ELSEVIER Mutation Research 305 (1994) 197-209 Fundamental and Molecular Mechanisms of Mutagenesis Possible integration of Trypanosoma cruzi kDNA minicircles into the host cell genome by infection Antonio R.L. Teixeira a,,, Enrique R. Argafiaraz a, Lucio H. Freitas Jr. a, Zulmira G. M. Lacava b, Jaime M. Santana a, Helena Luna b a Laboratory for Multidisciplinary Research on Chagas' Disease, Department of Pathology, Faculty of Health Sciences, b Laboratory of Genetics, Department of Genetics and Morphology, Institute of Biology, Universityof Brasilia, 70910.900 Brasilia, Brazil (Received 25 January 1993) (Revision received 5 September 1993) (Accepted 22 September 1993) Abstract Infection with Trypanosoma cruzi is known to induce the division of peritoneal macrophages in BALB/c mice. We have demonstrated, by cytogenetic analysis, that accessory DNA elements are associated with the metaphase macrophage chromosomes of such infected macrophages. The identification of these accessory DNA elements with T. cruzi DNA is strongly supported by the association of 3H-label with some chromatids in macrophages previously infected with T. cruzi which had been labelled with 3H-methyl-thymidine. The karyotyping consistently showed preferential associations of T. cruzi DNA with chromosomes 3, 6 and 11. A conclusive demonstration of the parasite origin of the integrated DNA came from fluorescein in situ hybridization studies using specific parasite DNAs as probes. In order to determine the identity of the inserted DNA and to investigate the nature of the integration mechanism, Southern blot analyses were performed on DNA extracted from both uninfected and infected (but parasite-free) macrophages. Hybridizations of BamHI, EcoRI and TaqI digests of DNA from T. cruzi-infected host cells all revealed the presence of a 1.7-kb DNA fragment when probed with kDNA. The covalent association of kDNA with that of the host was confirmed by the fact that AluI and Hinf-I digests of DNA from infected host cells produced a number of bands, in a size range of 0.8-3.6 kb, which hybridized with kDNA minicircles. None of these bands was found in DNA purified from cell-free preparations of the parasite and thus it must be concluded that they represent insertion fragments between parasite and host cell DNA. These results strongly suggest that kDNA minicircles from T. cruzi have been integrated into the genome of the host cell following infection. Key words: Trypanosoma cruzi; kDNA transfection; Gene rearrangement; Chagas' disease 1. Introduction Trypanosoma cruzi is a flagellate protozoan of * Corresponding author, the order Kinetoplastid, whose life cycle in the 0027-5107/94/$07.00 © 1994 Elsevier Science B.V. All rights reserved SSDI 0027-5107(93)E0190-2

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ELSEVIER Mutation Research 305 (1994) 197-209
Fundamental and Molecular Mechanisms of Mutagenesis
Possible integration of Trypanosoma cruzi kDNA minicircles into the host cell genome by infection
Antonio R.L. Teixeira a,,, Enrique R. Argafiaraz a, Lucio H. Freitas Jr. a, Zulmira G. M. Lacava b, Jaime M. Santana a, Helena Luna b
a Laboratory for Multidisciplinary Research on Chagas' Disease, Department of Pathology, Faculty of Health Sciences, b Laboratory of Genetics, Department of Genetics and Morphology, Institute of Biology, University of Brasilia,
70910.900 Brasilia, Brazil
(Received 25 January 1993) (Revision received 5 September 1993)
(Accepted 22 September 1993)
Abstract
Infection with Trypanosoma cruzi is known to induce the division of peritoneal macrophages in BALB/c mice. We have demonstrated, by cytogenetic analysis, that accessory DNA elements are associated with the metaphase macrophage chromosomes of such infected macrophages. The identification of these accessory DNA elements with T. cruzi DNA is strongly supported by the association of 3H-label with some chromatids in macrophages previously infected with T. cruzi which had been labelled with 3H-methyl-thymidine. The karyotyping consistently showed preferential associations of T. cruzi DNA with chromosomes 3, 6 and 11. A conclusive demonstration of the parasite origin of the integrated DNA came from fluorescein in situ hybridization studies using specific parasite DNAs as probes.
In order to determine the identity of the inserted DNA and to investigate the nature of the integration mechanism, Southern blot analyses were performed on DNA extracted from both uninfected and infected (but parasite-free) macrophages. Hybridizations of BamHI, EcoRI and TaqI digests of DNA from T. cruzi-infected host cells all revealed the presence of a 1.7-kb DNA fragment when probed with kDNA. The covalent association of kDNA with that of the host was confirmed by the fact that AluI and Hinf-I digests of DNA from infected host cells produced a number of bands, in a size range of 0.8-3.6 kb, which hybridized with kDNA minicircles. None of these bands was found in DNA purified from cell-free preparations of the parasite and thus it must be concluded that they represent insertion fragments between parasite and host cell DNA. These results strongly suggest that kDNA minicircles from T. cruzi have been integrated into the genome of the host cell following infection.
Key words: Trypanosoma cruzi; kDNA transfection; Gene rearrangement; Chagas' disease
1. Introduct ion
Trypanosoma cruzi is a flagellate p ro tozoan of * Corresponding author, the order Kinetoplastid, whose life cycle in the
0027-5107/94/$07.00 © 1994 Elsevier Science B.V. All rights reserved SSDI 0027-5107(93)E0190-2

198 A.R.L. Teixeira et aL /Mutation Research 305 (1994) 197-209
mammalian host includes blood trypomastigotes and intracellular amastigotes. High parasitemias in acutely infected individuals last 3-5 months. Chagas' disease is a complex of clinical manifes- tations that may result from infections with T. cruzi (Teixeira et al., 1978). The acute phase of the infection in infants and children is usually asymptomatic and subsides spontaneously. Cell- mediated immunity to the parasite antigen is lacking in these patients. In contrast, symp- tomatic acute T. cruzi infections (fever, enlarged spleen and lymph nodes, myocarditis and heart failure) are seen in a minority of individuals, in whom strong delayed-type skin reactions against the parasite antigen can be elicited. Thus, mor- bidity and mortality in Chagas' disease appear not to be related to the presence of T. cruzi forms in the blood and tissues of mammalian hosts (Teixeira, 1987). Instead, severe tissue dam- age that accompanies the chronic myocarditis of the disease appears to be associated with de- layed-type hypersensitivity.
The lack of an animal model of Chagas' dis- ease led us to standardize T. cruzi infection in rabbits (Teixeira et al., 1983). These experimental animals survived the acute infection but they died of chronic Chagas' disease with the same features as seen in the human disease. The rabbit model led us to other experiments, in which we evalu- ated the benefits of treatment of T. cruzi infec- tions with the trypanocidal nitroarene Benznida- zole (Teixeira et al., 1990a). We showed that the administration of Benznidazole in acutely and chronically T. cruzi-infected rabbits did not stop the progressively destructive autoimmune my- ocarditis, a hallmark of Chagas' disease, although the parasite was no longer demonstrable in the body of the treated animals. What could be sus- taining so active a host anti-self immune response in Benznidazole-treated Chagasic rabbits? We postulate that T. cruzi DNA is retained within the host cells after the overt infection has ended.
Although current notions in biology indicate that natural bai~riers would not allow trans-king- dom gene transfer, it has been shown that hori- zontal DNA conjugation is common among prokaryotes (Trieu-Cuot et al., 1988). Further, DNA transfer from prokaryotes to eukaryotes has
been reported (Stachel and Zambriski, 1989). Heinemann and Sprague (1989) have shown that conjugative plasmids of Escherichia coli can mo- bilize DNA transmission from this bacterium to the yeast Saccharomyces cerevisiae. In fact, bac- terium-yeast conjugation is not the only example of prokaryote-eukaryote gene transfer. Bakkeren et al. (1989) have shown direct DNA transfer from the soil-borne phytopathogen Agrobac- terium tumefaciens into plant ceils. During this cell-cell interaction a specific transfer DNA (TDNA) is mobilized from a donor bacterium to the plant cell nucleus and stabilized by chromoso- mal integration. Expression of TDNA genes in the plant cell leads to its transformation and the production of specialized nutrients (reviewed in Stachel and Zambriski, 1989). Furthermore, in certain cases of virus integration in human and bird liver cells, insertional activation of genes involved in specific cell functions has been ob- served (Yamazoe et al., 1991; Aufiero et al., 1990; Caselmann et al., 1990). More recently, the hori- zontal transfer of Drosophila genes by the mite Proctolaelaps regalis (Houck et al., 1991) has been suggested. If ectoparasites can accomplish inter- species gene transfer, there seems no reason why the endoparasitic T. cruzi should not.
We have first sought evidence for our postu- late of DNA transfer between T. cruzi and its mammalian host cell by examining the macrophages of acutely infected mice. It has been shown that T. cruzi chromatin can be present in macrophage chromosomes obtained at different periods of the acute infection (Teixeira et al., 1991). In this study we show further, by in situ hybridization and Southern blot analyses, that specific T. cruzi-DNA probes hybridize to ge- nomic DNA from diverse mammalian host cells in vitro, and in vivo in the acute and chronic phases of infection.
2. Material and methods
2.1. Chemical agents
The culture media used were RPMI and MEM with L-glutamine and antibiotics purchased from

A.R.L. Teixeira et al. / Mutation Research 305 (1994) 197-209 199
Flow Laboratories, USA. The fetal calf serum, mycoplasma and virus free, was from Laborclin, Curitiba, Brazil. The nitroarene Benznidazole (N-benzyl-2-nitroimidazoleacetamide) was a gift of Roche Pharmaceutical Industries, Switzerland. The radiochemicals 3H-methyl-thymidine (5.1 Ci/mmole specific activity), and a-32p-dATP (3000 Ci/mmole specific activity) were purchased from Amersham, UK. Ilford L4 nuclear emulsion was employed. The genomic DNA isolation kit and the nonradioactive nucleic acid detection sys- tem were from BRL, USA. The nick-translation kit was obtained from Boehringer-Mannheim, Germany. The biotin-ll-dUTP was from Biorad, whereas the anti-biotin antibody was obtained from Vector Laboratories, USA. The FITC-con- jugated rabbit anti-goat antibody was obtained from Nordic Laboratories, The Netherlands. The restriction enzymes were from Pharmacia, Swe- den. The remaining chemicals and reagents were all purchased from Sigma Chemical Co., USA.
2.4. Cell culture, T. cruzi infection and treatment
The U-937 human macrophage and the L-929 mouse fibroblast-like cell lines were grown in RPMI and MEM, respectively, supplemented with 250 mM L-glutamine and antibiotics. Either macrophages in the supernatant medium or fi- broblasts in monolayers were inoculated with 105 blood trypomastigotes of the Berenice stock of T. cruzi. Ten days after infection, the cultures were treated with the trypanocidal nitroarene Ben- znidazole (100 /~g/ml). The eradication of the parasite forms was shown by the absence of the intracellular amastigote or the trypomastigote in the supernatant medium of cultures kept in serial passages for 4 months at 37°C. To confirm the eradication, T. cruzi-infected, Benznidazole- treated ceils were also co-cultured in liver infu- sion tryptose (LIT) medium at 27°C. The epi- mastigote forms of the parasite were not recov- ered after eight serial passages.
2.2. Parasites
The Berenice stock of T. cruzi isolated from a patient with acute Chagas' disease, was used. The parasite has been maintained in our Laboratory through serial passages in mice, since infection usually causes death in 3 weeks' time. Tissue parasitism with amastigote forms of T. cruzi in- volves mostly the mononuclear phagocytic cells and the cardiac and skeletal muscle fibers.
2.3. Inoculation of mice with trypomastigotes of T. cruzi
Experiments were carried out in groups of 25 3-month-old female BALB/c mice, intraperi- toneally (i.p.) inoculated with 105 blood trypo- mastigote forms of the Berenice stock of T. cruzi. Parasitemia was detected in mice 4 days after inoculation and reached its highest level at days 8-12 of the infection. Surviving mice with unde- tected parasitemias remained chronically infected several months after the acute infection. Control groups consisted of non-infected BALB/c mice of the same age and sex.
2.5. Collection of peritoneal macrophages for cyto- genetic analysis
Peritoneal macrophages were collected from T. cruzi-infected mice at days 4, 7, 9, 12 and 15 of the acute infection. The non-immune yeast phagocytosis assay (Silverstein et al., 1977) was used to show these cells to be macrophages, whose division occurs in the peritoneal cavity. Briefly, groups of five mice received i.p. 20 ~g of colchicine in 1 ml of PBS, in order to interrupt cell division induced by the intracellular infec- tion. Two hours later, the animals received i.p. 10 IU of heparin in 3 ml of PBS, and the mac- rophages were collected using a Pasteur pipette. Peritoneal macrophages were also collected from normal mice. In these control experiments, mice were immunized with 200 mg of crude ovalbumin, emulsified in Freund's complete adjuvant (1:1 v/v). Three weeks after immunization, the ani- mals were challenged with 200 mg of ovalbumin in saline, and their peritoneal macrophages were collected 30 h later (Luna and Mariano, 1991). The metaphase macrophage chromosome spreads were prepared according to standard procedures (Viegas-Pequignot, 1989a). The cytogenetic anal-

200 A.R.L. Teixeira et aL /Muta t ion Research 305 (1994) 197-209
ysis and G-banding karyotyping were carried out in Giemsa-stained spreads.
2. 6. Autoradiography
In these experiments, a fetal mice skeletal muscle cell culture was established and infected with 105 T. cruzi trypomastigotes. At days 2 and 4 after the infection, dividing intracellular amastig- otes were labelled by addition of 0.5 mCi of 3H-methyl-thymidine (specific activity= 5.1 Ci / mmole) to the cell culture. Trypomastigotes that emerged in the supernatant culture medium were collected by centrifugation. 105 thoroughly washed 3H-thymidine-labelled trypomastigotes contained 5 x 104 cpm. 3H-DNA-labelled trypo- mastigotes were used to infect mice and peri- toneal macrophages were collected at several pe- riods of the infection. The absence of muscle cell contamination in thoroughly washed labelled try- pomastigotes, as well as in the peritoneal mac- rophage spreads, was monitored by light mi- croscopy. The chromosome spreads were ob- tained as usual from metaphase macrophages from mice infected with either 3H-DNA-labelled or unlabelled T. cruzi. The slides were dipped in autoradiograph emulsion, air-dried, stored in the dark for 2-3 weeks, and developed as described elsewhere (Harper and Saunders, 1981). Autora- diographs were analyzed directly from the micro- scope.
2. 7. In situ hybridization
The macrophage chromosome spreads used in this study were obtained from acutely and chroni- cally T. cruzi-infected mice and from control mice. The slides were air-dried and treated RNase (100 /xg/ml) for 1 h, at 37°C. After three washes in 2 x SSC, the chromosomes were denaturated in 70% formamide for 2 min, at 70°C, as described elsewhere (Viegas-Pequignot et al., 1989b).
The T. cruzi-DNA probes used were prepared from epimastigote forms of the parasite grown in LIT medium. The cells were harvested in the exponential growth phase and their total DNA extracted according to standard protocol (Mania- tis et al., 1989). Samples of DNA were cleaved
with KLAI endonuclease according to the manu- facturer's protocol. Kinetoplast DNA was ex- tracted using the shearing method (Simpson and Berliner, 1974; Gibson et al., 1985). The nDNA and the kDNA probes were analyzed by elec- trophoresis and their bands eluted from agarose gels. Genomic (tDNA), nuclear (nDNA) and kinetoplast (kDNA) probes were labelled with biotin-ll-dUTP by nick translation, according to the manufacturer's protocol, and purified in a Sephadex G50 column.
The biotin-labelled probes (10 ng/~l) were diluted in a hybridization mixture containing salmon DNA (1:200 ratio), and denatured at 100°C for 5 min. Then, 20 /xl of each probe was placed on the slides at the chromosome spread sites, covered with plastic film and incubated overnight in a moist chamber at 42°C. After three washes in 50% formamide in 2 x SSC, the spreads were flooded with a 1:250 dilution of rabbit anti-biotin antibody, and incubated for 45 min, at 37°C. Again, the slides were washed three times and then incubated with a 1 : 40 dilution of fluo- rescein-conjugated goat anti-rabbit IgG. The in- terphase and metaphase nuclear elements were counterstained with 1% propidium iodide solu- tion. The slides were analyzed using a Zeiss Axio- phot microscope with an HBO-50 ultraviolet light source and the microphotographs were taken with tungsten light (Scotch chrome 640T.)
2.8. Probe characterization
The T. cruzi nDNA probe used was shown to cross-hybridize with several plasmids encoding single and multicopy genes: pTC4, a full-length cDNA encoding a 40-kDa T. cruzi protein; pTC1, a partial-length eDNA encoding a protein of a 70-kDa mitochondrial heat-shock protein (En- gman et al., 1989, 1992); pK03, a full-length eDNA encoding a flagellar calcium binding protein (Gonzalez et al., 1985; Engman et al., 1989). These plasmids were kindly donated by Dr. D.M. Engman, Northwestern University, Illinois.
The T. cruzi kDNA probe used was shown to cross-hybridize with cloned kDNA minicircle in phage m13mp8 (the donation of Dr. S. Golden- berg, Instituto Oswaldo Cruz, Rio de Janeiro). In

A.R.L. Teixeira et al. / Mutation Research 305 (1994) 197-209 201
contrast, the kDNA probe did not cross-hybridize with the housekeeping genes in plasmids pTC4, pTC1 and pK03. All these probes were employed in the Southern blot analyses.
2.9. DNA extraction, cleavage, analytical agarose gel electrophoresis, Southern blotting and hy- bridization
DNA was extracted from several established cell lines: (1) U-937, human macrophages; (2) L-929, mouse fibroblast-like cells; (3) Benznida- zole-treated U-937 macrophages; (4) Benznida- zole-treated L-929 fibroblasts; (5) T. cruzi-in- fected U-937 cells; (6) T. cruzi-infected L-929 cells; (7) Benznidazole-treated T. cruzi-infected U-937 cells and (8) Benznidazole-treated T. cruzi-infected L-929 cells. A pool of cells har- vested from three 750-ml Falcon flasks was used. Briefly, the cells were removed with a scraper and suspended in lysis buffer. The extraction proceeded in accordance with the BRL genomic DNA isolation system. DNA was then purified by several extractions with phenol and chloroform and also ethanol-precipitated in 0.3 M Na-acetate. Restriction digestions were performed with EcoRI and BamHI endonucleases, according to the manufacturer's recommendation.
Electrophoresis was performed routinely in 0.8% agarose gels, containing 0.5 t~g/ml ethid- ium bromide, under a constant voltage of 35 V overnight. Gels were blotted on nylon filters and DNA bands were vacuum-transferred (Vacugene, Pharmacia). The filters were pre-hybridized and washed as described by Southern (1975). The tDNA, nDNA, and kDNA probes used were ob- tained as described above. The probes were ei- ther biotinylated or a-aEp-dATP (specific activity 3000 Ci/mMole)-labelled by nick translation. In typical experiments, the labelled probes' specific activities ranged from 7 × 107 to 5 x 108 cpm//~g. Hybridization took place at 65°C overnight in a mixture containing a specific probe, each at a final concentration of 50 ng/ml. The filters were washed in 0.1 x SSC with 0.1% SDS twice at room temperature, and at 60 and 65°C, and then exposed to Fuji RX films with intensifying screen at -80°C.
3. Results
Cytogenetic analysis of metaphase chromo- some spreads obtained from peritoneal cells of T. cruzi-infected BALB/c mice indicated an associ- ation between parasite chromatin and macro- phage chromosomes. Fig. 1 shows round-shaped accessory elements possibly associated with macrophage chromosomes through thin fila- ments. The identification of two DNA-containing organelles in these elements was taken as evi- dence of the nucleus and of the kinetoplast con- tained in the parasite genome. Therefore, free amastigote DNA was identified among chromo- somes, and associations occurred throughout in- fection. In addition, T. cruzi chromatin was ob- served in macrophage chromosome spreads ob- tained at different periods of infection. Fig. 2a shows accessory chromatin bridging two or more chromosomes. Fig. 2b shows close contact be- tween densely packed DNA elements and mouse chromosomes in metaphase macrophages. These associations were seen in several metaphase plates obtained from T. cruzi-infected mice and were never observed with mock-infected control mice.
_A
4
V B 1 Fig. 1. Giemsa-stained metaphase of a chromosome macrophage chromatin from Trypanosoma cruzi-infected mice. Arrows show amastigote forms that can be identified by two DNA-containing organelles: nucleus and kinetoplast. Insert shows DNA element-bridged chromosome through a thin filament. The peritoneal cell was collected on day 4 of infec- tion and subjected to standard chromosome spread prepara- tion. 1000×.

202 A.R.L. Teixeira et al. / Mutation Research 305 (1994) 197-209
2 a ¸
t • !
Fig. 2. Giemsa-stained chromosomes from Trypanosoma cruzi-inoculated mice on day 11 of infection. Arrows show accessory DNA bridging some chromosomes (a) and closely associated with others as densely packed chromatin (b) 1000 ×.
We analyzed autoradiograph spreads of peri- toneal cells, collected from 3H-DNA-labelled T. cruzi-infected mice. As shown in Fig. 3, 3H-DNA silver grains were found in macrophage nuclei, including those in metaphase chromosomes. Spe- cific silver dots were present in cells collected at day 4 of infection, but reached a peak at day 12 when 6.2% of the peritoneal cells showed radio- labelled nuclei, as shown in Fig. 4. At this stage of the infection, 9.1% of the metaphase mac- rophages showed radiolabelled chromatids. The absence of background silver dots elsewhere in the spreads could be explained by the small
Fig. 3. Autoradiographs of spreads of peritoneal cells col- lected from mice infected with 3H-DNA-labelled Try- panosoma cruzi, subjected to G-banding and Giemsa staining. 3H-DNA silver grains (arrows) are seen in interphase (a) and in metaphase chromatids (b), on day 4 of infection. 1000x.
i
,
Fig. 4. Autoradiograph of metaphase macrophage chromo- somes collected from the peritoneal cavity of mice infected with 3H-DNA-labelled Trypanosoma cruzi. 3H-DNA silver grains in chromatids of two chromosomes on day 12 of infec- tion (arrows). 1000×.
amount of 3H-DNA-labelled trypomastigotes in- oculated into mice. These studies will be reported in detail elsewhere. Here we concentrate on the karyotype analysis which showed the regular presence of T. cruzi DNA in specific host cell chromosomes. In control experiments, the au- toradiographs of chromosome spreads from mice infected with unlabelled T. cruzi showed a com- plete absence of silver dots in chromatids.
The G-banding and karyotyping of metaphase macrophages from mice infected with T. cruzi were used to determine whether the parasite DNA associated preferentially with particular host chromosomes. We observed preferential as- sociation of T. cruzi DNA with chromosomes 3, 6 and 11 (Table 1). In contrast, the cytogenetic analysis of metaphase macrophage spreads ob- tained from control, non-infected mice showed none of these findings.
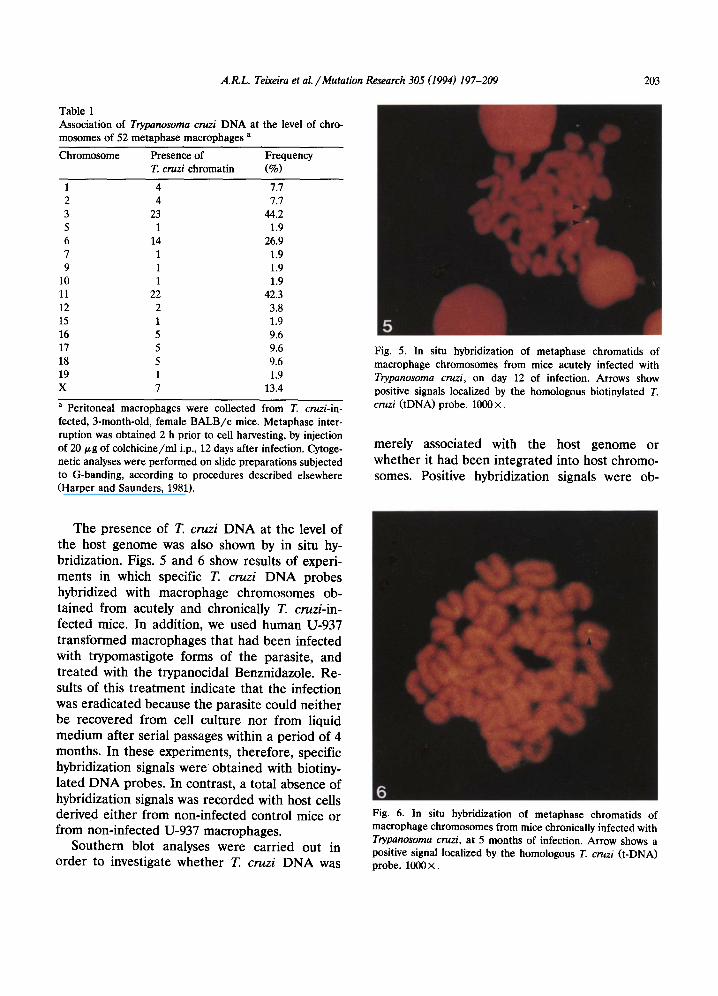
Table 1 Association of Trypanosoma cruzi DNA at the level of chro- mosomes of 52 metaphase macrophages a
Chromosome Presence of Frequency T. cruzi chromatin (%)
5
1 4 7.7 2 4 7.7 3 23 44.2 5 1 1.9 6 14 26.9 7 1 1.9 9 1 1.9
10 1 1.9 11 22 42.3 12 2 3.8 15 1 1.9 16 5 9.6 17 5 9.6 18 5 9.6 19 1 1.9 X 7 13.4
a Peritoneal macrophages were collected from T. cruzi-in- fected, 3-month-old, female BALB/c mice. Metaphase inter- ruption was obtained 2 h prior to cell harvesting, by injection of 20 ~g of coichicine/ml i.p., 12 days after infection. Cytoge- netic analyses were performed on slide preparations subjected to G-banding, according to procedures described elsewhere (Harper and Saunders, 1981).
A.R.L. Teixeira et al . / Mutation Research 305 (1994) 197-209 203
Fig. 5. In situ hybridization of metaphase chromatids of macrophage chromosomes from mice acutely infected with Trypanosoma cruzi, on day 12 of infection. Arrows show positive signals localized by the homologous biotinylated T. cruzi (tDNA) probe. 1000 X.
merely associated with the host genome or whether it had been integrated into host chromo- somes. Positive hybridization signals were ob-
The presence of T. cruzi DNA at the level of the host genome was also shown by in situ hy- bridization. Figs. 5 and 6 show results of experi- ments in which specific T. cruzi DNA probes hybridized with macrophage chromosomes ob- tained from acutely and chronically T. cruzi-in- fected mice. In addition, we used human U-937 transformed macrophages that had been infected with trypomastigote forms of the parasite, and treated with the trypanocidal Benznidazole. Re- suits of this treatment indicate that the infection was eradicated because the parasite could neither be recovered from cell culture nor from liquid medium after serial passages within a period of 4 months. In these experiments, therefore, specific hybridization signals were obtained with biotiny- lated DNA probes. In contrast, a total absence of hybridization signals was recorded with host cells derived either from non-infected control mice or from non-infected U-937 macrophages.
Southern blot analyses were carried out in order to investigate whether T. cruzi DNA was
6 Fig. 6. In situ hybridization of metaphase chromatids of macrophage chromosomes from mice chronically infected with Trypanosoma cruzi, at 5 months of infection. Arrow shows a positive signal localized by the homologous T. cruzi (t-DNA) probe. 1000 ×.
204 A.R.L. Teixeira et al. / Mutation Research 305 (1994) 197-209
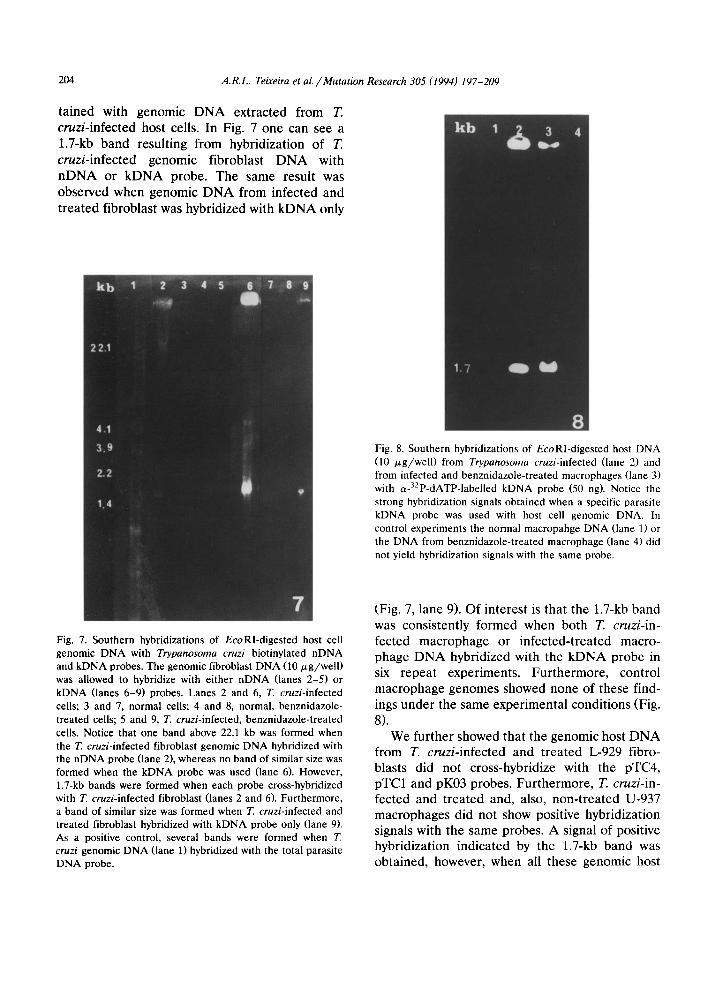
ta ined with genomic D N A extracted from T. cruz i - in f ec ted host cells. In Fig. 7 one can see a
1.7-kb b a n d resul t ing from hybridizat ion of T.
cruz i - in f ec t ed genomic f ibroblast D N A with n D N A or k D N A probe. The same result was observed when genomic D N A from infected and
t rea ted fibroblast was hybridized with k D N A only
7
Fig. 8. Southern hybridizations of EcoRl-digested host DNA (10 /xg/well) from Trypanosoma cruzi-infected (lane 2) and from infected and benznidazole-treated macrophages (lane 3) with a-32p-dATP-labelled kDNA probe (50 ng). Notice the strong hybridization signals obtained when a specific parasite kDNA probe was used with host cell genomic DNA. In control experiments the normal macropahge DNA (lane 1) or the DNA from benznidazole-treated macrophage (lane 4) did not yield hybridization signals with the same probe.
Fig. 7. Southern hybridizations of EcoRI-digested host cell genomic DNA with Trypanosoma cruzi biotinylated nDNA and kDNA probes. The genomic fibroblast DNA (10 p,g/well) was allowed to hybridize with either nDNA (lanes 2-5) or kDNA (lanes 6-9) probes. Lanes 2 and 6, T. cruzi-infected cells; 3 and 7, normal cells; 4 and 8, normal, benznidazole- treated cells; 5 and 9, T. cruzi-infected, benznidazole-treated cells. Notice that one band above 22.1 kb was formed when the T. cruzi-infected fibroblast genomic DNA hybridized with the nDNA probe (lane 2), whereas no band of similar size was formed when the kDNA probe was used (lane 6). However, 1.7-kb bands were formed when each probe cross-hybridized with T. cruzi-infected fibroblast (lanes 2 and 6). Furthermore, a band of similar size was formed when T. cruzi-infected and treated fibroblast hybridized with kDNA probe only (lane 9). As a positive control, several bands were formed when T cruzi genomic DNA (lane 1) hybridized with the total parasite DNA probe.
(Fig. 7, lane 9). Of interest is that the 1.7-kb band was consistent ly formed when both T. cruzi- in-
fected macrophage or infec ted- t rea ted macro- phage D N A hybridized with the k D N A probe in six repeat experiments . Fur the rmore , control macrophage genomes showed none of these find- ings unde r the same exper imenta l condi t ions (Fig.
8). We fur ther showed that the genomic host D N A
from T. c ruz i - in fec ted and t rea ted L-929 fibro- blasts did not cross-hybridize with the pTC4, pTC1 and pK03 probes. Fur the rmore , T. cruzi- in-
fected and t rea ted and, also, non- t r ea t ed U-937 macrophages did not show positive hybridizat ion signals with the same probes. A signal of positive hybridizat ion indicated by the 1.7-kb b a n d was obta ined, however, when all these genomic host
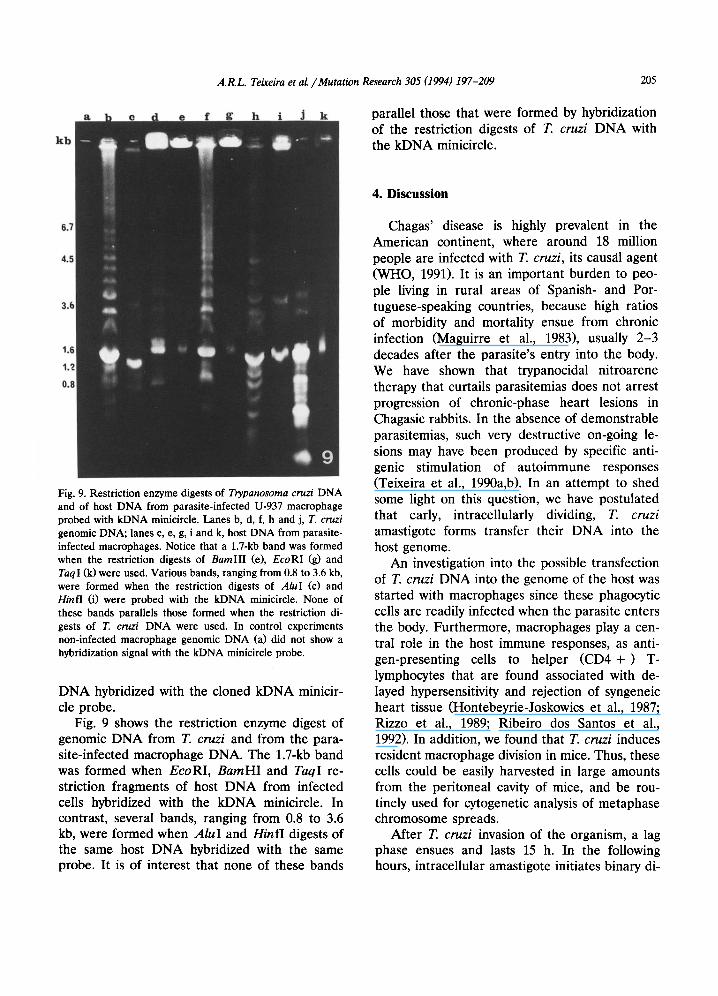
A.I~L. Teixeira et al. / Mutation Research 305 (1994) 197-209 205
l tb
& It parallel those that were formed by hybridization of the restriction digests of T. cruzi DNA with the kDNA minicircle.
6.7
4.5
3.~
1A
1A
0.i
Fig. 9. Restriction enzyme digests of Trypanosoma cruzi DNA and of host DNA from parasite-infected U-937 macrophage probed with kDNA minicircle. Lanes b, d, f, h and j, T. cruzi genomic DNA; lanes c, e, g, i and k, host DNA from parasite- infected macrophages. Notice that a 1.7-kb band was formed when the restriction digests of BamHI (e), EcoRl (g) and TaqI (k)were used. Various bands, ranging from 0.8 to 3.6 kb, were formed when the restriction digests of AluI (c) and Hinfl (i) were probed with the kDNA minicircle. None of these bands parallels those formed when the restriction di- gests of T. cruzi DNA were used. In control experiments non-infected macrophage genomic DNA (a) did not show a hybridization signal with the kDNA minicircle probe.
DNA hybridized with the cloned kDNA minicir- cle probe.
Fig. 9 shows the restriction enzyme digest of genomic DNA from T. cruzi and from the para- site-infected macrophage DNA. The 1.7-kb band was formed when EcoRI, BarnHI and TaqI re- striction fragments of host DNA from infected ceils hybridized with the kDNA minicircle. In contrast, several bands, ranging from 0.8 to 3.6 kb, were formed when AluI and HinfI digests of the same host DNA hybridized with the same probe. It is of interest that none of these bands
4. Discussion
Chagas' disease is highly prevalent in the American continent, where around 18 million people are infected with T. cruzi, its caus~il agent (WHO, 1991). It is an important burden to peo- ple living in rural areas of Spanish- and Por- tuguese-speaking countries, because high ratios of morbidity and mortality ensue from chronic infection (Maguirre et al., 1983), usually 2-3 decades after the parasite's entry into the body. We have shown that trypanocidal nitroarene therapy that curtails parasitemias does not arrest progression of chronic-phase heart lesions in Chagasic rabbits. In the absence of demonstrable parasitemias, such very destructive on-going le- sions may have been produced by specific anti- genic stimulation of autoimmune responses (Teixeira et al., 1990a,b). In an attempt to shed some light on this question, we have postulated that early, intracellularly dividing, T. cruzi amastigote forms transfer their DNA into the host genome.
An investigation into the possible transfection of T. cruzi DNA into the genome of the host was started with macrophages since these phagocytic cells are readily infected when the parasite enters the body. Furthermore, macrophages play a cen- tral role in the host immune responses, as anti- gen-presenting cells to helper (CD4 + ) T- lymphocytes that are found associated with de- layed hypersensitivity and rejection of syngeneic heart tissue (Hontebeyrie-Joskowics et al., 1987; Rizzo et al., 1989; Ribeiro dos Santos et al., 1992). In addition, we found that T. cruzi induces resident macrophage division in mice. Thus, these cells could be easily harvested in large amounts from the peritoneal cavity of mice, and be rou- tinely used for cytogenetic analysis of metaphase chromosome spreads.
After T. cruzi invasion of the organism, a lag phase ensues and lasts 15 h. In the following hours, intracellular amastigote initiates binary di-

206 A.R.L. Teixeira et aL /Mutation Research 305 (1994) 197-209
vision, with a doubling time of 12 h (Chrane and Dvorak, 1979). We observed early association of accessory DNA elements with metaphase chro- mosomes 48 h after infection (data not shown), when the parasite approximated three cycles of division. An increased number of associations was seen in macrophages collected in the follow- ing days, reaching a peak at day 14 of infection. Several patterns of association were seen with various groups of chromosomes in which DNA elements bridged mouse chromosomes, also ap- pearing to be in close contact with them. It is of interest that karyotyping of metaphase chromo- somes showed a consistent preferential associa- tion of these DNA elements with chromosomes 3, 6 and 11. It has been reported that human a- satellite DNA introduced in African green mon- key cells can be associated through bridges be- tween separate sets of chromatids (Haaf et al., 1992). This feature was considered consistent with the integrated a-satellite DNA disrupting normal chromosome segregation.
Further information was obtained by autora- diograph analysis of spreads, in which macro- phages were collected from mice acutely infected with 3H-DNA-labelled T. cruzi. 3H-DNA dots were seen in nuclei of some ceils, and also in chromatids of metaphase chromosomes. An ex- planation of the 3H-labelled material found asso- ciated with host chromosomes based on the cellu- lar uptake of 3H-DNA after the death of the parasite is unlikely. This explanation would re- quire a salvage pathway of dead T. cruzi DNA, which has not been described. Moreover, such a route of incorporation would lead to the dis- tributed incorporation of the label into all newly synthesized DNA. Instead, we observed 3H-DNA silver grains in selected murine chromosomes. The possible contamination of the murine macrophages by the muscle ceils that had been used for labelling the parasite in vitro is not supported by microscopic searches. Nevertheless, further experiments were undertaken in order to demonstrate that the incorporated DNA was re- ally of Trypanosoma origin.
We analyzed insertions of exogenous DNA into macrophage genomes by means of in situ hybridization. Metaphase macrophage chromo-
some spreads showed segments of chromosomal DNA hybridizing with homologous kDNA and also with nDNA probes, albeit with a lesser fre- quency. These findings were recorded on several occasions, when macrophages were collected from acutely and chronically T. cruzi-infected mice. None of these findings was seen in macrophage chromosomes obtained from control, non-in- fected mice.
In vitro experiments were carried out in hu- man U-937 macrophages that had been inocu- lated with T. cruzi and treated with a trypanoci- dal nitroarene. Eradication of the infection was accomplished by this treatment, as indicated by the repeated failure to recover T. cruzi from the drug-treated cell culture, or by co-culturing treated ceils in LIT medium after serial passages over 4 months. In situ hybridizations carried out on metaphase macrophage chromosome spreads derived from these cultures showed insertions of T. cruzi DNA identified with both nDNA and kDNA probes. The presence of T. cruzi DNA in macrophage chromosomes of chronically infected mice, and in the chromosomes of human macrophage whose infection had been eradi- cated, suggests that the foreign DNA was indeed been transfected into the genome of the host cell. None of these findings was seen in control experi- ments.
In addition to the U-937 macrophages, we also used a fibroblast-like, L-929 cell line to show T. cruzi DNA insertion in its genome. BamHI and EcoRI fragments of macrophage and fibroblast genomes showed a 1.7-kb DNA sequence that cross-hybridizes with a kDNA probe. This finding was consistently shown in various sources of host cells, from which the infection had been eradi- cated. Furthermore, the absence of nDNA in the same preparations was recorded. This observa- tion ruled out the possibility that the foreign material was a consequence of living parasites remaining in the host cell genome. Moreover, the 1.7-kb band that was formed when the host DNA from T. cruzi-infected macrophages was probed with kDNA, but not with pTC4, PTC1 and pK03, suggests that the phagocytic cell had eliminated the infection.
The restriction enzymes digests of genomic

A.R.L. Teixeira et aL / Mutation Research 305 (1994) 197-209 207
DNA from T. cruzi and from the parasite-in- fected macrophages showed striking differences. For a single band was formed when BamHI, EcoRI and TaqI fragments were used, and vari- ous bands larger than 1.7 kb were formed with Alub and Hinfl fragments, when probed with kDNA minicircles. These differences were ex- plained by flanking sequence differences. These results suggest that kDNA minicircles were inte- grated into the host cell genome by infection.
T. cruzi kDNA is a disk-like structure within the mitochondrial matrix. Intact kDNA contains about 10,000 minicircles and 50 maxicircles cate- nated into a large network (reviewed in Borst, 1991). Maxicircles contain genes for mitochon- drial proteins and rRNA, whereas minicircles are associated with RNA editing (Sturm and Simp- son, 1990). The reported heterogeneity of mini- circles appears to be required to provide suffi- cient genetic information for extensive editing (Weiner and Maizels, 1990). Further, it has been shown that recombination contributes to minicir- de heterogeneity (Gibson and Garside, 1990). Thus, in this study we show kDNA insertions into the genome of host ceils and, therefore, raise the question: is kDNA a 'homing DNA'?
Despite all these efforts, there still exists the suspicion that the extranuclear DNA in the kine- toplast is of bacterial origin (Margulis, 1981). It has been argued, however, that bacteria serve as vectors for horizontal transmission of genes be- tween eukaryotes (Stachel and Zambriski, 1989). Here, we show eukaryote-eukaryote transfer of genetic material by means of infections which normally take place in nature. However, the transfer mechanism whereby kDNA integrates into the host genome remains largely unexplored.
It has been postulated that trans-kingdom gene transfer may contribute to a flow of genetic mate- rial and so may influence the evolutionary pro- cess. We consider that the true implication of transxenogene (Gk., ~evoo" = strange) rearrange- ment in the biology of cell differentiation, host- parasite interactions and autoimmune disease needs further investigation. Despite the fact that kDNA minicircles do not encode proteins, a com- plex mechanism of guide RNA and RNA editing has been described (Sturm and Simpson, 1990).
We suggest that the integration of T. cruzi DNA into the host cell genome might lead to alter- ations in the regulation of host gene expression. In this respect, it is important to emphasize that the gene encoding the /3 chain of the T-cell receptor is present at locus B of chromosome 6. Furthermore, the striated muscle a-actin and heavy chain myosin genes are also present at chromosomes 3 and 11, respectively (Searle, 1987). Of particular interest is that some autoim- mune lesions in Chagas' disease appear to be related to T-cell cytotoxicity against heart and skeletal muscle myosin (Rizzo et al., 1989). Fur- ther investigations, aimed at unravelling these intricate autoimmune reactions, will be carried out taking into account the possibility of foreign genes being inserted into the host genome by means of infection. To substantiate this latter possibility, and the nature and site of T. cruzi DNA insertion, a detailed molecular investigation is presently being pursued.
5. Acknowledgements
We are indebted to E. Viegas-Pequignot, T.I. Scherrer and C. Martins de Sa for helpful sugges- tions. This work has received financial aid from FINEP and from PADCT/CNPq, Brazil.
6. References
Aufiero, B., and R.J. Schneider (1990) The hepatitis B virus X-gene product transactivates both RNA polymerase II and III promoters, EMBO J, 9, 497-504.
Bakkeren, G., Z. Koukolikov~i-Nicola, N. Grimsley and B. Hohn (1989) Recovery of Agrobacterium tumefaciens T- DNA molecules from whole plants early after transfer, Cell, 57, 847-857.
Borst, P. (1991) Why kinetoplast networks? Trends Genet. 7, 139-141.
Caselmann, W.H., M. Meyer, A.S. Kekul6, U. Lauer, P.H. Hofschneider and R. Koshy (1990) A transactivator func- tion is generated by integration of hepatitis B virus pre s /s sequences in human hepatocellular carcinoma DNA, Proc. Natl. Acad. Sci. USA, 87, 2970-2974.
Chrane, M. St. J., and J.A. Dvorak (1979) Trypanosoma cruzi: interaction with vertebrate cells. DNA synthesis and

208 A.R.L. Teixeira et al. / Mutation Research 305 (1994) 197-209
growth of intracellular amastigotes and their relationship to host cell DNA synthesis and growth, J. Protozool., 26, 599-604.
Engman, D.M., K.-H. Krause, J.H. Blumin, K.S. Kim, L.V. Kirchhoff and J.E. Donelson (1989a) A novel Ca2+-bind - ing protein in trypanosomes, J. Biol. Chem., 264, 18627- 18631.
Engman, D.M., L.V. Kirchhoff and J.E. Donelson (1989b) Molecular cloning of mtp 70, a mitochondrial member of hsp 70 family, Mol. Cell. Biol., 9, 5163-5168.
Engman, D.M., S.C. Fehr and J.E. Donelson (1992) Specific functional domains of mitochondrial hsp 70s suggested by sequence comparison of the trypanosome and yeast pro- teins, Mol. Biochem. Parasitol., 51, 153-156.
Gibson, W., and L. Garside (1990) Kinetoplast DNA minicir- cles are inherited from both parents in genetic hybrids of Trypanosoma brucei, Mol. Biochem. Parasitol., 42, 45-54.
Gibson, W., P. Borst and F. Fase-Fowler (1985) Further analysis of intra-specific variation in Trypanosoma brucei using restriction site polymorphisms in the maxicircle of kinetoplast DNA, Mol. Biochem. Parasitol., 15, 21-36.
Gonzalez, A., T.J. Lerner, M. Huecas, B. Sosa-Pineda, N. Nogueira and P.M. Lizardi (1985) Apparent generation of a segmented mRNA from two separate tandem gene fami- lies in Trypanosoma cruzi, Nucleic Acids Res., 13, 5789- 5804.
Haaf, T., P.E. Warburton and H.F. Willard (1992) Integration of human a-satellite DNA into simian chromosomes. Cen- trome protein binding and disruption of normal chromo- some segregation, Cell, 70, 681-696.
Harper, M.E., and G.F. Saunders (1981) Localization of single copy DNA sequences on G-banded chromosomes by in situ hybridization, Chromosome, 83, 431-439.
Heineman, J.A., and G.F. Sprague Jr. (1989) Bacterial con- jugative plasmids mobilize DNA transfer between bacteria and yeast, Nature, 340, 205-209.
Hontebeyrie-Joskowics, M., G. Said, G. Millon, G. Marchall and H. Eisen (1987) L3T4+ T cells able to mediate parasite specific delayed type hypersensitivity play a role in the pathology of experimental Chagas' disease, Eur. J. Immunol., 17, 1027-1031.
Houck, M.A., S.B. Clark, K.R. Petersen and M.G. Kidwel (1991) Possible horizontal transfer of Drosophila genes by the mite Proctolaelaps regalis, Science, 253, 1125-1128.
Luna, H., and M. Mariano (1991) Hyperdiploidy ant trisomy of chromosome 15 in hyperimmune, older female mice, J. Gerontol., 46, B148-151.
Maguirre, J.H., K.E. Mott, J.S. Lehman, R. Roff, T. Muniz, A.C. Guimarfies, I. Sherlock and R.H. Marrow (1983) Relationship of electrocardiographic abnormalities and seropositivity to Trypanosoma cruzi within a rural commu- nity in Northeast Brazil, Am. Heart J., 105, 287-293.
Maniatis, T., E.F. Fritsch and J. Sambrook (1989) Molecular Cloning. A Laboratory Manual, Cold Spring Harbor Labo- ratory, Cold Spring Harbor, NY.
Margulis, L. (1981) Several prokaryotes make a eukaryote, in: Symbiosis in Cell Evolution, Freeman, San Francisco, CA.
Ribeiro dos Santos, R., M.A. Rossi, J.L. Laus, J. Santana Silva, W. Savino and J. Mengel (1992) Anti-CD4 + abro- gates rejection and reestablishes long-term tolerance to syngeneic newborn hearts grafted in mice chronically in- fected with Trypanosoma cruzi, J. Exp. Med., 175, 29-39.
Rizzo, L.V., E. Cunha Neto and A.R.L. Teixeira (1989) Au- toimmunity in Chagas' disease: Specific inhibition of reac- tivity of CD4 + T cells against myosin in mice chronically infected with Trypanosoma cruzi, Infect. Immunol., 57, 2640-2644.
Searle, A.G. (1987) Mouse chromosome mapping, Genomics, 1, 3-18.
Silverstein, S., R.M. Steinman and Z.A. Cohn (1977) Endocy- tosis, Annu. Rev. Biochem., 46, 669-722.
Simpson, L., and J. Berliner (1974) Isolation of kDNA from Leishmania tarentolae in the form of a network, J. Proto- zool., 21,382-393.
Southern, E.M. (1975) Detection of specific sequences among DNA fragments separated by gel electrophoresis, J. Mol. Biol., 98, 503-517.
Stachel, S.E., and C. Zambryski (1989) Generic trans-kingdom sex? Nature, 340, 190-191.
Sturm, N.R., and L. Simpson (1990) Kinetoplast DNA mini- circles encode guide RNAs for editing of cytochrome oxidase subunit III mRNA, Cell, 61,879-884.
Teixeira, A.R.L. (1987) The Stercorarian trypanosomes, in: E.S.L. Soulsby (Ed.), Immune Responses in Parasitic In- fections: Immunology, Immunopathology, and Immuno- prophylaxis, CRC Press, Boca Raton, FL.
Teixeira, A.R.L., G. Teixeira, V. Macedo and A. Prata (1978) Acquired cell-mediated immune-depression in acuty Cha- gas' disease, J. Clin. Invest., 62, 1132-1141.
Teixeira, A.R.L., F. Figueiredo, J. Rezende Filho and V. Macedo (1983) Chagas' disease: a clinical, parasitological, immunological and pathological study in rabbits, Am. J. Trop. Med. Hyg., 32, 258-272.
Teixeira, A.R.L., J.C. C6rdoba, I. Souto-Maior and E. Solorzano (1990a) Chagas' disease: Lymphoma growth in rabbits treated with benznidazole, Am. J. Trop. Med. Hyg., 43, 146-158.
Teixeira, A.R.L., R. Silva, E. Cunha Neto, J.M. Santana and L.V. Rizzo (1990b) Malignant, non-Hodgkin's lymphomas in Trypanosoma cruzi-infected rabbits treated with ni- troarenes, J. Comp. Pathol., 103, 37-48.
Teixeira, A.R.L., Z. Lacava, J.M. Santana and H. Luna (1991) Inser~o de DNA de Trypanosoma cruzi no genoma de c61ula hospedeira de mamifero por meio de infe~sfio, Rev. Soc. Bras. Med. Trop., 24, 55-58.
Trieu-Cuot, P., C. Carlier and P. Courvalin (1988) Conjugative plasmid transfer from Enterococcus faecalis to Escherichia coli, J. Bacteriol., 170, 4388-4391.
Viegas-Pequignot, E., L.Z. Lin, B. Dutrillanx, F. Apiou and D. Panlin (1989a) Assignmenf of human desmin gene to

A.R.L. Teixeira et al. / Mutation Research 305 (1994) 197-209 209
band 2935 by non-radioactive in situ hybridization, Hum. Genet., 83, 33-36.
Viegas-Pequignot, E., B. Dutrillaux, H. Magdalenat and M. Coppey-Moisan (1989b) Mapping of single copy DNA sequences on human chromosomesby in sitn hybridization with biotinylated probes: Enhancement of detection sensi- tivity by intensified-fluorescence digital-imaging mi- croscopy. Proc. Natl. Acad. Sci. USA, 86, 582-586.
Weiner, A.M., and N. Maizels (1990) RNA editing: guided but non templated? Cell, 61,917-920.
World Health Organization (1991) Control of Chagas' Dis- ease, WHO Technical Report Series 811, WHO, Geneva.
Yamazoe, M., S. Nakai, N. Ogasawara and H. Yoshikawa (1991) Integration of woodchuck hepatitis virus (WHV) DNA at two chromosomal sites (VK and gag-like) in a hepatocellular carcinoma, Gene, 100, 139-146.
Related Documents











