Population Densities of Poison Dart Frogs in a Regenerating Tropical Forest as Measured by the Hayne Estimator A Thesis Presented by Jennifer Rose Bunnell Miller To the Joint Science Department Of The Claremont Colleges In partial fulfillment of The degree of Bachelor of Arts Senior Thesis in Organismal Biology April 2007

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Population Densities of Poison Dart Frogs in a Regenerating Tropical Forest
as Measured by the Hayne Estimator
A Thesis Presented
by
Jennifer Rose Bunnell Miller
To the Joint Science Department
Of The Claremont Colleges
In partial fulfillment of
The degree of Bachelor of Arts
Senior Thesis in Organismal Biology
April 2007

TABLE OF CONTENTS
ABSTRACT................................................................................................................................. 4
INTRODUCTION ......................................................................................................................... 5
MATERIALS AND METHODS ................................................................................................... 14
Study Area ........................................................................................................................... 14
Study Species....................................................................................................................... 17
Population Density.............................................................................................................. 19
Distribution ......................................................................................................................... 22
Abiotic Factors.................................................................................................................... 22
RESULTS ................................................................................................................................. 25
Population Densities and Distributions.............................................................................. 25
Rainfall................................................................................................................................ 32
Temperature ........................................................................................................................ 33
Time of day.......................................................................................................................... 34
DISCUSSION............................................................................................................................. 37
Hayne Estimator Validity.................................................................................................... 37
Population Densities and Distributions.............................................................................. 41
Temperature ........................................................................................................................ 44
Time of Day......................................................................................................................... 45
The Hayne Estimator as a Tool for Monitoring Amphibians ............................................. 46
Conclusions......................................................................................................................... 47
ACKNOWLEDGEMENTS........................................................................................................... 49
LITERATURE CITED................................................................................................................ 50
2

APPENDICES............................................................................................................................ 56
3

ABSTRACT
With amphibian populations declining throughout the world, there is an increasing
demand for effective tools to measure species responses to environmental change. This study
investigates the effectiveness of the Hayne Estimator in evaluating the densities of two
species of poison dart frogs in three Costa Rican lowland forest habitats with varying degrees
of recovery from deforestation (selectively-logged riparian forest, post-pasture secondary
forest and non-native bamboo plantation forest). Population densities of Dendrobates
granuliferus and Dendrobates auratus were significantly highest in riparian forest,
substantially lower in bamboo, and very low in secondary forest. This trend corresponds to
previous research on species recolonization after deforestation and subsequent regrowth and
indicates that the Hayne Estimator is well suited for the evaluation of poison dart frogs.
Abiotic factors such as proximity to water, rainfall, temperature and time of day were found
to have some effect on frog sighting frequency. Individuals of both species tended to
aggregate near water, but the proportional distribution of transects according to all habitat
water presence likely negated this effect. Rainfall was unrelated to the sighting frequency of
D. auratus but correlated with the sighting frequency of D. granuliferus. Air temperature did
not impact sighting frequency. Time of day, however, was found to influence the sighting
frequencies of both species, with peaks occurring in the early morning and late afternoon.
The robustness of the Hayne Estimator when used to monitor poison dart frogs suggests that
the technique may be a valuable tool for future conservation research.
4

INTRODUCTION
Since scientists gathered at the First World Congress of Herpetology in 1989 to
address the worldwide decline of amphibian populations, concern for these creatures has
increased at an accelerating rate (Phillips, 1990; Stuart et al., 2004). Now, nearly 2 decades
later, as populations continue to decrease in size and weaken in stability, scientists are calling
for the unprecedented cooperation of all to prevent further loss of amphibian diversity. In
2006, 49 accomplished herpetologists co-authored a forum in Science that announced the
disappearance of 122 species and identified 32.5% of known amphibians as threatened
(Mendelson III et al.). The group asserted that only through the union of “individuals,
governments, foundations, and the wider conservation community” would the escalating rate
of extinctions slow. Their recommendations echo the suggestions of other scientists and
necessitate the implementation of monitoring, surveys, habitat protection and breeding
research colonies in an international effort to ensure the continued existence of amphibians.
Long-term monitoring programs serve as a foundation of restoration ecology planning
because they reflect the responses of at-risk species to environmental change. Knowledge
about the population health of sensitive species in recovering habitats is invaluable to the
conservation community. These studies not only increase the likelihood of successful land
management of protected areas, but they also guide policy towards accurately prioritizing the
protection of land in regions and habitats that are greatly valued for preserving biodiversity.
Without a doubt, Latin America has been more severely impacted by amphibian
declines than any other region in the world (International Union for Conservation of Nature
and Natural Resources [IUCN], 2006). In Central and South America, over 30 genera, 9
families and 1,157 species of amphibians have declined or gone extinct (Young et al., 2001;
5

6
IUCN, 2006; Pounds et al., 2006). The tropical regions of this area have supported a high
diversity of amphibian species for millennia, leading the region to be classified as a
biodiversity hotspot (Figure 1; Myers et al., 2000; Brooks et al., 2002). When one considers
the increasing rate of new amphibian species discoveries (Donoghue and Alverson, 2000),
the potential number of amphibians that may have been harmed by human activities is
astounding.
Because of the extensive body of tropical ecological research conducted in Costa
Rica, this country has become a paradigm for understanding patterns of species decline in
other tropical locations. Although Costa Rica covers only 0.03% of the planet’s surface, it is
home to 4% of the world’s biodiversity and 3% of the world’s threatened amphibians (IUCN,
2006; World Resources Institute, 2006; National Biodiversity Institute, 2007). To date, one
amphibian species has gone extinct in the country (Bufo periglenes, the golden toad) and 64
species are considered threatened (IUCN, 2006). Although more than one-fifth of Costa
Rican land is protected, it is clear that further action must be taken in order to raise, or at
least sustain, the current level of biodiversity (World Resources Institute, 2006).
Like many other countries throughout the world, Costa Rica has been the site of
rampant deforestation over the past few centuries. However, human habitation has followed a
unique trend within the last several decades. With increasing job opportunities in the urban-
based tourism and textile industries, workers have begun to migrate away from agricultural
areas (Aide and Grau, 2004). Since 1960, the rural population in Latin America and the
Caribbean has dropped by 30%, a trend driven in part by a 20 million person decrease in the
population whose livelihood is based on agriculture, hunting, fishing or

7
Figure 1. The hotspots of the world. Costa Rica is classified as part of the Mesoamerica hotspot, which extends from southern
Mexico to Panama (image from Myers et al., 2000).

forestry since 1980 (Food and Agriculture Organization of the United Nations, 2004). While
much of the vacant land has been sold for the expansion of other farms, a substantial amount
has been abandoned, providing an opportunity for regrowth, recolonization and the
reestablishment of natural ecosystems.
While ecologists have extensively explored species’ responses to the degradation of
native habitat, less work has been done on recovering habitats. With the current trend in
Costa Rica favoring natural regrowth, and with the increasing public awareness about the
causes of global warming, a large movement to restore farmed and developed land across the
planet could occur within the next century. Species monitoring programs must be
implemented in order to predict, prepare and assist the associated changes in biodiversity.
In 2001, Pitzer College acquired the Firestone Center, a parcel of land in Costa Rica
that had previously been selectively stripped of forest and converted into a cattle farm and
Figure 2. The Firestone
Reserve, otherwise known as
the Finca la Isla del Cielo, is
owned by Pitzer College and
located near Dominical in
southwestern Costa Rica
(Firestone Center for
Restoration Ecology, 2006).
8

monoculture plantation (Figure 2). A series of changes in ownership has allowed the
Firestone Reserve to have 13 years of continuous natural regeneration. Today, the preserve is
an ideal study site for investigating the process of regrowth. The variation in land patch
quality permits the juxtaposition of species in native riparian habitat versus secondary and
non-native plantations that have had over a decade to recover. The Joint Science Department
of the Claremont Colleges launched a student research program in the summer of 2005 and
has plans to establish long-term monitoring programs to track the regeneration progress.
Since it is unrealistic to study the impacts of human activities on all species in a given
area, indicator species are studied as representatives of a larger group. Indicators may belong
to any taxonomic group, but are commonly characterized by a degree of sensitivity to
disturbance that mirrors the responses of a wide variety of other species (Landres et al.,
1988). Anurans are ideal indicators because all stages of their life cycles are highly
dependent on environmental conditions. Most frogs and toads require a permanent water
source for reproduction, the development of young, and a source of food. Their skins are
permeable to permit survival in water and on land, leaving their bodies vulnerable to the
chemical balance around them. The majority of anurans also consume insects, a group known
to shift radically with a change in vegetation (Gibbs and Stanton, 2001). Based on these
characteristics and others, anurans are especially susceptible to habitat loss, chemical
contamination, climate change and the introduction of exotic species and disease, factors
known as the leading causes of decline in other amphibian species as well (Young et al.,
2001).
Although anurans are prized for the insight they provide regarding the health of an
ecosystem, the creatures are also the arch nemeses of many field scientists. Cryptic and
9

nocturnal, the typical frog or toad is a challenge to study in its natural environment. The
endemic poison dart frogs of Costa Rica, however, provide a colorful alternative to studying
cryptic indicators in the tropics. All members of the family Dendrobatidae are
aposematically colored, diurnally active and easily identified to species. Seventeen species of
Dendrobates have been identified in Central America and more than 100 species are known
to South America (Maxson and Myers, 1985). In addition to the two-continent family
distribution, many poison dart frog species have a range that spans several countries. Data
collected on frogs in one region can thus be readily applied to an entirely different area.
In an effort to measure the biodiversity status in the regenerating Firestone Reserve
habitats, the abundances of two poison dart frog species were measured. Both the granular
Error!
Figure 3. Many frogs utilize camouflage to hide from predators and field scientists alike
(left, Hyla versicolor), whereas Dendrobatids have conspicuous skin color and patterns to
contrast against their background (right, Dendrobates azureus). Their aposematic coloration
conveys to predators the consequences of a quick snack. Photos courtesy of
www.livingunderworld.org and www.webshots.com
10

Figure 4. The study species: Dendrobates granuliferus (top) and Dendrobates auratus
(bottom). Photographs by Keith Christenson.
11

(Dendrobates granuliferus) and the green and black (Dendrobate auratus) poison dart frog
occur naturally on the preserve (Figure 4). The population densities of these species were
assessed in riparian, secondary and bamboo forest habitats during the early wet season of
October 2006. Data were collected and calculated using the Hayne Estimator technique
(Hayne, 1949), which utilizes measurements taken from observations of sighting angle and
distance to each animal. The densities were then applied to approximate frog recolonization
in the secondary and bamboo habitats as compared to the more pristine riparian habitat.
Because the method assumes that an individual will flush and be readily noticeable as
the observer approaches, the Hayne Estimator is not well designed for the cryptic, nocturnal
habits of most amphibians but has been repeatedly employed to evaluate populations of birds
and mammals (Coulson and Raines, 1985; Pelletier and Krebs, 1997). The conspicuous
coloration and diurnal activity periods of Dendrobatid frogs makes them potentially
appropriate for the Hayne Estimator technique. This study explores the utility of poison dart
frogs as subjects for the Hayne Estimator while investigating the quality of vegetation
regrowth at the Firestone Reserve as a means of supporting native levels of biodiversity.
To account for the impact of abiotic factors on frog sighting frequencies, proximity to
water, as well as correlations with rainfall, air temperature and time of day, were considered.
Because past studies indicate that poison dart frogs do not depend on large bodies of water
(reviewed by Savage, 1968; Vences et al., 2000; Jowers and Downie, 2005), random
distribution was expected. Rainfall and time of day have both been identified as influential
factors, with some Dendrobatids occurring in larger quantities in the presence of rain and in
the early morning and late afternoon (Graves, 1999). Finally, the air temperature was not
12

anticipated to affect sighting frequencies because of its small range due to the tropical
climate.
13

MATERIALS AND METHODS
Study Area
Field research was conducted at the Firestone Center for Restoration Ecology with the
permission of Pitzer College and the Joint Science Department. Claremont Colleges. The
Firestone Reserve is a 60 ha protected preserve of lowland (15m – 303 m) Pacific Moist
Forest in southwestern Costa Rica near Dominical (16.684 N, 51.643 W). The reserve has a
unique history that makes the area a suitable research site for an examination of poison dart
frog populations in regenerating habitats. Beginning around 1950, the property was
completed deforested, with the exception of two precipitous stream canyons within which
circa 100 m wide strips of riparian forest were only selectively logged (Firestone Center for
Restoration Ecology, 2006). The land was utilized as a cattle farm until 1993, when the
property was purchased by Ms. Firestone and converted into a combined sustainable farm
and private biological preserve. At this time, livestock were removed and parts of the land
were replanted with monoculture crops, including 5.9 ha of bamboo (Guadua aculeata, G.
angustifolia, Dendrocalamus asper, and D. latiflorus), 1 ha of bananas (Musa acuminata), 1
ha of black palm (Bactris gasipaes), and 24.7 ha of mixed hardwood tree species.1 The
remaining 27.4 ha were allowed to regenerate naturally. In 2005, the property was donated to
Pitzer College and farming maintenance was abandoned. The land has since been left alone
to regrow and is currently used as a biological reserve for education and research by the
Pitzer Study Abroad Program and the Claremont Colleges Joint Science Department.
The division of the reserve into multiple sub-habitats makes it an ideal location for
the study of biodiversity in recovering natural and non-native vegetation. The Firestone
1 Refer to http://costarica.jsd.claremont.edu/biodiversity/trees.shtml for an up-to-date listing of identified species.
14

15
Reserve borders the Hacienda Baru National Wildlife Refuge to form a contiguous 390 ha
sanctuary dedicated to scientific study and eco-tourism with minimal biological impact
(Hacienda Baru National Wildlife Refuge, accessed 2006).
This study focuses on Dendrobatid presence in three types of habitat on the Firestone
Reserve: selectively-logged riparian forest (from hereon referred to as “riparian”), abandoned
pasture (“secondary”) and bamboo plantation (“bamboo”; see Figure 5). The riparian regions
consist of primary forest with tall vegetation and dense canopy cover. The ground in this
habitat is shaded during most of the day and was usually covered by thick leaf litter. No
records are available that describe the method of selective logging in this habitat, leaving no
way to judge whether the land is an accurate standard of natural vegetation. However, the
riparian habitat of the Firestone Reserve is visually indistinguishable from the primary forest
of the Hacienda Baru National Wildlife Refuge. Therefore, the riparian habitat was used in
this study as a representative of natural forest conditions, although it should be recognized
that there are potential influences of the past selective logging that are unmeasured in this
study.
The secondary forest is comprised of lower, thinner trees than the riparian habitat.
Large patches of sunlight are often observed on the ground, causing grasses to replace damp
leaf litter throughout much of the secondary forest (Figure 5). The bamboo habitat features
thick groves of tall culms that provide moderate amounts of shade. Sunlight filters through
the vegetation at a lower intensity than in the secondary forest, and dense, knee-high
vegetation covers the ground around the bamboo. Multiple sources of water exist in all three
sampling habitats. Small and moderate, 1-10 m wide streams run through both the riparian
and secondary forests, while three large ponds border the bamboo habitat.
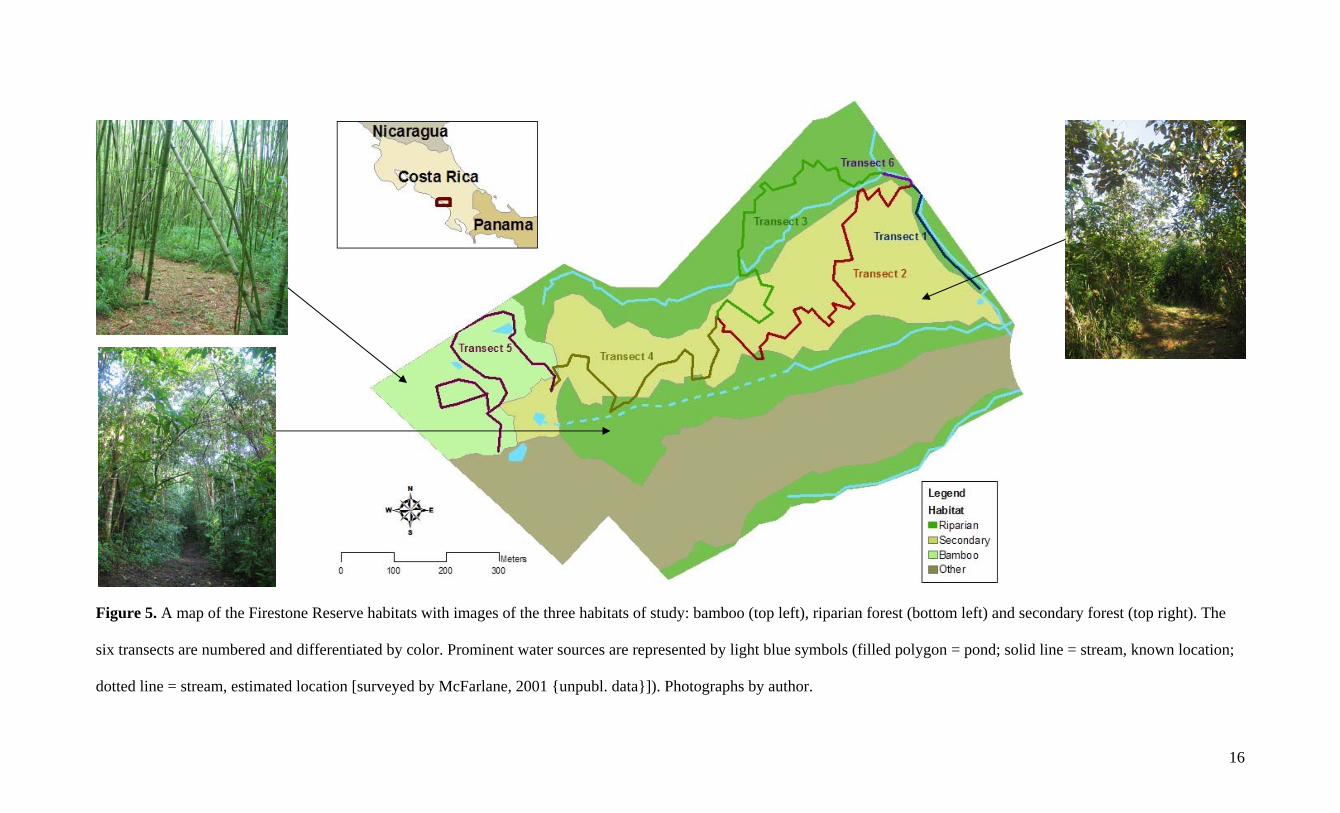
16
Figure 5. A map of the Firestone Reserve habitats with images of the three habitats of study: bamboo (top left), riparian forest (bottom left) and secondary forest (top right). The
six transects are numbered and differentiated by color. Prominent water sources are represented by light blue symbols (filled polygon = pond; solid line = stream, known location;
dotted line = stream, estimated location [surveyed by McFarlane, 2001 {unpubl. data}]). Photographs by author.
Observations were systematically made within each habitat by following the pre-
established trails of the reserve as transects for observation. The use of four maintained trails
permitted an accessible and repeatable loop through the forest and covered all three types of
habitat. The trails were divided into six transects of varying lengths, with most transects
covering multiple habitats (Table 1). The order that transects were walked was randomized
when possible so as not to cause unintentional correlations with time. Transects 1 and 4 could
not be shuffled because they provided starting and ending access or connected path loops,
respectively.
Table 1. Length distributions for study transects.
Habitat length (m) Transect
Trail ID*
Total length (m) Riparian Secondary Bamboo
1 WT 275.9 275.9 0 0 2 B 949.9 84.1 865.8 0 3 C 838.6 721.3 117.3 0 4 B 646.2 194.5 451.7 0 5 BB 985.6 0 236.6 749.0 6 C 100.5 100.5 0 0
*Trail ID corresponding to the survey by McFarlane, 2001 (unpubl. data).
Study Species
The combined Firestone-Hacienda Baru area hosts at least 28 known species of
anurans (M. Ryan, pers. comm.), including Dendrobates granuliferus, the granular poison
dart frog, and D. auratus, the green and black poison dart frog. While both species are
abundant on the Firestone Reserve, D. granuliferus is internationally recognized as a
threatened species due to habitat loss and degradation as well as human harvesting of the
species (IUCN, 2006). The range of D. granuliferus is also limited, covering 5,579 km2 from
17

the mid-western coastal lowlands of Costa Rica to the northern border of Panama (Global
Amphibian Assessment, 2006; IUCN, 2006). Dendrobates auratus is considered to be of
lesser concern, largely because of its greater range of 11,944 km2 from northern Costa Rica
through northern Columbia and higher tolerance of habitat degradation.
Dendrobates granuliferus and D. auratus were selected as study subjects because of
their relevance to amphibian declines and their conspicuous appearances in the field.
Dendrobatids have many natural history characteristics typical of tropical amphibians. All
species are diurnal and commonly live among the low vegetation and leaf litter of moist
forests below elevations of 3,000 m (Savage, 1968). They are considered terrestrial anurans
because their life cycles are independent of large water sources. Dendrobatid eggs are laid on
land and tadpoles are carried on the backs of their parents to temporary puddles of water
among vegetation. Dendrobatids specialize in eating ants but also consume a large quantity
of mites, insects that are also characteristic of the diets of other tropical amphibians such as
Atelopus, Bufo and Bolitoglossus (Toft, 1981; Anderson and Mathis, 1999). They mate
during the wet season like many other tropical amphibians, and they are most active between
May and November (reviewed by Savage, 2002). Because many of the human impacts that
threaten D. granuliferus and D. auratus also affect other tropical amphibians and potentially
other groups of organisms, these two species serve well as indicators of the status of tropical
wildlife populations.
In addition, these species were chosen because their unique aposematic coloration
makes them convenient to study in the field. While many anuran species are nocturnal and
camouflaged to their environments, Dendrobatids are diurnal and have brilliantly colored
skin markings. The coloration serves as a signal to predators, warning them of the toxic
18

alkaloids that can be released from the frogs’ skin glands as a mechanism of defense
(Saporito et al., 2004). The distinctive patterns of D. granuliferus and D. auratus permit easy
sighting of individuals and allowed for a high confidence in the accuracy of the field
techniques used in this study.
Frogs were observed on the Firestone Reserve during the wet season between 6
October and 14 October 2006. The research period corresponded to the Dendrobatid mating
season and the peak of their activity throughout the year (reviewed by Savage, 2002).
Observing at this time guaranteed the highest number of frog sightings possible, leading to
elevated estimates of population densities and an overall optimistic perspective of the
Dendrobatid presence on the Firestone Reserve.
Population Density
The population density of frogs was measured with the Hayne Estimator (Hayne,
1949). To keep measurement technique consistent, all observations were made by the author.
Two sessions of observations typically occurred each day. The first session began at
approximately 7:00 and ended around 11:00 and the second began at approximately 13:30
and ended around 16:30. Transects were walked at a constant speed from start to stop without
pause, except to record frog measurements. Consequentially, transects with many frog
sightings took longer to walk than transects with few sightings.
Each observation followed the same protocol. When a frog was sighted, the observer
immediately took three measurements (Figure 6):
(1) The distance from the observer to the frog’s location at first sighting. Measurements
were made using a Leica Geosytems laser rangefinder accurate to ± 3mm and later
19

trigonomically corrected from incline distances (i.e. from the height of the hand-held
rangefinder) to true plan distances.
(2) The magnetic bearings of the transect and frog, using a Suunto sighting compass
readable to ± 0.5 degrees.
(3) The time of the sighting.
Occasionally, when a frog was observed well beyond the first possible point of contact, the
observer back-tracked her steps until she reached the location where the frog first came into
view. For example, if a frog was first noticed when the observer was directly beside it, the
observer retraced her steps until she could first view the frog amidst the vegetation.
Obscurities due to vegetation occasionally caused sighting difficulties, but errors were most
likely not frequent enough to largely impact data. This technique corrected for the limitation
Figure 6. A visual representation of the
Hayne Estimator data collection
technique, showing the distance from the
observer to the frog (ri) and the
corresponding measured sighting area
(shaded red).
20

of being able to view only one side of a transect at a time.
The location of each frog was recorded relative to a surveyed map of the reserve
paths. A survey of current habitat borders was mapped during the study period and overlaid
on the original path survey. Survey information was used to relate frog sighting to habitat
type for use in the Hayne Estimator. Total transect length and average segment (i.e. the
distance between transect turns) length were also collected from the survey.
The population densities of D. granuliferus and D. aruatus for each observation
session were calculated for each habitat using the unmodified Hayne Estimator:
Dh =n
2L1n
1rii= t
n
∑⎛
⎝ ⎜
⎞
⎠ ⎟ ,
where Dh is the Hayne density estimate, n is the number of animals observed, L is the
transect length, and ri is the sighting distance to the ith animal. The standard deviation was
calculated by taking the square root of the variance, calculated as:
Variance(DH ) ≈ DH2 var(n)
n2 +
1r,
− R⎛
⎝ ⎜
⎞
⎠ ⎟
2
i= t
n
∑R2n n −1( )
⎡
⎣
⎢ ⎢ ⎢ ⎢ ⎢
⎤
⎦
⎥ ⎥ ⎥ , ⎥ ⎥
where R is the mean of the reciprocal of the sighting distances and calculated as:
R =1n
1rii= t
n
∑ ,
Circular statistics on sighting angles were computed using the StatistiXL Excel Add-
In (http://www.statistixl.com/). Population densities were analyzed with VassarStats (Lowry,
2007) for statistical differences between the species and habitats using One-Way Independent
ANOVA and Tukey HSD tests.
21

Distribution
To relate frog sightings to actual geographical features, ArcGIS Version 9.1 was used
to project the Firestone trail survey into a satellite image of the Firestone Reserve (obtained
from Digital Globe, Inc.) with reference to GPS coordinates collected at the site. Habitat
zones were constructed using several older habitat maps of the reserve as well as records of
current habitat boundaries taken during the study. Frog sighting points were imported and
displayed with graduated symbols to represent point densities. Water sources (streams and
ponds) were approximated and drawn by hand according to the trail survey (McFarlane,
2001, unpubl. data) and satellite image.
The proximity of each sighting to water was determined using COMPASS software
(Version 5.05; Fish, 2005) The straight-line distance between each sighting location and the
closest water source was measured and then correlated to the number of frogs sighted at the
location using a linear regression calculated with VassarStats (Lowry, 2007).
Abiotic Factors
To determine whether the transect distribution proportionally represented the amount
of water in each habitat, an analysis of transect-resource proportionality was conducted using
measurements from the Firestone maps created with ArcGIS to compare the ratio of the
habitat area within 50 m of a water source to the total habitat area versus the transect length
(by habitat) to the total transect length (by habitat). In other words,
area of habitat within 50m of watertotal area of habitat
: tran sec t length in habitat within 50m of watertotal length of tran sec t in habitat
22

Rainfall and air temperature (from now on referred to as “reserve temperature”) were
measured every 2 hours by a Davis Weatherlink meteorological station on the Firestone
Reserve. Average reserve temperature was calculated for each increment as the arithmetic
mean of the high and low temperatures. To test for a correlation between reserve temperature
and the frog sighting frequency, data were analyzed using a linear regression calculated with
Vassar Stats (Lowry, 2007). VassarStats was also used to determine whether a correlation
existed between rainfall and frog sighting frequency with a Pearson’s chi-square 2x2
contingency table test. To evaluate overall trends in frog sighting frequency, data from both
species were combined and compared to the time of sighting.
The air temperature in each habitat (from now on referred to as “habitat temperature”)
was measured using four temperature loggers (Stow Away XTI). One logger was attached to
a tree in each habitat and the sensor was oriented to hang freely (Figure 7). The loggers were
positioned so that they received light levels typical of the particular habitat (i.e. not in full
sunlight). A fourth control logger was set in a deforested meadow on the reserve to measure
the highest possible daily temperature (i.e. full sunlight). Loggers were set to record data for
each day and night of the study period and measured the habitat temperature every 5 or 20
minutes, depending on the format available on the logger. Temperature data from all the days
in the study period were averaged to find the 24-hour mean temperature fluctuation for each
habitat. The fluctuations of all habitats were then compared to determine whether a large
difference in temperature in any of the habitats may have influenced poison dart frog activity
levels.
23

Figure 7. Locations of the temperature loggers in each habitat: bamboo (left top), secondary
(bottom left), riparian (top right) and exposed meadow (bottom right). Photographs by
author.
24

RESULTS
Inconsistencies in data collected at the start of the study period have led to the
exclusion of several days of data from the final analysis. The number of frog sightings in the
first three days was significantly lower than sightings during the remainder of the observation
days (an average of 6 ± 6 observed frogs/km in contrast with 85 ± 39 observed frogs/km).
Additionally, no significant differences were found in abiotic factors such as rainfall or
temperature between the first 3 days and the subsequent days of observation. The lack of
disparity suggests that the initial low number of sightings was likely a result of the observer’s
learning period. A test run was not conducted ahead of time, leading the observer to learn the
sighting and measurement techniques during the official study period. Therefore, only data
from 9 October through 14 October 2006 were included in the analysis (Appendix A). Data
collected from 6 October to 8 October are listed in Appendix B but are not considered valid
data or incorporated into the thesis.
Population Densities and Distributions
A total of 166 D. granuliferus and 109 D. auratus were observed, resulting in a total
sample size of 275 frogs. The average population densities for both species were larger in the
riparian forest than in the secondary or bamboo habitats (Figure 8). For D. granuliferus, the
riparian density was estimated to be 68 times greater than the secondary density and 23 times
greater than the bamboo density. The D. auratus riparian density estimate was 155 times
greater than the secondary density but only three times greater than the bamboo density.
Bamboo densities were larger than secondary densities for both species, with density for D.
granuliferus in bamboo reaching an estimate that was three times larger than for secondary
25

forest and the density for D. auratus in bamboo estimated to be 47 times larger than in
secondary forest. ANOVA analysis indicated significant effects of habitats on densities for
both D. granuliferus and D. auratus (F = 20.31, df = 2, P < 0.0001; F = 28.62, df = 2, P <
0.0001, respectively).
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
2.2
Riparian Secondary Bamboo
Habitat Type
Den
sit
y (
fro
gs/
ha)
Figure 8. Average population densities of D. granuliferus (solid) and D. auratus (open) by
habitat. Vertical lines represent one standard deviation.
26

Population density trends differed through habitats between the two poison dart frog
species. The presence of D. granuliferus was nearly twice that of D. auratus in the riparian
forest, while the density of the D. auratus was more than four times larger than in the
bamboo. The population densities of both species in the secondary forest were very low,
although results indicated that D. granuliferus was sighted more often than D. auratus.
Tukey HSD tests found densities of riparian versus secondary habitats and riparian versus
bamboo habitats to be significantly different, but secondary versus bamboo habitats to be not
significant (P < 0.01, P < 0.01, P > 0.5, respectively).
A clear correlation between the number of frog sightings and proximity to water was
apparent for both species (Figure 9). Analysis with a linear regression indicated a significant
negative relationship between the number of sightings and the distances of the sighting
locations to a water source (y = -0.0476x + 13.478, df = 1, r2 = 0.207, P < 0.01). Three
particularly dense clusters of frog sightings are apparent in the riparian and secondary
habitats near streams, while sightings in the bamboo habitat did not appear to be correlated to
water (Figure 10).
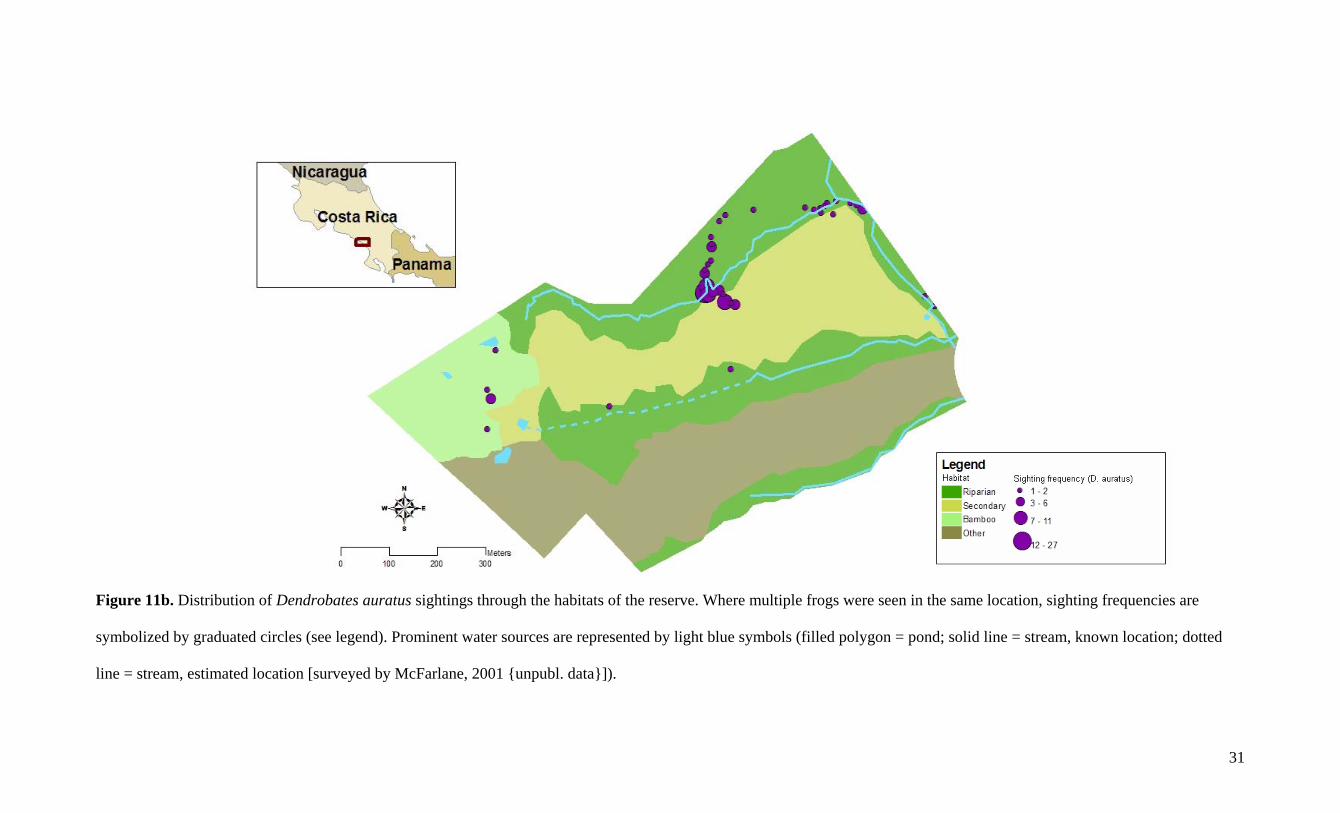
The distributions of D. granuliferus and D. auratus were generally very similar.
Individuals of both species were found simultaneously at the same locations on multiple
occasions. Only two locations throughout the study site indicated the dominating presence of
one species without the other (Figure 11). For one, there is a distinct difference in the number
of D. auratus (n = 8) found in bamboo compared to D. granuliferus (n = 2). However, the
small sample size undermines the strength of this disparity. A second conspicuous
dissimilarity in distribution occurred on the southernmost stream where the trail dips towards
the southern stream.
27
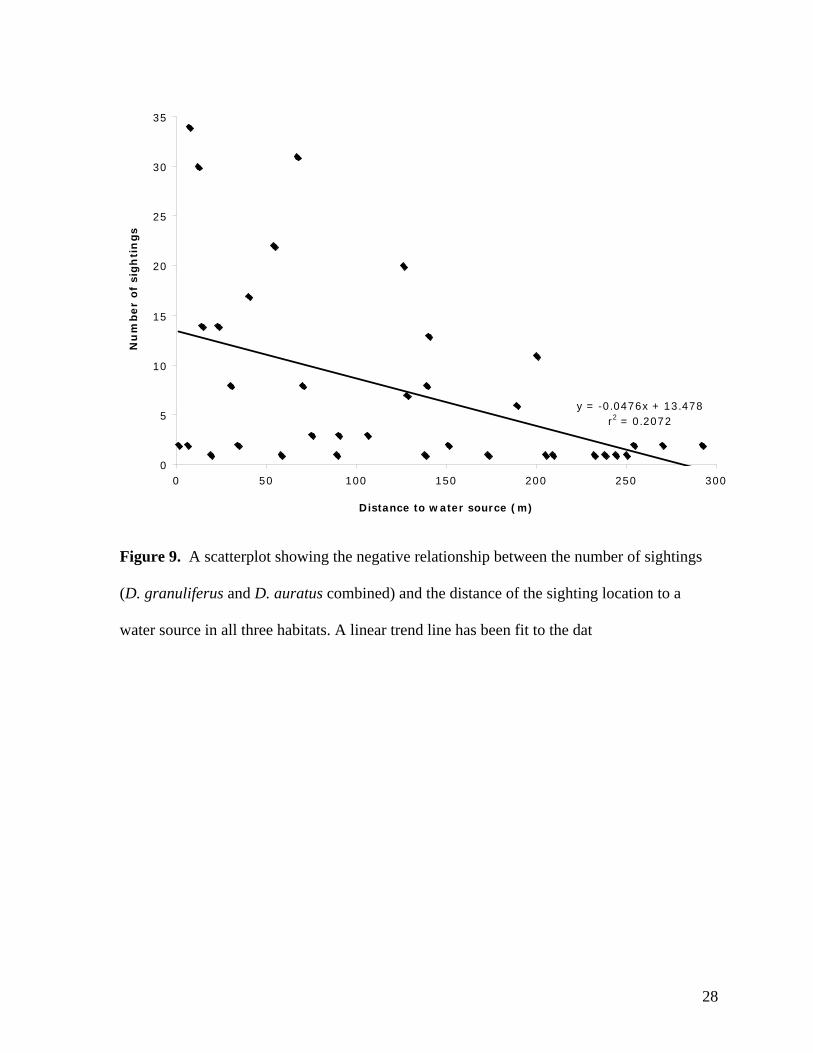
Figure 9. A scatterplot showing the negative relationship between the number of sightings
(D. granuliferus and D. auratus combined) and the distance of the sighting location to a
water source in all three habitats. A linear trend line has been fit to the dat
y = -0.0476x + 13.478r2 = 0.2072
20
25
30
35
200 250 300
)
of
sigh
tin
g
0
5
10
15
0 50 100 150
Distance to water source (m
Nu
mber
s
28

Figure 10. Distribution of all frog sightings through the habitats of the reserve (D. granuliferus and D. auratus combined). Where multiple frogs were seen in the same location,
sighting frequencies are symbolized by graduated circles (see legend). Prominent water sources are represented by blue symbols (filled polygon = pond; solid line = stream, known
location; dotted line = stream, estimated location [surveyed by McFarlane, 2001 {unpubl. data}]).
29

Figure 11a. Distribution of Dendrobates granuliferus sightings through the habitats of the reserve. Where multiple frogs were seen in the same location, sighting frequencies are
symbolized by graduated circles (see legend). Prominent water sources are represented by light blue symbols (filled polygon = pond; solid line = stream, known location; dotted
line = stream, estimated location [surveyed by McFarlane, 2001 {unpubl. data}]).
30
31
Figure 11b. Distribution of Dendrobates auratus sightings through the habitats of the reserve. Where multiple frogs were seen in the same location, sighting frequencies are
symbolized by graduated circles (see legend). Prominent water sources are represented by light blue symbols (filled polygon = pond; solid line = stream, known location; dotted
line = stream, estimated location [surveyed by McFarlane, 2001 {unpubl. data}]).
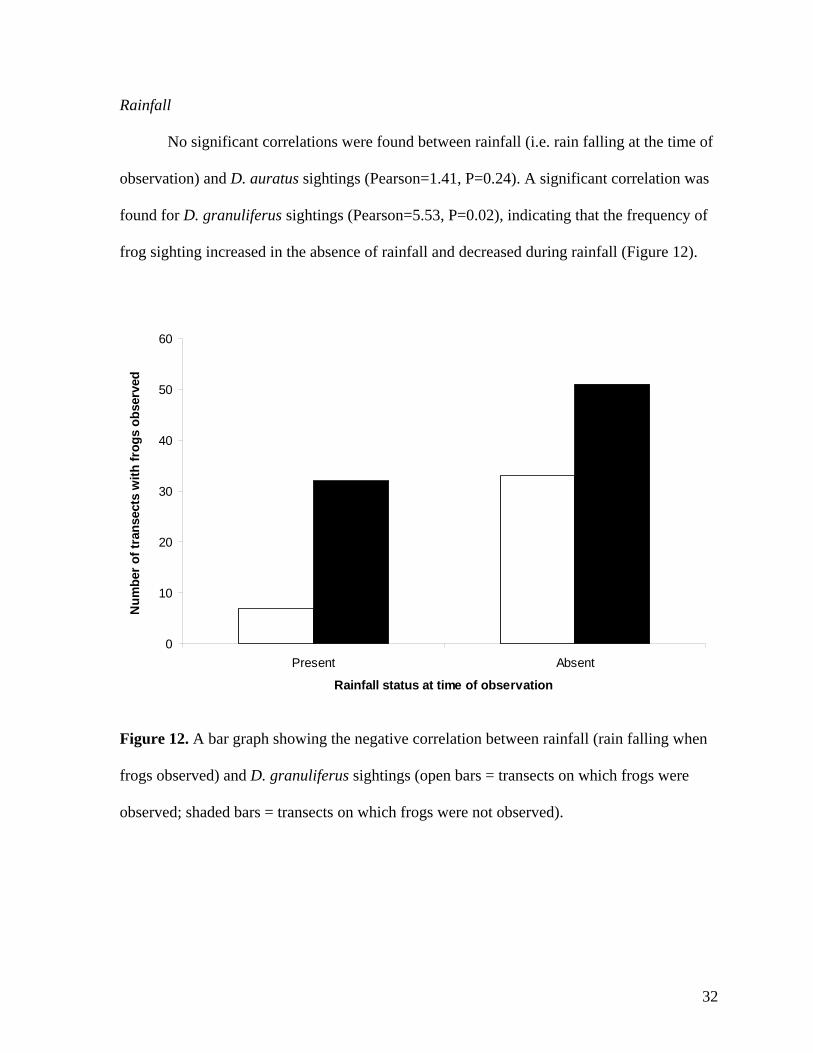
Rainfall
No significant correlations were found between rainfall (i.e. rain falling at the time of
observation) and D. auratus sightings (Pearson=1.41, P=0.24). A significant correlation was
found for D. granuliferus sightings (Pearson=5.53, P=0.02), indicating that the frequency of
frog sighting increased in the absence of rainfall and decreased during rainfall (Figure 12).
0
10
20
30
40
50
60
Present Absent
Rainfall status at time of observation
Num
ber o
f tra
nsec
ts w
ith fr
ogs
obse
rved
Figure 12. A bar graph showing the negative correlation between rainfall (rain falling when
frogs observed) and D. granuliferus sightings (open bars = transects on which frogs were
observed; shaded bars = transects on which frogs were not observed).
32

Temperature
Regression analyses found no significant relationship between temperature and frog
sightings for either D. granuliferus or D. auratus (y = -0.0031x + 0.0962, df = 17, P > 0.05,
r2 = 0.1378; y = -0.0012x + 0.0434, df = 17, P > 0.05, r2 = 0.0587, respectively). Temperature
varied only 7ºC according to the Firestone meteorological station during the time of
observation and ranged from 23ºC and 30ºC.
No large temperature differences were found between the riparian, secondary and
bamboo habitats. The temperatures of the study habitats consistently remained within 1˚ of
each other (Figure 13). In contrast, temperatures recorded in the deforested meadow
remained higher than in the study habitats, peaking at 6.6˚C higher than in the other habitats.
The temperature loggers indicated that the temperature in the three study habitats ranged
from 22.5˚C to 28˚C while the temperature in the deforested meadow ranged from 23.5˚C to
34.0˚C.
33

20
22
24
26
28
30
32
34
36
0:00
2:00
4:00
6:00
8:00
10:0
0
12:0
0
14:0
0
16:0
0
18:0
0
20:0
0
22:0
0
Time
Avera
ge a
ir t
em
pera
ture
(˚
C)
BambooSecondaryRiparianDeforested meadow
Figure 13. Mean daily temperature fluctuations in each study habitat and control
environment (deforested meadow).
Time of day
To evaluate overall trends in frog sighting frequency, data from both species were
combined and compared to the time of sighting (Figure 14). A large increase in the sighting
frequency was observed in the early morning (7:00 to 8:00), followed by relatively constant
rates in the later morning and early afternoon (8:00 to 12:00 and 13:00 to 14:00; no data was
collected from 12:00 to 13:00). From 14:00 to 15:00, a sharp decrease in sighting frequency
34
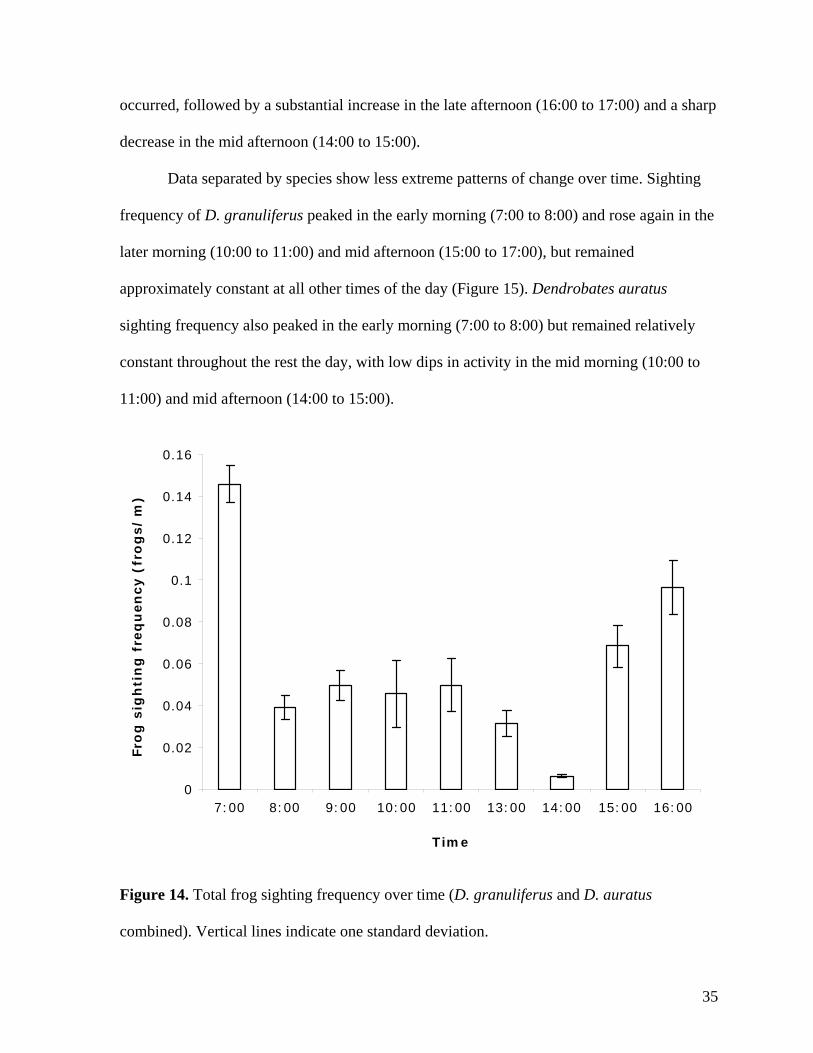
occurred, followed by a substantial increase in the late afternoon (16:00 to 17:00) and a sharp
decrease in the mid afternoon (14:00 to 15:00).
Data separated by species show less extreme patterns of change over time. Sighting
frequency of D. granuliferus peaked in the early morning (7:00 to 8:00) and rose again in the
later morning (10:00 to 11:00) and mid afternoon (15:00 to 17:00), but remained
approximately constant at all other times of the day (Figure 15). Dendrobates auratus
sighting frequency also peaked in the early morning (7:00 to 8:00) but remained relatively
constant throughout the rest the day, with low dips in activity in the mid morning (10:00 to
11:00) and mid afternoon (14:00 to 15:00).
0
0.02
0.04
0.06
0.08
0.1
0.12
0.14
0.16
7:00 8:00 9:00 10:00 11:00 13:00 14:00 15:00 16:00
Time
Fro
g s
igh
tin
g f
req
uen
cy (
fro
gs/
m)
Figure 14. Total frog sighting frequency over time (D. granuliferus and D. auratus
combined). Vertical lines indicate one standard deviation.
35

Gran/m vs. Time
0
0.01
0.02
0.03
0.04
0.05
0.06
0.07
0.08
0.09
7:00 8:00 9:00 10:00 11:00 13:00 14:00 15:00 16:00
Time
Fro
g
sig
hti
ng
fre
qu
en
cy (
fro
gs/
m)
0
0.01
0.02
0.03
0.04
0.05
0.06
0.07
0.08
0.09
7:00 8:00 9:00 10:00 11:00 13:00 14:00 15:00 16:00
Time
Fro
g s
igh
tin
g f
req
uen
cy (
fro
gs/
m)
Figure 15. Sighting frequency of D. granuliferus (top) and D. auratus (bottom) over time.
Vertical lines indicate one standard deviation.
36

DISCUSSION
Hayne Estimator Validity
The Hayne Estimator makes several assumptions that were met to the fullest extent
possible (Hayne, 1949):
(i) Animals are distributed randomly and independently throughout the area of
study.
(ii) Animals on the transect line are observed with a probability of one.
(iii) Sightings are independent events.
(iv) Animals are motionless until stimulated to flush by the observer.
(v) Each animal has a specific circle of detection in which the animal will flush
when stimulated by the observer’s presence.
(vi) The mean sighting angle between the observer’s path and the animal is 32.7˚.
(vii) Animals are not counted more than once.
(viii) Distances are measured without error.
(ix) Sighting conditions remain consistent during the study.
Assumption i challenges the instinctive nature of wild animals to cluster near
beneficial resources and when mating. Since breeding pairs of D. granuliferus and D. auratus
only come into contact for a short period during courting and amplexus (Dunn, 1941;
Summers et al., 1999), mating was not believed to substantially impact overall frog
distribution during the study. Additionally, because most organisms prefer to have easy
access to water, food, shelter and other similar necessities, animals disperse non-randomly.
However, the quantity of a given resource in a study site can be proportionally represented by
the amount of transect length that includes the particular resource. Both D. granuliferus and
37
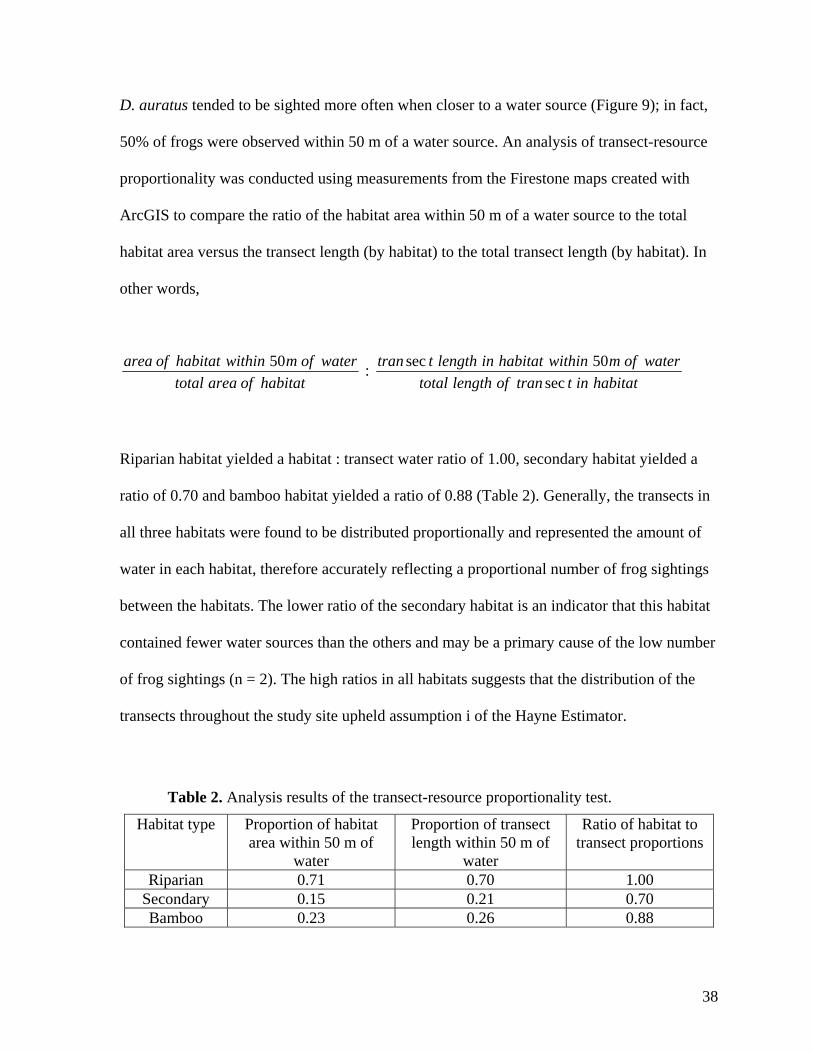
D. auratus tended to be sighted more often when closer to a water source (Figure 9); in fact,
50% of frogs were observed within 50 m of a water source. An analysis of transect-resource
proportionality was conducted using measurements from the Firestone maps created with
ArcGIS to compare the ratio of the habitat area within 50 m of a water source to the total
habitat area versus the transect length (by habitat) to the total transect length (by habitat). In
other words,
area of habitat within 50m of watertotal area of habitat
: tran sec t length in habitat within 50m of watertotal length of tran sec t in habitat
Riparian habitat yielded a habitat : transect water ratio of 1.00, secondary habitat yielded a
ratio of 0.70 and bamboo habitat yielded a ratio of 0.88 (Table 2). Generally, the transects in
all three habitats were found to be distributed proportionally and represented the amount of
water in each habitat, therefore accurately reflecting a proportional number of frog sightings
between the habitats. The lower ratio of the secondary habitat is an indicator that this habitat
contained fewer water sources than the others and may be a primary cause of the low number
of frog sightings (n = 2). The high ratios in all habitats suggests that the distribution of the
transects throughout the study site upheld assumption i of the Hayne Estimator.
Table 2. Analysis results of the transect-resource proportionality test.
Habitat type Proportion of habitat area within 50 m of
water
Proportion of transect length within 50 m of
water
Ratio of habitat to transect proportions
Riparian 0.71 0.70 1.00 Secondary 0.15 0.21 0.70 Bamboo 0.23 0.26 0.88
38

The consistency of sighting method maintained assumption ii. Because this project
was carried out in a tropical rainforest, the density of vegetation was unpredictable and
variable in different areas of the transects. Consequentially, the probability of sighting a frog
declined quickly as one moved farther away from the center of the trail. However, brush did
not tend to obstruct the transect itself and frogs sitting directly on the path were visually and,
often, audibly conspicuous.
Sightings were treated as independent events (iii) by counting each frog as an
individual observation, even if frogs were within close proximity to each other. However, the
study was conducted during the wet season, when poison dart frogs mate (Savage, 2002).
The aggregating of frogs may have altered the normal distribution of frogs throughout the
study site, but the effect of mating on the data has not been explored.
Assumptions iv and v are well suited for the study of poison dart frogs. When
approached, both D. granuliferus and D. auratus typically remained motionless or hopped
slowly until the observer was within about 3 m. At this point, the frog increased its rate of
movement until hidden by vegetation or at a location determined safe by the frog.
Occasionally frogs remained stationary despite the close proximity of the observer;
measurements in these cases were taken from the first possible sighting location as
determined by the observer. The density of vegetation may have had some impact on the
observer’s ability to see frogs, e.g. the bamboo habitat had less vegetation within 2 m of the
ground than the riparian or secondary forests. This may have biased the data, causing more
sightings to occur in the bamboo. However, only 10 frogs were observed in the bamboo,
while 261 and 4 frogs were seen in the riparian and secondary forests, respectively. The large
39

difference in frog sightings between riparian forest and bamboo habitats suggests that overall
trends are apparent regardless of the potential sighting error due to vegetation densities.
Assumption vi has been the cause of much debate because limitations in the field
cause many experiments to yield a mean sighting angle larger than 32.7˚ (Robinette et al.,
1974; Burnham et al., 1980). Alterations to the Hayne Estimator have been explored in a
number of studies (Gates, 1969; Burnham and Anderson, 1976; Burnham, 1979; Hayes and
Buckland, 1983). The Modified Hayne’s Estimator proposed by Burnham and Anderson
(1976) allows mean sighting angles of up to 45˚ through the introduction of a scalar into the
traditional Hayne Estimator. Because the mean sighting angle of all frogs in my study was
29.2 ± 4.1˚, which is not substantially different than the theoretical value of 32.7˚, the
unmodified Hayne Estimator was selected for density calculations.
Assumption vii also corresponds to the ideal transect for the Hayne Estimator: a
perfectly straight line on which animals are not counted more than once due to overlap in
observation area. While the transects were composed of curves, the sighting distances were
much smaller than segment lengths. The mean frog sighting distance was 1.2 ± 2.0 m, while
the mean surveyed trail segment length was 17.9 ± 8.1 m (COMPASS mapping software;
Fish, 2005). Since the mean sighting distance and mean trail segment length are more than
eight standard deviations apart, the trails can be treated as a series of short, straight transects.
Assumptions viii and xi were met by the study technique. Mean sighting distance was
measured using a Leica Geosystems laser rangefinder accurate to ± 3 mm from a constant
height. Magnetic bearings of the transect and frog were measured with a Suunto sighting
compass readable to ± 0.5 degrees. The low and consistent errors of these instruments
ensured the credibility of their measurements. All observations were carried out under the
40

same relative conditions. The effects of rainfall, humidity, temperature and time have been
accounted for and their influences will be discussed.
Population Densities and Distributions
Statistical results indicated that the population densities of both D. granuliferus and
D. auratus were significantly higher in riparian habitat than they were in secondary or
bamboo habitats. This is not surprising that population. Primary forest provides the natural
flora for which poison dart frogs are best adapted. Leaf litter is plentiful and provides
opportunities for water conservation, protection and foraging. Arthropod communities are
known to shift with habitat fragmentation (Gibbs and Stanton, 2001); perhaps the riparian
forest supports more preferred prey as compared to the secondary or bamboo habitats. While
in the field, I observed that the riparian forest tended to have higher canopy growth due to the
presence of more mature trees (original growth as compared to 13-year growth in secondary
and bamboo habitats) and provide more shade than did secondary or bamboo forests. These
factors may have been favored by poison dart frogs because of the decreased rate of
evaporative water loss, as well as greater buildup of leaf litter.
The densities recorded in the secondary and bamboo habitats were too low to detect a
statistically significant difference between these habitats. While no formal conclusions can be
reached, higher densities were measured in the bamboo than the secondary forest. This may
have been due to lower light penetration in the bamboo than the secondary forest, causing
leaf litter to be moister and more highly preferred by poison dart frogs. Frogs might avoid
dryer areas in favor of moister conditions where thermoregulation and moisture retention
would be less energetically expensive. The fact that the distribution of both frogs favored
41

areas close to water further supports this idea (Figure 9). Additionally, it is possible that the
bamboo habitat may provide appropriate nutrients for prey species that the secondary forest
vegetation lacks. Further study of frog densities in secondary and non-native bamboo habitats
is needed to test these hypotheses. If the difference in frog densities increases between these
two habitats with a greater sample size, then bamboo should be further explored as a
sustainable crop in Costa Rica.
The bias of both species’ distributions towards areas near water is intriguing. Three
particularly dense clusters of sightings occurred on the transects: at the intersection of several
streams near the eastern reserve border of the reserve; around the midpoint of the
northernmost east-west stream in the northern region of the reserve; and at the western end of
the southernmost east-west stream (Figure 10). All of these clusters occur near or
immediately adjacent to water sources. Previous studies have not documented poison dart
frogs as requiring large water sources for survival; rather, poison dart frogs are thought to be
unique among anurans because they utilize water from small pools of rainwater among the
leaf litter (reviewed by Savage 1968; Vences et al., 2000; Jowers and Downie, 2005). Instead
of laying eggs in permanent ponds, Dendrobatids are thought to lay their eggs on land and
carry their larvae to small pools of water in the folds of leaves, where tadpoles remain until
they become adults. This lifestyle supports random and independent distribution of poison
dart frogs throughout the Firestone Reserve. My data, however, indicate a strong correlation
between the number of sightings and the proximity to a permanent water source (Figure 9).
Other observations of poison dart frogs aggregating near large water sources have not been
previously documented in published literature.
42

A severe shortage of rainfall during October 2006 may have also influenced frog
behavior. When in Costa Rica, the author encountered many native Costa Ricans who
remarked on the atypical lack of rain. Their observations were confirmed by a simple
comparison of the total precipitation in October of 2005 and October 2006. A 502.2 cm
difference occurred between the two years, with 2005 receiving 772.4 cm and 2006 receiving
270.2 cm of rainfall (Firestone Reserve Weatherlink). The unseasonably dry weather may
have led poison dart frogs to alter their behavior, perhaps motivating them to congregate
around large water sources. The dryer environment could have stimulated some frogs to
withdraw beneath damp leaf litter as during the dry season of the year, thus reducing the
number of exposed, observable frogs and the density estimates recorded in this study.
Rainfall
Insignificant correlations between rainfall and sighting frequency for D. auratus and
significant negative correlations for D. granuliferus are inconsistent with previous
conclusions about tropical anuran species. Extensive research indicates that most frog species
increase activity during periods of rainfall (Aichinger, 1987; Duellman, 1995; Gottsberger
and Gruber, 2004). Our results conversely suggest that the activity of D. granuliferus
increased without rain and decreased with rain. The disparity between conclusions may be
caused by the limitations of study conditions. The inconsistency of rainfall during the study
period led to twice the number of transects to be observed when rain was absent than when
rain was present, which may account for the greater probability for frogs observed in the
absence of rainfall.
43

Temperature
The relative consistency of temperature during the study was expected of the tropical
setting. The average daily reserve temperature ranged only 7°C during the study period.
Observations were conducted during the daylight hours, during which the reserve
temperature range was merely 4°C. The temperature range during the observation period was
stable and moderate enough to not affect frog activity.
Considering the limited potential for temperature differences, it is not surprising that
the average daily habitat temperature of all three study habitats remained within 1°C of each
other. The only point at which a difference occurred was for the riparian habitat around 13:00
(Figure 13). At this point the temperature decreased 2°C within 2 hours, a trend that the other
two study habitats as well as the control habitat followed at a more gradual rate. The sudden
change in riparian temperature was likely due to the sun shifting and creating a completely
shaded environment around the temperature logger, or some other similar situation
unrepresentative of the overall temperature in the habitat.
The fact that the riparian, secondary and bamboo regions had approximately the same
average habitat temperature while the deforested meadow temperature averaged a higher
temperature suggests that the loggers received the same amount of sunlight in each of the
study habitats. Although field observations by the author indicated that vegetation was
densest in the riparian forest and sparsest in the secondary forest, results imply that the
amount of light penetration was approximately the same in all of the study habitats. This
finding provides an intriguing conclusion: that sunlight, or at least temperature, differences
were not the main cause of distribution bias towards the riparian forest and away from the
secondary forest. Perhaps there were substantial differences between vegetation coverage to
44

cause moisture and plant species composition differences, but not enough to cause
differences in temperature.
Time of Day
Differences in sighting frequency over the day echo the conclusions of other studies
on poison dart frog activity. Studies have shown that D. auratus has bimodal peaks of
activity around 7:00 and 17:00 (Jaeger and Hailman, 1981; Graves, 1999). High activity
levels in D. pumilio, a species whose biology is often compared to D. granuliferus, were
previously found to be limited to the morning between 8:00 and 9:00 (Graves, 1999). The
activity levels (represented by sighting frequency) of both D. granuliferus and D. auratus in
my study were found to be higher in the early morning when examined on the species level
(Figure 15). When species data were combined, a rise in activity in the late afternoon was
also observed (Figure 14).
The increase in activity during the early morning could be product of multiple factors.
Environmental conditions may be more favorable to frog activity due to higher ground
moisture levels from unevaporated nightly rainfall, lower light levels or decreased
temperature levels, leading to slower rates of evaporative water loss. Additionally, arthropod
activity may be higher during this time of the day, allowing frogs to expend less energy when
foraging (Basset et al., 2001). Finally, frogs could simply be hungry from a night spent
beneath leaf litter and commence feeding with the first morning light.
45

The Hayne Estimator as a Tool for Monitoring Amphibians
Based on the results of the study, the Hayne Estimator appears to be a useful tool for
measuring population densities of poison dart frog species. The pattern of density estimates
among habitats is consistent with the results of past studies on deforestation and population
densities. Although this study did not deal directly with positive or negative estimate bias,
many case studies have found the Hayne Estimator to overestimate population densities due
to its inability to account for a larger detection angle than 32.7º (Gates, 1969; Burnham and
Anderson, 1976; Hayes and Buckland, 1983). The results presented here did not reflect these
restrictions, further confirming the success of the Hayne Estimator field technique with
poison dart frogs in the forests of Costa Rica.
Additional studies using the Hayne Estimator and similar tools are needed to further
explore the effects of deforestation and subsequent re-growth on amphibian populations. The
results of this project suggest that poison dart frog population densities are higher in
selectively-logged riparian forests than secondary or non-native bamboo forests. Despite over
a decade of unrestricted natural regrowth, secondary forests showed substantially lower
population densities of both D. granuliferus and D. auratus. Nevertheless, the higher
presence of Dendrobatids in bamboo compared to the lack of presence in secondary forest
suggest that bamboo plantations may provide an interim solution in the process of restoring
canopy cover to deforested lands. Future studies could evaluate similar parameters in other
poison dart frog species to expand our understanding of the sensitivity of this group to habitat
destruction. Similarly, it would be relevant to examine areas of recovering forest of various
ages to investigate the length of time necessary for frogs to repopulate areas at normal
densities.
46

Conclusions
A powerful result of this study is that 13 years is an inadequate amount of time to
regenerate suitable habitat for poison dart frogs. Riparian forest most likely supported the
strongest frog presence because the selective logging preserved enough native vegetation to
support frog populations. As past studies have shown, secondary regrowth tends to feature a
distinct species composition and, as a result of reduced competition with native organisms,
an abundance of exotic species (Aide et al., 2000; Walker, 2000).
Empty and fertile from years with manure, livestock pastures are particularly
vulnerable to invasive species. A study on the vegetation species composition of 71 tropical
abandoned cattle pastures in Puerto Rico found that the density, basal area, aboveground
biomass and species richness of the secondary forest sites matched old growth forest areas
after 40 years of regeneration (Aide et al., 2000). Of the colonists, exotic species were some
of the most abundant species in the secondary forests, although not all maintained their
presence permanently. Similarly in New Zealand, an abandoned sheep and rabbit pasture
showed increased species richness and biodiversity over 4 years of monitoring, a
consequence of the introduction of exotic species (Walker, 2000). In both cases, the long-
term effect of the invasive flora depended on the species’ life-history characteristics and
abilities to persist through the rigorous competition of succession.
Despite the drastic impacts that exotic species can have on an ecosystem, there are
certain situations in which invasive species are the best alternative. In a site so badly affected
by human activity that native vegetation refuses to grow, exotics can prepare the earth for
native recolonization by increasing and stabilizing topsoil organic matter and boosting
nitrogen levels (Lamb, 1998). Such may have been the case with the bamboo plantation on
47

the Firestone Reserve, since more poison dart frogs were found in the bamboo than in the
secondary forest. Innumerable factors contribute to the outcome of forest recovery, the most
significant often being land-use history, time since abandonment, vegetation cover, rate and
type of seed dispersal, and spatial variables such as elevation and slope (Aragon and Morales,
1988; Holl, 1999). Managers must carefully weigh the unique qualities of each site, for they
may have important effects on the presence of sensitive groups such as poison dart frogs and
should serve as important criteria in predicting the success of recovering ecosystems to
maintain biodiversity.
Perhaps the most significant message from this study’s results is that the
reestablishment of biodiversity takes time. If the densities and distributions of D.
granuliferus and D. auratus serve as accurate indicators of the overall fauna of the reserve,
then it is clear that the Firestone Reserve is at the very beginning of a long process, despite
having been dedicated to natural regeneration for over a decade. How long must we wait
before we can accurately reclassify an area as natural? The answer will most likely never be
finite or universal, but rather unique to each restoration site. With the help of monitoring
programs that track regeneration over time, we will continue to refine our understanding of
reforestation and restore or, at the very least, stabilize biodiversity in these areas.
48

ACKNOWLEDGEMENTS
I am deeply appreciative of Professor McFarlane for his inspiration, guidance, humor
and endless support through my intellectual journey. I am also thankful of Professor Preest
for her assistance with editing, methodology and equipment and Professor Thomson for her
guidance with statistical analysis. My intimacy with Costa Rican poison dart frogs could not
have occurred without financial support from the Claremont McKenna College Dean of
Students, the Roberts Environmental Center, the Claremont Colleges Joint Science
Department and the Firestone Reserve. Additionally, I express my thanks to Carol Brandt of
the Pitzer College Costa Rica Study Abroad Program for accommodating me at the Firestone
Center while I conducted research. I extend my appreciation to the staff of Joint Science and,
in particular, the Organismal Biology Department for providing me with the biological
foundation to create this thesis. Finally, thanks to my family and friends for their
encouragement through all.
49

LITERATURE CITED
Aichinger, M. 1987. Annual activity patterns of anurans in a seasonal neotropical
environment. Oecologia 71(4):583-592.
Aide, T. M., and H. R. Grau. 2004. Globalization, Migration, and Latin American
Ecosystems. Science 305:1915-1916.
Aide, T. M., J. K. Zimmerman, J. B. Pascarella, L. Rivera, and H. Marcano-Vega. 2000.
Forest regeneration in a chronosequence of tropical abandoned pastures: Implications
for restoration ecology. Restor. Ecol. 8(4):328-338.
Anderson, M. T., and A. Mathis. 1999. Diets of two sympatric neotropical salamanders,
Bolitoglossa mexicana and B. rufescens, with notes on reproduction for B. rufescens.
J. Herpetol. 33(4):601-607.
Aragon, R., and J. M. Morales. 2003. Species composition and invasion in Argentinean
secondary forests: Effects of land use history, environment and landscape. J. Veget.
Sci. 14:185-204.
Basset, Y., H. Alberlenc, H. Barrios, G. Curletti, J. Berenger, J. Vesco, P. Causse, A. Haug,
A. Hennion, L. Lesobre, F. Marques, and R. O’Meara. 2001. Stratification and diel
activity of arthropods in a lowland rainforest in Gabon. Biological Journal of the
Linnean Society 72:585-607.
Brooks, T. M., R. A. Mittermeier, C. G. Mittermeier, G. A. B. da Fonseca, A. B. Rylands, W.
R. Konstant, P. Flick, J. Pilgrim, S. Oldfield, G. Magin, and C. Hilton-Taylor. 2002.
Habitat Loss and Extinction in the Hotspots of Biodiversity. Conservat. Biol.
16(4):909.
50

Burnham, K.P. 1979. A parametric generalization of the Hayne Estimator of line transect
sampling. Biometrics 35(3):587-595.
Burnham, K.P., and D.R. Anderson. 1976. Mathematical models for nonparametric
inferences from line transect data. Biometrix 32(2):325-336.
Burnham, K.P., D.R. Anderson, and J.L. Laake. 1980. Estimation of density from line
transect sampling of biological populations. Wildlife Monograph No. 72 Supplement
to Journal of Wildlife Management 44.
Conservation International. 2007. <http://www.biodiversityhotspots.org>. Accessed February
2007.
Coulson, G. M., and J. A. Raines. 1985. Methods of small-scale surveys of grey kangaroo
populations. Australian Wildlife Research 12(2):119-125.
Donoghue, M. J., and W. S. Alverson. 2000. A new age of discovery. Ann. Mo. Bot. Gard.
87(1):110-126.
Duellman, W.E. 1995. Temporal fluctuations in abundances of Anuran amphibians in a
seasonal Amazonian rainforest. Journal of Herpetology 29(1):13-21.
Dunn, E. R. 1941. Notes on Dendrobates auratus. Copeia 2:88-93.
Firestone Center for Restoration Ecology. 2006. <http://costarica.jsd.claremont.edu>.
Accessed 6 January 2006.
Fish, L. 2005. COMPASS v5.05. Fountain Computer. <http://fountainware.com/compass/>.
Food and Agriculture Organization of the United Nations. 2004. <http://faostat.fao.org>.
Accessed May 2004.
Gates, C.E. 1969. Simulation study of estimators for the line sampling method. Biometrics
25(2):317-328.
51

Gibbs, J. P., and E. J. Stanton. 2001. Habitat fragmentation and arthropod community
change: carrion beetles, phoretic mites, and flies. Ecol. Appl. 11(1):79-85.
Global Amphibian Assessment 2006. <http://www.globalamphibians.org>. Accessed
December 2006.
Gottsberger, B., and E. Gruber. 2004. Temporal partitioning of reproductive activity in a
neotropical anuran community. Journal of Tropical Ecology 20:271-280.
Graves, B.M. 1999. Diel activity patterns of the sympatric Poison Dart Frogs, Dendrobates
auratus and D. pumilio, in Costa Rica. Journal of Herpetology 33(3):375-381.
Hacienda Baru National Wildlife Refuge. <http://www.haciendabaru.com>. Accessed 6
January 2006.
Hayes, R.J., and S.T. Buckland. 1983. Radial-distance models for the line-transect method.
Biometrics 39(1):29-42.
Hayne, D. W. 1949. An examination of the strip census method for estimating and animal
populations. J. Wildl. Mange. 13:145.
Holl, K. D. 1999. Factors limiting tropical rain forest regeneration in abandoned pasture:
Seed rain, seed germination, microclimate, and soil. Biotropica 31(2):229-242.
International Union for conservation of Nature and Natural Resources, Conservation
International and NatureServe. 2006. Global Amphibian Assessment.
<www.globalamphibians.org>. Accessed December 2006.
International Union for Conservation of Nature and Natural Resources. 2006. 2006 IUCN
Red List of Threatened Species. < http://www.iucnredlist.org/>. Accessed 6 January
2006.
52

Jaeger, R.G., and J.P. Hailman. 1981. Activity of neotropical frogs in relation to ambient
light. Biotropica 13(1):59-65.
Jowers, M. J., and J. R. Downie. 2005. Tadpole deposition behaviour in male stream frogs
Mannophryne trinitatis (Anura: Dendrobatidae). J. Nat. Hist. 39(32):3013-3027.
Lamb, D. 1998. Large-scale ecological restoration of degraded tropical forest lands: The
potential role of timber plantations. Restor. Ecol. 6(3):271-279.
Landres, P. B., J. Verner, and J. W. Thomas. 1988. Ecological uses of vertebrate indicator
species: A critique. Conservat. Biol. 2(4):316-328.
Lowry, R. 2007. VassarStats: Web Site for Statistical Computation.
<http://faculty.vassar.edu/lowry/VassarStats.html>. Accessed February 2007.
Maxon, L. R., and C. W. Myers. 1985. Albumin evolution in tropical poison frogs
(Dendrobatidae): A preliminary report. Biotropica 17(1):50-56.
Mendelson III, J. R., K. R. Lips, R. W. Gagliardo, G. B. Rabb, J. P. Collins, J. E.
Diffendorfer, P. Daszak, R. Ibáñez, K. C. Zippel, D. P. Lawson, K. M. Wright, S. N.
Stuart, C. Gascon, H. R. da Silva, P. A. Burrowes, R. L. Joglar, E. La Marca, S.
Lötters, L. H. du Preez, C. Weldon, A. Hyatt, J. V. Rodriguez-Mahecha, S. Hunt, H.
Robertson, B. Lock, C. J. Raxworthy, D. R. Frost, R. C. Lacy, R. A. Alford, J. A.
Campbell, G. Parra-Olea, F. Bolaños, J. J. C. Domingo, T. Halliday, J. B. Murphy, M.
H. Wake, L. A. Coloma, S. L. Kuzmin, M. S. Price, K. M. Howell, M. Lau, R.
Pethiyagoda, M. Boone, M. J. Lannoo, A. R. Blaustein, A. Dobson, R. A. Griffiths,
M. L. Crump, D. B. Wake, and E. D. Brodie Jr. 2006. Confronting amphibian
declines and extinctions. Science 313:48.
Myers, N., R. A. Mittermeier, C. G. Mittermeier, G. A. B. da Fonseca, and J. Kent. 2000.
53

Biodiversity hotspots for conservation priorities. Nature 403:853–858.
National Biodiversity Institute. 2007. Biodiversity in Costa Rica.
<http://www.inbio.ac.cr/en/biod/bio_biodiver.htm>. Accessed January 2007.
Pelletier, L., and C. J. Krebs. 1997. Line-transect sampling for estimating ptarmigan
(Lagopus spp.) density. Canadian Journal of Zoology 75(8):1185-1192.
Phillips, K. 1990. Where have all the frogs and toads gone? BioScience 40(6):422-424.
Pounds, J.A., M. R. Bustamante, L. A. Coloma, J. A. Consuegra, M. P. L. Fogden, P. N.
Foster, E. La Marca, K. L. Masters, A. Merino-Viteri, R. Puschendorf, S. R. Ron, G.
A. Sánchez-Azofeifa, C. J. Still, and B. E. Young. 2006. Widespread amphibian
extinctions from epidemic disease driven by global warming. Nature 439:161-
167.
Robinette, W.L., C.M. Loveless, and D.A. Jones. 1974. Field tests of strip census methods.
Journal of Wildlife Management 38:81-96.
Saporito, R. A., H. M. Garraffo, M. A. Donnelly, A. L. Edwards, J. T. Longino, and J. W.
Daly. 2004. Formicine ants: an arthropod source for the pumiliotoxin alkaloids of
dendrobatid poison frogs. Proc. Natl. Acad. Sci. USA 101(25): 8045-8050.
Savage, J. M. 1968. The Dendrobatid frogs of Central America. Copeia 2:745-776.
Savage, J. M. 2002. The amphibians and reptiles of Costa Rica. London: The University of
Chicago Press. 382-386.
Stuart, S. N., J. S. Chanson, N. A. Cox, B. E. Young, A. S. L. Rodrigues, D. L. Fischman,
and R. W. Waller. 2004. Status and trends of amphibian decline sand extinctions
worldwide. Science 306(5702):1783-1786.
54

55
Summers, K., L. A. Weigt, P. Boag, and Bermingham, E. 1999. The evolution of female
parental care in poison frogs of the genus Dendrobates: evidence from mitochrondrial
DNA sequences. Herpetological 55(2):254-263.
Toft, C. A. 1981. Feeding ecology of Panamanian litter anurans: Patterns in diet and foraging
mode. J. Herpetol. 15(2):139-144.
Vences, M., J. Kosuch, S. Lotters, A. Widmer, K.-H. Jungfer, J. Kohler, and M. Veith. 2000.
Phylogeny and classification of poison frogs (Amphibia: Dendrobatidae), based on
mitochondrial 16S and 12S ribosomal RNA gene sequences. Mol. Phylogenet Evol.
15(1):34-40.
Walker, S. 2000. Post-pastoral changes in composition and guilds in a semi-arid conservation
area, Central Otago, New Zealand. N. Z. J. Ecol. 24(2):123-137.
Wildlife Conservation Society. 2007. The new monkey and Dr. Rob Wallace.
<http://www.wcs.org/international/latinamerica/centralandes/nwbolivia/madidimonke
y/madidimonkey_description>. Accessed 11 January 2007.
World Resources Institute. 2006. Biodiversity and protected areas: country profile – Costa
Rica. <http://earthtrends.wri.org/text/biodiversity-protected/country-profile-43.html>.
Accessed January 2007.
Young, B. E., K. R. Lips, J. K. Reaser, R. Ibáñez, A. W. Salas, J. R. Cedeño, L. A. Coloma,
S. Ron, E. LaMarca, J. R. Meyer, A. Muñoz, F. Bolaños, G. Chavez, and D. Romo.
2001. Population declines and priorities for amphibian conservation in Latin America.
Conservat. Biol. 15:1213–1223.
APPENDICES
Appendix A. Frog sighting data from 9 October to 14 October 2006. For species, Gran = Dendrobates granuliferus (granular poison
dart frog) and GB = Dendrobates auratus (green and black poison dart frog).
Date
Transect
Start Time
End Time
Observ. Time
Species
Frog Bearing (˚)
Path Bearing (˚)
Sighting Angle (˚)
Modified Dist. (m) Location Habitat
10/9 1 7:00 7:1310/9 6 7:13 7:20 7:17 GB 313 284 29 1.18 C71 Riparian10/9 6 7:13 7:20 Gran 320 311 9 0.90 C71 Riparian10/9 3 7:20 8:13 7:28 Gran 36 53 17 1.18 C25 Riparian10/9 3 7:20 8:13 7:46 Gran 26 53 27 1.08 C25 Riparian10/9 3 7:20 8:13 7:46 Gran 138 225 87 0.54 C26 Riparian10/9 3 7:20 8:13 7:48 Gran 241 216 25 1.21 C26 Riparian
10/9 3 7:20 8:13 7:49 Gran 144 186 42 0.90west of
C26 Riparian10/9 3 7:20 8:13 7:50 Gran 172 153 19 1.04 C27 Riparian10/9 3 7:20 8:13 7:52 GB 193 156 37 0.84 C27 Riparian10/9 3 7:20 8:13 7:53 Gran 258 208 50 0.95 C28 Riparian10/9 3 7:20 8:13 8:00 Gran 110 120 10 0.38 C29 Riparian10/9 3 7:20 8:13 8:01 Gran 195 185 10 1.26 C29-C30 Riparian10/9 3 7:20 8:13 8:02 GB 229 203 26 1.29 C31 Riparian10/9 3 7:20 8:13 8:05 GB 221 203 18 1.59 C31 Riparian10/9 4 8:15 8:47 8:24 Gran 181 232 51 0.99 B24 Riparian10/9 4 8:15 8:47 8:26 Gran 198 230 32 1.07 B24 Riparian10/9 4 8:15 8:47 8:27 Gran 286 230 56 1.07 B24 Riparian10/9 4 8:15 8:47 8:28 Gran 293 230 63 1.38 B24 Riparian10/9 4 8:15 8:47 8:30 Gran 262 230 32 1.61 B24 Riparian10/9 4 8:15 8:47 8:32 Gran 196 240 44 1.04 B23 Riparian
56

Date Transect Start Time
End Time
Observ. Time Species
Frog Bearing (˚)
Path Bearing (˚)
Sighting Angle (˚)
Modified Dist. (m) Location Habitat
10/9 4 8:15 8:47 8:32 Gran 196 240 44 1.04 B23 Riparian10/9 4 8:15 8:47 8:34 Gran 216 248 32 1.24 B23 Riparian10/9 5 8:44 9:02 8:47 GB 239 265 26 1.22 BB24 Bamboo
10/9 4 9:02 9:30 9:10 Gran 255 213 42 1.27B23 (10m
uphill) Riparian10/9 4 9:02 9:30 9:12 Gran 85 65 20 1.29 B23 Riparian10/9 4 9:02 9:30 9:14 Gran 7 29 22 1.24 B24 Riparian10/9 4 9:02 9:30 9:15 Gran 3 52 49 1.15 B24 Riparian
10/9 4 9:02 9:30 9:16 Gran 90 59 31 1.34B24 (20m downhill)
Riparian
10/9 4 9:02 9:30 9:18 Gran 53 73 20 1.24 B28 Secondary10/9 4 9:02 9:30 9:18 Gran 9 73 64 0.96 B28 Secondary10/9 2 9:30 9:50 9:47 GB 319 12 53 1.51 B92 Riparian10/9 2 9:30 9:50 9:48
GB 43 68 25 1.20 B92 Riparian
10/9 1 9:50 10:0210/9 1 16:16 16:28 16:20 GB 234 300 66 1.46 WT2 Riparian10/9 6 16:28 16:35 16:30 Gran 14 326 48 1.17 C72 Riparian10/9 6 16:28 16:35 16:31 Gran 340 322 18 1.11 C72 Riparian10/9 6 16:28 16:35 16:31 Gran 343 293 50 0.96 C72 Riparian10/9 3 16:35 17:14 16:35 GB 221 203 18 1.54 C31 Riparian10/9 3 16:35 17:14 16:40 Gran 221 203 18 1.63 C31 Riparian10/9 3 16:35 17:14 16:44 Gran 240 178 62 1.39 C30 Riparian10/9 3 16:35 17:14 16:49 Gran 132 170 38 1.27 C31 Riparian10/9 3 16:35 17:14 16:50 GB 230 258 28 1.36 C40 Riparian10/9 3 16:35 17:14 16:51 GB 288 279 9 1.15 C53 Riparian10/9 3 16:35 17:14 16:53 GB 320 305 15 0.97 C56 Riparian10/9 3 16:35 17:14 17:00 Gran 322 7 45 1.22 C26 Riparian10/9 3 16:35 17:14 17:00 Gran 324 340 16 1.68 C26 Riparian
57
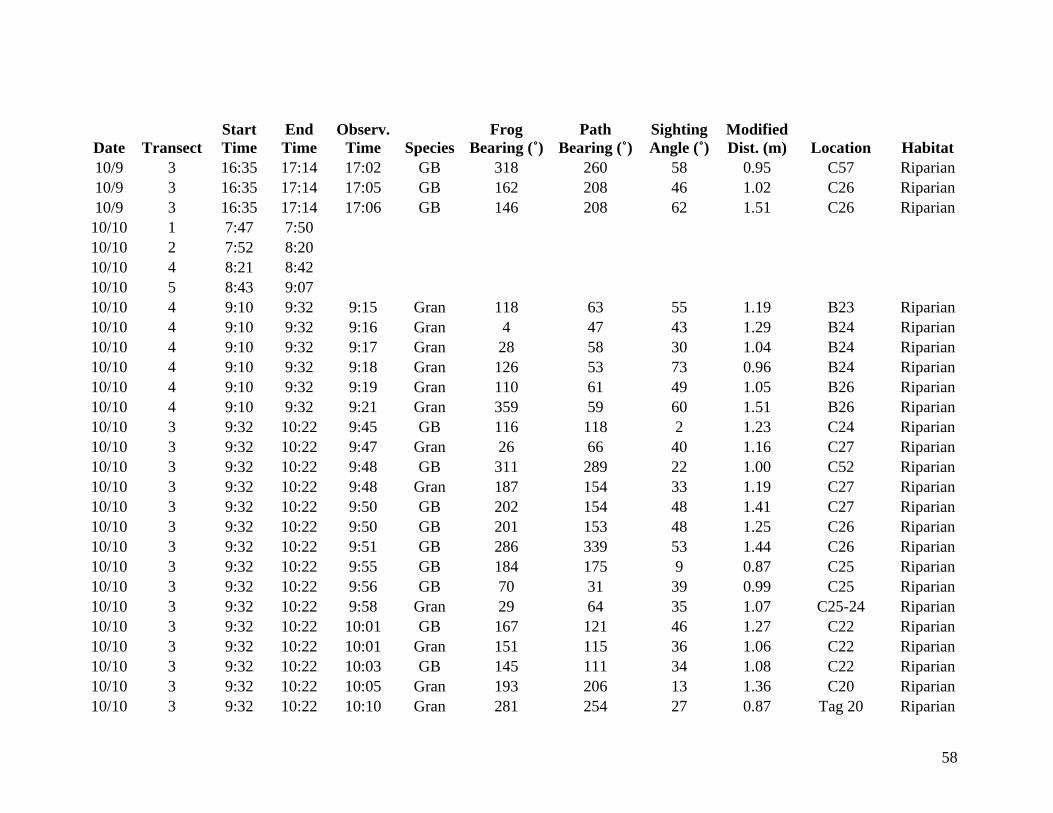
Date Transect Start Time
End Time
Observ. Time Species
Frog Bearing (˚)
Path Bearing (˚)
Sighting Angle (˚)
Modified Dist. (m) Location Habitat
10/9 3 16:35 17:14 17:02 GB 318 260 58 0.95 C57 Riparian10/9 3 16:35 17:14 17:05 GB 162 208 46 1.02 C26 Riparian10/9 3 16:35 17:14 17:06
GB 146 208 62 1.51 C26 Riparian
10/10 1 7:47 7:5010/10 2 7:52 8:2010/10 4 8:21 8:4210/10 5 8:43 9:0710/10 4 9:10 9:32 9:15 Gran 118 63 55 1.19 B23 Riparian10/10 4 9:10 9:32 9:16 Gran 4 47 43 1.29 B24 Riparian10/10 4 9:10 9:32 9:17 Gran 28 58 30 1.04 B24 Riparian10/10 4 9:10 9:32 9:18 Gran 126 53 73 0.96 B24 Riparian10/10 4 9:10 9:32 9:19 Gran 110 61 49 1.05 B26 Riparian10/10 4 9:10 9:32 9:21 Gran 359 59 60 1.51 B26 Riparian10/10 3 9:32 10:22 9:45 GB 116 118 2 1.23 C24 Riparian10/10 3 9:32 10:22 9:47 Gran 26 66 40 1.16 C27 Riparian10/10 3 9:32 10:22 9:48 GB 311 289 22 1.00 C52 Riparian10/10 3 9:32 10:22 9:48 Gran 187 154 33 1.19 C27 Riparian10/10 3 9:32 10:22 9:50 GB 202 154 48 1.41 C27 Riparian10/10 3 9:32 10:22 9:50 GB 201 153 48 1.25 C26 Riparian10/10 3 9:32 10:22 9:51 GB 286 339 53 1.44 C26 Riparian10/10 3 9:32 10:22 9:55 GB 184 175 9 0.87 C25 Riparian10/10 3 9:32 10:22 9:56 GB 70 31 39 0.99 C25 Riparian10/10 3 9:32 10:22 9:58 Gran 29 64 35 1.07 C25-24 Riparian10/10 3 9:32 10:22 10:01 GB 167 121 46 1.27 C22 Riparian10/10 3 9:32 10:22 10:01 Gran 151 115 36 1.06 C22 Riparian10/10 3 9:32 10:22 10:03 GB 145 111 34 1.08 C22 Riparian10/10 3 9:32 10:22 10:05 Gran 193 206 13 1.36 C20 Riparian10/10 3 9:32 10:22 10:10 Gran 281 254 27 0.87 Tag 20 Riparian
58

Date Transect Start Time
End Time
Observ. Time Species
Frog Bearing (˚)
Path Bearing (˚)
Sighting Angle (˚)
Modified Dist. (m) Location Habitat
(50m uphill)
10/10 3 9:32 10:22 10:11 Gran 203 279 76 0.93 C52 Riparian10/10 3 9:32 10:22 10:20 Gran 340 330 10 1.05 C46 Riparian10/10 3 9:32 10:22 10:21 Gran 150 185 35 0.80 C37 Riparian10/10 3 9:32 10:22 10:21 Gran 162 191 29 0.66 C37 Riparian
10/10 3 9:32 10:22 9:41 Gran 132 157 25 1.05Tag 18
(50m past)
Riparian 10/10 3 9:32 10:22 9:44 Gran 217 118 99 0.86 C29 Riparian10/10 6 10:23 10:35 10:25 Gran 93 105 12 1.10 C71 Riparian10/10 6 10:23 10:35 10:28 Gran 73 105 32 0.80 C71-C72 Riparian
10/10 6 10:23 10:35 10:30 Gran 62 111 49 0.93
C72-71(closer
to 72) Riparian 10/10 6 10:23 10:35 10:31 Gran 100 125 25 0.48 C72-71 Riparian10/10 3 10:35 11:00 10:50 Gran 183 150 33 1.03 C26 Riparian10/10 3 11:01 11:50 11:10 Gran 45 55 10 1.15 C25 Riparian10/10 3 11:01 11:50 11:13 Gran 150 141 9 0.94 C23 Riparian10/10 3 11:01 11:50 11:14 Gran 202 176 26 0.50 C22 Riparian10/10 3 11:01 11:50 11:15 GB 113 103 10 0.85 C22-21 Riparian10/10 3 11:01 11:50 11:16 GB 348 311 37 0.83 C23 Riparian
10/10 3 11:01 11:50 11:17 Gran 276 247 29 0.51
C24-C25 (closer to
C24) Riparian10/10 3 11:01 11:50 11:17 Gran 279 257 22 1.00 C25 Riparian10/10 3 11:01 11:50 11:17 Gran 335 32 57 0.97 C26 Riparian10/10 3 11:01 11:50 11:20 GB 66 54 12 1.44 C26 Riparian10/10 3 11:01 11:50 11:20 Gran 65 54 11 1.43 C26 Riparian
59

Date Transect Start Time
End Time
Observ. Time Species
Frog Bearing (˚)
Path Bearing (˚)
Sighting Angle (˚)
Modified Dist. (m) Location Habitat
10/10 3 11:01 11:50 11:23 Gran 46 22 24 1.12C27 (100m
down) Riparian10/10 3 11:01 11:50 11:23 GB 333 351 18 1.23 C27 Riparian10/10 3 11:01 11:50 11:28 Gran 52 45 7 1.32 C27 Riparian10/10 3 11:01 11:50 11:30 GB 53 28 25 1.27 C27 Riparian10/10 3 11:01 11:50 11:30 GB 282 308 26 1.00 C28 Riparian10/10 3 11:01 11:50 11:31 GB 355 357 2 0.86 C29-C30
Riparian
10/10 3 11:01 11:50 11:39 Gran 59 44 15 1.62 C31 Riparian10/10 3 11:01 11:50 11:39 GB 49 47 2 1.62 C31 Riparian10/10 3 11:01 11:50 11:39 GB 51 22 29 1.23 C31-Tag18 Riparian10/10 3 11:01 11:50 11:39 GB 72 49 23 0.85 Tag18-C32
Riparian
10/10 3 11:01 11:50 11:40 Gran 52 47 5 1.04 C38 Riparian10/10 3 11:01 11:50 11:40 GB 61 51 10 1.30 C28 Riparian
10/10 3 11:01 11:50 11:40 GB 76 92 16 0.89
C51-C53 (30m from
C53) Riparian10/10 3 11:01 11:50 11:47 Gran 142 121 21 1.04 C56 Riparian10/10 3 11:01 11:50 11:49 Gran 149 121 28 0.91 C56 Riparian10/10 6 11:50 11:56 11:51 GB 349 323 26 0.94 C70 Riparian10/10 6 11:50 11:56 11:55
Gran 247 300 53 0.91 C72-71 Riparian
10/10 1 11:56 12:0210/10 1 14:57 15:0510/10 6 15:12 15:16 15:14 Gran 320 298 22 1.00 C72-71 Riparian10/10 6 15:12 15:16 15:16 GB 187 222 35 0.97 Tag 20 Riparian10/10 3 15:16 15:50 15:40 Gran 317 317 0 0.70 C55 Riparian10/10 3 15:16 15:50 15:41 GB 151 211 60 0.99 C26 Riparian10/10 3 15:16 15:50 15:43 Gran 49 98 49 0.90 C26 Riparian10/10 3 15:16 15:50 15:46 GB 75 111 36 1.20 C25 Riparian
60
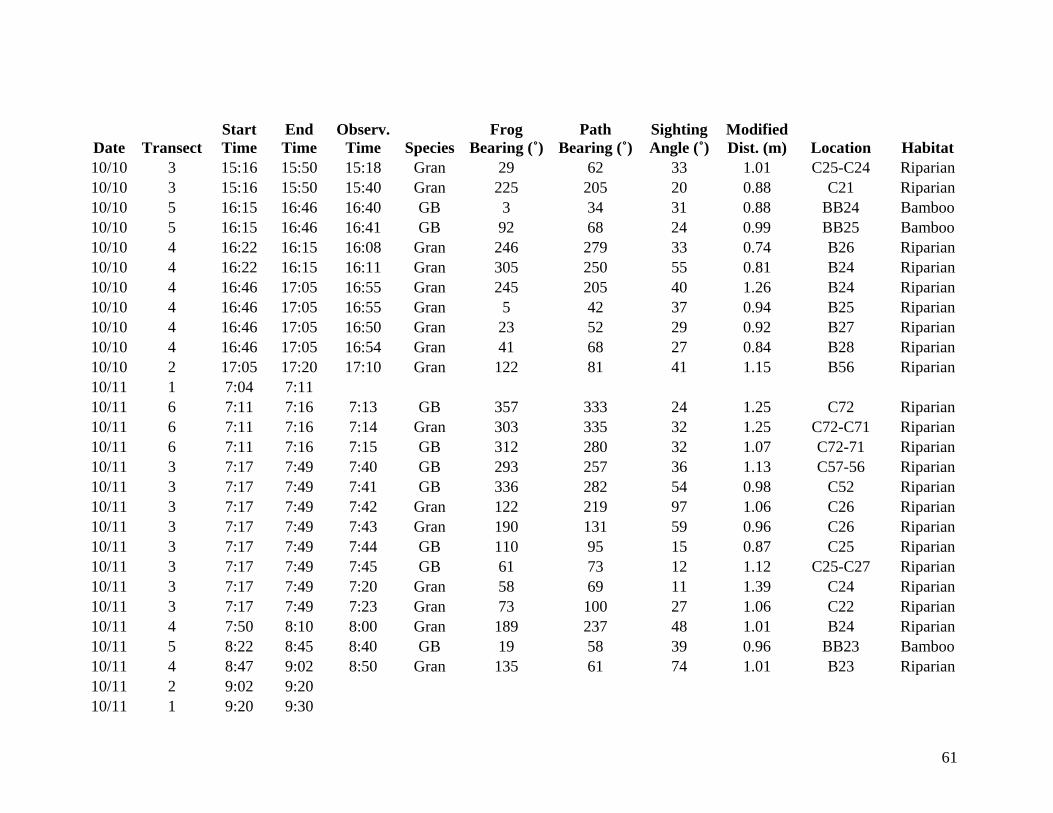
Date Transect Start Time
End Time
Observ. Time Species
Frog Bearing (˚)
Path Bearing (˚)
Sighting Angle (˚)
Modified Dist. (m) Location Habitat
10/10 3 15:16 15:50 15:18 Gran 29 62 33 1.01 C25-C24
Riparian10/10 3 15:16 15:50 15:40 Gran 225 205 20 0.88 C21 Riparian10/10 5 16:15 16:46 16:40 GB 3 34 31 0.88 BB24 Bamboo10/10 5 16:15 16:46 16:41 GB 92 68 24 0.99 BB25 Bamboo10/10 4 16:22 16:15 16:08 Gran 246 279 33 0.74 B26 Riparian10/10 4 16:22 16:15 16:11 Gran 305 250 55 0.81 B24 Riparian10/10 4 16:46 17:05 16:55 Gran 245 205 40 1.26 B24 Riparian10/10 4 16:46 17:05 16:55 Gran 5 42 37 0.94 B25 Riparian10/10 4 16:46 17:05 16:50 Gran 23 52 29 0.92 B27 Riparian10/10 4 16:46 17:05 16:54 Gran 41 68 27 0.84 B28 Riparian10/10 2 17:05 17:20 17:10
Gran 122 81 41 1.15 B56 Riparian
10/11 1 7:04 7:1110/11 6 7:11 7:16 7:13 GB 357 333 24 1.25 C72 Riparian10/11 6 7:11 7:16 7:14 Gran 303 335 32 1.25 C72-C71 Riparian10/11 6 7:11 7:16 7:15 GB 312 280 32 1.07 C72-71 Riparian10/11 3 7:17 7:49 7:40 GB 293 257 36 1.13 C57-56 Riparian10/11 3 7:17 7:49 7:41 GB 336 282 54 0.98 C52 Riparian10/11 3 7:17 7:49 7:42 Gran 122 219 97 1.06 C26 Riparian10/11 3 7:17 7:49 7:43 Gran 190 131 59 0.96 C26 Riparian10/11 3 7:17 7:49 7:44 GB 110 95 15 0.87 C25 Riparian10/11 3 7:17 7:49 7:45 GB 61 73 12 1.12 C25-C27
Riparian
10/11 3 7:17 7:49 7:20 Gran 58 69 11 1.39 C24 Riparian10/11 3 7:17 7:49 7:23 Gran 73 100 27 1.06 C22 Riparian10/11 4 7:50 8:10 8:00 Gran 189 237 48 1.01 B24 Riparian10/11 5 8:22 8:45 8:40 GB 19 58 39 0.96 BB23 Bamboo10/11 4 8:47 9:02 8:50
Gran 135 61 74 1.01 B23 Riparian
10/11 2 9:02 9:2010/11 1 9:20 9:30
61

Date
3
Start Time
End Time
Observ. Time
Species
Frog Bearing (˚)
Path Bearing (˚)
Sighting Angle (˚)
Modified Transect Dist. (m) Location Habitat
10/11 1 13:37 13:4210/11 6 13:42 13:46
10/11 3 13:46 14:15 13:46 GB 193 203 10 1.02
Tag 20 (10m on
transect 3)
Riparian 10/11 13:46 14:15 14:02 Gran 170 201 31 0.90 C26 Riparian10/11 3 13:46 14:15 14:02 Gran 113 181 68 0.94 C25 Riparian10/11 3 13:46 14:15 14:03 Gran 72 78 6 1.06 C24 Riparian10/11 3 13:46 14:15 14:10
GB 20 84 64 1.12 C24 Riparian
10/11 4 14:16 14:3110/11 5 14:31 14:58 14:43
GB 76 50 26 1.16 BB44 Bamboo
10/11 4 14:58 15:1210/11 2 15:12 15:2610/11 1 15:26 15:3110/12 1 7:05 7:1210/12 2 7:12 7:3310/12 4 7:35 7:56 7:46 Gran 220 251 31 1.15 B27 Riparian10/12 4 7:35 7:56 7:46 Gran 212 265 53 1.24 B27-B26
Riparian
10/12 4 7:35 7:56 7:47 Gran 340 261 79 0.80 B26 Riparian10/12 4 7:35 7:56 7:48 Gran 336 261 75 1.17 B26 Riparian10/12 4 7:35 7:56 7:44 Gran 189 233 44 1.07 C26 Riparian10/12 4 7:35 7:56 7:45 Gran 290 230 60 1.09 B26 Riparian10/12 5 8:00 8:27 8:14 Gran 193 193 0 0.70 Bamboo10/12 4 8:27 8:46 8:35 Gran 89 59 30 1.24 B24-B26
Riparian
10/12 4 8:27 8:46 8:37 Gran 114 65 49 1.12 B26 Riparian10/12 4 8:27 8:46 8:38 Gran 28 86 58 0.97 B27 Riparian10/12 3 8:47 9:43 9:00 GB 252 278 26 0.65 C21 Riparian10/12 3 8:47 9:43 9:00 Gran 252 278 26 0.62 C21 Riparian
62

Date
Riparian
Transect
Start Time
10/12
End
3
Time
8:47
Observ.
9:43
Time
8:55
Species
Gran
Frog
74
Bearing (˚)
105
Path
31
Bearing (˚)
0.95
Sighting
C51-C52
Angle (˚)
Riparian
Modified Dist. (m)
10/12
Location
3
Habitat
8:47
10/12 3 8:47 9:43 9:03 GB 332 290 42 1.30 C22 Riparian10/12 3 8:47 9:43 9:04 GB 313 290 23 1.09 C22 Riparian10/12 3 8:47 9:43 9:10 Gran 262 271 9 1.69 C24 Riparian10/12 3 8:47 9:43 9:12 GB 263 250 13 1.70 C24 Riparian10/12 3 8:47 9:43 9:12 Gran 299 248 51 1.32 C24-C25
Riparian
10/12 3 8:47 9:43 9:16 GB 85 48 37 1.32 C26 Riparian10/12 3 8:47 9:43 9:16 GB 62 48 14 1.56 C26 Riparian10/12 3 8:47 9:43 9:16 Gran 342 347 5 1.05 C27 Riparian10/12 3 8:47 9:43 9:16 Gran 356 347 9 1.73 C27 Riparian10/12 3 8:47 9:43 9:16 Gran 348 247 101 1.87 C27 Riparian10/12 3 8:47 9:43 9:16 Gran 344 347 3 2.18 C27 Riparian10/12 3 8:47 9:43 9:22 GB 325 347 22 0.71 C27 Riparian10/12 3 8:47 9:43 9:25 GB 45 6 39 1.53 C27 Riparian10/12 3 8:47 9:43 9:40 Gran 312 313 1 0.73 C29-C30 Riparian10/12 3 8:47 9:43 9:41 Gran 53 11 42 1.14 Tag 18 Riparian10/12 3 8:47 9:43 9:43 Gran 61 94 33 0.92 C51
9:43 8:55 Gran 342 349 7 1.38 Tag 20 Riparian10/12 6 9:43 9:46 9:45 GB 97 97 0 1.04 C71
Riparian
10/12 6 9:43 9:46 9:46
GB 85 181 96 1.05 Riparian10/12 1 9:46 9:5210/13 1 7:17 7:2510/13 6 7:25 7:2910/13 3 7:29 8:14 7:31 Gran 301 279 22 1.10 C52-C51
Riparian
10/13 3 7:29 8:14 7:46 Gran 158 196 38 1.16 C26 Riparian10/13 3 7:29 8:14 7:46 Gran 147 196 49 1.15 C26 Riparian10/13 3 7:29 8:14 7:46 GB 155 196 41 1.52 C26 Riparian10/13 3 7:29 8:14 7:26 GB 176 212 36 1.41 C26 Riparian
63

Date
Start Time
End Time
Observ. Time Species
Frog Bearing (˚)
Path Bearing (˚)
Sighting Angle (˚)
Modified Transect
Dist. (m)
Location
Habitat
10/13 3 7:29 8:14 7:46 GB 186 212 26 1.29 C26 Riparian10/13 3 7:29 8:14 7:48 Gran 186 210 24 0.87 C26 Riparian10/13 3 7:29 8:14 7:51 Gran 258 219 39 1.26 C26 Riparian10/13 3 7:29 8:14 7:51 Gran 263 219 44 1.48 C26 Riparian10/13 3 7:29 8:14 7:51 Gran 138 171 33 1.10 C26 Riparian10/13 3 7:29 8:14 7:57 Gran 59 75 16 0.89 C25-C24 Riparian10/13 3 7:29 8:14 7:57 Gran 306 75 129 0.38 C25-24 Riparian10/13 3 7:29 8:14 8:00 GB 64 69 5 1.14 C24 Riparian10/13 3 7:29 8:14 8:02 GB 121 100 21 0.86 C22 Riparian10/13 3 7:29 8:14 8:13
Gran 273 213 60 1.18 C2-C1 Riparian
10/13 4 8:15 8:3110/13 5 8:31 8:53 8:43 GB 351 313 38 1.31 BB3 Bamboo10/13 5 8:31 8:53 8:50
GB 341 21 40 0.61 BB24 Bamboo
10/13 4 9:14 9:2310/13 2 9:25 9:4310/13 1 9:43 9:4510/13 1 14:02 14:1210/13 2 14:12 14:32 14:14 GB 324 291 33 1.08 B96 Riparian10/13 4 14:32 14:47 14:42
Gran 251 279 28 1.04 B24 Riparian
10/13 5 14:48 15:1310/13 4 15:13 15:35 15:16 Gran 84 61 23 1.07 B24-26 Riparian10/13 4 15:13 15:35 15:17 Gran 80 62 18 1.23 B26 Riparian10/13 4 15:13 15:35 15:17 Gran 73 62 11 1.47 B26 Riparian10/13 4 15:13 15:35 15:17 Gran 65 62 3 1.78 B26 Riparian10/13 4 15:13 15:35 15:17 Gran 150 90 60 1.37 B27 Riparian10/13 4 15:13 15:35 15:17 Gran 150 90 60 1.37 B27 Riparian10/13 4 15:13 15:35 15:19 Gran 99 80 19 1.61 B27-B28 Secondary10/13 4 15:13 15:35 15:19 GB 135 85 50 1.60 B27-B28 Secondary
64
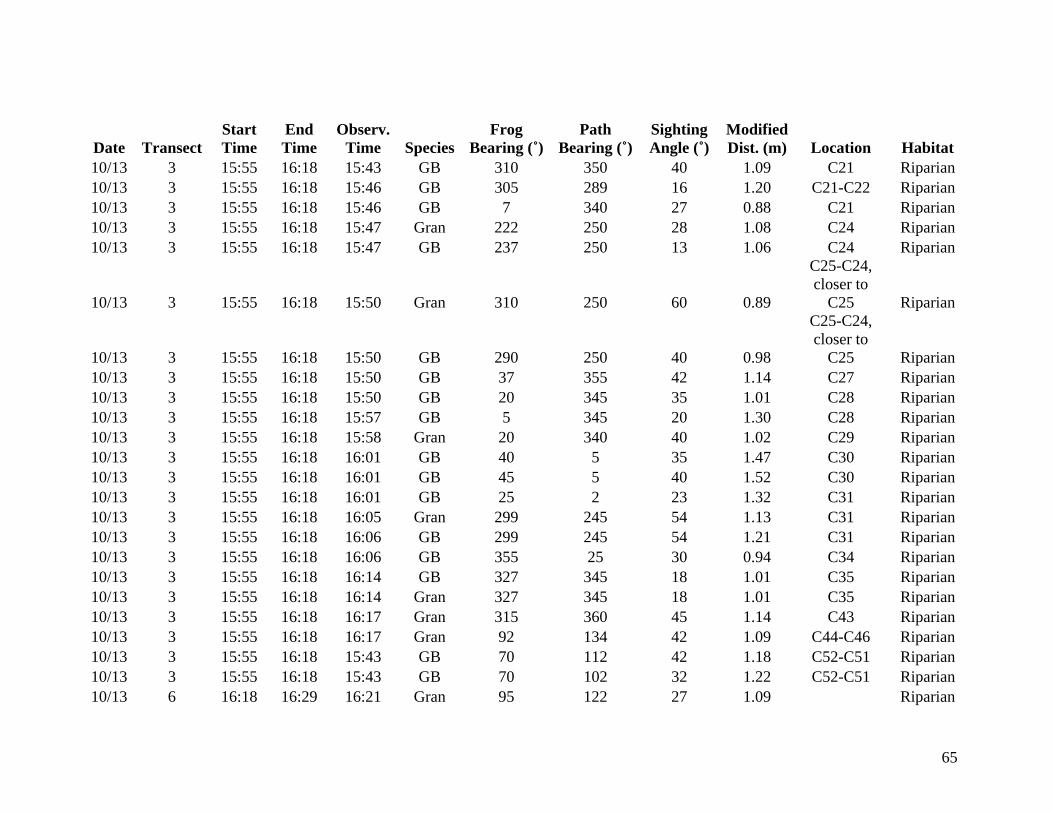
Date
Start Time
End Time
Observ. Time Species
Frog Bearing (˚)
Path Bearing (˚)
Sighting Angle (˚)
Modified Transect
Dist. (m)
Location
Habitat
10/13 3 15:55 16:18 15:43 GB 310 350 40 1.09 C21 Riparian10/13 3 15:55 16:18 15:46 GB 305 289 16 1.20 C21-C22
Riparian
10/13 3 15:55 16:18 15:46 GB 7 340 27 0.88 C21 Riparian10/13 3 15:55 16:18 15:47 Gran 222 250 28 1.08 C24 Riparian10/13 3 15:55 16:18 15:47 GB 237 250 13 1.06 C24 Riparian
10/13 3 15:55 16:18 15:50 Gran 310 250 60 0.89
C25-C24, closer to
C25 Riparian
10/13 3 15:55 16:18 15:50 GB 290 250 40 0.98
C25-C24, closer to
C25 Riparian10/13 3 15:55 16:18 15:50 GB 37 355 42 1.14 C27 Riparian10/13 3 15:55 16:18 15:50 GB 20 345 35 1.01 C28 Riparian10/13 3 15:55 16:18 15:57 GB 5 345 20 1.30 C28 Riparian10/13 3 15:55 16:18 15:58 Gran 20 340 40 1.02 C29 Riparian10/13 3 15:55 16:18 16:01 GB 40 5 35 1.47 C30 Riparian10/13 3 15:55 16:18 16:01 GB 45 5 40 1.52 C30 Riparian10/13 3 15:55 16:18 16:01 GB 25 2 23 1.32 C31 Riparian10/13 3 15:55 16:18 16:05 Gran 299 245 54 1.13 C31 Riparian10/13 3 15:55 16:18 16:06 GB 299 245 54 1.21 C31 Riparian10/13 3 15:55 16:18 16:06 GB 355 25 30 0.94 C34 Riparian10/13 3 15:55 16:18 16:14 GB 327 345 18 1.01 C35 Riparian10/13 3 15:55 16:18 16:14 Gran 327 345 18 1.01 C35 Riparian10/13 3 15:55 16:18 16:17 Gran 315 360 45 1.14 C43 Riparian10/13 3 15:55 16:18 16:17 Gran 92 134 42 1.09 C44-C46 Riparian10/13 3 15:55 16:18 15:43 GB 70 112 42 1.18 C52-C51 Riparian10/13 3 15:55 16:18 15:43 GB 70 102 32 1.22 C52-C51
Riparian
10/13 6 16:18 16:29 16:21 Gran 95 122 27 1.09 Riparian
65

Date
Start Time
End Time
Observ. Time Species
Frog Bearing (˚)
Path Bearing (˚)
Sighting Angle (˚)
Modified Transect
Dist. (m)
Location
Habitat
10/13 6 16:18 16:29 16:26 GB 125 154 29 1.13 Riparian10/13 6 16:18 16:29 16:26 GB 125 154 29 1.71 Riparian10/13 6 16:18 16:29 16:26
Gran 145 205 60 1.10 Riparian
10/13 1 16:29 16:3310/14 1 7:15 7:2010/14 6 7:20 7:29 7:25 Gran 320 268 52 1.14 Riparian10/14 6 7:20 7:29 7:26 GB 203 234 31 1.08 Riparian10/14 3 7:30 8:02 9:31 Gran 204 250 46 0.79 C56 Riparian10/14 3 7:30 8:02 7:44 GB 162 162 0 1.07 C28 Riparian10/14 3 7:30 8:02 7:46 GB 203 182 21 1.16 C27-28 Riparian10/14 3 7:30 8:02 7:46 GB 166 182 16 2.00 C27-28 Riparian10/14 3 7:30 8:02 7:47 Gran 180 158 22 1.22 C27 Riparian10/14 3 7:30 8:02 7:48 Gran 129 168 39 1.07 C27 Riparian10/14 3 7:30 8:02 7:48 GB 232 208 24 0.88 C26 Riparian10/14 3 7:30 8:02 7:49 GB 125 148 23 1.02 C26 Riparian10/14 3 7:30 8:02 7:50 Gran 53 71 18 0.88 C25 Riparian10/14 3 7:30 8:02 7:50 Gran 52 72 20 1.40 C25-24 Riparian10/14 3 7:30 8:02 7:55 Gran 182 117 65 1.18 C22 Riparian10/14 3 7:30 8:02 7:56 GB 94 109 15 1.20 C22 Riparian10/14 3 7:30 8:02 7:56
Gran 114 99 15 1.20 C22-21 Riparian
10/14 4 8:03 8:1610/14 5 8:16 8:4010/14 4 8:40 8:51 8:44
Gran 13 58 45 1.23 B24-26 Riparian
10/14 2 8:57 9:1510/14 1 9:15 9:1810/14 1 13:30 13:3510/14 6 13:35 13:43 13:40 Gran 310 284 26 0.59 bridge Riparian10/14 3 13:40 14:19 14:02 Gran 102 192 90 1.25 C30 Riparian
66
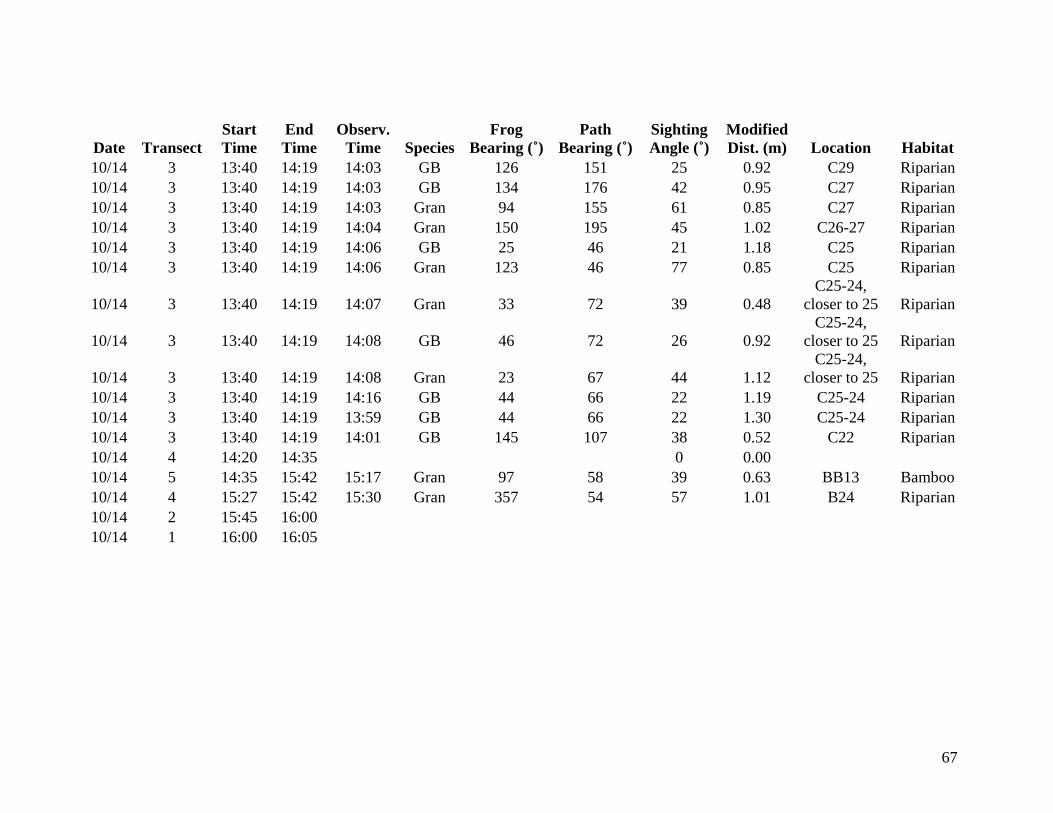
Date
Start Time
End Time
Observ. Time Species
Frog Bearing (˚)
Path Bearing (˚)
Sighting Angle (˚)
Modified Transect
Dist. (m)
Location
Habitat
10/14 3 13:40 14:19 14:03 GB 126 151 25 0.92 C29 Riparian10/14 3 13:40 14:19 14:03 GB 134 176 42 0.95 C27 Riparian10/14 3 13:40 14:19 14:03 Gran 94 155 61 0.85 C27 Riparian10/14 3 13:40 14:19 14:04 Gran 150 195 45 1.02 C26-27 Riparian10/14 3 13:40 14:19 14:06 GB 25 46 21 1.18 C25 Riparian10/14 3 13:40 14:19 14:06 Gran 123 46 77 0.85 C25 Riparian
10/14 3 13:40 14:19 14:07 Gran 33 72 39 0.48C25-24,
closer to 25 Riparian
10/14 3 13:40 14:19 14:08 GB 46 72 26 0.92C25-24,
closer to 25 Riparian
10/14 3 13:40 14:19 14:08 Gran 23 67 44 1.12C25-24,
closer to 25 Riparian 10/14 3 13:40 14:19 14:16 GB 44 66 22 1.19 C25-24 Riparian10/14 3 13:40 14:19 13:59 GB 44 66 22 1.30 C25-24 Riparian10/14 3 13:40 14:19 14:01
GB 145 107 38 0.52 C22
Riparian
10/14 4 14:20 14:35 0 0.0010/14 5 14:35 15:42 15:17 Gran 97 58 39 0.63 BB13 Bamboo10/14 4 15:27 15:42 15:30
Gran 357 54 57 1.01 B24 Riparian
10/14 2 15:45 16:0010/14 1 16:00 16:05
67
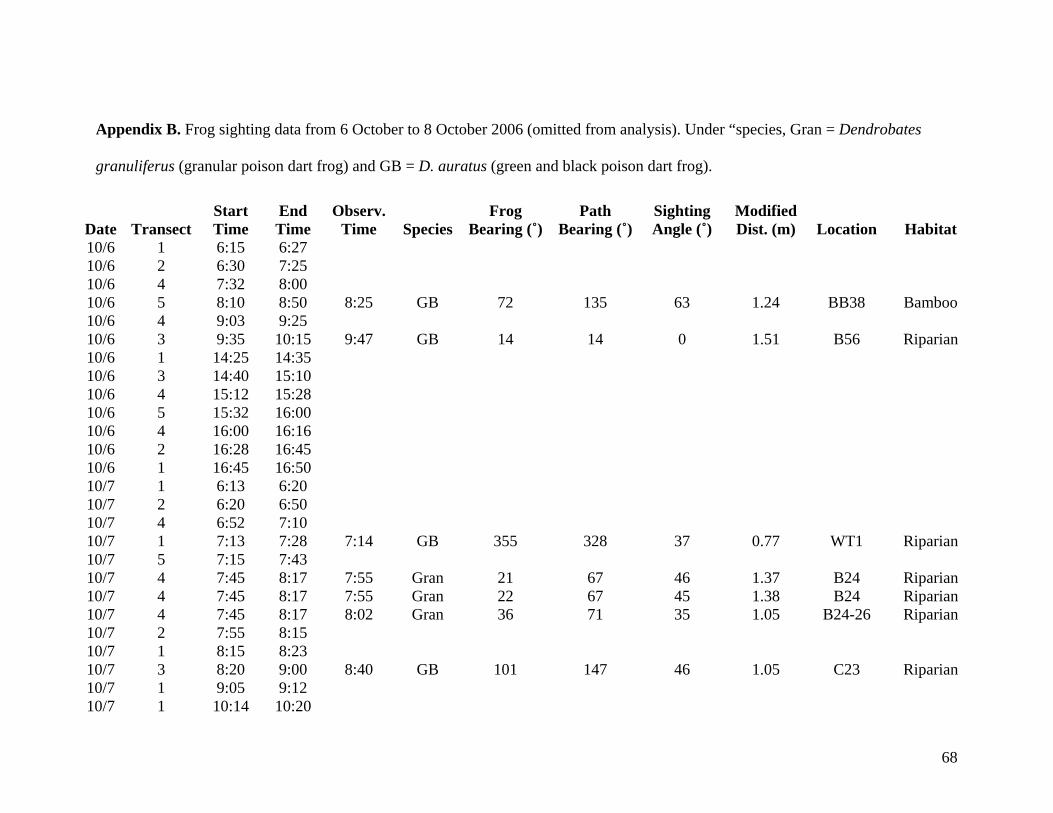
Appendix B. Frog sighting data from 6 October to 8 October 2006 (omitted from analysis). Under “species, Gran = Dendrobates
granuliferus (granular poison dart frog) and GB = D. auratus (green and black poison dart frog).
Date
Transect
Start Time
End Time
Observ. Time
Species
Bearing (˚)
Frog Path Bearing (˚)
Sighting Angle (˚)
Modified Dist. (m)
Location
Habitat
10/6 1 6:15 6:2710/6 2 6:30 7:2510/6 4 7:32 8:0010/6 5 8:10 8:50 8:25
GB 72 135 63 1.24
BB38
Bamboo
10/6 4 9:03 9:2510/6 3 9:35 10:15 9:47
GB 14 14 0 1.51
B56 Riparian
10/6 1 14:25 14:3510/6 3 14:40 15:1010/6 4 15:12 15:2810/6 5 15:32 16:0010/6 4 16:00 16:1610/6 2 16:28 16:4510/6 1 16:45 16:5010/7 1 6:13 6:2010/7 2 6:20 6:5010/7 4 6:52 7:1010/7 1 7:13 7:28 7:14
GB 355 328 37 0.77
WT1
Riparian
10/7 5 7:15 7:4310/7 4 7:45 8:17 7:55 Gran 21 67 46 1.37 B24 Riparian10/7 4 7:45 8:17 7:55 Gran 22 67 45 1.38 B24 Riparian10/7 4 7:45 8:17 8:02
Gran
36 71 35 1.05
B24-26
Riparian
10/7 2 7:55 8:1510/7 1 8:15 8:2310/7 3 8:20 9:00 8:40
GB 101 147 46 1.05
C23 Riparian
10/7 1 9:05 9:1210/7 1 10:14 10:20
68
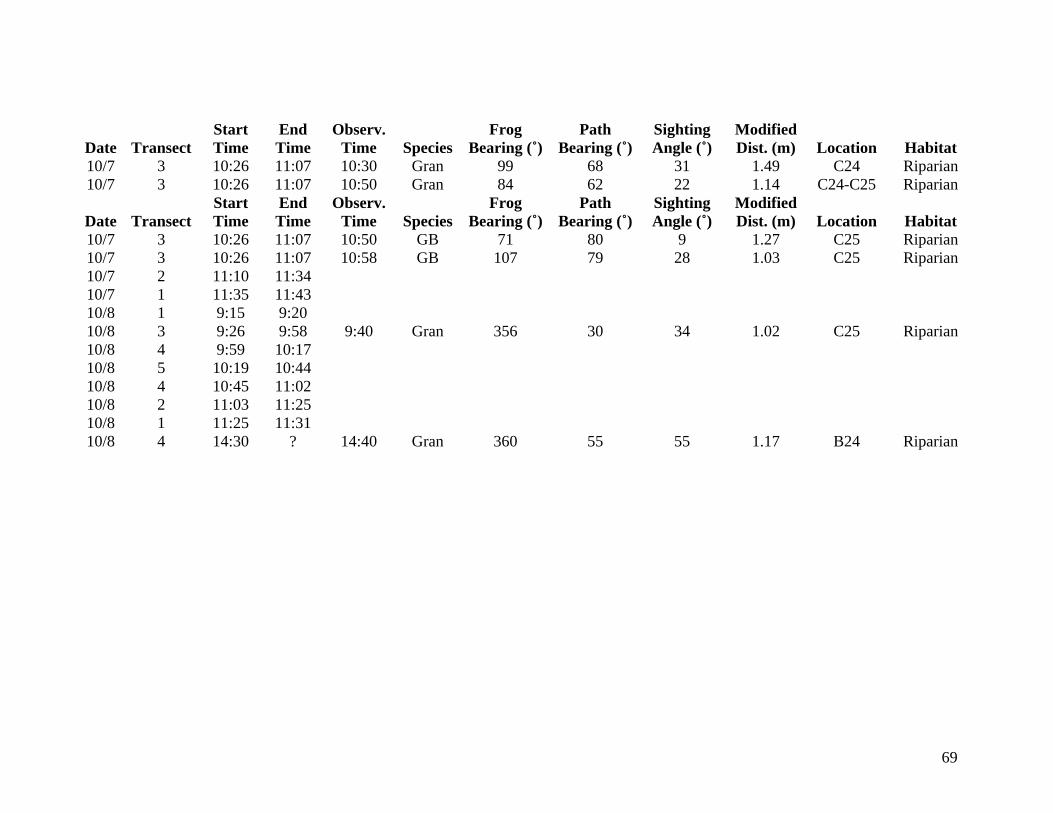
Date
Transect
Start Time
End Time
Observ. Time Species Bearing (˚)
Frog Path Bearing (˚)
Sighting Angle (˚)
Modified Dist. (m)
Location
Habitat
10/7 3 10:26 11:07 10:30 Gran 99 68 31 1.49 C24 Riparian10/7 3 10:26 11:07 10:50 Gran 84 62 22 1.14 C24-C25 Riparian
Date Transect
Start Time
End Observ. Time Species
Bearing (˚)
Frog Path Bearing (˚)
Sighting Angle (˚)
Modified Dist. (m)
Location
Time Habitat
10/7 3 10:26 11:07 10:50 GB 71 80 9 1.27 C25 Riparian10/7 3 10:26 11:07 10:58
GB 107 79 28 1.03
C25 Riparian
10/7 2 11:10 11:3410/7 1 11:35 11:4310/8 1 9:15 9:2010/8 3 9:26 9:58 9:40
Gran
356 30 34 1.02
C25 Riparian
10/8 4 9:59 10:1710/8 5 10:19 10:4410/8 4 10:45 11:0210/8 2 11:03 11:2510/8 1 11:25 11:31
10/8 4 14:30 ? 14:40 Gran 360 55 55 1.17 B24 Riparian
69
Related Documents











