Polymorphisms and resistance mutations in the protease and reverse transcriptase genes of HIV-1 F subtype Romanian strains § Simona Paraschiv, Dan Otelea * , Magdalena Dinu, Daniela Maxim, Mihaela Tinischi Molecular Diagnostics Laboratory, ‘Prof. Dr. Matei Bals’ Institute for Infectious Diseases, Str. Calistrat Grozovici, nr.1, sector 2, 021105 Bucharest, Romania Received 15 July 2005; received in revised form 2 November 2005; accepted 15 November 2005 Corresponding Editor: Salim S. Abdool Karim, Durban, South Africa International Journal of Infectious Diseases (2007) 11, 123—128 http://intl.elsevierhealth.com/journals/ijid KEYWORDS HIV-1 F subtype; Resistance mutations; Natural polymorphisms Summary Objective: To evaluate the prevalence of resistance mutations in the genome of HIV-1 F subtype strains isolated from Romanian antiretroviral (ARV) treatment-naı ¨ve patients and to assess the phylogenetic relatedness of these strains with other HIV-1 strains. Methods: Twenty-nine HIV-1 strains isolated from treatment-naı ¨ve adolescents (n = 15) and adults (n = 14) were included in this study. Resistance genotyping was performed by using Big Dye Terminator chemistry provided by the ViroSeq Genotyping System. The sequences of the protease and reverse transcriptase genes were aligned (ClustalW) and a phylogenetic tree was built (MEGA 3 software). For subtyping purposes, all the nucleotide sequences were submitted to the Stanford database. Results: All the studied strains were found to harbor accessory mutations in the protease gene. The most frequent mutation wasM36I (29 of 29 strains), followed by L63T, K20R, and L10V. The number of polymorphisms associated with protease inhibitor resistance was different for the two age groups. Intraphylogenetic divergence was greater for adults than for adolescents infected in childhood. All the strains were found to belong to the F1 subtype. The phylogenetic analysis revealed that Romanian strains clustered together, but distinctly from F1 HIV-1 strains isolated in other parts of the world (Brazil, Finland, and Belgium). Conclusion: Protease secondary mutations are present with high frequency in the HIV-1 F subtype strains isolated from Romanian ARV treatment-naı ¨ve patients, but no major resistance mutations were found. # 2006 International Society for Infectious Diseases. Published by Elsevier Ltd. All rights reserved. § This study was presented in part as a poster at the 14th European Congress of Clinical Microbiology and Infectious Diseases, Prague, May 2004. * Corresponding author. Tel.: +40 213186100x3084; fax: +40 213186090. E-mail address: [email protected] (D. Otelea). 1201-9712/$32.00 # 2006 International Society for Infectious Diseases. Published by Elsevier Ltd. All rights reserved. doi:10.1016/j.ijid.2005.11.006

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

International Journal of Infectious Diseases (2007) 11, 123—128
Polymorphisms and resistance mutations in theprotease and reverse transcriptase genes of HIV-1 Fsubtype Romanian strains§
Simona Paraschiv, Dan Otelea *, Magdalena Dinu,Daniela Maxim, Mihaela Tinischi
http://intl.elsevierhealth.com/journals/ijid
Molecular Diagnostics Laboratory, ‘Prof. Dr. Matei Bals’ Institute for Infectious Diseases,Str. Calistrat Grozovici, nr.1, sector 2, 021105 Bucharest, Romania
Received 15 July 2005; received in revised form 2 November 2005; accepted 15 November 2005Corresponding Editor: Salim S. Abdool Karim, Durban, South Africa
KEYWORDSHIV-1 F subtype;Resistance mutations;Natural polymorphisms
Summary
Objective: To evaluate the prevalence of resistance mutations in the genome of HIV-1 F subtypestrains isolated from Romanian antiretroviral (ARV) treatment-naı̈ve patients and to assess thephylogenetic relatedness of these strains with other HIV-1 strains.Methods: Twenty-nine HIV-1 strains isolated from treatment-naı̈ve adolescents (n = 15) andadults (n = 14) were included in this study. Resistance genotyping was performed by using BigDye Terminator chemistry provided by the ViroSeq Genotyping System. The sequences of theprotease and reverse transcriptase genes were aligned (ClustalW) and a phylogenetic tree wasbuilt (MEGA 3 software). For subtyping purposes, all the nucleotide sequences were submitted tothe Stanford database.Results: All the studied strains were found to harbor accessory mutations in the protease gene.The most frequent mutation was M36I (29 of 29 strains), followed by L63T, K20R, and L10V. Thenumber of polymorphisms associated with protease inhibitor resistance was different for the twoage groups. Intraphylogenetic divergence was greater for adults than for adolescents infected inchildhood. All the strains were found to belong to the F1 subtype. The phylogenetic analysisrevealed that Romanian strains clustered together, but distinctly from F1 HIV-1 strains isolated inother parts of the world (Brazil, Finland, and Belgium).Conclusion: Protease secondary mutations are present with high frequency in the HIV-1 F subtypestrains isolated from Romanian ARV treatment-naı̈ve patients, but no major resistance mutationswere found.# 2006 International Society for Infectious Diseases. Published by Elsevier Ltd. All rights reserved.
§ This study was presented in part as a poster at the 14th European Congress of Clinical Microbiology and Infectious Diseases, Prague, May 2004.* Corresponding author. Tel.: +40 213186100x3084; fax: +40 213186090.E-mail address: [email protected] (D. Otelea).
1201-9712/$32.00 # 2006 International Society for Infectious Diseases. Published by Elsevier Ltd. All rights reserved.doi:10.1016/j.ijid.2005.11.006

124 S. Paraschiv et al.
Introduction
With the advent of highly active antiretroviral therapy(HAART) the life expectancy of HIV infected patients has beensignificantly prolonged. However, it is now clear that muta-tions accumulate in treated patients, limiting the efficacy ofthe therapy and requiring adjustments of the antiretroviralregimen. Resistance genotyping has become standard of carein HIV infection management.1 Resistance mutations arereported with increasing frequencies in treatment-naı̈ve indi-viduals.2—5 The prevalence of major resistance mutations forany antiretroviral drug observed in recent seroconverters canrange from 2% to 26% in North America, 23% to 26% in Spain, 4%to 17% in France, and 5% to 11% in Switzerland.3 Informationhas accumulated so far mainly on type M subtype F strainswhicharepredominant in theWesternhemisphere.An increas-ing number of communications have shown that resistancemutations can occur spontaneously in the genome of virusesbelonging to subtypes other than B isolated from untreatedpatients.6—8 Most of these mutations are polymorphisms andare considered secondary mutations that by themselves play alimited, if any, role in resistance.However, their pre-existencemight favor a more rapid evolution towards resistance whenadditional mutations are selected under therapy.
Since 1992, the HIV-1 strains isolated from patients world-wide have been classified into three distinct genetic groups: M(major) group, O (outlier) group, and N (non-M, non-O) group.Within the M group nine subtypes have been identified so far(A, B, C, D, F, G, H, J, and K) and intersubtype circulatingrecombinant forms (CRFs) and unique recombinant forms(URFs) have been described. Since most of the therapeuticexperience has been derived from HIV-1 subtype B infections,the management of patients infected with HIV-1 strains ofsubtypes other than B is mostly the result of extrapolation.
In this study we present data obtained from the analysis ofstrains isolated from Romanian treatment-naı̈ve adolescentsand adults. Previous anecdotal information has suggested thatsubtype F strains are rather common in Romania.9,10 Ourfindings have confirmed these observations (the vast majorityof the strains tested so far belong to the F subtype), althoughthe strains displayed several dissimilarities with subtype Fstrains originating from other geographical regions. A recentstudy11 has evaluated the subtype impact on polymorphismand resistance mutation development under ARV treatmentand it has concluded that the positions associated with drugresistance for theB subtypeare not significantlydifferent fromthose found in other HIV-1 subtypes; however, the number ofpolymorphisms was higher in non-B subtype HIV-1 strains.Preliminary data indicated that theHIV-1 strains fromRomaniaclustereddistinctlywithin theF subtype.12 Itwas thereforeourpurpose to assess the frequency of resistance mutations spon-taneously occurring in the genome of the Romanian subtype Fstrains and to compare the relatedness of several subtype Fstrains isolated from naı̈ve patients in Romania with otherstrains of subtype F and other subtypes.
Methods
Study population
We studied 29 HIV-1 strains isolated from Romanian patients,adolescents (n = 15) and adults (n = 14), originating from dif-
ferent regions of the country. All of the adolescents were thesame age (14—15 years old) and most of the available epide-miological data suggest that they were infected by contami-nated blood products of improper sterilization of needles usedfor parenteral treatment. The adults were all infected by theheterosexual route. The studiedpatientswereARV treatment-naı̈ve at the time when the blood samples were drawn.
Clinical specimens
Plasma samples were obtained from the 29 HIV-1 infectedpatients. For all the patients the viral loads were higher than10 000 copies/mL. Five mL of blood were collected in EDTAand the plasma was separated by centrifugation at 850 g for15 min. The samples were frozen at �80 8C for storage untilthe RNA extraction.
Sequencing
The ViroSeq HIV-1 genotyping system (Celera Diagnostics,Alameda, USA) was used according to the manufacturer’srecommendations, with the exception of the RNA extractionwhich was performed with the Abbott m1000
TM
automaticnucleic acid extraction instrument. Briefly, after the deter-mination of the viral load by LCx HIV RNA quantitative assay(Abbott Laboratories, Abbott Park, USA), the extracted RNAwas reverse transcribed and then amplified by PCR. Thethermal cycling was performed on a GeneAmp System 9700(Applied Biosystems, Foster City, USA) thermal cycler. The1.8 kb RT-PCR product was purified using MicroCon YM-100concentrators (spin columns). The purity and the amount ofthe DNA amplification product were evaluated by running analiquot on a 1% agarose electrophoresis gel. The ampliconwas diluted to a standard 20—40 ng/sequencing reaction.
Big Dye Terminator chemistry was used for DNA sequen-cing: seven sequencing reactions were set up for covering theentire HIV-1 protease (PR) gene and two-thirds of the reversetranscriptase (RT) gene.
Capillary electrophoresis of the samples was performed onan ABI Prism 3100-Avant Genetic Analyzer (Applied Biosys-tems). Prior to loading, the purified products of the sequen-cing reactions were resuspended in formamide, denaturatedat 95 8C/2 min and chilled on ice. The raw analysis of thesequences was made using Sequencing Analysis softwareversion 3.7 and then they were assembled with ViroSeq2.5 HIV-1 Genotyping System software. The correctness ofeach electropherogram interpretation was validated by theoperator and the sequences were saved in Fasta format. Theresistance report generated by ViroSeq provides a list ofnucleotide mutations detected in the PR (codons 1—99)and the RT gene (codons 1—335) known to determine resis-tance to antiretroviral drugs and infers a resistance profilefor each strain. The Fasta files were then used to interpretthe results by the HIV Seq Program (HIV-1 RT and proteasesearch engine of queries), available at the Stanford database(http://hivdb.stanford.edu). The same resistance mutationswere identified by both Viroseq 2.5 and Stanford software.
Phylogenetic and sequence analysis
The subtyping of all the Romanian strains was done by usingthe publicly available algorithm at the Stanford database
Polymorphisms and resistance mutations in HIV-1 F subtype 125
Table 1 Amino acid substitutions in the protease gene sequences of HIV-1 F subtype Romanian strains at positions associated withresistance to protease inhibitors
Codon position Subtype B amino acid Amino acid substitution No. of strains Percentage
10 L V 13 44.8I 2 6.9IL/VL 2 6.9
20 K R 21 72.4M 1 3.4
36 M I 29 100
63 L T 27 93.1AT 1 3.4
77 V I 1 3.4
82 V I 1 3.4
93 I L 4 13.8
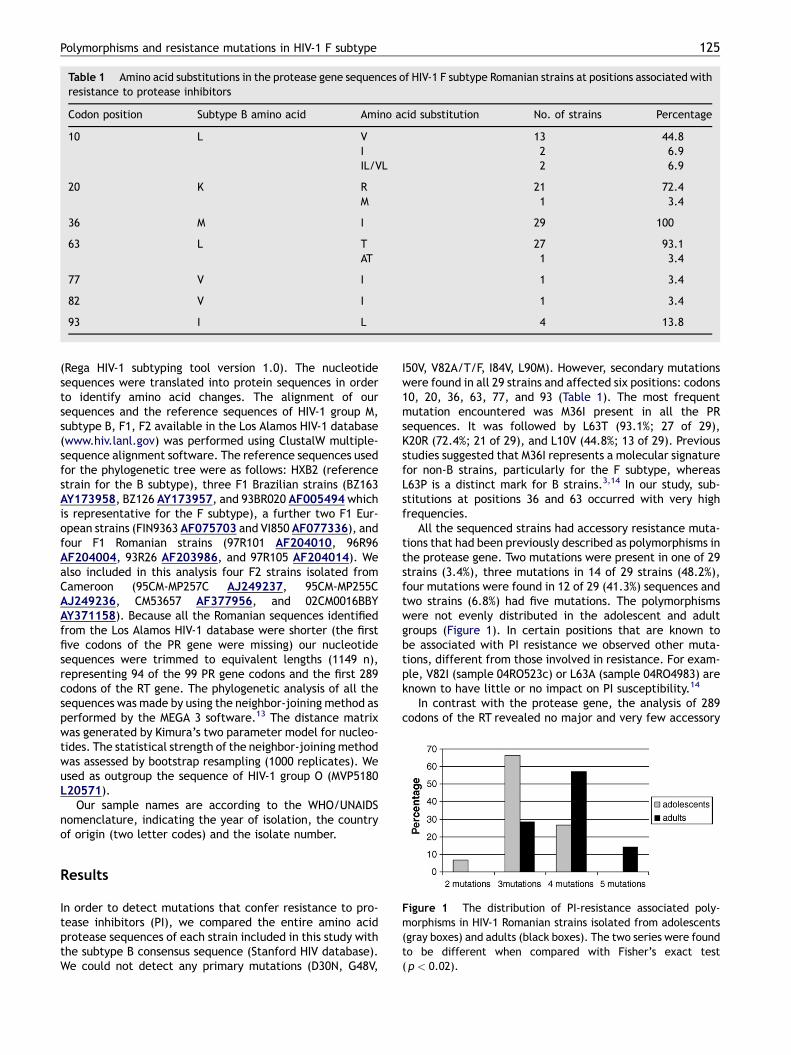
Figure 1 The distribution of PI-resistance associated poly-morphisms in HIV-1 Romanian strains isolated from adolescents(gray boxes) and adults (black boxes). The two series were foundto be different when compared with Fisher’s exact test(p < 0.02).
(Rega HIV-1 subtyping tool version 1.0). The nucleotidesequences were translated into protein sequences in orderto identify amino acid changes. The alignment of oursequences and the reference sequences of HIV-1 group M,subtype B, F1, F2 available in the Los Alamos HIV-1 database(www.hiv.lanl.gov) was performed using ClustalW multiple-sequence alignment software. The reference sequences usedfor the phylogenetic tree were as follows: HXB2 (referencestrain for the B subtype), three F1 Brazilian strains (BZ163AY173958, BZ126 AY173957, and 93BR020 AF005494whichis representative for the F subtype), a further two F1 Eur-opean strains (FIN9363 AF075703 and VI850 AF077336), andfour F1 Romanian strains (97R101 AF204010, 96R96AF204004, 93R26 AF203986, and 97R105 AF204014). Wealso included in this analysis four F2 strains isolated fromCameroon (95CM-MP257C AJ249237, 95CM-MP255CAJ249236, CM53657 AF377956, and 02CM0016BBYAY371158). Because all the Romanian sequences identifiedfrom the Los Alamos HIV-1 database were shorter (the firstfive codons of the PR gene were missing) our nucleotidesequences were trimmed to equivalent lengths (1149 n),representing 94 of the 99 PR gene codons and the first 289codons of the RT gene. The phylogenetic analysis of all thesequences was made by using the neighbor-joining method asperformed by the MEGA 3 software.13 The distance matrixwas generated by Kimura’s two parameter model for nucleo-tides. The statistical strength of the neighbor-joining methodwas assessed by bootstrap resampling (1000 replicates). Weused as outgroup the sequence of HIV-1 group O (MVP5180L20571).
Our sample names are according to the WHO/UNAIDSnomenclature, indicating the year of isolation, the countryof origin (two letter codes) and the isolate number.
Results
In order to detect mutations that confer resistance to pro-tease inhibitors (PI), we compared the entire amino acidprotease sequences of each strain included in this study withthe subtype B consensus sequence (Stanford HIV database).We could not detect any primary mutations (D30N, G48V,
I50V, V82A/T/F, I84V, L90M). However, secondary mutationswere found in all 29 strains and affected six positions: codons10, 20, 36, 63, 77, and 93 (Table 1). The most frequentmutation encountered was M36I present in all the PRsequences. It was followed by L63T (93.1%; 27 of 29),K20R (72.4%; 21 of 29), and L10V (44.8%; 13 of 29). Previousstudies suggested that M36I represents a molecular signaturefor non-B strains, particularly for the F subtype, whereasL63P is a distinct mark for B strains.3,14 In our study, sub-stitutions at positions 36 and 63 occurred with very highfrequencies.
All the sequenced strains had accessory resistance muta-tions that had been previously described as polymorphisms inthe protease gene. Two mutations were present in one of 29strains (3.4%), three mutations in 14 of 29 strains (48.2%),four mutations were found in 12 of 29 (41.3%) sequences andtwo strains (6.8%) had five mutations. The polymorphismswere not evenly distributed in the adolescent and adultgroups (Figure 1). In certain positions that are known tobe associated with PI resistance we observed other muta-tions, different from those involved in resistance. For exam-ple, V82I (sample 04RO523c) or L63A (sample 04RO4983) areknown to have little or no impact on PI susceptibility.14
In contrast with the protease gene, the analysis of 289codons of the RT revealed no major and very few accessory

126 S. Paraschiv et al.
mutations. The mutations associated with resistance tonucleoside reverse transcriptase inhibitors (NRTIs) andnon-nucleoside reverse transcriptase inhibitors (NNRTIs)have been well characterized and differ between thesetwo classes of RT inhibitors: resistance mutations to NRTIsare localized in ‘fingers’, ‘thumb’, and ‘palm’ domains of thereverse transcriptase while mutations responsible for resis-tance to NNRTIs are localized into the hydrophobic cavity(NNRTI binding site). The V179D mutation appeared in onlytwo of the samples (04RO2165 and 04RO2301) isolated fromadults. Low-level resistance to each NNRTI has been attrib-uted to this mutation, but its effect is considered insignif-icant in the absence of other mutations. In the RT gene weidentified some atypical mutations at key positions known tobe linked with drug resistance, namely M41R (03RO155),T69AT (03RO1436), Y318I (03RO155), and K101RK(04RO4420), but these mutations have not been reportedas important for resistance to NRTIs or NNRTIs.
Phylogenetic analysis
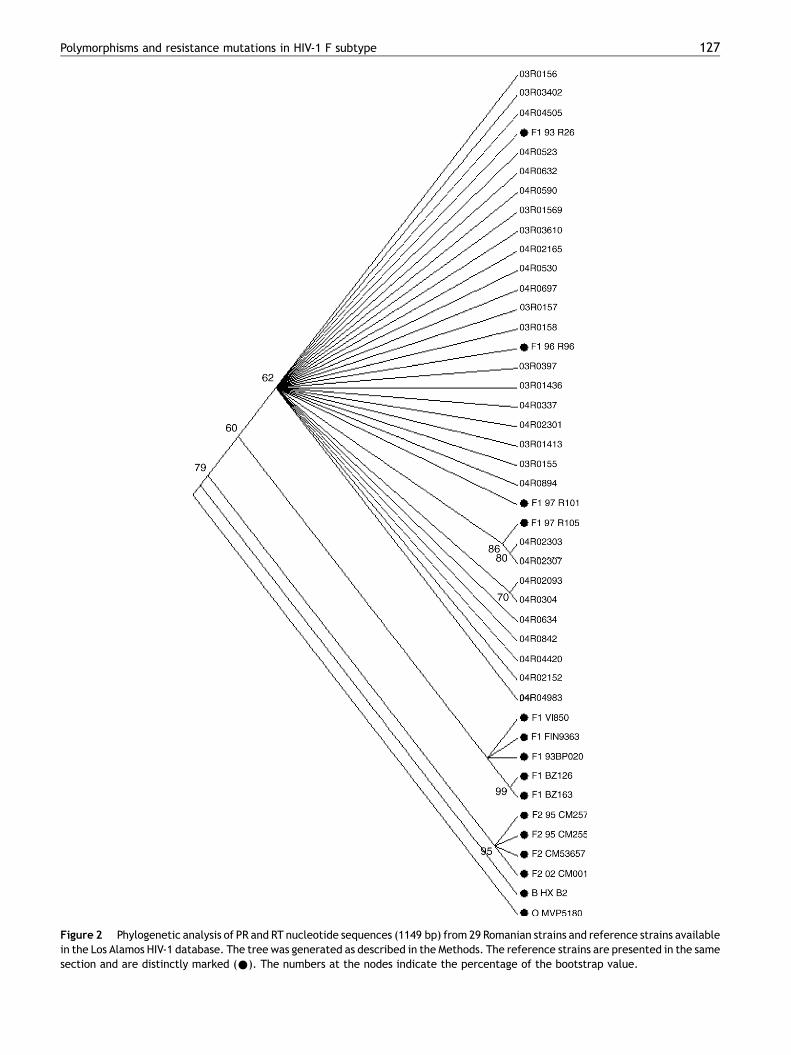
The subtype analysis of the PR and RT sequences indicatedthat all 29 HIV-1 strains belonged to the F1 subtype, with abootstrap support ranging between 76% and 100%. The 1149nucleotide sequences of the 29 Romanian strains represent-ing the PR gene and half of the RT gene were used to build aphylogenetic tree. Our strains were compared with referencestrains of different subtypes (B, F1, F2) and sequences of HIV-1 strains isolated between 1993 and 1997 from Romanianpatients, available in the Los Alamos HIV-1 sequence data-base. We used as outgroup the MVP5180 sequence (HIV-1group O). The result of the phylogenetic analysis is presentedin Figure 2. As expected, the algorithm could efficientlydiscriminate among different subtypes. The studied strainsclustered with the F1 reference strains and were distinctfrom the F2 strains. Moreover, all the Romanian strainsclustered together with the other Romanian strains identifiedfrom the Los Alamos HIV-1 sequence database, but weredistinct from F1 Brazilian strains or F1 European strains(Belgium and Finland), suggesting a different epidemiologi-cal context for the infection.
The phylogenetic analysis did not discriminate betweenthe HIV-1 strains isolated from the two age groups. However,the average sequence divergence was higher in the adultgroup (6.49%) in comparison with the adolescent group(5.11%).
It has been shown that although the pol gene is veryconserved, it can be used for subtyping purposes. Our dataconfirm this observation and, furthermore, show that thegene can discriminate between groups of strains belonging tothe same subtype (Romanian and Brazilian F strains clustereddistinctly).
Discussion
Recent studies have tried to assess the influence that differ-ent HIV-1 subtypes exert on the efficacy of ARV treatment andthe progression to AIDS.14—16 The results have often beencontradictory, but there is evidence that strains belonging togroup O are naturally resistant to NNRTIs and some G subtypestrains are less susceptible to PIs.17,18
Most of the RT and PR resistance data reported to dateoriginate from HIV-1 B subtype studies — this being thepredominant subtype in Western Europe and North America.The dominant subtype responsible for HIV infections in Roma-nian children and adults is the F subtype.9,19 It has also beenreported, with lower prevalence, in other countries likeBrazil, Argentina, Cameroon, France, and Belgium. Phyloge-netic analysis of the HIV-1 strains isolated from these differ-ent countries suggest that separate epidemiological factorscontribute to the particular distribution of HIV-1 F subtype indistinct regions of the world.2,12,20
In Romania, epidemiological data suggest that childrenhave been infected by contaminated blood products and/orimproper sterilization of needles used for parenteral treat-ment (vaccination). Most of the viral transmission in adults isby the heterosexual route. To better understand the epide-miological context of the HIV infection in Romania, weperformed a phylogenetic analysis of strains isolated fromtwo distinct age groups. Phylogenetic trees are powerfultools that evaluate the relatedness of field strains. Ourresults show that all the HIV strains isolated from adultsand adolescents infected in childhood clustered togetherinto a distinct group. For comparison purposes, PR and RTsequences of F1 and F2 subtypes collected from SouthAmerica and Europe were included in our analysis. Theresistance-associated mutations in the PR gene were differ-ently distributed in the adolescent and adult groups: moremutations were detected in strains from adult patients thanin those from adolescents (Figure 1). Differences betweenstrains isolated from these two age groups were alsoobserved when intrasubtype F sequence divergence wascalculated: the divergence was higher in strains originatingfrom adults. These data correlate well with the distinctinfective events in teenagers vs. adults: the adolescentcohort, all between 14 and 15 years of age have presumablybeen infected within a short span of time by the parenteralroute whereas the adults have been infected over a longerperiod of time duringwhich the virus has had the opportunityto evolve through bottleneck events during interhumantransmission.
None of the subjects included in this study had receivedany antiretroviral treatment. In order to investigate theimpact of HIV-1 F subtype on resistance before the onsetof ARV treatment, we searched mutations known to beassociated with resistance to protease and reverse transcrip-tase inhibitors. Overall, the protease gene appears to be lessconserved than the RT gene. No primary resistance mutationswere found, but every HIV strain analyzed was harboring atleast two accessory mutations. The M36I mutation in theprotease gene (associated with resistance to ritonavir andnelfinavir in B subtype viruses) was found in all the Romanianstrains. This finding supports the suggestion that methioninein this position may represent a polymorphism specific to theB subtype. Substitutions were also observed in other proteasegene positions, some of them with rather high frequency: 10,20, 63, 77, and 93.
Accessory mutations may not result in a significantdecrease of sensitivity to ARV drugs, but have been reportedto be associated with increased viral fitness in viruses withprimary mutations. It can therefore be speculated that theselection of primary mutations under selective pressurecould precipitate the development of highly resistant HIV
Polymorphisms and resistance mutations in HIV-1 F subtype 127
Figure 2 Phylogenetic analysis of PR and RT nucleotide sequences (1149 bp) from 29 Romanian strains and reference strains availablein the Los Alamos HIV-1 database. The tree was generated as described in the Methods. The reference strains are presented in the samesection and are distinctly marked (*). The numbers at the nodes indicate the percentage of the bootstrap value.

128 S. Paraschiv et al.
variants during ARV treatment in viruses where one or moresecondary mutations are already present.
In this study direct sequencing of PCR products was per-formed. Therefore, each sequence represents the consensusgenotype; it is possible that additional mutations may exist inthese HIV isolates, but PCR cloning followed by sequencingwould be necessary to detect them.
Kantor et al.11 have recently evaluated the frequency ofpolymorphisms and resistance mutations in protease andreverse transcriptase genes in non-B subtypes. Our findingsshow a higher frequency of protease polymorphisms that aresecondary resistance mutations as well. We interpret thisobservation as due to the higher homogeneity of the strainswe examined (only 18% of the subtype F strains analyzed byKantor et al. were isolated from Romanian patients).Furthermore, this higher frequency correlates with theincreased frequency of the same polymorphisms as detectedin viruses isolated from Romanian treated patients (data notshown).
In conclusion, no major mutations associated with resis-tance to NRTIs, NNRTIs, and PIs were detected in subtype Fstrains isolated from Romanian patients, but many accessorysubstitutions were found as natural variants, mostly in posi-tions associated with resistance to protease inhibitors. Theirincidence is partly different from that reported for othersubtype F HIV-1 strains. It is not yet clear if the high fre-quency of accessory protease mutations may contribute to amore rapid crossing of the genetic resistance barrier. Sequen-tial phenotypic analyses and long-term follow-up of patientsunder antiretroviral treatment will be necessary to evaluatethe clinical significance of these minor PR mutations.
Acknowledgements
The authors wish to thank Adriana Hristea, Gratiela Tardei,Sorin Petrea, and Mihai Rosca for technical support.
Conflict of interest: No conflict of interest to declare.
References
1. Tural C, Ruiz L, Holtzer C, Schapiro J, Viciana P, Gonzales J, et al.Clinical utility of HIV-1 genotyping and expert advice: the Havanatrial. AIDS 2002;16:209—18.
2. Pieniazek D, Rayfield M, Hu D, Nkengasong J, Wiktor S, DowningR, et al. Protease sequences from HIV-1 group M subtypes A—Hreveal distinct amino acid mutation patterns associated withprotease resistance in protease inhibitor-naı̈ve individualsworldwide. AIDS 2000;14:1489—95.
3. Dumans AT, Soares MA, Pieniazek D, Kalish ML, De Vroey V,Hertogs K, et al. Prevalence of protease and reverse transcrip-tase drug resistance mutations over time in drug-naı̈ve humanimmunodeficiency virus type 1-positive individuals in Rio deJaneiro, Brazil. Antimicrob Agents Chemother 2002;46:3075—9.
4. Vergne L, Peeters M, Mpoudi-Ngole E, Bourgeois A, Liegeois F,Toure-Kane C, et al. Genetic diversity of protease and reversetranscriptase sequences in non-subtype B human immunodefi-ciency virus type 1 strains: evidence of many minor drug resis-tance mutations in treatment-naive patients. J Clin Microbiol2000;38:3919—25.
5. Little SJ, Holte S, Routy JP, Daar ES, Markowitz M, Collier AC,et al. Antiretroviral-drug resistance among patients recentlyinfected with HIV. N Engl J Med 2002;347:385—94.
6. Turner D, Brenner B, Moisi D, Detorio M, Cesaire R, Kurimura T,et al. Nucleotide and amino acid polymorphism at drug resis-tance sites in non-B subtype variants of human immunodefi-ciency virus type 1. Antimicrob Agents Chemother 2004;48:2993—8.
7. Pires IL, Soares MA, Speranza FA, Ishii SK, Vieira M, Gouvea M,et al. Prevalence of human immunodeficiency virus drug resis-tance mutations and subtypes in drug-naive, infected individualsin the army health service of Rio de Janeiro, Brazil. J ClinMicrobiol 2004;42:426—30.
8. Cornelissen M, van der Burg R, Zorgdrager F, Lukashov V, Gouds-mit J. pol gene diversity of five human immunodeficiency virustype 1 subtypes: evidence for naturally occurring mutations thatcontribute to drug resistance, limited recombination patterns,and common ancestry for subtypes B and D. J Virol 1997;71:6348—58.
9. Apetrei C, Necula A, Holm-Hansen C, Lousert-Ajaka I, Pandrea I,Cozmei C, et al. HIV-1 diversity in Romania. AIDS 1998;12:1079—85.
10. Apetrei C, Descamps D, Collin G, Loussert-Ajaka I, Damond F,Duca M, et al. Human immunodeficiency virus type 1 subtype Freverse transcriptase sequence and drug susceptibility. J Virol1998;72:3534—8.
11. Kantor R, Katzenstein DA, Efron B, Carvalho AP, Wynhoven B,Cane P, et al. Impact of HIV-1 subtype and antiretroviral therapyon protease and reverse transcriptase genotype: results of aglobal collaboration. PLoS Med 2005;2:e112.
12. Otelea D, Oprisan G, Dinu M, Manaila S, Banica D, Tinischi M,et al. Phylogenetic analysis of subtype F HIV-1 strains isolated inRomania. Clin Microbiol Infect 2004;10(Suppl. 3):160. (poster667 abstract).
13. Kumar S, Tamura K, Nei M. MEGA3: Integrated software forMolecular Evolutionary Genetics Analysis and sequence align-ment. Brief Bioinform 2004;5:150—63.
14. Tanuri A, Vicente ACP, Otsuki K, Ramos CA, Ferreira Jr OC,Schechter M, et al. Genetic variation and susceptibilities toprotease inhibitors among subtype B and F isolates in Brazil.Antimicrob Agents Chemother 1999;43:253—8.
15. Pillay D, Walker AS, Gibb DM, de Rossi A, Kaye S, Ait-Khaled M,et al. Impact of immunodeficiency virus type 1 subtypes onvirologic response and emergence of drug resistance amongchildren in the Paediatric European Network for Treatment ofAIDS (PENTA) 5 trial. J Infect Dis 2002;186:617—25.
16. Spira S, Wainberg MA, Loemba H, Turner D, Brenner BG. Impactof clade diversity on HIV-1 virulence, antiretroviral drug sensi-tivity and drug resistance. J Antimicrob Chemother 2003;51:229—40.
17. Descamps D, Collin G, Letourneur F, Apetrei C, Damond F,Loussert-Ajaka I, et al. Susceptibility of human immunodefi-ciency virus type 1 group O isolates to antiretroviral agents:in vitro phenotypic and genotypic analysis. J Virol 1997;71:8893—8.
18. Descamps D, Apetrei C, Collin G, Damond F, Simion F, Brun-Vezinet F. Naturally occurring decreased susceptibility of HIV-1subtype G to protease inhibitors. AIDS 1998;12:1109—11.
19. Otelea D, Manaila S, Dinu M, Banica D, Tinischi M, Maxim D.Prevalence of resistance mutations in subtype F strains isolatedfrom Romanian naı̈ve children. Clin Microbiol Infect2004;10(Suppl. 3):160. (poster 666 abstract).
20. Brindeiro R, Vanderborght B, Caride E, Correa L, Oravec RM,Berro O, et al. Sequence diversity of reverse transcriptase ofhuman immunodeficiency virus type 1 from untreated Brazilianindividuals. Antimicrob Agents Chemother 1999;43:1674—80.
Related Documents











