This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Population-level lateralized feeding behaviour in North Atlantic humpbackwhales, Megaptera novaeangliae
Claire Canning a,b,1, Danielle Crain a,c,1, T. Scott Eaton, Jr a,1, Kathryn Nuessly a,1, Ari Friedlaender c,*,Tom Hurst d, Susan Parks e, Colin Ware f, David Wiley g, Mason Weinrich a
a The Whale Center of New England, Gloucester, MA, U.S.A.bUniversity of Exeter, Cornwall Campus, Penryn, Cornwall, U.K.cDuke University Marine Laboratory, Beaufort, NC, U.S.A.dWoods Hole Oceanographic Institution, Woods Hole, MA, U.S.A.eApplied Research Laboratory, The Pennsylvania State University, State College, PA, U.S.A.fCenter for Coastal and Ocean Mapping, University of New Hampshire, Durham, NH, U.S.A.g Stellwagen Bank National Marine Sanctuary, NOAA National Ocean Service, Scituate, MA, U.S.A.
a r t i c l e i n f o
Article history:Received 1 May 2011Initial acceptance 5 May 2011Final acceptance 6 July 2011Available online 17 August 2011MS. number: A11-00353R
Keywords:DTAGfeeding behaviourhandedness indexhumpback whalejaw scuffinglateralized behaviourMegaptera novaeangliaeNorth Atlantic
Lateralized behaviour refers to a preference for using a specific limb or direction of movement that isconsistent across events. We investigated apparent lateralized behaviour in North Atlantic humpbackwhales, Megaptera novaeangliae, in the southern Gulf of Maine using digital acoustic tag (‘DTAG’) tech-nology to analyse feeding behaviours during 2004e2009. An ideal tool to examine lateralized behav-iours, DTAGs allowed continuous measurements of whale depth, heading, pitch and roll throughout theattachment period (less than 24 h). Side preference (right or left) in rolling behaviours (e.g. bottomrolling and side lunging) and directional preference (clockwise or anticlockwise) in feeding behaviourssuch as looping and spiraling were determined from visual examination of DTAG data. A ‘handednessindex’ (HI) was used to quantify side bias and strength of preference. We found a population-level right-side bias similar to the 90/10 right-hand bias in humans. Furthermore, photos of jaw scuffing and directtag data on rolling behaviour were used to assign a bottom-rolling side preference for 11 individuals forwhom both of these data were available, 10 (90.9%) of which demonstrated a right-side bias. Consistentlaterality was observed between individuals that performed bottom rolling and directional feedingbehaviours, which was biased in the right/clockwise direction. Younger individuals tended to showweaker lateralization scores than older individuals, even among adults (P ¼ 0.048), suggesting thatlateralization develops, and continues to do so, beyond sexual maturity.� 2011 The Association for the Study of Animal Behaviour. Published by Elsevier Ltd. All rights reserved.
‘Lateralization’ refers to a particular preference for usinga specific side of the body (e.g. a limb or direction of movement inmechanized behaviour, or a particular sensory organ such as an eyeor ear) that is consistent across events. However, the origin of lat-eralized behaviour remains under debate. Denenberg (1981) statesthat individual preferences could be ontogenetic, suggesting thatthese preferences are learned and therefore lack evolutionarysignificance. For instance, immature chimpanzees, Pan troglodytes,show weaker laterality in hand use than adults, especially adultfemales (Humle &Matsuzawa 2009). However, Versace et al. (2007)demonstrated that lambs, Ovis aries, are more strongly lateralizedin obstacle avoidance than adult sheep. Others suggest that in at
least some cases laterality is driven by a similar neurologicalasymmetry across individuals in a population (McGrew &Marchant1997; Rogers 2000).
Lateralization has evolved at both the individual and populationlevels. This behaviour has been commonly demonstrated at theindividual level in various animal studies (i.e. side preference oftrunk movements in Asian elephants, Elephas maximus:Haakonsson & Semple 2009; limb preference while performingroutine daily tasks, such as locomotion, feeding, grooming and tooluse, in several species of primates: McGrew & Marchant 1997;Peters & Rogers 2008). Lateralized behaviour at the populationlevel may have evolved as an evolutionary stable strategy (MaynardSmith 1982) in order to coordinate behaviour using ‘socialconstraints’ among asymmetric individuals, providing higherfitness for the individual in the group (Vallortigara & Rogers 2005).
The most recognizable example of laterality at the populationlevel is ‘handedness’ in humans, where approximately 90%
* Correspondence: A. Friedlaender, Duke University Marine Laboratory, 135Pivers Island Rd, Beaufort, NC 28516, U.S.A.
E-mail address: [email protected] (A. Friedlaender).1 First four authors contributed equally to the manuscript.
Contents lists available at ScienceDirect
Animal Behaviour
journal homepage: www.elsevier .com/locate/anbehav
0003-3472/$38.00 � 2011 The Association for the Study of Animal Behaviour. Published by Elsevier Ltd. All rights reserved.doi:10.1016/j.anbehav.2011.07.031
Animal Behaviour 82 (2011) 901e909

Author's personal copy
of individuals demonstrate a right-hand bias (Annett 1972).Population-level consistencies of lateralized behaviour are alsoapparent in nonhuman species (i.e. left-turn bias in the giant waterbug, Belostoma flumineum, while swimming through a T-maze:Kight et al. 2008; right- pectoral-fin bias in the sound production ofchannel catfish, Ictalurus punctatus: Fine et al. 1996; right-side biasin obstacle avoidance in flocks of sheep: Versace et al. 2007;anticlockwise-circling bias in herds of free-ranging reindeer,Rangifer tarandus: Espmark & Kinderås 2002). Furthermore, a pop-ulation-level right-sided preference for search and capture of preyhas also been shown in teleost fish (Miklósi & Andrew 1999),lungfish (Lippolis et al. 2009), frogs and toads (Vallortigara et al.1998; Robins & Rogers 2006), and birds (Ventolini et al. 2005;Rogers 2008).
Lateralization at the population level has also been well docu-mented in numerous marine mammal species. Some of this infor-mation is derived from studies conducted in captive settings (e.g.studies of bottlenose dolphins, Tursiops truncatus, beluga whales,Delphinapterus leucas, and California sea lions, Zalophuscalifornianus), although it is inherently limited due to constraints ofboth the setting and number of individuals studied (Sobel et al.1994; Marino & Stowe 1997a,b; Wells et al. 2006). Lateralizedbehaviour in marine mammals has also been examined usingscarring on carcasses of grey whales, Eschrichtius robustus (Kasuya& Rice 1970), photographs of scarring of live humpback whales,Megaptera novaeangliae (Clapham et al. 1995; Hain et al. 1995), andmeasurements of the density and length of pectoral appendages todetermine limb bias based on increased mechanical stress inwalruses, Odobenus rosmarus, and harbour porpoises, Phocoenaphocoena (Levermann et al. 2003; Galatius 2005). In all of thesecases, physical asymmetries were used as proxies to infer lateralitywith an overall bias towards the right side in the underlyingbehaviours. In North Atlantic humpback whales, 80% of individualsexamined showed predominantly right side rostrum and mandiblescarring, which was used as a proxy for laterality in subsurfacefeeding behaviours at the population level (Clapham et al. 1995).Surface behavioural observations, such as intentional beaching inthe bottlenose dolphin (Hoese 1971), aerial displays (e.g. breaching,flipper slapping) in humpback whales (Clapham et al. 1995), andobservations of wild beluga calfemother interactions (Kareninaet al. 2010) have also been used to determine population-levellaterality in wild cetaceans. More recently, digital acoustic tagtechnology (Johnson & Tyack 2003) has been used to describe lat-eralized behaviour in feeding grey whales (Woodward & Winn2006).
In this study we used photographic and tag-derived data toexamine lateralization across a number of feeding behaviours inNorth Atlantic humpback whales in the southern Gulf of Maine.Humpback whales are large (12e16 m, 22 000e26 000 kg)baleen whales that typically spend their winters in low-latitudebreeding grounds and their summers in higher-latitude feedinggrounds (Clapham 2000). Feeding ground aggregations in thisspecies appear to be maternally directed, with offspringmimicking the distribution of their mothers (Clapham et al.1993). Prey for humpback whales in this area is primarilyschooling fish including sand lance, Ammodytes spp. (Hain et al.1982; Payne et al. 1990) and herring, Clupea harengus (Weinrichet al. 1997).
In the present study, (1) we examined whether scarring on therostrum and mandibles of humpback whales could be used toinfer the laterality of subsurface feeding behaviours, (2) we testedthe hypothesis that humpback whales show consistent lateralitywithin and between individuals, and (3) we examined whetherthe strength of laterality in feeding behaviours varies with ageclass.
METHODS
Study Population
Humpback whale fine-scale movement patterns and under-water behaviour were recorded using digital acoustic tags, orDTAGs (Johnson & Tyack 2003; Friedlaender et al. 2009). The DTAGis a noninvasive tag with four suction cup attachments points thatis deployed from small inflatable rigid-hulled vessels. The tag isapplied as the whale begins to dive, which results in a short-livedreaction, causing minimal disruption to the animals’ activity. Tagsare programmed to detach within 24 h and record an animal’sheading, pitch, roll, acceleration and depth continuously usinga sampling rate of 50 Hz. A VHF signal is emitted when the tag is atthe surface to aid in its recovery. Data is archived on the tag anddownloaded upon retrieval.
These tags have been successfully deployed on multiple speciesof baleen whales (Nowacek et al. 2003; Woodward & Winn 2006;Friedlaender et al. 2009). Whale tag data were collected underNational Marine Fisheries Permits numbers 605-1607 and605-1904 issued to The Whale Center of New England and DukeUniversity Institutional Animal Care and Use Permit A041-09-02.During 2004e2009, 67 DTAGs were deployed on 57 individualhumpback whales, resulting in over 456 h of on-animal data (10whales were tagged multiple times). Five tag deployments wereunusable for analysis (in one case the tag was not recovered; theremaining four fell off quickly); data for this analysis were thereforeobtained from 52 individual humpback whales observed in 62usable tag deployments (Table 1).
Tagged whales were categorized by age and sex class using theWhale Center of New England (WCNE) humpback whale cataloguein Gloucester, MA, U.S.A. Sex was determined by molecular tech-niques applied to biopsy samples (Baker et al. 1991; Berube &Palsbøll 1996), photographing males and females in the genitalregion (Glockner 1983), or previous observations of female indi-viduals with a calf. In our sample of 52 individual tagged whales, 33were female, 9 were male and 10 were of unknown sex. Age wasdetermined either by seeing the animal initially as a young-of-the-year calf (exact age), the number of years since first sighted(minimum age), or seeing the whale with a calf (unknown ageadult). Animals were classified as calves (<1 year), juveniles(1e4 years), subadults (5e8 years), adults (9þ years) and unknown(Sardi et al. 2005). Our sample consisted of 4 calves, 0 juveniles,0 subadults, 43 adults and 5 whales of unknown age for which wecalculated minimum age. This sampling bias is due to specificallytargeting adults for tagging. Therefore, whales were split intocategories by age. Category 1 included calves, category 2 includedwhales 1e8 years of age, category 3 included whales 9e16 years ofage, category 4 included whales 17þ years of age, and category 5
Table 1DTAG deployment information on the 62 humpback whales organized by year andlocation
Year Location No. of whales tagged Total tag time (h)
2004 GSC 4 48:02:582005 JL 4 62:07:002006 SB 15 100:34:252007 SB 10 36:32:472008 SB 13 92:14:382009 SB, JL 16 116:52:28
Total 62 456:24:16
Total tag time for each year is provided. Locations include the Great South Channel(GSC), Jeffreys Ledge (JL) and Stellwagen Bank (SB). In 2009, whales were tagged intwo locations: 9 in SB, 7 in JL.
C. Canning et al. / Animal Behaviour 82 (2011) 901e909902

Author's personal copy
included whales of unknown age. Adults were further split intocategories 3 and 4 in order to test whether laterality is influencedby postmaturational ageing.
Study Area
Whales were tagged in three different habitats in the southernGulf of Maine off the New England, U.S.A. coastline: the Great SouthChannel (2004; N ¼ 4), Jeffreys Ledge (2005, 2009; N ¼ 11), andStellwagen Bank (2006e2009; N ¼ 47). Stellwagen Bank, part of theStellwagen Bank National Marine Sanctuary, is a submarine plateau39 km long and 13 km at the widest point (Meyer et al. 1979). Thebank ranges in depth from 18 to 36 m and contains a predominatelysandy bottom substrate with smaller areas of mud and gravel(Meyer et al. 1979). Deep-water basins that range from 45 to 140 msurround the bank with more complex bottom substrates. JeffreysLedge is a complex, winding glacial deposit found 19 km north ofStellwagen Bank. The Ledge is 54 km long with a depth range of45e61 m and the bottom substrate is a mixture of rock and mud(Appolonio 1979). The Great South Channel is encompassed by theeastern end of the Nantucket Shoals and the northwest threshold ofGeorges Bank and is 170 by 222 km (CeTAP 1982).
Primary prey differs for humpback whales among these threelocations. Sand lance, Ammodytes spp. is the preferred prey onStellwagen Bank and in the Great South Channel (Overholtz &Nicolas 1979; Hain et al. 1982; Payne et al. 1990), while onJeffreys Ledge, herring, C. harengus, is predominant (Weinrich et al.1997; Chase 2002; M. Weinrich, unpublished data).
TrackPlot Software
To visualize the subsurface feeding behaviours of tagged whales,we utilized ‘TrackPlot’, a custom software tool for DTAG data thatshows the whale’s path as a ribbon track, displaying heading, pitch,roll, depth and time at a sampling rate of approximately 0.5 sintervals (Ware et al. 2006). Rolling behaviours are indicated inTrackPlot by twists of the ribbon along its central axis, driven by theroll data gathered from the DTAG’s instrumentation (Ware et al.2006; Friedlaender et al. 2009). Tracks were specifically pro-grammed to indicate any roll of greater than 45� off the dor-soeventral axis (as determined by the tag’s roll data, corrected fortag placement as described in Ware et al. (2011)) as the colouryellow to enhance visual observations. Visual examination of theserolls was used to determine left (i.e. anticlockwise) or right(i.e. clockwise) direction of roll.
Feeding Behaviours
For this study, we examined lateral preferences inwhales duringa variety of feeding behaviours.
Rolling feeding behavioursBottom rolling. Thewhale dives to the bottom and completes one ormore off-axis rolls of greater than 45�, presumably feeding alongthe bottom with each roll (Goodyear 1989; Hain et al. 1995; Wareet al. 2006; Friedlaender et al. 2009; Fig. 1a).
Figure 1. Humpback whale feeding behaviours visualized by TrackPlot using DTAG data. Each tick mark on the green line attached to the whale graphic represents 1 m below thesurface: (a) bottom rolling, whale moving from left to right, (b) looping, whale at the bottom of its second loop about to lunge towards the surface, (c) side lunging, whale lungingtowards the surface, and (d) spiraling, whale moving from right to left.
C. Canning et al. / Animal Behaviour 82 (2011) 901e909 903

Author's personal copy
Side lunging. The whale rolls (� � 45�) at the surface or mid-waterwith an increase in fluke stroke amplitude preceding the roll (Wareet al. 2011; Fig. 1b).
Directional feeding behaviours (consistent with Wiley et al. 2011)Looping. The whale dives and corrals prey using variations ofa vertical circling technique. Looping was scored as clockwise (CW)or anticlockwise (ACW; Fig. 1c).
Spiraling. The whale uses variations of a horizontal corkscrewwhile ascending. Spiraling was scored as CW or ACW (Fig. 1d).
Only whales that performed more than 10 feeding events wereused in order to control for sample size limitations (sample sizesare given in Tables 3 and 4). Behaviours recorded for whales taggedmultiple times within and between years were both combined andanalysed separately for consistency between tagging sessions.
Jaw Scuffing
Jaw scuffing refers to the scarring or openwounds located on therostrum and mandibles, presumably due to repeated contact withthe bottom substrate (Hain et al. 1995; Clapham et al. 1995; Fig. 2).Photographs of tagged individuals were accessed from the long-term photographic databases at the Whale Center of NewEngland (WCNE), Gloucester, MA, U.S.A. (Weinrich et al. 1997;Weinrich & Corbelli 2009) and the Provincetown Center forCoastal Studies, Provincetown, MA, U.S.A. (Robbins 2007). We usedphotographs of animals where individual identification (based ondorsal fin and fluke pattern; Katona et al. 1979) and photographs ofboth sides of the mandibles and rostrum were available. Claphamet al. (1995) suggested that prevalence of jaw scuffing by side wasconsistent for periods of up to 12 years; for this study we used jawscuffing photographs from within 4 years of tagging to be conser-vative. Photographs were scored as right-side scuffed, left-sidescuffed, neither side scuffed, or both sides scuffed based on the
coloration and condition of skin around the whale’s rostrum andmandible, carefully excluding the white pigmentation that is oftenfound on the tip of humpback whale rostrums, which is sometimesconfused with jaw scuffing.
Statistical Analysis
The distributions of data on lateralization preferences extractedfrom tag tracks were tested for normality using chi-square good-ness-of-fit tests, and were determined to be nonparametric. To testfor a significant relationship between bottom-rolling and jaw-scuffed side, we used Microsoft Excel (2010) for statistical analysis(binomial distribution, Fisher’s exact and chi-square tests). Todetermine the side preference and strength of preference, we useda ‘handedness index’ (HI), as in Bard et al. (1990):
ðR� LÞ=ðRþ LÞ ¼ HI
where R is the number of right-biased feeding events and L is thenumber of left-biased feeding events. A positive HI indicates anoverall right bias, and a negative HI indicates an overall left bias.Wecategorized an HI of 0.9 or greater as a ‘right-biased’ whale and anHI of �0.9 or less as a ‘left-biased’ whale; any whale with an HIbetween �0.89 and þ0.89 was considered an ‘unbiased’ whale(Bard et al. 1990; McGrew & Marchant 1997). We tested forconsistent laterality within and between individuals by comparingthe HI of rolling behaviours with the HI of the directional behav-iours. We compared the absolute value of HI across age class (noanimals of unknown age were used in analyses), sex, yeartagged and the study area where the whale was tagged usinga KruskaleWallis test in SPSS v.17.0 (SPSS, Inc., Chicago, IL, U.S.A.),with a ManneWhitney U test used as a post hoc analysis. Finally,we compared the strength of the HI against the exact age (whereknown) and minimum age of tagged whales using Spearman’s rankcorrelation (SPSS v.17.0).
RESULTS
Handedness Index (HI) Consistency across Variables
Data from 37 individual whales were used in these analyses,eight of which were tagged multiple times within and/or acrossyears (five tagged twice in the same year; and four tagged multipletimes across years, one of which was tagged three times; Table 2).There was no significant difference in the HI between directionalfeeding behaviours (looping and spiraling; ManneWhitney U test:Z ¼ �1.356, N1 ¼15, N2 ¼ 10, P ¼ 0.261); therefore, the HIs for thesebehaviours were calculated using combined data. No differenceswere found between the HI and the portion of the study area inwhich the whales were tagged (KruskaleWallis test: c2
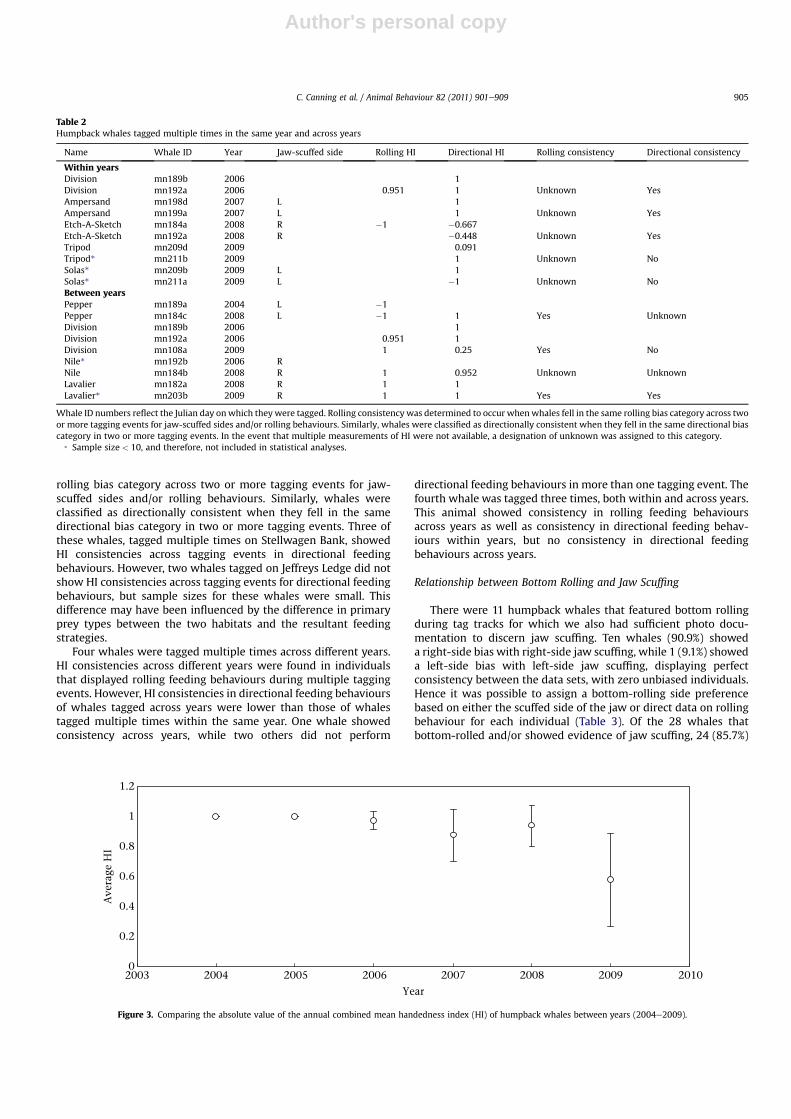
2 ¼ 4.380,N ¼ 49, P ¼ 0.112) or sex (ManneWhitney U test: Z ¼ �0.097,N1 ¼ 30, N2 ¼ 6, P ¼ 0.951). However, a significant difference wasfound between the HI of whales tagged between the years 2004and 2009 (KruskaleWallis test: c2
2 ¼ 11.984, N ¼ 49, P ¼ 0.03).Analysing this further, there was no difference in HI between theyears 2004 and 2008 (KruskaleWallis test: c2
4 ¼ 4.579, N ¼ 37,P ¼ 0.333; Fig. 3), but 2009 was significantly different from2004e2008 combined (ManneWhitney U test: Z ¼ �3.883,N1 ¼ 36, N2 ¼ 12, P < 0.001).
Handedness Index (HI) Consistency in Whales Tagged MultipleTimes
Five whales were tagged twice within the same year. Rollingconsistency was determined to occur when whales fell in the same
Figure 2. Example of an individual humpback whale that showed right-side jawscuffing with no signs of scuffing on the left side. Photographs: the Whale Center ofNew England (Gloucester, MA, U.S.A.).
C. Canning et al. / Animal Behaviour 82 (2011) 901e909904
Author's personal copy
rolling bias category across two or more tagging events for jaw-scuffed sides and/or rolling behaviours. Similarly, whales wereclassified as directionally consistent when they fell in the samedirectional bias category in two or more tagging events. Three ofthese whales, tagged multiple times on Stellwagen Bank, showedHI consistencies across tagging events in directional feedingbehaviours. However, two whales tagged on Jeffreys Ledge did notshow HI consistencies across tagging events for directional feedingbehaviours, but sample sizes for these whales were small. Thisdifference may have been influenced by the difference in primaryprey types between the two habitats and the resultant feedingstrategies.
Four whales were tagged multiple times across different years.HI consistencies across different years were found in individualsthat displayed rolling feeding behaviours during multiple taggingevents. However, HI consistencies in directional feeding behavioursof whales tagged across years were lower than those of whalestagged multiple times within the same year. One whale showedconsistency across years, while two others did not perform
directional feeding behaviours in more than one tagging event. Thefourth whale was tagged three times, both within and across years.This animal showed consistency in rolling feeding behavioursacross years as well as consistency in directional feeding behav-iours within years, but no consistency in directional feedingbehaviours across years.
Relationship between Bottom Rolling and Jaw Scuffing
There were 11 humpback whales that featured bottom rollingduring tag tracks for which we also had sufficient photo docu-mentation to discern jaw scuffing. Ten whales (90.9%) showeda right-side bias with right-side jaw scuffing, while 1 (9.1%) showeda left-side bias with left-side jaw scuffing, displaying perfectconsistency between the data sets, with zero unbiased individuals.Hence it was possible to assign a bottom-rolling side preferencebased on either the scuffed side of the jaw or direct data on rollingbehaviour for each individual (Table 3). Of the 28 whales thatbottom-rolled and/or showed evidence of jaw scuffing, 24 (85.7%)
Table 2Humpback whales tagged multiple times in the same year and across years
Name Whale ID Year Jaw-scuffed side Rolling HI Directional HI Rolling consistency Directional consistency
Within yearsDivision mn189b 2006 1Division mn192a 2006 0.951 1 Unknown YesAmpersand mn198d 2007 L 1Ampersand mn199a 2007 L 1 Unknown YesEtch-A-Sketch mn184a 2008 R �1 �0.667Etch-A-Sketch mn192a 2008 R �0.448 Unknown YesTripod mn209d 2009 0.091Tripod* mn211b 2009 1 Unknown NoSolas* mn209b 2009 L 1Solas* mn211a 2009 L �1 Unknown NoBetween yearsPepper mn189a 2004 L �1Pepper mn184c 2008 L �1 1 Yes UnknownDivision mn189b 2006 1Division mn192a 2006 0.951 1Division mn108a 2009 1 0.25 Yes NoNile* mn192b 2006 RNile mn184b 2008 R 1 0.952 Unknown UnknownLavalier mn182a 2008 R 1 1Lavalier* mn203b 2009 R 1 1 Yes Yes
Whale ID numbers reflect the Julian day onwhich they were tagged. Rolling consistency was determined to occur whenwhales fell in the same rolling bias category across twoor more tagging events for jaw-scuffed sides and/or rolling behaviours. Similarly, whales were classified as directionally consistent when they fell in the same directional biascategory in two or more tagging events. In the event that multiple measurements of HI were not available, a designation of unknown was assigned to this category.
* Sample size < 10, and therefore, not included in statistical analyses.
0
0.2
0.4
0.6
0.8
1
1.2
2003 2004 2005 2006 2007 2008 2009 2010
Year
Ave
rage
HI
Figure 3. Comparing the absolute value of the annual combined mean handedness index (HI) of humpback whales between years (2004e2009).
C. Canning et al. / Animal Behaviour 82 (2011) 901e909 905

Author's personal copy
showed a right-side bias and 4 (14.3%) showed a left-side bias(Fig. 4).
Consistency of Laterality between Behaviours
Of the 22 whales that performed rolling during feeding behav-iours (side lunging and/or bottom rolling), 18 individuals wereright-side biased, 1 individual was left-side biased, and 3 individ-uals were unbiased, of which the number of right-sided individualssignificantly exceeded the number of left-sided individuals (chi-square test: c2
2 ¼ 15.2, P < 0.001). Twenty-six whales showeddirectional feeding behaviours (looping and spiraling), of which 17were biased in a CW direction, 0 were biased in an ACW direction,and 9 were unbiased, of which the number of CW-biased individ-uals significantly exceeded the number of ACW-biased individuals(chi-square test: c2
2 ¼ 17, P < 0.001; Table 4). This CW bias wassignificant across individuals, that is, at the population level(binomial distribution: P < 0.001).
No behavioural consistency was found in the 16 whales thatshowed both directional feeding behaviours and bottom rolling. Ananimal is considered consistent in laterality between behaviours ifit shows a right-side bias for rolling as well as a clockwise bias indirectional behaviours, or, similarly, if it shows a left-side bias forrolling along with an anticlockwise bias in directional behaviours.Thirteen (81%) right-biased whales performed clockwise direc-tional behaviours, displaying consistent laterality between thebehaviours, whereas three (19%) left-biased whales also performed
clockwise behaviours. There was no difference in directional biaswhether the whale was right- or left-side biased (Fisher’s exacttest: two-tailed P >> 0.05; Table 5). Two whales that wereambiguous in their consistency were not included in this analysis.
Relationship between Laterality and Age
A significant difference was found in HI between age classes forthe 37 individuals that showed both directional and/or rollingfeeding behaviours (KruskaleWallis test: c2
3 ¼ 12.386, P ¼ 0.002).Spearman rank correlations showed a significant correlation
Table 3Humpback whales that showed a jaw-scuffed side bias (R ¼ Right, L ¼ Left), whichcould be used to infer a side preference for bottom rolling and vice versa
Whale name Whale ID Jaw-scuffed side Bottom-rolling preference
Bilbo mn200c R R (38)Dyad mn196a R R (162)Fulcrum mn195a R R (259)Ivee mn197a R R (184)Cardhu mn190a R R (10)Lavalier mn182a R R (189)Nile mn184b R R (240)Pepper mn184c L L (107)Perseid mn183b R R (232)Entropy mn201a R R (339)Samovar mn206b R R (369)
Sample sizes for bottom rolls are given in parentheses.
81.890.9 85.7
4.59.1
14.313.6
0
10
20
30
40
50
60
70
80
90
100
Bottom rolling
% W
hal
es s
how
ing
sid
e bi
as
Bottom rolling and jaw scuffing Bottom rolling and/or jawscuffing
Tagged humpback whales
Right
Left
Unbiased
Figure 4. Percentage of tagged humpback whales that showed a side bias when bottom rolling. Side bias was calculated using whales that (1) bottom-rolled, as a proxy (N ¼ 22); (2)had jaw-scuffed sides that also bottom-rolled (N ¼ 11); and (3) that had jaw-scuffed sides and/or bottom-rolled (N ¼ 28).
Table 4Humpback whales that showed directional feeding behaviours
Name Whale ID Year CW ACW HI Agecategory
Lavalier 09Calf
mn203a 2009 3 24 �0.777 1
Grackle mn198c 2007 9 2 0.636 3Geometry mn202a 2007 19 0 1 3Etch-A-Sketch mn184a,
mn192a2008 39 131 �0.541 3
Falcon mn189a 2008 15 0 1 3Epee mn195b 2006 32 0 1 3Venom mn193c 2008 9 1 0.8 3Draco mn206a 2009 3 12 �0.625 3Solas mn209b,
mn211a2009 7 3 0.4 3
Bungee mn177a 2005 14 0 1 4Division mn192a,
mn108a2006,2009
136 30 0.639 4
Dyad mn196a 2006 56 0 1 4Underline mn188a 2006 35 0 1 4Zipper mn189c 2006 36 0 1 4Ampersand mn198d,
mn199a2007 63 0 1 4
Anvil mn197c 2007 28 2 0.867 4Cardhu mn190a 2008 13 0 1 4Isthmus mn196a 2008 47 0 1 4Nile mn184b 2008 46 1 0.957 4Pepper mn184c 2008 39 0 1 4Tripod mn211b,
mn209d2009 13 5 0.444 4
Lavalier mn182a,mn203b
2008,2009
97 0 1 5
CW: clockwise; ACW: anticlockwise; HI: handedness index. Age categories were asfollows: 1 ¼ calf; 2 ¼ 1e8 years of age; 3 ¼ 9e16 years of age; 4 ¼ 17þ years of age;and 5 ¼ unknown age.
C. Canning et al. / Animal Behaviour 82 (2011) 901e909906

Author's personal copy
between HI and exact age (rS ¼ 0.619, N ¼ 19, P ¼ 0.005) andbetween HI and minimum age (rS ¼ 0.307, N ¼ 42, P ¼ 0.048). In allcases the older age classes showed a stronger index of handedness(Fig. 5).
DISCUSSION
Our results confirm previous studies (Clapham et al. 1995; Hainet al. 1995) suggesting that North Atlantic humpback whales in thesouthern Gulf of Maine display a strong population-level right-sidebias in rolling feeding behaviours. Furthermore, our data confirmprevious hypotheses that scarring on the rostrum and/or mandi-bles, termed jaw scuffing, is a reliable indicator through whichlateralized behaviour in this population can be determined. Ourresults provide further evidence that the handedness index forhumpback whales approaches a ratio similar to the 90:10 right-hand bias found in humans (Annett 1972). Additionally, ourresults show a population-level bias to swim clockwise duringdirectional feeding behaviours. However, true consistency was notfound between lateralization across rolling and directional feedingbehaviours. Finally, our results indicate that, as whales age, thestrength of their laterality in feeding behaviours increases, which
could result from either learning or forming habits from pastfeeding events (Weinrich et al. 1992).
Handedness Index (HI) Consistency across Categories
We found no difference in laterality among individuals using theHI when comparing different sexes, portions of the study area inwhich the whales were tagged, and tagging years 2004e2008.However, in 2009, there was a significant decrease in HI. This couldbe due to variability in prey type, abundance, density, or otherenvironmental factors beyond the scope of our analysis. Therefore,we cannot rule out that this is a biologically insignificant artefact ofrandom sampling.
Relationship between Bottom Rolling and Jaw Scuffing
Previous work (Clapham et al. 1995; Hain et al. 1995) hypothe-sized that jaw scuffing is an indication of bottom rolling whilefeeding, which our results confirm. Clapham et al. (1995) founda population-level right side:left side rolling ratio of 80:20 forhumpback whales by observing photographic evidence of jawscuffing; our results present a very similar 85:15 ratio. A left- or
Table 5Humpback whales that showed both directional feeding behaviours and bottom rolling
Name Whale ID Year HI for directionalbehaviours
Bottom-rolling side and/orjaw-scuffed side
Consistency acrossbehaviours
Deuce mn191a 2004 1 1 YesDivision mn108a 2009 0.231 1 AmbiguousDivision mn192a 2006 1 1 YesDyad mn196a 2006 0.966 1 YesEpee mn195b 2006 1 1 YesUnderline mn188a 2006 1 1 YesZipper mn189c 2006 1 1 YesAmpersand mn198d, mn199a 2007 1 �1 NoAnvil mn197c 2007 1 1 YesGeometry mn202a 2007 1 1 YesCardhu mn190a 2008 1 1 YesEtch-A-Sketch mn184a, mn192a 2008 �0.523 1 AmbiguousFalcon mn189a 2008 1 1 YesIsthmus mn196a 2008 1 1 YesLavalier mn182a 2008 1 1 YesNile mn184b 2008 0.96 1 YesPepper mn184c 2008 1 �1 NoVenom mn193c 2008 0.8 �1 No
The handedness index (HI) of directional behaviours was compared to bottom-rolling side bias and/or jaw-scuffed side bias to determine consistency of laterality acrossbehaviours.
0
0.2
0.4
0.6
0.8
1
1.2
1 3 4
Age class
Ave
rage
HI
Figure 5. Absolute mean handedness index (HI) values between age classes of humpback whales. No known juveniles or subadults were observed in this study. Age categories wereas follows: 1 ¼ calf; 2 ¼ 1e8 years of age; 3 ¼ 9e16 years of age; 4 ¼ 17þ years of age; 5 ¼ unknown age.
C. Canning et al. / Animal Behaviour 82 (2011) 901e909 907

Author's personal copy
right-side bias could be correctly assigned to all 11 individuals thatperformed both bottom rolling and jaw scuffing (based on photo-graphic evidence). However, one whale showed no evidence of jawscuffing on the rostrum and/or mandibles (based on photographicevidence), but was observed bottom rolling 640 times, 97.1% to theright side. While this result does not refute the other findings, thelack of any jaw scuffing on this individual is surprising. This absencecould be a result of behaviours that occur close to, but not in contactwith, the bottom (e.g. ‘prey flushing’). Prey flushing was hypothe-sized by Goodyear (1989), who suggested that whales swimming inclose proximity to the bottom substrate could create a ‘bow-wave’disturbance, flushing the sand lance burrowed in the sand withoutactually contacting the ocean floor. However, given the prevalenceof jaw scuffing on most whales in our study, bottom contact, assuggested by Clapham et al. (1995) and Hain et al. (1995), isprobably far more common than prey flushing.
Consistency of Laterality between Behaviours
Whales that were tagged multiple times displayed consistencyin rolling and directional feeding behaviours. We determinedconsistency between rolling and directional feeding behaviours tobe when a right-side biased whale displayed a clockwise direc-tional bias, or if the whale was left-side biased, to display an anti-clockwise directional bias. However, while there appeared to beconsistency of laterality between behaviours, all whales demon-strated a clockwise directional bias, despite their side bias in rollingbehaviour. This suggests that directional feeding behaviours mightnot be an accurate representation of a true lateral bias, despite theobservation of an overall CW preference shown by the population.Nevertheless, this is the first study of a wild marine mammal thathas demonstrated a population-level directional preference (CW/ACW) during feeding events. Consistent laterality at the populationlevel may prove to yield a social benefit for individuals by means ofimproving feeding efficiency, as in Asian elephants, or by coordi-nating feeding behaviour in the individuals of a population (Rogers2002; Martin & Niemitz 2003). While a population bias towardsCW directional feeding behaviours in humpback whales couldmake it easier to collaborate, it could also lead to a niche forming inthis area to effectively parasitize on the feeding behaviour ofanother whale (Weinrich et al., in press).
Strength of Laterality and Age Class
Our results suggest that laterality increases across large-scaleage classes (i.e. younger animals display weaker laterality thanolder animals, even among adults). Calves had significantly weakerHIs than all adults, and younger adults had significantly weaker HIsthan older adults. However, the data set did not contain a sufficientsample size of calves or juveniles to fully interpret this difference. Itappears that the strength of laterality in North Atlantic humpbackwhales intensifies with age, even after reaching physical maturity.These findings parallel those of Humle & Matsuzawa (2009), whoexamined tool use and laterality in wild Bossou chimpanzees, Pantroglodytes verus, where younger individuals tended to be less lat-eralized than adults. Perhaps a lack of fixed laterality in younganimals can allow maximum flexibility for behavioural trans-mission throughout a population (Weinrich et al. 1992).
Until now, most examples of lateralized behaviour in marinemammals have been documented in captive studies (Sobel et al.1994; Marino & Stowe 1997a, b; Wells et al. 2006). There arerelatively few examples with substantial sample sizes thatdemonstrate lateralized behaviour at the population level in wildcetaceans. This study confirms previously hypothesized behav-ioural laterality studies by adding details on underwater behaviour.
Further studies should incorporate a broader repertoire ofbehaviours (i.e. playing, nursing, breaching, flipper slapping, etc.) inorder to determine a true bias similar to the one found in rollingfeeding behaviours of humpback whales. However, the possibilityremains that increasedmechanical stress resulting from repetitiouslimb usemay affect some lateralized characteristics (e.g. rolling anddirectional feeding behaviours), which could enhance individualpreferences (Rogers 2002; Levermann et al. 2003; Galatius 2005).Therefore, skeletons of humpback whales should be examined forphysical bilateral directional asymmetries similar to those found inharbour porpoises and walruses (Levermann et al. 2003; Galatius2005). Such asymmetries would provide a link between behav-ioural lateralization and its physical expression, as well as providingfurther evidence for lateralized behaviour in humpback whales.
Acknowledgments
We thank the officers and crew of the NOAA research vesselsNancy Foster and Auk for their capable assistance during fieldoperations. We also thank the various members of our field teamover the years including: Roland Arsenault, Pat Halpin, Elliot Hazen,Tom Hurst, Just Moller, Cara Pekarcik, Allison Rosner, Kate Sardi,Jamison Smith, Alison Stimpert, Jennifer Tackaberry, Becky Wood-ward and Jeremy Winn. Pete Raimondi gave invaluable help withstatistics. Furthermore, we thank two anonymous referees forinsightful comments on the manuscript. Funding was provided bythe Stellwagen Bank National Marine Sanctuary, Office of NationalMarine Sanctuaries and the National Oceanographic PartnershipProgram. Jooke Robbins and the Provincetown Center for CoastalStudies graciously provided additional jaw scuffing photographs fortagged whales. Whale tag data were collected under permits605-1607 and 605-1904, issued to The Whale Center of NewEngland by the National Marine Fisheries Service and DukeUniversity Institutional Animal Care and Use Permit A041-09-02.
References
Annett, M. 1972. The distribution of manual asymmetry. British Journal ofPsychology, 63, 343e358.
Appolonio, S. 1979. The Gulf of Maine. Rockland: Courier of Maine.Baker, C. S., Lambertsen, R. H., Weinrich, M. T., Calambokidis, J., Early, G. &
O’Brien, S. J. 1991. Molecular genetic identification of the sex of humpbackwhales (Megaptera novaeangliae). Reports of the International WhalingCommission, Special Issue, 13, 105e111.
Bard, K. A., Hopkins, W. D. & Fort, C. L. 1990. Lateral bias in infant chimpanzees(Pan troglodytes). Journal of Comparative Psychology, 104, 309e321.
Berube, M. & Palsbøll, P. 1996. Identification of sex in cetaceans by multiplexingwith three ZFX and ZFY specific primers. Molecular Ecology, 5, 283e287.
Cetacean and Turtle Assessment Program 1982. A Characterization of MarineMammals and Turtles in the Mid and North Atlantic Regions of the U.S. OuterContinental Shelf: Final Report. No. AA551-CT8-48. Washington, D.C: Bureau ofLand Management.
Chase, B. C. 2002. Differences in diet of Atlantic bluefin tuna (Thunnus thynnus) atfive seasonal feeding grounds on the New England continental shelf. FisheryBulletin, 100, 168e180.
Clapham, P. J. 2000. The humpback whale: seasonal feeding and breeding ina baleen whale. In: Cetacean Societies: Field Studies of Dolphins and Whales (Ed.by J. Mann, R. C. Connor, P. L. Tyack & H. Whitehead), pp. 173e196. Chicago:University of Chicago Press.
Clapham, P. J., Baraff, L. S., Carlson, C. A., Christian, M. A., Mattila, D. K.,Mayo, M. A. & Pittman, S. 1993. Seasonal occurrence and annual return ofhumpback whales, Megaptera novaeangliae, in the southern Gulf of Maine.Canadian Journal of Zoology, 71, 440e443.
Clapham, P. J., Leimkuhler, E., Gray, B. K. & Mattila, D. K. 1995. Do humpbackwhales exhibit lateralized behaviour? Animal Behaviour, 50, 73e82.
Denenberg, V. H. 1981. Hemispheric laterality in animals and the effects of earlyexperience. Behavioral and Brain Sciences, 4, 1e49.
Espmark, Y. & Kinderås, K. 2002. Behavioural lateralisation in reindeer. Rangifer,22, 51e59.
Fine, M. L., McElroy, D., Rafi, J., King, C. B., Loesser, K. E. & Newton, S. 1996.Lateralization of pectoral stridulation sound production in the channel catfish.Physiology and Behavior, 60, 753e757.
C. Canning et al. / Animal Behaviour 82 (2011) 901e909908

Author's personal copy
Friedlaender, A. S., Hazen, E. L., Nowacek, D. P., Halpin, P. N., Ware, C.,Weinrich, M. T., Hurst, T. & Wiley, D. 2009. Diel changes in humpback whaleMegaptera novaeangliae feeding behavior in response to sand lance Ammodytesspp. behavior and distribution. Marine Ecology Progress Series, 395, 91e100.
Galatius, A. 2005. Bilateral directional asymmetry of the appendicular skeleton ofthe harbor porpoise (Phocoena phocoena).Marine Mammal Science, 21, 401e410.
Glockner, D. A. 1983. Determining the sex of humpback whales (Megapteranovaeangliae) in their natural environment. In: Behavior and Communication ofWhales. American Association for the Advancement of Science Symposium No. 76(Ed. by R. S. Payne), pp. 447e464. Boulder, Colorado: Westview Press.
Goodyear, J. D. 1989. Night behavior and ecology of humpback whales (Megapteranovaeangliae) in the western North Atlantic. M.S. thesis, San Jose StateUniversity.
Haakonsson, J. E. & Semple, S. 2009. Lateralisation of trunk movements in captiveAsian elephants (Elephas maximus). Laterality, 14, 413e422.
Hain, J. H. W., Carter, G. R., Kraus, S. D., Mayo, C. A. & Winn, H. E. 1982. Feedingbehavior of the humpback whale, Megaptera novaeangliae, in the western NorthAtlantic. Fishery Bulletin, 80, 259e268.
Hain, J. H. W., Ellis, S. L., Kenney, R. D., Clapham, P. J., Gray, B. K., Weinrich, M. T.& Babb, I. G. 1995. Apparent bottom feeding by humpback whales on stell-wagen bank. Marine Mammal Science, 11, 464e479.
Hoese, H. D. 1971. Dolphin feeding out of water in a salt marsh. Journal ofMammalogy, 52, 222e223.
Humle, T. & Matsuzawa, T. 2009. Laterality in hand use across four tool-usebehaviors among wild chimpanzees of Bossou, Guinea, West Africa. AmericanJournal of Primatology, 71, 40e48.
Johnson, M. P. & Tyack, P. L. 2003. A digital acoustic recording tag for measuringthe response of wild marine mammals to sound. IEEE Journal of Oceanic Engi-neering, 28, 3e12.
Karenina,K., Giljov,A., Baranov,V.,Osipova, L.,Krasnova,V.&Malashichev,Y.2010.Visual laterality of calfemother interactions in wild whales. PLoS One, 5, e13787.
Kasuya, T. & Rice, D. W. 1970. Notes on baleen plates and on arrangement ofparasitic barnacles of gray whales. Scientific Reports of the Whales ResearchInstitute, 22, 39e43.
Katona, S., Baxter, B., Brazier, O., Kraus, S., Perkins, J. & Whitehead, H. 1979.Identification of humpback whales by fluke photographs. In: Behavior of MarineAnimals: Current Perspectives in Research (Ed. by H. E. Winn & B. L. Olla),pp. 33e44. New York: Plenum.
Kight, S. L., Steelman, L., Coffey, G., Lucente, J. & Castillo, M. 2008. Evidence ofpopulation-level lateralized behaviour in giant water bugs, Belostoma flumi-neum Say (Heteroptera: Belostomatidae): T-maze turning is left biased.Behavioural Processes, 79, 66e69.
Levermann, N., Galatius, A., Ehlme, G., Rysgaard, S. & Born, E. W. 2003. Feedingbehaviour of free-ranging walruses with notes on apparent dextrality of flipperuse. BMC Ecology, 3, 1e13.
Lippolis, G., Joss, J. M. P. & Rogers, L. J. 2009. Australian lungfish (Neoceratodus for-steri): a missing link in the evolution of complementary side biases for predatoravoidance and prey capture. Brain Behavior and Evolution, 73, 295e303.
McGrew, W. C. & Marchant, L. F. 1997. On the other hand: current issues in andmeta-analysis of the behavioral laterality of hand function in nonhumanprimates. Yearbook of Physical Anthropology, 40, 201e232.
Marino, L. & Stowe, J. 1997a. Lateralized behavior in two captive bottlenosedolphins (Tursiops truncatus). Zoo Biology, 16, 173e177.
Marino, L. & Stowe, J. 1997b. Lateralized behavior in a captive beluga whale(Delphinapterus leucas). Aquatic Mammals, 23, 101e103.
Martin, F. & Niemitz, C. 2003. ‘Right-trunkers’ and ‘left-trunkers’: side preferencesof trunk movements in wild Asian elephants (Elephas maximus). Journal ofComparative Psychology, 117, 371e379.
Maynard Smith, J. 1982. Evolution and the Theory of Games. Cambridge: CambridgeUniversity Press.
Meyer, T. L., Cooper, R. A. & Langton, R. W.1979. Relative abundance, behavior, andfood habits of the American sand lance, Ammodytes americanus, from the Gulfof Maine. Fishery Bulletin, 77, 243e253.
Miklósi, Á & Andrew, R. J. 1999. Right eye use associated with the decision to bite.Behavioural Brain Research, 105, 199e205.
Nowacek, D. P., Johnson, M. P. & Tyack, P. L. 2003. North Atlantic right whales(Eubalaena glacialis) ignore ships but respond to alerting stimuli. Proceedings ofthe Royal Society B, 271, 227e231.
Overholtz, W. J. & Nicolas, J. R. 1979. Apparent feeding by the fin whale,Balaenoptera physalus, and humpback whale, Megaptera novaeangliae, on theAmerican sand lance, Ammodytes americanus, in the northwest Atlantic. FisheryBulletin, 77, 285e287.
Payne, P. M.,Wiley, D. N., Young, S. B., Pittman, S., Clapham, P. J. & Jossi, J. W.1990.Recent fluctuations in the abundance of baleen whales in the southern Gulf ofMaine in relation to changes in selected prey. Fishery Bulletin, 88, 687e696.
Peters, H. H. & Rogers, L. J. 2008. Limb use and preferences in wild orang-utansduring feeding and locomotor behavior. American Journal of Primatology, 70,261e270.
Robbins, J. 2007. Structure and dynamics of the Gulf of Maine humpback whalepopulation. Ph.D. thesis, University of St Andrews.
Robins, A. & Rogers, L. J. 2006. Lateralized visual and motor responses in the greentree frog, Litoria caerulea. Animal Behaviour, 72, 843e852.
Rogers, L. J. 2000. Evolution of hemispheric specialization: advantages and disad-vantages. Brain and Language, 73, 236e253.
Rogers, L. J. 2002. Lateralization in vertebrates: its early evolution, general patternand development. Advances in the Study of Behavior, 31, 107e162.
Rogers, L. J. 2008. Development and function of lateralization in the avian brain.Brain Research Bulletin, 76, 235e244.
Sardi, K. A., Weinrich, M. T. & Connor, R. C. 2005. Social interactions of humpbackwhale (Megaptera novaeangliae) mother/calf pairs on a North Atlantic feedingground. Behaviour, 142, 731e750.
Sobel, N., Supin, A. Y. & Myslobodsky, M. S. 1994. Rotational swimming tendenciesin the dolphin (Tursiops truncatus). Behavioural Brain Research, 65, 41e45.
Vallortigara, G. & Rogers, L. J. 2005. Survival with an asymmetrical brain:advantages and disadvantages of cerebral lateralization. Behavioral and BrainSciences, 28, 575e633.
Vallortigara, G., Rogers, L. J., Bisazza, A., Lippolis, G. & Robins, A. 1998. Comple-mentary right and left hemifield use for predatory and agonistic behaviour intoads. NeuroReport, 9, 3341e3344.
Ventolini, N., Ferrero, E. A., Sponza, S., Della Chiesa, A., Zucca, P. & Vallortigara, G.2005. Laterality in the wild: preferential hemifield use during predatory andsexual behaviour in the black-winged stilt. Animal Behaviour, 69, 1077e1084.
Versace, E., Morgante, M., Pulina, G. & Vallortigara, G. 2007. Behavioural later-alization in sheep (Ovis aries). Behavioural Brain Research, 184, 72e80.
Ware, C., Arsenault, R. & Plumlee, M. 2006. Visualizing the underwater behavior ofhumpback whales. IEEE Computer Graphics and Applications, 26, 14e18.
Ware, C., Friedlaender, A. S. & Nowacek, D. 2011. Shallow and deep lunge feedingof humpback whales in fjords of the West Antarctic Peninsula. Marine MammalScience, 227, 587e605.
Weinrich, M. T. & Corbelli, C. 2009. Does whale watching in southern New Englandimpact humpback whale (Megaptera novaeangliae) calf production or calfsurvival. Biological Conservation, 142, 2931e2940.
Weinrich, M. T., Schilling, M. R. & Belt, C. R. 1992. Evidence for acquisition ofa novel feeding behaviour: lobtail feeding in humpback whales, Megapteranovaeangliae. Animal Behaviour, 44, 1059e1072.
Weinrich, M. T., Martin, M., Griffiths, R., Bove, J. & Schilling, M. 1997. A shift indistribution of humpback whales, Megaptera novaeangliae, in response to preyin the southern Gulf of Maine. Fishery Bulletin, 95, 826e836.
Weinrich, M., Friedlaender, A., Cholewiak, D., Hurst, T., Parks, S., Thompson, M.,Ware, C., Wiley, D. In press. Underwater behavior of associated humpbackwhales on their feeding grounds in the southern Gulf of Maine: cooperation,commensalism, or parasitism? Behavioral Ecology and Sociobiology.
Wells, D. L., Irwin, R. M. & Hepper, P. G. 2006. Lateralised swimming behaviour inthe California sea lion. Behavioural Processes, 73, 121e123.
Wiley, D., Ware, C., Bocconcelli, A., Cholewiak, D., Friedlaender, A.,Thompson, M. & Weinrich, M. 2011. Underwater components of humpbackwhale bubble-net feeding behaviour. Behaviour, 148, 575e602.
Woodward, B. L. & Winn, J. P. 2006. Apparent lateralized behavior in gray whalesfeeding off the central British Columbia coast. Marine Mammal Science, 22,64e73.
C. Canning et al. / Animal Behaviour 82 (2011) 901e909 909
Related Documents











![Nonlinear Bubble Dynamics And The Effects On Propagation ... · the mechanism by which humpback whales (Megaptera novaeangliae) exploit bubble nets to catch fish [8]. It has been](https://static.cupdf.com/doc/110x72/5f0462157e708231d40db3e8/nonlinear-bubble-dynamics-and-the-effects-on-propagation-the-mechanism-by-which.jpg)