1 Posttranslational Modification and Targeting of Proteins Graduate Biochemistry Term 2/2016 Assist. Prof. Dr. Panida Khunkaewla School of Chemistry, Institute of Science Suranaree University of Technology

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

11
Posttranslational Modification
and Targeting of Proteins
Graduate Biochemistry
Term 2/2016
Assist. Prof. Dr. Panida Khunkaewla
School of Chemistry, Institute of Science
Suranaree University of Technology

22
What is posttranslational Modification?
“Modification of nascent protein by adding groups or cleavage
of some parts to get mature protein”
Key wards:
Adding
Deleting

33
Diversity of Posttranslational modification of proteins
Cleavage of signal peptides
Phosphorylation
Amidation
Glycosylation
Hydroxylation
Ubiquitination
Addition of prosthetic groups
Iodination
Adenylation
Sulfonation
Prenylation
Myristoylation
Acylation
Acetylation
Methylation
Oxidative crosslinking
N-Glutamyl cyclization
Carboxylation

4
Nucleus acetylation, phosphorylation
Lysosome mannose-6-phosphate labelled N-linked sugar
Mitochondria N-formyl acylation
Golgi N- and O-linked ologosaccharide, sulfation,
palmitoylation
ER N-linked oligosaccharide, GPI-anchor
Cytosol acetylation, methylation, phosphorylation,
Ribosome myristoylation
Plasma membrane N- and O-glycosylation, GPI-anchor
Extracelullar fluid N- and O-glycosylation, acetylation,
phosphorylation
Extracellular matrix N- and O-glycosylation, phosphorylation,
hydroxylation
Location Modification
5
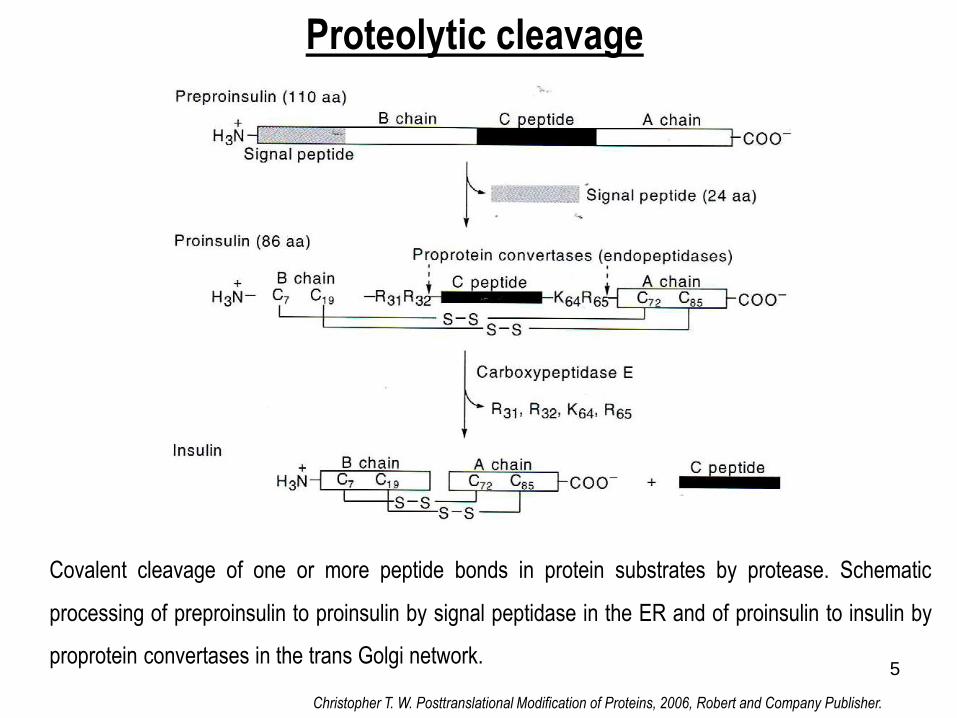
Proteolytic cleavage
Covalent cleavage of one or more peptide bonds in protein substrates by protease. Schematic
processing of preproinsulin to proinsulin by signal peptidase in the ER and of proinsulin to insulin by
proprotein convertases in the trans Golgi network.
Christopher T. W. Posttranslational Modification of Proteins, 2006, Robert and Company Publisher.
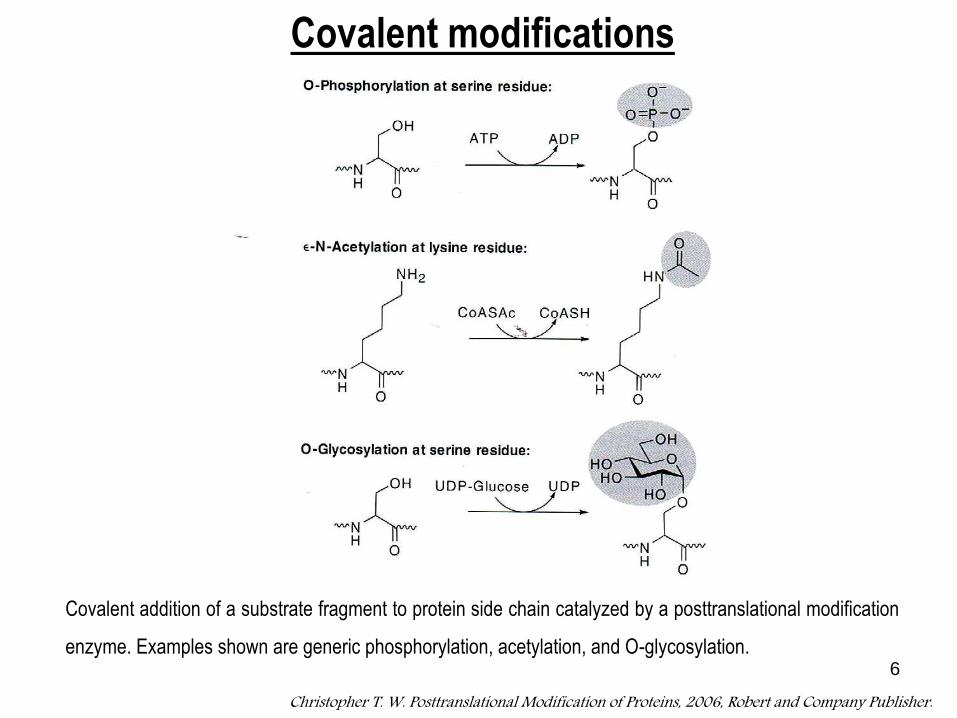
6
Covalent addition of a substrate fragment to protein side chain catalyzed by a posttranslational modification
enzyme. Examples shown are generic phosphorylation, acetylation, and O-glycosylation.
Christopher T. W. Posttranslational Modification of Proteins, 2006, Robert and Company Publisher.
Covalent modifications
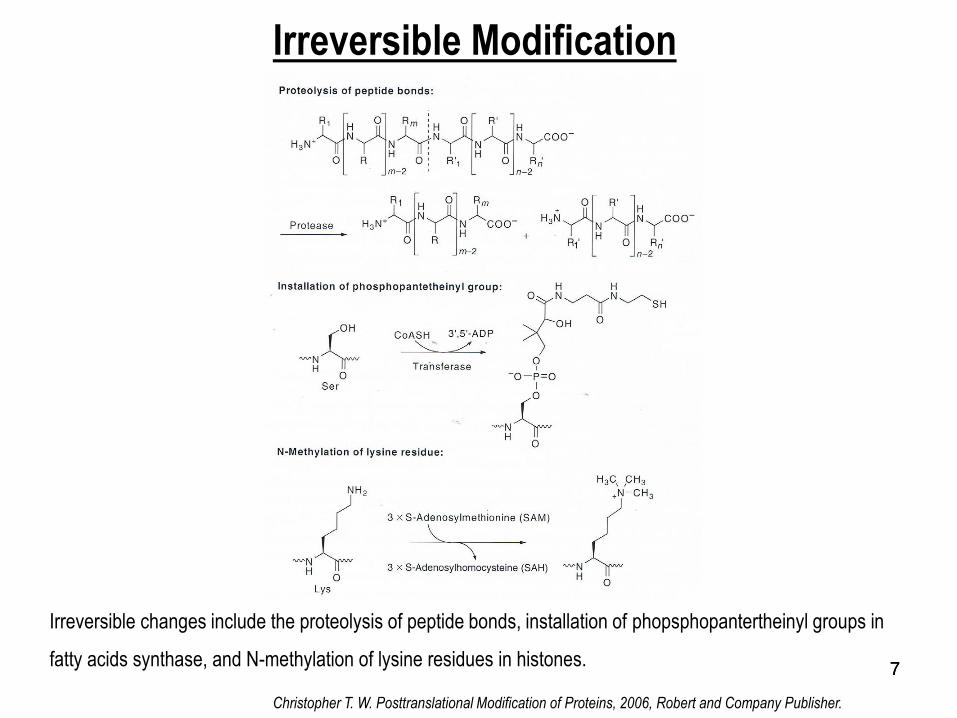
77
Irreversible Modification
Irreversible changes include the proteolysis of peptide bonds, installation of phopsphopantertheinyl groups in
fatty acids synthase, and N-methylation of lysine residues in histones.
Christopher T. W. Posttranslational Modification of Proteins, 2006, Robert and Company Publisher.

8
Reversible Modifications
Reversible covalnt modifications including protein phosphorylations (due to balance of kinase and phosphatase),
histone acetylation (due to the balance of histone acetyltransferase and histone deacetylase), and protein
ubiquitylations (due to the balance of ligases and deubiquitylating hydrolase
Christopher T. W. Posttranslational Modification of Proteins, 2006, Robert and Company Publisher.

9
Purpose of posttranslational modification.
Targeting of protein
Stability of protein
Function of protein
Control protein activity

10
Targeting of proteins
“How are synthesized proteins directed to their final cellular destination?”
Secreted protein, membrane protein, inclusion protein in lysosomes
Protein destines to mitochondria Protein destines to chloroplast
Protein destines to nucleus Cytoplasmic proteins
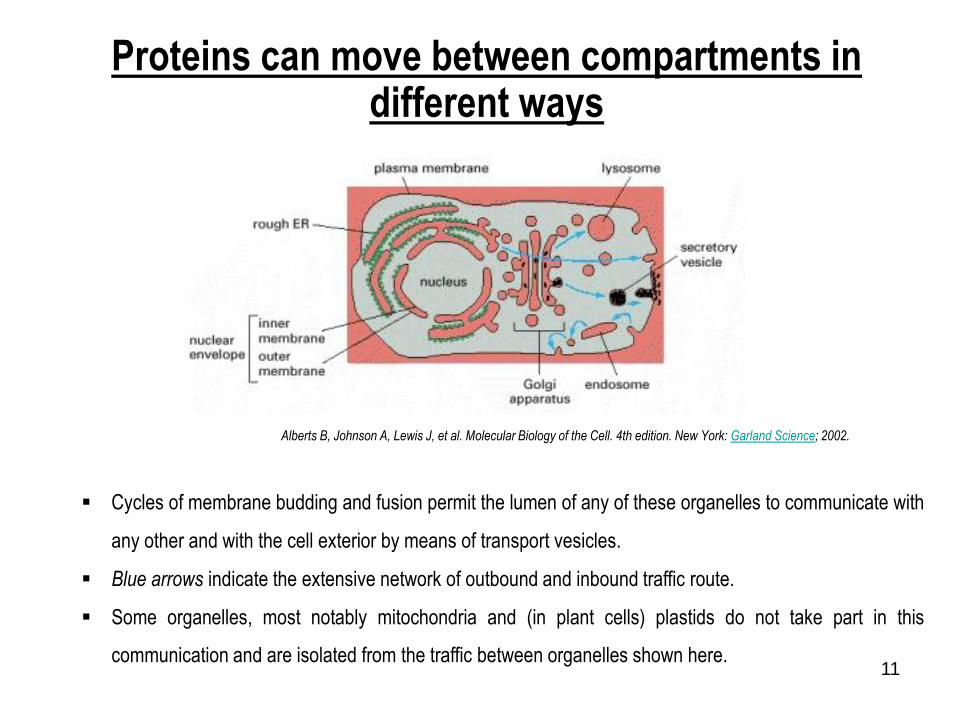
11
Proteins can move between compartments in different ways
Cycles of membrane budding and fusion permit the lumen of any of these organelles to communicate with
any other and with the cell exterior by means of transport vesicles.
Blue arrows indicate the extensive network of outbound and inbound traffic route.
Some organelles, most notably mitochondria and (in plant cells) plastids do not take part in this
communication and are isolated from the traffic between organelles shown here.
Alberts B, Johnson A, Lewis J, et al. Molecular Biology of the Cell. 4th edition. New York: Garland Science; 2002.

12
Three major ways of protein moving between different compartments
1. In gated transport, the protein traffic between the cytosol and nucleus occurs through
the nuclear pore complexes. The nuclear pore complexes function as selective gates
that actively transport specific macromolecules and macromolecular assemblies,
although they also allow free diffuse of smaller molecules.
2. In transmembrane transport, membrane-bound protein translocators directly transport
specific proteins across a membrane from the cytosol into a space that is topologically
distinct. The transported protein molecules usually must unfold to snake through the
translocator. The initial transport of selected proteins from the cytosol into the ER lumen
or from the cytosol into mitochondria, for example, occurs in this way.
3. In vesicular transport, membrane-enclosed transport intermediates—which may be
small, spherical transport vesicles or larger, irregularly shaped organelle fragments-ferry
protein from one compartment to another.
Alberts B, Johnson A, Lewis J, et al. Molecular Biology of the Cell. 4th edition. New York: Garland Science; 2002.

13
Signal sequence
Signal sequence is a short
sequence of amino acids that directed a
protein to it appropriate location in the
cell and, for many protein, is removed
during transport or after the protein has reached to its final destination.
1970

14
Albert B. et. al. Molecular Biology of the Cell, 4th ed, 2000
The Nobel Prize in Physiology or Medicine 1999
"for the discovery that proteins have intrinsic signals that govern their transport and localization in
the cell” PROTEIN ZIP CODES

15
Signal sequence directed to ER
The carboxyl terminus of the signal sequence is defined by a cleavage site, where protease
action removes the sequence after the protein is imported into the ER.
Signal sequences vary in length from 13 to 36 amino acid residues.
All signal sequences have the following features:
1) about 10 to 15 hydrophobic amino acid residues
2) one or more positively charged residues, usually near the amino terminus, preceding
the hydrophobic sequence
3) a short sequence at the carboxyl terminus (near the cleavage site) that is relatively
polar, typically having amino acid residues with short side chains (especially Ala) at
the positions closest to the cleavage site.

16
Examples of Signal sequences for protein translocation into ER
Nelson & Cox, Lehninger Principles of Biochemistry, 4th ed., 2005

17
Targeting of nuclear proteins
Nelson & Cox, Lehninger Principles of Biochemistry, 4th ed., 2005
18
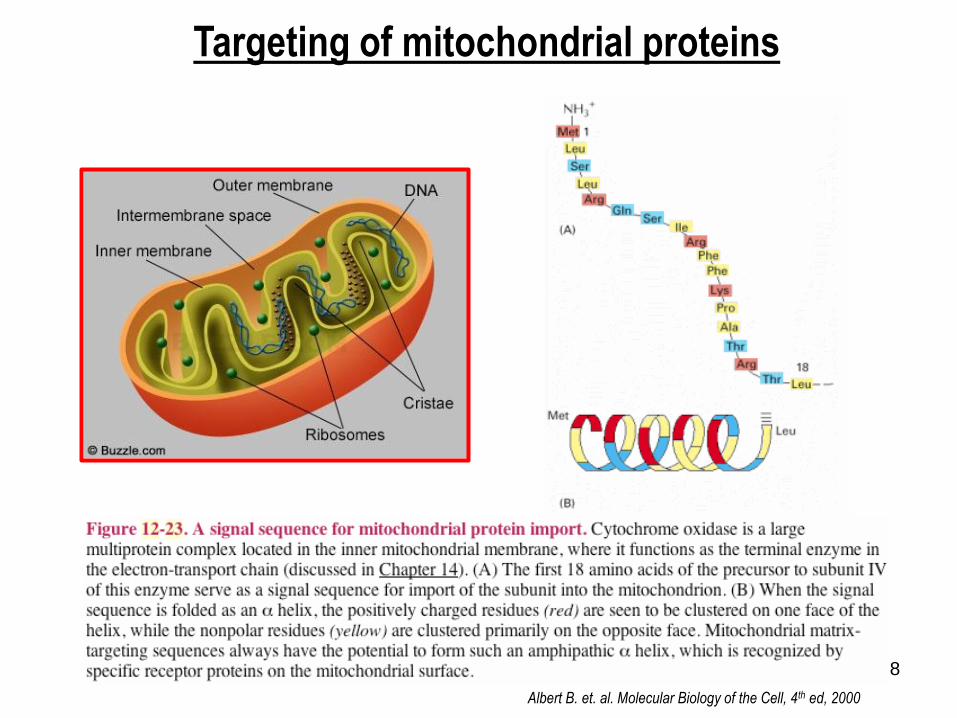
Targeting of mitochondrial proteins
Albert B. et. al. Molecular Biology of the Cell, 4th ed, 2000
19
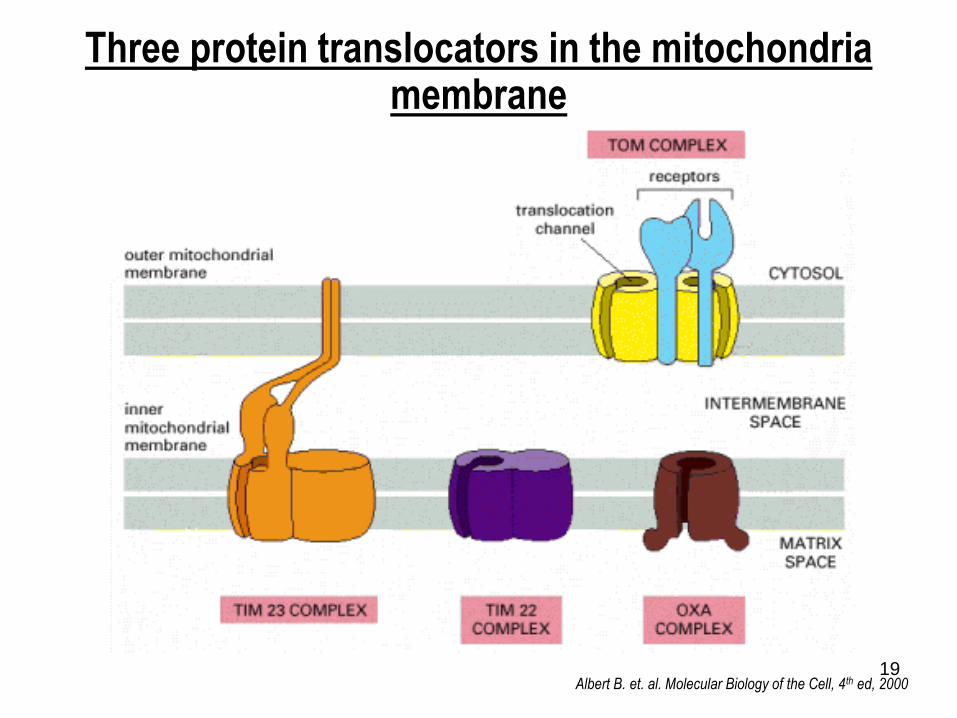
Three protein translocators in the mitochondria membrane
Albert B. et. al. Molecular Biology of the Cell, 4th ed, 2000
20
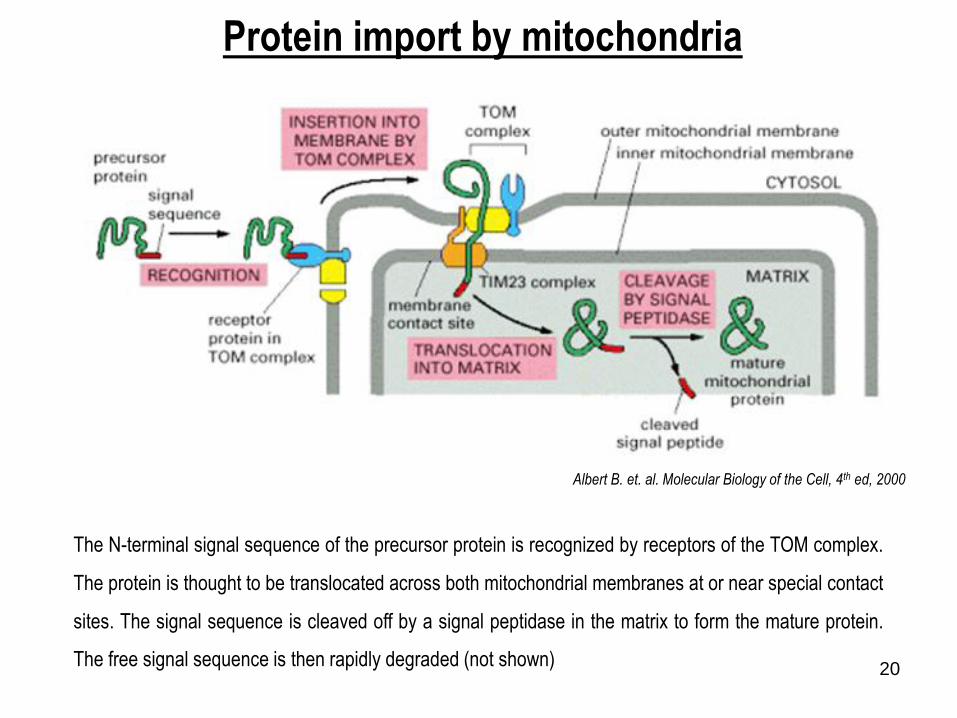
Albert B. et. al. Molecular Biology of the Cell, 4th ed, 2000
Protein import by mitochondria
The N-terminal signal sequence of the precursor protein is recognized by receptors of the TOM complex.
The protein is thought to be translocated across both mitochondrial membranes at or near special contact
sites. The signal sequence is cleaved off by a signal peptidase in the matrix to form the mature protein.
The free signal sequence is then rapidly degraded (not shown)

21
Albert B. et. al. Molecular Biology of the Cell, 4th ed, 2000
The role of energy in protein import into the mitochondrial matrix
Albert B. et. al. Molecular Biology of the Cell, 4th ed, 2000
22
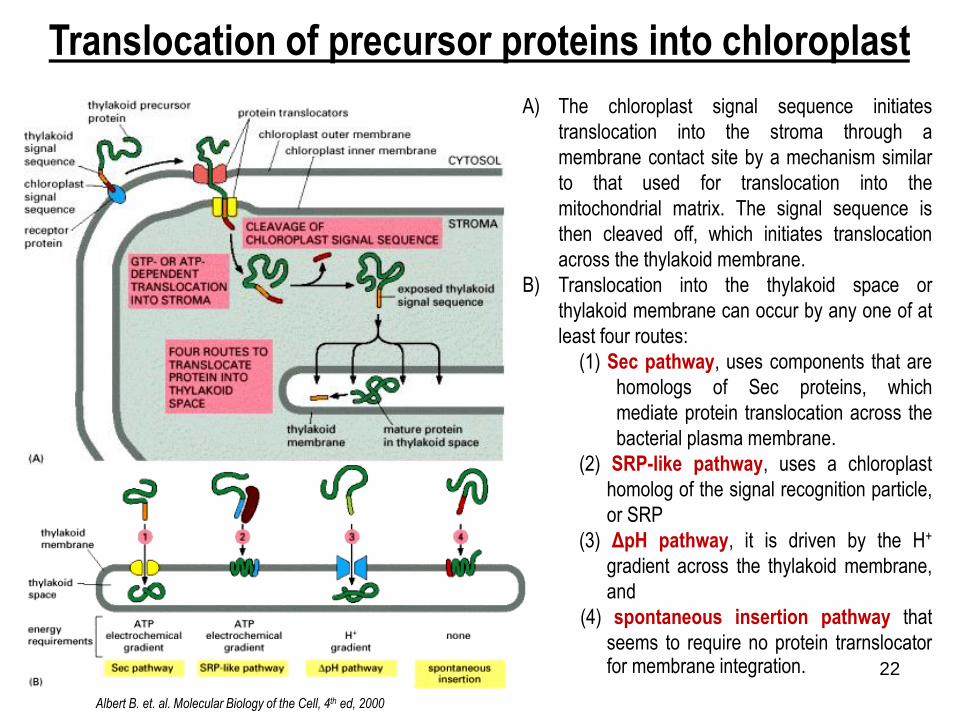
Translocation of precursor proteins into chloroplast
Albert B. et. al. Molecular Biology of the Cell, 4th ed, 2000
A) The chloroplast signal sequence initiates
translocation into the stroma through a
membrane contact site by a mechanism similar
to that used for translocation into the
mitochondrial matrix. The signal sequence is
then cleaved off, which initiates translocation
across the thylakoid membrane.
B) Translocation into the thylakoid space or
thylakoid membrane can occur by any one of at
least four routes:
(1) Sec pathway, uses components that are
homologs of Sec proteins, which
mediate protein translocation across the
bacterial plasma membrane.
(2) SRP-like pathway, uses a chloroplast
homolog of the signal recognition particle,
or SRP
(3) ΔpH pathway, it is driven by the H+
gradient across the thylakoid membrane,
and
(4) spontaneous insertion pathway that
seems to require no protein trarnslocatorfor membrane integration.
23
23
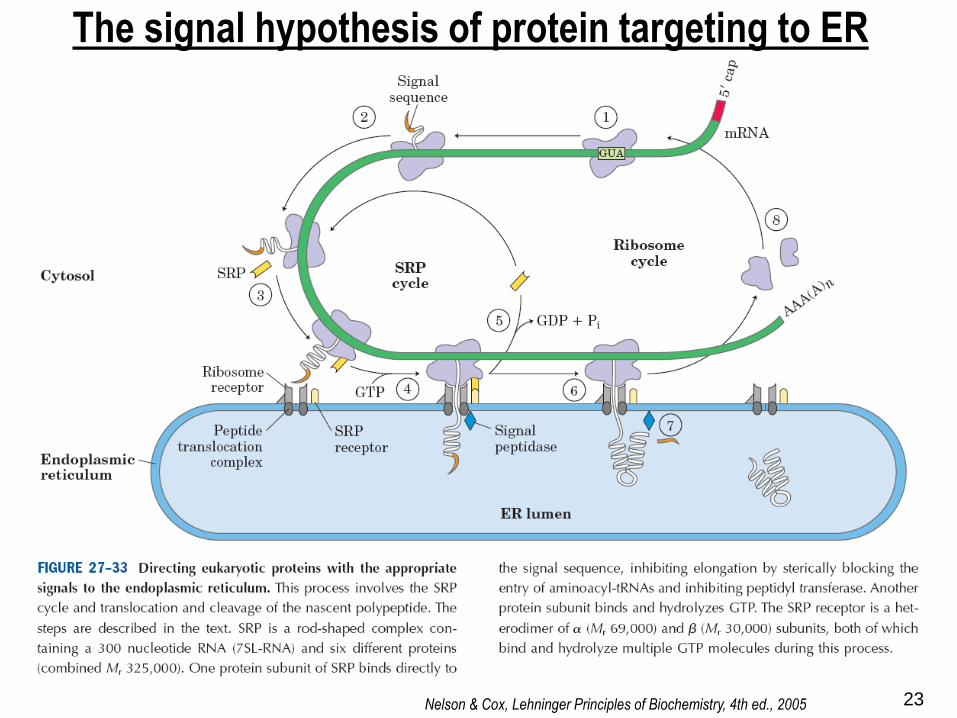
The signal hypothesis of protein targeting to ER
Nelson & Cox, Lehninger Principles of Biochemistry, 4th ed., 2005
24
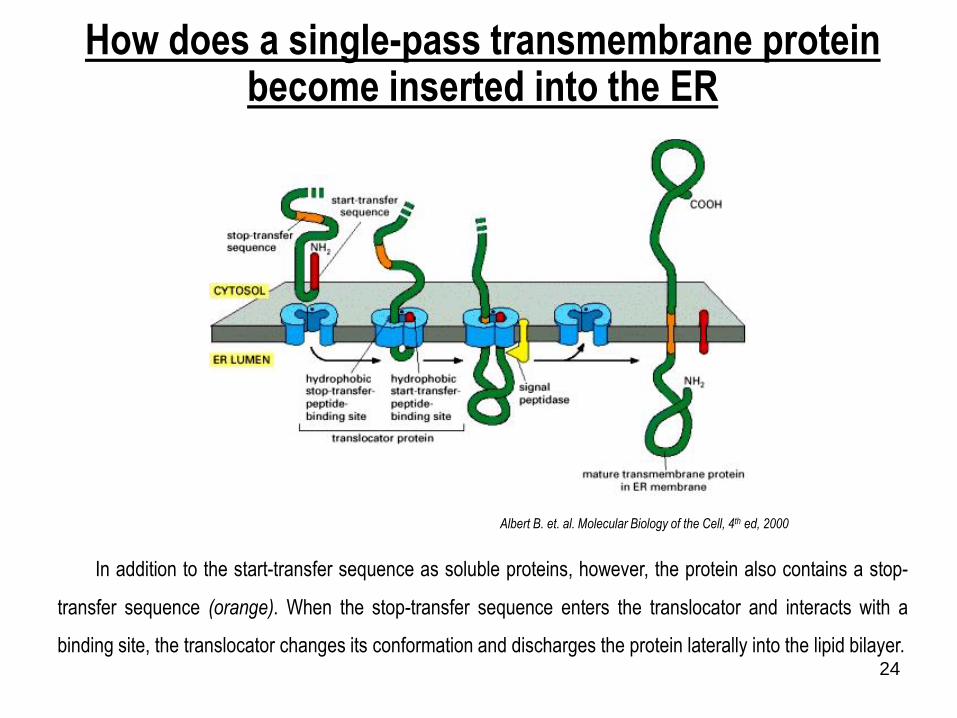
How does a single-pass transmembrane protein become inserted into the ER
In addition to the start-transfer sequence as soluble proteins, however, the protein also contains a stop-
transfer sequence (orange). When the stop-transfer sequence enters the translocator and interacts with a
binding site, the translocator changes its conformation and discharges the protein laterally into the lipid bilayer.
Albert B. et. al. Molecular Biology of the Cell, 4th ed, 2000
25
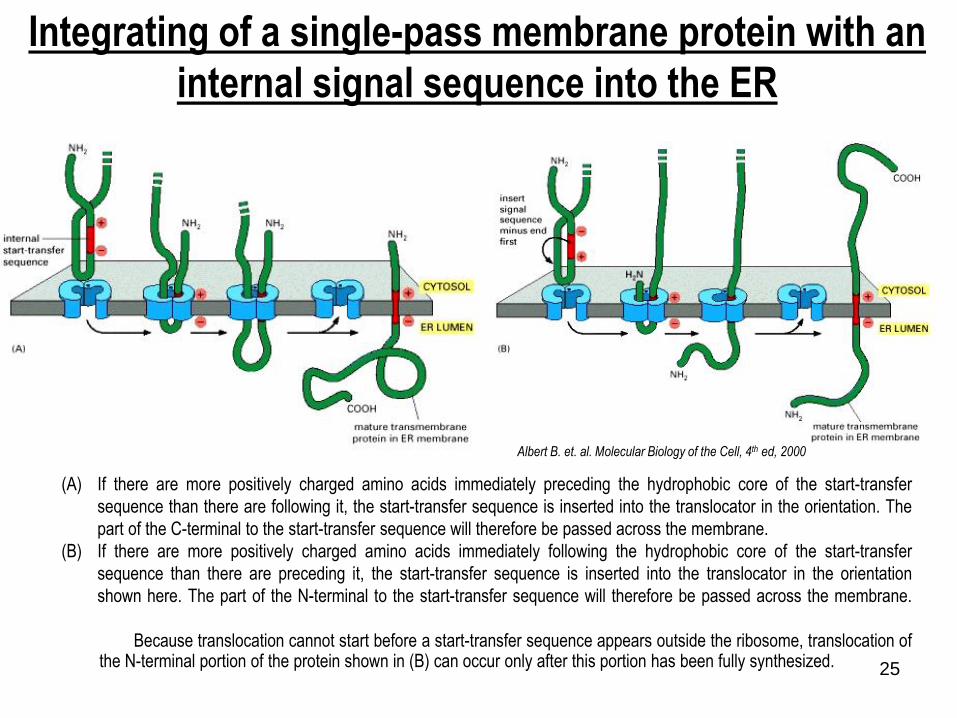
Integrating of a single-pass membrane protein with an
internal signal sequence into the ER
(A) If there are more positively charged amino acids immediately preceding the hydrophobic core of the start-transfer
sequence than there are following it, the start-transfer sequence is inserted into the translocator in the orientation. The
part of the C-terminal to the start-transfer sequence will therefore be passed across the membrane.
(B) If there are more positively charged amino acids immediately following the hydrophobic core of the start-transfer
sequence than there are preceding it, the start-transfer sequence is inserted into the translocator in the orientation
shown here. The part of the N-terminal to the start-transfer sequence will therefore be passed across the membrane.
Because translocation cannot start before a start-transfer sequence appears outside the ribosome, translocation ofthe N-terminal portion of the protein shown in (B) can occur only after this portion has been fully synthesized.
Albert B. et. al. Molecular Biology of the Cell, 4th ed, 2000
26
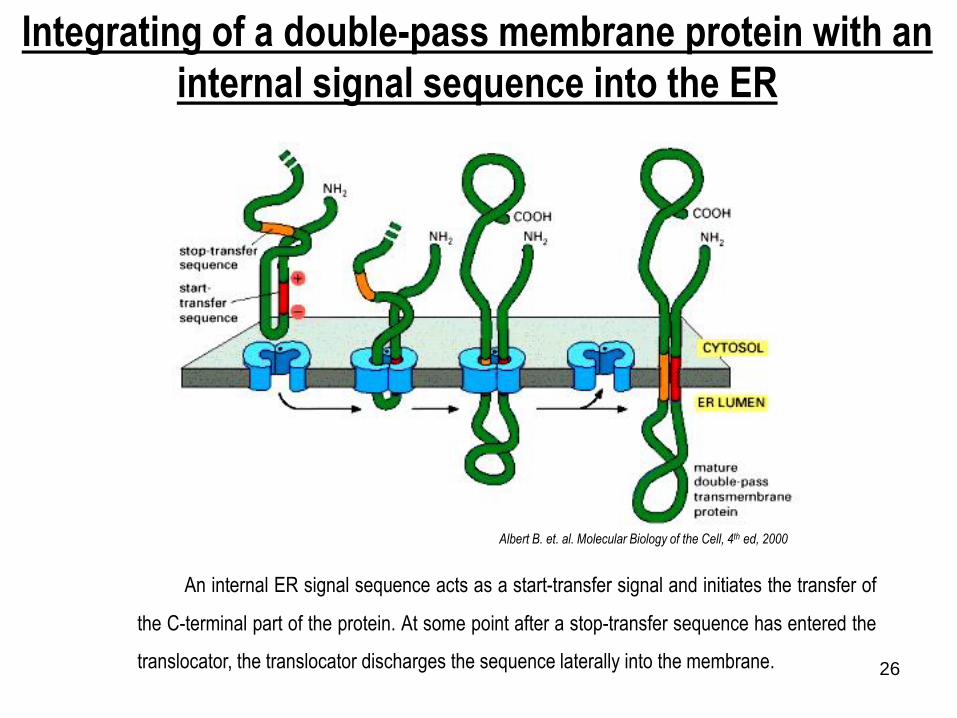
Integrating of a double-pass membrane protein with an
internal signal sequence into the ER
An internal ER signal sequence acts as a start-transfer signal and initiates the transfer of
the C-terminal part of the protein. At some point after a stop-transfer sequence has entered the
translocator, the translocator discharges the sequence laterally into the membrane.
Albert B. et. al. Molecular Biology of the Cell, 4th ed, 2000

27
The attachment of a GPI anchor to a protein in the ER
1) Immediately after the completion of protein synthesis, the precursor protein remains anchored in the ER membrane
by a hydrophobic C-terminal sequence of 15–20 amino acids; the rest of the protein is in the ER lumen.
2) Within less than a minute, an enzyme in the ER cuts the protein free from its membrane-bound C terminus and
simultaneously attaches the new C terminus to an amino group on a preassembled GPI intermediate.
3) The signal that specifies this modification is contained within the hydrophobic C-terminal sequence and a few amino
acids adjacent to it on the lumenal side of the ER membrane; if this signal is added to other proteins, they too
become modified in this way.
Because of the covalently linked lipid anchor, the protein remains membrane-bound, with all of its amino acidsexposed initially on the lumenal side of the ER and eventually on the cell exterior.
Albert B. et. al. Molecular Biology of the Cell, 4th ed, 2000

28
“What happen to the newly synthesized in the ER lumen?”
Removing of signal sequences
Folding of polypeptides
Forming of disulfide bond
Glycoprotein formation
Play a key role in protein targeting

2929
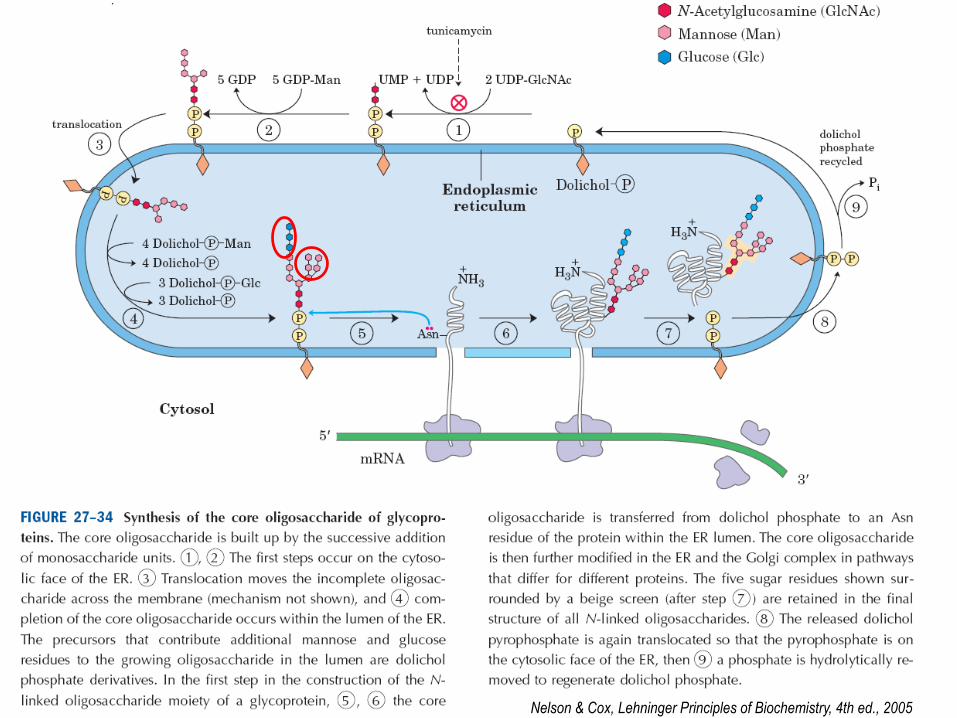
Glycosylation
“Adding of carbohydrate part into protein and there are 2 types of glycosylation”
1. O-linked GalNac 2. N-linked GlcNac
(Ser, Thr)
(Asn)
3030
Nelson & Cox, Lehninger Principles of Biochemistry, 4th ed., 2005

31
A.
B.
Nelson & Cox, Lehninger Principles of Biochemistry, 4th ed., 2005
3232
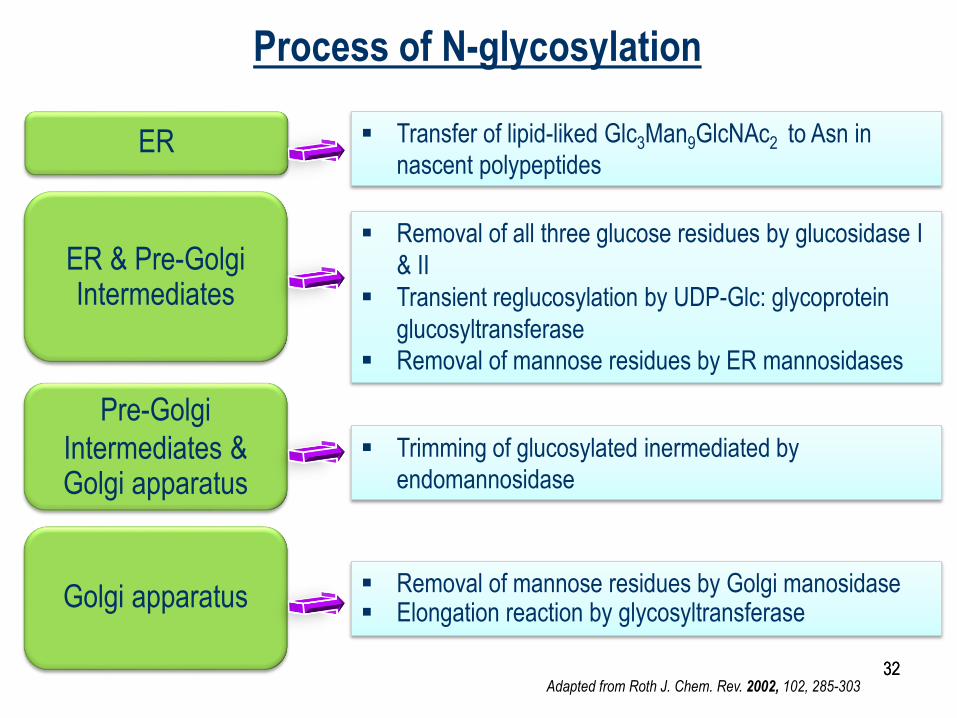
ER
ER & Pre-Golgi Intermediates
Pre-Golgi
Intermediates & Golgi apparatus
Golgi apparatus
Transfer of lipid-liked Glc3Man9GlcNAc2 to Asn in
nascent polypeptides
Removal of all three glucose residues by glucosidase I
& II
Transient reglucosylation by UDP-Glc: glycoprotein
glucosyltransferase
Removal of mannose residues by ER mannosidases
Trimming of glucosylated inermediated by
endomannosidase
Removal of mannose residues by Golgi manosidase Elongation reaction by glycosyltransferase
Adapted from Roth J. Chem. Rev. 2002, 102, 285-303
Process of N-glycosylation
33
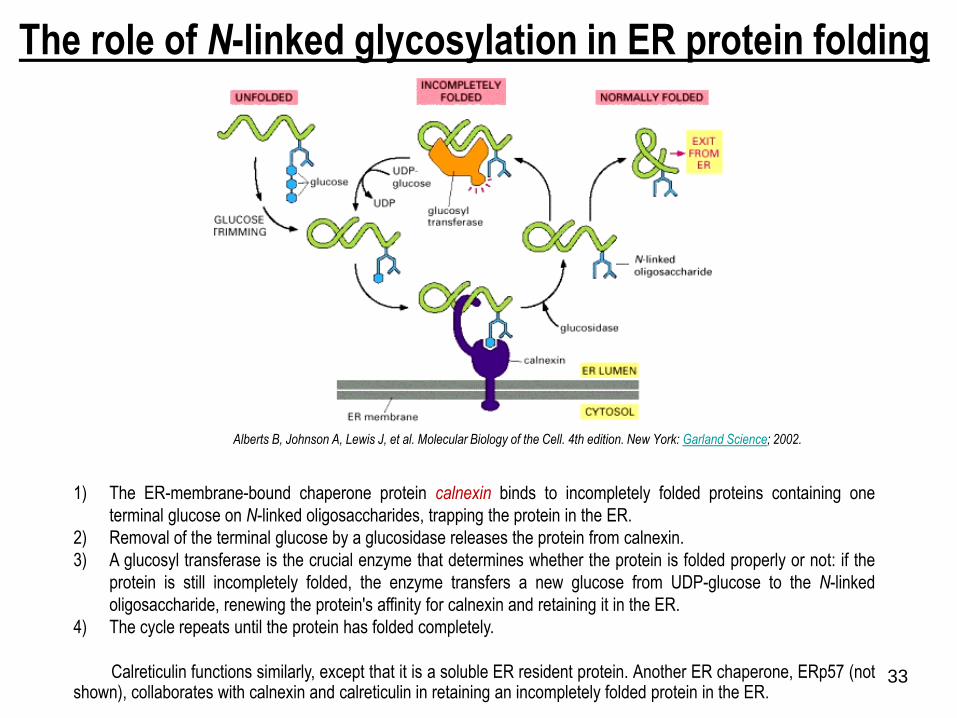
The role of N-linked glycosylation in ER protein folding
1) The ER-membrane-bound chaperone protein calnexin binds to incompletely folded proteins containing one
terminal glucose on N-linked oligosaccharides, trapping the protein in the ER.
2) Removal of the terminal glucose by a glucosidase releases the protein from calnexin.
3) A glucosyl transferase is the crucial enzyme that determines whether the protein is folded properly or not: if the
protein is still incompletely folded, the enzyme transfers a new glucose from UDP-glucose to the N-linked
oligosaccharide, renewing the protein's affinity for calnexin and retaining it in the ER.
4) The cycle repeats until the protein has folded completely.
Calreticulin functions similarly, except that it is a soluble ER resident protein. Another ER chaperone, ERp57 (notshown), collaborates with calnexin and calreticulin in retaining an incompletely folded protein in the ER.
Alberts B, Johnson A, Lewis J, et al. Molecular Biology of the Cell. 4th edition. New York: Garland Science; 2002.
34
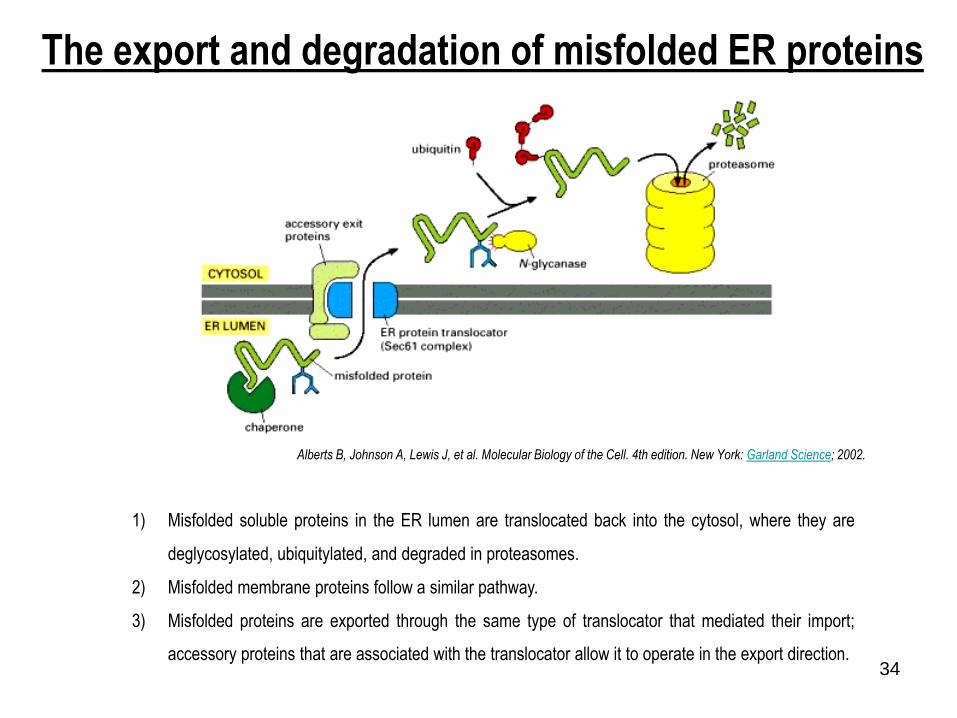
1) Misfolded soluble proteins in the ER lumen are translocated back into the cytosol, where they are
deglycosylated, ubiquitylated, and degraded in proteasomes.
2) Misfolded membrane proteins follow a similar pathway.
3) Misfolded proteins are exported through the same type of translocator that mediated their import;
accessory proteins that are associated with the translocator allow it to operate in the export direction.
The export and degradation of misfolded ER proteins
Alberts B, Johnson A, Lewis J, et al. Molecular Biology of the Cell. 4th edition. New York: Garland Science; 2002.

3535
O-glycosylation
No consensus sequence for Ser and Thr
Consensus for Hyl
Gly – X – Hyl – Y – Arg
Begins with GalNac transferase (N-
acetylgalactosamine)
Mannose common addition to core

36
Usually be extracellular proteins or located at cell surface
High structural information content
• molecular recognition
Occurs along the secretory pathway
Often stabilizes structure
Difficult to get crystal structure for more than one or two
carbohydrate residues
Notes for glycosylation

37
Utilization of different coats in vesicular traffic

38
The postulated role of SNAREs in guiding vesicular
transport
There are at least 20 different SNAREs in an animal cell, each
associated with a particular membrane-enclosed organelles
involved in the biosynthetic-secretory or endocytic pathway.
These transmembrane proteins exist as complementary sets
vesicle membrane SNAREs, called v-SNAREs
target membrane SNAREs, called t-SNAREs,
v-SNARESs and t-SNAREs have characteristic helical
domains.
When a v-SNARES interacts with a t-SNARE, the helical
domains of one wrap around the helical domains of the other
to form stable trans-SNARE complexes, which lock the two
membranes together.
The specificity with which SNAREs interact determines the
specificity of vesicle docking and fusion.
SNAREs specify compartment identity and govern the orderly
transfer of material during vesicle transport.
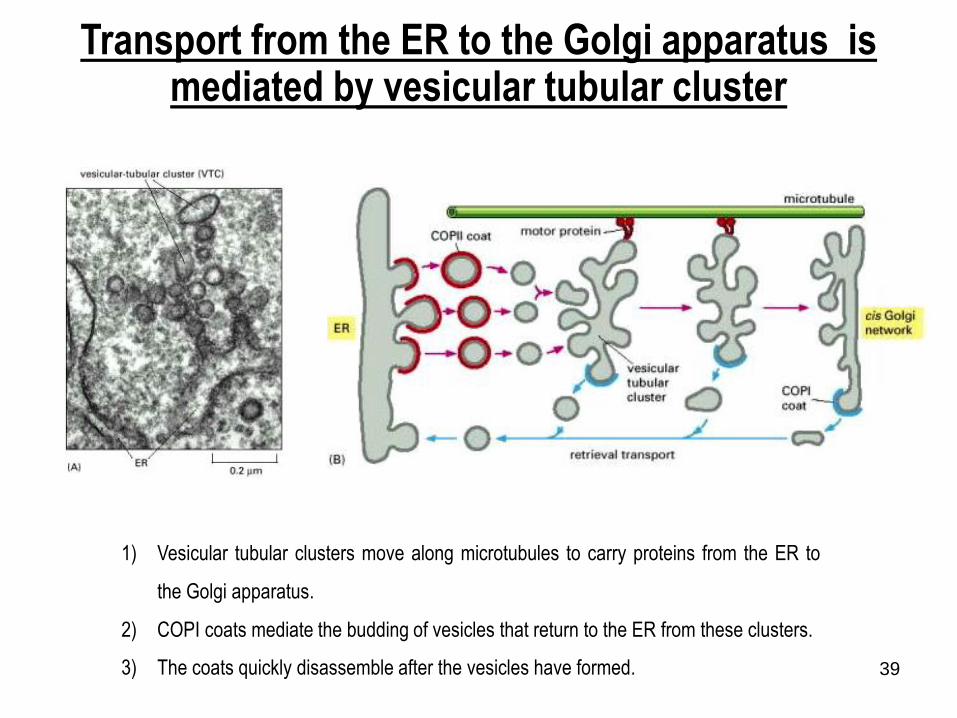
39
Transport from the ER to the Golgi apparatus is mediated by vesicular tubular cluster
1) Vesicular tubular clusters move along microtubules to carry proteins from the ER to
the Golgi apparatus.
2) COPI coats mediate the budding of vesicles that return to the ER from these clusters.
3) The coats quickly disassemble after the vesicles have formed.
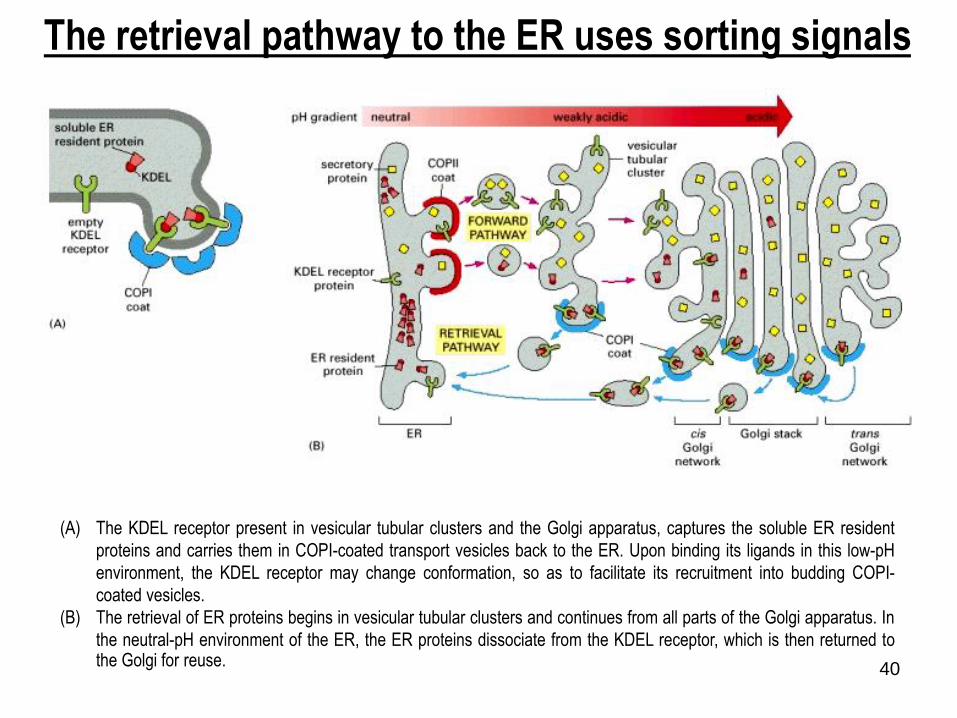
40
The retrieval pathway to the ER uses sorting signals
(A) The KDEL receptor present in vesicular tubular clusters and the Golgi apparatus, captures the soluble ER resident
proteins and carries them in COPI-coated transport vesicles back to the ER. Upon binding its ligands in this low-pH
environment, the KDEL receptor may change conformation, so as to facilitate its recruitment into budding COPI-
coated vesicles.
(B) The retrieval of ER proteins begins in vesicular tubular clusters and continues from all parts of the Golgi apparatus. In
the neutral-pH environment of the ER, the ER proteins dissociate from the KDEL receptor, which is then returned tothe Golgi for reuse.

4141

42
Alberts B, Johnson A, Lewis J, et al. Molecular Biology of the Cell. 4th edition. New York: Garland Science; 2002.
Oligosaccharide processing in the ER and the Golgi
apparatus
Processing begins in the ER with the removal of the glucoses from the oligosaccharide initially transferred to the protein. A mannosidase in the ER membrane
removes a specific mannose. Golgi mannosidase I removes three more mannoses and N-acetylglucosamine transferase I then adds an N-acetylglucosamine,
which enables mannosidase II to remove two additional mannoses. This yields the final core of three mannoses that is present in a complex oligosaccharide.
At this stage, the bond between the two N-acetylglucosamines in the core becomes resistant to attack by a highly specific endoglycosidase (Endo H). Since all
later structures in the pathway are also Endo H-resistant, treatment with this enzyme is widely used to distinguish complex from high-mannose
oligosaccharides. Additional N-acetylglucosamines, galactoses, and sialic acids are added. These final steps in the synthesis of a complex oligosaccharide
occur in the cisternal compartments of the Golgi apparatus. Three types of glycosyl transferase enzymes act sequentially, using sugar substrates that have
been activated by linkage to the indicated nucleotide. The membranes of the Golgi cisternae contain specific carrier proteins that allow each sugar nucleotide to enter in exchange for the nucleotide phosphates that are released after the sugar is attached to the protein on the lumenal face.

43
Albert B. et. al. Molecular Biology of the Cell, 4th ed, 2000
Related Documents











