Pomolic acid triggers mitochondria-dependent apoptotic cell death in leukemia cell line Janaina Fernandes a,1 , Ricardo Weinlich b,1 , Rachel Oliveira Castilho c , Maria Auxiliadora Coelho Kaplan c , Gustavo Pessini Amarante-Mendes b , Cerli Rocha Gattass a, * a Instituto de Biofı ´sica Carlos Chagas Filho, UFRJ, Ilha do Fundao, IBCCF, CCS Bl.G, Rio de Janeiro 21949-900, Brazil b Departamento de Imunologia, Instituto de Cie ˆncias Biome ´dicas, USP, Sa ˜o Paulo, e Instituto de Investigac ¸a ˜o em Imunologia, Instituto do Mile ˆnio, Brazil c Nu ´cleo de Pesquisas em Produtos Naturais, UFRJ, Rio de Janeiro, Brazil Received 2 July 2004; received in revised form 28 August 2004; accepted 1 September 2004 Abstract One of the major goals in chemotherapy is to circumvent anti-apoptotic strategies developed by tumor cells. In a previous paper, we showed that pomolic acid (PA) is able to kill the leukemia cell line K562 and its MDR derivative, Lucena 1. Here, we demonstrated that PA-induced apoptosis of HL-60 cells is dependent on the activation of caspases-3 and -9 and dissipation of the mitochondrial transmembrane potential (Dj m ). Disruption of Dj m precedes caspase activation and is not inhibited by zVAD-fmk indicating mitochondria as the main target of PA. Our data pointed to the potential use of PA to overcome apoptosis resistance. q 2004 Elsevier Ireland Ltd. All rights reserved. Keywords: Pomolic acid; Apoptosis; Mitochondria membrane potential; Cancer therapy; Leukemia; MDR 1. Introduction One of the major causes of chemotherapeutic failures in cancer treatment is the development of different kinds of resistance. The search for new drugs able to overcome the resistance mechanisms and leading to tumor cell death is of utmost importance for cancer therapy. In a previous study [1] we showed that pomolic acid (PA), a pentacyclic triterpene isolated from Chrysobalanus icaco, was highly effective in inhibiting the growth of both 0304-3835/$ - see front matter q 2004 Elsevier Ireland Ltd. All rights reserved. doi:10.1016/j.canlet.2004.09.001 Cancer Letters 219 (2005) 49–55 www.elsevier.com/locate/canlet Abbreviations: Dj m , mitochondrial transmembrane potential; ANT, adenine nucleotide translocator; Apaf-1, apoptotic protease activating factor-1; CD95-L, CD95-ligand; CsA, cyclosporin; DiOC 6 (3), 3,3 0 -dihexyloxacarbocyanide iodide; MTT, 3-(4,5- dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide; PI, propi- dium iodide; PTP, permeability transition pore; zVAD-fmk, benzyloxycarbonyl-Val-Ala-Asp(OMe)-fluoromethylketone. * Corresponding author. Tel.: C55 21 2562 6564; fax: C55 21 2280 8193. E-mail address: [email protected] (C.R. Gattass). 1 Janaina Fernandes and Ricardo Weinlich contributed equally to this paper.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Pomolic acid triggers mitochondria-dependent apoptotic
cell death in leukemia cell line
Janaina Fernandesa,1, Ricardo Weinlichb,1, Rachel Oliveira Castilhoc,Maria Auxiliadora Coelho Kaplanc, Gustavo Pessini Amarante-Mendesb,
Cerli Rocha Gattassa,*
aInstituto de Biofısica Carlos Chagas Filho, UFRJ, Ilha do Fundao, IBCCF, CCS Bl.G, Rio de Janeiro 21949-900, BrazilbDepartamento de Imunologia, Instituto de Ciencias Biomedicas, USP, Sao Paulo, e Instituto de Investigacao em Imunologia,
Instituto do Milenio, BrazilcNucleo de Pesquisas em Produtos Naturais, UFRJ, Rio de Janeiro, Brazil
Received 2 July 2004; received in revised form 28 August 2004; accepted 1 September 2004
Abstract
One of the major goals in chemotherapy is to circumvent anti-apoptotic strategies developed by tumor cells. In a previous
paper, we showed that pomolic acid (PA) is able to kill the leukemia cell line K562 and its MDR derivative, Lucena 1. Here, we
demonstrated that PA-induced apoptosis of HL-60 cells is dependent on the activation of caspases-3 and -9 and dissipation of
the mitochondrial transmembrane potential (Djm). Disruption of Djm precedes caspase activation and is not inhibited by
zVAD-fmk indicating mitochondria as the main target of PA. Our data pointed to the potential use of PA to overcome apoptosis
resistance.
q 2004 Elsevier Ireland Ltd. All rights reserved.
Keywords: Pomolic acid; Apoptosis; Mitochondria membrane potential; Cancer therapy; Leukemia; MDR
0304-3835/$ - see front matter q 2004 Elsevier Ireland Ltd. All rights re
doi:10.1016/j.canlet.2004.09.001
Abbreviations: Djm, mitochondrial transmembrane potential;
ANT, adenine nucleotide translocator; Apaf-1, apoptotic protease
activating factor-1; CD95-L, CD95-ligand; CsA, cyclosporin;
DiOC6(3), 3,3 0-dihexyloxacarbocyanide iodide; MTT, 3-(4,5-
dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide; PI, propi-
dium iodide; PTP, permeability transition pore; zVAD-fmk,
benzyloxycarbonyl-Val-Ala-Asp(OMe)-fluoromethylketone.
* Corresponding author. Tel.: C55 21 2562 6564; fax: C55 21
2280 8193.
E-mail address: [email protected] (C.R. Gattass).1 Janaina Fernandes and Ricardo Weinlich contributed equally to
this paper.
1. Introduction
One of the major causes of chemotherapeutic
failures in cancer treatment is the development of
different kinds of resistance. The search for new
drugs able to overcome the resistance mechanisms
and leading to tumor cell death is of utmost
importance for cancer therapy. In a previous study
[1] we showed that pomolic acid (PA), a pentacyclic
triterpene isolated from Chrysobalanus icaco, was
highly effective in inhibiting the growth of both
Cancer Letters 219 (2005) 49–55
www.elsevier.com/locate/canlet
served.

J. Fernandes et al. / Cancer Letters 219 (2005) 49–5550
K562, a leukemia cell line and Lucena-1, a MDR
resistant leukemia line that overexpresses P-glyco-
protein (Pgp 170) [2]. However, apart from its
capacity to induce apoptosis [1], the mechanisms of
PA activity are presently unclear. As part of the
evaluation of the potential of new anti-cancer
compounds, this paper investigated the molecular
pathways of PA-induced apoptosis.
Several compounds used in anticancer chemother-
apy are believed to induce cell death via activation of
key elements of the apoptotic program. Apoptosis is
characterized by distinct morphological features such
as plasma membrane blebbing, cell shrinkage, chro-
matin condensation, DNA fragmentation, and break-
down of the cell into apoptotic bodies [3–5]. Recently,
it has been shown that mitochondria play an essential
role in the so-called intrinsic pathway of apoptosis [6],
the major route used by several chemotherapeutic
drugs [7,8]. Activation of the mitochondrial per-
meability transition pore (PTP) leads to dissipation of
mitochondria membrane potential (Djm) [9–11]. In
the cytosol, the complex formed by cytochrome c,
Apaf-1 and pro-caspase-9 (apoptosome) triggers the
activation of caspase-9 [12]. This caspase promotes
the activation of a cascade of caspases including
caspase-3, with the consequent cleavage of specific
caspase substrates and cell death [7,10]. Because
caspase activities can be detected in all cells under-
going apoptosis, regardless of their origin or death
stimuli, and inhibitors of caspases prevent the hall-
marks of apoptosis, these proteases are considered the
main executioners of apoptosis [13,14].
Due to the relevance of the potential anti MDR-
activity of PA and considering that characterization of
their mechanisms of action is a preliminary and
fundamental step to future clinical application/use of
new drugs, the aim of this paper is to clarify the
mechanism of action of this triterpene. Investigation
of PA-induced death pathways demonstrated that
induction of apoptosis by this triterpene involves
activation of caspases-9 and -3 in a process mediated
through loss of Djm, an unusual mechanism for drug-
induced apoptosis in HL-60 cells. Indeed, treatment of
cells with CSA an inhibitor of mitochondria PTP,
completely abrogates apoptosis suggesting that mito-
chondria is the main target for PA. Since it has been
proposed that drugs that act on mitochondria
may bypass MDR resistance mechanisms, our data
reinforce the potential of pomolic acid as a new anti-
MDR drug.
2. Materials and methods
2.1. Chemicals and cell culture
Pomolic acid isolated from Chrysobalanus
icacoL. as described previously [1] was dissolved
in dimethyl sulfoxide (DMSO, SIGMA, St Louis)
and diluted in RPMI 1640 for use. DiOC6(3) was
from Molecular Probes, Inc. (Eugene, OR), cyclos-
porin A was from Calbiochem (San Diego, CA,
USA), caspase inhibitor zVAD-fmk was from
Enzyme Systems (Livermore, USA), mAbs for
caspases-3 and -9 were from Pharmingen (San
Diego, CA, USA), anti-actin mAb was from ICN
(Costa Mesa, USA) and anti-mouse IgG-HRP from
Amersham Biosciences (Arlington, IL). The human
leukemia cell line HL-60 was cultured at 37 8C and
5% CO2 in RPMI 1640 (Life Technologies, Inc.,
USA) supplemented with heat inactivated 10% fetal
calf serum (FCS; Life Technologies, Inc., USA),
2 mM L-glutamine, 100 U/ml of penicillin and
100 mg/ml of streptomycin (Life Technologies,
Inc., USA).
2.2. Cell viability assay
Cell viability was assessed by MTT [15]. Cells
(104/well) were plated in 96 well tissue culture plates
for 24 h and then treated with different concentrations
of PA (1, 10, 25, 100 mg/ml) or DMSO (in the same
concentrations carried by the drug). After another
48 h, 20 ml (5 mg/ml) MTT was added to each well
and the plate was incubated at 37 8C in the dark for at
least 4 h. The formazan crystals were solubilized in
DMSO (200 ml/well) and the reduction of MTT was
quantified by absorbance (A570 nm, with 630 nm-
reference filter). Effects of the drug on cell viability
were calculated using cells treated with DMSO as
control.
2.3. Detection of apoptosis
Apoptosis was evaluated by cell morphology and
cell cycle analysis [16]. After 24 h resting, plated cells

J. Fernandes et al. / Cancer Letters 219 (2005) 49–55 51
(2!105/well) were treated with medium or different
concentrations (1, 10, 25, 100 mg/ml) of PA and
incubated for another 24 h. After this time, cells were
harvested and alternatively pelleted onto glass slides
for Giemsa staining or suspended in 300 ml of HFS-
Hypotonic Fluorescent Solution (50 mg/ml propidium
iodide (PI) and 0.1% Triton X-100 in 0.1% Na citrate
buffer). After 1 h incubation in the dark at 4 8C the
DNA content was measured by flow cytometry (FL-2)
(FACSCalibur, Becton Dickinson, San Jose, CA).
Data acquisition and analysis were controlled by
Cellquest software version 3.1f. Subdiploid popu-
lations were considered apoptotic. In some exper-
iments, cells were pre-incubated during 1 h with
zVAD-fmk (100 mM) or CSA (5 mM) before the
addition of PA to the cultures.
2.4. Measurement of mitochondrial transmembrane
potential (Djm)
Variations of mitochondrial membrane potential
(MMP) was assessed in cells plated and treated with
PA under the same conditions described for apoptosis
detection. Then cells were stained with the fluor-
ochrome DiOC6(3) (40 nM) as described previously
[17] and analyzed by flow cytometry (FL-1), accord-
ing to Ormerod [18].
2.5. Western blot analysis
SDS-PAGE and western-blot analysis were per-
formed as described elsewhere [19]. Briefly, plated
cells (2!105/well), treated with 25 mg/ml PA for 4, 8,
and 12 h, were collected by centrifugation, washed
once in ice-cold PBS, lysed directly in SDS-sample
buffer (50 mM Tris–HCl, pH 6.8, 2% SDS, 10%
glycerol, 2.5% 2-ME) and boiled for 5 min. Equal
amounts of protein were separated by SDS-PAGE and
transferred to polyvinylidene difluoride membranes at
150 mA for 5 h. Blots were blocked for 2 h in TBST
(10 mM Tris–HCl, pH 7.4, 150 mM NaCl, 0.05%
Tween) containing 5% nonfat dried milk, probed with
anti inactive caspases-3 or -9 for 1 h at RT and then
with anti-mouse IgG-HRP. Blots were developed
using the enhanced chemiluminescence system (ECL,
Amersham, Arlington, IL).
2.6. Statistical analysis
All of the experiments were performed in triplicate
and repeated at least three times. Results are
expressed as meanGSE. Statistical comparisons
were made by Turkey test. P%0.05 was considered
significant.
3. Results
3.1. Pomolic acid induces apoptosis of HL-60
In a previous paper, we showed that pomolic acid
inhibited the proliferation and induced apoptosis of
leukemia cell lines [1]. In this paper, HL-60 cells were
used to investigate the intracellular pathways involved
in PA cytotoxic activity.
Use of the MTT assay revealed that PA inhibited
the viability of HL-60 cells in a dose-dependent
manner, reaching a plateau at 10 mg/ml after 48 h of
treatment (Fig. 1A). To further evaluate the nature of
this inhibition, we followed the kinetics of PA action
on cell morphology. Cells incubated with 25 mg/ml
PA and analyzed 4 and 8 h later by optical
microscopy, showed characteristic apoptotic mor-
phology with cell shrinkage, DNA fragmentation and
the presence of apoptotic bodies (results not shown).
Cell-cycle analysis was used to quantify this effect.
Appearance of the sub-diploid nuclei population,
indicative of DNA fragmentation, demonstrated
that PA-induced DNA fragmentation was dose- and
time-dependent (Fig. 1B). Although maximal induc-
tion of apoptosis was observed after 48 treatment with
25 mg/ml PA, at this concentration of triterpene, a
significant DNA fragmentation was already seen
around 12 h of treatment (Fig. 1C).
3.2. Pomolic acid induces caspase activation
To investigate the biochemical features that lead to
apoptosis induced by PA, we examined the effect of
this triterpene on activation of caspases-3 and -9. HL-
60 cells were treated with pomolic acid (25 mg/ml)
and samples collected at 4, 8 and 12 h were blotted
with mAbs that recognize the inactive form of the
caspases. Compatible with the DNA fragmentation
shown in Fig. 1c insert, after 8 h a reduction of
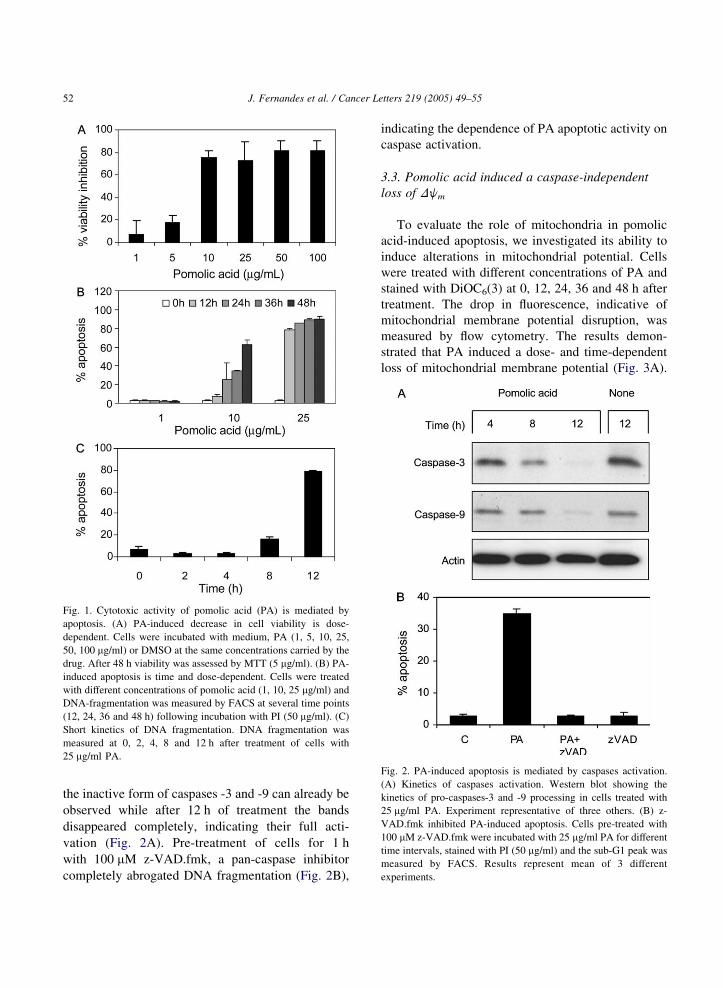
Fig. 1. Cytotoxic activity of pomolic acid (PA) is mediated by
apoptosis. (A) PA-induced decrease in cell viability is dose-
dependent. Cells were incubated with medium, PA (1, 5, 10, 25,
50, 100 mg/ml) or DMSO at the same concentrations carried by the
drug. After 48 h viability was assessed by MTT (5 mg/ml). (B) PA-
induced apoptosis is time and dose-dependent. Cells were treated
with different concentrations of pomolic acid (1, 10, 25 mg/ml) and
DNA-fragmentation was measured by FACS at several time points
(12, 24, 36 and 48 h) following incubation with PI (50 mg/ml). (C)
Short kinetics of DNA fragmentation. DNA fragmentation was
measured at 0, 2, 4, 8 and 12 h after treatment of cells with
25 mg/ml PA.
Fig. 2. PA-induced apoptosis is mediated by caspases activation.
(A) Kinetics of caspases activation. Western blot showing the
kinetics of pro-caspases-3 and -9 processing in cells treated with
25 mg/ml PA. Experiment representative of three others. (B) z-
VAD.fmk inhibited PA-induced apoptosis. Cells pre-treated with
100 mM z-VAD.fmk were incubated with 25 mg/ml PA for different
time intervals, stained with PI (50 mg/ml) and the sub-G1 peak was
measured by FACS. Results represent mean of 3 different
experiments.
J. Fernandes et al. / Cancer Letters 219 (2005) 49–5552
the inactive form of caspases -3 and -9 can already be
observed while after 12 h of treatment the bands
disappeared completely, indicating their full acti-
vation (Fig. 2A). Pre-treatment of cells for 1 h
with 100 mM z-VAD.fmk, a pan-caspase inhibitor
completely abrogated DNA fragmentation (Fig. 2B),
indicating the dependence of PA apoptotic activity on
caspase activation.
3.3. Pomolic acid induced a caspase-independent
loss of Djm
To evaluate the role of mitochondria in pomolic
acid-induced apoptosis, we investigated its ability to
induce alterations in mitochondrial potential. Cells
were treated with different concentrations of PA and
stained with DiOC6(3) at 0, 12, 24, 36 and 48 h after
treatment. The drop in fluorescence, indicative of
mitochondrial membrane potential disruption, was
measured by flow cytometry. The results demon-
strated that PA induced a dose- and time-dependent
loss of mitochondrial membrane potential (Fig. 3A).
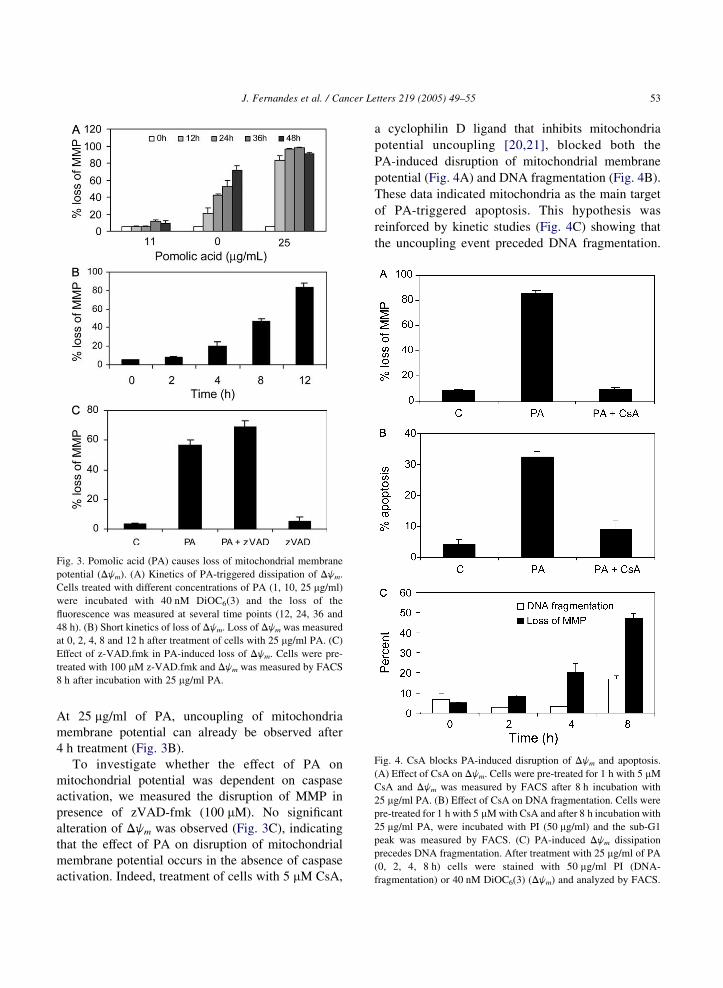
Fig. 3. Pomolic acid (PA) causes loss of mitochondrial membrane
potential (Djm). (A) Kinetics of PA-triggered dissipation of Djm.
Cells treated with different concentrations of PA (1, 10, 25 mg/ml)
were incubated with 40 nM DiOC6(3) and the loss of the
fluorescence was measured at several time points (12, 24, 36 and
48 h). (B) Short kinetics of loss of Djm. Loss of Djm was measured
at 0, 2, 4, 8 and 12 h after treatment of cells with 25 mg/ml PA. (C)
Effect of z-VAD.fmk in PA-induced loss of Djm. Cells were pre-
treated with 100 mM z-VAD.fmk and Djm was measured by FACS
8 h after incubation with 25 mg/ml PA.
Fig. 4. CsA blocks PA-induced disruption of Djm and apoptosis.
(A) Effect of CsA on Djm. Cells were pre-treated for 1 h with 5 mM
CsA and Djm was measured by FACS after 8 h incubation with
25 mg/ml PA. (B) Effect of CsA on DNA fragmentation. Cells were
pre-treated for 1 h with 5 mM with CsA and after 8 h incubation with
25 mg/ml PA, were incubated with PI (50 mg/ml) and the sub-G1
peak was measured by FACS. (C) PA-induced Djm dissipation
precedes DNA fragmentation. After treatment with 25 mg/ml of PA
(0, 2, 4, 8 h) cells were stained with 50 mg/ml PI (DNA-
fragmentation) or 40 nM DiOC6(3) (Djm) and analyzed by FACS.
J. Fernandes et al. / Cancer Letters 219 (2005) 49–55 53
At 25 mg/ml of PA, uncoupling of mitochondria
membrane potential can already be observed after
4 h treatment (Fig. 3B).
To investigate whether the effect of PA on
mitochondrial potential was dependent on caspase
activation, we measured the disruption of MMP in
presence of zVAD-fmk (100 mM). No significant
alteration of Djm was observed (Fig. 3C), indicating
that the effect of PA on disruption of mitochondrial
membrane potential occurs in the absence of caspase
activation. Indeed, treatment of cells with 5 mM CsA,
a cyclophilin D ligand that inhibits mitochondria
potential uncoupling [20,21], blocked both the
PA-induced disruption of mitochondrial membrane
potential (Fig. 4A) and DNA fragmentation (Fig. 4B).
These data indicated mitochondria as the main target
of PA-triggered apoptosis. This hypothesis was
reinforced by kinetic studies (Fig. 4C) showing that
the uncoupling event preceded DNA fragmentation.

J. Fernandes et al. / Cancer Letters 219 (2005) 49–5554
Indeed, 4 h after pomolic treatment when no DNA
fragmentation was observed, the loss of Djm is
already 20% (Fig. 4C).
4. Discussion
Pomolic acid has recently been shown to inhibit the
growth and to induce apoptosis of leukemia cell lines,
including one that overexpresses Pgp, a multidrug
resistance protein [1]. Using HL-60 cells to character-
ize the molecular pathways involved in this effect, this
paper showed that apoptosis induced by PA is triggered
by alteration of mitochondria membrane potential.
In recent years, several studies performed in vitro
have shown that the cytotoxic effects of several types
of drugs on human tumor cell lines are mediated by
apoptosis [7,22]. In agreement with these obser-
vations, DNA-fragmentation analysis presented here
showed that PA-induced reduction of HL-60 cell
viability is also due to apoptosis.
Caspases represent the core to which most of the
death signals converge to induce apoptosis. Thus,
independent of which signal initiated the events that
lead to apoptosis, caspases play a fundamental role in
the apoptotic process [14,23]. Induction of apoptosis
by PA is also associated with activation of caspases,
as both initiator (caspase-9) and effector (caspase-3)
caspases are activated during this process. The
dependence of caspases activation on PA-triggered
apoptosis was evidenced by the complete abrogation
of apoptosis in cells treated with the broad-range
peptide inhibitor of caspases, zVAD-fmk.
Although apoptosis can be activated by different
signaling pathways, opening of the permeability
transition pore (PTP) and a subsequent drop in
mitochondrial membrane potential have been
suggested as the main mechanisms involved in
cytochrome c release and apoptosome formation
during apoptosis induced by a series of chemo-
therapeutic agents [7,8]. Assembly of the apoptosome,
a complex formed by cytochrome c, Apaf-1, procas-
pase-9 and dATP, triggers activation of caspases-9, -3
and others, leading to cleavage of specific substrates
and cell death [24]. Our data showed that loss of Djm
is an early event in PA-induced apoptosis. In fact,
disruption of mitochondrial membrane potential
started 4 h after drug exposition while DNA-
fragmentation was only observed 8 h after PA
treatment, indicating the involvement of the mito-
chondrial pathway on PA-induced death. A depen-
dency of the loss of mitochondrial potential on
caspase activity has also been reported [25]. However,
our data, showing that blockage of caspases activation
by zVAD-fmk, which completely abrogated PA-
triggered apoptosis, did not impair the Djm disrup-
tion, indicated that PA-induced loss of mitochondrial
potential is independent of caspase activation.
Investigation on the mechanism of PTP opening has
helped to elucidate its role on apoptosis. It has been
proposed that the basic structure of PTP is composed by
a voltage-dependent anion channel (VDAC), a matrix
isomerase cyclophilin-D (CyP-D) and an adenine
nucleotide translocator (ANT) [21,26]. Herein we
showed that inhibition of mitochondrial pore opening
by the ANT inhibitor, CsA, a ligand of cyclophilin-D,
prevented both mitochondrial membrane potential loss
and DNA fragmentation induced by PA. Thus, in HL-60
treated with PA, disruption of Djm occurs upstream of
apoptosome formation and effector caspases activation,
as already shown by studies involving other cytotoxic
drugs in different cell lines [27,28].
Due to the modulatory role of pre-mitochondrial
transduction signals on PTP opening and mitochon-
dria depolarization, substances whose activity is
mediated through mitochondria have been proposed
as important tools to overcome resistance mediated by
pre-mitochondrial signals [11]. Data presented in this
paper showing that (1) blockage of caspase activation
abrogates DNA-fragmentation but had no effect on
PA-induced loss of Djm and (2) treatment with CSA,
which prevented loss of MMP also blocked PA-
induced DNA-fragmentation, pointed to mitochondria
as the main target of PA. Thus, our data emphasizes
PA as a promising experimental cytotoxic agent to
develop new strategies directed to overcome apopto-
sis resistance due to defects in the intrinsic death
receptor-signaling pathway.
Acknowledgements
J.F is a recipient of a PhD fellowship from CNPq
and R.W. was a recipient of a MSc fellowship from
FAPESP. This research was supported by grants from
CNPq, FAPERJ, PRONEX (MCT), and FAPESP.

J. Fernandes et al. / Cancer Letters 219 (2005) 49–55 55
References
[1] J. Fernandes, R.O. Castilho, M.R. Costa, K. Wagner-Souza,
M.A.C. Kaplan, C.R. Gattass, Pentacyclic triterpenes from
Chrysobalanaceae species: cytotoxicity on multidrug resistant
and sensitive leukemia cell lines, Cancer Lett. 190 (2003)
165–169.
[2] V.M. Rumjanek, G.S. Trindade, K. Wagner-Souza,
M.C. Meletti-de-Oliveira, L.F. Marques-Santos, R.C. Maia,
et al., Multidrug-resistance in tumor cells: characterization of
the multidrug resistant cell line K562-Lucena 1, Ann. Acad.
Bras. Ci. 73 (2001) 57–69.
[3] J.F. Kerr, A.H. Wyllie, A.R. Currie, Apoptosis: a basic
biological phenomenon with wide-ranging implications in
tissue kinetics, Br. J. Cancer 26 (1972) 239–257.
[4] W.C. Earnshaw, Nuclear changes in apoptosis, Curr. Opin.
Cell Biol. 7 (1995) 337–343.
[5] M. Zornig, A-O. Hueber, W. Baum, G. Evan, Apoptosis
regulators and their role in tumorigenesis, Biochim. Biophys.
Acta 1551 (2001) F1–F37.
[6] X. Wang, The expanding role of mitochondria in apoptosis,
Gene Dev. 15 (2001) 2922–2933.
[7] R. Kim, K. Tanabe, Y. Uchida, M. Emim, U. Inoue, T. Toge,
Current status of the molecular mechanisms of anticancer
drug-induced apoptosis. The contribution of molecular-level
analysis to cancer chemotherapy, Cancer Chemother. Phar-
macol. 50 (2002) 343–352.
[8] P. Constantini, E. Jacotot, D. Decaudin, G. Kroemer, Mito-
chondrion as a novel target of anticancer drugs, J. Natl. Cancer
Inst. 5 (2000) 1042–1053.
[9] J. Cai, J. Yang, T.P. Joens, Mitochondrial control of apoptosis:
the role of cytochrome c, Biochim. Biophys. Acta 1366 (1998)
139–149.
[10] G.P. Amarante-Mendes, D.R. Green, The regulation of
apoptotic cell death, Braz. J. Med. Biol. Res. 32 (1999)
1053–1061.
[11] K-M. Debatin, D. Poncet, G. Kroemer, Chemotherapy,
targeting the mitochondrial cell death pathway, Oncogene 21
(2002) 8786–8803.
[12] H. Zou, Y. Li, X. Liu, X. Wang, An APAF-1.cytochrome c
multimeric complex is a functional apoptosome that activates
pro-caspase-9, J. Biol. Chem. 274 (1999) 11549–11556.
[13] E.A. Slee, M.T. Herte, B.B. Wolf, C.A. Casiano,
D.D. Newmyer, H.G. Wang, et al., Ordering the cytochrome
c-initiated caspase cascade: hierarchical activation of caspase-
2, -3, -6, -7, -8 and -10 in a caspase-9-dependent manner,
J. Cell. Biol. 144 (1999) 281–292.
[14] E.M. Creagh, J.S. Martin, Caspases: cellular demolition
experts, Biochem. Soc. Trans. 29 (2001) 696–702.
[15] T. Mosmann, Rapid colorimetric assay for cellular growth and
survival: application to proliferation and cytotoxicity assays,
J. Immunol. Methods 65 (1983) 55–63.
[16] I. Nicoletti, G. Megliorati, M.C. Pagliacci, F. Grignani,
C. Riccardi, A rapid and simple method for measuring
thymocytes apoptosis by propidium iodide staining and flow
cytometry, J. Immunol. Methods 139 (1991) 271–279.
[17] G.P. Amarante-Mendes, D.M. Finucane, S.J. Martin,
T.G. Cotter, G.S. Salvesen, D.R. Green, Anti-apoptotic
oncogenes prevent caspase-dependent and independent cell
death, Cell Death Differ. 5 (1998) 298–306.
[18] M.C. Ormerod, Flow cytometry in the study of apoptosis in:
M.C. Ormerod (Ed.),, Flow Cytometry-A Practical Approach,
Oxford University Press, Oxford, 2000, pp. 235–247.
[19] A.E.B. Bueno da Silva, G. Brumatti, F.O. Russo, D.R. Green,
G.P. Amarante-Mendes, Bcr-Abl-mediated resistance to
apoptosis is independent of constant tyrosine-kinase activity,
Cell. Death Differ. 10 (2003) 592–598.
[20] M. Crompton, The mitochondrial permeability transition pore
and its role in cell death, Biochem. J. 341 (1999) 233–249.
[21] M.J. Hansson, T. Persson, H. Friberg, M.F. Keep, A. Rees,
T. Wieloch, et al., Powerful cyclosporin inhibition of calcium-
induced permeability-transition in brain mitochondria, Brain
Res. 960 (2003) 99–111.
[22] S.H. Kaufmann, W.C. Earnshaw, Induction of apoptosis by
cancer chemotherapy, Exp. Cell Res. 256 (2000) 42–49.
[23] G.M. Cohen, Caspases: the executioners of apoptosis,
Biochem J. 326 (1997) 1–16.
[24] P. Li, D. Nijhawan, I. Budihardjo, S.M. Srinivasula,
M. Ahmad, E.S. Alnemri, et al., Cytochrome c and dATP-
dependent formation of Apaf-1/caspase-9 complex initiates an
apoptotic protease cascade, Cell 91 (1997) 479–489.
[25] I. Marzo, P. Perez-Galan, P. Giraldo, D. Rubio-Felix, A. Anel,
J. Naval, Cladribine induces apoptosis in human leukaemia
cells by caspase dependent and -independent pathways acting
on mitochondria, Biochem. J. 359 (2001) 537–546.
[26] G. Beutner, A. Ruck, B. Riede, D. Brdiczka, Complexes
between porin, hexokinase, mitochondrial creatine kinase and
adenylate translocator displays properties of the permeability
transition pore. Implication for regulation of permeability
transition by the kinases, Biochim. Biophys. Acta 1368 (1998)
7–18.
[27] Y. Chen, D.L. Kramer, P. Diegelman, S. Vujcic, C.W. Porter,
Apoptotic Signaling in Polyamine Analogue-treated SK-
MEL-28 Human Melanoma Cells, Cancer Res. 61 (2001)
6437–6444.
[28] S.A. Susin, N. Zamzami, M. Castedo, E. Daugas, H-G. Wang,
S. Geley, et al., The central executioner of apoptosis: multiple
connections between protease activation and mitochondria in
Fas/Apo-1/CD-95 and ceramide-induced apoptosis, J. Exp.
Med. 186 (1997) 25–37.
Related Documents











