Fax +41 61 306 12 34 E-Mail [email protected] www.karger.com Original Paper Cells Tissues Organs DOI: 10.1159/000320572 Portrayal of the Notch System in Embryonic Stem Cell-Derived Embryoid Bodies Isabel Alcobia a, b Andreia Gomes a Pedro Saavedra a, b Ricardo Laranjeiro a Sónia Oliveira a Leonor Parreira a, b António Cidadão a a Unidade de Biologia da Hematopoiese, Instituto de Histologia e Biologia do Desenvolvimento, Faculdade de Medicina, Universidade de Lisboa, Lisboa, e b Instituto Gulbenkian de Ciência, Oeiras, Portugal Our results show that Notch system components are con- tinuously and abundantly expressed in the multicellular en- vironments arising in differentiating EBs. In such an active Notch system, receptors and ligands do not accumulate ex- tensively at the cell surface but instead localize at cytoplas- mic foci, an observation that fits current knowledge on en- docytic modulation of Notch signaling. Our data thus sug- gest that Notch may function as a territorial modulator during early development, where it may eventually influ- ence YS hematopoiesis. Copyright © 2010 S. Karger AG, Basel Introduction Notch signaling is of utmost importance for numerous phenomena during embryonic development and in adult organisms, namely differentiation, proliferation, survival and cell fate decisions [Bray, 2006; Chiba, 2006; Fiúza and Arias, 2007]. It starts with a cell surface-signaling event occurring between adjacent cells which elicits -secre- tase-dependent proteolytic activation (S3 cleavage) on transmembrane Notch receptors (Notch1–4 in mam- mals). This activation step usually follows the interaction of the extracellular (EC) domain of Notch receptors lo- cated at the plasma membrane of a given cell, with the EC Key Words Yolk sac hematopoiesis Notch signaling Embryoid bodies Embryonic stem cells S3 cleavage Mouse Abstract We portrayed the Notch system in embryonic stem cell (ESC)-derived embryoid bodies (EBs) differentiating under the standard protocols used to assess yolk sac (YS) hemato- poiesis in vitro. Notch receptors and Notch ligands were de- tected in virtually all cells throughout EB development. Notch1 and Notch 2, but not Notch4, were visualized in the nucleus of EB cells, and all these receptors were also ob- served as patent cytoplasmic foci. Notch ligands (Delta-like1 and 4, Jagged1 and 2) were immunodetected mostly as cy- toplasmic foci. Widespread Notch1 activation was evident at days 2–4 of EB differentiation, the time window of heman- gioblast generation in this in vitro system. EBs experienced major spatial remodeling beyond culture day 4, the time point coincident with the transition between primitive and multilineage waves of YS hematopoiesis in vitro. At day 6, where definitive YS hematopoiesis is established in EBs, these exhibit an immature densely packed cellular region (DCR) surrounded by a territory of mesodermal-like cells and an outer layer of endodermal cells. Immunolabeling of Notch receptors and ligands was usually higher in the DCR. Accepted after revision: August 20, 2010 Published online: November 30, 2010 Dr. António Cidadão, Unidade de Biologia da Hematopoiese Instituto de Histologia e Biologia do Desenvolvimento Faculdade de Medicina da Universidade de Lisboa Av. Professor Egas Moniz, PT–1649-028 Lisboa (Portugal) Tel. +351 21 799 9522, Fax +351 21 799 9527, E-Mail a.cidadao @ fm.ul.pt © 2010 S. Karger AG, Basel 1422–6405/11/0000–0000$38.00/0 Accessible online at: www.karger.com/cto

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Fax +41 61 306 12 34E-Mail [email protected]
Original Paper
Cells Tissues Organs DOI: 10.1159/000320572
Portrayal of the Notch System in Embryonic Stem Cell-Derived Embryoid Bodies
Isabel Alcobia a, b Andreia Gomes a Pedro Saavedra a, b Ricardo Laranjeiro a Sónia Oliveira a Leonor Parreira a, b António Cidadão a
a Unidade de Biologia da Hematopoiese, Instituto de Histologia e Biologia do Desenvolvimento, Faculdade de Medicina, Universidade de Lisboa, Lisboa , e b Instituto Gulbenkian de Ciência, Oeiras , Portugal
Our results show that Notch system components are con-tinuously and abundantly expressed in the multicellular en-vironments arising in differentiating EBs. In such an active Notch system, receptors and ligands do not accumulate ex-tensively at the cell surface but instead localize at cytoplas-mic foci, an observation that fits current knowledge on en-docytic modulation of Notch signaling. Our data thus sug-gest that Notch may function as a territorial modulator during early development, where it may eventually influ-ence YS hematopoiesis. Copyright © 2010 S. Karger AG, Basel
Introduction
Notch signaling is of utmost importance for numerous phenomena during embryonic development and in adult organisms, namely differentiation, proliferation, survival and cell fate decisions [Bray, 2006; Chiba, 2006; Fiúza and Arias, 2007]. It starts with a cell surface-signaling event occurring between adjacent cells which elicits � -secre-tase-dependent proteolytic activation (S3 cleavage) on transmembrane Notch receptors (Notch1–4 in mam-mals). This activation step usually follows the interaction of the extracellular (EC) domain of Notch receptors lo-cated at the plasma membrane of a given cell, with the EC
Key Words
Yolk sac hematopoiesis � Notch signaling � Embryoid bodies � Embryonic stem cells � S3 cleavage � Mouse
Abstract
We portrayed the Notch system in embryonic stem cell (ESC)-derived embryoid bodies (EBs) differentiating under the standard protocols used to assess yolk sac (YS) hemato-poiesis in vitro. Notch receptors and Notch ligands were de-tected in virtually all cells throughout EB development. Notch1 and Notch 2, but not Notch4, were visualized in the nucleus of EB cells, and all these receptors were also ob-served as patent cytoplasmic foci. Notch ligands (Delta-like1 and 4, Jagged1 and 2) were immunodetected mostly as cy-toplasmic foci. Widespread Notch1 activation was evident at days 2–4 of EB differentiation, the time window of heman-gioblast generation in this in vitro system. EBs experienced major spatial remodeling beyond culture day 4, the time point coincident with the transition between primitive and multilineage waves of YS hematopoiesis in vitro. At day 6, where definitive YS hematopoiesis is established in EBs, these exhibit an immature densely packed cellular region (DCR) surrounded by a territory of mesodermal-like cells and an outer layer of endodermal cells. Immunolabeling of Notch receptors and ligands was usually higher in the DCR.
Accepted after revision: August 20, 2010 Published online: November 30, 2010
Dr. António Cidadão , Unidade de Biologia da HematopoieseInstituto de Histologia e Biologia do DesenvolvimentoFaculdade de Medicina da Universidade de Lisboa Av. Professor Egas Moniz, PT–1649-028 Lisboa (Portugal) Tel. +351 21 799 9522, Fax +351 21 799 9527, E-Mail a.cidadao @ fm.ul.pt
© 2010 S. Karger AG, Basel1422–6405/11/0000–0000$38.00/0
Accessible online at:www.karger.com/cto

Alcobia /Gomes /Saavedra /Laranjeiro /Oliveira /Parreira /Cidadão
Cells Tissues Organs 2
domain of transmembrane Notch ligands [Delta-like 1, 3 and 4(Dll1–4), and Jagged1–2 (J1–2), in mammals], ex-pressed at the surface of neighboring cells. The S3-cleaved intracellular (IC) domain of activated Notch receptors translocates to the nucleus where it binds to transcription corepressors/coactivators, inducing the expression of Notch target genes [Kopan and Ilagan, 2009]. The high diversity of Notch receptor and ligand expression, the ex-istence of multiple Notch system modulators, the variabil-ity of Notch target genes in distinct contexts, plus a con-firmed cross-talk between Notch and other key signaling pathways [Hurlbut et al., 2007; Hayward et al., 2008], add to the complexity of biological readouts of this system. The role of Notch signaling in early embryonic develop-ment is, however, poorly understood. An active Notch sys-tem may be dispensable for early embryonic development [Shi and Stanley, 2006]. Yet, both in vivo and in vitro data show that Notch signaling influences major developmen-
tal events like germ layer segregation [Croce and McKay, 2010; Revinski et al., 2010], embryonic lineage commit-ment [Yu et al., 2008], and germ layer differentiation in a ligand-specific manner [Ramasamy and Lenka, 2010].
Paradigms often used to emphasize the significance of Notch signaling are late-embryonic [Gering and Patient, 2010] and postnatal hematopoiesis [Jaleco et al., 2001; Neves et al., 2006; Orkin and Zon, 2008]. The influence of Notch on early-embryonic hematopoiesis is, in con-trast, still largely unexplored. The earliest developmental stages of hematopoiesis occur in yolk sac (YS) blood is-lands, apparently from an Flk-1-positive mesoderm-de-rived precursor, the hemangioblast [Cumano and Godin, 2007]. Notch1 activation occurs in the nascent mesoderm and subsequently in YS blood islands [Del Monte et al., 2007], i.e. at the proper sites and time frame to influence hematopoiesis. As the earliest phases of embryonic hema-topoiesis are difficult to functionally assess in vivo, em-bryoid bodies (EBs) generated from murine embryonic stem cells (ESCs) are routinely used as a standard in vitro model of YS hematopoiesis [Daley, 2003; Kyba et al., 2003; Baron, 2005; Murry and Keller, 2008]. In this study, we portray the active Notch system prevailing during the differentiation of such EBs.
Our results show early and widespread Notch activa-tion in EBs harboring YS-like hematopoiesis, suggesting that Notch may, directly or indirectly, influence such cell differentiation events. In the active Notch system of dif-ferentiating EBs, Notch receptors and ligands do not ac-cumulate extensively at the cell surface but instead local-ize at cytoplasmic foci, in agreement with current models of Notch endocytic modulation [Bray, 2006; Fiúza and Arias, 2007; Hurlbut et al., 2007; Sorensen and Conner, 2010]. Finally, as Notch components were found to be un-equally distributed in the microenvironments that arise during EB differentiation, we suggest that Notch signal-ing might function as a territorial modulator during ear-ly embryonic development.
Materials and Methods
Study Design To exclude any possible line-dependent bias on the dynamics of
emerging EB multicellular compartments as well as on the distri-bution patterns of Notch system components, three ESC lines, R1, AINV15 ESC lines (provided by A. Duarte and G. Daley, respec-tively) and ESiHoxB4, a derivate from the AINV15 line in which HoxB4 can be induced by the addition of doxycycline (DOXY, Sig-ma) (provided by G. Daley), were used in this study. Since the re-sults were similar for all ESC lines, representative examples ob-tained from the different cell lines are shown in figures 1–6 .
Abbreviations used in this paper
AINV15 AINV15 embryonic stem cell line�-CAT �-cateninCS chondroitin sulfateDCR Densely packed cellular regionDll1–4 Delta-like 1–4DOXY doxycyclineEB embryoid bodyEC extracellularE-CAD E-cadherinEDTA ethylenediaminetetraacetic acidEMA-1 epithelial membrane antigen-1ESC embryonic stem cellESiHoxB4 HoxB4-inducible embryonic stem cell lineFN fibronectinHS heparan sulfateIC intracellularICN Notch IC domain J1, J2 Jagged 1 or Jagged 2LAMP-1 lysosome markerLN lamininNICD notch intracellular domainNotch1-S3 �-secretase-dependent S3 cleavage of Notch1OPN osteopontinPEST signal peptide for protein degradationR1 R1 embryonic stem cell lineRab7 late-endosome markerRER rough endoplasmic reticulumSSEA-1 stage-specific embryonic antigen-1TROMA-I cytokeratin Endo-Aw/o DOXY without doxycyclineYS yolk sac

Notch System in ESC-Derived Embryoid Bodies
Cells Tissues Organs 3
EBs were generated from the three ESC lines using stan-dard protocols for hematopoietic differentiation (see below and online suppl. fig. 1; for all online supplementary material, see www.karger.com/doi/10.1159/000320572). Moreover, as HoxB4 is known to expand multilineage precursors (online suppl. fig. 1, online suppl. table 1) and affect the expression of some Notch li-gands and Notch target genes [Schiedlmeier et al., 2007] (online suppl. fig. 2), EBs derived from a HoxB4-inducible embryonic stem cell line (ESiHoxB4) in which HoxB4 was induced from days 4–6 of differentiation (the in vitro time window encompassing the transition between primitive to multilineage YS hematopoiesis) [Daley, 2003; Baron, 2005; Keller, 2005] were further analyzed. At specific time points of differentiation, EBs were either fixed for immunocytochemistry or extracted for Western blot analysis of active Notch1 (S3 cleaved). EBs at days 4 and 6 of differentiation (see above) were always used for immunocytochemistry. For the detection of active Notch1 (S3 cleaved), and also of Notch1 and Notch2 IC domains, labeling experiments were extended to days 2 and 3 of EB differentiation (the R1 and AINV15 lines), the time window that spans the peak of EB hemangioblast potential [Mur-ry and Keller, 2008]. Western blot analysis was also performed at days 2–6 of differentiation.
Growth and Differentiation of ESCs ESC lines were used as described [Daley, 2003; Kyba et al.,
2003; Baron, 2005; Schiedlmeier et al., 2007], with modifications. Undifferentiated ESCs were cultured in a feeder layer of mitomy-cin-C-inactivated mouse embryonic fibroblasts and maintained in Dulbecco’s modified Eagle’s medium (Sigma) supplemented with 1,000 U/ml recombinant leukemia inhibitor factor (ESGRO, Chemicon), 15% ES-qualified fetal bovine serum (Invitrogen), 1% L -glutamine (Invitrogen), 1% penicillin-streptomycin (Pen/Strep, Invitrogen), 1% nonessential amino acids (Invitrogen), 0.1 m M � -mercaptoethanol (Invitrogen), 1% nucleosides (Sigma) (ESC me-dium). Cultures were maintained at 37 ° C in humidified air with 5%CO 2 and the medium replaced daily. For ESC lines R1 and AINV15, and prior to initiating ESC differentiation into EBs, ESCs were dissociated with 0.25% trypsin-EDTA (Sigma) and cultured for 2 days in a predifferentiation medium (ESC medium with Iscove’s modified Dulbecco’s medium instead of Dulbecco’s modified Eagle’s medium) without feeder cells. For differentia-tion, 1 ! 10 5 ESCs per 90-mm plate were plated in Iscove’s mod-ified Dulbecco’s medium (Sigma) supplemented with 15% fetal bovine serum (Hyclone or Invitrogen), 1% L -glutamine (Invitro-gen), 1% Pen/Strep (Invitrogen), 1% nonessential amino acids (In-vitrogen), 0.1 m M � -mercaptoethanol (Invitrogen), 1% nucleo-sides (Sigma), 4.5 m M monothioglycerol (Sigma), 50 � g/ml ascor-bic acid (Sigma) and 200 � g/ml transferrin (Invitrogen). The medium was replaced every 2 days. Induction of HoxB4 in the ESiHoxB4 cell line [Daley, 2003; Baron, 2005; Keller, 2005] was performed at days 4–6 of EB differentiation by adding 1 � g/ml of DOXY to the medium.
Immunocytochemistry EBs at days 2–4 and 6 of differentiation were fixed in 3.7%
paraformaldehyde in phosphate-buffered saline (pH 7.2). Immu-nolabeling was performed in polyethylene glycol (Sigma) on 1- � m sections. This water/ethanol miscible medium, developed for electron microscopy [Wolosewick, 1980; Cidadão and David-Ferreira, 1986], allows very rapid infiltration and removal times,
avoids organic solvents like xylene, and is adaptable to light mi-croscopy and high-resolution immunocytochemistry [Mowery et al., 1989]. Sections were obtained with an ultramicrotome and attached to poly- L -lysine (Sigma)-covered glass slides. Such sec-tions correspond to a small fraction of the thickness of a cell, thereby providing excellent accessibility to antigens located in both cytoplasmic and nuclear subcellular compartments without need for detergent permeabilization. In addition, misinterpreta-tion of data due to tissue superimposition is minimized. Antige-nicity is usually better preserved than with paraffin embedding [Klosen et al., 1993]. Sections were washed in phosphate-buffered saline and then incubated for 10 min at room temperature in a blocking solution (5% bovine serum albumin in phosphate-buff-ered saline). Primary antibodies ( table 1 ) were diluted in phos-phate-buffered saline and incubated overnight at 4 ° C. Fluoro-chrome-conjugated secondary antibodies were incubated for 1 h at 37 ° C. Controls included omission of primary antibody or use of irrelevant antibody. Specificity of the Notch1 (Val1744) antibody (Cell Signaling; demonstrating S3 cleavage of the Notch1 recep-tor) was further confirmed in immunoblots. Moreover, immu-nostaining with Notch1 (Val1744) antibody and other antibodies was performed simultaneously on EBs derived from distinct ESC lines to minimize possible sources of artifactual staining inten-sity variation. Tissue sections were washed in phosphate-buff-ered saline, mounted in VectaShield (Vector Laboratories) con-taining 4 � ,6-diamidino-2-phenylindole, a nuclear counterstain, observed with a wide-field epifluorescence microscope (Leica DM5000B), and imaged with a refrigerated CCD camera (Leica). For single-labeling experiments, images of the irrelevant f luores-cence channel were always obtained to discriminate tissue auto-fluorescence.
Western Blots 293T cells and EBs at different days of development were lysed
in 50 m M Tris-HCl, pH 8.0, 150 m M NaCl, 5 m M EDTA, 1% (v/v) NP-40, 1 m M Na 3 VO 4 , 10 m M NaF, 10 m M NaPyroph, 1 m M 4-(2-aminoethyl) benzenesulfonyl fluoride hydrochloride), 10 � g/ml leupeptin, 10 � g/ml aprotinin, 1 � g/ml pepstatin. Equal amounts of protein (50 or 75 � g/sample) from 293T cells and day 2–6 EB extracts were resolved by 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis, transferred onto nitrocellu-lose membranes and immunoblotted overnight at 4 ° C with Notch1 (Val1744) or Notch11-IC (C20) antibodies ( table 1 ), followed by incubation with horseradish peroxidase-conjugated antibodies (Promega) for 1 h at room-temperature and detection by en-hanced chemiluminescence (Pierce). Densitometry analysis of Notch1 (Val1744) and � -actin was performed on 12-bit digitized im-ages using ImageJ software (gel analyzer module). Two indepen-dent Western blots for each ESC line-derived EBs were used for the densitometric analysis (bar graphs on fig. 2 j depict averaged results from those independent Western blots). A specificity con-trol for the Notch1 (Val1744) antibody reactivity was performed on 293T cells transfected with � E- or Notch IC domain (ICN)-trun-cated forms of Notch1, and is depicted on the top frame of figure 2 j. � E is a membrane-tethered form of the Notch1 receptor con-stitutively cleaved (i.e. independently of ligand) by � -secretase, at the canonic Val1744 S3 site to produce biologically active ICN. The � E construct was used as a positive control for Notch1 (Val1744) . Conversely, the ICN construct is a constitutively active, non-membrane-tethered form of Notch1 that lacks the epitope recog-

Alcobia /Gomes /Saavedra /Laranjeiro /Oliveira /Parreira /Cidadão
Cells Tissues Organs 4
nized by the Notch1 (Val1744) antibody, and thus was used as a neg-ative control. Both � E and ICN should be recognized by the Notch1-IC (C20) antibody, whose epitope maps to the PEST domain of Notch1 (information provided by Santa Cruz Biotechnology).
Results
EBs Undergo Spatial Reorganization between Days 4–6 of Differentiation To identify the cellular territories where Notch system
activation might occur, we performed a detailed immu-nocytochemical study of EB microenvironments.
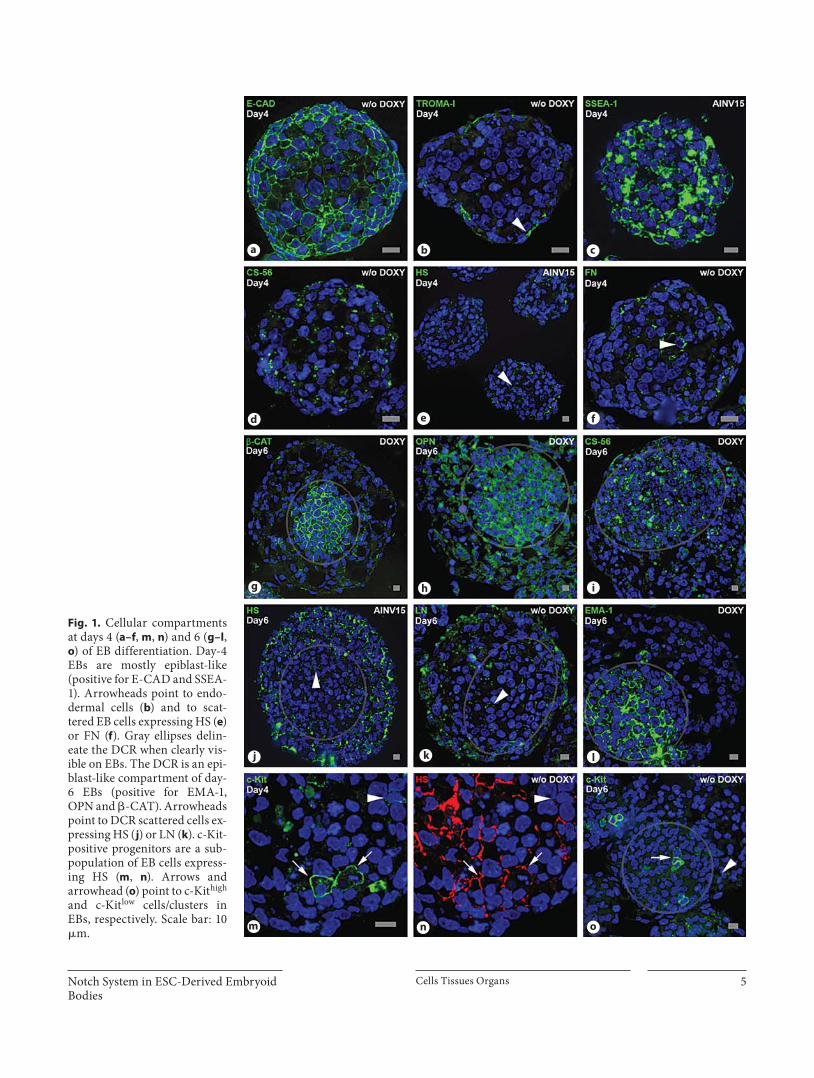
Day-4 EBs ( fig. 1 a–f, m, n) are compact and rather ho-mogeneous, with tightly packed cells globally expressing E-cadherin (E-CAD) ( fig. 1 a) and � -catenin ( � -CAT)
(online suppl. fig. 3a), and surrounded by flattened en-dodermal cells positive for cytokeratin Endo-A (TRO-MA-I) ( fig. 1 b). Day-4 EBs are also globally positive for osteopontin (OPN) (online suppl. fig. 3b), an important component of the hematopoietic niche in the adult [Stier et al., 2005], whose expression during early embryonic development is under the control of octamer-4 [Botquin et al., 1998], a well-known stemness marker. Day-4 EBs are highly positive for stage-specific embryonic anti-gen-1 (SSEA-1) ( fig. 1 c), as well as epithelial membrane antigen-1 (EMA-1) (online suppl. fig. 3c), two standard markers of immature cells. The immunolocalization patterns of 6 relevant EC matrix molecules were also as-sessed. Chondroitin sulfate (CS) and heparan sulfate (HS) are upregulated upon EB differentiation [Nairn et
Table 1. List of primary antibodies used in this study
Antibody (clone/ref. No.) Species/class Dilution Supplier Antigen retrieval
�-CAT (18-8226) mouse IgG 1:50 Zymed CBc-Kit (AF1356) goat 1:50 R&D Systems BA or T-EDTACS-56 mouse IgM 1:200 Sigma noneDll1+4 (C-20; sc-8155) goat 1:10 SCBT BA or noneDll4 (C-20; sc-18640) goat 1:10 SCBT BA or noneE-CAD mouse IgG 1:50 BD Pharmingen BAEMA-1 (germ cells) mouse IgM 1:50 DSHB BA or noneFN (F-3648) rabbit 1:100 Sigma noneHS (10E4; #370255) mouse IgM 1:100 Seikagaku noneHes-1 (H-140; sc-25392) rabbit 1:50 SCBT BA or T-EDTAJ1 (H-114; sc-8303) rabbit 1:50 SCBT BA or noneJ2 (R-19; sc-8158) goat 1:50 SCBT BA or noneLaminin rabbit 1:100 BRL noneLAMP-1 (1D4B) rat IgG 1:50 DSHB BA or noneNedd4 (611480) mouse IgG 1:100 BD Pharmingen BANotch1-EC(20–150) (H131; sc-9170) rabbit 1:50 SCBT BA or noneNotch1-EC(381–853) (07-218) rabbit 1:100 Upstate Biotech BA or T-EDTANotch1-IC(bTAN 20) (bTAN 20) rat IgG 1:200 DSHB BANotch1-IC(C20) (C-20; sc-6014) rabbit 1:100 SCBT BANotch1(Val1744) (Val 1744; N1-S3) rabbit 1:50 Cell Signaling T-EDTANotch2-IC (C651.6DbHN) rat IgG 1:100 DSHB BANotch4-IC (C-19; sc-8644) goat 1:50 SCBT BANumb (Cmnb-1) mouse IgG 1:10 DSHB BAOPN (MPIIIB10) mouse IgG 1:100 DSHB None, CB or BAPofut1 (H00023509-A01) mouse poly 1:50 Abnova BA or noneRab7 (H-50; sc-10767) rabbit 1:100 SCBT BARER (H-69) rat IgG 1:50 DSHB BASSEA-1 (Mc-480) mouse IgM 1:200 DSHB none, BA or T-EDTATROMA-I (SP2/0) rat IgG 1:100 DSHB BA
D SHB = Developmental Studies Hybridoma Bank, University of Iowa; SCBT = Santa Cruz Biotechnology; BA = boric acid, pH 7.0, microwave antigen retrieval; CB = citrate buffer, pH 6.0, microwave antigen retrieval; T-EDTA = Tris-EDTA, pH 8.0, microwave an-tigen retrieval. Whenever antigen retrieval was needed, one 15-min MW cycle, 800 W, was used.
Notch System in ESC-Derived Embryoid Bodies
Cells Tissues Organs 5
Fig. 1. Cellular compartments at days 4 ( a–f , m , n ) and 6 ( g–l , o ) of EB differentiation. Day-4 EBs are mostly epiblast-like (positive for E-CAD and SSEA-1). Arrowheads point to endo-dermal cells ( b ) and to scat-tered EB cells expressing HS ( e ) or FN ( f ). Gray ellipses delin-eate the DCR when clearly vis-ible on EBs. The DCR is an epi-blast-like compartment of day-6 EBs (positive for EMA-1, OPN and � -CAT). Arrowheads point to DCR scattered cells ex-pressing HS ( j ) or LN ( k ). c-Kit-positive progenitors are a sub-population of EB cells express-ing HS ( m , n ). Arrows and arrowhead ( o ) point to c-Kit high and c-Kit low cells/clusters in EBs, respectively. Scale bar: 10 � m.
a b c
d e f
g h i
j k l
m n o
Alcobia /Gomes /Saavedra /Laranjeiro /Oliveira /Parreira /Cidadão
Cells Tissues Organs 6
N1-IC(C20) Val1744
�E
�E
�E
2 3 4 6
2 3 4 6
2 3 4 6
�EICN
ICN
Tubulin
N1-IC(Val1744)
Val1744
�-Actin
Val1744
�-Actin
110 kDa
48 kDa
110 kDa
48 kDa
2.0
1.5
1.0
0.5
02 3 4 6
Ratio
N1-
S3/�
-act
in
2.0
1.5
1.0
0.5
0
Ratio
N1-
S3/�
-act
in
R1
AINV15
specificity controla b c
d e f
g h i
k l m n
j
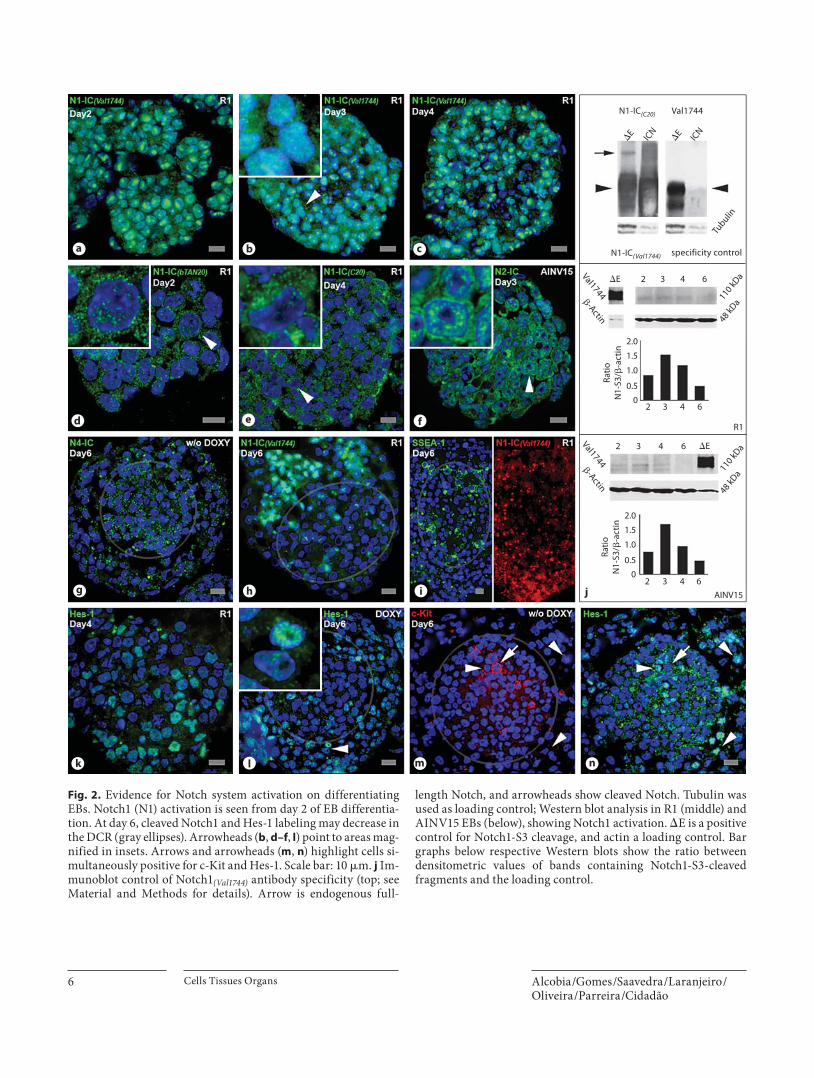
Fig. 2. Evidence for Notch system activation on differentiating EBs. Notch1 (N1) activation is seen from day 2 of EB differentia-tion. At day 6, cleaved Notch1 and Hes-1 labeling may decrease in the DCR (gray ellipses). Arrowheads ( b , d–f , l ) point to areas mag-nified in insets. Arrows and arrowheads ( m , n ) highlight cells si-multaneously positive for c-Kit and Hes-1. Scale bar: 10 � m. j Im-munoblot control of Notch1 (Val1744) antibody specificity (top; see Material and Methods for details). Arrow is endogenous full-
length Notch, and arrowheads show cleaved Notch. Tubulin was used as loading control; Western blot analysis in R1 (middle) and AINV15 EBs (below), showing Notch1 activation. � E is a positive control for Notch1-S3 cleavage, and actin a loading control. Bar graphs below respective Western blots show the ratio between densitometric values of bands containing Notch1-S3-cleaved fragments and the loading control.
Notch System in ESC-Derived Embryoid Bodies
Cells Tissues Organs 7
al., 2007]. Day-4 EBs are moderately reactive with CS-56 ( fig. 1 d), an antibody which recognizes D-type CS [Ito et al., 2005]. Some scattered cells in day-4 EBs express an HS epitope ( fig. 1 e, n) highly enriched in Flk1-positive mesodermal cells, from which hemangioblasts derive [Cumano and Godin, 2007; Baldwin et al., 2008]. A mi-nority of cells from day-4 EBs also label for fibronectin (FN) ( fig. 1 f) and laminin (LN) (online suppl. fig. 3d, ar-rowheads), EC molecules that, together with HS, are in-volved in ESC differentiation [Hayashi et al., 2007; Lan-ner et al., 2010].
Day-4 EBs undergo profound spatial reorganization which results, at day 6 ( fig. 1 g–l, o), in the segregation of
a densely packed cellular region (DCR) from a surround-ing mesoderm-like territory. The DCR is positive for � -CAT ( fig. 1 g), E-CAD (online suppl. fig. 3e), OPN ( fig. 1 h) and CS-56 ( fig. 1 i), whereas the mesodermal-like terri-tory stains intensely for HS ( fig. 1 j), LN ( fig. 1 k) and FN (online suppl. fig. 3f, arrowheads). Reminiscent of what happens at day 4, the DCR of day 6 EBs contains a few scattered HS ( fig. 1 j)-, LN ( fig. 1 k)- and FN-positive cells (online suppl. fig. 3f, arrowheads). EMA-1 ( fig. 1 l) and SSEA-1 ( fig. 2 i, below), known markers of cellular imma-turity, are highly expressed in the DCR and virtually ab-sent from adjacent mesodermal territories. As on day 4, a layer of TROMA-I-positive endodermal cells borders
a b c d
e f g h
i j k l
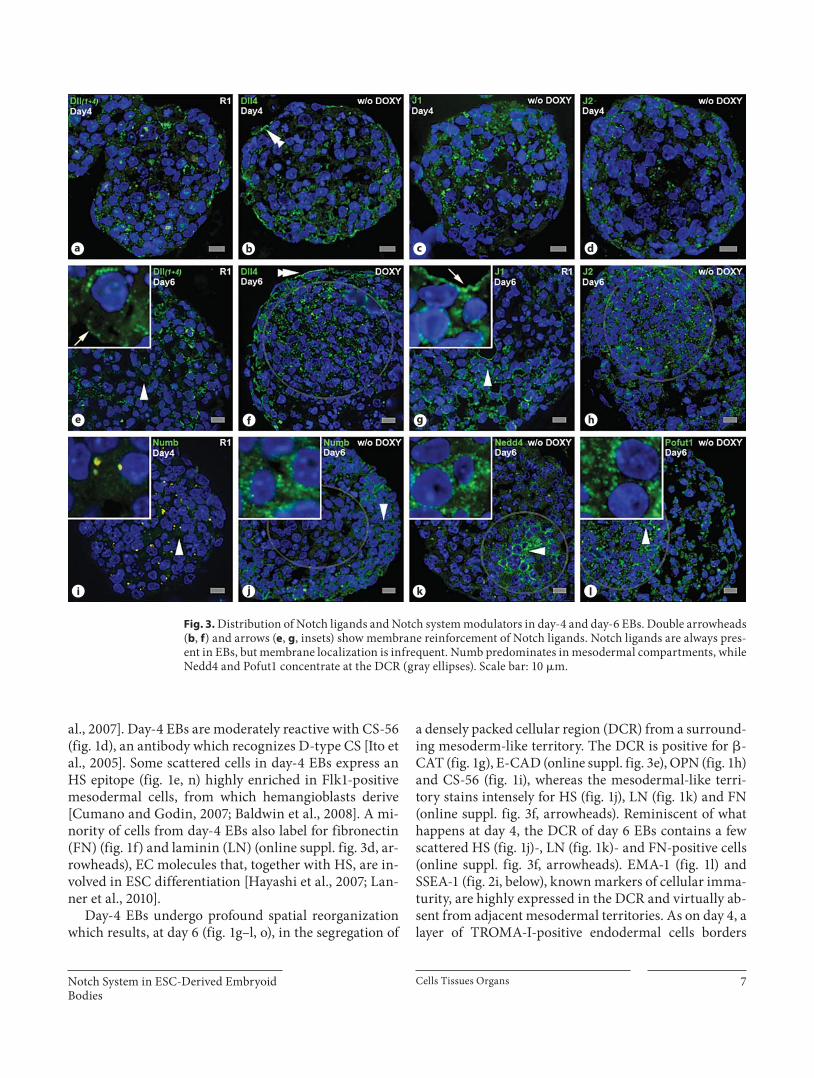
Fig. 3. Distribution of Notch ligands and Notch system modulators in day-4 and day-6 EBs. Double arrowheads ( b , f ) and arrows ( e , g , insets) show membrane reinforcement of Notch ligands. Notch ligands are always pres-ent in EBs, but membrane localization is infrequent. Numb predominates in mesodermal compartments, while Nedd4 and Pofut1 concentrate at the DCR (gray ellipses). Scale bar: 10 � m.
Alcobia /Gomes /Saavedra /Laranjeiro /Oliveira /Parreira /Cidadão
Cells Tissues Organs 8
day-6 EBs (online suppl. fig. 3e, arrowhead). c-Kit is ex-pressed in ESCs and is dynamically regulated throughout EB differentiation, where it may be useful to identify pre-cursors of various cell lineages, including hematopoietic precursors [Ling and Neben, 1997, and references there-in]. c-Kit-positive cells are present on day-4 ( fig. 1 m) and day-6 EBs ( fig. 1 o), either isolated or as clusters. Staining intensity for c-Kit is variable on these cells. On day-6 EBs, c-Kit high cells often concentrate in the DCR ( fig. 1 o). Both on day-4 (compare fig. 1 m, n) and day-6 EBs (online sup-pl. fig. 3g), c-Kit-positive cells are a subset of HS-positive cells. Since the HS antibody we used maps to an epitope
highly enriched in Flk1-positive cells [Baldwin et al., 2008], the c-Kit-positive cells that it detects may well be-long to such a cell population.
Together, our results highlight the profound territo-rial remodeling that occurs in developing EBs.
The Notch System Is Active throughout EB Differentiation Notch activation ( fig. 2 ) is detectable from day 2 of EB
differentiation as revealed by nuclear staining for Notch1 with an antibody (Notch1 (Val1744) ) that recognizes the ca-nonic Notch1 � -secretase-dependent S3 (Notch1-S3)
Ligand-binding (EGF repeats 11–12)
Plasma membrane
EC (2
0–15
0)
EGF repeatsEC (381–853)
S3(Val1744)
2220
–237
7 (b
TAN
20)
2410
–246
0 (C
20)
TAD P
b
c d e f
g h i j
a
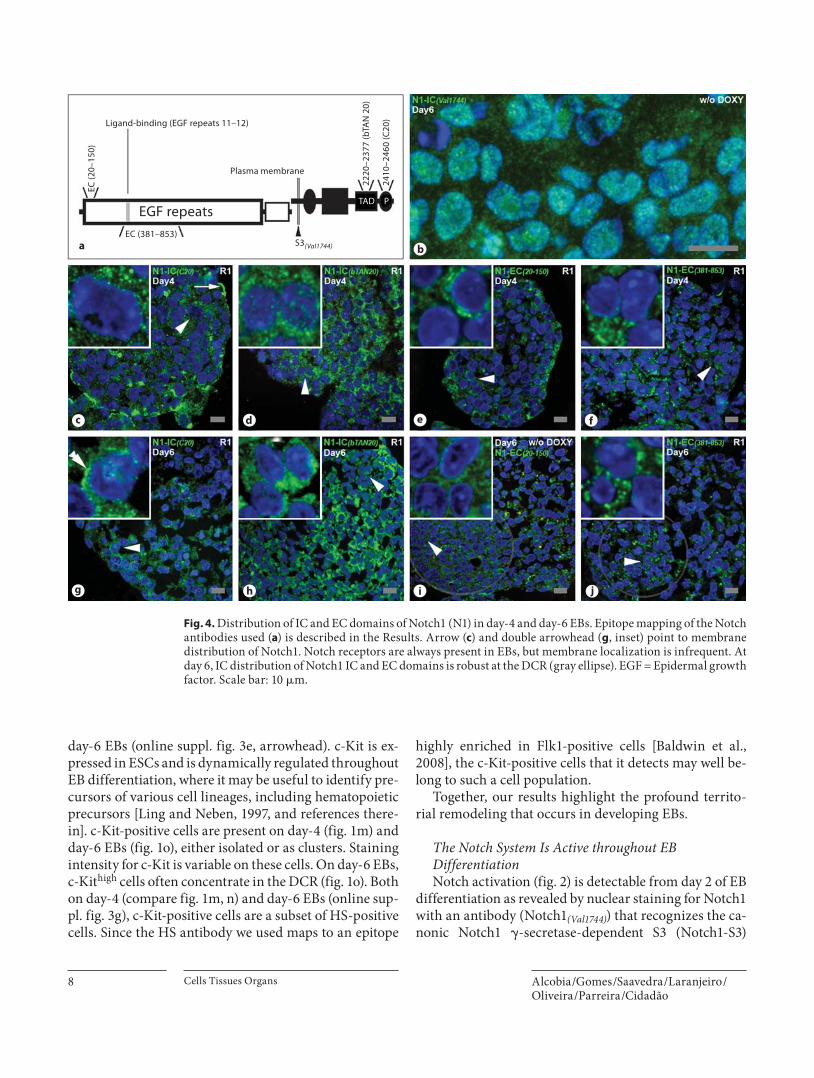
Fig. 4. Distribution of IC and EC domains of Notch1 (N1) in day-4 and day-6 EBs. Epitope mapping of the Notch antibodies used ( a ) is described in the Results. Arrow ( c ) and double arrowhead ( g , inset) point to membrane distribution of Notch1. Notch re ceptors are always present in EBs, but membrane localization is infrequent. At day 6, IC distribution of Notch1 IC and EC domains is robust at the DCR (gray ellipse). EGF = Epidermal growth factor. Scale bar: 10 � m.
Notch System in ESC-Derived Embryoid Bodies
Cells Tissues Organs 9
cleavage [Kopan and Ilagan, 2009]. Importantly, virtu-ally all nuclei from day-2 to day-4 EBs ( fig. 2 a–c) are strongly reactive with this antibody, suggesting that Notch1 signaling is globally active at this early period of EB differentiation. Notch1-S3 cleavage is also detected in Western blots of EB extracts from day 2 onwards, display-ing a transient upregulation at days 3–4 ( fig. 2 j; lines R1 and AINV15). Extensive nuclear staining was also evi-dent using two other antibodies reactive with the IC do-main of Notch1, namely Notch1-IC (bTAN20) , recognizing the transactivation domain [Zagouras et al., 1995] ( fig. 2 d), and Notch1-IC (C20) mapping to the PEST do-
main [Kopan and Ilagan, 2009; information provided by Santa Cruz Biotechnology] ( fig. 2 e). Strong nuclear stain-ing is also observed with an antibody against the IC do-main of Notch2 ( fig. 2 f). In contrast, Notch4-IC labeling is virtually absent from nuclei, and only observed in the cytoplasm as large bright foci ( fig. 2 g). Notch3 immuno-labeling was absent or inconclusive, and thus it is not in-cluded in this study.
The widespread Notch1-S3 staining found until day-4 changes on day-6 EBs, where some DCR cells exhibit faint nuclear reactivity with the Notch1 (Val1744) antibody ( fig. 2 h, i). While lacking the spatial information provid-
a b c d
e f g h
i j k l
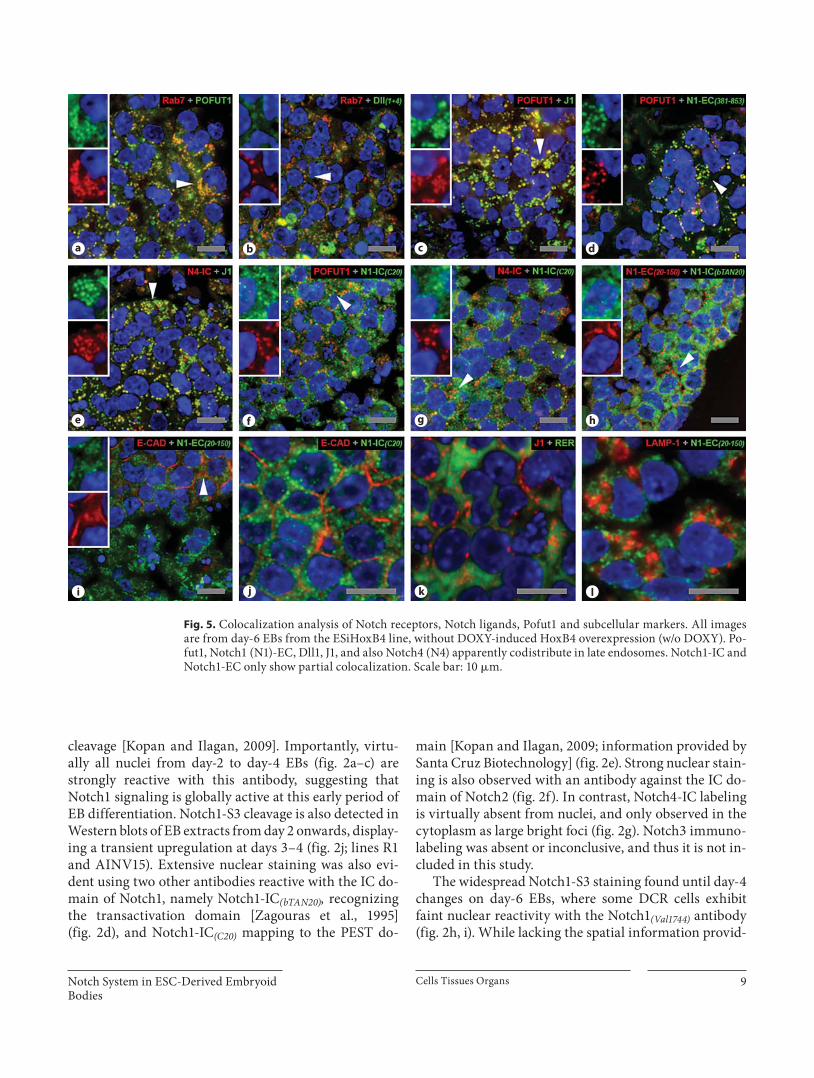
Fig. 5. Colocalization analysis of Notch receptors, Notch ligands, Pofut1 and subcellular markers. All images are from day-6 EBs from the ESiHoxB4 line, without DOXY-induced HoxB4 overexpression (w/o DOXY). Po-fut1, Notch1 (N1)-EC, Dll1, J1, and also Notch4 (N4) apparently codistribute in late endosomes. Notch1-IC and Notch1-EC only show partial colocalization. Scale bar: 10 � m.

Alcobia /Gomes /Saavedra /Laranjeiro /Oliveira /Parreira /Cidadão
Cells Tissues Organs 10
ed by in situ studies, lower Notch1-S3 reactivity is also seen in Western blots of day-6 EB extracts ( fig. 2 j, mid and bottom frames; lines R1 and AINV15).
The distribution pattern and the labeling intensity of Notch receptors on EBs derived from the ESiHoxB4 line are unchanged after HoxB4 induction, suggesting that the expansion of hematopoietic progenitors described to occur following HoxB4 expression [Daley, 2003; Baron, 2005; Keller, 2005 (online suppl. fig. 1)] is not accompa-nied by major immunocytochemical changes of Notch receptors in the context of the whole EB.
Hes-1 expression in undifferentiated ESCs seems to be controlled by leukemia inhibitory factor and bone mor-phogenetic protein, transiently suppressing Notch signal-ing, downregulating Notch ligands and delaying ESC dif-ferentiation. At later stages of ESC differentiation, Hes-1 apparently becomes a Notch target gene, leading to, and modulating, mesodermal differentiation [Kobayashi and Kageyama, 2010]. Hes-1 protein is immunodetected in both day-4 and day-6 EBs as a ‘salt-and-pepper’ pattern of unequally positive nuclei ( fig. 2 k, l, n). Similarly to the
lower Notch1-S3 immunoreactivity of the DCR with re-spect to the surrounding territories, Hes-1 is often de-tected therein with lower intensity ( fig. 2 l). c-Kit-positive cells in differentiating EBs (see above, and fig. 1 m, o) may show low ( fig. 2 m, n) or high levels of Hes-1 nuclear im-munoreactivity ( fig. 2 m, n). The latter, mostly c-Kit low , are often found outside the DCR. Globally, c-Kit-positive cells are also reactive with Notch1-IC antibodies (online suppl. fig. 3h, I, arrows), as expected from the generalized expression of Notch receptors in EBs (see below).
In short, we present morphological and biochemical evidence that Notch activation occurs in EBs harboring hematopoietic differentiation. The emerging pattern is that, up to day 4, Notch1-S3 cleavage is widespread in all EB cells, and that by day 6 it may be downregulated at the DCR.
Notch Ligands Are Mostly Expressed as Cytoplasmic Foci in Differentiating EBs To verify whether the territorial expression and/or
subcellular distribution of Notch ligands could be cor-related with specific germ layers (see Introduction and references therein) or with the lower Notch1-S3 immu-nolabeling sometimes exhibited by DCR cells of day-6 EBs, we performed in situ detection of Notch ligands Dll1, Dll4, J1 and J2. All antibodies used identify the IC domain of these molecules, but one of them recognizes an epitope common to Dll1 and Dll4 (Dll1+4). Thus, im-munoreactivity obtained with the Dll1+4 antibody was ascribed to Dll1 only when it was different from that originating from the Dll4 antibody. Dll3 was not ana-lyzed.
Notch ligands Dll1 ( fig. 3 a, e), Dll4 ( fig. 3 b, f), J1 ( fig. 3 c, g) and J2 ( fig. 3 d, h) are always present in EBs, mainly as bright cytoplasmic foci, particularly abundant at the DCR ( fig. 3 f, h). Dll1 and J1 cytoplasmic foci are usually larger and clustered around nuclei ( fig. 3 e, g; insets). Notch ligands are also detected as cell surface reinforce-ments, here interpreted as membrane localization, most-ly in EB cells which are not in close contact with a neigh-boring cell. Examples are the apical surface of endoder-mal cells (detected by the Dll4 antibody; fig. 3 b, f) and a few cells from mesodermal territories (detected by both Dll1 + 4 and by J1 antibodies; fig. 3 e, g).
HoxB4 induction during EB differentiation is known to upregulate Dll3 and J2 [Schiedlmeier et al., 2007] (on-line suppl. fig. 2). The former was not analyzed here, and we were not able to discriminate any immunocytochem-ical changes on J2 or the distribution of any other ligand, even though an increase in J2 mRNA upon HoxB4 induc-
Fig. 6. Interpretation of colocalization patterns found on EBs in the context of an active Notch system (see Discussion for details). ER = Endoplasmic reticulum; Nu = Nucleus; L-End = late endo-some; N = notch; N-EC = notch extracellular domain; N-TMD = notch transmembrane domain; NICD = notch intracellular do-main.

Notch System in ESC-Derived Embryoid Bodies
Cells Tissues Organs 11
tion has been detected by the real-time quantitative re-verse transcription polymerase chain reaction (online suppl. fig. 2).
In short, our results point to a strong and apparently maintained distribution of Notch ligands in differentiat-ing EBs, as cytoplasmic foci, mostly abundant in the DCR and less frequently as membrane reinforcement.
Notch Modulators Are Expressed in Differentiating EBs To further characterize the active Notch system pre-
vailing in EBs harboring YS-like hematopoiesis, namely the lower Notch1-S3 immunoreactivity that may be seen in the DCR of day-6 EBs, the expression of Notch modu-lators was analyzed.
Numb, a Notch signaling inhibitor relevant for hem-angioblast differentiation [Cheng et al., 2008] is weakly expressed in day-4 EBs ( fig. 3 i), thereafter becoming en-riched at the mesodermal compartment that surrounds the DCR in day-6 EBs ( fig. 3 j). Nedd4, another Notch signaling inhibitor [Sorensen and Conner, 2010], is also present in differentiating EBs. Yet, and in contrast to what occurs with Numb, it predominates in the DCR of day-6 EBs ( fig. 3 k). Therefore, the distinct distribution of Numb and Nedd4 within the EB precludes any straightforward correlation with the lower Notch1-S3 labeling that is observed at the DCR of day-6 EBs (see above). Nevertheless, the data suggest that Nedd4 may act preferentially in the more immature cells of the DCR. Pofut1, an essential modulator and chaperon for Notch receptor ligand recognition and Notch signaling for instance in mouse ESCs [Fiúza et al., 2007; Stahl et al., 2008; Kopan and Ilagan, 2009], was detected in dif-ferentiating EBs being highly expressed at the DCR at day 6 ( fig. 3 l).
In short, and as expected from an active Notch sys-tem, key Notch modulators are present in differentiat-ing EBs.
Cytoplasmic Portrayal of the Active Notch System Prevailing in Differentiating EBs As Notch1 signaling is particularly important during
embryonic development, especially for YS hematopoiesis (see Introduction and references therein), Notch1 was studied with a panel of five antibodies of known specific-ity ( fig. 4 ). Epitope mapping for these antibodies is de-picted in figure 4 a. Specifically, two antibodies recogniz-ing the Notch1 EC domain were used, one mapping to the C terminus (at amino acids 20–150; EC (20–150) ), the other with an epitope that spans the epidermal growth factor-like repeats 11–12 that are crucial for ligand binding
(amino acids 381–853; EC (381–853) ). Canonic � -secretase Notch1-S3 cleavage at the conserved valine 1744 was also assessed (S3 (Val1744) ). Notch1 IC domain was probed with two antibodies, one mapping to the transactivation do-main at amino acids 2220–2377 (IC (bTAN20) ), and the oth-er to the C-terminal PEST domain at amino acids 2410–2460 (information kindly provided by Santa Cruz Bio-technology; Notch1-IC (C20) ). Also, due to the relevance of the endocytic modulation of Notch signaling (see Intro-duction and references therein), markers of subcellular compartments like rough endoplasmic reticulum (RER), late endosomes (Rab7-positive foci) and lysosomes (LAMP-1) ( fig. 5 ) were used together with antibodies against Notch receptors, Notch ligands, and Pofut1 (dou-ble-labeling experiments) to probe for the existence of subcellular codistributions. E-CAD was used as a mem-brane marker.
Besides its nuclear localization ( fig. 2 ), S3-cleaved Notch1 ( fig. 4 a) is also detected as small cytoplasmic foci ( fig. 4 b), supporting the hypothesis that endocytic vesi-cles may be involved in � -secretase-dependent Notch ac-tivation [Kopan and Ilagan, 2009]. Small cytoplasmic foci are also elicited by the Notch1-IC (C20) antibody ( fig. 4 a, c, g). Membrane staining is rare, except for the apical domain of some TROMA-1-positive endodermal cells ( fig. 4 c) and a few scattered cells in mesodermal ter-ritories ( fig. 4 g). Such infrequent membrane staining is reminiscent of what was found for Notch ligands (see above; fig. 3 ) and, together with the robust cytoplasmic distribution of both Notch receptors and ligands, is sug-gestive of an active Notch system [Kopan and Ilagan, 2009]. Apical distribution of Notch receptors has already been described for other epithelia [Sasaki et al., 2007; Tanaka et al., 2007]. Notch1 membrane reinforcement is never seen with the Notch1-IC (bTAN 20) antibody ( fig. 4 a, d, h) which, otherwise, generally behaves as the Notch1-IC (C20) antibody. The reason for this different behavior was not addressed. A possibility is that steric hindrance of the Notch1-IC (bTAN 20) epitope, which maps to the transactivation domain [Zagouras et al., 1995], might oc-cur in Notch1 molecules located at/near the cell surface.
With respect to antibodies recognizing the N1 EC do-main, Notch1-EC (20–150) ( fig. 4 a, e, i) produces a RER-like pattern, without any nuclear labeling, as expected, and also lacking membrane reinforcement, especially in endodermal cells where Notch1-IC (C20) demonstrates it. Globally, membrane reinforcement in EB cells is rare with the Notch1-EC (20–150) antibody. The same occurs with the Notch1-EC (381–853) antibody ( fig. 4 a, f, j), which maps to epidermal growth factor-like repeats, crucial for

Alcobia /Gomes /Saavedra /Laranjeiro /Oliveira /Parreira /Cidadão
Cells Tissues Organs 12
ligand binding [Ge et al., 2008]. As suggested for the Notch1-IC (bTAN20) antibody, perhaps steric hindrance in Notch molecules located at the cell surface impedes its proper detection in fixed samples. Notch1 has been iden-tified at the cell membrane of ESCs and EB cells by flow cytometry in live cells [Ge et al., 2008; Hadland et al., 2004; Stahl et al., 2008].
The morphology and cellular distribution of Rab7-positive cytoplasmic spots (here interpreted as late endo-somes) resembles that of large foci reactive for Pofut1, Dll1+4, J1, Notch1-EC (381–853) and also Notch4-IC ( fig. 2 , 3 ). This suggests territorial colocalization, which was as-sessed and confirmed with double-labeling experiments ( fig. 5 ). Large and bright Pofut1 cytoplasmic foci show extensive colocalization with Rab7-positive late endo-somes ( fig. 5 a), the same occurring for Dll1+4 ( fig. 5 b). J1 shows widespread overlap with Pofut1-positive foci ( fig. 5 c), indirect evidence that it concentrates in late en-dosomes (antibody species-related issues prevented Rab7-J1 double labeling). Similar evidence exists for Notch1-EC, as recognized by the Notch1-EC (381–853) antibody ( fig. 5 d), and for N4-IC ( fig. 5 e), since both molecules co-localize with Rab7, Pofut1 and J1, respectively. In contrast to the extensive overlap found to occur between Pofut1 and Notch1-EC (381–853) , Pofut1 and Notch1-IC (C20) are only partially colocalized ( fig. 5 f), the same occurring with Notch1-IC (C20) and Notch4-IC ( fig. 5 g). As Pofut1 binds to the EC domain of Notch, the partial overlap be-tween Pofut1 and Notch1-IC suggests some degree of cy-toplasmic segregation between IC and EC Notch1 do-mains, which was indeed observed ( fig. 5 h). Globally, these results seem to be compatible with an active Notch system (see Introduction and references therein), whose dynamics would perhaps explain why both Notch1-EC and Notch1-IC are mostly found inside the cell and not at the membrane, as shown by double labeling with E-CAD ( fig. 5 i, j). The small cytoplasmic puncta of Notch1-IC ( fig. 5 j) differ considerably from the immunolabeling pat-tern typical of RER ( fig. 5 k), late endosomes ( fig. 5 a) as well as of lysosomes ( fig. 5 l). In contrast, the IC distribu-tion of Notch4-IC ( fig. 5 e) resembles that of late endo-somes. The virtual absence of Notch4 nuclear staining in EBs (see above) together with extensive colocalization with late endosomes apparently suggests that Notch4 may be largely targeted to endocytosis without proteo-lytic activation.
In short, the observed complex IC distribution pat-terns of Notch receptors and Notch ligands are consistent with current models of Notch activation and endocytic modulation of Notch signaling.
Discussion
We show that Notch activation is an event that occurs throughout EB differentiation, as EB microenviron-ments arise and remodel with time. Notch activation is robust and widespread in early EB development, becom-ing territorially attenuated at later stages. While our data were obtained from EBs harboring YS-like hematopoie-sis, the fact that Notch signaling is known to influence other cell lineages, both in vivo and in vitro (see Intro-duction and references therein), makes it plausible that our findings may also apply to other EB differentiation protocols.
EBs display self-organizing properties which enable them to establish polarity and develop primitive streak domains, where an epithelial-to-mesenchymal transi-tion occurs under the influence of Wnt signaling [Berge et al., 2008]. EB differentiation thus emulates early em-bryonic development, supporting its use as a standard in vitro model of YS hematopoiesis, and validating func-tional assessments of Notch signaling at that develop-mental window (see Introduction and references there-in).
According to the model of Notch/Wnt functional inte-gration [Hayward et al., 2008], Wnt signaling stimulates the expression of Notch ligands, while the resulting Notch signaling activates Wnt antagonists to reach a dynamic equilibrium. Berge et al. [2008] suggested that a nonauto-nomous cellular process might control the expansion of the Wnt-positive primitive-streak domain of differentiat-ing EBs. Our finding that Notch activation is global in early EBs, becoming territorially downregulated later on, raises the possibility that Notch signaling may be involved in such a phenomenon, not as an ‘essential’ pathway but as a territorial modulator. This hypothesis would fit the reported influence of Notch on germ layer differentiation [Yu et al., 2008; Croce and McClay, 2010; Ramasamy and Lenka, 2010; Revinski et al., 2010], as well as data showing that Notch signaling is dispensable for mouse early em-bryonic development [Shi and Stanley, 2006].
Our results agree with the reported canonical Notch1 signaling during early mouse development though EBs apparently exhibit an earlier and more generalized Notch1 activation than observed in vivo [Hadland et al., 2004; Del Monte et al., 2007]. Thus, while Notch is un-doubtedly active in the mouse EB model, the precise bio-logical significance of Notch signaling at this develop-mental stage must still be considered an open question. For instance, the Notch system was reported to be inac-tive in undifferentiated human ESCs [Noggle et al., 2006;

Notch System in ESC-Derived Embryoid Bodies
Cells Tissues Organs 13
Yu et al., 2008], while our data and other published data [Ramasamy and Lenka, 2010] demonstrate the opposite in the mouse. Also, as mouse ESCs express an isoform of the Notch coactivator RBP-Jk that is downregulated up-on differentiation [Li and Leder, 2007], roles ascribed to Notch at early developmental stages may differ from those occurring later or in the adult. Moreover, as Notch integrates chromatin-remodeling complexes which in-teract with key molecules like RBP-Jk [Kadam and Em-erson, 2003; Das et al., 2007; Takeuchi et al., 2007], chro-matin conformations related to ‘stemness’ may impose constraints on which are the major Notch target genes at a given time point.
Together, our results point to the existence of robust Notch activation at the microenvironments that arise and remodel with EB differentiation and show that Notch activation therein is an early and global event. In EBs exhibiting such an active system, Notch receptors and ligands do not accumulate extensively at cell sur-faces, but instead (co)localize at cytoplasmic foci, an ob-servation that fits current models of endocytic modula-tion of Notch signaling [Bray, 2006; Fiúza and Arias, 2007; Hurlbut et al., 2007; Sorensen and Conner, 2010]. As depicted in figure 6 , our data are compatible with a codistribution of Notch receptors, Notch ligands and Po-fut1, in the endoplasmic reticulum (1), and with a tran-sient expression of Notch receptors/ligands at the plasma membrane (2) until interaction with Notch ligands/re-ceptors present on adjacent cells eventually occurs (3; e.g. infrequent membrane reinforcement seen with Notch1-IC (C20) , Dll1 or J1 antibodies). Our results also suggest that � -secretase-dependent S3 cleavage of Notch1, most likely a plasma membrane event, may as well occur in small endocytic vesicles (4). As expected, we found ex-tensive immunoreactivity for NICD in the nucleus (5),
where it transiently induces the expression of Notch tar-get genes and is eventually degraded (6). Our data fur-ther point out that late endosomes are organelles where Pofut1, Notch EC domain and Notch ligands codistrib-ute, possibly following postactivation endocytosis (7; e.g. Notch1-EC, Dll1, J1) and/or internalization-mediated downregulation of nonactivated Notch receptors (8, e.g. Notch4). While the above results apply to all EB cells, we did observe higher cytoplasmic immunoreactivity for Notch receptors/ligands, for the Notch modulator Po-fut1, and for the Notch signaling inhibitor Nedd4, in the DCR of day-6 EBs. Conversely, we often found evidence of decreased N1 activation in the DCR (lower reactivity with the Notch1 (Val1744) antibody). Perhaps Nedd4 has a major role in downregulation of Notch signaling [So-rensen and Conner, 2010]. In short, although our results cannot be directly linked to specific Notch functions during EB differentiation, they point to a physiologically important role of Notch as a territorial modulator during early development.
Acknowledgments
ES cell lines R1 and AINV15 were provided by Prof. A. Duarte (Faculdade de Medicina Veterinária, Universidade Nova de Lis-boa, Portugal) and ESiHoxB4, by Prof. G. Daley (Harvard Medi-cal School, Boston, Mass., USA). The monoclonal antibodies EMA-1 (germ cells), LAMP-1 (1D4B), Notch1 (bTAN20) , Notch2 (C651.6DbHN), Numb (Cmnb-1), OPN (MPIIIB10), RER (H-69), SSEA-1 (Mc480) and TROMA-I (SP2/0) were obtained from the Developmental Studies Hybridoma Bank developed under the auspices of the NICHD and maintained by The University of Iowa, Department of Biological Sciences, Iowa City, USA. This work was supported by Fundação para a Ciência e Tecnologia, Portugal (grant PTDC/SAU-OBD/64165/2006).
References
Baldwin, R.J., G.B. Ten Dam, T.H. van Kuppe-velt, G. Lacaud, J.T. Gallagher, V. Kouskoff, C.L. Merry (2008) A developmentally regu-lated heparan sulfate epitope defines a sub-population with increased blood potential during mesodermal differentiation. Stem Cells 26: 3108–3118.
Baron, M.H. (2005) Developmental Hematopoi-esis: Methods and Protocols. Totowa, Huma-na Press, p 105.
Berge, D., W. Koole, C. Fuerer, M. Fish, E. Ero-glu, R. Nusse (2008) Wnt signaling mediates self-organization and axis formation in em-bryoid bodies. Cell Stem Cell 3: 508–518.
Botquin, V., H. Hess, G. Fuhrmann, C. Anas-tassiadis, M.K. Gross, G. Vriend, H.R. Schoeler (1998) New POU dimer configura-tion mediates antagonistic control of an os-teopontin preimplantation enhancer by Oct-4 and Sox-2. Genes Dev 12: 2073–2090.
Bray, S.J. (2006) Notch signaling: a simple path-way becomes complex. Nat Rev Mol Cell Biol 7: 678–689.
Cheng, X., T.L. Huber, V.C. Chen, P. Gadue, G.M. Keller (2008) Numb mediates the interaction between Wnt and Notch to modulate primi-tive erythropoietic specification from the he-mangioblast. Development 135: 3447–3458.
Chiba, S. (2006) Notch Signaling in Stem Cell Systems. Stem Cells 24: 2437–2447.
Cidadão, A.J., J.F. David-Ferreira (1986) A meth-od for TEM visualization of the extracellular matrix three-dimensional organization in tissues. J Microsc 142: 49–62.
Croce, J.C., D.R. McClay (2010) Dynamics of Delta/Notch signaling on endomesoderm segregation in the sea urchin embryo. Devel-opment 137: 83–91.
Cumano, A., I. Godin (2007) Ontogeny of the he-matopoietic system. Annu Rev Immunol 25: 745–785.

Alcobia /Gomes /Saavedra /Laranjeiro /Oliveira /Parreira /Cidadão
Cells Tissues Organs 14
Daley, G.Q. (2003) From embryos to embryoid bodies. Generating blood from embryonic stem cells. Ann NY Acad Sci 996: 122–131.
Das, A.V., J. James, S. Bhattacharya, A.N. Imbal-zano, M.L. Antony, G. Hedge, X. Zhao, K. Mallya, F. Ahmad, E. Knudsen, I. Ahmad (2007) SWI/SNF chromatin remodeling ATPase Brm regulates the differentiation of early retinal stem cells progenitors by influ-encing Brn3b expression and Notch signal-ing. J Biol Chem 282: 35187–35201.
Del Monte, G., J. Grego-Bessa, A. González-Ra-jal, V. Bolós, J.L. De La Pompa (2007) Moni-toring Notch1 activity in development: evi-dence for a feedback regulatory loop. Dev Dyn 236: 2594–2614.
Fiúza, U.M., A.M. Arias (2007) Cell and molecu-lar biology of Notch. J Endocrinol 194: 459–474.
Ge, C., T. Liu, X. Hou, P. Stanley (2008) In vivo consequences of deleting EGF repeats 8–12 including the ligand binding domain of mouse Notch1. BMC Dev Biol 8: 48.
Gering, M., R. Patient (2010) Notch signalling and haematopoietic stem cell formation dur-ing embryogenesis. J Cell Physiol 222: 11–16.
Hadland, B.K., S.S. Huppert, J. Kanungo, Y. Xue, R. Jiang, T. Gridley, R.A. Conlon, A.M. Cheng, R. Kopan, G.D. Longmore (2004) A requirement for Notch1 distinguishes 2 phases of definitive hematopoiesis. Blood 104: 3097–3105.
Hayashi, Y., M.K. Furue, T. Okamoto, K. Ohnu-ma, Y. Myoishi, Y. Fukuhara, T. Abe, J.D. Sato, R. Hata, M. Asashima (2007) Integrins regulate mouse embryonic stem cell self-re-newal. Stem Cells 25: 3005–3015.
Hayward, P., T. Kalmar, A. Martinez, A. Arias (2008) Wnt/Notch signaling and informa-tion processing during development. Devel-opment 135: 411–424.
Hurlbut, G., M.W. Kankel, R.J. Lake, S. Artava-nis-Tsakonas (2007) Crossing paths with Notch in the hyper-network. Curr Opin Cell Biol 19: 1–10.
Ito, Y., M. Hikino, Y. Yajima, T. Mikami, S. Sirko, A. von Holst, A. Faissner, S. Fukui, K. Suga-hara (2005) Structural characterization of the epitopes of the monoclonal antibodies 473HD, CS-56, and MO-225 specific for chondroitin sulfate D-type using the oligo-saccharide library. Glycobiology 15: 593–603.
Jaleco, A.C., H. Neves, E. Hooijberg, P. Gameiro, N. Clode, M. Haury, D. Henrique, L. Parreira (2001) Differential effects of Notch ligands Delta-1 and Jagged-1 in human lymphoid differentiation. J Exp Med 194: 991–1002.
Kadam, S., B.M. Emerson (2003) Transcription-al specificity of human SWI/SNF BRG1 and BRM chromatin remodeling complexes. Moll Cell 11: 377–389.
Keller, G. (2005) Embryonic stem cell differen-tiation: emergence of a new era in biology and medicine. Genes Dev 19: 1129–1155.
Klosen, P., X. Maessen, B.P. Aguilar (1993) PEG embedding for immuno-cytochemistry: ap-plication to the analysis of immunoreactivity loss during histological processing. J Histo-chem Cytochem 41: 455–463.
Kobayashi, T., R. Kageyama (2010) Hes1 regu-lates embryonic stem cell differentiation by suppressing Notch signaling. Genes Cells 15: 689–698.
Kopan, R., M.X.G. Ilagan (2009) The canonical Notch signaling pathway: unfolding the ac-tivation mechanism. Cell 137: 216–232.
Kyba, M., R.C. Perlingeiro, G.Q. Daley (2003) Development of hematopoietic repopulating cells from embryonic stem cells. Methods Enzymol 365: 114–129.
Lanner, F., K.L. Lee, M. Sohl, K. Holmborn, H. Yang, J. Wilbertz, L. Poellinger, J. Rossant, F. Farnebo (2010) Heparan sulfation-depen-dent fibroblast growth factor signaling maintains embryonic stem cells primed for differentiation in a heterogeneous state. Stem Cells 28: 191–200.
Li, X., P. Leder (2007) Identifying genes prefer-entially expressed in undifferentiated em-bryonic stem cells. BMC Cell Biol 8: 37.
Ling, V., S. Neben (1997) In vitro differentiation of embryonic stem cells: immunophenotypic analysis of cultured embryoid bodies. J Cell Physiol 171: 104–115.
Mowery, J., J. Chesner, S. Spangenberger, D.C. Hixson (1989) Rapid low molecular weight polyethylene glycol embedding protocol for immunocytochemistry. J Histochem Cyto-chem 37: 1549–1552.
Murry, C.E., G. Keller (2008) Differentiation of embryonic stem cells to clinically relevant populations: lessons from embryonic devel-opment. Cell 132: 661–680.
Nairn, A.V., A. Kinoshita-Toyoda, H. Toyoda, J. Xie, K. Harris, S. Dalton, M. Kulik, J.M. Pierce, T. Toida, K.W. Moremen, R.J. Lin-hardt (2007) Glycomics of proteoglycan bio-synthesis in murine embryonic stem cell dif-ferentiation. J Proteome Res 6: 4374–4387.
Neves, H., F. Weerkamp, A.C. Gomes, B.A. Naber, P. Gameiro, J.D. Becker, P. Lúcio, N. Clode, J.J. van Dongen, F.J. Staal, L. Parreira (2006) Effects of Delta1 and Jagged1 on early human hematopoiesis: correlation with ex-pression of Notch signaling-related genes in CD34+ cells. Stem Cells 24: 1328–1337.
Noggle, S.A., D. Weiler, B.G. Condie (2006) Notch signaling is inactive but inducible in human embryonic stem cells. Stem Cells 24: 1646–1653.
Orkin, S.H., L.I. Zon (2008) Hematopoiesis: an evolving paradigm for stem cell biology. Cell 132: 631–644.
Ramasamy, S.K., N. Lenka (2010) Notch exhibits ligand bias and manoeuvres stage specific steering of neural differentiation in ESCs. Mol Cell Biol 30: 1946–1957.
Revinski, D.R., A.R. Paganelli, A.E. Carrasco, S.L. López (2010) Delta-Notch signalingis involved in the segregation of the three germ layers in Xenopus laevis. Dev Biol 339: 477–492.
Sasaki, N., T. Sasamura, H.O. Ishikawa, M. Ka-nai, R. Ueda, K. Saigo, K. Matsuno (2007) Po-larized exocytosis and transcytosis of Notch during its apical localization in Drosophila epithelial cells. Genes Cells 12: 89–103.
Schiedlmeier, B., A.C. Santos, A. Ribeiro, N. Moncaut, D. Lesinski, H. Auer, K. Korn-acker, W. Ostertag, C. Baum, M. Mallo, H. Klump (2007) HOXB4’s road map to stem cell expansion. Proc Natl Acad Sci USA 104: 16952–16957.
Shi, S., P. Stanley (2006) Evolutionary origins of Notch signaling in early development. Cell Cycle 5: 274–278.
Sorensen, E.B., S.D. Conner (2010) Gamma secretase-dependent cleavage initiates Notch signaling from the plasma membrane. Traf-fic 11: 1234–1245.
Stahl, M., K. Uemura, C. Ge, S. Shi, Y. Tashima, P. Stanley (2008) Roles of Pofut1 and O -fu-cose in mammalian Notch signaling. J Biol Chem 283: 13638–13651.
Stier, S., Y. Ko, R. Forkert, C. Lutz, T. Neuhaus, E. Grünewald, T. Cheng, D. Dombkowski, L.M. Calvi, S.R. Rittling, D.T. Scadden (2005) Osteopontin is a hematopoietic stem cell niche component that negatively regu-lates stem cell pool size. J Exp Med 201: 1781–1791.
Takeuchi, J.K., H. Lickert, B.W. Bisgrove, X. Sun, M. Yamamoto, K. Chawengsaksophak, H. Hamada, H.J. Yost, J. Rossant, B.G. Bruneau (2007) Baf60c is a nuclear Notch signaling component required for the establishment of left-right asymmetry. Proc Natl Acad Sci USA 104: 846–851.
Tanaka, M., M. Kokubo, T. Marunouchi (2007) Asymmetric localization of Notch2 on the microvillous surface of choroid plexus epi-thelial cells. Histochem Cell Biol 127: 449–456.
Wolosewick, J.J. (1980) The application of poly-ethylene glycol (PEG) to electron microsco-py. J Cell Biol 86: 661–675.
Yu, X., J. Zou, Z. Ye, H. Hammond, G. Chen,A. Tokunaga, P. Mali, Y.M. Li, C. Civin, N. Gaiano, L. Cheng (2008) Notch signalingactivation in human embryonic stem cellsis required for embryonic, but not tropho-blastic, lineage commitment. Cell Stem Cell 2: 461–471.
Zagouras, P., S. Stifani, C.M. Blaumueller, M.L. Carcangiu, S. Artavanis-Tsakonas (1995) Al-terations of Notch signaling in neoplastic le-sions of the human cervix. Proc Natl Acad Sci USA 92: 6414–6418.
Related Documents











