editor sally a. moody volume 48 no. 4 april 2010 The Journal of Genetics and Development ISSN 1526-954X genesis pages 213–280 volume 48, number 4, april 2010 genesis

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

editor sally a. moody
vo
lum
e 4
8 n
o. 4
april 2010
The Journal of Genetics and Development
ISSN 1526-954X
genesis
pa
ge
s 2
13–2
80
vo
lum
e 4
8, n
um
be
r 4, a
pril 2
010
ge
nesis

ARTICLE
Polyploidy-Associated Genomic Instability inArabidopsis thalianaYixing Wang,1 Ajay K. Jha,1 Rujin Chen,2 John H. Doonan,3 and Ming Yang1*1Department of Botany, Oklahoma State University, Stillwater, Oklahoma2Plant Biology Division, The S. R. Noble Foundation, Ardmore, Oklahoma3Department of Cell and Developmental Biology, John Innes Center, Norwich, United Kingdom
Received 22 November 2009; Accepted 12 January 2010
Summary: Formation of polyploid organisms by fertiliza-tion of unreduced gametes in meiotic mutants is believedto be a common phenomenon in species evolution.However, not well understood is how species in naturegenerally exist as haploid and diploid organisms in a longevolutionary time while polyploidization must have repeat-edly occurred via meiotic mutations. Here, we show thatthe ploidy increased for two consecutive generations dueto unreduced but viable gametes in the Arabidopsis cyclina1;2-2 (also named tardy asynchronousmeiosis-2) mutant,but the resultant octaploid plants produced progeny of ei-ther the same or reduced ploidy via genomic reductionsduring meiosis and pollen mitosis. Ploidy reductionsthrough sexual reproduction were also observed in inde-pendently generated artificial octaploid and hexaploidArabidopsis plants. These results demonstrate that octa-ploid is likely the maximal ploidy produced through sexualreproduction in Arabidopsis. The polyploidy-associatedgenomic instability may be a general phenomenon thatconstrains ploidy levels in species evolution. genesis48:254–263, 2010. VVC 2010Wiley-Liss, Inc.
Key words: cell cycle; flow cytometry; cyclin; aneuploid;generative cell; dyad
INTRODUCTION
Polyploidization is a common phenomenon in the evolu-tionary history of species and undoubtedly a major fac-tor shaping the genomic structures of many species(Otto, 2007). Analyses of plant (Fawcett et al., 2009;Jiang et al., 2009; Rousseau-Gueutin et al., 2008) andanimal (Evans et al., 2004; Macqueen and Johnston,2008; Moghadam et al., 2009; Semon and Wolf, 2007)genomes have uncovered both ancient and recent poly-ploidization events, underscoring the prevalence of pol-yploidization. However, the prevalence of polyploidy isalso contrasted with the fact that haploidy and diploidyare the dominant ploidy levels in nature, implicating theexistence of counter-ploidy-increase mechanisms.
A reasonable hypothesis for explaining the dominanceof low ploidy levels in nature is that high ploidy levelsare unstable, that is, they quickly revert to lower ploidy
levels if they ever arise from polyploidization events.Many studies involving mitosis in a polyploid back-ground support this hypothesis. In fact, polyploidy-asso-ciated genomic instability in the form of deletion of chro-mosomes and/or chromosomal fragments has been stud-ied extensively in animals and humans for its connectionwith tumorigenesis (Davis et al., 2008; Nguyen et al.,2009; Roh et al., 2008; Sliwinska et al., 2009). Studies inyeast also suggest that triploidy and tetraploidy pose geo-metric constraints for chromosome segregation, leadingto genomic instability (Mayer and Aguilera, 1990; Storch-ova et al., 2006). In plants, perhaps because of technicaldifficulties and mostly benign consequences, genomicinstability during mitosis in a polyploid background hasseldom been directly investigated. In contrast, genomicinstability during meiosis in polyploid plants has beenreported, because it is often detrimental to plant fertility(Madlung et al., 2005). Studies in plants also indicatethat polyploidy and segmental duplication followed bygene losses are standard phenomena in the evolutionaryhistory of all flowering plants (Bennetzen, 2007).
A majority of modern plant species are predicted tohave a polyploid ancestry (Matzke et al., 1999). Poly-ploid plant species are more common than polyploidanimal species probably because plants lack the pachy-tene checkpoint, which renders plants tolerant to aber-rant chromosome pairing in polyploid meiocytes (Liet al., 2009). Plants thus are attractive systems for inves-tigating the dynamics of polyploidy over multiple genera-tions. Allopolyploid plants resulting from hybridizationof related species have been studied, which uncovereddramatic and rapid genetic and epigenetic changes inseveral generations (Buggs et al., 2009; Chaudhary et al.,2009; Eilam et al., 2008; Madlung et al., 2005). However,
* Correspondence to: Ming Yang, Department of Botany, Oklahoma State
University, 104 Life Sciences East, Stillwater, OK 74078, USA.
E-mail: [email protected]
Contract grant sponsors: The Underwood Fellowship, BBSRC, The
Samuel Roberts Noble Foundation, Oklahoma State University.Published online 8 February 2010 in
Wiley InterScience (www.interscience.wiley.com).
DOI: 10.1002/dvg.20610
' 2010 Wiley-Liss, Inc. genesis 48:254–263 (2010)

compared to allopolyploid plants, much less is knownabout genomic changes over generations in autopolyploidplants although reductions in genomic content weredetected in some autotetraploid plants (Eilam et al., 2008).
Unreduced gametes due to meiotic mutations arebelieved to be the major cause of autopolyploidy. Differ-ing from allopolyploid organisms, autopolyploid organ-isms arising from meiotic mutations have the potentialto double their genomes in every generation throughsexual reproduction. Several such mutants in Arabidop-sis, including dyad/switch1 (Ravi et al., 2008), atps1
(d’Erfurth et al., 2008), and osd1 (d’Erfurth et al., 2009),have been recently reported. In the case of osd1, octa-ploidy was observed after two rounds of genome dupli-cation, but the ploidy in the subsequent generation isunknown. Thus, the crucial question of whether the ge-nome continues to increase its size in the postoctaploidgenerations has not been answered.
We previously reported that the partial loss-of-func-tion tam (now designated as tam-1) mutant exhibitsdelayed cell-cycle progression in male meiosis, so thatdyads instead of tetrads are first formed but in generalthe dyads can continue to divide and form tetrads (Mag-nard et al., 2001; Wang et al., 2004). Here, we reportthat a null allele, tam-2, is completely blocked fromentry into meiosis II, producing unreduced gametes. Wealso present evidence that polyploid genomes derivedfrom the unreduced gametes or artificial means under-went frequent reductions in ploidy due to aberrant chro-mosome segregation and/or cell budding during meiosis,and in octaploid tam-2 the formation of three nuclei ofreduced size during mitosis in the generative cell. Theseresults collectively indicate that autopolyploid genomesare intrinsically unstable, limiting the ability of Arabidop-sis to increase ploidy through sexual reproduction.
RESULTS
tam-2 Is a Null Mutant Whose MeiocytesProduce Unreduced Gametes
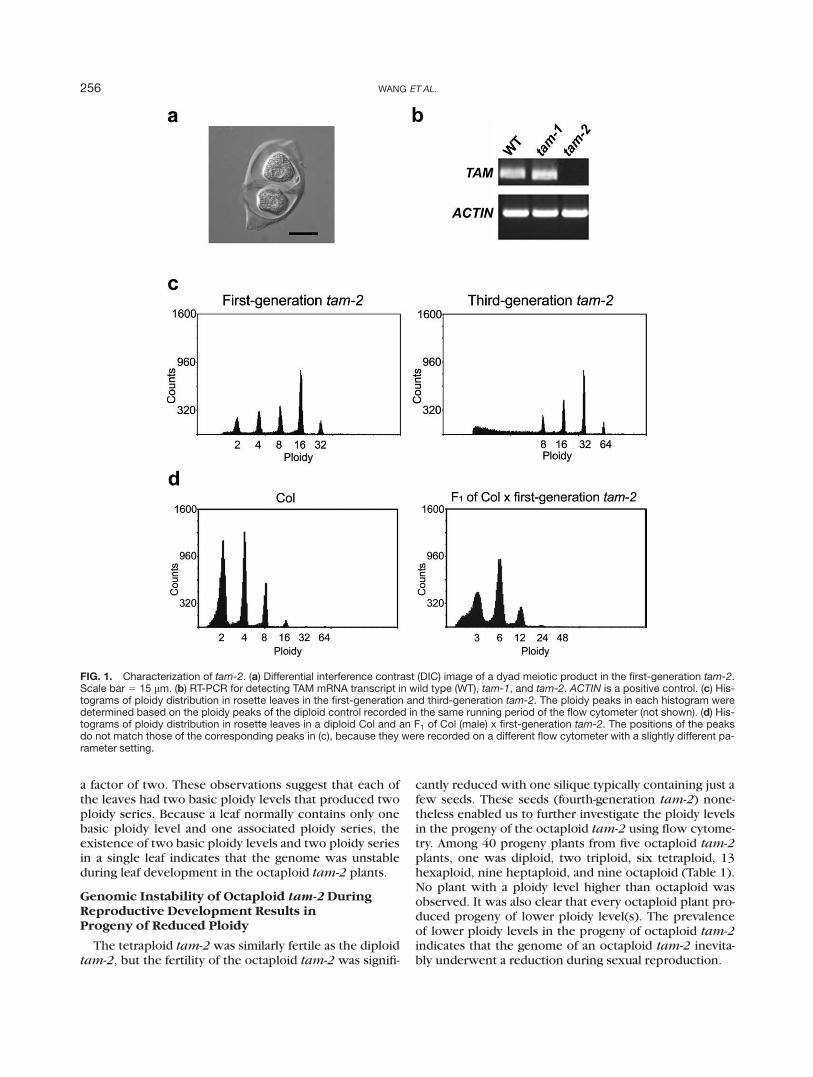
To further understand the meiotic function of TAM, anull mutant of tam was sought among the Salk lines thatharbor a T-DNA insertion in or near the coding region ofTAM in www.arabidopsis.org. We selected Salk_080686that contains the T-DNA insertion in the third last intronof TAM as a candidate line. The location of the insertionwas verified with genomic polymerase chain reactions(PCR) and sequencing of the PCR products (data notshown). The homozygous mutant plants as the first-gen-eration progeny of the heterozygous parent did not ex-hibit an obvious defect in overall development, but pro-duced only dyad meiotic products in male meiosis (Fig.1a); triads and tetrads were not found. This meioticdefect was similar to, but more severe than, that of thepreviously reported tam mutant (Magnard et al., 2001).The new mutant was then designated as tam-2 and theprevious one tam-1. The TAM mRNA transcript couldnot be detected in tam-2 by reverse transcription-PCR
(RT-PCR), indicating that tam-2 is a null mutant (Fig.1b). However, remarkably, these tam-2 plants were fer-tile with siliques of apparently normal length althoughsome of the seeds shrank and died before maturity.
The dyad phenotype suggests that at least the male ga-mete may be unreduced in tam-2, which would lead topolyploidization in the next generation. To test this possi-bility, ploidy levels of tam-2 of the first and the next twogenerations were first determined using flow cytometry.The first-generation tam-2 was found to be diploid, thesecond-generation tetraploid, and the third-generationoctaploid (Fig. 1c and data not shown). We then did re-ciprocal crosses between wild-type Arabidopsis plantsand the first-generation tam-2 plants and performed flowcytometric analysis on nine of the F1 plants, four fromCol (male) 3 tam-2 and five from tam-2 (male) 3 Col.All these plants turned out to be triploid (Fig. 1d and datanot shown). Consistent with the ploidy levels, seeds fromthe mutant plants were larger than seeds from normaldiploid plants (data not shown). These results indicatethat both the male and female gametes were unreduced,and the unreduced gametes had twice resulted in ge-nome doubling over two reproductive cycles.
tam-2 Undergoes Meiosis I But Skips Meiosis II
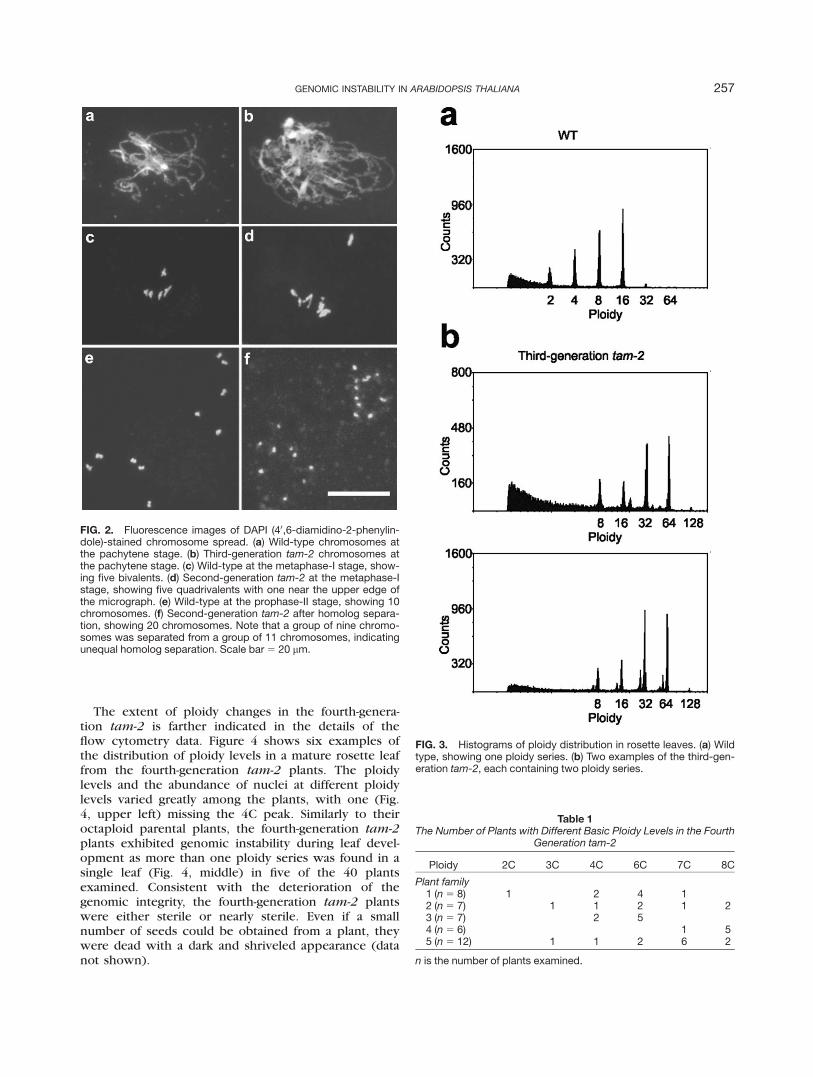
To further understand the nature of the meiotic defectin tam-2, we performed chromosome spread in malemeiosis in tetraploid and octaploid tam-2 plants. Figure 2bshows that homologous chromosomes in octaploid tam-2
underwent synapsis in prophase I, resembling the packy-tene chromosomes in the wild type (Fig. 2a). Five groupsof chromosomes were observed in both wild type (Fig. 2c)and tetraploid (Fig. 2d) and octaploid tam-2 (not shown)at metaphase I. Each chromosome group in Figure 2d waspredicted to be a quadrivalent, because the plant was tet-raploid. Indeed, 20 chromosomes were identified in thisplant at a stage near the end of meiosis I (Fig. 2f), in con-trast with the 10 chromosomes observed at prophase II inthe wild type. These chromosomal features indicate thathomologous chromosomes in tam-2 behaved similarly asthose of wild type in meiosis I, and the formation of dyadsis a result of the absence of meiosis II.
The Genome of Octaploid tam-2 Is UnstableDuring Leaf Development
In a wild-type leaf with one basic ploidy such as dip-loidy, endoreduplications always result in one ploidy se-ries typically consisting of 2C, 4C, 8C, 16C, 32C, and64C (Fig. 3a). In fact, the consistency of one ploidyseries per leaf was confirmed in 130 diploid plantsincluding tam-1 and first-generation tam-2 (Fig. 1c, left),and in four wild-type tetraploid plants. In these cases,the adjacent ploidy levels always differ by a factor oftwo. However, in two of nine third-generation tam-2
plants examined, some of the adjacent ploidy levels con-sistently differed by a factor less than two (Fig. 3b).These extra ploidy peaks were at the positions that wereconsistent with the existence of an additional ploidyseries whose own adjacent ploidy levels also differed by
255GENOMIC INSTABILITY IN ARABIDOPSIS THALIANA
a factor of two. These observations suggest that each ofthe leaves had two basic ploidy levels that produced twoploidy series. Because a leaf normally contains only onebasic ploidy level and one associated ploidy series, theexistence of two basic ploidy levels and two ploidy seriesin a single leaf indicates that the genome was unstableduring leaf development in the octaploid tam-2 plants.
Genomic Instability of Octaploid tam-2 DuringReproductive Development Results inProgeny of Reduced Ploidy
The tetraploid tam-2 was similarly fertile as the diploidtam-2, but the fertility of the octaploid tam-2 was signifi-
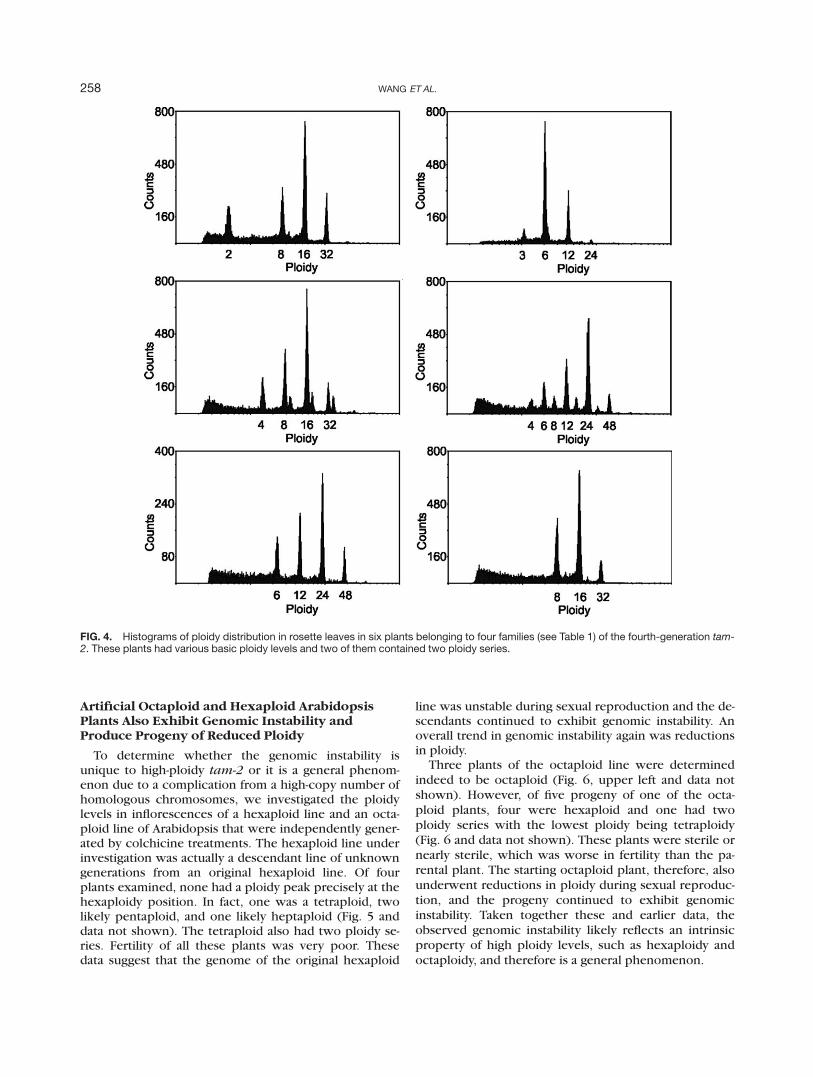
cantly reduced with one silique typically containing just afew seeds. These seeds (fourth-generation tam-2) none-theless enabled us to further investigate the ploidy levelsin the progeny of the octaploid tam-2 using flow cytome-try. Among 40 progeny plants from five octaploid tam-2
plants, one was diploid, two triploid, six tetraploid, 13hexaploid, nine heptaploid, and nine octaploid (Table 1).No plant with a ploidy level higher than octaploid wasobserved. It was also clear that every octaploid plant pro-duced progeny of lower ploidy level(s). The prevalenceof lower ploidy levels in the progeny of octaploid tam-2indicates that the genome of an octaploid tam-2 inevita-bly underwent a reduction during sexual reproduction.
FIG. 1. Characterization of tam-2. (a) Differential interference contrast (DIC) image of a dyad meiotic product in the first-generation tam-2.Scale bar 5 15 lm. (b) RT-PCR for detecting TAM mRNA transcript in wild type (WT), tam-1, and tam-2. ACTIN is a positive control. (c) His-tograms of ploidy distribution in rosette leaves in the first-generation and third-generation tam-2. The ploidy peaks in each histogram weredetermined based on the ploidy peaks of the diploid control recorded in the same running period of the flow cytometer (not shown). (d) His-tograms of ploidy distribution in rosette leaves in a diploid Col and an F1 of Col (male) x first-generation tam-2. The positions of the peaksdo not match those of the corresponding peaks in (c), because they were recorded on a different flow cytometer with a slightly different pa-rameter setting.
256 WANG ET AL.
The extent of ploidy changes in the fourth-genera-tion tam-2 is farther indicated in the details of theflow cytometry data. Figure 4 shows six examples ofthe distribution of ploidy levels in a mature rosette leaffrom the fourth-generation tam-2 plants. The ploidylevels and the abundance of nuclei at different ploidylevels varied greatly among the plants, with one (Fig.4, upper left) missing the 4C peak. Similarly to theiroctaploid parental plants, the fourth-generation tam-2
plants exhibited genomic instability during leaf devel-opment as more than one ploidy series was found in asingle leaf (Fig. 4, middle) in five of the 40 plantsexamined. Consistent with the deterioration of thegenomic integrity, the fourth-generation tam-2 plantswere either sterile or nearly sterile. Even if a smallnumber of seeds could be obtained from a plant, theywere dead with a dark and shriveled appearance (datanot shown).
FIG. 2. Fluorescence images of DAPI (40,6-diamidino-2-phenylin-dole)-stained chromosome spread. (a) Wild-type chromosomes atthe pachytene stage. (b) Third-generation tam-2 chromosomes atthe pachytene stage. (c) Wild-type at the metaphase-I stage, show-ing five bivalents. (d) Second-generation tam-2 at the metaphase-Istage, showing five quadrivalents with one near the upper edge ofthe micrograph. (e) Wild-type at the prophase-II stage, showing 10chromosomes. (f) Second-generation tam-2 after homolog separa-tion, showing 20 chromosomes. Note that a group of nine chromo-somes was separated from a group of 11 chromosomes, indicatingunequal homolog separation. Scale bar5 20 lm.
FIG. 3. Histograms of ploidy distribution in rosette leaves. (a) Wildtype, showing one ploidy series. (b) Two examples of the third-gen-eration tam-2, each containing two ploidy series.
Table 1The Number of Plants with Different Basic Ploidy Levels in the Fourth
Generation tam-2
Ploidy 2C 3C 4C 6C 7C 8C
Plant family1 (n 5 8) 1 2 4 12 (n 5 7) 1 1 2 1 23 (n 5 7) 2 54 (n 5 6) 1 55 (n 5 12) 1 1 2 6 2
n is the number of plants examined.
257GENOMIC INSTABILITY IN ARABIDOPSIS THALIANA
Artificial Octaploid and Hexaploid ArabidopsisPlants Also Exhibit Genomic Instability andProduce Progeny of Reduced Ploidy
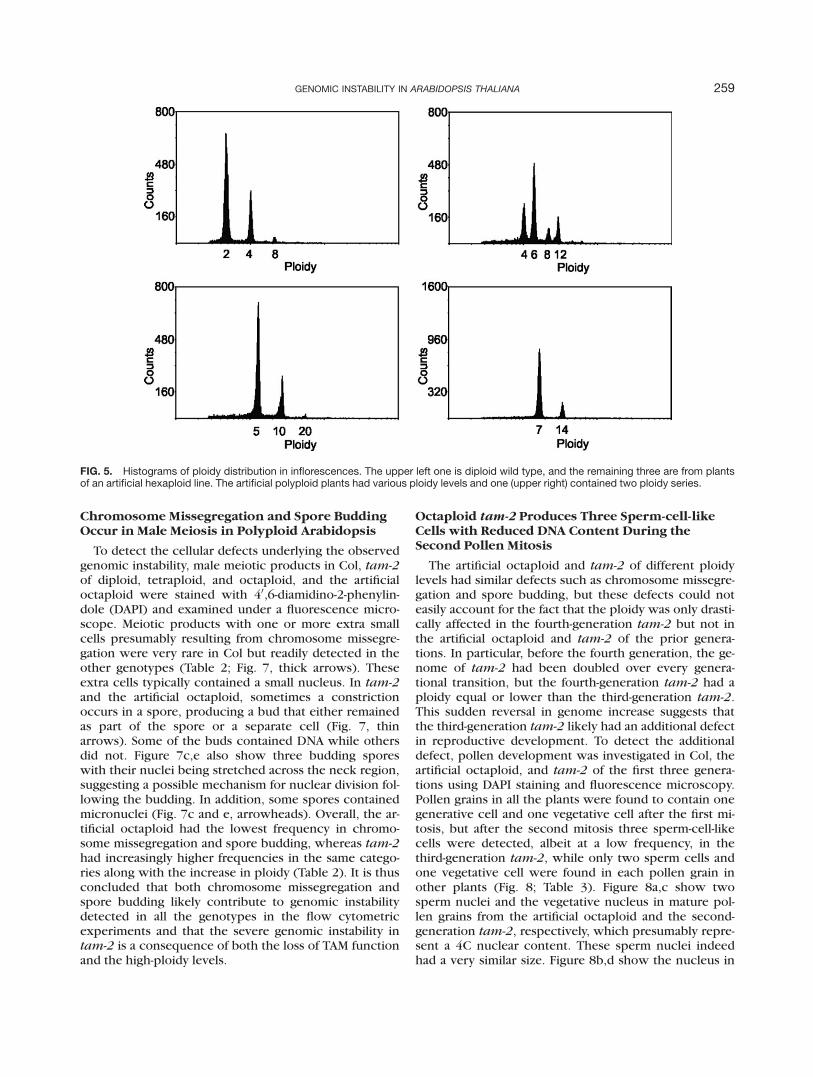
To determine whether the genomic instability isunique to high-ploidy tam-2 or it is a general phenom-enon due to a complication from a high-copy number ofhomologous chromosomes, we investigated the ploidylevels in inflorescences of a hexaploid line and an octa-ploid line of Arabidopsis that were independently gener-ated by colchicine treatments. The hexaploid line underinvestigation was actually a descendant line of unknowngenerations from an original hexaploid line. Of fourplants examined, none had a ploidy peak precisely at thehexaploidy position. In fact, one was a tetraploid, twolikely pentaploid, and one likely heptaploid (Fig. 5 anddata not shown). The tetraploid also had two ploidy se-ries. Fertility of all these plants was very poor. Thesedata suggest that the genome of the original hexaploid
line was unstable during sexual reproduction and the de-scendants continued to exhibit genomic instability. Anoverall trend in genomic instability again was reductionsin ploidy.
Three plants of the octaploid line were determinedindeed to be octaploid (Fig. 6, upper left and data notshown). However, of five progeny of one of the octa-ploid plants, four were hexaploid and one had twoploidy series with the lowest ploidy being tetraploidy(Fig. 6 and data not shown). These plants were sterile ornearly sterile, which was worse in fertility than the pa-rental plant. The starting octaploid plant, therefore, alsounderwent reductions in ploidy during sexual reproduc-tion, and the progeny continued to exhibit genomicinstability. Taken together these and earlier data, theobserved genomic instability likely reflects an intrinsicproperty of high ploidy levels, such as hexaploidy andoctaploidy, and therefore is a general phenomenon.
FIG. 4. Histograms of ploidy distribution in rosette leaves in six plants belonging to four families (see Table 1) of the fourth-generation tam-2. These plants had various basic ploidy levels and two of them contained two ploidy series.
258 WANG ET AL.
Chromosome Missegregation and Spore BuddingOccur in Male Meiosis in Polyploid Arabidopsis
To detect the cellular defects underlying the observedgenomic instability, male meiotic products in Col, tam-2
of diploid, tetraploid, and octaploid, and the artificialoctaploid were stained with 40,6-diamidino-2-phenylin-dole (DAPI) and examined under a fluorescence micro-scope. Meiotic products with one or more extra smallcells presumably resulting from chromosome missegre-gation were very rare in Col but readily detected in theother genotypes (Table 2; Fig. 7, thick arrows). Theseextra cells typically contained a small nucleus. In tam-2
and the artificial octaploid, sometimes a constrictionoccurs in a spore, producing a bud that either remainedas part of the spore or a separate cell (Fig. 7, thinarrows). Some of the buds contained DNA while othersdid not. Figure 7c,e also show three budding sporeswith their nuclei being stretched across the neck region,suggesting a possible mechanism for nuclear division fol-lowing the budding. In addition, some spores containedmicronuclei (Fig. 7c and e, arrowheads). Overall, the ar-tificial octaploid had the lowest frequency in chromo-some missegregation and spore budding, whereas tam-2
had increasingly higher frequencies in the same catego-ries along with the increase in ploidy (Table 2). It is thusconcluded that both chromosome missegregation andspore budding likely contribute to genomic instabilitydetected in all the genotypes in the flow cytometricexperiments and that the severe genomic instability intam-2 is a consequence of both the loss of TAM functionand the high-ploidy levels.
Octaploid tam-2 Produces Three Sperm-cell-likeCells with Reduced DNA Content During theSecond Pollen Mitosis
The artificial octaploid and tam-2 of different ploidylevels had similar defects such as chromosome missegre-gation and spore budding, but these defects could noteasily account for the fact that the ploidy was only drasti-cally affected in the fourth-generation tam-2 but not inthe artificial octaploid and tam-2 of the prior genera-tions. In particular, before the fourth generation, the ge-nome of tam-2 had been doubled over every genera-tional transition, but the fourth-generation tam-2 had aploidy equal or lower than the third-generation tam-2.This sudden reversal in genome increase suggests thatthe third-generation tam-2 likely had an additional defectin reproductive development. To detect the additionaldefect, pollen development was investigated in Col, theartificial octaploid, and tam-2 of the first three genera-tions using DAPI staining and fluorescence microscopy.Pollen grains in all the plants were found to contain onegenerative cell and one vegetative cell after the first mi-tosis, but after the second mitosis three sperm-cell-likecells were detected, albeit at a low frequency, in thethird-generation tam-2, while only two sperm cells andone vegetative cell were found in each pollen grain inother plants (Fig. 8; Table 3). Figure 8a,c show twosperm nuclei and the vegetative nucleus in mature pol-len grains from the artificial octaploid and the second-generation tam-2, respectively, which presumably repre-sent a 4C nuclear content. These sperm nuclei indeedhad a very similar size. Figure 8b,d show the nucleus in
FIG. 5. Histograms of ploidy distribution in inflorescences. The upper left one is diploid wild type, and the remaining three are from plantsof an artificial hexaploid line. The artificial polyploid plants had various ploidy levels and one (upper right) contained two ploidy series.
259GENOMIC INSTABILITY IN ARABIDOPSIS THALIANA
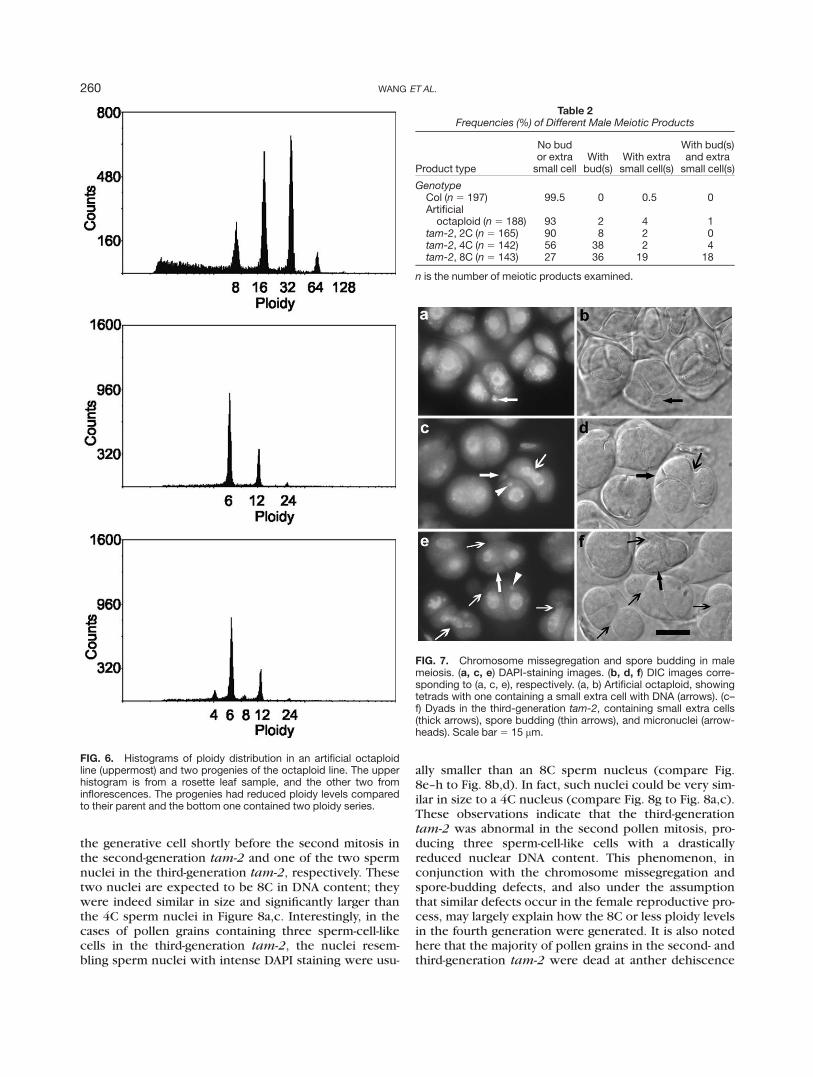
the generative cell shortly before the second mitosis inthe second-generation tam-2 and one of the two spermnuclei in the third-generation tam-2, respectively. Thesetwo nuclei are expected to be 8C in DNA content; theywere indeed similar in size and significantly larger thanthe 4C sperm nuclei in Figure 8a,c. Interestingly, in thecases of pollen grains containing three sperm-cell-likecells in the third-generation tam-2, the nuclei resem-bling sperm nuclei with intense DAPI staining were usu-
ally smaller than an 8C sperm nucleus (compare Fig.8e–h to Fig. 8b,d). In fact, such nuclei could be very sim-ilar in size to a 4C nucleus (compare Fig. 8g to Fig. 8a,c).These observations indicate that the third-generationtam-2 was abnormal in the second pollen mitosis, pro-ducing three sperm-cell-like cells with a drasticallyreduced nuclear DNA content. This phenomenon, inconjunction with the chromosome missegregation andspore-budding defects, and also under the assumptionthat similar defects occur in the female reproductive pro-cess, may largely explain how the 8C or less ploidy levelsin the fourth generation were generated. It is also notedhere that the majority of pollen grains in the second- andthird-generation tam-2 were dead at anther dehiscence
FIG. 6. Histograms of ploidy distribution in an artificial octaploidline (uppermost) and two progenies of the octaploid line. The upperhistogram is from a rosette leaf sample, and the other two frominflorescences. The progenies had reduced ploidy levels comparedto their parent and the bottom one contained two ploidy series.
Table 2Frequencies (%) of Different Male Meiotic Products
Product type
No budor extrasmall cell
Withbud(s)
With extrasmall cell(s)
With bud(s)and extrasmall cell(s)
GenotypeCol (n5 197) 99.5 0 0.5 0Artificialoctaploid (n 5 188) 93 2 4 1
tam-2, 2C (n5 165) 90 8 2 0tam-2, 4C (n5 142) 56 38 2 4tam-2, 8C (n5 143) 27 36 19 18
n is the number of meiotic products examined.
FIG. 7. Chromosome missegregation and spore budding in malemeiosis. (a, c, e) DAPI-staining images. (b, d, f) DIC images corre-sponding to (a, c, e), respectively. (a, b) Artificial octaploid, showingtetrads with one containing a small extra cell with DNA (arrows). (c–f) Dyads in the third-generation tam-2, containing small extra cells(thick arrows), spore budding (thin arrows), and micronuclei (arrow-heads). Scale bar5 15 lm.
260 WANG ET AL.

according to Evans blue staining (not shown; Gaff andOkong’o-Ogola, 1971), which is consistent with the lowfertility observed in these plants.
DISCUSSION
We have demonstrated that hexaploid and octaploidgenomes are unstable during Arabidopsis vegetative andreproductive development. We have also showed an evo-lutionary path that starts from a diploid mutant produc-ing unreduced gametes to limited ploidy increases tooctaploidy and to inevitable breakdown of the octaploidyinto lower ploidy levels. These plants of high-ploidy lev-els, regardless of their origins, ultimately have few chan-ces to reproduce the next generation due to severelyreduced fertility. The frequencies of cellular defects caus-ing genomic instability, namely chromosome missegrega-
tion, spore budding, and usual formation of three sperm-cell-like cells, are clearly increased along with theincrease in ploidy in tam-2. These findings have an impli-cation to mechanisms whereby organisms attain low-ploidy levels in nature over a long evolutionary time.Indeed, organisms undergoing sexual reproduction rarelyhave a ploidy level exceeding hexaploidy or octaploidy.
We observed a new phenomenon, that is, spore bud-ding, which most frequently occurs in octaploid tam-2.A nucleus can be stretched across the neck region of thebudding spore, possibly also causing the budding of thenucleus. This phenomenon appears to resemble the ‘‘nu-clear budding’’ fashion of cell division, recently termedneosis, in human cancer cell lines (Rajaraman et al.,2005). The cancer cells first become polyploid upon adrug treatment, and then some of them undergo karyoki-nesis via nuclear budding followed by asymmetric, intra-
FIG. 8. DAPI-stained pollen grains. (a) Mature pollen grain from the artificial octaploid, showing the two sperm nuclei with intense DAPI stain-ing and the vegetative nucleus with faint DAPI staining. The ploidy of the nuclei should be 4C. (b) Pollen grain from the second-generationtam-2 after the first mitosis, showing the generative nucleus with an expected 8C nuclear DNA content as the 4C genome should have beenduplicated at this stage. (c) Mature pollen grain from the second-generation tam-2, showing the two sperm nuclei and the vegetative nuclei.The ploidy of the nuclei is expected to be 4C, and indeed the sperm nuclei are similar in size to those in (a). (d) Mature pollen grain from thethird-generation tam-2, showing one sperm nucleus (the other is out of the focal plane) and the vegetative nucleus. The ploidy of the nuclei isexpected to be 8C, which is consistent with the sperm nucleus being similar in size to the generative nucleus in (b). (e–g) Three focal planes ofone mature pollen grain from the same third-generation tam-2 as in (d), showing three nuclei resembling a sperm nucleus. These nuclei arepredicted to have a ploidy significantly lower than 8C. Note the presence of the vegetative nucleus in (e). (h) Another mature pollen grain fromthe same plant as in (d–g), showing one of the three nuclei resembling a sperm nucleus (the others not shown). Scale bar5 15 lm.
Table 3Abnormal Second Mitosis in Pollen in Octaploid tam-2
Genotype Cells from first division Cells from second division
Col Generative and vegetative cells Two sperm cellsArtificial octaploid Generative and vegetative cells Two sperm cellstam-2, 2C Generative and vegetative cells Two sperm cellstam-2, 4C Generative and vegetative cells Two sperm cellstam-2, 8C Generative and vegetative cells Three sperm-cell-like cells
For each developmental stage in each genotype, pollen grains from at least 18 anthers were carefully examined. No unusual cell numberwas observed in Col, the artificial octaploid, and tetraploid tam-2 after both mitoses and in octaploid tam-2 after the first mitosis. However,several pollen grains containing three sperm-cell-like cells were observed in anthers from each bud of octaploid tam-2 after the secondmitosis.
261GENOMIC INSTABILITY IN ARABIDOPSIS THALIANA

cellular cytokinesis, producing several small mononu-clear cells, called the Raju cells. Some of the smallbudded cells become drug-resistant, inherit genomicinstability, and regain the ability to divide mitotically(Mansilla et al., 2009; Rajaraman et al., 2005), indicatingthat a new genetic makeup has been attained in thesecells. The spore budding phenomenon most frequentlyoccurring in octaploid tam-2 raises the possibility thatneosis may be a common phenomenon in polyploids ofboth animals and plants. In Storchova et al.’s (2006)work on screening for mutants with polyploid-specificlethality, almost all the mutations identified affectgenomic stability by impairing homologous recombina-tion, sister chromatid cohesion, or mitotic spindle func-tion. These aspects of cellular functions are expected tobe conserved in all eukaryotes, which may be the basisfor neosis to be a common phenomenon in polyploid eu-karyotic cells. It also remains to be investigated whetherthe formation of three sperm-cell-like cells in octaploidtam-2 is via a process similar to the spore budding pro-cess. It can be speculated that because the generativecells in octaploid tam-2 have a nuclear DNA content ofpossibly eight times of that in the generative cells in dip-loid wild type, the genomes in generative cells in octa-ploid tam-2 are unstable. Further investigation into thespore budding and generative cell division processes inoctaploid tam-2 may yield useful information on themechanism of neosis, possibly providing an insightabout the process of cancer formation.
As shown in this report, the evolutionary path mostlikely leads to sterility in a meiotic mutant undergoingautopolyploidization. However, considering the possibil-ities that it takes several generations to reach the deadend for most plants, that recessive mutations can lurkaround for a long time, and that new genetic composi-tions may emerge through neosis, some spores may ulti-mately escape the death trap and produce new varietiesor even new species. Furthermore, as such plantsapproaching sterility in a natural habitat, they dramati-cally increase the chances of being pollinated by neigh-boring fertile plants. Such hybridizations could also dra-matically alter the evolutionary fate of these plants.
Apomixis is a mode of reproduction by special genera-tive tissues without fertilization. One apomictic mecha-nism in plant reproduction is that an unreduced eggdevelops into an embryo. An apomict thus has agenomic content identical to that of its mother. Suchstrict inheritance of genetic materials may be beneficialto agricultural productions. It is not understood howapomixis has evolved in nature, given that more thanone genetic modification is needed to bring about all thenecessary traits associated with an apomictic reproduc-tive process. Our finding indicates that a meiotic mutantcan become autopolyploids that can further undergoneosis-like cell divisions to potentially produce gametesof different genetic compositions. These linked dramaticevents suggest that achieving the right combinations ofgenetic composition for apomixis may not insurmount-able after all.
The result that no ploidy levels higher than octaploidwere found in the progeny of octaploid plants is intrigu-ing. At least some of the large pollen grains in the octa-ploid tam-2 appear viable at anthesis (see Fig. 8). There-fore, the gametes that can potentially give rise to ploidylevels higher than octaploid might be somehowexcluded from contribution to the formation of the nextgeneration at stages after pollen release from the antherbut before fertilization. Alternatively, the eliminationmight occur after fertilization, that is, the embryo and itsassociated endosperm might not be able to develop intomaturity when their ploidy levels exceed those of theoctaploid plants. Further investigation into the fertiliza-tion and embryo developmental processes may revealhow the ploidy maximum is controlled.
METHODS
Plant Materials and Growth Conditions
All Arabidopsis plants are in the Columbia (Col) eco-type background. The tetraploid and hexaploid wild-type lines are gifts from Rod Scott at University of Bath.The octaploid wild-type line is a gift from Hirokazu Tsu-kaya at University of Tokyo. tam-2 was obtained fromthe Arabidopsis Biological Resource Center in Ohio.Plants were grown either in growth chambers with a 16h light/8-h dark photoperiod at about 228C or a green-house in the John Innes Center, Norwich, UK, with natu-ral light supplemented with artificial light at somewhatvariable temperatures around 208C. In all cases, an artifi-cial soil was used. Because the T-DNA insertion containsa kanamycin selection marker, homozygous and hetero-zygous tam-2 plants in a segregation population from aheterozygous parental plant were also selected on anagar medium containing kanamycin before transplantingthe resistant plants to the soil.
T-DNA Insertion Confirmation and GeneExpression Analysis
For confirmation of the T-DNA insertion in tam-2,genomic DNA was isolated from tam-2 and the wild-typeplants, and PCR was performed using the DNA samplesas templates, with an annealing temperature of 588C and35 cycles. PCR primers: GAAGAGTATAGGCTTTCGCCC,and TGCAACCACATCAGATGAGAG for amplification of agenomic fragment when the insertion is absent; and theT-DNA left border primer GCGTGGACCGCTTGCTGCAACT and TGCAACCACATCAGATGAGAG for a frag-ment when the insertion is present. The PCR productbearing the left border of the T-DNA was sequencedusing capillary electrophoresis (3730 DNA Analyzer,Applied Biosystem).
For RT-PCR for detecting the mRNA transcript, theRNA samples in Figure 1b were isolated using RNeasyPlant Mini kit (Qiagen, Valencia, CA) from the corre-sponding plants and used as templates. The SuperScriptOne-step RT-PCR kit (Invitrogen, Carlsbad, CA) and thesame PCR primers for amplification of the above
262 WANG ET AL.

genomic fragment were used. The PCR was conductedwith an annealing temperature at 588C and 40 cycles.PCR for the ACTIN control was conducted at the anneal-ing temperature of 588C and for 30 cycles with the fol-lowing primers: AAGCTGTTCTCTCCTTGTACGCC, andGTTGCACCACCTGAAAGGAAG.
Chromosome Spread, DAPI Staining, andLight Microscopy
Sample preparation for chromosome spread, DAPIstaining, light microscopy, and image capture and manip-ulation were essentially as previously described (Wanget al., 2004). The images were captured on either aNikon Eclipse E600 or E800 microscope fitted with a dig-ital camera using image acquisition software.
Flow Cytometry
Each sample for ploidy analysis was prepared fromone mature or two-three developing rosette leaves or anapex of an inflorescence bearing newly opened flowersand closed floral buds. The flow cytometers were PartecPloidy Analyzers. The experimental procedure wasaccording to, and the reagents were supplied by Partec.On each continuous run of the flow cytometer, diploidwild-type, or other Arabidopsis plants with knownploidy levels were included as the ploidy standard.
ACKNOWLEDGMENTS
The authors thank Max Bush, Vasilis Gegas, and AndyHopkins for assistance with the flow cytometric work.
LITERATURE CITED
Bennetzen JL. 2007. Patterns in grass genome evolution. Curr OpinPlant Biol 10:176–181.
Buggs RJ, Doust AN, Tate JA, Koh J, Soltis K, Feltus FA, Paterson AH, Sol-tis PS, Soltis DE. 2009. Gene loss and silencing in Tragopogon mis-
cellus (Asteraceae): Comparison of natural and synthetic allotetra-ploids. Heredity 103:73–81.
Chaudhary B, Flagel L, Stupar RM, Udall JA, Verma N, Springer NM,Wendel JF. 2009. Reciprocal silencing, transcriptional bias andfunctional divergence of homeologs in polyploid cotton (gos-sypium). Genetics 182:503–517.
Davis E, Teng H, Bilican B, Parker MI, Liu B, Carriera S, Goding CR,Prince S. 2008. Ectopic Tbx2 expression results in polyploidy andcisplatin resistance. Oncogene 27:976–984.
d’Erfurth I, Jolivet S, Froger N, Catrice O, Novatchkova M, Simon M,Jenczewski E, Mercier R. 2008. Mutations in AtPS1 (Arabidopsisthaliana parallel spindle 1) lead to the production of diploid pol-len grains. PLoS Genet 4: e1000274.
d’Erfurth I, Jolivet S, Froger N, Catrice O, Novatchkova M, Mercier R.2009. Turning meiosis into mitosis. PLoS Biol 7:e1000124.
Eilam T, Anikster Y, Millet E, Manisterski J, Feldman M. 2008. Nuclear DNAamount and genome downsizing in natural and synthetic allopoly-ploids of the genera Aegilops and Triticum. Genome 51:616–627.
Evans BJ, Kelley DB, Tinsley RC, Melnick DJ, Cannatella DC. 2004. A mi-tochondrial DNA phylogeny of African clawed frogs: Phylogeogra-phy and implications for polyploid evolution. Mol Phylogenet Evol33:197–213.
Fawcett JA, Maere S, Van de Peer Y. 2009. Plants with double genomesmight have had a better chance to survive the Cretaceous-Tertiaryextinction event. Proc Natl Acad Sci USA 106:5737–5742.
Gaff DF, Okong’o-Ogola O. 1971. The use of non-permeating pigmentsfor testing the survival of cells. J Exp Bot 22:756–758.
Jiang LY, Qian ZQ, Guo ZG, Wang C, Zhao GF. 2009. Polyploid originsin Gynostemma pentaphyllum (Cucurbitaceae) inferred frommultiple gene sequences. Mol Phylogenet Evol 52:183–191.
Li XC, Barringer BC, Barbash DA. 2009. The pachytene checkpoint andits relationship to evolutionary patterns of polyploidization andhybrid sterility. Heredity 102:24–30.
Macqueen DJ, Johnston IA. 2008. An update on MyoD evolution in tele-osts and a proposed consensus nomenclature to accommodate thetetraploidization of different vertebrate genomes. PLoS One 3:e1567.
Madlung A, Tyagi AP, Watson B, Jiang H, Kagochi T, Doerge RW, Mar-tienssen R, Comai L. 2005. Genomic changes in synthetic Arabi-dopsis polyploids. Genomic changes in synthetic Arabidopsis poly-ploids. Plant J 41:221–230.
Magnard JL, Yang M, Chen YC, Leary M, McCormick S. 2001. The Arabi-dopsis gene tardy asynchronous meiosis is required for the normalpace and synchrony of cell division during male meiosis. PlantPhysiol 127:1157–1166.
Mansilla S, Bataller M, Portugal J. 2009. A nuclear budding mechanismin transiently arrested cells generates drug-sensitive and drug-re-sistant cells. Biochem Pharmacol 78:123–132.
Matzke MA, Mittelsten Scheid O, Matzke AJ. 1999. Rapid structural andepigenetic changes in polyploid and aneuploid genomes. Bioessays21:761–767.
Mayer VW, Aguilera A. 1990. High levels of chromosome instability inpolyploids of Saccharomyces cerevisiae. Mutat Res 231:177–186.
Moghadam HK, Ferguson MM, Danzmann RG. 2009. Comparativegenomics and evolution of conserved noncoding elements (CNE)in rainbow trout. BMC Genom 10:278.
Nguyen HG, Makitalo M, Yang D, Chinnappan D, St Hilaire C, Ravid K.2009. Deregulated Aurora-B induced tetraploidy promotes tumori-genesis. FASEB J 23:2741–2748.
Otto SP. 2007. The evolutionary consequences of polyploidy. Cell131:452–462.
Rajaraman R, Rajaraman MM, Rajaraman SR, Guernsey DL. 2005. Neo-sis–a paradigm of self-renewal in cancer. Cell Biol Int 29:1084–1097.
Ravi M, Marimuthu MP, Siddiqi I. 2008. Gamete formation withoutmeiosis in Arabidopsis Nature 451:1121–1124.
Roh M, Franco OE, Hayward SW, van der Meer R, Abdulkadir SA. 2008.A role for polyploidy in the tumorigenicity of Pim-1-expressinghuman prostate and mammary epithelial cells. LoS One 3:e2572.
Rousseau-Gueutin M, Lerceteau-Kohler E, Barrot L, Sargent DJ, MonfortA, Simpson D, Arus P, Guerin G, Denoyes-Rothan B. 2008. Compar-ative genetic mapping between octaploid and diploid Fragaria spe-cies reveals a high level of colinearity between their genomes andthe essentially disomic behavior of the cultivated octaploid straw-berry. Genetics 179:2045–2060.
Semon M, Wolfe KH. 2007. Rearrangement rate following the whole-ge-nome duplication in teleosts. Mol Biol Evol 24:860–867.
Sliwinska MA, Mosieniak G, Wolanin K, Babik A, Piwocka K, MagalskaA, Szczepanowska J, Fronk J, Sikora E. 2009. Induction of senes-cence with doxorubicin leads to increased genomic instability ofHCT116 cells. Mech Ageing Dev 130:24–32.
Storchova Z, Breneman A, Cande J, Dunn J, Burbank K, O’Toole E, Pell-man D. 2006. Genome-wide genetic analysis of polyploidy in yeast.Nature 443:541–547.
Walen KH. 2008. Genetic stability of senescence reverted cells: Ge-nome reduction division of polyploidy cells, aneuploidy and neo-plasia. Cell Cycle 7:1623–1629.
Wang Y, Magnard JL, McCormick S, Yang M. 2004. Progression throughmeiosis I and meiosis II in Arabidopsis anthers is regulated by anA-type cyclin predominately expressed in prophase I. Plant Physiol136:4127–4135.
263GENOMIC INSTABILITY IN ARABIDOPSIS THALIANA
Related Documents











