K. Erzini, L. Bentes, P.G. Lino, J. Ribeiro, R. Coelho, P. Monteiro, C. Correia and J.M.S. Gonçalves: Universidade do Algarve, CCMAR/FCMA Campus de Gambelas, 8000 Faro, Portugal [tel: +351 289 800 900, fax: +351 289 818 353, e-mail: [email protected]] ICES CM 2001/J:16 Population structure of the Dover sole, Solea solea L., in the Atlantic using randomly amplified polymorphic DNA (PCR-RAPD) marking Athanasios K. Exadactylos, Andreas Y. Troumbis, Audrey J. Geffen, and John P. Thorpe The primary objective of this paper is to compare patterns of allozyme and RAPD variation in six Atlantic populations of Solea solea. Sixteen individuals were randomly selected from each of the six Atlantic populations of S. solea sampled during 1994 and 1995. In total, 96 individuals were screened from three Irish Sea, two North Sea and one Bay of Biscay populations. Alllozyme and RAPD markers produced results which were in general agreement with each other. However, RAPD markers detected more variation in the sampled sole populations than that observed by using allozyme markers. The percentage of polymorphism and genetic diversity estimates were very high. There was clearly no geographic trend to any RAPD or allozyme allele frequencies at all polymorphic loci. The chi-square contingency analyses (allele frequencies and hierarchical F ST and non-hierarchical F DT ) indicated a highly significant heterogeneity among populations and between groupings. This conclusion is consistent with the life history of Dover sole, the homing behaviour they exhibit and the discrete spawning areas. A north-to-south pattern of significant population differentiation in the allozyme study was replaced by a northwest-to-southeast significant variation. Natural selection for differences in spawning habitat may explain the genetic isolation between these groupings. Another reason could be the significant absence of some alleles, present in the southern European populations. Evidence in favour of the divergence between these regions is shown by the cluster analyses (UPGMA, Wagner procedure) of both allozyme and RAPD data. However, allozymes provided a slightly better fit of data to predictions about genetic distance (higher values of cophenetic correlation of clusters from the “goodness-of-fit” statistics) and better correlation between genetic and geographic distances (Mantel’s r). In both studies a strong similarity between the Bay of Biscay and the German Bight populations was evident, judging from their clustering positions. It was apparent that the trend of probable movement of migrants through the English Channel suggested by the allozyme study was supported using RAPD markers. The apparent absence of isolation-by-distance (non-significant correlation of genetic with geographic distance) in both studies suggests that S. solea may not yet be at genetic equilibrium. Overall, S. solea conforms to the type of species with either a broad variety of geographic and ecological habitats, or large and stable population sizes, or substantial gene flow at a regional scale. A. Exadactylos: Biodiversity Conservation Laboratory, Department of Environmental Studies, University of the Aegean, Mytilene, GR81100, Hellas [tel: +0251–36247, e-mail: [email protected]]. A.Y. Troumbis: Biodiversity Conservation Laboratory, Department of Environmental Studies, University of the Aegean, Mytilene, GR81100, Hellas [tel: +0251–36230, e-mail: [email protected]]. J.P. Thorpe: Port Erin Marine Laboratory, School of Biological Sciences, The University of Liverpool, Port Erin, Isle of Man, IM9 6JA, UK [tel: +01624–831010, e-mail: J.P.Thorpe@ liverpool.ac.uk] ICES CM 2001/J:17 – WITHDRAWN ICES CM 2001/J:18 Automated fecundity and egg-sizing for marine species from scanned images: rapid sample processing and large data volumes Kevin Friedland, D. Ama-Abasi, L. Clarke, D. Sigourney, and A. Stein One of the most fundamental parameters in fisheries management and ecology is the population production of eggs. Management decisions are largely based on the dynamics of spawning stock biomass, and it is essential to know the contribution of viable gametes by age, which is affected at younger ages by growth and maturity rates and at older ages by the physiology of reproductive senescence. In addition to population modelling, recruitment dynamics are also strongly influenced by the size and quality of eggs. Egg quality can be characterized by a number of different qualitative and quantitative approaches; however, the most attractive methods developed to date rely on techniques that are efficient and lend themselves to wide application. The measurement of fecundity and egg size for fish and invertebrate fishery resources has been plagued by the cumbersome nature of the traditional approaches. We have developed an imaging-based technique that allows for the reduction of samples to electronic counts in relative little time and with a favourable costing. The technique is dependant upon high resolution scans of plated egg samples and the 7

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

K. Erzini, L. Bentes, P.G. Lino, J. Ribeiro, R. Coelho, P. Monteiro, C. Correia and J.M.S. Gonçalves: Universidade do Algarve, CCMAR/FCMA Campus de Gambelas, 8000 Faro, Portugal [tel: +351 289 800 900, fax: +351 289 818 353, e-mail: [email protected]]
ICES CM 2001/J:16
Population structure of the Dover sole, Solea solea L., in the Atlantic using randomly amplified polymorphic DNA (PCR-RAPD) marking
Athanasios K. Exadactylos, Andreas Y. Troumbis, Audrey J. Geffen, and John P. Thorpe
The primary objective of this paper is to compare patterns of allozyme and RAPD variation in six Atlantic populations of Solea solea. Sixteen individuals were randomly selected from each of the six Atlantic populations of S. solea sampled during 1994 and 1995. In total, 96 individuals were screened from three Irish Sea, two North Sea and one Bay of Biscay populations. Alllozyme and RAPD markers produced results which were in general agreement with each other. However, RAPD markers detected more variation in the sampled sole populations than that observed by using allozyme markers. The percentage of polymorphism and genetic diversity estimates were very high. There was clearly no geographic trend to any RAPD or allozyme allele frequencies at all polymorphic loci. The chi-square contingency analyses (allele frequencies and hierarchical FST and non-hierarchical FDT) indicated a highly significant heterogeneity among populations and between groupings. This conclusion is consistent with the life history of Dover sole, the homing behaviour they exhibit and the discrete spawning areas. A north-to-south pattern of significant population differentiation in the allozyme study was replaced by a northwest-to-southeast significant variation. Natural selection for differences in spawning habitat may explain the genetic isolation between these groupings. Another reason could be the significant absence of some alleles, present in the southern European populations. Evidence in favour of the divergence between these regions is shown by the cluster analyses (UPGMA, Wagner procedure) of both allozyme and RAPD data. However, allozymes provided a slightly better fit of data to predictions about genetic distance (higher values of cophenetic correlation of clusters from the “goodness-of-fit” statistics) and better correlation between genetic and geographic distances (Mantel’s r). In both studies a strong similarity between the Bay of Biscay and the German Bight populations was evident, judging from their clustering positions. It was apparent that the trend of probable movement of migrants through the English Channel suggested by the allozyme study was supported using RAPD markers. The apparent absence of isolation-by-distance (non-significant correlation of genetic with geographic distance) in both studies suggests that S. solea may not yet be at genetic equilibrium. Overall, S. solea conforms to the type of species with either a broad variety of geographic and ecological habitats, or large and stable population sizes, or substantial gene flow at a regional scale.
A. Exadactylos: Biodiversity Conservation Laboratory, Department of Environmental Studies, University of the Aegean, Mytilene, GR81100, Hellas [tel: +0251–36247, e-mail: [email protected]]. A.Y. Troumbis: Biodiversity Conservation Laboratory, Department of Environmental Studies, University of the Aegean, Mytilene, GR81100, Hellas [tel: +0251–36230, e-mail: [email protected]]. J.P. Thorpe: Port Erin Marine Laboratory, School of Biological Sciences, The University of Liverpool, Port Erin, Isle of Man, IM9 6JA, UK [tel: +01624–831010, e-mail: J.P.Thorpe@ liverpool.ac.uk]
ICES CM 2001/J:17 – WITHDRAWN
ICES CM 2001/J:18
Automated fecundity and egg-sizing for marine species from scanned images: rapid sample processing and large data volumes
Kevin Friedland, D. Ama-Abasi, L. Clarke, D. Sigourney, and A. Stein
One of the most fundamental parameters in fisheries management and ecology is the population production of eggs. Management decisions are largely based on the dynamics of spawning stock biomass, and it is essential to know the contribution of viable gametes by age, which is affected at younger ages by growth and maturity rates and at older ages by the physiology of reproductive senescence. In addition to population modelling, recruitment dynamics are also strongly influenced by the size and quality of eggs. Egg quality can be characterized by a number of different qualitative and quantitative approaches; however, the most attractive methods developed to date rely on techniques that are efficient and lend themselves to wide application. The measurement of fecundity and egg size for fish and invertebrate fishery resources has been plagued by the cumbersome nature of the traditional approaches. We have developed an imaging-based technique that allows for the reduction of samples to electronic counts in relative little time and with a favourable costing. The technique is dependant upon high resolution scans of plated egg samples and the
7

CM 2001/J:16 The Life History, Dynamics and Exploitation of Living
Marine Resources: Advances in knowledge and methodology
1
�OT TO BE CITED WITHOUT PRIOR REFERE�CE TO THE AUTHOR
POPULATIO� STRUCTURE OF THE DOVER SOLE, SOLEA
SOLEA L., I� THE ATLA�TIC USI�G RA�DOMLY
AMPLIFIED POLYMORPHIC D�A (PCR-RAPD) MARKI�G
ATHANASIOS EXADACTYLOS1, ANDREAS Y. TROUMBIS
1, AUDREY J. GEFFEN
2,
JOHN P. THORPE2
1Biodiversity Conservation Laboratory, Department of Environmental Studies, University of the
Aegean, Mytilene, GR 81100, E-mail: [email protected].
2Port Erin Marine Laboratory, School of Biological Sciences, The University of Liverpool, Port Erin,
Isle of Man, IM9 6JA.

CM 2001/J:16 The Life History, Dynamics and Exploitation of Living
Marine Resources: Advances in knowledge and methodology
2
ABSTRACT
The primary objective of this work is to compare patterns of allozyme and RAPD variation in six
Atlantic populations of Solea solea. Sixteen individuals were randomly selected from each of the six
Atlantic populations of S. solea sampled during 1994 and 1995. In total, 96 individuals were
screened from three Irish Sea, two North Sea and one Bay of Biscay populations. Allozyme and
RAPD markers produced results, which were in general agreement with each other. However, RAPD
markers detected more variation in the sampled sole populations than that observed by using allozyme
marker. Percentage of polymorphism and genetic diversity estimates were very high. There was
clearly no geographic trend to any RAPD or allozyme allele frequencies at all polymorphic loci. The
chi-square contingency analyses (allele frequencies and hierarchical FST and non- hierarchical FDT)
indicated a highly significant heterogeneity among populations and between groupings. This
conclusion is consistent with the life history of Dover sole, the homing behaviour they exhibit and the
discrete spawning areas. A north to south pattern of significant population differentiation in the
allozyme study was replaced by a northwest to southeast significant variation. Natural selection for
differences in spawning habitat may explain the genetic isolation between these groupings. Another
reason could be the significant absence of some alleles, present in the southern European populations.
Evidence in favour of the divergence between these regions is shown by the cluster analyses
(UPGMA, Wagner procedure) of both allozyme and RAPD data. However, allozymes provided a
slightly better fit of data to predictions about genetic distance (higher values of cophenetic correlation
of clusters from the Goodness of fit statistics) and better correlation between genetic and geographic
distances (Mantel’s r). In both studies a strong similarity between the Bay of Biscay and the German
Bight populations was evident, judging from their clustering positions. It was apparent that the trend
of probable movement of migrants through the English Channel suggested by the allozyme study was
supported using RAPD markers. The apparent absence of isolation-by-distance (non significant
correlation of genetic with geographic distance) in both studies suggests that S. solea may not yet be
at genetic equilibrium. Overall, S. solea conforms to the type of species with either a broad variety of
geographic and ecological habitats, or large and stable population sizes, or substantial gene flow at a
regional scale.

CM 2001/J:16 The Life History, Dynamics and Exploitation of Living
Marine Resources: Advances in knowledge and methodology
3
1. I�TRODUCTIO�
Estimates of genetic variation are increasingly being based upon information at the DNA level
as distinct from allozyme electrophoresis. Random amplification of polymorphic DNA (RAPD) by
polymerase chain reaction (PCR), or PCR-RAPD, is a means of detecting polymorphisms rapidly for
genetic mapping and strain identification (Welsh and McClelland, 1990, 1991; Williams et al., 1990;
Hadrys et al., 1992). The method can detect extensive polymorphisms and does not require a priori
sequence information on the organism studied. Polymorphisms result from either size changes in the
amplified region or base changes that alter primer binding (Lynch and Milligan, 1994). Such
polymorphisms are inherited in a Mendelian fashion and can be used as genetic markers (Bardakci
and Skibinski, 1994). PCR-RAPD is simpler, considerably faster and less expensive than other
methods for detecting DNA sequence variation (Caetano-Anolles et al., 1991a, b). The fact that
RAPD scan numerous loci in the genome makes the method generally attractive for analysis of
genetic distance, genetic structure at a population level, breeding applications, or phylogeny
reconstruction (Clark and Lanigan, 1993).
Practical problems for RAPD applications include the difficulty in distinguishing products of
different loci with similar molecular weights (comigration) (Grosberg et al., 1996). Polymorphism is
detected as band presence versus absence after PCR amplification, since RAPD markers are usually
dominant characters (Williams et al., 1990). These phenotypically dominant markers in diploid
organisms make it impossible to distinguish between the genotypes of homozygous individuals and
those heterozygous for the dominant allele (Lynch and Milligan, 1994). This problem does not
prevent the estimation of allele frequencies necessary for population genetic analysis, but it does
reduce the accuracy of such estimation compared to analysis with codominant markers (i.e.
allozymes) (Lynch and Milligan, 1994). Each RAPD locus must be treated as a two allele system,
with the assumption that only one of the alleles per locus is amplifiable by the PCR. The existence of
multiple amplifiable alleles at a locus is reported to be relatively rare (Clark and Lanigan, 1993).
Under random mating the inbreeding index FIS equals to zero and the expected genotype frequencies
follow the Hardy-Weinberg expectations. This is another assumption that has to be considered when
carrying out population-genetic structure analyses using dominant genetic markers (Stewart and
Excoffier, 1996).

CM 2001/J:16 The Life History, Dynamics and Exploitation of Living
Marine Resources: Advances in knowledge and methodology
4
Little is known about the capacity of RAPD to detect variation in a broader variety of natural
populations, or how such variation compares with that of allozyme-based analysis. A comparison of
allozyme and RAPD diversity is of interest. The allozyme database, although carrying its limitations,
is quite large and for some types of questions allozymes may still provide sufficient information. For
other questions, DNA based methods may prove superior (Grosberg et al., 1996). The primary
objective of this work is to compare patterns of allozyme and RAPD variation in six Atlantic
populations of Solea solea, using some of the more recently developed statistical methods. We have
already reported a preliminary allozyme study (Exadactylos et al., 1998), and we follow here in this
work with a RAPD survey to estimate levels of genetic diversity within populations and genetic
divergence between populations based on the same material.
2. MATERIALS A�D METHODS
Sixteen individuals were randomly selected from each of six Atlantic populations of S. solea
sampled during 1994 and 1995. In total, 96 individuals were screened from three Irish Sea, two
North Sea and one Bay of Biscay populations. Frozen tissue samples of skeletal muscle stored at -75
oC were used for DNA extractions. Preliminary trials were contacted on juvenile sole to evaluate the
better of two DNA extraction techniques with phenol-chloroform (Exadactylos, 1997).
2.1. DATA ANALYSIS
The entire data set of 96 individuals was used for the analysis, including the observed OPF15-
B monomorphic marker. The computer program RapdBios (Black, 1993) provides input files suitable
for Biosys-2 (Swofford and Selander, 1989), which was used to calculate allele frequencies,
heterozygosity estimates and standard genetic analyses of population structure. Heterogeneity χ2
analyses of allele frequencies among populations and within geographic basins were performed by
hierarchical analysis using Biosys-2. The sequential Bonferroni procedure was employed to control
the probability of incorrectly rejecting null hypotheses at the 0.005 level of significance. Calculation
of Wright’s (1978) non-hierarchical FDT was carried out in Biosys-2 to facilitate comparison of this
paper’s data set with that obtained from the allozyme study (Exadactylos et al., 1998). Weir and
Cockerham’s (1984) unbiased estimates of F-statistics were calculated using the RapdFST program,
version 3.0 (Black, 1995), using the weighted mean frequency of alleles. Contemporary estimates of
gene flow (�m) were calculated using Slatkin’s (1993) approximation, assuming current equilibrium
between migration and genetic drift. Two methods of cluster analyses were performed (UPGMA and
CM 2001/J:16 The Life History, Dynamics and Exploitation of Living
Marine Resources: Advances in knowledge and methodology
5
Wagner procedure), in order to reveal the better fitted dendrogram, according to the Goodness of fit
statistics, using Modified Rogers Distance (DT) and Prevosti’s Distance (Wright, 1978) respectively.
The relation between matrices of genetic and linear geographic distances was examined with a
Mantel’s (1967) test of time-spacing clustering using the Ntsys program (Rohlf, 1993).
3. RESULTS
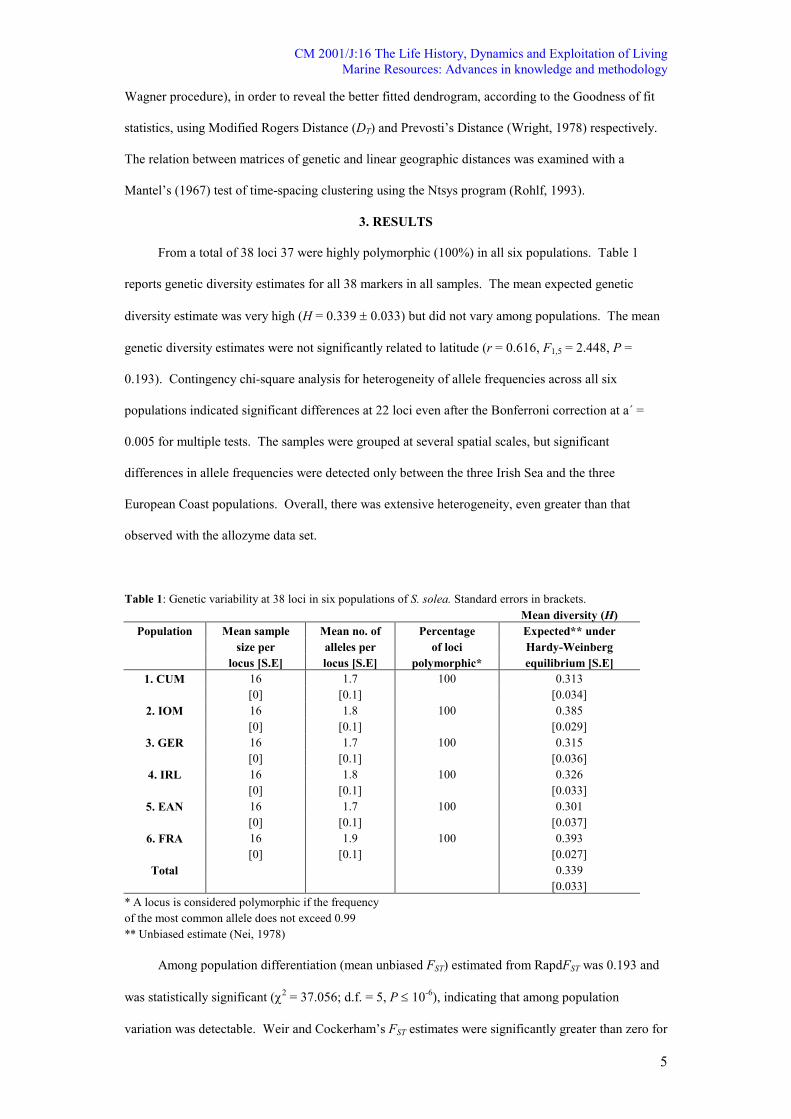
From a total of 38 loci 37 were highly polymorphic (100%) in all six populations. Table 1
reports genetic diversity estimates for all 38 markers in all samples. The mean expected genetic
diversity estimate was very high (H = 0.339 ± 0.033) but did not vary among populations. The mean
genetic diversity estimates were not significantly related to latitude (r = 0.616, F1,5 = 2.448, P =
0.193). Contingency chi-square analysis for heterogeneity of allele frequencies across all six
populations indicated significant differences at 22 loci even after the Bonferroni correction at a´ =
0.005 for multiple tests. The samples were grouped at several spatial scales, but significant
differences in allele frequencies were detected only between the three Irish Sea and the three
European Coast populations. Overall, there was extensive heterogeneity, even greater than that
observed with the allozyme data set.
Table 1: Genetic variability at 38 loci in six populations of S. solea. Standard errors in brackets.
Mean diversity (H)
Population Mean sample Mean no. of Percentage Expected** under
size per alleles per of loci Hardy-Weinberg
locus [S.E] locus [S.E] polymorphic* equilibrium [S.E]
1. CUM 16 1.7 100 0.313
[0] [0.1] [0.034]
2. IOM 16 1.8 100 0.385
[0] [0.1] [0.029]
3. GER 16 1.7 100 0.315
[0] [0.1] [0.036]
4. IRL 16 1.8 100 0.326
[0] [0.1] [0.033]
5. EA� 16 1.7 100 0.301
[0] [0.1] [0.037]
6. FRA 16 1.9 100 0.393
[0] [0.1] [0.027]
Total 0.339
[0.033]
* A locus is considered polymorphic if the frequency
of the most common allele does not exceed 0.99
** Unbiased estimate (Nei, 1978)
Among population differentiation (mean unbiased FST) estimated from RapdFST was 0.193 and
was statistically significant (χ2 = 37.056; d.f. = 5, P ≤ 10
-6), indicating that among population
variation was detectable. Weir and Cockerham’s FST estimates were significantly greater than zero for

CM 2001/J:16 The Life History, Dynamics and Exploitation of Living
Marine Resources: Advances in knowledge and methodology
6
21 loci, indicating a high level of population differentiation. Among them, OPF17-C was the locus
with the greatest overall genetic variation; the A allele had the greatest FST value and was therefore
the most variable between the sampling locations. It was evident that its frequency increased
gradually from north-west (Irish Sea) to south-east (European Coast). The mean estimate of Wright’s
non-hierarchical FDT calculated from Biosys-2 was 0.162 and was also significant (χ2 = 31.104; d.f. =
5, P ≤ 10-4). Gene flow estimates of approximately one immigrant into the average deme per
generation were calculated from both these FST values. Gene flow was considerably less when
estimated from RAPD (one migrant) as opposed to allozyme (11 migrants) data (Table 2).
Table 2: Estimates of among population differentiation (F-statistics), gene flow and correlation
of genetic distance with approximate linear geographic distance for RAPD and allozyme data sets.
Technique Populations Individuals FST m
1 FDT m
2 r Mantel's P
RAPD 6 96 0.193 1.045 0.162 1.293 0.37116 0.917
Allozymes 7 303 0.021 11.655 0.0262 9.292 0.54204 0.976
1 Gene flow calculated from Weir and Cockerham's FST using RapdFST (Black, 1993).
2 Gene flow calculated from Wright's (1978) FDT using Biosys-2.
The Wagner procedure tree gave a better fit according to the Goodness of fit statistics (Fig 2).
Despite the observed high cophenetic correlation of 0.887, the allozyme cophenetic correlation was
even higher 0.96, suggesting a better agreement between the original matrix and the phenogram than
with RAPD data. DT ranged from 0.232 between the Isle of Man and Eastern Irish Coast populations
to 0.342 between the two North Sea populations, indicating the small degree of genetic divergence
between these populations. Both trees produced a cluster of the geographically proximate Irish Sea
populations, which were related to the East Anglian Coast population. The Bay of Biscay population
was closely related to the German Bight population, in agreement with their clustering position in the
allozyme survey. A Mantel’s test shows that genetic distance was correlated with geographical
distance (r = 0.37; t = 1.385). However, this correlation was not statistically significant; the one-
tailed probability that the observed Z was greater than random Z in 1000 permutations was 0.917.
Correlation of geographic distance with genetic distance was also positive for the allozyme data set
yielding a higher r and t (r = 0.54; t = 1.978), but was also statistically non-significant (P = 0.976).

CM 2001/J:16 The Life History, Dynamics and Exploitation of Living
Marine Resources: Advances in knowledge and methodology
7
4. DISCUSSIO�
The chi-square contingency analyses (allele frequencies and hierarchical FST and non-
hierarchical FDT) indicated a highly significant heterogeneity among populations and between
groupings; results from both techniques. This conclusion is consistent with the life history of Dover
sole, the homing behaviour they exhibit (e.g. Rijnsdorp et al., 1992) and the discrete spawning areas
(e.g. Symonds and Rogers, 1995). These characteristics serve to reduce the homogenisation of
population allele frequencies between spawning stocks. A northwest to southeast significant variation
replaced a north to south pattern of significant population differentiation in the allozyme study.
Natural selection for differences in spawning habitat may explain the genetic isolation between these
groupings. Another reason could be the significant absence of some alleles, present in the southern
European populations. Therefore, the progressive decline in allozyme allelic diversity northwards
was interpreted as a possible consequence of a population bottleneck, founder effects, genetic drift, or
a combination of these. This interpretation is supported by the finding that populations sampled in
southern Europe are more variable than northern European populations; a finding consistent with the
general tendency for terrestrial populations to exhibit reduced genetic variability in areas glaciated
during the Pleistocene (e.g. Highton and Webster, 1976; Bellemin et al., 1978; Vaisanen and
Lehvasliho, 1984; Singh and Rhomberg, 1987). Such a stepping-stone pattern, that led to a loss of
genetic diversity even northern (Irish Sea) could also have been involved with colonisation of this
0 0.02 0.04 0.07 0.09 0.11 0.13
FRA
GER
IRL
EAN
CUM
IOM
Goodness of fit statistics: Farris (1972) “f” = 0.148
Prager and Wilson (1976) “F” = 4.426
% standard deviation (Fitch and Margoliash, 1976) = 5.978
Cophenetic correlation = 0.887
Figure 1: Wagner procedure demdogram using Prevosti’s Distance (Wright, 1978) and Goodness of fit statistics
between the six Atlantic Solea solea populations.

CM 2001/J:16 The Life History, Dynamics and Exploitation of Living
Marine Resources: Advances in knowledge and methodology
8
area by S. solea. This observed disjunction between the Irish Sea and the European Coast populations
indicates reduced exchange of migrants between these regions, in agreement with the conclusions in
the allozyme study.
Evidence in favour of the divergence between these regions is shown by the cluster analyses
(UPGMA, Wagner procedure) of both allozyme and RAPD data. However, allozymes provided a
slightly better fit of data to predictions about genetic distance (higher values of cophenetic correlation
of clusters from the Goodness of fit statistics) and better correlation between genetic and geographic
distances (Mantel’s r). In this regard, allozymes remain valuable for many types of population studies
despite the development of new genetic markers (Schaal et al., 1991). In both studies a strong
similarity between the Bay of Biscay and the German Bight populations was evident, judging from
their clustering positions. Cluster analyses also suggested a similarity between the three Irish Sea and
the East Anglian populations. It was apparent that the trend of probable movement of migrants
through the English Channel suggested by Exadactylos et al. (1998) was supported using RAPD
markers. Tagging experiments carried out by Greer-Walker and Emerson (1990) reported a seasonal
migration of soles through the Dover strait on both north-going and south-going tides. The apparent
absence of isolation-by-distance (non significant correlation of genetic with geographic distance) in
both studies suggests that S. solea may not be at genetic equilibrium (Slatkin, 1993). If populations
have not reached an equilibrium the F-statistics will underestimate the degree of differentiation
expected at equilibrium and the estimates of gene flow will overestimate the true levels (Slatkin,
1985). Estimated �m values of approximately one migrant into the average deme per generation
indicates gene flow among populations sufficient to deter population differentiation, if populations are
currently at an equilibrium between migration and genetic drift.
Several factors may explain these trends. Population fragmentation for S. solea may have been
a relatively recent event. This result is in agreement with the hypothesis reported in Exadactylos and
Thorpe (2001) that the family Soleidae and within it Solea is the most recently evolved group among
North Atlantic flatfishes (see also Lauder and Liem, 1983). Perhaps S. solea population
fragmentation has occurred sporadically throughout the species range, thus, there are no large
geographic areas that have become isolated. Furthermore, dispersal may be more restricted by
specific habitat types, rather than by distance to the next closest population. Genetic variability
depends on whether the species is a habitat specialist or a habitat generalist (Smith and Fujio, 1982).

CM 2001/J:16 The Life History, Dynamics and Exploitation of Living
Marine Resources: Advances in knowledge and methodology
9
Hence, adaptation for local selection pressures are possible. White (1978) proposed that effective
population size (effective number of individuals contributing to the next generation) may be related to
the extent of geographic habitat in a species undergoing substantial gene flow between its local
populations, but it can also reflect the ability of a species to occupy broad ecological habitats. S.
solea conforms to the type of species with either a broad variety of geographic and ecological
habitats, or large and stable population sizes (Kotoulas et al., 1995), or substantial gene flow at a
regional scale. Thus, FST / FDT results may be artificially significantly high from relatively poor
estimates of within-population variation. Resulting gene flow values from such FST estimates should
thus be interpreted with great caution, although they may serve as a useful relative measure with
which to compare between populations (Larcson et al., 1989). Overall, on the basis of the RAPD loci
examined, the population differentiation observed in the Atlantic Ocean is likely to be primarily the
result of random genetic drift or differential selection. Alternative means of detecting polymorphism
in S. solea including the use of single-locus minisatellite probes and mtDNA analysis may help to
further clarify the picture.
5. REFERE�CES
Bardakci, F., Skibinski, D.O.F., 1994. Application of the RAPD technique in tilapia fish: species and
subspecies identification.- Heredity 73: 117-123
Bellemin, J., Adest, G., Gorman, G. C., 1978. Genetic uniformity in northern populations of
Thamnopsis sirtalis (Serpentes: Colubridae).- Copeia 1978: 150-151
Black, B., 1993. RAPDBIOS - A FORTRAN program to convert RAPD-PCR files to BIOSYS
(DATYPE=3) files. Colorado State University, Department of Microbiology, Fort Collins, U.S.A
Black, B., 1995. RAPDFST 3.0 - A FORTRAN program to estimate Fst and effective migration rates
among subpopulations using RAPD-PCR files. Colorado State University, Department of
Microbiology, Fort Collins, U.S.A
Caetano-Anolles, G., Bassam, B.J., Gresshoff, P.M., 1991a. DNA amplification fingerprinting using
very short arbitrary oligonucleotide primers.- Biotechnology 9: 553-556
Caetano-Anolles, G., Bassam, B.J., Gresshoff, P.M., 1991b. DNA amplification fingerprinting: a
strategy for genome analysis.- Plant Mol. Biol. Rep. 9: 294-307
Clark, A.G., Lanigan, C.M.S., 1993. Prospects for estimating nucleotide divergence with RAPDs.- J.
Mol. Biol. Evol. 10: 1096-1111
Exadactylos, A., 1997. Population Genetics of the Dover sole, Solea solea L. (Linneaus, 1758)
(Teleostei: Soleidae). Ph.D. Thesis. University of Liverpool, School of Biological Sciences, Port
Erin Marine Lab.
Exadactylos, A., Geffen, A.J., Thorpe, J.P., 1998. Population structure of the Dover sole, Solea solea
L. in a background of high gene flow.- J. Sea Res. 40: 117-129

CM 2001/J:16 The Life History, Dynamics and Exploitation of Living
Marine Resources: Advances in knowledge and methodology
10
Exadactylos, A., Thorpe, J.P., 2001. Allozyme variation and genetic inter-relationships between seven
flatfish species of the order Pleuronectiformes. Zool. J. Linn. Soc. 132: 487-499
Farris, J.S., 1972. Estimating phylogenetic trees from distance matrices.- Amer. Nat. 106: 645-668
Fitch, W.M., Margoliash, E., 1967. Construction of phylogenetic trees.- Science 155: 279-284
Greer-Walker, M., Emerson, L., 1990. The seasonal migration of soles (Solea solea) through the
Dover strait.- Netherlands journal of Sea Research 25 (3): 417-422
Grosberg, R.K., Levitan, D.R., Cameron, B.B., 1996. Characterization of genetic structure and
genealogies using RAPD-PCR markers: A random primer for the novice and nervous. In J.D.
Ferraris, S.R. Palumbi. Molecular Zoology: Advances, Strategies and Protocols. Wiley-Liss. Inc.,
California: 67-100
Hadrys, H., Balick, M., Scierwater, B., 1992. Applications of random amplified polymorphic DNA
(RAPD) in molecular ecology.- Mol. Ecol. 1: 55-63
Highton, R., Webster, T.P., 1976. Geographic protein variation and divergence in populations of the
salamander, Plethodon cinereus.- Evolution 30: 33-45
Kotoulas, G., Bonhomme, F., Borsa, P., 1995. Genetic structure of the common sole Solea vulgaris at
different geographical scales.- Mar. Biol. 122: 361-375
Larcson, J.M., Riccardi, V.M., Calhoun, S.W., Morizot, D.C., 1989. Genetic differentiation of
bicolour damselfish (Eupomacentrus partitus) populations in the Florida Keys. - Mar. Biol. 103:
445-451
Lauder, G.V., Liem, K.F., 1983. The evolution and interrelationships of the actinopterygian fishes.-
Bulletin of the Museum of Comparative Zoology 150: 95-197
Lynch, M., Milligan, B.G., 1994. Analysis of population genetic structure with RAPD markers.-
Molecular Ecology 3: 91-99
Mantel, N., 1967. The detection of disease clustering and a generalised regression approach.- Cancer
Research 27: 209-220
Nei, M., 1987. Molecular Evolutionary Genetics. Columbia University Press, New York
Prager, E.M., Wilson, A.C., 1976. Congruency of phylogenies derived from different proteins.- J.
Mol. Evol. 9: 45-57
Rijnsdorp, A.D., Van Beek, F.A., Flatman, S., Millner, R.M., Riley, J.D., Giret, M., De Clerck, R.,
1992. Recruitment of sole stocks Solea solea L. in the Northeast Atlantic.- Netherlands Journal of
Sea Research 29 (1-3): 1173-192
Rohlf, F.J., 1993. NTSYS-pc Numerical taxonomy and multivariate analysis system, Version 1.80,
Applied Biostatistics, Setauket, NY
Schaal, B.A., O’Kane-Jr, S.L., Rogstad, S.H., 1991. DNA variation in plant populations.- Trends in
Ecology and Evolution 6: 329-336
Singh,R.S., Rhomberg, L.R., 1987. A comprehensive study of genic variation in natural populations
of Drosophila melanogaster. II. Estimates of heterozygosity and patterns of geographic variation.
Genetics 117: 255-271
Slatkin, M., 1985. Gene flow in natural populations.- A. Rev. Ecol. Syst. 16: 393-430

CM 2001/J:16 The Life History, Dynamics and Exploitation of Living
Marine Resources: Advances in knowledge and methodology
11
Slatkin, M., 1993. Isolation by distance in equilibrium and non-equilibrium populations.- Evolution
47: 264-279
Smith, P.J., Fujio, Y., 1982. Genetic variation in marine teleosts: high variability in habitat specialists
and low variability in habitat generalists.- Mar. Biol. 69: 7-20
Sneath, P.H.A., Sokal, R.R., 1973. Numerical Taxonomy. Freeman & Co., San Francisco
Stewart, C.N., Excoffier, L., 1996. Assessing population genetic structure and variability with RAPD
data: Application to Vaccinium macrocarpon (American Cranberry).- J. Evol. Biol. 9: 153-171
Swofford, D.L., Selander, R.B., 1989. Biosys-2: A computer program for the analysis of allelic
variation in population genetics and biochemical systematics. Champaign, Illinois 61801 USA,
D.L. Swofford, Illinois Natural History Survey
Symonds, D.J., Rogers, S.I., 1995. The influence of spawning and nursery grounds on the distribution
of sole, Solea solea (L.) in the Irish Sea, Bristol Channel and adjacent areas. - Journal of
experimental Marine Biology and Ecology 190: 243-261
Vaisanen, R.A., Lehvasliho, H., 1984. Absence of genic polymorphism in a northern population of the
house sparrow, Passer domesticus. Hereditas 100: 161-162
Weir, B.S., Cockerham, C.C., 1984. Estimating F-statistics for the analysis of population structure.-
Evolution 38 (6): 1358-1370
Welsh, J., McClelland, M., 1990. Fingerprinting genomes using PCR with arbitrary primers.- Nucleic
Acids Research 18: 7213-7218
Welsh, J., McClelland, M., 1991. Genomic fingerprinting using arbitrarily primed PCR and a matrix
of pairwise combinations or primers.- Nucleic Acids Research 19: 5275-5279
White, M, J.D., 1978. Models of speciation. W.H. Freeman and Co., San Francisco
Williams, J.G.K., Kubelik, A.R., Livak, K.J., Rafalski, J.A., Tingrey, S.V., 1990. DNA
polymorphisms amplified by arbitrary primers are useful as genetic markers.- Nucleic Acids
Research 18: 6531-6535
Wright, S., 1978. Variability within and among natural populations. Evolution and the Genetics of
populations. Vol. 4. University of Chicago Press, Chicago 580pp
Related Documents











