic potential (Chen
and Murata 2002), combined with their primary location
within chloroplasts (Papageorgiou and Murata 1995) plus
the fact that cytosolic Na1 concentrations rarely exceed
30 mM, even under severe salinity stress (Tester andDavenport 2003), it leaves K1 to fulfil the role of the major
osmoticum used in cell osmotic adjustment. Indeed,
onset of hypertonic stress does lead to an increased
uptake of K1 in various plant tissues (Shabala et al. 2000,
Teodoro et al. 1998, Zingarelli et al. 1999). We have
previously shown in Arabidopsis root epidermal cells that
>90% of the cell turgor is recovered by inorganic ion
uptake within 40 min after the onset of hyperosmoticstress (Shabala and Lew 2002), in good agreement with
whole-plant data (Azaizeh et al. 1992). This leads to the
question of whether such osmotic adjustment can occur
under saline conditions.
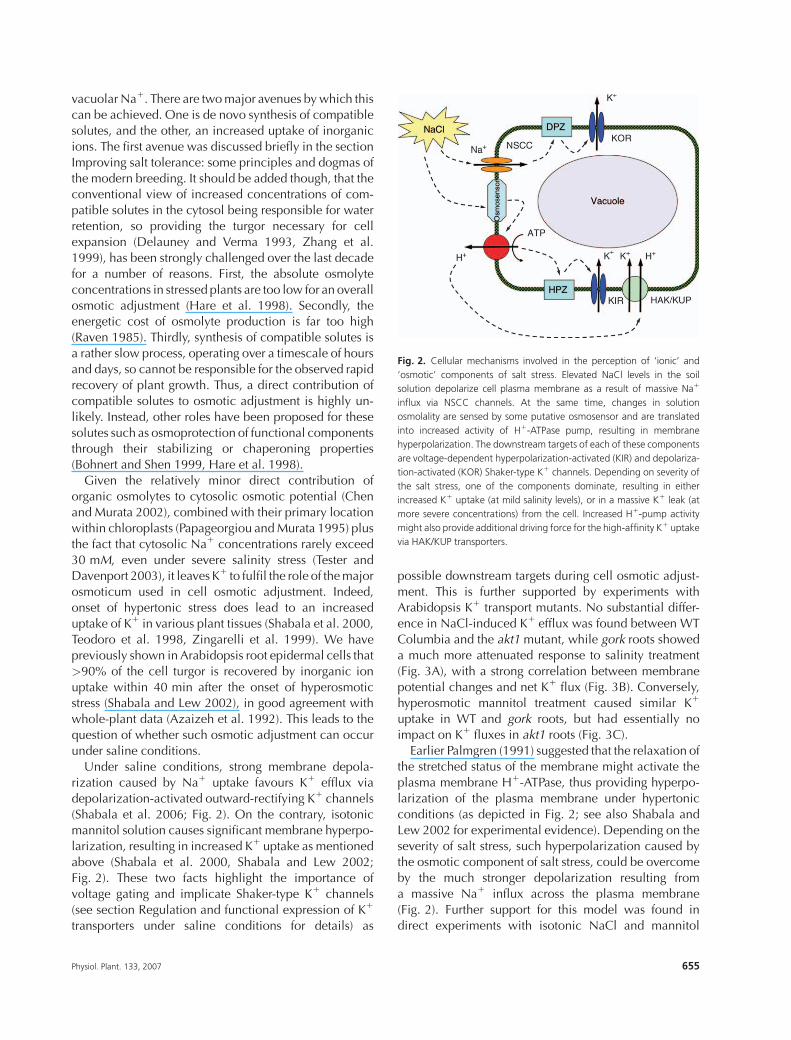
Under saline conditions, strong membrane depola-
rization caused by Na1 uptake favours K1 efflux via
depolarization-activated outward-rectifying K1 channels
(Shabala et al. 2006; Fig. 2). On the contrary, isotonicmannitol solution causes significant membrane hyperpo-
larization, resulting in increased K1 uptake as mentioned
above (Shabala et al. 2000, Shabala and Lew 2002;
Fig. 2). These two facts highlight the importance of
voltage gating and implicate Shaker-type K1 channels
(see section Regulation and functional expression of K1
transporters under saline conditions for details) as
possible downstream targets during cell osmotic adjust-
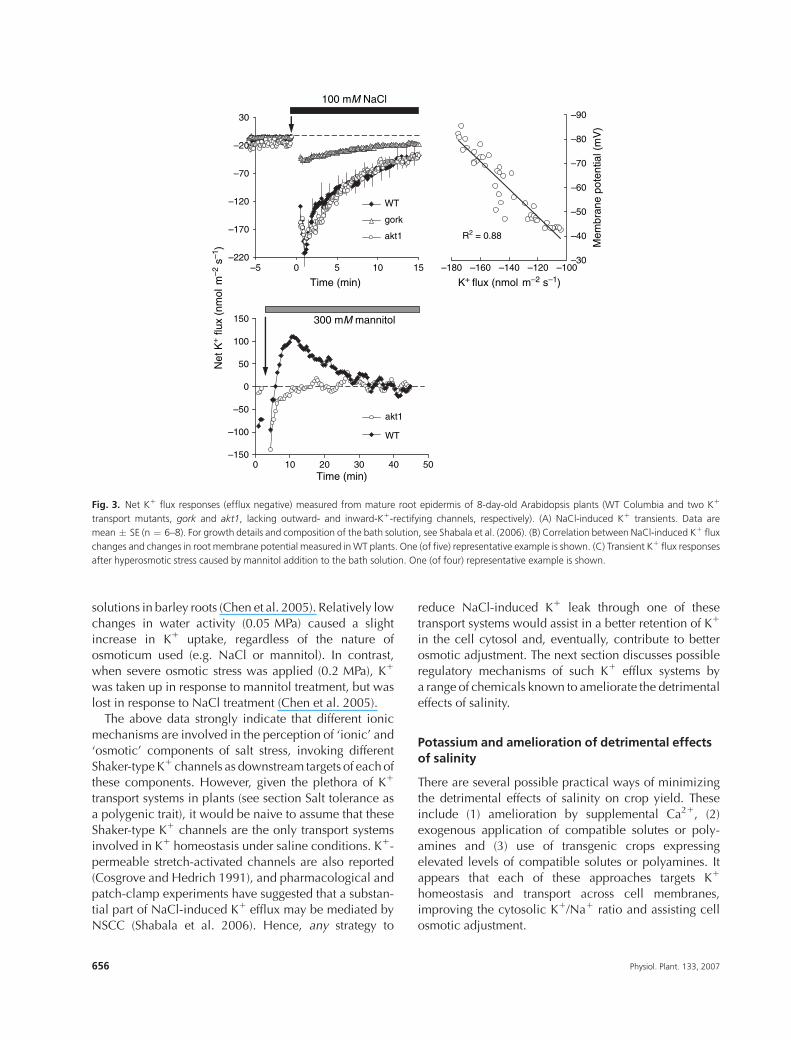
ment. This is further supported by experiments withArabidopsis K1 transport mutants. No substantial differ-
ence in NaCl-induced K1 efflux was found between WT
Columbia and the akt1 mutant, while gork roots showed
a much more attenuated response to salinity treatment
(Fig. 3A), with a strong correlation between membrane
potential changes and net K1 flux (Fig. 3B). Conversely,
hyperosmotic mannitol treatment caused similar K1
uptake in WT and gork roots, but had essentially noimpact on K1 fluxes in akt1 roots (Fig. 3C).
Earlier Palmgren (1991) suggested that the relaxation of
the stretched status of the membrane might activate the
plasma membrane H1-ATPase, thus providing hyperpo-
larization of the plasma membrane under hypertonic
conditions (as depicted in Fig. 2; see also Shabala and
Lew 2002 for experimental evidence). Depending on the
severity of salt stress, such hyperpolarization caused bythe osmotic component of salt stress, could be overcome
by the much stronger depolarization resulting from
a massive Na1 influx across the plasma membrane
(Fig. 2). Further support for this model was found in
direct experiments with isotonic NaCl and mannitol
Fig. 2. Cellular mechanisms involved in the perception of ‘ionic’ and
‘osmotic’ components of salt stress. Elevated NaCl levels in the soil
solution depolarize cell plasma membrane as a result of massive Na1
influx via NSCC channels. At the same time, changes in solution
osmolality are sensed by some putative osmosensor and are translated
into increased activity of H1-ATPase pump, resulting in membrane
hyperpolarization. The downstream targets of each of these components
are voltage-dependent hyperpolarization-activated (KIR) and depolariza-
tion-activated (KOR) Shaker-type K1 channels. Depending on severity of
the salt stress, one of the components dominate, resulting in either
increased K1 uptake (at mild salinity levels), or in a massive K1 leak (at
more severe concentrations) from the cell. Increased H1-pump activity
might also provide additional driving force for the high-affinity K1 uptake
via HAK/KUP transporters.
Physiol. Plant. 133, 2007 655
solutions in barley roots (Chen et al. 2005). Relatively low
changes in water activity (0.05 MPa) caused a slight
increase in K1 uptake, regardless of the nature ofosmoticum used (e.g. NaCl or mannitol). In contrast,
when severe osmotic stress was applied (0.2 MPa), K1
was taken up in response to mannitol treatment, but was
lost in response to NaCl treatment (Chen et al. 2005).
The above data strongly indicate that different ionic
mechanisms are involved in the perception of ‘ionic’ and
‘osmotic’ components of salt stress, invoking different
Shaker-type K1 channels as downstream targets of each ofthese components. However, given the plethora of K1
transport systems in plants (see section Salt tolerance as
a polygenic trait), it would be naive to assume that these
Shaker-type K1 channels are the only transport systems
involved in K1 homeostasis under saline conditions. K1-
permeable stretch-activated channels are also reported
(Cosgrove and Hedrich 1991), and pharmacological and
patch-clamp experiments have suggested that a substan-tial part of NaCl-induced K1 efflux may be mediated by
NSCC (Shabala et al. 2006). Hence, any strategy to
reduce NaCl-induced K1 leak through one of these
transport systems would assist in a better retention of K1
in the cell cytosol and, eventually, contribute to betterosmotic adjustment. The next section discusses possible
regulatory mechanisms of such K1 efflux systems by
a range of chemicals known to ameliorate the detrimental
effects of salinity.
Potassium and amelioration of detrimental effectsof salinity
There are several possible practical ways of minimizing
the detrimental effects of salinity on crop yield. These
include (1) amelioration by supplemental Ca21, (2)
exogenous application of compatible solutes or poly-
amines and (3) use of transgenic crops expressing
elevated levels of compatible solutes or polyamines. It
appears that each of these approaches targets K1
homeostasis and transport across cell membranes,improving the cytosolic K1/Na1 ratio and assisting cell
osmotic adjustment.
R2 = 0.88
–90
–80
–70
–60
–50
–40
–30–180 –160 –140 –120 –100
–220
–170
–120
–70
–20
30
–5 0 5 10 15
WT
gork
akt1
Net
K+ fl
ux (
nmol
m–2
s–1
)
Time (min) K+ flux (nmol m–2 s–1)
Mem
bran
e po
tent
ial (
mV
)
100 mM NaCl
Time (min)
–150
–100
–50
0
50
100
150
0 10 20 30 40 50
akt1
WT
300 mM mannitol
Fig. 3. Net K1 flux responses (efflux negative) measured from mature root epidermis of 8-day-old Arabidopsis plants (WT Columbia and two K1
transport mutants, gork and akt1, lacking outward- and inward-K1-rectifying channels, respectively). (A) NaCl-induced K1 transients. Data are
mean � SE (n ¼ 6–8). For growth details and composition of the bath solution, see Shabala et al. (2006). (B) Correlation between NaCl-induced K1 flux
changes and changes in root membrane potential measured inWT plants. One (of five) representative example is shown. (C) Transient K1 flux responses
after hyperosmotic stress caused by mannitol addition to the bath solution. One (of four) representative example is shown.
656 Physiol. Plant. 133, 2007

Application of supplemental Ca21 significantly ameli-
orates salinity stress symptoms in many species (Cramer
et al. 1987, Reid and Smith 2000, Shabala et al. 2003) and
is a widely used agricultural practice. For a long time,
such amelioration was attributed to Ca21 restriction of
Na1 uptake via NSCC: a major route for Na1 uptakeunder saline conditions (Apse and Blumwald 2007,
Demidchik and Tester 2002, Tester and Davenport
2003). Recently, we have provided evidence for an
additional mechanism of Ca21 action on salt toxicity in
plants: inhibiting Na1-induced K1 efflux through out-
wardly directed, K1-permeable channels (Shabala et al.
2006). Patch-clamp experiments have shown the pres-
ence of two populations of Ca21-sensitive K1 effluxchannels, showing different voltage dependence, activa-
tion kinetics and sensitivity to Na1. We have also shown
that not only Ca21 but also other divalent cations (Mg21,
Ba21, Zn21) may prevent K1 leakage in response to
salinity (Shabala et al. 2005b), thus assisting in maintain-
ing the high K1/Na1 ratio required for optimal plant
growth and leaf photosynthesis.
It has also been shown that salt-resistant plant varietiescontain higher polyamine levels under saline conditions
(Basu and Ghosh 1991, Erdei et al. 1990) and that their
exogenous application can ameliorate detrimental salin-
ity effects (Ndayiragije and Lutts 2006). However, the
underlying cellular mechanisms remain elusive. We have
recently shown that one of the modes of action of
polyamines may be through their blockage of NSCC
(Shabala et al. 2007). This reduces the magnitude ofNaCl-induced plasma membrane depolarization and, in
turn, the depolarization-induced K1 efflux through KOR.
In addition to this mechanism, polyamines may directly
block outward K1 currents through NSCC and activate
the plasma membrane H1-ATPase, so restoring the
membrane potential (Shabala et al. 2007). Once again,
cytosolic K1/Na1 ratio will be the main beneficiary of
such polyamine regulation of membrane transportactivity.
Molecular engineering of transgenic species over-
expressing genes responsible for biosynthesis of various
compatible solutes has long been, and remains, a major
avenue for improving plant salt tolerance (Bohnert et al.
1995, Bray 1997). In addition, numerous reports have
suggested that crop performance under salinity may be
improved by exogenous application of compatiblesolutes in the form of either aerial sprays or seed priming
(Harinasut et al. 1996). It has been proposed that the role
of compatible solutes in cytosolic osmotic adjustment
may be indirect through the plethora of regulatory or
osmoprotective functions (Hare et al. 1998, Hasegawa
et al. 2000). One such regulatory function appears to be
the maintenance of cytosolic K1 homeostasis by prevent-
ing NaCl-induced K1 leakage from the cell (Cuin and
Shabala 2005), most likely achieved through the
enhanced activity of the H1-ATPase. This would control
voltage-dependent outward-rectifying K1 channels and
create the electrochemical gradient necessary for sec-
ondary ion transport processes. Indeed, the activation ofproton pumps by salt stress is positively correlated with
salt tolerance (Golldack and Dietz 2001), and the effect is
stronger in salt-tolerant than in salt-sensitive species (Kefu
et al. 2003). Furthermore, such an increase in proton
pumping would, at the same time, provide the driving
force for a plasma membrane Na1/H1 exchanger to move
Na1 from the cytoplasm into the apoplast (Ayala et al.
1996), thereby reducing the cytosolic Na1 load. This,alongside increased K1 uptake, would further improve
the K1/Na1 ratio.
The above findings were further supported by experi-
ments testing 26 protein and non-protein amino acids (a
major class of compatible solutes) applied to plant roots at
physiologically relevant exogenous concentrations.
Twenty one of these caused a significant mitigation of
NaCl-induced K1 efflux (Cuin and Shabala 2007). It wasalso shown that the potential mitigating effect of amino
acids on salt-induced K1 efflux is dependent on the type
of amino acid supplied, shows strong dose dependency,
acts from the cytosolic side and correlates with the extent
of membrane depolarization. Evidence for the additive
effects of amino acids on NaCl-induced K1 efflux was
also presented, suggesting that it is not the increase in the
total amino acid content of plants per se but the actualratio between different types of amino acids that has the
mitigating effect on K1 homeostasis in salinized plant
tissues.
Our understanding of the processes behind the
mitigating effect of these compatible solutes is severely
limited and can be attributed to literally thousands of
components of a complex signalling network within the
cell. Only by showing the specific ionic and molecularmechanisms underlying their regulation and influence
over K1 (and Na1) transport will we be able to increase
the efficiency of the mode of action, possibly through
genetic engineering of salt-tolerant crops.
Breeding for salt tolerance by targetingK1 homeostasis
Salt tolerance as a polygenic trait
There is little doubt that salt tolerance is a complex
multigenic trait showing heterosis, dominance and
additive effects (Flowers 2004, Fooland 1997). Salt
tolerance is also multifaceted physiologically, with
numerous tissue- and age-specific components involved.
Physiol. Plant. 133, 2007 657

As such, salt tolerance will be determined by a number of
subtraits (specific for each particular species), any of
which might, in turn, be determined by any number
of genes. It is estimated that salinity affects the level of
transcription of approximately 8% of all genes (Tester and
Davenport 2003). To add to this complexity, fewer than25% of the salt-regulated genes are salt stress specific
(Ma et al. 2006), with a complex set of pathways observed
in response to salinity stress. Alterations in the regulation
of gene expression and metabolic adjustment in response
to salinity share common elements tootherabiotic stresses,
and it is very difficult (or even impossible – at least to our
current knowledge) to separate these components. Using
the microarray technique in the aforementioned study,Ma et al. (2006) analysed 1000 highly upregulated and
the 500 most downregulated salt-responsive genes,
grouped in 22 clusters, of which only one (!) contained
what could be called salt-specific upregulation. The latter
is, strictly speaking, an overstatement, as some moderate
induction under drought conditions was also observed.
And even in this case, the above cluster contained 171
genes, 60 of which had no known function! Not sur-prising, improving salt tolerance has always been, and
remains a great challenge for generations of breeders
(Flowers 2004, Flowers and Yeo 1995).
As a consequence of having a large number of
component traits of comparable importance conferring
salt tolerance, several points should be raised (Garcia
et al. 1997). First, no individual trait will necessarily
correlate with overall plant performance. Secondly,tolerance could be improved by a favourable recombi-
nation of the trait. Finally, selection should be for the
component trait and not for overall performance. The last
point is especially important, because most traditional
breeding is carried out based on plant vigour or yield.
Unsurprisingly, all attempts at improving crops’ salt
tolerance through conventional breeding programmes
have met with very limited success (Flowers 2004). Theprospect of changing a phenotype through genetic
manipulation or through conventional breeding is much
greater if one or a few defined regions of chromosome are
targeted rather than by generating a desired phenotype by
changing a large number of genes, each with a small
effect, scattered all over the genome (Koyama et al. 2001).
As discussed below, there is some cautious optimism that
this may be the case if breeding for salt tolerance isachieved through targeting K1 homeostasis.
NaCl-inducedK1 leak as ameasureof salt tolerance
Using seven barley cultivars contrasting in their salt
tolerance, a comprehensive study was undertaken to
correlate whole-plant responses to salinity (changes in
growth rate, biomass, net CO2 assimilation, chlorophyll
fluorescence, root and leaf elemental and water content)
and the ability of plant roots to retain K1 in the cytosol.
The latter trait was estimated by the magnitude of NaCl-
induced K1 efflux from root epidermis measured by non-
invasive ion selective microelectrodes (Chen et al. 2005).A very strong negative correlation (r2 > 0.8) was found
between the magnitude of K1 efflux from the root and salt
tolerance of a particular cultivar. The average K1 efflux
40 min after salt application ranged between 20 and
25 nmol m22 s21 for salt-tolerant cultivars, and between
150 and 180 nmol m22 s21 for salt-sensitive cultivars
(for 80 mM NaCl treatment). The effect showed a clear
dose dependence and was retained in seedlings ofdifferent ages (Chen et al. 2005). Thus, although hundreds
of different genes may be involved in conferring salt
tolerance, it appears that over 80% of the genetic
variability may be attributed to just one physiological
trait, namely a cell’s ability to prevent NaCl-induced K1
leak. Surprising as it may be, such an unexpectedly high
correlation may be explained by the fact that the observed
K1 efflux may be an integral characteristic of manydifferent factors controlling intracellular K1 homeostasis.
In other words, NaCl-induced K1 leak can be judged as
the ‘integrated’ measurement of tissue Na1 tolerance. It
is also important to emphasize that the difference in
the magnitude of K1 efflux between salt-tolerant and
susceptible genotypes was six-fold – much wider than the
genotypic dispersion for any of the other physiological
parameters measured (root Na1 content, 1.2-fold; flagleaf sap osmolality, 1.7; plant biomass, 2.2; net CO2
assimilation, 2.3) (Chen et al. 2005).
Further validation of K1 flux measurements as a screen-
ing tool for salt tolerance in barley was performed on
crosses between tolerant and sensitive barley cultivars
(Chen et al. 2005). The lowest K1 efflux was measured
from the two tolerant cultivars and the cross between
them, while the highest K1 efflux was observed fromsensitive cultivars and the cross between them. Crosses
between tolerant and sensitive cultivars showed inter-
mediate K1 efflux. These intermediate responses of F1s
suggest additive genetic control of salinity tolerance and
indicate inheritance of the K1 retention trait. The latter
issue is discussed in detail in the section Inheritance of K1
transport ‘markers’.
A large-scale trial was conducted to follow-up thiswork (Chen et al. 2007a). Nearly 70 barley cultivars of
various agronomic and genetic traits (e.g. winter vs
spring, feed vs malt, six-rowed vs two-rowed, husked vs
huskless, awned vs awnless) were grown to harvest
under saline conditions in a glasshouse over two con-
secutive years. In a parallel set of experiments, plant salt
tolerance was evaluated by non-invasive microelectrode
658 Physiol. Plant. 133, 2007

measurements of net K1 flux from roots of 3-day-old
seedlings of each cultivar following 1-h treatment in
80 mM NaCl. Sixty two of 69 cultivars followed an
inverse relationship between K1 efflux and salt tolerance
(Chen et al. 2007), with r2 ¼ 0.71 found between the
magnitude of K1 efflux and the overall ranking score(judged by six physiological traits such as relative grain
yield, shoot biomass, plant height, CO2 assimilation,
survival rate and thousand seed weight). Interestingly, the
few remaining ‘exceptions’ had a superior ability to
prevent Na1 accumulation in plant leaves, thus main-
taining a higher K1/Na1 ratio by this means. Therefore, it
appears that in 100% of plants, salt tolerance was
determined by the ability to maintain a higher K1/Na1
ratio; 90% of genotypes (62 of 69) achieved this by
retaining cytosolic K1, while about 10% (7 of 69)
achieved this by efficiently excluding Na1 from the
shoot. Taken together, our results show that a plant’s
ability to maintain high K1/Na1 ratio (either through
retention of K1 or by preventing Na1 from accumulating
in leaves) is a key feature for salt tolerance in barley, and
that in most genotypes this is achieved by K1 retentionrather then Na1 exclusion. This questions the conven-
tional view of Na1 exclusion being the major avenue to
optimize intracellular K1/Na1 ratio (Munns 2005, Tester
and Davenport 2003). Furthermore, it is consistent with
reports of Dvorak et al. (1994) who found that a novel
tetraploid wheat germplasm with enhanced K1/Na1
discrimination (via Kna1 locus) showed leaf Na1 con-
centrations not significantly different from its durumparents, whereas K1/Na1 ratios were substantially higher.
It also appears that similar patterns are characteristic in
other species (e.g. lucerne, unpublished results from our
laboratory). A large-scale experiment on other cereals
(durum and bread wheat) is currently in progress to
validate this concept.
Inheritance of K1 transport ‘markers’
Wide hybridization remains a false hope in terms of
practical breeding, as the damage to the crop genome by
alien genetic material almost always outweighs any
advantage (if there was one) of introducing alien genes
in the first place (Flowers and Yeo 1995). Therefore,
pyramiding component physiological traits remains the
most efficient avenue to improve plant salt tolerance,assuming such traits are inheritable.
Unexpectedly, there have been only a limited number
of studies in which the heritability of salt tolerance have
been reported, as well as very few measurements of the
heritability of particular traits for which the underlying
physiological or anatomical basis is understood (Garcia
et al. 1997). Indeed, most breeders focus on such
unspecific characteristics as seed germination (Mano
and Takeda 1997) or shoot or root growth (Ashraf et al.
1986). Nonetheless, high heritability of salt tolerance
based on Na1 and K1 uptake and the K1/Na1 ratio has
been reported (Gregorio and Senadhira 1993). K1/Na1
discrimination has also been a subject of QTL analysis forsalt tolerance in some cereals (Koyama et al. 2001,
Lindsay et al. 2004), and these reports have suggested that
the overall K1/Na1 ratio is heritable, although not in all
species (Garcia et al. 1997).
The genetics of salt tolerance is rather complex, with
additive, dominance and overdominance gene effects
reported (Flowers 2004). For example, Gregorio and
Senadhira (1993) found that the Na1/K1 ratio in rice wasgoverned by both additive and dominance gene effects.
In addition, Mano and Takeda (1997) found that salt
tolerance of barley at the germination stage was con-
trolled by overdominant alleles, and non-additive genetic
variance was larger than additive genetic variance.
In contrast, at the seedling stage, tolerance was pre-
dominantly controlled by additive genes, with also some
effects of dominance. The extent to which K1 transport‘markers’ are inherited, and which of above patterns are
attributed to this trait, remains to be answered.
A half diallel cross was made in our laboratory between
six barley cultivars contrasting in salt tolerance to test the
genetic model and also to study the inheritance of the K1
efflux trait in plant roots under saline conditions.
Significant differences were found between different
parents or F1 populations. The average values of root K1
flux of F1 progenies were very close to the midparent
values, while those of F2 populations were significantly
lower (i.e. greater efflux) than the midparent values,
indicating the dominance of salt tolerance (Z. Chen,
M. Zhou, N. Mendham, I. Newman and S. Shabala,
University of Tasmania, Hobart, unpublished data).
Variance and covariance analysis showed the existence
of epistatic effects, which was confirmed by further testsusing six different populations (parents, F1, F2, BC1 and
BC2) from two different crosses. However, the tolerance
was controlled mainly by additive effects, with relatively
smaller contributions from dominant and epistatic effects.
Based on a single MIFE (UTas Innovation, Hobart,
Australia) measurement, the estimated heritability was
relatively low (between 0.73 and 0.82). Using the average
values from around 10 measurements, the heritability ofthe trait increased dramatically, being 0.97, 0.98 and
0.97, respectively, from ANOVA of parents and F1s, diallel
analysis of F1s and diallel analysis of F2s (Z. Chen,
M. Zhou, N. Mendham, I. Newman and S. Shabala,
unpublished data). Such extremely high heritability of
NaCl-induced root K1 flux may also be the result of the
relatively smaller environmental effects of the MIFE
Physiol. Plant. 133, 2007 659

measurement compared with the many factors affecting
field experiments. Overall, our results confirm a high
heritability for salt tolerance in barley and show that
plants could be effectively selected and bred for salt
tolerance based on the NaCl-induced root K1 flux
measurements.
Molecular identity of ‘salt tolerant’ genescontrolling potassium homeostasis
Regulation and functional expression of K1
transporters under saline conditions
Potassium transport across plant membranes is mediated
by at least seven major families of cation transporters (75
genes in total in Arabidopsis). As this topic has been
extensively reviewed over the last few years (Cherel 2004,
Cuin and Shabala 2006, Gierth and Maser 2007, Lebaudyet al. 2007, Maser et al. 2001, Very and Sentenac 2002,
2003), only a brief summary is given here; the major
emphasis is on their functional expression under saline
conditions.
In general, two major groups are distinguished:
(1) three families of potassium permeable channels and
(2) three families of potassium transporters. The first group
includes (a) Shaker-type potassium channels (9 genesin Arabidopsis), (b) ‘two-pore’ potassium channels
(6 genes), as well as (c) NSCCs. The latter are further split
into (i) cyclic nucleotide-gated channels (CNGC)
(20 genes) and (ii) glutamate receptors (iGluRs) (20 genes).
The second group (potassium transporters) consists of
(d) KUP/HAK/KT transporters (13 genes), (e) HKT trans-
porters (1 gene), and (f) K1/H1 antiporters (6 genes). All
these show rather complex tissue- and organ-specific
expression (Fig. 4).
Shaker-type potassium channels
Shaker-type potassium channels are located at the plasma
membrane and mediate most of the K1-selective voltage-
gated currents at both hyperpolarized and depolarized
membrane potentials (Very and Sentenac 2002). These
channels are expressed in various cell types and operatein the millimolar range (low-affinity passive transport).
Based on their voltage dependency, the channels are
grouped into three functional subfamilies: (1) hyper-
polarization-activated inward-rectifying channels (AKT1,
KAT1, KAT2 and SPIK), (2) weakly inward-rectifying
channels (AKT2/3) and (3) depolarization-activated out-
ward-rectifying channels (SKOR and GORK). The phys-
iological roles and functional expression of the Shakerfamily channels are diverse. Both AKT1 and AtKC1 are
expressed in roots (Bertl et al. 1994, Ivashikina et al.
2001, Pilot et al. 2003), with AKT1 being a major player in
K1 acquisition by the plant (Hirsch et al. 1998). The
function of AKT2/AKT3 (which enables bi-directional K1
transport) is attributed to phloem loading and/or unload-
ing (Lacombe et al. 2000, Marten et al. 1999) and K1
Fig. 4. Tissue-specific expression of K1 transporters in plants. Most of the data come from experiments with Arabidopsis, although results from other
plant species are also included. With the exception of KCO channels, all other transporters shown are likely to be expressed at cell plasma membrane.
Reproduced from Cuin and Shabala (2006), with kind permission of Springer Science and Business Media.
660 Physiol. Plant. 133, 2007

uptake into leaf mesophyll cells (Dennison et al. 2001).
KAT1 and KAT2 are guard cell-specific channels mediat-
ing K1 influx for turgor-dependent regulation of stomatal
aperture (Szyroki et al. 2001). Potassium release from
most cells (including guard cells, root hairs and root
epidermis) is mediated by the depolarization-activatedGORK channel (Ivashikina et al. 2001), while K1 release
into the xylem occur through the SKOR channel
(Gaymard et al. 1998).
There are a number of reports of adverse effects of Na1
on the functioning of Shaker channels. Sodium may have
both direct (reducing a channel’s open probability) and
indirect (reducing the total number of expressed chan-
nels) effects on the AKT channel (Qi and Spalding 2004).Reports have shown that the AKT2/3 K1 channel is
upregulated during salinity (Marten et al. 1999), suggest-
ing that AKT2/3 may be involved in the recirculation of K1
through the phloem. Similarly the stelar-root tissue-
located SKOR is also upregulated by salinity (Maathuis
2006). The substantial salt-induced upregulation of both
SKOR in roots and AKT2/3 in shoots would result in
increased rates of K1 circulation through the vasculartissue, pointing towards a long distance redistribution of
K1 between the roots and shoots. Interestingly, the
opposite, i.e. a downregulation of SKOR and AKT2/3
occurs during K1 deprivation (Pilot et al. 2003).
Other Shaker-type channels though may have a much
larger role in K1homeostasis under saline conditions. The
large Na1 influx that occurs upon the subjection of plants
to salinity results in membrane depolarization (Babourinaet al. 2000, Horie et al. 2001, Shabala et al. 2003). Such
a voltage change would result in the activation of
depolarization-activated outward-rectifying K1 chan-
nels. Indeed, we have found, using K1-specific channel
blockers, that the massive K1 efflux resulting from salt
application is mediated through such channels in both
Arabidopsis (Shabala et al. 2006) and barley (Z. Chen,
S. Shabala and I. Pottosin, unpublished data). Further-more, measurements on a variety of K1 transporter
mutants have shown that this efflux is strongly reduced
in Arabidopsis gork1 mutants (Fig. 3A). As this NaCl-
induced efflux corresponds to decreases in cellular K1
content (Shabala et al. 2006), it strongly implicates a role
for GORK in plant salt tolerance.
‘Two-pore’ potassium channels
‘Two-pore’ potassium channels have two families in
Arabidopsis (KCO-1P and KCO2-P), with one and five
members in each, respectively (Czempinski et al. 2002).
These channels appear to be located predominantly at
tonoplast and show strong Ca21 dependence and out-
ward rectification (Schonknecht et al. 2002). Although
the involvement of KCO channels in plant adaptive
responses to salinity is mostly unknown, a recent report
has indicated that the transcript level of the K1-selective,
largely voltage-independent, vacuolar-located TPK5
(KCO5) decreases under saline conditions, possibly
signifying its involvement in maintaining cytoplasmicK1 levels and/or the exchange of vacuolar K1 for Na1
(Maathuis 2006).
Non-selective cation channels
NSCCs are ubiquitous in both plasma and endomem-
branes (specifically, tonoplast) of plant cells, with 40
putative NSCCs shown in the Arabidopsis genomesequence (Maser et al. 2001, Very and Sentenac 2002,
2003). NSCCs show a high selectivity for cations over
anions but, as a rule, do not discriminate strongly
between cations, and have K1/Na1 selectivity ratios
between 0.3 and 3 (Demidchik and Maathuis 2007,
Demidchik et al. 2002). The physiological role of NSCCs
is to function in low-affinity uptake. NSCCs may be gated
by a large number of factors and may be classified asdepolarization-activated, hyperpolarization-activated,
voltage-insensitive, calcium-activated, mechanosensi-
tive, cyclic nucleotide-gated and glutamate-gated chan-
nels (Demidchik et al. 2002). The last two groups are
considered in more detail below.
It is well accepted that NSCC are the prime mediators of
Na1 influx into plant roots (Demidchik and Maathuis
2007, Demidchik et al. 2002, Maathuis and Sanders2001, Tyerman et al. 1997), and transcript levels of a
number of members of this family are modified by salinity
(Maathuis 2006). This may be beneficial during salt stress
because it will help decrease the osmotic potential of
tissues, thereby alleviating water stress. However, if the
excess Na1 is not compartmentalized into the vacuole,
the result can be severe toxicity in the cytosol.
Recently, work in our laboratory has implicated NSCCin mediating at least a part of the NaCl-induced K1 efflux
from the cell, alongside K1 outward-rectifying channels
(Shabala et al. 2006). However, the full contribution of
NSCC towards K1 acquisition and transport within plants
under saline conditions remains to be quantified.
CNGCs are ligand-gated channels that are activated by
either cAMP or cGMP (Leng et al. 2002), resulting in an
influx of cations into the cell (Balague et al. 2003). Thesechannels are widely present across the plant kingdom,
with 20 members of the CNGC family found in
Arabidopsis genome (Maser et al. 2001). In contrast to
animal CNGCs, plant CNGCs are also gated by calcium
and calmodulin (Maser et al. 2001, Very and Sentenac
2002). Being classified as non-selective, they have almost
equal permeability for K1 and Na1 (Balague et al. 2003).
Physiol. Plant. 133, 2007 661

CNGCs are expressed ubiquitously in all tissues (Talke
et al. 2003) and are localized to the plasma membrane
(Arazi et al. 1999).
The large size of the plant CNGC family indicates
a wide diversity of physiological functions, ranging from
cell signalling (Very and Sentenac 2002) to root cationuptake (Maathuis and Sanders 2001). Their contribution
towards K1 acquisition and transport within plants under
saline conditions remains to be quantified. Notwith-
standing, for several members of the CNGC family,
functional data are available (Balague et al. 2003, Leng
et al. 2002). All characterized CNGCs are capable of
conducting K1 and, apart from CNGC2, Na1 as well.
Transcriptome analysis has revealed that transcript levelsfor CNGC1, CNGC19 and CNGC20 increased under
saline conditions in Arabidopsis roots, while in shoots,
CNG19, CNGC3 and CNGC8 transcript levels were
increased (Maathuis 2006). However, the significance of
this is yet to be revealed.
iGluRs are expressed ubiquitously throughout the
plants, with some of them being most abundant in root
tissues (Davenport 2002). The precise physiological roleof the plant glutamate receptor family is unknown
(Lacombe et al. 2001), and it remains to be proven
whether GluRs are actually capable of forming functional
ion channels. GluRs3.2 overexpression creates hyper-
sensitivity to K1 and Na1 (Kim et al. 2001). Thus,
glutamate receptors (GLR) activity during salt stress may
not only influence Na1 fluxes per se, but also affect K1
nutrition. In contrast to GLR2.3, GLR2.5 is downregu-lated in root tissue, possibly pointing towards a function
in the redistribution of ions such as K1 (Maathuis 2006).
KUP/HAK/KT transporters
KUP/HAK/KT transporters are involved in both high- and
low-affinity K1 uptake (Santa-Marıa et al. 1997, Vallejo
et al. 2005). They form a large family, with 13 members inArabidopsis (Maser et al. 2001), at least 17 members in
rice (Banuelos et al. 2002) and 5 in barley (Santa-Marıa
et al. 1997). They are ubiquitously expressed in plant
tissues, functioning both at the plasma membrane and the
tonoplast (Banuelos et al. 2002, Serrano and Rodrıguez-
Navarro 2001).
While the KUP/HAK/KT family of K1 transporters might
mediate some low-affinity Na1 influx at high externalNa1 concentration, the full extent of this is not known.
Nevertheless, the transcript levels and activity of a num-
ber of these transporters from this group are modified
under saline conditions, thus could be targets for genetic
improvement. When expressed in yeast, HvHAK1 from
barley mediates low-affinity Na1 transport in addition to
high-affinity K1 uptake (Santa-Marıa et al. 1997).
Elevated Na1 also inhibits K1 transport through heterol-
ogously expressed AtKUP1 (Fu and Luan 1998) and
AtHAK5 (Rubio et al. 2000) and transcript expression of
KUP2 is decreased in shoots of NaCl-treated plants
(Maathuis 2006). Also reported is the upregulation
of Mesembryanthemum crystallinum McHAK1 andMcHAK2 under both K1 starvation and NaCl stress (Su
et al. 2002). These isoforms are proposed to be involved in
maintaining cytoplasmic K1 levels and/or turgor regula-
tion under conditions where external Na1 inhibits K1
uptake and cellular Na1 replaces K1.
HKT transporters
The main function of plant HKT transporters is to facilitate
K1/Na1 symport under high-affinity conditions (Rubio
et al. 1995, Schachtman and Schroeder 1994). Further
studies have suggested that this transporter may also
mediate low-affinity Na1 transport into roots (Laurie et al.
2002), especially under low K1 to Na1 ratios (Rubio et al.
1995). The HKT transporters are proposed to play a major
role in Na1 uptake and recirculation within salt-stressedplants (Apse and Blumwald 2007, Rus et al. 2001).
The HKT family of proteins has been implicated in Na1
transport in a number of species. Members of this family
have been shown to function as Na1/K1 symporters and
as Na1-selective transporters at both high and low affinity
(Garciadeblas et al. 2003, Rubio et al. 1995). Certainly,
decreased expression of HKT under salt stress often
correlates with plant salt tolerance (Golldack et al. 1997),suggesting that this transporter may be a determinant of
salt sensitivity in plants (Rubio et al. 1995). However, its
role in salt tolerance is far from resolved. The rice
OsHKT1:5 functions as a Na1-selective transporter in
oocytes and is hypothesized to control shoot Na1 and
influence shoot K1 by withdrawing Na1 from the xylem
stream into the xylem parenchyma cells (Ren et al. 2005).
A similar recirculating role has been proposed inArabidopsis for AtHKT1:1 in mediating the loading of
Na1 from the shoot into the phloem and then unloading it
into the roots (Berthomieu et al. 2003, Sunarpi et al.
2005). HKT has been also implicated in the regulation of
K1 transport (Uozumi et al. 2000) and homeostasis (Ren
et al. 2005, Sunarpi et al. 2005) under saline conditions
in several species.
K1/H1 antiporters
The Arabidopsis genome contains six putative K1 efflux
antiporters (named KEA1 through KEA6) that belong to
the monovalent cation:proton antiporter family 2 (CPA2
family). Overall, three major families of such antiporters
are known: CPA1 (a monovalent cation:proton antiporter
662 Physiol. Plant. 133, 2007

family; eight members in Arabidopsis), CPA2 or cation/
proton exchanger family [(CHX), 28 members] and NhaD
(two members) (Maser et al. 2001). CPA1 transporters
appear to be crucial for regulating cytosolic sodium level.
The role of these transporters in K1 homeostasis is poorly
understood, although there are suggestions that thetonoplast-located Na1/H1 antiporter also mediates K1
transport (Apse et al. 2003, Zhang and Blumwald 2001).
Cellier et al. (2004) suggested that at least one member
of CPA2 family: the root and leaf-expressed AtCHX17,
may contribute towards K1 acquisition and homeostasis
under saline conditions, as its transcript level increases
under salinity (Maathuis 2006). However, in chx17
mutants, no difference in Na1 content was observedfrom WT plants when grown under saline conditions,
while the K1 levels decreased in the mutant. This
indicates that CHX17 transports K1 rather than Na1
(Cellier et al. 2004), suggesting its involvement in root K1
homeostasis, particularly during NaCl stress.
Members of the CHX family also play a well-known
and significant role in cytosolic Na1 extrusion from
the cell. These transporters display Na1/H1 antiporteractivity, both at the plasma (AtSOS1; Shi et al. 2000) and
vacuolar (AtNHX1; Apse et al. 1999, 2003) membranes.
Overexpression of both these transporters has been
shown to improve salt tolerance in several plant species
(Blumwald et al. 2000, Shi et al. 2003). In addition,
CHX21 has been functionally characterized as a Na1
transporter in the xylem parenchyma (Hall et al. 2006).
Other transporters
Two other types of transporters may potentially contribute
to regulation of cellular ionic and osmotic homeostasis
under saline conditions. These include the wheat LCT1
low-affinity transporter mediating uptake of a wide range
of monovalent cations, including K1 and Na1 (Amtmann
et al. 2001) and a putative members of the cation chlo-ride co-transporter family (Very and Sentenac 2003,
Colmenero-Flores et al. 2007).
Major targets for genetic improvement
Given the fact that plant salt tolerance correlates with its
ability to retain K1 in the cell and that NaCl-induced K1
efflux is mediated predominantly by GORK channels (seeabove), it would be very tempting to suggest that knocking
outGORK genes would increase salt tolerance. However,
given the multiple roles of these channels in plants, any
benefits of such modification may be outweighed by the
potential numerous physiological disturbances caused by
such mutation. Consistent with this notion, no substantial
difference in the K1-permeable outward-rectifying
channel (e.g. GORK) density was found between two
contrasting barley cultivars, CM72 and Gairdner, show-
ing a six-fold difference in their ability to retain K1 after
salt imposition (Z. Chen, S. Shabala, I. Pottosin, unpub-
lished results). Therefore, it appears to be the ability of
a cell to regulate a channel’s activity as opposed to thenumber of expressed channels per se that determines
plant salt tolerance. Hence, factors contributing to post-
transcriptional regulation of GORK channels should be
targeted. Recent work in our laboratory has explicitly
proven that multiple mechanisms determine salt toler-
ance in barley. These mechanisms include (Chen et al.
2007b): (1) better control of membrane voltage, so re-
taining a more negative membrane potential, (2) intrin-sically higher H1 pump activity, (3) enhanced ability of
root cells to pump Na1 from the cytosol to the external
medium and (4) higher sensitivity to supplemental Ca21.
The first two mechanisms appear to be closely linked
and explain most of genetic variation in the ability to
retain K1 in barley roots. Importantly, plasma membrane
H1 ATPase activity is under control of relatively few genes
(Gaxiola et al. 2007), making this trait relatively easy tomanipulate by molecular means. The potential drawback
of increased intrinsic high H1 pump activity would be an
unproportionally high amount of energy drawn from
other metabolic process, resulting in possible penalties
in terms of reduced growth rate and/or yield. Theoretically,
this problem may be overcome by introducing stress-
specific promoters, causing post-translational activation of
H1-ATPase activity only under stress conditions.It would be naive and misleading to assume that the
above modification of H1-ATPase activity leading to an
improved ability of plant to retain K1, as the only possible
way of improving salt tolerance. Without any doubt, the
latter trait should be complemented by other mechanisms
proven to also significantly contribute to tolerance.
Among these, overexpression of the tonoplast NHX1
H1/Na1 exchanger is probably the most important. Thiswould allow plants to use Na1 as a cheap osmoticum,
pumping it into vacuole, thus maintaining cell turgor. At
the same time, increased vacuolar Na1 would allow the
available K1 to be used in the cytosol rather than stored in
the vacuole for osmotic purposes. The importance of Na1
compartmentation to the vacuole by overexpressing
NHX1 has been shown by improved performance in
salt-stressed transgenic tomato (Zhang and Blumwald2001), Brassica (Zhang et al. 2001), rice (Fukuda et al.
2004), wheat (Saqib et al. 2005, Xue et al. 2004) and
cotton (Wu et al. 2004). Additional evidence supporting
the role of vacuolar transport in salt tolerance has been
provided by Arabidopsis plants overexpressing a vacuolar
H1-PPase (Gaxiola et al. 2001). In this work, transgenic
plants overexpressing AVP1 coding for the vacuolar
Physiol. Plant. 133, 2007 663

H1-PPase showed enhanced salt tolerance that was cor-
related with the ionic K1 content of the plants.
Importantly, the tonoplast-located Na1/H1 antiporter
also mediates K1 transport (Apse et al. 2003, Zhang and
Blumwald 2001), so might assist in increasing the
vacuolar K1 pool under control conditions, making itavailable when plants are hit by salt stress.
Another possible target could be the homologue of
the yeast HAL genes. Overexpressing HAL1 and HAL3
genes improved salt tolerance by increasing the cellular
K1/Na1 ratio (Gaxiola et al. 1992, Serrano 1996). Both
a reduction in K1 loss and a decrease in intracellular Na1
accumulation were observed. Genes homologous to the
yeast HAL could also be present in higher plants and maybe relevant to salt tolerance (Bordas et al. 1997, Espinosa-
Ruiz et al. 1999). However, the relationship between
these genes and K1 homeostasis in plants is unknown and
needs further study.
Conclusions and prospects
Our understanding of the importance of K1 transportsystems and its cytosolic homeostasis under saline
conditions has increased considerably in recent years.
Indeed, it is becoming more and more accepted that the
ability of a plant to maintain a high cytosolic K1/Na1 ratio
is crucial in plant salt tolerance mechanisms. In spite of
this, only a small number of genes responsible for K1 as
well as for Na1 transport have been fully characterized
physiologically. Furthermore, the majority of thesestudies on plants have been mostly carried out on the
model Arabidopsis species. The extension of some of this
work to the salt-tolerant Arabidopsis-related species
Thellungiella halophila opens up prospects for extending
such work onto a halophytic species, although, neither
this species nor Arabidopsis are useful agriculturally.
Research in our laboratory conducted over the last few
years has significantly contributed to understanding thegenetic aspects of the inheritability of cytosolic K1
homeostasis on the agriculturally important barley crop
and have highlighted the crucial importance of main-
taining an optimal cytosolic K1/Na1 ratio in salt tolerance
mechanisms (Chen et al. 2005, 2007). This research is
currently being extended to wheat. The recent sequenc-
ing of the rice genome and a progressive build-up of
genomic information regarding other crop species willundoubtedly facilitate our understanding of potential
transport mechanisms for K1 within plants, enabling the
application of information concerning K1 and Na1
transport gained from Arabidopsis to economically
important plant species. This will allow the pinpointing
of particular genes in the control of K1 homeostasis under
adverse environmental conditions. Although much is still
to be discovered concerning the mechanisms of the
maintenance of optimal K1/Na1 ratios under saline
conditions, the research outlined in this review does
allow us a certain amount of optimism regarding the
prospect of developing plants with increased salt toler-
ance that are useful to the farmer, enabling an increase inarable production under salt-stressed conditions.
Acknowledgements – This work was supported by ARS
Discovery and GRDC grants to Dr S.Shabala.
References
Amtmann A, Fischer M, Marsh EL, Stefanovic A, Sanders D,
Schachtman DP (2001) The wheat cDNA LCT1 generates
hypersensitivity to sodium in a salt sensitive yeast strain.
Plant Physiol 126: 1061–1071
Apse MP, Blumwald E (2007) Na1 transport in plants. FEBS
Lett 581: 2247–2254
Apse MP, Aharon GS, Snedden WA, Blumwald E (1999)
Salt tolerance conferred by overexpression of a vacuolar
Na1/H1 antiport in Arabidopsis. Science 285: 1256–1258
Apse MP, Sottosanto JB, Blumwald E (2003) Vacuolar cation/
H1 exchange, ion homeostasis, and leaf development are
altered in a T-DNA insertional mutant of AtNHX1, the
Arabidopsis vacuolar Na1/H1 antiporter. Plant J 36:
229–239
Arazi T, Sunkar R, Kaplan B, Fromm H (1999) A tobacco
plasma membrane calmodulin-binding transporter confers
Ni21 tolerance and Pb21 hypersensitivity in transgenic
plants. Plant J 20: 171–182
Ashraf M, Khanum A (1997) Relationship between ion
accumulation and growth in two spring wheat lines
differing in salt tolerance at different growth stages. J Agron
Crop Sci 178: 39–51
Ashraf M, McNeilly T, Bradshaw AD (1986) Heritability of
NaCl tolerance in seven grass species. Euphytica 35:
935–940
Ayala F, Oleary JW, Schumaker KS (1996) Increased vacuolar
and plasma membrane H1-ATPase activities in Salicornia
bigelovii Torr in response to NaCl. J Exp Bot 47: 25–32
Azaizeh H, Gunne B, Steudle E (1992) Effects of NaCl and
CaCl2 on water transport across root cells of maize
(Zea mays L.) seedlings. Plant Physiol 99: 886–894
Babourina O, Leonova T, Shabala S, Newman I (2000) Effect
of sudden salt stress on ion fluxes in intact wheat
suspension cells. Ann Bot 85: 759–767
Bajaj S, Targolli J, Liu LF, Ho THD, Wu R (1999) Transgenic
approaches to increase dehydration-stress tolerance in
plants. Mol Breed 5: 493–503
Balague C, Lin BQ, Alcon C, Flottes G, Malmstrom S, Kohler
C, Neuhaus G, Pelletier G, Gaymard F, Roby D (2003)
HLM1, an essential signaling component in the
hypersensitive response, is a member of the cyclic
664 Physiol. Plant. 133, 2007

nucleotide-gated channel ion channel family. Plant Cell
15: 365–379
Banuelos MA, Garciadeblas B, Cubero B, Rodrıguez-Navarro
A (2002) Inventory and functional characterization of the
HAK potassium transporters of rice. Plant Physiol 130:
784–795
Basu R, Ghosh B (1991) Polyamines in various rice (Oryza
sativa) genotypes with respect to sodium-chloride salinity.
Physiol Plant 82: 575–581
Berthomieu P, Conejero G, Nublat A, Brackenbury WJ,
Lambert C, Savio C, Uozumi N, Oiki S, Yamada K, Cellier
F, Gosti F, Simonneau T, Essah PA, Tester M, Very A-A,
Sentenac H, Casse F (2003) Functional analysis of AtHKT1
in Arabidopsis shows that Na1 recirculation by the phloem
is crucial for salt tolerance. EMBO J 22: 2004–2014
Bertl A, Anderson JA, Slayman CL, Sentenac H, Gaber RF
(1994) Inward and outward rectifying potassium currents
in Saccharomyces cerevisiae mediated by endogenous and
heterologously expressed ion channels. Folia Microbiol
(Praha) 39: 507–509
Blumwald E, Aharon GS, Apse MP (2000) Sodium transport
in plant cells. Biochim Biophys Acta 1465: 140–151
Bohnert HJ, Shen B (1999) Transformation and compatible
solutes. Sci Hortic 78: 237–260
Bohnert HJ, Nelson DE, Jensen RG (1995) Adaptation to
environmental stresses. Plant Cell 7: 1099–1111
Bordas M, Montesinos C, Dabauza M, Salvador A, Roig LA,
Serrano R, Moreno V (1997) Transfer of the yeast salt
tolerance gene HAL1 to Cumcumis melo L. cultivats and in
vitro evaluation of salt tolerance. Transgenic Res 6: 41–50
Botella MA, Martinez V, Pardines J, Cerda A (1997) Salinity
induced potassium deficiency in maize plants. J Plant
Physiol 150: 200–205
Bray EA (1997) Plant responses to water deficit. Trends Plant
Sci 2: 48–54
Byrt CS, Platten JD, Spielmeyer W, James RA, Lagudah ES,
Dennis ES, Tester M, Munns R (2007) HKT1;5-like cation
transporters linked to Na1 exclusion loci in wheat, Nax2
and Kna1. Plant Physiol 143: 1918–1928
Cellier F, Conejero G, Ricaud L, Luu DT, Lepetit M, Gosti F,
Casse F (2004) Characterization of AtCHX17, a member of
the cation/H1 exchangers, CHX family, from Arabidopsis
thaliana suggests a role in K1 homeostasis. Plant J 39:
834–846
Chandler SF, Thorpe TA (1987) Characterization of growth,
water relations, and proline accumulation in
sodium-sulfate tolerant callus of Brassica napus L cv
Westar (Canola). Plant Physiol 84: 106–111
Chen THH, Murata N (2002) Enhancement of tolerance of
abiotic stress by metabolic engineering of betaines and
other compatible solutes. Curr Opin Plant Biol 5: 250–257
Chen Z, Newman I, Zhou M, Mendham N, Zhang G, Shabala
S (2005) Screening plants for salt tolerance by measuring
K1 flux: a case study for barley. Plant Cell Environ 28:
1230–1246
Chen ZH, Zhou MX, Newman IA, Mendham NJ, Zhang GP,
Shabala S (2007a) Potassium and sodium relations in
salinised barley tissues as a basis of differential salt
tolerance. Funct Plant Biol 34: 150–162
Chen ZH, Pottosin II, Cuin TA, Fuglsang AT, Tester M, Jha D,
Zepeda-Jazo I, Zhou MX, Palmgren MG, Newman IA,
Shabala S. 2007b).Root plasma membrane transporters
controlling K1/Na1 homeostasis in salt stressed barley.
Plant Physiol Doi:10.1104/pp.107.110262
Cherel I (2004) Regulation of K1 channel activities in plants:
from physiological to molecular aspects. J Exp Bot 55:
337–351
Colmenero-Flores JM, Martinez G, Gamba G, Vazquez N,
Iglesias DJ, Brumos J, Talon M (2007) Identification and
functional characterization of cation-chloride
cotransporters in plants. Plant J 50: 278–292
Colmer TD, Flowers TJ, Munns R (2006) Use of wild relatives
to improve salt tolerance in wheat. J Exp Bot 57:
1059–1078
Cosgrove DJ, Hedrich R (1991) Stretch-activated chloride,
potassium, and calcium channels coexisting in plasma
membranes of guard cells of Vicia faba L. Planta 186:
143–153
Cramer GR, Lynch J, Lauchli A, Epstein E (1987) Influx of
Na1, K1, and Ca21 into roots of salt-stressed cotton
seedlings. Effects of supplemental Ca21. Plant Physiol 83:
510–516
Cramer GR, Epstein E, Lauchli A (1991) Effects of sodium,
potassium and calcium on salt-stressed barley. 2.
Elemental analysis. Physiol Plant 81: 197–202
Cuin TA, Shabala S (2005) Exogenously supplied compatible
solutes rapidly ameliorate NaCl-induced potassium efflux
from barley roots. Plant Cell Physiol 46: 1924–1933
Cuin TA, Shabala S (2006) Potassium homeostasis in salinised
plant tissues. In: Volkov A (ed) Plant Electrophysiology –
Theory and Methods. Springer, Heidelberg, Germany,
pp 287–317
Cuin TA, Shabala S (2007) Amino acids regulate
salinity-induced potassium efflux in barley root epidermis.
Planta 225: 753–761
Cuin TA, Miller AJ, Laurie SA, Leigh RA (2003) Potassium
activities in cell compartments of salt-grown barley leaves.
J Exp Bot 54: 657–661
Czempinski K, Frachisse JM, Maurel C, Barbier-Brygoo H,
Muller-Rober B (2002) Vacuolar membrane localization of
the Arabidopsis ‘two-pore’ K1 channel KCO1. Plant J 29:
809–820
Davenport R (2002) Glutamate receptors in plants. Ann Bot
90: 549–557
Davenport RJ, Munoz-Mayor A, Jha D, Essah PA, Rus A,
Tester M (2007) The Na1 transporter AtHKT1;1 controls
retrieval of Na1 from the xylem in Arabidopsis. Plant Cell
Environ 30: 497–507
Delauney AJ, Verma DPS (1993) Proline biosynthesis and
osmoregulation in plants. Plant J 4: 215–223
Physiol. Plant. 133, 2007 665

Demidchik V, Tester M (2002) Sodium fluxes through
nonselective cation channels in the plasma membrane of
protoplasts from Arabidopsis roots. Plant Physiol 128:
379–387
Demidchik V, Maathuis FJM (2007) Physiological roles of
nonselective cation channels in plants: from salt stress to
signalling and development. New Phytol 175: 387–404
Demidchik V, Davenport RJ, Tester M (2002) Nonselective
cation channels in plants. Annu Rev Plant Biol 53: 67–107
Dennison KL, Robertson WR, Lewis BD, Hirsch RE, Sussman
MR, Spalding EP (2001) Functions of AKT1 and AKT2
potassium channels determined by studies of single
and double mutants of Arabidopsis. Plant Physiol
127: 1012–1019
Ding L, Zhu JK (1997) Reduced Na1 uptake in the
NaCl-hypersensitive sos1 mutant of Arabidopsis thaliana.
Plant Physiol 113: 795–799
Dubcovsky J, Maria GS, Epstein E, Luo MC, Dvorak J (1996)
Mapping of the K1/Na1 discrimination locus Kna1 in
wheat. Theor Appl Genet 92: 448–454
Dvorak J, Noaman MM, Goyal S, Gorham J (1994)
Enhancement of the salt tolerance of Triticum turgidum L.
by the kna1 locus transferred from the Triticum aestivum L.
chromosome 4D by homoeologous recombination. Theor
Appl Genet 87: 872–877
Erdei L, Trivedi S, Takeda K, Matsumoto H (1990) Effects of
osmotic and salt stresses on the accumulation of
polyamines in leaf segments from wheat-varieties differing
in salt and drought tolerance. J Plant Physiol 137: 165–168
Espinosa-Ruiz A, Belles JM, Serrano R, Culianez-Macia FA
(1999) Arabidopsis thaliana AAtHAL3: a flavoprotein
related to salt and osmotic tolerance and plant growth.
Plant J 20: 529–539
Flowers TJ (2004) Improving crop salt tolerance. J Exp Bot 55:
307–319
Flowers TJ, Yeo AR (1995) Breeding for salinity resistance in
crop plants – where next? Aust J Plant Physiol 22: 875–884
Flowers TJ, Hajibagheri MA, Yeo AR (1991) Ion
accumulation in the cell walls of rice plants growing under
saline conditions: evidence for the Oertli hypothesis. Plant
Cell Environ 14: 319–325
Fooland MR (1997) Genetic basis of physiological traits
related to salt tolerance in tomato, Lycopersicon
esculentum Mill. Plant Breed 116: 53–58
Fu HH, Luan S (1998) AtKUP1: a dual-affinity K1 transporter
from Arabidopsis. Plant Cell 10: 63–73
Fukuda A, Nakamura A, Tagiri A, Tanaka H, Miyao A,
Hirochika H, Tanaka Y (2004) Function, intracellular
localization and the importance in salt tolerance of
a vacuolar Na1/H1 antiporter from rice. Plant Cell Physiol
45: 146–159
Garcia A, Rizzo CA, UdDin J, Bartos SL, Senadhira D,
Flowers TJ, Yeo AR (1997) Sodium and potassium
transport to the xylem are inherited independently in rice,
and the mechanism of sodium: potassium selectivity
differs between rice and wheat. Plant Cell Environ 20:
1167–1174
Garciadeblas B, Senn ME, Banuelos MA, Rodrıguez-Navarro
A (2003) Sodium transport and HKT transporters: the rice
model. Plant J 34: 788–801
Garthwaite AJ, von Bothmer R, Colmer TD (2005) Salt
tolerance in wild Hordeum species is associated with
restricted entry of Na1 and Cl2 into the shoots. J Exp Bot
56: 2365–2378
Gaxiola R, Delarrinoa IF, Villalba JM, Serrano R (1992) A
novel and conserved salt-induced protein is an important
determinant of salt tolerance in yeast. EMBO J 11:
3157–3164
Gaxiola RA, Li JS, Undurranga S, Dang LM, Allen GJ, Alper
SL, Fink GR (2001) Drought- and salt-tolerant plants results
from overexpression of the AVP1 H1-pump. Proc Natl
Acad Sci USA 98: 11444–11449
Gaxiola RA, Palmgren MG, Schumacher K (2007) Plant
proton pumps. FEBS Lett 571: 2204–2214
Gaymard F, Pilot G, Lacombe B, Bouchez D, Bruneau D,
Boucherez J, Michaux-Ferriere N, Thibaud JB, Sentenac H
(1998) Identification and disruption of a plant shaker-like
outward channel involved in K1 release into the xylem
sap. Cell 94: 647–655
Genc Y, McDonald GK, Tester M (2007) Reassessment of
tissue Na1 concentration as a criterion for salinity
tolerance in bread wheat. Plant Cell Environ. doi:10.1111/
j.1365–3040.2007.01726.x
Gierth M, Maser P (2007) Potassium transporters in plant –
involvement in K1 acquisition. Redistribution and
homeostasis. FEBS Lett 581: 2348–2356
Golldack D, Dietz KJ (2001) Salt-induced expression of the
vacuolar H1-ATPase in the common ice plant is
developmentally controlled and tissue specific. Plant
Physiol 125: 1643–1654
Golldack D, Kamasani UR, Quigley F, Bennett J, Bohnert HJ
(1997) Salt stress-dependent expression of a HKT1-type
high affinity potassium transporter in rice. Plant Physiol
114: 529–529
Gorham J, Bristol A, Young EM, Jones RGW (1991) The
presence of the enhanced K/Na discrimination trait in
diploid triticum species. Theor Appl Genet 82: 729–736
Gregorio GB, Senadhira D (1993) Genetic analysis of salinity
tolerance in rice (Oryza sativa L.). Theor Appl Genet 86:
333–338
Hall D, Evans AR, Newbury HJ, Pritchard J (2006) Functional
analysis of CHX21: a putative sodium transporter in
Arabidopsis. J Exp Bot 57: 1201–1210
Hare PD, Cress WA, Van Staden J (1998) Dissecting the roles
of osmolyte accumulation during stress. Plant Cell Environ
21: 535–553
Harinasut P, Tsutsui K, Takabe T, Nomura M, Takabe T,
Kishitani S (1996) Exogenous glycinebetaine accumulation
and increased salt-tolerance in rice seedlings. Biosci
Biotechnol Biochem 60: 366–368
666 Physiol. Plant. 133, 2007

Hasegawa PM, Bressan RA, Zhu JK, Bohnert HJ (2000) Plant
cellular and molecular responses to high salinity. Annu
Rev Plant Physiol Plant Mol Biol 51: 463–499
Hassan NS, Wilkins DA (1988) In vitro selection for salt
tolerant lines in Lycopersicon peruvianum. Plant Cell Rep
7: 463–466
Haw M, Cocklin C, Mercer D (2000) A pinch of salt:
landowner perception and adjustment to the salinity
hazard in Victoria, Australia. J Rural Stud 16: 155–169
Hirsch RE, Lewis BD, Spalding EP, Sussman MR (1998) A role
for the AKT1 potassium channel in plant nutrition. Science
280: 918–921
Horie T, Yoshida K, Nakayama H, Yamada K, Oiki S, Shinmyo
A (2001) Two types of HKT transporters with different
properties of Na1 and K1 transport in Oryza sativa. Plant J
27: 129–138
Ivashikina N, Becker D, Ache P, Meyerhoff O, Felle HH,
Hedrich R (2001) K1 channel profile and electrical
properties of Arabidopsis root hairs. FEBS Lett 508:
463–469
Jacobsen SE, Mujica A, Jensen CR (2003) The resistance of
quinoa (Chenopodium quinoa Willd.) to adverse abiotic
factors. Food Rev Int 19: 99–109
Kefu Z, Hai F, San Z, Jie S (2003) Study on the salt and
drought tolerance of Suaeda salsa and Kalanchoe
claigremontiana under iso-osmotic salt and water stress.
Plant Sci 165: 837–844
Khan MA, Ungar IA, Showalter AM (2005) Salt stimulation
and tolerance in an intertidal stem-succulent halophyte.
J Plant Nutr 28: 1365–1374
Kim SA, Kwak JM, Jae SK, Wang MH, Nam HG (2001)
Overexpression of the AtGluR2 gene encoding an
Arabidopsis homolog of mammalian glutamate
receptors impairs calcium utilization and sensitivity to
ionic stress in transgenic plants. Plant Cell Physiol 42:
74–84
Koyama ML, Levesley A, Koebner RMD, Flowers TJ, Yeo AR
(2001) Quantitative trait loci for component physiological
traits determining salt tolerance in rice. Plant Physiol 125:
406–422
Lacombe B, Pilot G, Michard E, Gaymard F, Sentenac H,
Thibaud JB (2000) A shaker-like K1 channel with weak
rectification is expressed in both source and sink phloem
tissues of Arabidopsis. Plant Cell 12: 837–851
Lacombe B, Becker D, Hedrich R, DeSalle R, Hollmann M,
Kwak JM, Schroeder JI, Le Novere N, Nam HG, Spalding
EP, Tester M, Turano FJ, Chiu J, Coruzzi G (2001) The
identity of plant glutamate receptors. Science 292:
1486–1487
Laurie S, Feeney KA, Maathuis FJM, Heard PJ, Brown SJ,
Leigh RA (2002) A role for HKT1 in sodium uptake by
wheat roots. Plant J 32: 139–149
Lebaudy A, Very A-A, Sentenac H (2007) K1 channel activity
in plants: genes, regulations and functions. FEBS Lett 581:
2357–2366
Leng Q, Mercier RW, Hua BG, Fromm H, Berkowitz GA
(2002) Electrophysiological analysis of cloned cyclic
nucleotide-gated ion channels. Plant Physiol 128:
400–410
Lindsay MP, Lagudah ES, Hare RA, Munns R (2004) A locus
for sodium exclusion (Nax1), a trait for salt tolerance,
mapped in durum wheat. Funct Plant Biol 31: 1105–1114
Ludlow MM, Muchow RC (1990) A critical-evaluation of
traits for improving crop yields in water-limited
environments. Adv Agron 43: 107–153
Ma SS, Gong QQ, Bohnert HJ (2006) Dissecting salt stress
pathways. J Exp Bot 57: 1097–1107
Maathuis FJM (2006) The role of monovalent cation
transporters in plant responses to salinity. J Exp Bot 57:
1137–1147
Maathuis FJM, Amtmann A (1999) K1 nutrition and Na1
toxicity: the basis of cellular K1/Na1 ratios. Ann Bot 84:
123–133
Maathuis FJM, Sanders D (2001) Sodium uptake in
Arabidopsis roots is regulated by cyclic nucleotides. Plant
Physiol 127: 1617–1625
Mano Y, Takeda K (1997) Diallel analysis of salt tolerance at
germination and the seedling stage in barley (Hordeum
vulgare L.). Breed Sci 47: 203–209
Marschner H (1995) The Mineral Nutrition of Higher Plants.
Academic Press, London
Marten I, Hoth S, Deeken R, Ache P, Ketchum KA,
Hoshi T, Hedrich R (1999) AKT3, a phloem-localized K1
channel, is blocked by protons. Proc Natl Acad Sci
USA 96: 7581–7586
Maser P, Thomine S, Schroeder JI, Ward JM, Hirschi K,
Sze H, Talke IN, Amtmann A, Maathuis FJM, Sanders D,
Harper JF, Tchieu J, Gribskov M, Persans MW, Salt DE,
Kim SA, Guerinot ML (2001) Phylogenetic relationships
within cation transporter families of Arabidopsis. Plant
Physiol 126: 1646–1667
Morgan JM (1983) Osmoregulation as a selection criterion
for drought tolerance in wheat. Aust J Agric Res 34:
607–614
Munns R (2002) Comparative physiology of salt and water
stress. Plant Cell Environ 25: 239–250
Munns R (2005) Genes and salt tolerance: bringing them
together. New Phytol 167: 645–663
Munns R, Hare RA, James RA, Rebetzke GJ (1999) Genetic
variation for improving the salt tolerance of durum wheat.
Austr J Agric Res 51: 69–74
Munns R, James RA, Lauchli A (2006) Approaches to
increasing the salt tolerance of wheat and other cereals.
J Exp Bot 57: 1025–1043
Ndayiragije A, Lutts S (2006) Exogenous putrescine reduces
sodium and chloride accumulation in NaCl-treated calli of
the salt-sensitive rice cultivar I Kong Pao. Plant Growth
Regul 48: 51–63
Palmgren MG (1991) Regulation of plasma membrane
H1-ATPase activity. Physiol Plant 83: 314–323
Physiol. Plant. 133, 2007 667

Papageorgiou GC, Murata N (1995) The unusually strong
stabilizing effects of glycine betaine on the structure and
function of the oxygen-evolving photosystem II complex.
Photosynth Res 44: 243–252
Pilot G, Gaymard F, Mouline K, Cherel I, Sentenac H (2003)
Regulated expression of Arabidopsis Shaker K1 channel
genes involved in K1 uptake and distribution in the plant.
Plant Mol Biol 51: 773–787
Qi Z, Spalding EP (2004) Protection of plasma membrane K1
transport by the salt overly sensitive1 Na1/H1 antiporter
during salinity stress. Plant Physiol 136: 2548–2555
Raven JA (1985) Regulation of pH and generation of
osmolarity in vascular plants: a cost-benefit analysis in
relation to efficiency of use of energy, nitrogen and water.
New Phytol 101: 25–77
Reid RJ, Smith FA (2000) The limits of sodium/calcium
interactions in plant growth. Aust J Plant Physiol 27:
709–715
Ren ZH, Gao JP, Li LG, Cai XL, Huang W, Chao DY, Zhu MZ,
Wang ZY, Luan S, Lin HX (2005) A rice quantitative trait
locus for salt tolerance encodes a sodium transporter.
Nat Genet 37: 1141–1146
Rengasamy P (2006) World salinization with emphasis on
Australia. J Exp Bot 57: 1017–1023
Rubio F, Gassmann W, Schroeder JI (1995) Sodium-driven
potassium uptake by the plant potassium transporter HKT1
and mutations conferring salt tolerance. Science 270:
1660–1663
Rubio F, Santa-Marıa GE, Rodrıguez-Navarro A (2000)
Cloning of Arabidopsis and barley cDNAs encoding HAK
potassium transporters in root and shoot cells. Physiol
Plant 109: 34–43
Rus A, Yokoi S, Sharkhuu A, Reddy M, Lee BH,
Matsumomoto TK, Koiwa H, Zhu JK, Bressan RA,
Hasegawa PM (2001) AtHKT1 is a salt tolerance
determinant that controls Na1 entry into plant roots.
Proc Natl Acad Sci USA 98: 14150–14155
Rus A, Lee BH, Munoz-Mayor A, Sharkhuu A, Miura K, Zhu
JK, Bressan RA, Hasegawa PM (2004) AtHKT1 facilitates
Na1 homeostasis and K1 nutrition in planta. Plant Physiol
136: 2500–2511
Santa-Cruz A, Acosta M, Rus A, Bolarin MC (1999)
Short-term salt tolerance mechanisms in differentially salt
tolerant tomato species. Plant Physiol Biochem 37: 65–71
Santa-Marıa GE, Rubio F, Dubcovsky J, Rodriguez-Navarro A
(1997) The HAK1 gene of barley is a member of a large
gene family and encodes a high-affinity potassium
transporter. Plant Cell 9: 2281–2289
Saqib M, Zorb C, Rengel Z, Schubert S (2005) The expression
of the endogenous vacuolar Na1/H1 antiporters in wheat
(Triticum aestivum L.) Plant Sci 169: 959–965
Schachtman DP, Schroeder JI (1994) Structure and transport
mechanism of a high-affinity potassium uptake transporter
from higher plants. Nature 370: 655–658
Schonknecht G, Spoormaker P, Steinmeyer R, Bruggeman L,
Ache P, Dutta R, Reintanz B, Godde M, Hedrich R, Palme
K (2002) KCO1 is a component of the slow-vacuolar (SV)
ion channel. FEBS Lett 511: 28–32
Serraj R, Sinclair TR (2002) Osmolyte accumulation: can it
really help increase crop yield under drought conditions?
Plant Cell Environ 25: 333–341
Serrano R (1996) Salt tolerance in plants and
microorganisms: toxicity targets and defense responses. Int
Rev Cytol 165: 1–52
Serrano R, Rodrıguez-Navarro A (2001) Ion homeostasis
during salt stress in plants. Curr Opin Cell Biol 13:
399–404
Shabala S (2006) Non-invasive microelectrode ion flux
measurements in plant stress physiology. In: Volkov A (ed)
Plant Electrophysiology – Theory and Methods.
Springer-Verlag, Berlin, pp 35–71
Shabala S, Lew RR (2002) Turgor regulation in osmotically
stressed Arabidopsis epidermal root cells. Direct support
for the role of inorganic ion uptake as revealed by
concurrent flux and cell turgor measurements. Plant
Physiol 129: 290–299
Shabala S, Babourina O, Newman I (2000) Ion-specific
mechanisms of osmoregulation in bean mesophyll cells.
J Exp Bot 51: 1243–1253
Shabala S, Shabala L, Van Volkenburgh E (2003) Effect of
calcium on root development and root ion fluxes in
salinised barley seedlings. Funct Plant Biol 30: 507–514
Shabala L, Cuin TA, Newman IA, Shabala S (2005a)
Salinity-induced ion flux patterns from the excised roots of
Arabidopsis sos mutants. Planta 222: 1041–1050
Shabala S, Shabala L, Van Volkenburgh E, Newman I (2005b)
Effect of divalent cations on ion fluxes and leaf
photochemistry in salinised barley leaves. J Exp Bot 56:
1369–1378
Shabala S, Demidchik V, Shabala L, Cuin TA, Smith SJ, Miller
AJ, Davies JM, Newman IA (2006) Extracellular Ca21
ameliorates NaCl-induced K1 loss from Arabidopsis root
and leaf cells by controlling plasma membrane
K1-permeable channels. Plant Physiol 141: 1653–1665
Shabala S, Cuin TA, Pottosin I (2007) Polyamines prevent
NaCl-induced K1 efflux from pea mesophyll by blocking
non-selective cation channels. FEBS Lett 581: 1993–1999
Shi HZ, Ishitani M, Kim CS, Zhu JK (2000) The Arabidopsis
thaliana salt tolerance gene SOS1 encodes a putative Na1/
H1 antiporter. Proc Natl Acad Sci USA 97: 6896–6901
Shi HZ, Lee BH, Wu SJ, Zhu JK (2003) Overexpression of
a plasma membrane Na1/H1 antiporter gene improves salt
tolerance in Arabidopsis thaliana. Nat Biotechnol 21:
81–85
Su H, Golldack D, Zhao CS, Bohnert HJ (2002) The
expression of HAK-type K1 transporters is regulated in
response to salinity stress in common ice plant. Plant
Physiol 129: 1482–1493
668 Physiol. Plant. 133, 2007

Sunarpi, Horie T, Motoda J, Kubo M, Yang H, Yoda K, Horie
R, Chan WY, Leung HY, Hattori K, Konomi M, Osumi M,
Yamagami M, Schroeder JI, Uozumi N (2005) Enhanced
salt tolerance mediated by AtHKT1 transporter-induced
Na1 unloading from xylem vessels to xylem parenchyma
cells. Plant J 44: 928–938
Szyroki A, Ivashikina N, Dietrich P, Roelfsema MRG, Ache P,
Reintanz B, Deeken R, Godde M, Felle H, Steinmeyer R,
Palme K, Hedrich R (2001) KAT1 is not essential for stomatal
opening. Proc Natl Acad Sci USA 98: 2917–2921
Talke IN, Blaudez D, Maathuis FJM, Sanders D (2003)
CNGCs: prime targets of plant cyclic nucleotide
signalling? Trends Plant Sci 8: 286–293
Teodoro AE, Zingarelli L, Lado P (1998) Early changes of Cl2
efflux and H1 extrusion induced by osmotic stress in
Arabidopsis thaliana cells. Physiol Plant 102: 29–37
Tester M, Davenport R (2003) Na1 tolerance and Na1
transport in higher plants. Ann Bot 91: 503–527
Tyerman SD, Skerrett M, Garrill A, Findlay GP, Leigh RA
(1997) Pathways for the permeation of Na1 and Cl2 into
protoplasts derived from the cortex of wheat roots. J Exp
Bot 48: 459–480
Uozumi N, Kim EJ, Rubio F, Yamaguchi T, Muto S, Tsuboi A,
Bakker EP, Nakamura T, Schroeder JI (2000) The
Arabidopsis HKT1 gene homolog mediates inward Na1
currents in Xenopus laevis oocytes and Na1 uptake in
Saccharomyces cerevisiae. Plant Physiol 122: 1249–1259
Vallejo AJ, Peralta ML, Santa-Marıa GE (2005) Expression of
potassium-transporter coding genes, and kinetics of
rubidium uptake, along a longitudinal root axis. Plant Cell
Environ 28: 850–862
Very A-A, Sentenac H (2002) Cation channels in the
Arabidopsis plasma membrane. Trends Plant Sci 7:
168–175
Very A-A, Sentenac H (2003) Molecular mechanisms and
regulation of K1 transport in higher plants. Annu Rev Plant
Biol 54: 575–603
Volkov V, Wang B, Dominy PJ, Fricke W, Amtmann A (2003)
Thellungiella halophila, a salt-tolerant relative of
Arabidopsis thaliana, possesses effective mechanisms to
discriminate between potassium and sodium. Plant Cell
Environ 27: 1–14
Walker DJ, Leigh RA, Miller AJ (1996) Potassium homeostasis
in vacuolate plant cells. Proc Natl Acad Sci USA 93:
10510–10514
Wu YY, Chen GD, Meng QW, Cheng CC (2004) The cotton
GhNHX1 gene encoding a novel putative tonoplast Na1/
H1 antiporter plays an important role in salt stress. Plant
Cell Physiol 45: 600–607
Xue ZY, Zhi DY, Xue GL, Zhang H, Zhao YX, Xia GM (2004)
Enhanced salt tolerance of transgenic wheat (Triticum
sativum L.) expressing a vacuolar Na1/H1 antiporter gene
with improved grain yields in saline soils in the field and
a reduced level of leaf Na1. Plant Sci 167: 849–859
Yeo AR, Lee KS, Izard P, Boursier PJ, Flowers TJ (1991)
Short-term and long-term effects of salinity on leaf growth
in rice (Oryza sativa L). J Exp Bot 42: 881–889
Zhang HX, Blumwald E (2001) Transgenic salt-tolerant
tomato plants accumulate salt in foliage but not in fruit.
Nat Biotechnol 19: 765–768
Zhang JX, Nguyen HT, Blum A (1999) Genetic analysis of
osmotic adjustment in crop plants. J Exp Bot 50: 291–302
Zhang HX, Hodson JN, Williams JP, Blumwald E (2001)
Engineering salt tolerant Brassica plants: characterization
of yield and seed oil quality in transgenic plants with
increased vacuolar sodium accumulation. Proc Natl Acad
Sci USA 98: 12832–12836
Zhu JK (2003) Regulation of ion homeostasis under salt stress.
Curr Opin Plant Biol 6: 441–445
Zingarelli L, Marre MT, Massardi F, Lado P (1999) Effects of
hyper-osmotic stress on K1 fluxes, H1 extrusion,
transmembrane electric potential difference and
comparison with the effects of fusicoccin. Physiol Plant
106: 287–295
Edited by J. K. Schjørring
Physiol. Plant. 133, 2007 669
Related Documents











