185 J Vet Diagn Invest 13:185–194 (2001) Postweaning multisystemic wasting syndrome induced after experimental inoculation of cesarean-derived, colostrum- deprived piglets with type 2 porcine circovirus Steven R. Bolin, William C. Stoffregen, Gopi P. S. Nayar, Andre L. Hamel Abstract. Cesarean-derived, colostrum-deprived pigs (n 5 23) were inoculated intranasally and subcuta- neously with a low cell culture passage of type 2 porcine circovirus. In 11 pigs, a persistent fever that lasted 7–17 days began 12–15 days after inoculation with virus. Additional signs of disease in those 11 pigs included depression (11 of 11 pigs), palpable enlargement of inguinal, prefemoral, and popliteal lymph nodes (11 of 11), icterus (6 of 11), and hyperpnea (2 of 11). The remaining 12 pigs had fever that occurred intermittently for 2–4 days between days 12 and 20 postinoculation. Overt signs of disease in those pigs were limited to palpable enlargement of inguinal and popliteal lymph nodes (9 of 12 pigs). When compared with control pigs of similar age, the average daily rate of weight gain for all pigs inoculated with virus was less over a 2-week period that began 2 weeks post inoculation. At postmortem examination, lymph node enlargement was seen in 14 of 14 pigs euthanized between days 20 and 28 postinoculation. Lymph node enlargement was especially prominent in pigs that developed a persistent fever. Microscopic lesions noted in pigs that developed a persistent fever included cellular depletion in lymphoid tissues; hepatic cell necrosis; and lymphogranulomatous inflam- mation of lymph nodes, Peyer’s patches of the intestine, liver, kidney, and heart. Virus was isolated with varying frequency from nasal, rectal, or tonsil swab specimens, buffy coat, serum, urine, and lung lavage fluid obtained antemortem or postmortem. Virus was isolated from or viral DNA was detected in a variety of tissues obtained postmortem up to 125 days postinoculation. Antibody against type 2 porcine circovirus usually was detected in serum between 15 and 20 days postinoculation; however, antibody against virus was not detected in serum from 4 pigs euthanized 20–24 days postinoculation. Direct contact with pigs inoculated with virus 42 days previously resulted in transmission of virus to 3 of 3 control pigs. Postweaning multisystemic wasting syndrome (PMWS) is an emerging disease of swine first identi- fied in 1991 in high-health swine herds in western Canada. 1,13 Manifestations of PMWS include weight loss, respiratory distress, rough hair coat, pallor, diar- rhea, jaundice, and death. 17,18 Lymphadenopathy, gas- tric ulceration, hepatopathy, nephritis, and pneumonia are gross lesions associated with PMWS. 13,17,21 Micro- scopic lesions include cell depletion in lymphoid fol- licles, granulomatous inflammation of lymphoid tis- sues, homogenous, spherical, and basophilic cytoplas- mic inclusion bodies of various sizes in histiocytic cells in lymphoid follicles, bronchointerstitial pneu- monia, and lymphogranulomatous inflammation of the liver, kidney, and heart. 4,13,21,28,33 Small virus-like par- ticles have been observed in and isolated from tissues from swine that had PMWS. 4,13,21 Those virus-like par- ticles lack a lipid envelope and have an icosahedral From the Virus and Prion Diseases of Livestock Research Unit, National Animal Disease Center, Agricultural Research Service, USDA, PO Box 70, Ames, IA 50010 (Bolin, Stoffregen), and the Virology Laboratory, Veterinary Services, Manitoba Agriculture, 545 University Crescent, Winnipeg, Manitoba R3T 5S6, Canada (Nayar, Hamel). Received for publication June 12, 2000. capsid about 17 nm in diameter. Morphologically, the virus-like particles associated with PMWS resemble porcine circovirus (PCV), a known contaminant of the PK-15 cell line. 36,37,40 Analysis of the DNA nucleic acid sequence from the virus-like particles revealed high similarity with the DNA of the PCV from PK-15 cells. 15,26–28 Although related genetically, the PCV as- sociated with PMWS and the PCV in PK-15 cells dif- fer sufficiently in nucleic acid sequence to allow clas- sification as separate viral genotypes. The PCV in PK- 15 cells appears representative of genotype 1 PCV (PVC1), and the recently identified PCV associated with PMWS appears representative of genotype 2 PCV (PCV2). 27 This classification has support from mono- clonal and polyclonal antibody binding assays, which show that PCV1 and PCV2 are antigenically similar, but the viral genotypes can be segregated by serologic tests. 4,13,27 In previous studies, experimental inoculation of swine 1 day to 9 months of age with PCV1 did not induce signs of disease, and no overt gross or micro- scopic lesions were detected. 3,38 Viral antigen was de- tected in numerous tissues, most often in lung and lymphoid tissue from pigs euthanized 3–11 days post- inoculation (PI). 3 Virus was isolated from nasal swab by guest on February 5, 2015 vdi.sagepub.com Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

185
J Vet Diagn Invest 13:185–194 (2001)
Postweaning multisystemic wasting syndrome induced afterexperimental inoculation of cesarean-derived, colostrum-
deprived piglets with type 2 porcine circovirus
Steven R. Bolin, William C. Stoffregen, Gopi P. S. Nayar, Andre L. Hamel
Abstract. Cesarean-derived, colostrum-deprived pigs (n 5 23) were inoculated intranasally and subcuta-neously with a low cell culture passage of type 2 porcine circovirus. In 11 pigs, a persistent fever that lasted7–17 days began 12–15 days after inoculation with virus. Additional signs of disease in those 11 pigs includeddepression (11 of 11 pigs), palpable enlargement of inguinal, prefemoral, and popliteal lymph nodes (11 of11), icterus (6 of 11), and hyperpnea (2 of 11). The remaining 12 pigs had fever that occurred intermittentlyfor 2–4 days between days 12 and 20 postinoculation. Overt signs of disease in those pigs were limited topalpable enlargement of inguinal and popliteal lymph nodes (9 of 12 pigs). When compared with control pigsof similar age, the average daily rate of weight gain for all pigs inoculated with virus was less over a 2-weekperiod that began 2 weeks post inoculation. At postmortem examination, lymph node enlargement was seen in14 of 14 pigs euthanized between days 20 and 28 postinoculation. Lymph node enlargement was especiallyprominent in pigs that developed a persistent fever. Microscopic lesions noted in pigs that developed a persistentfever included cellular depletion in lymphoid tissues; hepatic cell necrosis; and lymphogranulomatous inflam-mation of lymph nodes, Peyer’s patches of the intestine, liver, kidney, and heart. Virus was isolated with varyingfrequency from nasal, rectal, or tonsil swab specimens, buffy coat, serum, urine, and lung lavage fluid obtainedantemortem or postmortem. Virus was isolated from or viral DNA was detected in a variety of tissues obtainedpostmortem up to 125 days postinoculation. Antibody against type 2 porcine circovirus usually was detectedin serum between 15 and 20 days postinoculation; however, antibody against virus was not detected in serumfrom 4 pigs euthanized 20–24 days postinoculation. Direct contact with pigs inoculated with virus 42 dayspreviously resulted in transmission of virus to 3 of 3 control pigs.
Postweaning multisystemic wasting syndrome(PMWS) is an emerging disease of swine first identi-fied in 1991 in high-health swine herds in westernCanada.1,13 Manifestations of PMWS include weightloss, respiratory distress, rough hair coat, pallor, diar-rhea, jaundice, and death.17,18 Lymphadenopathy, gas-tric ulceration, hepatopathy, nephritis, and pneumoniaare gross lesions associated with PMWS.13,17,21 Micro-scopic lesions include cell depletion in lymphoid fol-licles, granulomatous inflammation of lymphoid tis-sues, homogenous, spherical, and basophilic cytoplas-mic inclusion bodies of various sizes in histiocyticcells in lymphoid follicles, bronchointerstitial pneu-monia, and lymphogranulomatous inflammation of theliver, kidney, and heart.4,13,21,28,33 Small virus-like par-ticles have been observed in and isolated from tissuesfrom swine that had PMWS.4,13,21 Those virus-like par-ticles lack a lipid envelope and have an icosahedral
From the Virus and Prion Diseases of Livestock Research Unit,National Animal Disease Center, Agricultural Research Service,USDA, PO Box 70, Ames, IA 50010 (Bolin, Stoffregen), and theVirology Laboratory, Veterinary Services, Manitoba Agriculture,545 University Crescent, Winnipeg, Manitoba R3T 5S6, Canada(Nayar, Hamel).
Received for publication June 12, 2000.
capsid about 17 nm in diameter. Morphologically, thevirus-like particles associated with PMWS resembleporcine circovirus (PCV), a known contaminant of thePK-15 cell line.36,37,40 Analysis of the DNA nucleicacid sequence from the virus-like particles revealedhigh similarity with the DNA of the PCV from PK-15cells.15,26–28 Although related genetically, the PCV as-sociated with PMWS and the PCV in PK-15 cells dif-fer sufficiently in nucleic acid sequence to allow clas-sification as separate viral genotypes. The PCV in PK-15 cells appears representative of genotype 1 PCV(PVC1), and the recently identified PCV associatedwith PMWS appears representative of genotype 2 PCV(PCV2).27 This classification has support from mono-clonal and polyclonal antibody binding assays, whichshow that PCV1 and PCV2 are antigenically similar,but the viral genotypes can be segregated by serologictests.4,13,27
In previous studies, experimental inoculation ofswine 1 day to 9 months of age with PCV1 did notinduce signs of disease, and no overt gross or micro-scopic lesions were detected.3,38 Viral antigen was de-tected in numerous tissues, most often in lung andlymphoid tissue from pigs euthanized 3–11 days post-inoculation (PI).3 Virus was isolated from nasal swab
by guest on February 5, 2015vdi.sagepub.comDownloaded from

186 Bolin et al.
specimens obtained during the first week PI and fromfecal samples obtained on days 13 and 14 PI.38 Gno-tobiotic pigs 1 or 3 days of age, colostrum-deprivedpigs 1–2 days of age, and conventional pigs 8 weeksof age have been inoculated with PCV2.2,6,14,20,22 Mildto moderate disease was manifested by depression,low-grade fever, and reduced rate of weight gain,2,6 andsevere disease noted in 1 pig resulted in loss of bodyweight.2,20 The primary macroscopic lesion waslymphadenopathy, but edema of the colonic mesenteryand interstitial pulmonary edema was seen in 1pig.2,14,20 Microscopic lesions primarily were mild tomoderate; histiocytic infiltration in lymph nodes wasthe most prominent lesion.2,6,20,22 Simultaneous inocu-lation of gnotobiotic or colostrum-deprived pigs withPCV2 and porcine parvovirus (PPV) has induced se-vere clinical disease and microscopic lesions consis-tent with those described in pigs from outbreaks ofPMWS.2,14,20,22
In the current study, cesarean-derived, colostrum-deprived pigs 20 or 25 days of age were inoculatedwith the fourth cell culture passage of PCV2 strain 688(PCV2/688). One purpose was to reproduce clinicalsigns of disease and the gross and microscopic lesionsassociated with PMWS. A second purpose was to de-termine clinical specimens and tissues from which vi-rus could be isolated or in which viral DNA could bedetected and to acquire preliminary information of du-ration of viral persistence in pigs.
Materials and methods
Animals. Mixed-breed cesarean-derived, colostrum-de-prived pigs were obtained from date-mated sows. The pigswere housed individually in environmentally controlled iso-lation facilities and fed commercially available milk replaceruntil 10 days of age. On day 10 after birth, the pigs wereallotted into 4 groups containing 15 (group A), 7 (group B),8 (group C), or 5 (group D) pigs. Each group included pigsfrom 2 litters, and grouping was done by personnel that didnot participate in the experiment. The groups of pigs wereplaced in separate biocontainment rooms on elevated deckswith supplemental heat provided by heating pads and heatlamps. From days 10 to 17 after birth, the pigs were weanedgradually to a commercially available starter ration contain-ing 21% protein. Rectal body temperature was recorded dai-ly for each pig. Antibiotics were administered to pigs thathad a body temperature of .40 C, hyperpnea, or diarrhea.All groups of pigs were treated similarly.
Cells and virus. A PCV-free clone of the PK-15 cell linewas used for propagation, titration, and isolation of PCV2.The MARC-145 cell line was used for propagation and iso-lation of porcine reproductive and respiratory syndrome vi-rus (PRRSV). The PT-1 clonea of the ST cell line was usedfor propagation and isolation of PPV. The PK-15 cell line(CCL 33) that is contaminated with PCV1 was obtainedfrom the American Type Culture Collection and used forpropagation of PCV1. Bovine turbinate cells were used for
propagation and isolation of bovine viral diarrhea virus(BVDV). All cell cultures were grown in McCoy 5A me-dium containing 0.5 mg/ml kanamycin sulfate and supple-mented to 6% with fetal bovine serum that tested free ofBVDV and of antibody against BVDV.7,8 The PCV2 usedhere was isolated from a field outbreak of PMWS that oc-curred in Manitoba, Canada. That original viral isolate wascontaminated with PPV. To obtain PCV2 free of PPV, theoriginal viral isolate was passaged by inoculation into pigs.At 39 days PI, PCV2 was isolated from serum obtained froman inoculated pig that had detectable concentrations of an-tibody in serum against PPV for the previous 14 days. Thevirus was designated PCV2/688, and the fourth cell culturepassage was used in the current study. The NADC-9 strainof PRRSVb and a field isolate of PPVb were used to testserum for antibody against those viruses. The Singer strainof type 1 BVDV and the 125C strain of type 2 BVDV wereused to test for antibody against BVDV.
Viral inoculation, clinical specimens, and body weight. At20 days of age, the 15 pigs in group A were inoculated with3 ml of medium containing approximately 10,000 cell cul-ture infective doses of PCV2/688. One milliliter of inoculumwas injected into each nostril, and the remaining 1 ml ofinoculum was injected into the subcutaneous tissue behindthe right ear. At 25 days of age, the 8 pigs in group C weresimilarly inoculated with PCV2/688. At 20 or 25 days ofage, the 7 pigs in group B and the 5 pigs in group D wereinoculated with 3 ml of medium harvested from PCV-freePK-15 cells. Before inoculation, each pig was weighed anda sample of blood was obtained for hematology, viral iso-lation, and serology. Rectal, tonsil, and nasal swab speci-mens also were obtained for viral isolation. The swabs wereplaced in tubes containing 1 ml of medium for transport tothe laboratory. At 2–6-day intervals through day 28 PI, sam-ples of blood and rectal, tonsil, and nasal swab specimenswere obtained from several pigs in each group. After day 28PI, viral isolation was attempted periodically from serum orbuffy coat cells. Body weight was recorded each time clin-ical specimens were obtained through day 28 PI. At post-mortem, urine was aspirated from the bladder for viral iso-lation. Also, lung lavage was done to obtain alveolar mac-rophages for viral isolation.
Contact exposure. At 42 days PI, 3 pigs from group Cinoculated with PCV2/688 at 25 days of age were moved toa clean biocontainment room and placed in contact with 3control pigs of like age from group D that had been shaminoculated with medium from PCV-free PK-15 cells. Thepigs were observed daily for signs of disease. A sample ofserum was obtained at weekly intervals from the control pigsto monitor for antibody production against PCV2. Buffy coatcells and serum from those pigs were used for viral isolation;however, other clinical specimens were not obtained andbody weight was not recorded.
Viral isolation. Viral isolation was done by inoculation ofPCV-free PK-15 cells grown in 24-well tissue culture plateswith 0.1 ml of serum, urine, or filtered lung lavage fluids. Alike volume of filtered rectal, tonsil, or nasal swab specimenswas used for viral isolation. Cells in lung lavage fluid werepelleted by centrifugation for 15 min at 1,000 3 g. Buffycoat cells were aspirated off packed blood cells in tubes
by guest on February 5, 2015vdi.sagepub.comDownloaded from

187Experimentally induced postweaning multisystemic wasting syndrome
containing ethylenediaminetetraacetic acid (EDTA) aftercentrifugation for 10 min at 1,500 3 g. Buffy coat cells andlung lavage cell pellets were suspended in 1 ml of mediumand frozen at 280 C. Viral isolation was done by inoculationof 0.1 ml of thawed material onto PCV-free PK-15 cellsgrown in 24-well tissue culture plates. All viral isolationplates were incubated for 8 days at 39 C in a humidifiedatmosphere containing 5% CO2 and then were frozen at 220C. Isolation of PRRSV was attempted from lung lavage flu-ids and cell pellets using MARC-145 cells grown in 24-welltissue culture plates. The MARC-145 cells were observedfor cytopathic effect for 8 days. As an internal control forcross-contamination of wells in the 24-well plates, speci-mens from the sham-inoculated control pigs were includedin each plate and mixed among wells containing specimensfrom infected pigs.
Viral isolation also was attempted from tonsil, distal ile-um, spleen, liver, kidney, mesenteric lymph node, thymus,lung, midbrain, cerebellum, and cerebrum obtained fromsome pigs postmortem. Approximately 1 g of tissue wasminced with a razor blade and ground in a Tenbroeck tissuegrinder containing 7 ml of medium. The resulting suspensionwas clarified by centrifugation at 1,000 3 g for 15 min. Theclarified fluid was passaged through a 0.22-mm filter andinoculated onto PCV-free PK-15 cells seeded into a 25-cm2
flask. After 5 days of incubation at 39 C, cells and mediumwere removed from the 25-cm2 flask and placed into a 75-cm2 flask containing 15 ml of fresh medium. After 48 hr, themedium in the 75-cm2 flasks was removed and replaced with8 ml of Hanks balanced salt solution containing 300 mM D-glucosamine. After incubation at room temperature for 30min, the solution containing D-glucosamine. was replacedwith 20 ml of fresh medium, and the flasks were incubatedat 39 C for 72 hr. The flasks were then frozen and stored at220 C until tested for viral DNA.
Hematology and serology. White blood cell count, redblood cell (RBC) count, hematocrit, and platelet count wereobtained using an automated cell counter.c A differentialwhite blood cell count was obtained by hand. Sera weretested for antibody against PCV2/688, PCV1, PPV, andPRRSV in 96-well microtitration plates with minor modifi-cations of a described procedure.7,8 The PK-15 cells persis-tently infected with PCV1 or PCV2/688 were seeded intowells of a microtitration plate and grown at 39 C in a hu-midified atmosphere containing 5% CO2. After 48 hr, themedium was removed and 50 ml of Hanks balanced salt so-lution containing 300 mM D-glucosamine was added to eachwell. After a 30-min incubation at room temperature, thatsolution was replaced with 100 ml of fresh medium, and theplates were incubated at 39 C for 48 hr. The plates were thenprepared for immunoperoxidase staining of viral antigen aspreviously described.8 The initial dilution of serum was 1:8in binding buffer. Serial 5-fold dilutions of serum in bindingbuffer were made in wells of the microtitration plate. Re-combinant protein G–peroxidased conjugate was used to la-bel antibody bound to viral antigen. Minor modifications ofthe procedure were made to test for antibody against PRRSV,PPV, or BVDV. For PRRSV, 96-well microtitration plateswere seeded with MARC-145 cells and incubated at 37 Cuntil a cell monolayer was formed. The cell monolayer was
inoculated with the NADC-9 strain of PRRSV and incubatedfor 20 hr before staining. For PPV, PT-1 cells were mixedwith PPV and seeded into 96-well microtitration plates. Theplates were stained after incubation for 20 hr. For BVDV,bovine turbinate (BT) cells were seeded into 96-well micro-titration plates and incubated at 37 C until a cell monolayerhad formed. The BT cells were inoculated with the Singerstrain of type 1 BVDV or the 125C strain of type 2 BVDVand incubated at 37 C for 18 hr before the plates werestained.
Postmortem examination. Pigs inoculated with PCV2/688were euthanized for postmortem examination on PI days 20(n 5 3), 21 (n 5 2), 22 (n 5 2), 23 (n 5 1), 24 (n 5 2), 27(n 5 1), 28 (n 5 3), 35 (n 5 5), 52 (n 5 2), and 125 (n 52). Pigs sham inoculated with medium were euthanized forpostmortem examination on PI days 20 (n 5 1), 24 (n 5 2),and 28 (n 5 1). Gross lesions were noted, and the followingtissues were harvested for microscopic examination: inguinallymph node, internal iliac lymph node, testicle, ovary, uter-us, urinary bladder, kidney, adrenal gland, spleen, pancreas,liver, stomach, duodenum, jejunum, distal ileum, cecum, il-eocecal valve, colon, rectum, mesenteric lymph node, lung,bronchial lymph node, heart, thymus, thyroid, trachea,esophagus, superficial cervical lymph node, mandibularlymph node, mandibular salivary gland, tonsil, cerebellum,cerebrum, midbrain, pituitary gland, and bone marrow. Tis-sues were fixed in 10% neutral buffered formalin for 24–48hr, trimmed, and placed in 70% ethanol for routine process-ing and paraffin embedment. Tissues obtained at postmortemfor detection of viral DNA using the polymerase chain re-action technique (PCR) included kidney, spleen, liver, pan-creas, adrenal gland, mesenteric lymph node, distal ileum,lung, bronchial lymph node, heart, thymus, tonsil, bone mar-row, cerebrum, cerebellum, and midbrain. The tissues werestored frozen at 220 C until processed for PCR. The threecontact exposure pigs were euthanized for postmortem ex-amination on day 43 after initial contact with convalescentpigs. Tissues obtained from those pigs for microscopic ex-amination included liver, kidney, distal ileum, lung, heart,thymus, tonsil, mesenteric lymph node, and inguinal lymphnode.
Polymerase chain reaction. Aligned nucleic acid sequenc-es of PCV1 and PCV2 were used to design PCR primersthat amplified a 580-bp fragment of DNA from either PCV1or PCV2. The sequences of those primers were 286, 59-CCCGCTGCTACATCGAGAAA-39; 865, 59-TCTACA-GCTGGGACAGCAGTTG-39 (base positions from sequenceof PCV2). The PCR reaction mixture contained 200 mMdNTP, 1.5 mM MgCl2, 30 pM each primer, 13 Taq buffer,e
and 0.75 units Taq polymerasee in a final volume of 27 ml.Reaction conditions were 95 C for 10 min (1 cycle) followedby 35 cycles of 95 C for 20 sec, 55 C for 30 sec, and 72 Cfor 40 sec and then a final cycle at 72 C for 5 min. Toconfirm amplified DNA was from PCV2, a second PCR re-action was done on select samples to produce an ampliconof 256 bp. The PCR primers used were 610, 59-TTGCA-GACCCGGAAACCAC-39; 865, 59-TCTACAGCTGGGA-CAGCAGTTG-39 (base positions from sequence of PCV2).To detect possible contamination with PCV1, DNA harvest-ed from select samples of clinical specimens or tissues was
by guest on February 5, 2015vdi.sagepub.comDownloaded from

188 Bolin et al.
Table 1. Mean (range) weight gain (kg/day) for pigs inoculatedwith PCV2.
Piggroup*
Days postinoculation
0–14 14–20 20–28†
A (PF)A (IF)BC (PF)C (IF)D
0.23 (0.15–0.36)0.20 (0.14–0.28)0.22 (0.15–0.31)0.24 (0.20–0.27)0.34 (0.26–0.40)0.37 (0.35–0.39)
0.22 (0.05–0.34)0.34 (0.32–0.41)0.41 (0.31–0.49)0.12 (0.10–0.13)0.23 (0.14–0.33)0.58 (0.53–0.62)
.24 (0.23–0.24)0.34 (0.30–0.38)0.46 (0.36–0.49)
NA0.42 (0.34–0.50)
0.69 (0.60–0.75)
* PF 5 persistent fever; IF 5 intermittent fever. A (PF): n 5 9,aged 20 days old at inoculation; A (IF): n 5 6, 20 days old atinoculation; B: controls, n 5 7, 20 days old at sham inoculation; C(PF): n 5 2, 26 days old at inoculation; C (IF): n 5 6, 26 days oldat inoculation; D: controls, n 5 5, 26 days old at sham inoculation.
† Group A (PF): n 5 2; group A (IF): n 5 5; group B: controls,n 5 6; group C (IF): n 5 5; group D: controls, n 5 3. NA 5 notapplicable (no surviving pigs in this group).
amplified with PCR primers 174, 59-TGTTTGCGGAGAG-GAAGGTTTGG-39 (base positions from sequence of PCV1)and 865, 59-TCTACAGCTGGGACAGCAGTTG-39. Thislast set of primers produced a 692-bp fragment of DNA fromPCV1.
Isolation of PCV2 from clinical specimens was detectedusing the same primer sets to amplify viral DNA from frozenand thawed 24-well tissue culture plates. Medium plus celldebris (180 ml) was mixed with 20 ml lysis buffer (100 mMTris-HCl, pH 8.0, 10 mM EDTA, pH 8.0, 0.5% Triton X-100) and incubated at 100 C for 10 min. After a 5-min in-cubation on ice, the mixture was centrifuged for 3 min at1,200 3 g. Clarified supernatant (3 ml) was removed andadded to 27 ml of PCR reaction mixture for amplification ofviral DNA. To detect viral DNA in tissues, approximately50 mg of tissue was minced with a razor blade and placedin 450 ml of digestion solution (10 mM Tris-HCl, pH 8.3,50 mM KCl, 0.5% polyoxyethylenesorbitan monolaurate, 90mg proteinase K). The digestion mixture was incubated over-night with agitation at 37 C. The digested material was vor-texed briefly, centrifuged at 1,200 3 g for 1 min, incubatedin boiling water for 10 min, chilled on ice for 5 min, andcentrifuged at 18,000 3 g for 3 min. Clarified supernatant(3 ml) was removed and added to 27 ml of PCR reactionmixture for amplification of viral DNA.
In situ hybridization. A 580-bp fragment of DNA fromPCV1 was amplified by PCR using the first set of primers.The amplicon was blunt-end cloned into a plasmid vectorcontaining T7 and Sp6 promoters using a kit.f Using a sec-ond kit,g the cloned DNA was transcribed with either Sp6or T7 RNA polymerase in the presence of digoxigenin-la-beled UTP. This produced sense and antisense digoxigenin-labeled riboprobes. Sections of formalin-fixed tissue weredeparaffinized, treated with proteinase K, and prepared forin situ hybridization as previously described.26,32,34 The hy-bridization buffer was 50% deionized formamide, 33 stan-dard saline citrate (SSC) solution, 13 Denhardt solution, 0.2mg/ml yeast tRNA, 50 mM sodium phosphate (pH 7.4), and10% (w/v) dextran sulfate. The digoxigenin-labeled ribopro-bes were added to the hybridization buffer and heated to 80C for 2 min, cooled on ice, and applied to tissue sectionsfor overnight incubation at 50 C. The tissue sections werewashed twice for 5 min in 23 SSC at room temperature,flooded with a solution of 10 mM Tris-HCl and 0.5 M NaCl(pH 8.0) containing 20 mg/ml ribonuclease A,h and incubatedfor 30 min at 37 C in a humidified chamber. The tissuesections were washed with SSC solutions prewarmed to 50C as follows: 23 SSC for 5 min, 13 SSC for 5 min, and0.53 SSC for 1 hr. A final wash was done with 0.53 SSCat room temperature for 5 min. Kits were used to block non-specific reactionsi and for enzyme-linked immunoassay andenzyme-catalyzed color reactions.j
Results
Clinical signs and rate of weight gain. A body tem-perature of .40 C (fever) was recorded rarely fromday 0 to day 28 PI in the 12 sham-inoculated controlpigs in groups B and D. During that period, this ele-vated temperature was never recorded on more than 2
successive days nor on more than 3 days total for anypig. Other signs of disease were not observed in thecontrol pigs. For 12 of 23 pigs from groups A and Cinoculated with PCV2/688, fever was recorded inter-mittently between days 12 and 20 PI; however, fevernever was recorded on more than 2 successive daysnor on more than 4 days total. Palpable enlargementof the inguinal, prefemoral, and popliteal lymph nodeswas noted in 9 of those 12 pigs from about day 16 today 22 PI. Serum from 1 of those 12 pigs appearedicteric at postmortem on day 29 PI. A persistent feveroccurred in 11 of 23 pigs from groups A and C in-oculated with PCV2/688. In those pigs, fever was re-corded on 7–17 successive days, beginning on aboutday 12 PI. Those pigs became depressed, and mostwere euthanized for humane purposes between days20 and 28 PI. During the period of persistent fever,the temperature fluctuated from 40.1 C to 41.4 C. Thehair on those pigs became erect, and many pigs ap-peared to shiver. Palpable enlargement of inguinal,prefemoral, and popliteal lymph nodes was noted inall affected pigs at about day 16 PI and persisted for4–7 days. The skin of 6 of the 11 pigs became yel-lowish between days 21 and 26 PI. During that time,the serum from those 6 pigs had a distinct yellow toyellow-brown color. Hyperpnea was observed in 2pigs with persistent fever, but diarrhea was not ob-served. The control pigs placed in contact with in-fected pigs did not develop a fever and did not showclinical signs of disease.
The average daily rate of weight gain between days14 and 28 PI was lower in groups of pigs exposed toPCV2/688 than in groups of control pigs of similarages (Table 1). During that time, the amount of fecalmaterial in pens of pigs inoculated with virus was no-ticeably less than that in pens of control pigs. Three
by guest on February 5, 2015vdi.sagepub.comDownloaded from
189Experimentally induced postweaning multisystemic wasting syndrome
Table 2. Geometric mean (log10) titer of antibody against PCV2or PCV1 in pigs on select days after inoculation.
Virus*
Age atinocu-lation(days)
Antibody titer
Day15† Day 19 Day 24 Day 28 Day 35 Day 42
PCV2/688n
PCV1n
PCV2/688n
PCV1n
20
20
25
25
08081.6808
1.114
014
2.2808
ND
ND
3.480.98
2.470.974.051.95
3.041.844.152.65
4.142.144.432.63
* The number of samples of serum (n) tested at each time pointafter inoculation with virus varied due to experimental design anddeath of pigs.
† Antibodies against PCV1 and PCV2 were not detected on orbefore day 12 PI.
of 11 pigs that had persistent fever lost body weightin the last 2–3 days of life. Only 2 pigs that had per-sistent fever survived to day 28 PI, and rate of weightgain for those pigs was about half that of control pigs.
Viral isolation from clinical specimens. PCV1,PCV2, PPV, BVDV, and PRRSV were not isolatedfrom clinical specimens obtained from control pigs.Isolation of PCV2, as detected by PCR, was infrequentfrom nasal, rectal, or tonsil swab specimens obtainedfrom pigs inoculated with PCV2/688. From thosespecimens, virus was isolated between only days 12and 19 PI. During that period, virus was isolated from6 of 28 rectal swabs obtained from 5 of 14 pigs. Virusalso was isolated from 3 of 28 tonsil swabs and 1 of28 nasal swabs obtained from the same 14 pigs. Viralisolation from rectal, tonsil, or nasal swabs was notdetected before day 12 or after day 19 PI. Viral iso-lation from urine was detected from only 1 pig at 28days PI. Between days 20 and 28 PI, virus was isolatedfrom lung lavage fluid from 5 of 14 pigs and from thelung lavage cell pellet from 11 of 14 pigs. Virus wasfirst isolated from buffy coat or serum at 12 days PI.Between days 12 and 35 PI, virus was isolated from49 of 67 samples of buffy coat cells obtained from 23pigs. Between days 12 and 28 PI, virus was isolatedfrom 20 of 60 samples of serum obtained from 23 pigs.Virus was not isolated from serum after day 28 PI.
Hematology and serology. Most pigs inoculatedwith virus had normal hematologic profiles throughday 28 PI, regardless of severity of clinical disease.Four pigs that showed signs of disease developed neu-tropenia (,1,000 neutrophils/mm3), lymphopenia(,1,000 lymphocytes/mm3), or anemia (,4 3 106
RBC/mm3, hematocrit ,21 %). Those hematologic ab-normalities were seen after day 19 PI.
No antibodies against BVDV, PPV, or PRRSV weredetected in serum obtained from any pig. Antibodyagainst PCV2/688 was detected in serum between days15 and 19 PI in all pigs of group C that were inocu-lated with virus at 25 days of age. Antibody againstPCV2/688 was first detected in serum on day 19 PI inpigs of group A that were inoculated with virus at 20days of age. However, 4 of 15 pigs in group A failedto produce detectable concentrations of antibody in se-rum before death at days 20–24 PI. The titer of anti-body against PCV2/688 in serum increased steadily forabout 3 weeks after first detection. Antibody in serumthat cross-reacted with PCV1 was not detected untilseveral days after detection of antibody against PCV2/688. The titer in serum of antibody against PCV1 al-ways was less than the titer of antibody against PCV2/688 (Table 2).
Postmortem examination. Gross or microscopic le-sions were not observed in control pigs. Gross lesionswere most prominent in pigs that developed persistent
fever after inoculation with PCV2/688. Moderate tomarked lymphadenopathy of inguinal, prefemoral, in-ternal iliac, mesenteric, gastric, bronchial, and/or man-dibular lymph nodes was a common gross lesion ob-served from 20 to 35 days PI. Other gross lesions ob-served during that period included generalized icterusof skin, subcutis, adipose tissue, sclera of the eye, in-testines, and liver. Ascites was common in pigs thatwere icteric. Lesions in the kidney included scatteredwhite foci on the cortical surface, often accompaniedby pitting of the surface, and occasional hemorrhagein the medulla. After day 35 PI, scattered white fociand pitting on the surface of the kidney was the mostfrequent gross lesion. Scattered white foci and pittingon the surface of the kidney was seen in 2 of 3 controlpigs euthanized 43 days after being placed in contactwith convalescent pigs that had been inoculated withPCV2/688.
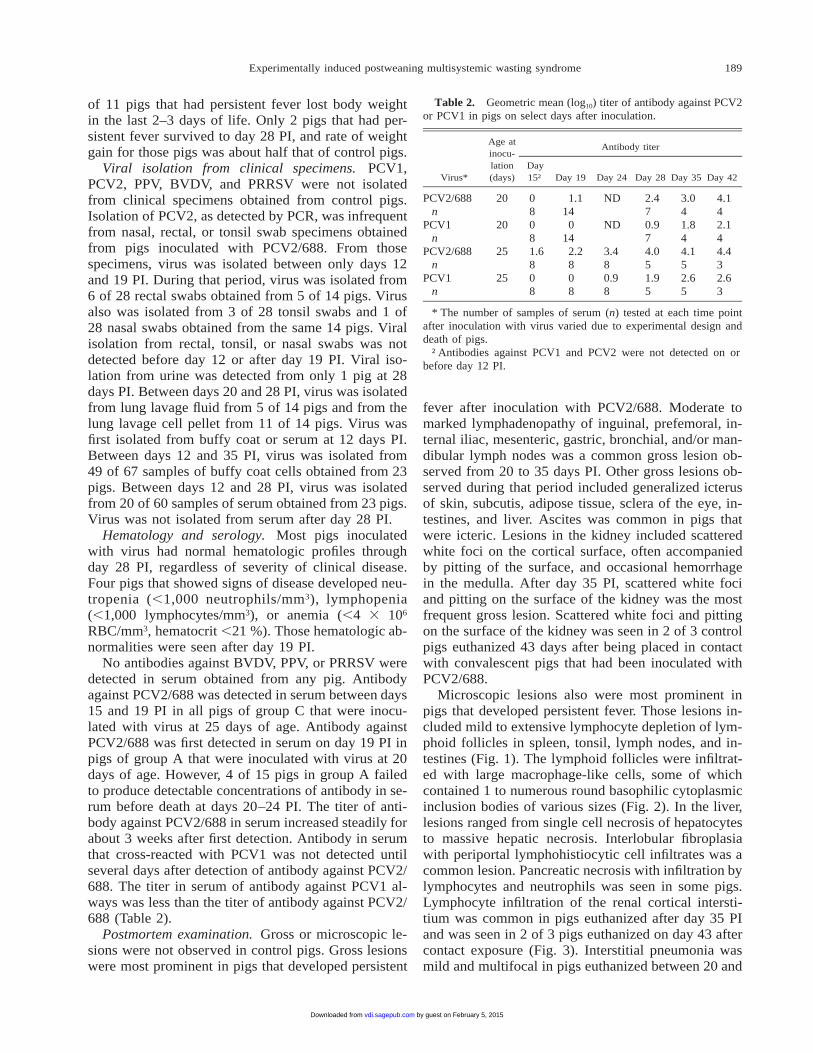
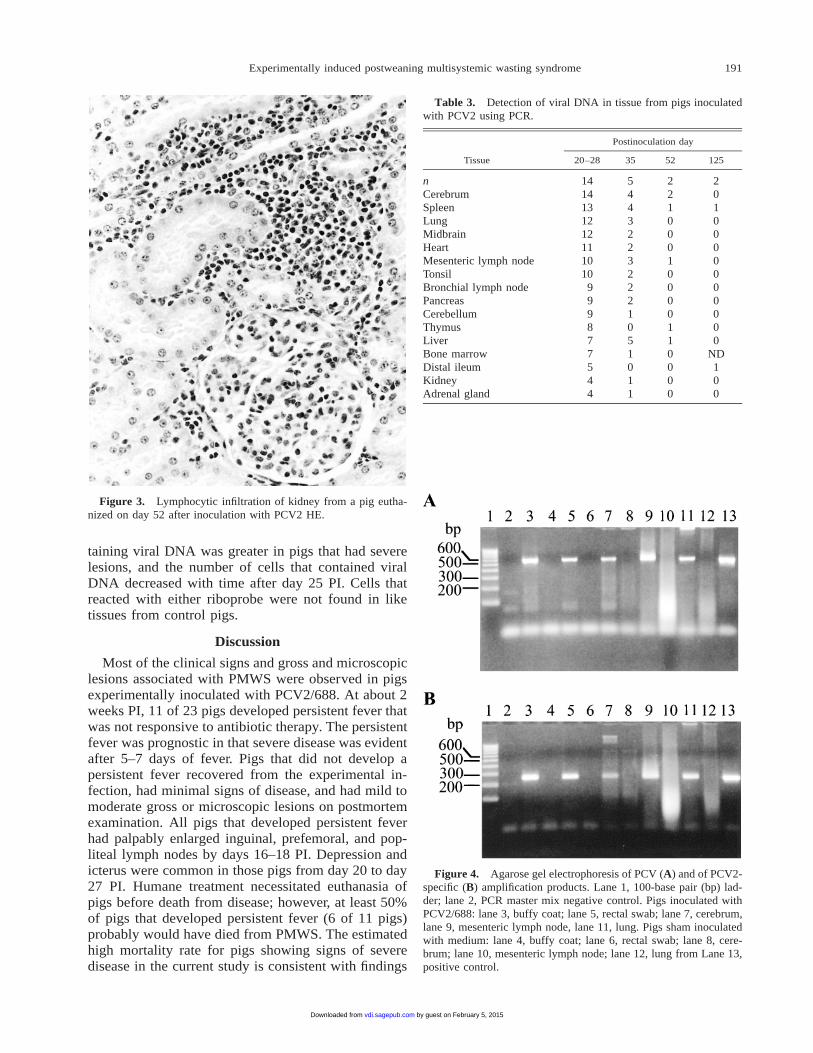
Microscopic lesions also were most prominent inpigs that developed persistent fever. Those lesions in-cluded mild to extensive lymphocyte depletion of lym-phoid follicles in spleen, tonsil, lymph nodes, and in-testines (Fig. 1). The lymphoid follicles were infiltrat-ed with large macrophage-like cells, some of whichcontained 1 to numerous round basophilic cytoplasmicinclusion bodies of various sizes (Fig. 2). In the liver,lesions ranged from single cell necrosis of hepatocytesto massive hepatic necrosis. Interlobular fibroplasiawith periportal lymphohistiocytic cell infiltrates was acommon lesion. Pancreatic necrosis with infiltration bylymphocytes and neutrophils was seen in some pigs.Lymphocyte infiltration of the renal cortical intersti-tium was common in pigs euthanized after day 35 PIand was seen in 2 of 3 pigs euthanized on day 43 aftercontact exposure (Fig. 3). Interstitial pneumonia wasmild and multifocal in pigs euthanized between 20 and
by guest on February 5, 2015vdi.sagepub.comDownloaded from
190 Bolin et al.
Figure 1. Cellular depletion in a Peyer’s patch of the distal il-eum from a pig euthanized on day 26 after inoculation with PCV2HE.
Figure 2. Cytoplasmic inclusion bodies (arrows) in a depletedB-cell region of a lymphoid follicle from a pig euthanized on day24 after inoculation with PCV2 HE.
28 days PI and rare in pigs euthanized after day 28PI. Lymphocytic myocarditis was mild and rare beforeday 35 PI but became a more common lesion after day35 PI.
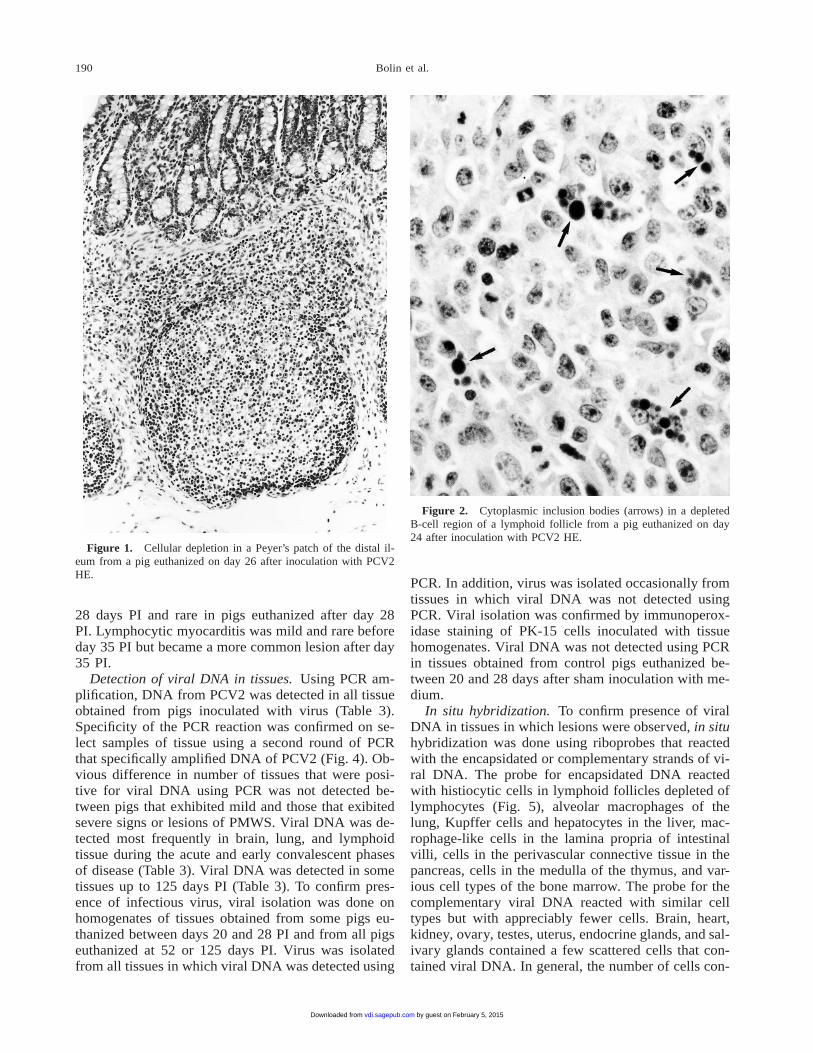
Detection of viral DNA in tissues. Using PCR am-plification, DNA from PCV2 was detected in all tissueobtained from pigs inoculated with virus (Table 3).Specificity of the PCR reaction was confirmed on se-lect samples of tissue using a second round of PCRthat specifically amplified DNA of PCV2 (Fig. 4). Ob-vious difference in number of tissues that were posi-tive for viral DNA using PCR was not detected be-tween pigs that exhibited mild and those that exibitedsevere signs or lesions of PMWS. Viral DNA was de-tected most frequently in brain, lung, and lymphoidtissue during the acute and early convalescent phasesof disease (Table 3). Viral DNA was detected in sometissues up to 125 days PI (Table 3). To confirm pres-ence of infectious virus, viral isolation was done onhomogenates of tissues obtained from some pigs eu-thanized between days 20 and 28 PI and from all pigseuthanized at 52 or 125 days PI. Virus was isolatedfrom all tissues in which viral DNA was detected using
PCR. In addition, virus was isolated occasionally fromtissues in which viral DNA was not detected usingPCR. Viral isolation was confirmed by immunoperox-idase staining of PK-15 cells inoculated with tissuehomogenates. Viral DNA was not detected using PCRin tissues obtained from control pigs euthanized be-tween 20 and 28 days after sham inoculation with me-dium.
In situ hybridization. To confirm presence of viralDNA in tissues in which lesions were observed, in situhybridization was done using riboprobes that reactedwith the encapsidated or complementary strands of vi-ral DNA. The probe for encapsidated DNA reactedwith histiocytic cells in lymphoid follicles depleted oflymphocytes (Fig. 5), alveolar macrophages of thelung, Kupffer cells and hepatocytes in the liver, mac-rophage-like cells in the lamina propria of intestinalvilli, cells in the perivascular connective tissue in thepancreas, cells in the medulla of the thymus, and var-ious cell types of the bone marrow. The probe for thecomplementary viral DNA reacted with similar celltypes but with appreciably fewer cells. Brain, heart,kidney, ovary, testes, uterus, endocrine glands, and sal-ivary glands contained a few scattered cells that con-tained viral DNA. In general, the number of cells con-
by guest on February 5, 2015vdi.sagepub.comDownloaded from
191Experimentally induced postweaning multisystemic wasting syndrome
Figure 3. Lymphocytic infiltration of kidney from a pig eutha-nized on day 52 after inoculation with PCV2 HE.
Table 3. Detection of viral DNA in tissue from pigs inoculatedwith PCV2 using PCR.
Tissue
Postinoculation day
20–28 35 52 125
nCerebrumSpleenLungMidbrainHeartMesenteric lymph nodeTonsilBronchial lymph nodePancreasCerebellumThymusLiverBone marrowDistal ileumKidneyAdrenal gland
1414131212111010
999877544
54432232221051011
22100010000110000
2010000000000
ND100
Figure 4. Agarose gel electrophoresis of PCV (A) and of PCV2-specific (B) amplification products. Lane 1, 100-base pair (bp) lad-der; lane 2, PCR master mix negative control. Pigs inoculated withPCV2/688: lane 3, buffy coat; lane 5, rectal swab; lane 7, cerebrum,lane 9, mesenteric lymph node, lane 11, lung. Pigs sham inoculatedwith medium: lane 4, buffy coat; lane 6, rectal swab; lane 8, cere-brum; lane 10, mesenteric lymph node; lane 12, lung from Lane 13,positive control.
taining viral DNA was greater in pigs that had severelesions, and the number of cells that contained viralDNA decreased with time after day 25 PI. Cells thatreacted with either riboprobe were not found in liketissues from control pigs.
Discussion
Most of the clinical signs and gross and microscopiclesions associated with PMWS were observed in pigsexperimentally inoculated with PCV2/688. At about 2weeks PI, 11 of 23 pigs developed persistent fever thatwas not responsive to antibiotic therapy. The persistentfever was prognostic in that severe disease was evidentafter 5–7 days of fever. Pigs that did not develop apersistent fever recovered from the experimental in-fection, had minimal signs of disease, and had mild tomoderate gross or microscopic lesions on postmortemexamination. All pigs that developed persistent feverhad palpably enlarged inguinal, prefemoral, and pop-liteal lymph nodes by days 16–18 PI. Depression andicterus were common in those pigs from day 20 to day27 PI. Humane treatment necessitated euthanasia ofpigs before death from disease; however, at least 50%of pigs that developed persistent fever (6 of 11 pigs)probably would have died from PMWS. The estimatedhigh mortality rate for pigs showing signs of severedisease in the current study is consistent with findings
by guest on February 5, 2015vdi.sagepub.comDownloaded from

192 Bolin et al.
Figure 5. A depleted lymphoid follicle from a pig euthanized26 days after inoculation with PCV2. A PCV riboprobe was usedfor in situ hybridization. The darkly stained cells contain viral DNA.Nuclear fast red counterstain, alkaline phosphatase stain for viralDNA.
from field outbreaks of PMWS, in which mortalityrates average about 80% for pigs showing disease.17
Lesions previously associated with PMWS that wereseen in the current study included lymphadenopathy,icteric tissues and ascitic fluid, hepatic cell necrosis,depletion of small lymphoid cells in lymphoid folli-cles, multifocal granulomatous inflammation of liver,kidney, and lymphoid tissue, and cytoplasmic inclu-sion bodies in histiocytic cells in depleted lymphoidfollicles.4,11,13,19–22,25,28,30,33 In contrast to reports fromfield outbreaks of PMWS,4,17,19,21,28,30,33 diarrhea andskin pallor were not observed in the current study. Hy-perpnea was observed in pigs that had fever, but re-spiratory distress was not observed. The interstitialpneumonia seen on microscopic examination of lungwas multifocal and mild, as compared with extensivebronchointerstitial pneumonia seen frequently in thefield. Multinucleate syncytial cells often found in lungand lymphoid tissue from pigs with naturally occurringPMWS were rarely observed in the currentstudy.10,11,13,19,21,25,33
In previous reports, experimental inoculation of pigswith PCV1 or PCV2 either failed to induce disease or
lesions or induced clinical disease and lesions that pri-marily varied from mild to moderate in the majorityof pigs and were moderate to severe in 1 pig.2–4,6,20,22,38
Those findings are consistent with the mild disease andmild to moderate lesions seen in the present study inpigs that had infrequent fever after inoculation withPCV2/688. The reduced rate of weight gain observedin pigs inoculated with PCV2/688 was reported in con-ventional pigs after intranasal inoculation with a ho-mogenate of tissues obtained from a field outbreak ofPMWS.6
Severe disease and extensive lesions consistent withPMWS have been reported after simultaneous infec-tion of pigs with PCV2 and PPV.2,14,20,22 Mixed infec-tions of PCV2 with PCV1, PPV, PRRSV, pseudorabiesvirus, and Chlamydia occur during field outbreaks ofPMWS.9,12,13,19,23,24,31,32 The contribution of infectiousagents other than PCV2 to PMWS is unknown, butpathogenic synergism between infectious agents maybe a factor in the severity of PMWS. During this study,the stock PCV2 used to inoculate pigs was tested foradventitious agents, but none were found. Serum ob-tained from pigs was tested for antibody against PPV,PRRSV, and BVDV, which were considered likelycontaminants of the stock virus. No antibodies againstthose viruses were detected in serum obtained through125 days PI.
The in vitro growth characteristics of PCV in cellculture makes viral isolation time consuming and de-tection of isolated virus difficult.39 These difficultiesprobably have prompted numerous reports in whichPCR was the primary method used for detection ofPCV in tissues or cell cultures.11,14–16,23,24,28,29,31 There-fore, the utility of clinical specimens and tissues forviral isolation or for detection of viral DNA was eval-uated in the present study. The duration of time afterinitial infection with PCV2 that viral DNA could bedetected using PCR also was evaluated.
Viral DNA was detected after inoculation of cellcultures with each type of clinical specimen tested, andviral DNA was detected in each type of tissues tested.The frequency of detection of viral DNA and lengthof time after date of infection that viral DNA was de-tected differed with the clinical specimen or tissue. Ofthe clinical specimens obtained ante mortem, PCV2was detected most frequently after viral isolation frombuffy coat cells. Virus was detected in multiple sam-ples of buffy coat cells obtained from each pig overan extended period of time. Detection of virus frombuffy coat cells was successful $2 weeks after detec-tion of antibody against PCV2 in serum and up to 5weeks after the date of experimental infection. Detec-tion of virus in serum was accomplished readily be-ginning about 2 weeks PI. Appearance of antibody inserum correlated with loss of cell culture infective vi-
by guest on February 5, 2015vdi.sagepub.comDownloaded from

193Experimentally induced postweaning multisystemic wasting syndrome
rus in serum. Although others have had success withisolation of PCV from nasal swab specimens and fecalsamples,38 in the current study virus was detected innasal, rectal, and tonsil swab specimens only infre-quently between 12 and 19 days PI. The methods usedin the current study for viral isolation and for PCRmay not have allowed efficient detection of virus inthose specimens. The rapid seroconversion of the con-tact exposure pigs in the current study certainly sug-gests that excretions and/or secretions from the bodycontain infectious virus.
The tissues in which viral DNA was detected mostfrequently using PCR were brain, lung, and lymphoidtissue. Viral DNA was detected regularly in those tis-sues from day 20 PI through day 35 PI and infrequent-ly through day 125 PI. In many studies that have usedPCR or in situ hybridization,6,10,12,15,19,23–25,28–33 lung andlymphoid tissue have been reported to contain viralDNA. Viral antigen has been reported in brain from apig experimentally coinfected with PCV2 and PPV, butPCV2 was not detected in brain tissue from pigs ex-perimentally infected with PCV2 alone.2
The presence of viral DNA in tissue was confirmedusing in situ hybridization. Numerous cells containingviral DNA were found in lymphoid tissue, liver, andlung of pigs euthanized between days 20 and 28 PI.Similar to previous reports on PMWS,4,6,9,10,25,28,32,33,35
in the current study the cell types most frequentlyfound to contain viral DNA were of monocyte/mac-rophage lineage. Although brain tissue frequently waspositive for PCV2 using PCR, few cells in the braincontained viral DNA when examined using in situ hy-bridization. Those cells usually were located in theperivascular connective tissue adjacent to small bloodvessels in the brain. The conditions used for in situhybridization, the sensitivity of the procedure, or thechoice of probe might account for limited detection ofviral DNA in brain. The distribution of cells and celltypes that contained viral DNA in pancreas, kidney,intestine, and bone marrow were consistent with thatin previous reports.4,6,9,10,25,28,32,33 In the heart, scatteredmyocytes and cells in the perivascular connective tis-sue adjacent to small blood vessels contained viralDNA. That finding is consistent with a report of de-tection of PCV2 in the heart of aborted porcine fetus-es.41
Viral DNA was detected using PCR in tissue ob-tained postmortem on days 52 and 125 PI. Attemptsat isolation of infective virus from those tissues weresuccessful. The extended duration after experimentalinoculation with PCV2 that infective virus was isolat-ed from tissue indicates that a persistent infection wasestablished. This finding has importance for diseasecontrol strategies because it suggests that apparentlyhealthy adult pigs may be carriers of PCV2. Contact
transmission of virus from older to younger pigs alsomay be an important means for spread of PCV2. At42 days PI, pigs in this study apparently still shed in-fectious virus; seronegative control pigs all producedantibody within 3 weeks of contact exposure.
The serology performed in the present study con-firmed results of previous studies in which PCV1 andPCV2 were found to be antigenically similar but couldbe differentiated by serologic tests.4,5,13 In the currentstudy, the titer of antibody against PCV2 always wasmuch greater than the titer of antibody against PCV1.Thus, differential serologic tests could be used to iden-tify the genotype of PCV in infected herds.
Acknowledgements
We thank Theresa Rahner and Shari Steadham for theirexcellent technical support and Mary Sue Brown and ChuckGreiner for photographic assistance.
Sources and manufacturers
a. American BioResearch Laboratories, Sevierville, TN.b. Dr. W. L. Mengeling, USDA, ARS, National Animal Disease
Center, Ames, IA.c. Coulter T-890, Coulter Corp., Hialeah, FL.d. Zymed Laboratories, San Francisco, CA.e. Boehringer Mannheim, Indianapolis, IN.f. Zero Blunt TOPO PCR cloning kit, Invitrogen, Carlsbad, CA.g. DIG RNA labeling kit (Sp6/T7), Boehringer Mannheim, Indi-
anapolis, IN.h. Sigma Chemical Co., St. Louis, MO.i. DIG wash and block buffer set, Boehringer Mannheim, Indian-
apolis, IN.j. DIG nucleic acid detection kit, Boehringer Mannheim, Indian-
apolis, IN.
References
1. Allan GM, Ellis JA: 2000, Porcine circoviruses: a review. J VetDiagn Invest 12:3–14.
2. Allan GM, Kennedy S, McNeilly F, et al.: 1999, Experimentalreproduction of severe wasting disease by co-infection of pigswith porcine circovirus and porcine parvovirus. J Comp Pathol121:1–11.
3. Allan GM, McNeilly F, Cassidy JP, et al.: 1995, Pathogenesisof porcine circovirus; experimental infections of colostrum de-prived piglets and examination of pig foetal material. Vet Mi-crobiol 44:49–64.
4. Allan GM, McNeilly F, Kennedy S, et al.: 1998, Isolation ofporcine circovirus-like viruses from pigs with a wasting diseasein the USA and Europe. J Vet Diagn Invest 10:3–10.
5. Allan GM, McNeilly F, Meehan BM, et al.: 1999, Isolation andcharacterisation of circoviruses from pigs with wasting syn-dromes in Spain, Denmark and Northern Ireland. Vet Microbiol66:115–123.
6. Balasch M, Segales J, Rosell C, et al.: 1999, Experimental in-oculation of conventional pigs with tissue homogenates frompigs with post-weaning multisystemic wasting syndrome. JComp Pathol 121:139–148.
7. Bolin SR, Matthews PJ, Ridpath JF: 1991, Methods for detec-tion and frequency of contamination of fetal bovine serum withbovine viral diarrhea virus and antibodies against bovine viraldiarrhea virus. J Vet Diagn Invest 3:199–203.
by guest on February 5, 2015vdi.sagepub.comDownloaded from

194 Bolin et al.
8. Bolin SR, Ridpath JF: 1998, Prevalence of bovine viral diarrheavirus genotypes and antibody against those genotypes in fetalbovine serum. J Vet Diagn Invest 10:135–139.
9. Carrasco L, Segales J, Bautista MJ, et al.: 2000, Intestinal chla-mydial infection concurrent with postweaning multisystemicwasting syndrome in pigs. Vet Rec 146:21–23.
10. Choi C, Chae C: 1999, In-situ hybridization for the detection ofporcine circovirus in pigs with postweaning multisystemic wast-ing syndrome. J Comp Pathol 121:263–270.
11. Choi C, Chae C, Clark EG: 2000, Porcine postweaning multi-systemic wasting syndrome in Korean pigs: detection of porcinecircovirus 2 infection by immunohistochemistry and polymerasechain reaction. J Vet Diagn Invest 12:151–153.
12. Ellis JA, Bratanich A, Clark EG, et al.: 2000, Coinfection byporcine circoviruses and porcine parvovirus in pigs with natu-rally acquired postweaning multisystemic wasting syndrome. JVet Diagn Invest 12:21–27.
13. Ellis J, Hassard L, Clark E, et al.: 1998, Isolation of circovirusfrom lesions of pigs with postweaning multisystemic wastingsyndrome. Can Vet J 39:44–51.
14. Ellis J, Krakowka S, Larimore M, et al.: 1999, Reproduction ofpostweaning multisystemic wasting syndrome in gnotobioticpigs. J Vet Diagn Invest 11:3–14.
15. Hamel AL, Lin LL, Nayar GPS: 1998, Nucleotide sequence ofporcine circovirus associated with postweaning multisystemicwasting syndrome in pigs. J Virol 72:5262–5267.
16. Hamel AL, Lin LL, Sachvie C, et al.: 2000, PCR detection andcharacterization of type-2 porcine circovirus. Can J Vet Res 64:44–52.
17. Harding JCS, Clark EG: 1997, Recognizing and diagnosingpostweaning multisystemic wasting syndrome (PMWS). SwineHealth Prod 5:201–203.
18. Harding JCS, Clark EG, Strokappe JH, et al.: 1998, Postweaningmultisystemic wasting syndrome: epidemiology and clinical pre-sentation. Swine Health Prod 6:249–254.
19. Illanes O, Lopez A, Miller L, et al.: 2000, Lesions associatedwith postweaning multisystemic wasting syndrome in pigs fromPrince Edward Island, Canada. J Vet Diagn Invest 12:146–150.
20. Kennedy S, Moffett D, McNeilly F, et al.: 2000, Reproductionof lesions of postweaning multisystemic wasting syndrome byinfection of conventional pigs with porcine circovirus type 2alone or in combination with porcine parvovirus. J Comp Pathol122:9–24.
21. Kiupel M, Stevenson GW, Mittal SK, et al.: 1998, Circovirus-like viral associated disease in weaned pigs in Indiana. VetPathol 35:303–307.
22. Krakowa S, Ellis JA, Meehan B, et al.: 2000, Viral wastingsyndrome of swine: experimental reproduction of postweaningmultisystemic wasting syndrome in gnotobiotic swine by coin-fection with porcine circovirus 2 and porcine parvovirus. VetPathol 37:254–263.
23. Larochelle R, Antaya M, Morin M, Magar R: 1999, Typing ofporcine circovirus in clinical specimens by multiplex PCR. JVirol Methods 80:69–75.
24. Larochelle R, Morin M, Antaya M, Magar R: 1999, Identifica-tion and incidence of porcine circovirus in routine field cases inQuebec as determined by PCR. Vet Rec 145:140–142.
25. McNeilly F, Kennedy S, Moffet D, et al.: 1999, A comparison
of in situ hybridization and immunohistochemistry for the de-tection of a new porcine circovirus in formalin-fixed tissuesfrom pigs with post-weaning multisystemic wasting syndrome(PMWS). J Virol Methods 80:123–128.
26. Meehan BM, Creelan JL, McNulty MS, Todd D: 1997, Se-quence of porcine circovirus DNA: affinities with plant circo-viruses. J Gen Virol 78:221–227.
27. Meehan BM, McNeilly F, Todd D, et al.: 1998, Characterizationof novel circovirus DNAs associated with wasting syndromesin pigs. J Gen Virol 79:2171–2179.
28. Morosov I, Sirinarumitr T, Sorden S, et al.: 1998, Detection ofa novel strain of porcine circovirus in pigs with postweaningmultisystemic wasting syndrome. J Clin Microbiol 36:2535–2541.
29. Nayar GPS, Hamel A, Lin L: 1997, Detection and characteriza-tion of porcine circovirus associated with postweaning multi-systemic wasting syndrome in pigs. Can Vet J 38:385–386.
30. Onuki A, Abe K, Togashi K, et al.: 1999, Detection of porcinecircovirus from lesions of a pig with wasting disease in Japan.J Vet Med Sci 61:1119–1123.
31. Ouardani M, Wilson L, Jette R, et al.: 1999, Multiplex PCR fordetection and typing of porcine circoviruses. J Clin Microbiol37:3917–3924.
32. Rodrı´guez-Arrioja GM, Segales J, Rossel C, et al.: 1999, Au-jeszky’s disease virus infection concurrent with postweaningmultisystemic wasting syndrome in pigs. Vet Rec 144:152–153.
33. Rosell C, Segales J, Plana-Duran J, et al.: 1999, Pathological,immunohistochemical, and in-situ hybridization studies of nat-ural cases of postweaning multisystemic wasting syndrome(PMWS) in pigs. J Comp Pathol 120:59–78.
34. Sirinarumitr T, Paul PS, Kluge JP, Halbur PG: 1996, In situhybridization technique for the detection of swine enteric andrespiratory coronaviruses, transmissible gastroenteritis virus(TGEV) and porcine respiratory coronavirus (PRCV), in for-malin-fixed paraffin-embedded tissues. J Virol Methods 56:149–160.
35. Sorden SD, Harms PA, Nawagitigul P, et al.: 1999, Developmentof a polyclonal-antibody-based immunohistochemical methodfor the detection of type 2 porcine circovirus in formalin-fixed,paraffin-embedded tissue. J Vet Diagn Invest 11:528–530.
36. Stevenson GW, Kuipel M, Mittal SK, Kanitz CL: 1999, Ultra-structure of porcine circovirus in persistently infected PK-15cells. Vet Pathol 36:368–378.
37. Tischer, I, Gelderblom H, Vettermann W, Koch MA, et al.: 1982,A very small porcine virus with circular single-stranded DNA.Nature 295:64–66.
38. Tischer I, Mields W, Wolff D, et al.: 1986, Studies on epide-miology and pathogenicity of porcine circovirus. Arch Virol 91:271–276.
39. Tischer I, Peters D, Rasch R, Pociuli S: 1987, Replication ofporcine circovirus: induction by glucosamine and cell cycle de-pendence. Arch Virol 96:39–57.
40. Tischer I, Rasch R, Tochtermann G: 1974, Characterization ofpapovavirus and picornavirus-like particles in permanent pigkidney cell lines. Zentralbl Bakteriol Mikrobiol Hyg Ser A 226:153–167.
41. West KH, Bystrom JM, Wojnarowicz C, et al.: 1999, Myocar-ditis and abortion associated with intrauterine infection of showswith porcine circovirus 2. J Vet Diagn Invest 11:530–532.
by guest on February 5, 2015vdi.sagepub.comDownloaded from
Related Documents











