Journal of Neurocytology 24, 478-486 (1995) Postsynaptic and extrasynaptic localization of K-opioid receptor in selected brain areas of young rat and chick using an anti-receptor monoclonal antibody KATALIN MADERSPACH 1., J. TAKACS 2, GRAZYNA NIEWIADOMSKA 2~ a n d A. CSILLAG 2 1BiologicaI Research Center, Hungarian Academy of Sciences, Temesv&i krt. 62, H-6701 Szeged, Hungary, and 21st Department of Anatomy, Semmelweis University Medical School, Tfizolt6 u. 58, H-1450 Budapest, Hungary Received 14 July 1994; revised 7 November 1994 and 1 February 1995; accepted 10 February 1995 Summary ~c-opioid receptors were visualized by light and electron microscopical immunohistochemistry in young rat and chick brains, using a monoclonal antibody KA8 (IgG1, K) raised against a lc-opioid receptor preparation from frog brain, which recognizes selectively the ~c-type receptor with preference for the lc-2 subtype. The most pronounced lc-opioid receptor-like immunoreactivity was observed in the hypothalamic nuclei of the rat brain and in the chick optic tectum, in regions where the functional significance of ~c-opioid receptors is well documented. Both neurons and glia were stained, the former on both somata and dendrites. At the ultrastructural level, the receptor-like immunoreactivity was similar in both species. Immunoprecipitate decorated the inner surface of the plasma membrane of glial cells, neuronal somata and dendrites, in a discontinuous arrangement. In the cytoplasm, labelling was associated with ribosomes, polyribosomes and rough endoplasmic reticulum membranes but not with Golgi cisternae. In the neuropil, the immunoprecipitate was observed along the dendritic microtubules and was also associated with postsynaptic sites. Nuclei and axons were devoid of label and immunoreactivity was never visible presynaptically. Our findings indicate that the antibody used in the present study marks various forms of the ~c-opioid receptor protein including those synthesised in ribosomes, transported along dendritic microtubules and incorporated into postsynaptic and non-synaptic membranes. The antibody also recognizes glial opioid receptors. The observed subcellular distribution appears to be conserved in phylogenetically distant species. Introduction Opioid receptors are heterogeneous with respect to ligand binding specificity and are classified as ~t, 6 and K types. This grouping, based primarily upon pharmacological experiments (Hughes, 1975; Martin et al., 1976; Zukin & Zukin, 1987; Goldstein, 1987; Goldstein & Naidu, 1989; Wollemann, 1993), was strongly supported by the recent molecular cloning results (Evans et al., 1992; Keiffer et al., 1992; Wang et al., 1993; Meng et al., 1993; Reisine et al., 1993; Mollereau et al., 1994). Several attempts have been made to determine the anatomical distribution of these receptor types in the brain tissue. Ligand autoradiographic studies have been done in avian (Csillag et al., 1989, 1990, 1993; Reiner et aI., 1989; Deviche et al., 1993) and mammalian (Blackburn et al., 1988; Mansour et al., 1988; Zukin et al., 1988; Slater et al., 1989; Maggi et al., 1989; Desjardins et al., 1990; Kujirai et al., 1991; Unterwald et al., 1991) species and in human brain (Blackburn et al., 1988; Vogt et al., 1990). These light microscopical findings indicated regional, species- and age-related differences. An inherent problem of the autoradiographic method is its low resolution, espe- cially at the ultrastructural level. Nevertheless, Hamel and Beaudet (1984) have localized ~t-opioid receptors electron-microscopically with a Met-enkephalin analo- gue in rat neostriatum and reported marked labelling on dendrites, axons and somata of neurons but rarely at synaptic junctions. Using the same radioligand, Beauvillain and colleagues (1992) have recently visualized ~t receptors on nerve endings and tanycytes in the median eminence of guinea-pig brain. K-opioid receptors have been mapped by electron microscopic autoradiography by Jomary and colleagues (1992). * To whom correspondenceshould be addressed. Present address: Department of Neurophysiology,Nencki Institute of Experimental Biology,3 Pasteur Street, 02-093 Warsaw, Poland. 0300-4864/95 1995 Chapman and Hall

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of Neurocytology 24, 478-486 (1995)
Postsynaptic and extrasynaptic localization of K-opioid receptor in selected brain areas of young rat and chick using an anti-receptor monoclonal antibody
K A T A L I N M A D E R S P A C H 1., J. T A K A C S 2, G R A Z Y N A N I E W I A D O M S K A 2~ and A . C S I L L A G 2
1BiologicaI Research Center, Hungarian Academy of Sciences, Temesv&i krt. 62, H-6701 Szeged, Hungary, and 21st Department of Anatomy, Semmelweis University Medical School, Tfizolt6 u. 58, H-1450 Budapest, Hungary
Received 14 July 1994; revised 7 November 1994 and 1 February 1995; accepted 10 February 1995
Summary ~c-opioid receptors were visualized by light and electron microscopical immunohistochemistry in young rat and chick brains, using a monoclonal antibody KA8 (IgG1, K) raised against a lc-opioid receptor preparation from frog brain, which recognizes selectively the ~c-type receptor with preference for the lc-2 subtype. The most pronounced lc-opioid receptor-like immunoreactivity was observed in the hypothalamic nuclei of the rat brain and in the chick optic tectum, in regions where the functional significance of ~c-opioid receptors is well documented. Both neurons and glia were stained, the former on both somata and dendrites. At the ultrastructural level, the receptor-like immunoreactivity was similar in both species. Immunoprecipitate decorated the inner surface of the plasma membrane of glial cells, neuronal somata and dendrites, in a discontinuous arrangement. In the cytoplasm, labelling was associated with ribosomes, polyribosomes and rough endoplasmic reticulum membranes but not with Golgi cisternae. In the neuropil, the immunoprecipitate was observed along the dendritic microtubules and was also associated with postsynaptic sites. Nuclei and axons were devoid of label and immunoreactivity was never visible presynaptically. Our findings indicate that the antibody used in the present study marks various forms of the ~c-opioid receptor protein including those synthesised in ribosomes, transported along dendritic microtubules and incorporated into postsynaptic and non-synaptic membranes. The antibody also recognizes glial opioid receptors. The observed subcellular distribution appears to be conserved in phylogenetically distant species.
Introduction
Opioid receptors are heterogeneous with respect to ligand binding specificity and are classified as ~t, 6 and K types. This grouping, based primarily upon pharmacological experiments (Hughes, 1975; Martin et al., 1976; Zukin & Zukin, 1987; Goldstein, 1987; Goldstein & Naidu, 1989; Wollemann, 1993), was strongly supported by the recent molecular cloning results (Evans et al., 1992; Keiffer et al., 1992; Wang et al., 1993; Meng et al., 1993; Reisine et al., 1993; Mollereau et al., 1994).
Several attempts have been made to determine the anatomical distribution of these receptor types in the brain tissue. Ligand autoradiographic studies have been done in avian (Csillag et al., 1989, 1990, 1993; Reiner et aI., 1989; Deviche et al., 1993) and mammalian (Blackburn et al., 1988; Mansour et al., 1988; Zukin et al., 1988; Slater et al., 1989; Maggi et al.,
1989; Desjardins et al., 1990; Kujirai et al., 1991; Unterwald et al., 1991) species and in human brain (Blackburn et al., 1988; Vogt et al., 1990). These light microscopical findings indicated regional, species- and age-related differences. An inherent problem of the autoradiographic method is its low resolution, espe- cially at the ultrastructural level. Nevertheless, Hamel and Beaudet (1984) have localized ~t-opioid receptors electron-microscopically with a Met-enkephalin analo- gue in rat neostriatum and reported marked labelling on dendrites, axons and somata of neurons but rarely at synaptic junctions. Using the same radioligand, Beauvillain and colleagues (1992) have recently visualized ~t receptors on nerve endings and tanycytes in the median eminence of guinea-pig brain. K-opioid receptors have been mapped by electron microscopic autoradiography by Jomary and colleagues (1992).
* To whom correspondence should be addressed. Present address: Department of Neurophysiology, Nencki Institute of Experimental Biology, 3 Pasteur Street, 02-093 Warsaw, Poland.
0300-4864/95 �9 1995 Chapman and Hall

n-opioid receptor in rat and chick brains
Working with a radiolabel!ed dynorphine analogue in the guinea-pig neostriatum, labelling was detected at membrane locations partly associated with synaptic specializations.
An alternative approach, the application of specific antibodies, has proved particularly favourable for subcellular receptor localization. Concerning opioid receptors, only a few reports are known. Myers and Glasel (1987) raised an anti-morphine (anti-idiotypic) antibody with g- and some 8-opioid specificity and visualized receptors on NG 108-15 cell membranes (Ornatovska & Glasel, 1991). Hassan and colleagues (1989) presented immunohistochemical results with an anti-[3-endorphin antibody (Gramsch et al., 1988) which recognizes g and 6, but no ~:, receptors. They found labelling in the somata and the proximal dendrites of neurons of the hypothalamus (preoptic nuclei and the ventromedial area) and striatum, and they suggested also the intracellular presence of the immunoreactivity.
Visualization of r receptors by light and electron microscopical immunohistochemistry was first successful in our laboratory using the monoclo- nal anti-receptor antibody KA8 (IgG1, ~:), (Maderspach et al., 1991). In primary cell cultures of embryonic chick brain, a polarized arrangement of the K-opioid receptor-like immunoreactivity (KLRI) was found in the somata of young neurons, and this labelling spread to the processes during differentiation (Maderspach & N6meth, 1993). Using the same antibody in the human brain cortex (Schmidt et al., 1994), we detected immunoprecipitate in pyramidal neurons of cortical layers II/III and V, and in ovoidal neurons of layer VI. At the ultrastructural level, the plasma membrane, free ribosomes and dendritic microtubules were immunoreactive. Some astrocytes were also labelled. The ultrastructural appearance of opioid receptors is expected to show common features across the different species and these characteristics may not be dissimilar from other receptors (Aoki et al., 1989; Naas et al., 1991; Schr6der, 1992).
The primary aim of the present study was to demonstrate the subcellular localization of the ~c- opioid receptor in different brain regions of phylo- genetically distant species. We chose to use young animals because the K-opioid signals through specific receptors are supposed to regulate neuronal and glial differentiation during ontogeny (Schmahl et al., 1989; Stiene-Martin et al., 1991; Maderspach & N6meth, 1993). A systematic comparison between species, age or brain regions was not intended.
479
young (30-35 days old, weighing about 100 g) female Wistar rats (n=3) were deeply anaesthetised with sodium pentobarbital and a cannula was introduced into the ascending aorta via the left ventrical of the heart. The animals were perfused with 20-25 ml of cold isotonic saline followed by 100 ml of freshly prepared Zamboni's fixative. In the case of chickens, the perfusion solution did not contain glutaraldehyde, whereas in the case of rats, 0.1% glutaral- dehyde was added to the fixative. The perfusion of rats was completed with 200ml of Zamboni's fixative without glutaraldehyde, at room temperature. The brains were removed from the skull and postfixed in Zamboni's fixative (without glutaraldehyde) for 34 h at 4 ~ C. From the brains 5- 6 mm thick blocks were cut in the horizontal plane and placed in 0.05M phosphate-buffered saline (PBS, pH 7.4) containing 20% sucrose (at 4 ~ C, overnight). The tissue blocks were frozen in iso-pentane (at -50 ~ C) and thawed to room temperature (repeated twice). After rinsing in PBS, 60-70 ~tm thick sections were cut from the blocks using a Vibratome (Oxford Instruments). The sections were collected in cold PBS and washed in the same buffer for 12-16h at 4 ~ C, changing the buffer at least six times.
IMMUNOHISTOCHEMISTRY
Tissue slices were immunolabelled with the antibody produced by the KA8 mouse hybridoma cell line (Mader- spach et al., 1991). Two alternative methods were used. In some cases, slices were incubated with the supernatant of KA8 hybridoma cell line diluted at 1 : 2-1 : 4 in PBS contain- ing 10% BSA, while, in other experiments, the ascitic fluid harvested from Balb/C mice (which had been injected i.p. with the same cell line) was applied, diluted 1:50 in the above buffer. To suppress non-specific immunoreactivity, slices were preincubated in PBS containing 10% BSA at room temperature for 90min. In order further to decrease the staining background, working dilutions of the antibody were pretreated with rat brain acetone powder (Arnel Co. Inc.) prior to application. 0.1 g brain powder was mixed with 1.0 ml of the diluted antibody and stirred gently for 1 h at room temperature; centrifuged at 1200g and filtered on Millipore filter (pore size 0.45 gin). Sections were incubated with the primary antibody for 24-36 h at 4-8 ~ C, then they were exposed to biotinylated goat-antimouse IgG (Vector) diluted I : 100, for 2 h, followed by treatment with peroxidase- labelled avidin-biotin complex (Vector ABC), diluted 1:100 for 90 rain. The immunocomplex was visualized using 0.05% 3-3'diamino-benzidine tetrahydrochloride (DAB; Sigma) in the presence of 0.01% H202 in Tris-HC1 (pH 7.6) at room temperature. Tissue slices for EM investigations were postfixed in 1% OsO4 (pH 7.3 in 0.1 MNa-phosphate buffer) for l h, dehydrated in a graded series of ethanol and propylene oxide, stained with 1% uranyl-acetate in 70% ethanol and embedded in Durcupan ACM (Fluka). Ultrathin sections were counterstained with lead citrate and photo- graphed in a JEOL 100-B electron microscope.
Materials and methods
PREPARATION OF SECTIONS
Two-day-old domestic chicks (Hunnia broilers n = 3) and
NOTES ON THE SPECIFICITY
Immunocytochemical procedure
In every run of immunostaining, parallel tissue sections were incubated in the absence of the primary antibody, but

480 M A D E R S P A C H , TAK,~CS, N I E W I A D O M S K A andCSILLAG
otherwise according to the above described protocol. Under these conditions, no immunoreactivity was detected.
Primary antibody The mAb KA8 was raised against the partially purified ~c- opioid receptor protein of frog brain as antigen, by the hybridoma technique. The antibody was found to be highly selective to the K-opioid receptor type on the basis of ligand binding experiments; moreover it has preference for the lc-2 receptor subtype (Maderspach et al., 1994). Since no published cloned sequences have been claimed to represent the K-2 subtype, preabsorption with the purified receptor protein, as a test of specificity, was considered premature at this stage.
Results
The mAb KA8 immunostaining of sections was feasible both with hybridoma supernatant and ascitic fluid. The advantage of supernatant application is the lower background. In the case of rat brain sections, the hybridoma supernatant resulted in convincing specific staining. Conversely, in the chick brain this form of antibody produced only weak labelling. Therefore, in this species, we had to opt for the ascitic fluid, containing the antibody in higher concentration. Both approaches led to comparable results.
LIGHT MICROSCOPIC OBSERVATIONS IN THE RAT BRAIN
The most marked lc-opioid receptor-like immuno- reactivity (ICRLI) appeared in the hypothalamic area
but labelling was detected also in other diencephalic regions, in the cerebral cortex and striatum. The somata of the nerve cells and, occasionally, the primary thick dendritic branches were stained (Figs 1-2). The immunoprecipitate gave a granular impres- sion in the cytoplasm, while the nuclei remained unstained. Occasionally, the immunoreactive astro- cytes were also visible (not shown).
ELECTRON MICROSCOPIC OBSERVATIONS
Rat brain
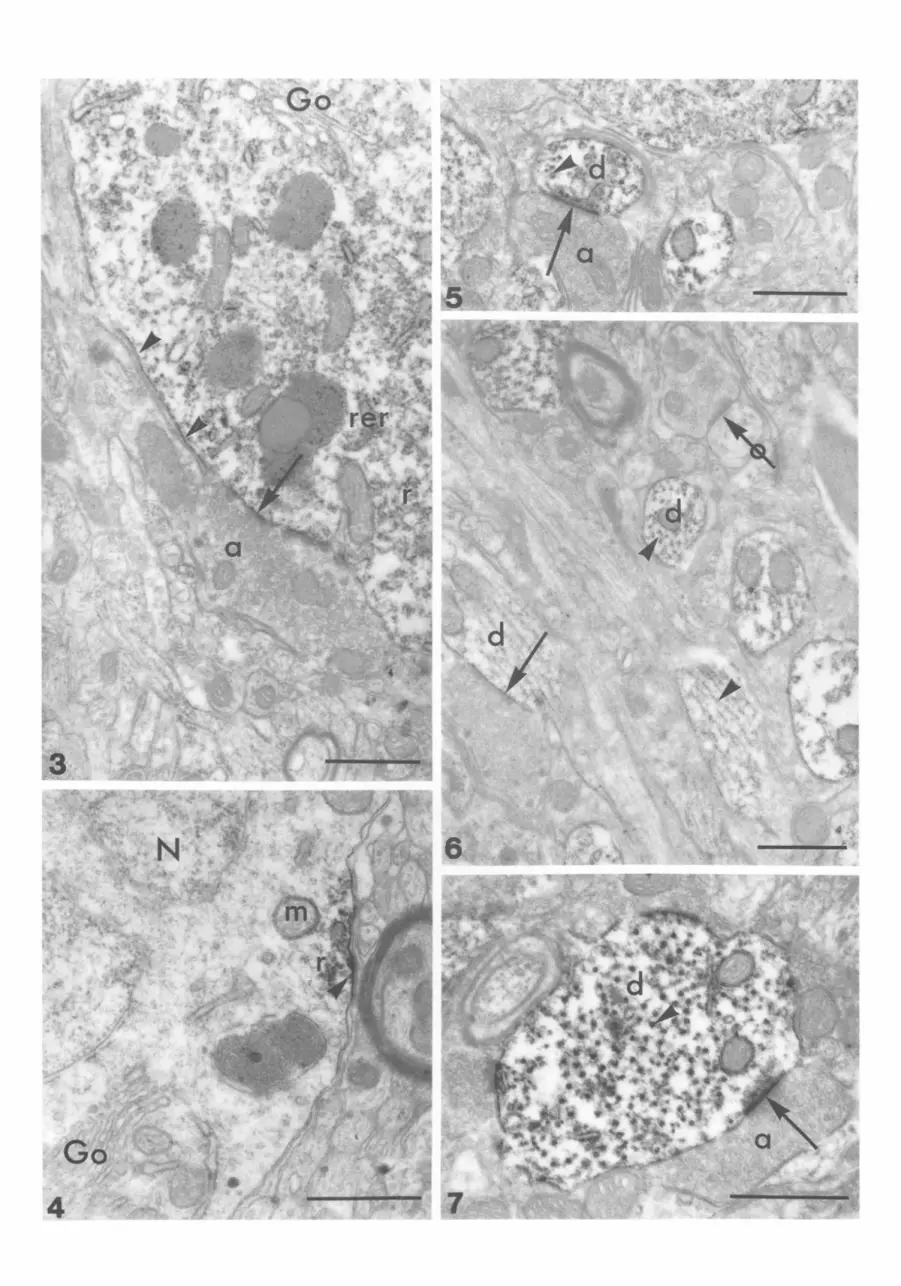
We focused EM investigations on the ventromedial hypothalamic nucleus, where the most conspicuous labelling had been found by LM. In most neuronal cell bodies, KRLI was visible in the cytoplasm. In particular, free ribosomes, polyribosomes and ribo- somes on the rough endoplasmic reticulum (RER) membrane were labelled (Figs 3-4). Fine granular deposits of immunoperoxidase endproduct in discontinuous arrangement were frequently observed along the neuronal plasma membrane (Figs 3-4). These deposits were associated either with axo- somatic synaptic appositions (Fig. 3) or they formed a dense undercoating attached closely to the inner surface of the plasma membrane apparently not facing synaptic sites (Figs 3-4). This undercoating was also seen in the vicinity of immunolabelled ribosomes (Fig. 4). In some cases, faint immunoprecipitate appeared to delineate the outer membranes of
Fig. 1. Coronal section of a rat hypothalamus. KLRI dominant in the nucleus paraventricularis (outlined by arrowheads). Scale bar = 350 ~tm. Fig. 2. Coronal section of rat hypothalamus from a region similar to that marked by arrowheads in Fig. 1. Neuronal cell bodies (arrows) are immunoreactive. The cells lining the IIIrd ventricle are also stained (ringed arrows). A labelled dendritic segment is also visible (arrowhead, insert). Scale bar = 75 ~tm.

~:-opioid receptor in rat and chick brains
mitochondria (Fig. 4). Nuclei and the cisternae of Golgi apparatus were devoid of labels (Figs 3-4).
In the neuropil, KLRI was observed in dendritic profiles (Figs 5~) . Most pronounced sites of labelling were the postsynaptic densities and microtubules (Figs 5, 7). Immunonegative synaptic junctions were also observed within densely immunoreactive areas (Fig. 6). KRLI adjacent to microtubules was often abundant, seemingly 'enlarging' the cross-sectional profiles (Fig. 7). Fine granular labelling was also observed on short segments of RER membranes and on the outer membranes of mitochondria (Fig. 7).
Chick brain
Specimens for EM were obtained from deep layers of the optic tectum. In addition to neuronal labelling, we observed some immunopositive glial cells. In glial perikarya, similarly to neuronal KRLI, polyribosomal clusters and membrane appositions of various length on the inner surface of the plasma membrane were immunoreactive (Figs 8, 9). Labelled polyribosomes and membrane appositions frequently showed close association (Fig. 9). In some glial cells, the juxtanuclear segments of RER also contained immunoprecipitate (Fig. 9). Other cytoplasmic organelles and cell nuclei were devoid of label.
The KRLI in the neuropil was essentially similar to the pattern found in the rat. Labelling was confined to dendritic profiles, mainly on and around microtubules, or in the form of submembraneous appositions (Figs 10, 11). However, the latter form of labelling was not restricted to sy~aptic sites (Fig. 11).
Discussion
The review of Mansour and colleagues (1988) summarized LM autoradiographic observations rele- vant to the mapping of opioid receptor types in the rat brain. The general conclusion from these data was that
481
all three types are abundant and overlapping in the different brain areas except the hypothalamic nuclei, where the r type opioid receptors predominate. By contrast, Desjardins and colleagues (1990) described ~t-, 6- and •-opioid receptors in the rat hypothalamus with different but essentially overlapping distribu- tions. These findings were supported recently by Allen and colleagues (1993). Several other authors have published autoradiographic mapping studies on K-opioid receptors and their subtypes in rat and guinea pig brains and found that most of the brain areas with the heaviest labelling belonged to the limbic system (Zukin et al., 1988; Slater et al., 1989; Unterwald et ai., 1991). This corresponds to the known ~c-opioid-related functions (see review by Wollemann et al., 1993). The K-opioid receptors proved to be abundant in the human brain (for reference see Schmidt et al., 1994) and also in avian species, as evident from autoradiographic mapping (Mansour et al., 1988). The presence of this receptor type has been confirmed in the optic tectum of pigeon (Reiner et al., 1989) and chicken (Csillag et al., 1990).
The present study offers the first cellular and subcellular immunolocalization of K-opioid receptors in rat and chick brains. The regional density of cells with KRLI, as evident from the LM results, appears rather low as compared to the previously listed autoradiographic data. The difference could be due, on the one hand, to an essentially smaller component of non-specific labelling. On the other hand, the mAb KA8 has high selectivity, does not cross-react with ~t or 6 receptors, and displays preference for the K-2- subtype of opioid receptors (Maderspach et al., 1991). The most pronounced neuronal labelling appeared in distinct areas of the hypothalamic nuclei of the rat brain and in the deep layers of chick optic tectum. Neuronal somata and dendrites were immunoposi- tive. Astroglial immunoreactivity was also detectable in both species, furnishing anatomical evidence for
Figs 3-7. Electron micrographs of rat hypothalamus. Fig. 3. Detail of an immunoreactive neuron. Discontinuous fine immunoprecipitate is visible along the plasma-membrane (arrowheads) often opposite to axon (a) terminals (arrow). The rough endoplasmic reticulum membranes (rer), free ribosomes and polyribosomes (r) are also stained while no labelling is observed in the Golgi apparatus (Go). Scale bar = 0.5 Ixm.
Fig. 4. Immunoreactive membrane area (arrowhead) of a neuron with subjacent labelled polyribosomes (r). Immunoprecipi- rate is associated occasionally with the outer surface of mitochondria (m). Nucleus (N) and Golgi apparatus (Go) are devoid of label. Scale bar = 0.5 ~tm. Fig. 5. Neuropil of the rat hypothalamus. KRLI in fine dendritic branches (d) is accumulated at post-synaptic membrane specializations (arrows) in contact with axon terminals (a). Cross-sections of microtubules are also labelled (arrowhead). Scale bar = 0.5 ~m. Fig. 6. Longitudinal and cross-sections of immunolabelled dendritic profiles (d) with microtubules (arrowheads). One finely labelled sub-synaptic apposition is marked by an arrow, whereas an immunonegative axodendritic synapse is indicated by a ringed arrow. Scale bar = 0.5 ~tm. Fig. 7. Cross-section of a dendrite (d) at higher magnification. Immunoreactive microtubules (arrowhead) and discrete membrane areas as well as labelled post-synaptic membrane specializations of axo-dendritic synapses (arrow) are visible. Scale bar = 0.5 Ixm.


484 M A D E R S P A C H , TAKACS, N I E W I A D O M S K A andCSILLAG
the presence of ~:-opioid receptors on mature glial cells, as suggested by earlier biochemical findings (Maderspach & Solomonia, 1988; Stiene-Martin et al., 1991, Hauser & Stiene-Martin; 1993).
The subcellular distribution of KRLI proved to be remarkably similar in the two species studied here and also in human specimens (Schmidt et al., 1994). A discontinuous arrangement of immunoprecipitate was visible along the inner surface of the plasma membrane of neuronal and glial cells. In some cases, this labelling in the neurons coincided with postsynaptic appositions; in other cases, no synaptic specialization was detectable at the sites of membrane-associated immunoreactivity. The extra- synaptic appositions in the neurons may represent past or future synaptic sites generated in the dynamically changing membrane. Alternatively, these immunoreactive sites, just like those on glial cells, may corrrespond to genuine non-synaptic receptors responding also to endogenous opiates released from neurons and perhaps also from glial cells (Zagon et aI., 1990; Stiene-Martin et al., 1991; Hauser & Stiene-Martin, 1993). This could be of special importance in the differentiating nervous system where synaptic specializations are not yet developed (Hauser & Stiene-Martin, 1993; Maderspach & N6meth, 1993).
The receptor-like immunoreactivity was often seen at postsynaptic axo-somatic or axo-dendritic sites but it was never observed presynaptically, in axons or axon terminals. Anatomical and physiological data evenly support presynaptic and postsynaptic localization of ~:-opioid receptors. A postsynaptic localization was suggested by Inenaga and colleagues (1994) who described a decrease in postsynaptic potentials caused by K selective agonists in the rat hypothalamus. Goldsmith and colleagues (1991) observed opioid peptides in rat hypothalamic para- ventricular and supraoptic nuclei presynaptically, while Fodor and colleagues (1990) found both presynaptic and postsynaptic dynorphin-A contain- ing neural elements in the rat nucleus solitarius. Mulder and colleagues (1984) as well as de Vries and colleagues (1990) reported a presynaptic modulation
of neurotransmission by ~-opioids. These results would imply that ~c-opioid receptors may function both pre- and postsynaptically. However, a fascinating novel interpretation appears from a recent study (Drake et al., 1994). Based on postsynaptically detected dynorphin immunolabelling and electro- physiological results in guinea-pig hippocampus, the authors suggested that the retrograde inhibition of neurotransmission was mediated by presynaptic opioid receptors of the ~:-1 subtype. Since the findings of the present study revealed a postsynaptic localiza- tion of KRLI, obtained by the K-2 preferring mAB KA8, we can hypothesize that the ~:-2 receptor subtype may be postsynaptic, as opposed to presynaptic localiza- tion of K-1 receptors. This hypothesis, however, needs further experimental support.
EM revealed KRLI in the cytoplasm of both glial cells and neuronal cell bodies and dendrites associated with ribosomes and polyribosomes. This may indicate that the antibody also recognizes an early synthetic form of the receptor protein. We did not find Golgi cisternae to be labelled, although post-translational modification of this receptor protein, especially by glycosylation, is very likely. Marked labelling was often seen along dendritic microtubules. This micro- tubular localization may be associated with the transport of the receptor protein. The labelled poly- ribosomes in close proximity to immunoreactive membrane domains may represent a receptor pool just being incorporated into the plasma membrane and, perhaps, providing a deposit for rapid receptor turnover.
These findings resemble the observations on other membrane receptors, such as nicotinic and muscarinic acetylcholine (Schr6der, 1992), GABA (Somogyi et al., 1989), ~-adrenergic (Aoki et al.,, 1989) or dopamine (Level et al., 1993) receptors. This is probably reflecting the similar fate of various receptor proteins inside the cell, including synthesis, transport and incorporation into the membrane at functional sites. However, certain differences are also evident, e.g. plasma membrane-associated glycine receptors have been found exclusively (Triller et al., 1985) or predomi- nantly (Naas et al., 1991) at synaptic sites, and we were
Figs 8-11. Electron micrographs of chick optic tectum. Fig. 8. Immunoreactivity in a glial cell along the inner surface of the plasma membrane (arrowhead) and on the polyribosomes (pr) in close contact with the immunoreactive membrane area. The nucleus (N) is devoid of label. E = endothelial cell; C = capillary lumen. Scale bar = 0.5 lxm. Fig. 9. Immunoreactive glial cell of the chick optic tectum at higher magnification. Labelled polyribosomes (pr) are associated with either an immunopositive plasma membrane segment (arrowhead) or the outer nuclear envelope. Scale bar = 0.5 Bin. Fig. 10. Neuropil of the chick optic tectum. KRLI in a dendrite synapsing with an axon terminal (a). Arrow marks the synaptic contact. Staining can be seen in non-synaptic membrane areas (arrowhead) and subjacent microtubules (small arrows). m = mitochondrion; sv = synaptic vesicles. Scale bar = 0.25 ~m. Fig. 11. Immunoreactive dendrite (d) in the neuropil. Staining is associated with non-synaptic membranes (arrowhead) and microtubules (small arrows). Scale bar = 0.25 Bin.

~c-opioid receptor in rat and chick brains
unable to detect KRLI in Golgi cisternae, a typical site of glycine receptor (Naas et aI., 1991).
The cellular distr ibution of KRLI, observed in neurons and glial cells in the present s tudy appears to be conserved in phylogenet ical ly distant species.
Acknowledgements
This w o r k was suppor t ed by OTKA (project no. 904)
485
and OMFB (project no. 92-97-07-0032) (to KM) and by OTKA (project no. 1097) and Minis t ry of Welfare (T04 369/93) (to AC). GN was the holder of a t ravel fel lowship unde r the aegis of a collaborative exchange agreement be tween the Polish and Hunga r i an Acad- emies. The skilled assistance of Mrs Zsuzsanna Magyar , Mrs Erzs6bet Bor6k and Mrs M~iria Szfisz is great ly appreciated.
References
ALLEN, D. L., JOHNSON, A. E., TEMPEL, A., ZUKIN, R. S., LUINE, V. N. & McEWEN, B. S. (1993) Serotoni- nergic lesions decrease ~- and 5-opiate receptor binding in discrete areas of the hypothalamus and the midbrain central gray. Brain Research 625, 269-75.
AOKI, C., ZEMCIK, A. B., STADER, C. D. & PICKEL, V. M. (1989) Cytoplasmic loop of ]3-adrenergic receptors: synaptic and inracellular localization and relation to catecholaminergic neurons in the nuclei of the solitary tracts. Brain Research 493, 331-467.
BEAUVILLAIN, J. C., MOYSE, E., DUTRIEZ, I., MITCHELL, V., POULAIN, P. & MAZZUCA, M. (1992) Localization of mu opioid receptors on the membranes of nerve endings and tanycytes in the guinea-pig median eminence by electron microscopic radioautography. Neuroscience 49, 925-36.
BLACKBURN, T. P., CROSS, A. J., HILLE, C. & SLATER, P. (1988) Autoradiographic localization of delta opiate receptors in rat and human brain. Neuroscience 27, 497- 506
CSILLAG, A., BOURNE, R. C., KJ~LMAN, M., BOXER, M. I. & STEWART, M. G. (1989) [3H]Naloxone binding in the brain of the domestic chick (GalIus domesticus) determined by in vitro quantitative autoradiography. Brain Research 479, 391-6.
CSILLAG, A., BOURNE, R. & STEWART, M. G. (1990) Distribution of mu, delta, and kappa opioid receptor binding sites in the brain of the one-day old domestic chick (Gallus domesticus): an in vitro quantitative autoradiographic study. Journal of Comparative Neurol- ogy, 302, 543-51.
CSILLAG, A., STEWART, M. G., SZEKELY, A. D., MAGLOCZKY, Z., BOURNE, R. C. & STEELE, R. J. (1993) Quantitative autoradiographic demonstration of changes in binding to delta opioid, but not mu or kappa receptors, in chick forebrain 30 minutes after passive avoidance training. Brain Research 613, 96-105.
DESJARDINS, G. C., BRAWER, J. R. & BEAUDET, A. (1990) Distribution of ~. 6 and K opioid receptors in the hypothalamus of the rat. Brain Research 536, 114-23.
DEVICHE, P., COTZER, P. & GULLEDGE, C.. C. (1993) Identification, partical characterization, and hypothala- mic distribution of K, g and 5 opoid receptors in a passerine songbird (Junco hyemalis). Brain Research 614, 220-6.
DE VRIES, T. J., HAGENBOOM, F., MULDER, A. H. & SCHOFFELMEER, A. N. M. (1990)Ontogeny of ~t-, 5-and K-opioid receptors mediating inhibition of neurotrans- mitter release and adenylate cyclase activity in rat brain.
Developmental Brain Research 54, 63-9. DRAKE, C. T., TERMAN, G. W., SIMMONS, M. L. MILNER, T. A.,
KUNKEL, D. D., SCHWARTZKROIN, P. A. & CHAVKIN, C. (1994) Dynorphin opioids present in dentate granule cells may function as retrograde inhibitory neurotransmitters. Journal of Neuroscience 14, 3736-50.
EVANS, C. J., KEITH, D. E., MORRISON, H., MAGENDZO, K. & EDWARDS, R. H. (1992) Closing of a delta opioid receptor by functional expression. Science 258, 1952-5.
FODOR, M., CSIFFARY, A., KISS, P. & PALKOVITS, M. (1990) Dyd~orphin A-containing neural elements in the nucleus of the solitary tract of the rat. Brain Research 522, 251-8.
GOLDSMITH, P.C., BOGGAN, J.E. & THIND, K.K. (1991) Opioid synapses on vasopressin neurons in the para- ventricular and supraoptic nuclei of juvenile monkeys. Neuroscience 45, 709-19.
GOLDSTEIN, A. (1987) Binding selectivity profiles for ligands of multiple receptor types: focus on opioid receptors. Trends in Pharmacological Sciences 8, 456-9.
GOLDSTEIN, A. & NAIDU, A. (1989) Multiple opioid receptors: ligand selectivity profiles and binding site signatures. Molecular Pharmacology 36, 265-72.
GRAMSCH, C., SCHULZ, R., KOSINS, S. & HERZ, A.(1988) Monoclonal anti-idiotypic antibodies to opioid recep- tors. Journal of Biological Chemistry 263, 5853-9.
HAMEL, E. & BEAUDET, A. (1984) Electron microscopic autoradiographic localization of opioid receptors in rat neostriatum. Nature 312, 155-7.
HASSAN, A. H. S., ALMEDIA, O. F. X., GRAMSCH, C. & HE RZ, A. (1989) Immunocytochemical demonstration of opioid receptors in selected rat brain areas and neuroblastoma X glioma hybrid (NG108-15) cells using a monodonal anti-idiotypic antibody. Neuroscience 32, 269-78.
HAUSER, K. F., & STIENE-MARTIN, A. (1993) Opiates and the regulation of nervous system development: evi- dence from in vitro studies. In The Nemvbiology of Opiates (edited by HAMMER, R. P.) pp. 23-61. Boca Raton: CRC Press.
INENAGA K., NAGATOMO, T., NAKAO, K., YANAIHARA, N. & YAMASHITA, H. (1994) Kappa-selective agonists decrease postsynaptic potentials and calcium compo- nents of action potentials in the supraoptic nucleus of rat hypothalamus in vitro. Neuroscience 58, 331-40.
JOMARY, C., GAIRIN, J. E. & BEAUDET, A. (1992) Synaptic localization of K opioid receptors in guinea pig neostriatum. Proceedings of the National Academy of Sciences (USA) 89, 564-8.

486 M A D E R S P A C H , T A K A C S , N I E W I A D O M S K A a n d C S I L L A G
KIEFFER, B. L., BEFORT, K., GAVERIAUX-RUFF, C. & HIRTH, C. G. (1992) The 3-opioid receptor: isolation of a cDNA by expression cloning and pharmacological characterization. Proceedings of the National Academy of Sciences (USA) 89, 12048-52.
KUJIRAI, K., FAHN, S. & CADET, J. L. (1991) Receptor autoradiography of ~t and 3 opioid peptide receptors in spontaneously hypertensive rats. Peptides 12, 779-85.
LEVEI, A. I., HERSCH, S. M., RYE, D. B., SUNAHARA, R. K., NIZNIK, H. B., KITT, C. A., PRICE, D. L., MAGGIO, R., BRANN, M. R. & CILIAX, B. J. (1993)Localization of D~ and D2 dopamine receptors in brain with subtype- specific antibodies. Proceedings of the National Academy of Sciences (USA) 90, 8861-5.
MADERSPACH, K. & NEMETH, K. (1993) Immunocyto- chemical visualization of kappa-opioid receptors on chick embryonic neurons differentiating in vitro. Neuroscience 57, 459-65.
MADERSPACH, K. & SOLOMONIA, R. (1988) Glial and neuronal opioid receptors: apparent positive coopera- tivity observed in intact cultured cells. Brain Research 441, 41-7.
MADDERSPACH, K., NI~METH, K., SIMON, J., BENYHE, S., SZUCS, M. & WOLLEMANN, M. (1991) A monoclonal antibody recognizing K- but not ~t- or 6-opioid receptors. Journal of Neurochemistry 56, 1897-904.
MAGGI, R.,LIMONTA, P.,DONDI, D.,MARTINI, L.&PIVA, F. (1989) Distribution of kappa opioid receptors in the brain of young and old male rats. Life Sciences 45, 2085-92.
MANSOUR, A., KHACHATURIAN, H., LEWIS, M. E., AKIL, H. & WATSON, S. J. (1988) Anatomy of CNS opioid receptor. Trends in Neurosciences 11, 308--14.
MARTIN, W. R., EADES, C. G., THOMPSON, J. A., HUPPLER, R. E. & GILBERT. P. E. (1976) The effects of morphin- and nalorphine-like drugs in the nondepen- dent and morphin-dependent chronic spinal dog. Journal of Pharmacology and Experimental Therapeutics 197, 517-32.
MENG, F., XIE, G.-X., THOMPSON, R. C., MANSOUR, A., GOLDSTEIN, A., WATSON, S. I. & AKIL, H. (1993) Cloning and pharmacological characterization of a rat ~c opioid receptor. Proceedings of the National Academy of Sciences (USA) 90, 9954-8.
MOLLERAU, C., PARMENTIER, M., MAILLEUX, P., BUTOUR, J.-L., MOISAND, C., CHALON, P., CAPUT, D., VASSART, G. & MEUNIER, J.-C. (1994) ORL1, a novel member of the opioid receptor family. Cloning, func- tional expression and localization. FEBS Letters 341, 33-8.
MULDER, A. H., WARDEH, G., HOGENBOOM, F. & FRANKHUYSEN, A. L. (1984) ~c- and 6-opioid receptor agonists differentially inhibit striatal dopamine and acetylcholine release. Nature 308, 278-80.
MYERS, W. E. & GLASEL, J. A. (1987) Subclass-specificity of antMdiotypic anti-opiate receptor antibodies in rat brain, guinea pig cerebellum, and neurobiastoma X glioma (NG 108-15). Life Sciences 38, 1783-8.
NAAS, E., ZILLES, K., GNAHN, H., BETZ, H., BECKER, C.-M. & SCHRODER, H. (1991) Glycine receptor immunoreactivity in rat and human cerebral cortex. Brain Research 561, 139-46.
ORNATOVSKA, M. & GLASEL, J. A. (1991) Stereospecificity of opiate receptors in a synchronized culture of NG 108-
15 cells probed with fluorescent anti-idiotypic anti- bodies. Molecular Pharmacology 40, 354--9.
REINER, A., BRAUTH, S. E., KITT, C. A. & QUIRION, R. (1989) Distribution of mu, delta and kappa opiate receptor types in the forebrain and midbrain of pigeons. Journal of Comparative Neurology 280, 359-82.
REISINE, T. & BELL, G. (1993) Molecular biology of opioid receptors. Trends in Neurosciences 16, 506-10.
SCHMAHL, W., FUNK, R., MIASKOWSKI, U. & PLEDL, J. (1989) Long-lasting effects of naltrexone, an opioid receptor antagonist, on cell proliferation in developing rat forebrain. Brain Research 486, 297-300.
SCHMIDT, P., SCHRODER, H., MADERSPACH, K. & STAAK, M. (1994) Immunohistochemical localization of lc opioid receptors in the human frontal cortex. Brain Research 654, 223-33.
s c H R 0 D E R, H. (1992) Immunohistochemistry of cholinergic receptors. Anatomy and Embryology 186, 407-29.
SLATER, P., CROSS, A. J., HEWITT, L. & SEVAGE, W. I. (1989) Autoradiographic visualization of kappa opioid receptors with labelled dynorphins in guinea pig brain. Journal of Receptor Research 9, 171-80.
SOMOGYI, P., TAKAGI, Hi, RICHARDS, J. G. & MOHLER, H. (1989) Subcellular localization of benzodiazepine/ GABA A receptors in the cerebellum of rat, cat and monkey using monoclonal antibodies. The Journal of Neuroscience 9, 2197-209.
STIENE-MARTIN, A., GURWELL, J. A. & HAUSER, K. F. (1991) Morphine alters astrocyte growth in primary cultures of mouse glial cells: evidence for a direct effect of opiates on neural maturation. Developmental Brain Research 60, 1-7.
TRILLER, A., CLUZEAUD, F., PFEIFFER, F., BETZ, H. & KORN, H. (1985) Distribution of glycine receptors at central synapses. An immunoelectronmicroscopy study. The Journal of Cell Biology 101, 683-8.
UNTERWALD, E. M., KNAPP, C, & ZUKIN, R. S. (1991) Neuroanatomical localization of K1 and ~2 opioid receptors in rat and guinea pig brain. Brain Research 562, 57-65.
VOGT, B. A., PLAGER, M. D., CRINO, P. B. & BIRD, E. D. (1990) Laminar distributions of muscarinic acetyl- choline, serotonin, GABA and opioid receptors in human posterior cingulate cortex. Neuroscience 36, 165- 74.
WANG, J. B., IMAI, Y., EPPLER, C. M., GREGOR, P., SPIVAC, C. E. & UHL, G. R. (1993) ~t opiate receptor: cDNA cloning and expression. Proceedings of the National Academy of Sciences (USA) 90, 10230-4.
WOLLEMANN, M., BENYHE, S. & SIMON, J. (1993) The kappa-opioid receptor: evidence for the different subtypes. Life Sciences 52, 599--611.
ZUKIN, R. S. & ZUKIN, S. R. (1987) The case for multiple opiate receptors. Trends in Neurosciences 7, 160-4.
ZUKIN, R. S., EGHBALI, M., OLIVE, D., UNTERWALD, E. M. & TEMPEL, A. (1988) Characterization and visualization of rat and guinea pig brain tc opioid receptors: evidence for K 1 and ~2 opioid receptors. Proceedings of the National Academy of Sciences (USA) 85, 4061-5.
ZAGON, I. S. & McLAUGHLIN, P. J. (1990) Ultrastructural localization of enkephalin-like immunoreactivity in developing rat cerebellum. Neuroscience 34, 479-89.
Related Documents











