Chem.-Biol. Interactions, 29 (1980) 277--289 © Elsevier/North-Holland Scientific Publishers Ltd. 277 POLYCHLORINATED BIPHENYL ISOMERS AND CONGENERS INDUCERS OF BOTH 3-METHYLCHOLANTHRENE- AND PHENOBARBITONE-TYPE MICROSOMAL ENZYME ACTIVITY AS A. PARKINSON, R. COCKERLINE and S. SAFE* Guelph-Waterloo Centre for Graduate Work in Chemistry, Department of Chemistry, University of Guelph, Guelph, Ontario NIG 2W1 (Canada) (Received August 30th, 1979) (Revision received October 24th, 1979) (Accepted November 2nd, 1979) SUMMARY Highly purified synthetic polychlorinated biphenyls substituted in the meta and para positions of both phenyl rings and at one ortho position were administered to male Wistar rats and the effects of these compounds on the microsomal drug-metabolising enzymes were evaluated. The in vivo effects of these compounds were determined by measuring the microsomal benzo[a]- pyrene hydroxylase, dimethylaminoantipyrine N~lemethylase and NADPH- cytochrome c reductase enzyme activities, the cytochrome b5 content and the relative peak intensities and spectral shifts of the reduced microsomal cytochrome P-450 : CO and ethylisocyanide binding difference spectra. The results were compared to the effects of administering phenobarbitone (PB), 3-methylcholanthrene (MC), 2,2',4,4'-tetrachlorobiphenyl (TCBP-II) (a PB- type inducer), 3,3',4,4'-tetrachlorobiphenyl (TCBP-I)(an MC-type inducer), PB plus MC (coadministered) and TCBP-II + TCBP-I (coadministered) to the test animals. At dosage levels of 30 and 150 ~mol • kg-~, pretreatment with 2,3,3',4,4'-pentachlorobiphenyl (PCBP-II), 2,3',4,4',5-pentachloro- biphenyl (PCBP-I), 2,3,3',4,4',5-hexachlorobiphenyl (HCBP-II) and 2,3,3',- 4,4',5-hexachlorobiphenyl (HCBP-III) gave hepatic microsomes with enzymic and spectral properties consistent with a mixed pattern of induction. These polychlorinated biphenyl (PCB) isomers and congeners have been identified • To whom reprint requests should be sent. Abbreviations: B [a ] P, benzo[a ] pyrene; CO, carbon monoxide; DMAP, 4-dimethylamino- antipyrine; EC, electron capture; EIC, ethylisocyanide; GLC, gas-liquid chromatography; HCBP-I, 2,3',4,4',5,5'-hexachlorobiphenyl; HCBP-H, 2,3,3°,4,4',5'-hexachlorobiphenyl; HCBP-III, 2,3,3',4,4',5-hexachlorobiphenyl; HCBP-IV, 2,2',4,4',5,5'-hexachlorobiphenyl; HpCBP, 2,3,3',4,4',5,5'-heptachlorobiphenyl; MC, 3omethylcholanthrene; NMR, nuclear magnetic resonance; PB, phenobarbitone; PBB, polybrominated biphenyl; PCB, poly- chlorinated biphenyl; PCBP-I, 2,3',4,4t,5-pentachlorobiphenyl; PCBP-II, 2,3,3',4,4'- pentachlorobiphenyl; TCBP-I, 3,3',4,4'-tetrachlorobiphenyl; TCBP-II, 2,2',4,4t-tetra - chlorobiphenyl; TLC, thin-layer chromatography.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Chem.-Biol. Interactions, 29 (1980) 277--289 © Elsevier/North-Holland Scientific Publishers Ltd.
277
POLYCHLORINATED BIPHENYL ISOMERS AND CONGENERS INDUCERS OF BOTH 3-METHYLCHOLANTHRENE- AND PHENOBARBITONE-TYPE MICROSOMAL ENZYME ACTIVITY
AS
A. PARKINSON, R. COCKERLINE and S. SAFE*
Guelph-Waterloo Centre for Graduate Work in Chemistry, Department of Chemistry, University of Guelph, Guelph, Ontario NIG 2W1 (Canada)
(Received August 30th, 1979) (Revision received October 24th, 1979) (Accepted November 2nd, 1979)
SUMMARY
Highly purified synthetic polychlorinated biphenyls substituted in the meta and para positions of both phenyl rings and at one ortho position were administered to male Wistar rats and the effects of these compounds on the microsomal drug-metabolising enzymes were evaluated. The in vivo effects of these compounds were determined by measuring the microsomal benzo[a]- pyrene hydroxylase, dimethylaminoantipyrine N~lemethylase and NADPH- cytochrome c reductase enzyme activities, the cytochrome b5 content and the relative peak intensities and spectral shifts of the reduced microsomal cytochrome P-450 : CO and ethylisocyanide binding difference spectra. The results were compared to the effects of administering phenobarbitone (PB), 3-methylcholanthrene (MC), 2,2',4,4'-tetrachlorobiphenyl (TCBP-II) (a PB- type inducer), 3,3',4,4'-tetrachlorobiphenyl (TCBP-I)(an MC-type inducer), PB plus MC (coadministered) and TCBP-II + TCBP-I (coadministered) to the test animals. At dosage levels of 30 and 150 ~mol • kg -~, pretreatment with 2,3,3',4,4'-pentachlorobiphenyl (PCBP-II), 2,3',4,4',5-pentachloro- biphenyl (PCBP-I), 2,3,3',4,4',5-hexachlorobiphenyl (HCBP-II) and 2,3,3',- 4,4',5-hexachlorobiphenyl (HCBP-III) gave hepatic microsomes with enzymic and spectral properties consistent with a mixed pattern of induction. These polychlorinated biphenyl (PCB) isomers and congeners have been identified
• To whom reprint requests should be sent. Abbreviations: B [a ] P, benzo[a ] pyrene; CO, carbon monoxide; DMAP, 4-dimethylamino- antipyrine; EC, electron capture; EIC, ethylisocyanide; GLC, gas-liquid chromatography; HCBP-I, 2,3',4,4',5,5'-hexachlorobiphenyl; HCBP-H, 2,3,3°,4,4',5'-hexachlorobiphenyl; HCBP-III, 2,3,3',4,4',5-hexachlorobiphenyl; HCBP-IV, 2,2',4,4',5,5'-hexachlorobiphenyl; HpCBP, 2,3,3',4,4',5,5'-heptachlorobiphenyl; MC, 3omethylcholanthrene; NMR, nuclear magnetic resonance; PB, phenobarbitone; PBB, polybrominated biphenyl; PCB, poly- chlorinated biphenyl; PCBP-I, 2,3',4,4t,5-pentachlorobiphenyl; PCBP-II, 2,3,3',4,4'- pentachlorobiphenyl; TCBP-I, 3,3',4,4'-tetrachlorobiphenyl; TCBP-II, 2,2',4,4t-tetra - chlorobiphenyl; TLC, thin-layer chromatography.

278
as either major or minor components of the commercial PCB mixtures and must contr ibute to their activity as MC-type inducers. The only PCB isomer in this series which was not a mixed type inducer was 2,3',4,4',5,5W-hexa - chlorobiphenyl (HCBP-I) which appeared to be a PB-type inducer. This con- trasted to the mixed-type activity observed for 2,3',4,4',5,5'-hexabromobi- phenyl which was isolated from a commercial polybrominated biphenyl (PBB) mixture.
INTRODUCTION
Chemicals which induce the activity of hepatic microsomal monooxygenase enzymes have been divided into two main categories [1] . One category, typi- fied by PB, induces the cytochrome P-450-dependent monooxygenase activi- ties and the other class, typif ied by MC, induces the cytochrome P-448 (or P1-450)~lependent monooxygenase enzyme activities. The division into these two classes of inducers is based on a variety of qualitative differences in the properties of the associated monooxygenase enzymes [2--5] . PB pretreat- ment stimulates an increase in the hepatic concentrations of cy tochrome P-450 haemoproteins similar to those which are constitutive in hepatic micro- somes from non-induced animals. The reduced forms of both PB-induced and non-induced cy tochrome P-450 haemoproteins bind with carbon mon- oxide (CO) to form a complex which exhibits an absorption maximum at 450 nm. The reduced haemoproteins also form a complex with ethyliso- cyanide (EIC) which exhibits absorption maxima at 428 and 455 nm with a 455 : 428 nm peak height ratio of about 1 : 2. In contrast, pretreatment, with MC stimulates the de novo synthesis of microsomal haemoproteins which are electrophoreticaily distinct from PB-induced or non-induced micro- somal haemoproteins. Moreover, the CO- or EIC-difference spectra with MC- induced microsomes absorb maximally at 448 nm or at 429.4 and 452 nm with a 452 : 429.4 nm peak height ratio of about 2 : 1.
The differences in PB- and MC-induced microsomai monooxygenase enzymes can also be observed in the substrate and oxidation site specificity of the two enzyme systems [6] . For example, MC pretreatment induces benzo[a ]pyrene (B[a]P) hydroxylase activity by more than an order of magnitude over controls but fails to stimulate the activity of 4-dimethyl- aminoantipyrine (DMAP) N<lemethylase. PB pretreatment results in a 2--5-fold enhancement of both activities. In addition PB, but not MC, induces the activity of NADPH-cytochrome P-450 reductase which mediates the transfer of electrons from NADPH to cytochrome P-450.
Polychlorinated biphenyls (PCBs) are industrial chemicals which can induce the hepatic drug-metabolising enzymes [7--9] . The spectral, enzymic and electrophoretic properties of the microsomal monooxygenases induced by commercial mixtures of PCBs, such as Aroclor 1254, are consistent with the pat tern of mixed induction produced by the simultaneous administration of PB and MC [10--13] . Since commercial PCBs are complex mixtures of chlori-

279
nated biphenyl isomers and congeners, their activity is a function of the pro- perties of the individual components of the mixture. Recently, purified, syn- thetic PCB isomers and congeners have been categorised on a structural basis into PB-type and MC-type inducers [14--16]. These studies with individual PCBs as hepatic microsomal enzyme inducers supported the following structure-activity relationships:
(1) Biphenyl chlorinated in the para and ortho positions of both phenyl rings (4,4' and 2,2') is a PB-type inducer of cytochrome P-450 and any addi- tional chloro substituents do not affect the type of induction but could alter the potency of the inducer.
(2) Biphenyl chlorinated in only the para and rneta positions of both phenyl rings (4,4' and 3,3') is an MC-type inducer of cytochrome/)-448 and any additional meta chloro substituents (5,5') do not change the type of activity. However, additional ortho substituents abolish the activity.
(3) Most of the lower chlorinated biphenyls and PCBs substituted in only one phenyl ring do not induce microsomal enzyme activity.
It has recently been reported that 2,3',4,4',5,5'-hexabromobiphenyl [17] isolated from a commercial polybrominated biphenyl mixture, purified hexa- chlorobenzene [18,19] and synthetic 2,3,3',4,4'5,5'-heptachlorobiphenyl (HpCBP) [20] were mixed-type inducers of hepatic microsomal monooxy- genase enzyme activity. This study reports the effects of all possible PCB congeners which are substituted at the rneta and para positions of both phenyl rings and at one ortho position as hepatic microsomal enzyme in- ducers in the rat (see Fig. 1).
(k
C7 ~
TCBP I
(k
C~ c~
TCBP I I
Ct C~ cL
c ~ 7 L CL
PCBP I
Ct
CK: PCBP II
CL c~c~
c ~ 7 t Ct
HCBP I
Ct C 11,
C+. HCBP I I
Ct
Cy ~
c~K: Ct
HCBP III
Fig. 1. PCB i somers and congeners used in this s tudy .
Ct
c~7 ~
c~ ~ct CL
HCBP IV
Ct
c~TL
CL HpCBP

280
METHODS
Synthesis and purification of PCB isomers and congeners 2,4,5-Trichloroaniline, 2,3,4-trichloroaniline, 3,4<lichloroaniline, 1,2-
dichlorobenzene, 1,2,3-trichlorobenzene and 1,2,3,4-tetrachlorobenzene were purchased from Aldrich Chemical Co. and isoamyl nitrite from Eastman Organic Chemicals. PCBP-I, PCBP-II, HCBP-I, HCBP-II and HCBP-III were synthesized by the Cadogan coupling of a chlorinated aniline (10--15 mmol) with excess chlorinated benzene (200--300 mmol) as indicated in Table I [21]. The isoamyl nitrite (20--25 mmol) was added to the stirred solution at 120--130°C for 10 h. After removal of the excess chlorinated benzene, the crude product was adsorbed onto neutral alumina, added to a column of Florisil with a layer of alumina on top and the chlorinated biphenyls eluted with petroleum spirit (b.p., 30°--60°C). HCBP-III was purified by thin-layer chromatography (TLC) on silica gel HF2s4 (BDH Chemicals) using petroleum spirit (b.p., 30°-60°C) as the eluant and crystallized from methanol. The coupling reactions which gave PCBP-I, PCBP-II, HCBP-I and HCBP-II each contained an additional major PCB product; namely, 2,2',3'4,5-pentachloro- biphenyl, 2,2',3,3'4-pentachlorobiphenyl, 2,2',3,4,4',5'-hexachlorobiphenyl and 2,2',3,3',4,4'-hexachlorobiphenyl, respectively, which were separated from the desired major products by repeated TLC. The purified isomers and congeners were crystallized from methanol and their purities determined by gas-liquid chromatography (GLC) using a Hewlett Packard model 5710 chro- matograph equipped with a 63Ni electron capture (EC) detector and an 0.6 cm X 1.2 M glass column packed with 3% OV 101 on Ultrabonded Carbowax 20 M, 80--100 mesh (RFR Corp.). The structures of the desired products were confirmed by their 220 MHz nuclear magnetic resonance (NMR) and mass spectra (determined on a VG Micromass 7070 mass spectro- meter) and the results are summarized in Table I. TCBP-II, TCBP-I and 2,2',4,4',5,5'-hexachlorobiphenyl (HCBP-IV) were synthesized from the corresponding chlorinated benzidines as described [22]. The PCB congeners were purified by alumina/Florisil column chromatography, silica gel TLC and crystallization from methanol as described above and their purity was >99% as determined by EC-GLC.
Biochemicals Cytochrome c (horse heart, type III), NADP ÷, NADPH, D-glucose-6-
phosphate, D-glucose-6-phosphate dehydrogenase (Baker's yeast), MC, B[a]P and EIC were purchased from Sigma Chemical Co.; DMAP from Aldrich Chemical Co; CO (research purity) from Matheson, and sodium phenobarbitone from the Ontario Veterinary College, Guelph. [aH]B[a]P was obtained from New England Nuclear Co. and purified by TLC using hexane as eluant.
Animals treatment and isolation of microsomes One-month-old male Wistar rats, average weight 100--120 g, were housed
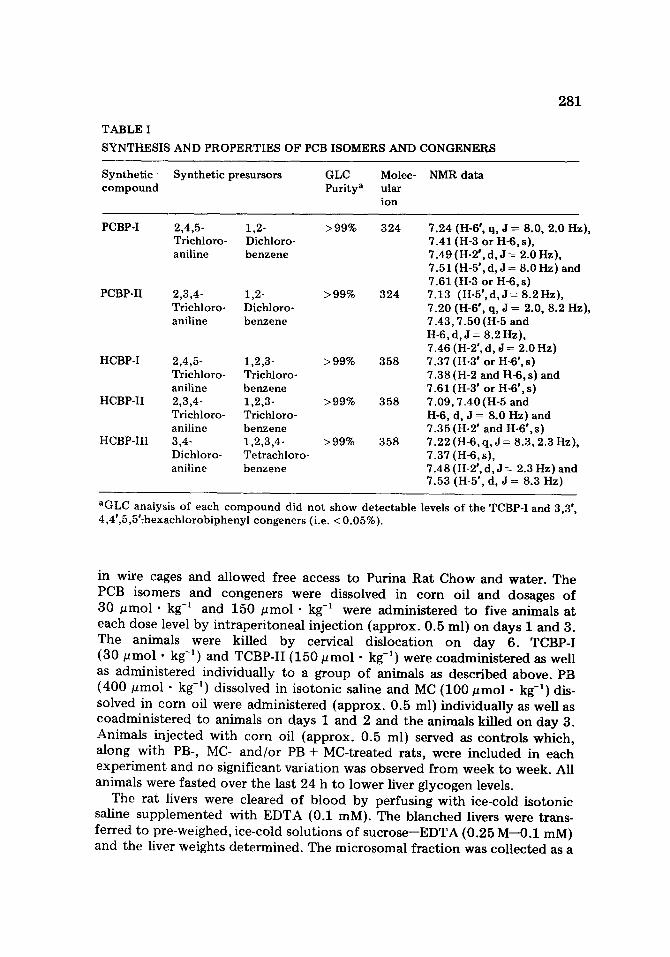
TABLE I
SYNTHESIS AND PROPERTIES OF PCB ISOMERS AND CONGENERS
281
Synthetic Synthetic presursors GLC Molec- NMR data compound Purity a ular
ion
PCBP-I 2,4,5- 1,2- >99% 324 Trichloro- Dichloro- aniline benzene
PCBP-II 2,3,4- 1,2- >99% 324 Trichloro- Dichloro- aniline benzene
HCBP-I 2,4,5- 1,2,3- >99% 358 Trichloro- Trichloro- aniline benzene
HCBP-II 2,3,4- 1,2,3- >99% 358 Trichloro- Trichloro- aniline benzene
HCBP-III 3,4- 1,2,3,4- >99% 358 Dichloro- Tetrachloro- aniline benzene
7.24 (H-6 °, q, J : 8.0, 2.0 Hz), 7.41 (H-3 or H-6, s), 7.49 (H-2',d,J = 2.0 Hz), 7.51 (H-5 °, d, J = 8.0 Hz) and 7.61 (H-3 or H-6, s) 7.13 (H-5°,d,J= 8.2Hz), 7.20 (H-6 °, q, J = 2.0, 8.2 Hz), 7.43, 7.50 (H-5 and H-6, d, J = 8.2 Hz), 7.46 (H-2', d, J = 2.0 Hz) 7.37 (H-3 ° or H-6', s) 7.38 (H-2 and H-6, s) and 7.61 (H-3 ° or H-6 °, s) 7.09, 7.40(H-5 and H-6, d, J -- 8.0 Hz) and 7.35 (H-2 ° and H-6',s) 7.22 (H-6, q,J = 8.3, 2.3 Hz), 7.37 (H-6, s), 7.48 (H-2 °, d, J = 2.3 Hz) and 7.53 (H-5', d, J -- 8.3 Hz)
aGLC analysis of each compound did not show detectable levels of the TCBP-I and 3,3 °, 4,4',5,5'-hexachlorobiphenyl congeners (i.e. <0.05%).
in wire cages and al lowed free access to Purina Rat Chow and water . The PCB isomers and congeners were dissolved in corn oil and dosages of 30 p m o l • kg- ' and 150 p m o l • kg -~ were adminis tered to five animals at each dose level by int raper i toneal inject ion (approx. 0.5 ml) on days 1 and 3. The animals were killed by cervical d is locat ion on day 6. TCBP-I (30 /~mol • kg - ' ) and TCBP-II (150 p m o l - kg -1) were coadminis te red as well as adminis tered individually to a group of animals as described above. PB (400 p m o l • kg -~) dissolved in isotonic saline and MC (100 p m o l - kg -1) dis- solved in corn oil were adminis tered (approx. 0.5 ml) individually as well as coadminis te red to animals on days 1 and 2 and the animals killed on day 3. Animals injected with corn oil (approx. 0.5 ml) served as cont ro ls which, along with PB-, MC- a n d / o r PB + MC-treated rats, were included in each expe r imen t and no significant variat ion was observed f rom week to week. All animals were fasted over the last 24 h to lower liver g lycogen levels.
The rat livers were cleared o f b lood by perfusing with ice-cold isotonic saline supp lemented with E D T A (0.1 mM). The b lanched livers were trans- ferred to pre-weighed, ice-cold solut ions o f suc rose - -EDTA (0.25 M--0,1 mM) and the liver weights de te rmined . The mic rosomal f rac t ion was col lected as a

282
100 000 × g pellet by further centrifugation of a 10 000 × g supernatant from the liver homogenate.
Assays In all assays the final concentration of mierosomal protein was
1.0 mg- ml-' as determined by the method of Lowry et al. [23]. The cyto- chrome P-450 content was determined by the method of Omura and Sato [24] , f rom the CO~lifference spectrum of dithionite-reduced microsomes using an extinction coefficient of 91 cm-' • raM-' between Area x and A490. The EIC~lifference spectrum was determined in a similar manner to the CO- difference spectrum except that EIC was added to the sample cuvette (final concentration 4.5 raM) instead of CO. A preliminary study revealed that contaminating haemoglobin decreased the 455 : 428 nm peak height ratio of the EIC~iifference spectrum (unpublished results). This was related to the ability of haemoglobin to bind such ligands as CO and EIC giving an absorb- ance peak at about 420 nm. There are two reasons why such haemoglobin interference cannot be compensated for by subtracting the difference spec- t rum between the haemoglobin/EIC complex and oxyhaemoglobin (deter- mined by adding EIC to the sample cuvette using oxidised microsomes) from the EIC<iifference spectrum. First, oxidised microsomes bind EIC and pro- duce a reverse type I spectrum with a peak at 433--434 nm [25,26]. This spectrum contributes to and complicates the spectrum produced by the haemoglobin/EIC complex. Second, methaemoglobin, which invariably com- prises a small fraction of the contaminating haemoglobin and which fails to bind either CO or EIC, is reduced to haemoglobin upon addition of dithi- onite. Therefore, the total interference from haemoglobin is unlikely to be represented by the haemoglobin/EIC complex minus oxyhaemoglobin spec- trum. In an effort to circumvent some of these problems, the amount of haemoglobin contaminating each microsomal preparation was estimated from the peak height at 420 nm of the difference spectrum between carbon- monoxyhaemoglobin and oxyhaemoglobin (determined by bubbling CO through the sample cuvette using oxidised microsomes).
In most samples the amount of contaminating haemoglobin was extremely snlall (A42o--A49o -- 0--0.005). Since almost identical spectra are produced when haemoglobin binds CO and EIC, the largest possible error in measure- ments of the 455 : 428 nm peak height ratio of the EIC-difference spectra was calculated to be approx. 15%. This calculation was based on the largest acceptable peak at 420 nm. due to haemoglobin (i.e. A = 0.005) and the smallest (control) peak at: 428 nm due to cytochrome P-450 (A428- A49o ~ 0.04). The concentration of cytochrome b5 was determined from the difference spectrum between NADH-reduced microsomes and oxidised micro- somes [27] using the corrected extinction coefficient of 185 cm-' • mM-' [24] . All spectra were recorded on a Cary 118C spectrophotometer with a repetitive scan accessory. Holmium oxide was used to calibrate all spectra. In all cases, identical peak maxima from at least four of the five animals were recorded, hence, these results are expressed as the mode value.

283
The rate of oxidative N<lemethylation of 4-dimethylaminoantipyrine (DMAP) was measured by quantifying the production of formaldehyde as described [28]. The formaldehyde, trapped as t h e semicarbazone, was developed in double-strength Nash reagent [29] . The rate of B[a] P hydroxy- lation was measured by the radiometric assay of DePierre et al. [30] as im- proved by Nesnow et al. [31] , by quantifying the base-soluble metabolites following hexane-extraction of the unreacted B[a]P. Because an NADPH- regenerating system was employed in both metabolic assays, all tubes were preincubated for 15 min. The reactions were begun by the addition of sub- strate which, in the case of the B[a] P hydroxylase assay, affords the highest rate of metabolism compared to other methods of initiating the reaction [ 32]. The activity of NADPH-cytochrome P-450 reductase was measured by the rate of reduction of cytochrome c [33]. To prolong the linearity of the cytochrome c reduction, samples with high NADPH-cytochrome P-450 re- ductase activity were diluted to a final microsomal protein concentration of 0.2 mg-m1-1
The statistical significance between the sample means of control and treated groups for each parameter studies was analyzed by the Student 's t-test at the 1% level of significance (P < 0.01).
RESULTS
The effects o f PB, MC and their coadministration (PB + ~4C) PB-treatment resulted in a proliferation of the endoplasmic reticulum (as
indicated by an increase in the mg microsomal protein per g liver) in addition to a general increase in the liver to body weight ratio. Neither of these two parameters was significantly (P < 0.01) altered by MC treatment. Both PB and MC enhanced the concentration of cytochromes bs and P-450. The CO- difference spectrum of reduced PB-induced microsomes displayed an absor~)- tion maximum at 450 nm. The corresponding EIC<lifference spectrum exhi- bited two major peaks at 428 and 455 nm with a 455 : 428 nm peak height ratio of about 0.6. The absorption maximum of the reduced MC-induced microsome/CO complex was displaced to 448 nm. Both absorption maxima of the reduced MC-induced microsome/EIC complex were shifted from 428 to 429.4 nm and from 455 to 452 nm. The 452 : 429.4 nm peak height ratio increased to 1.9.
PB induction resulted in an increase in the concentration of cytochrome P-450 which was accompanied by an increase in the activity of DMAP N-demethylase (X 2.5), B[a] P hydroxylase (X 3.4) and NADPH-cytochrome c reductase (X 2.7). In the case of MC, no increase in the activity of DMAP N-demethylase or NADPH-cytochrome c reductase was observed but the increase in the concentration of cytochrome P-448 was accompanied by a large increase in B[a] P hydroxylase activity (X 15.6).
In nearly all cases the coadministration of PB and MC resulted in increases in enzymic activity and cytochrome concentration which were greater than the corresponding increases evoked by either PB or MC alone. Following the

284
coadministration of PB and MC, the CO~ifference spectrum showed max- imum absorbance at 448.5 nm, whilst the EIC<lifference spectrum showed two peaks at 428.5 and 452.0 nm with a 452.0 : 428.5 nm peak height ratio of about 1.2. The spectral shifts elicited by the coadministration of PB and MC were slightly less than those evoked by the administration of MC alone. In general, the qualitative spectral characteristics displayed by PB + MC-induced microsomes were intermediate between PB- and MC-induced microsomes.
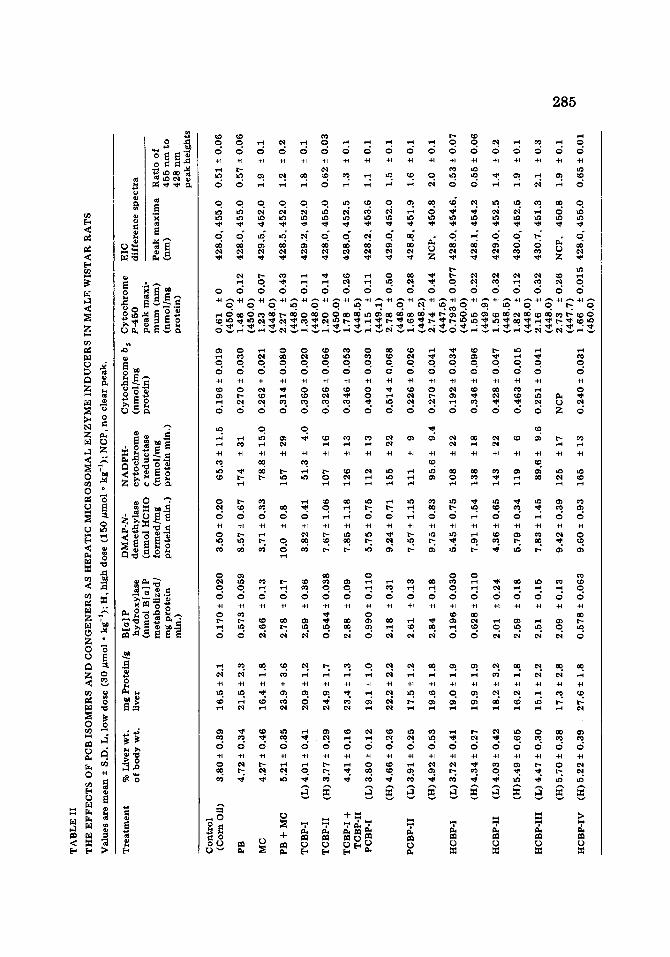
The effects o f TCBP-I, TCBP-II and their coadministration (TCBP I + II) The results shown in Table II confirmed that TCBP-II (with two para and
two ortho chlorine substituents) is a PB-type inducer of cytochrome P-450- dependent monooxygenase activity and that TCBP-I (with two para and two rneta chlorine substituents) is an MC-type inducer of cytochrome P-448- dependent monooxygenase activity. The coadministration of TCBP-I and II produced a pattern of mixed induction commensurate with that produced by the coadministration of PB and MC.
The effects o f PCBP-I, PCBP-II, HCBP-I, HCBP-II, HCBP-III and HCBP-IV At the high dosage level, the penta- and hexachlorobiphenyls tested all
stimulated the activity of DMAP N~lemethylase and NADPH-cytochrome c reductase. This response is a characteristic of PB-type inducers which distin- guishes them from MC-type inducers. These PCB isomers and congeners also enhanced the activity of B[a]P hydroxylase. With the exception of HCBP-I and HCBP-IV, the increase in B[a]P hydroxylase activity ( l l--17-fold) was significantly greater than that produced by either PB or TCBP-II (3--3.5- fold) and resembeled, in order of magnitude, the increase stimulated by MC or TCBP-I (15--16-fold). The enzyme activities induced by PCBP-I and II and by HCBP-II and III suggested that these PCBs possess both PB- and MC- type inducing properties. The effects described for the high dosage level of the penta- and hexachlorobiphenyls were also evident, albeit to varying degrees, at the low dosage level.
Like the enzymic properties, the spectral characteristics of microsomes from PCB-treated rats were dependent on both the structure of the PCB congener and the administered dosage level of the chemical. At the high dosage level, all PCB isomers and congeners increased the concentration of cytochromes bs and P-450. With the exception of HCBP-I and HCBP°IV, the quantitative changes in cytochrome P-450 were accompanied by qualita- tive alterations as indicated by hypsochromic shifts in the CO-difference spectra. Similarly, bathochromic and hypsochromic shifts in the 428 and 455 nm Sorer peaks, respectively, were observed in the EIC-difference spectra along with changes in the 455 : 428 nm peak height ratio. The quantitative and qualitative alterations in the spectral properties described for the high dosage level were also evident at the low dosage level. Like HpCBP, the effects of PCBP-I, PCBP-II, HCBP-II and HCBP-III suggest these PCB isomers and congeners possessed both PB- and MC-type inducing properties. HCBP-I, however, resembled a PB-type inducer. The results shown in Table II also
2 8 5
<
<
. <
z r~
N
r.r.1 ~
N ~ ~ z .< N
m ~ 0 °
g ~
Z T
Z °
~ a
r~
O ,I=
O ~
g
~ O ~ . ~ ~ .~
~ ®.~
~ 1 0 ~
O
o o o o o o o o o o o o o ~ o c~ o o 4-I +I + l 4-I -}-I +I 4-I +l -I-I -H +I +I +I +I 4-I -I-I -F-I 4-I
,~ o. ~ ,~ ~ ~ ~ ~ o. ~. ~. ,-~ 0~ ~. . o ~ o o ~ ~ o ~ o o o o o o o o o o
~+, . . . . . . . . ~'+~ ~+, ~+, ~+, ~+'~ +, ~+, ~+, ~ ~^~ ~+, . . . . . . ~" +, ~+, ~'+, ~,+, ~'+,~
~ ~ ~ A ~ A ~ , ~ "~ " "~ "~ " ~ " "~ ~ A ~ ~
o o. o. o o o o. o. o o o. o. o. o. o o. O O O O O O O O O O O O O O O O O +1 -H -I-I -H 4-1 + l +1 +1 4"t +1 + l +1 +1 +1 + l 4-1 +1
o o. ,~.
+I +I +I +I +l +I +I +I +I +I +I -H +I +I +I +I +I +l
+I ~-I -l-I +I +I +I +I 4-i -}-I +I +I +I 4-i +I +L +I +I +I
o. o . . ~ o. o. ~ , ~ . ~ o . ,-~ *~ ~ ~ . o o o o ~ o o o o o o ~ o o c; o o o o -H ~ +I +I +I 4-I -H 4-I 4-I -FI +I 4-I +I -H +I +I -l-f +I
• ~ . ~ r - ~. ~ ~ ~ ~. . ~ ,~ ~. . ~ . . o . .
-H -t4 +1 -I-I +1 +1 +1 -~-I 4-1 -I-I -H 4-1 4-1 4-1 4-1 -4-1 -.)-I +1
o o o o o o o o o o o ~ ~ o o o ~ o -I-I +1 -I-i 4-1 +1 +1 -H 4-1 +1 4-1 -I-I -H +1 -I-I 4-1 +1 -t-I -H
~ o o + ~ ~ :> o~

286
confirmed that HCBP-IV, when synthesized and purified by methods which eliminate chlorinated dibenzofurans, is a PB-type inducer of cytochrome P-450<iependent monooxygenase activity [34] .
DISCUSSION
Individual PCB isomers and congeners have been divided on a structural basis into PB-type and MC-type inducers [14--16] . It has been established that commercial mixtures of PCBs are mixed-type inducers of hepatic drug- metabolizing enzymes in the rat [10--13] . However, PCB congeners which would be expected to induce MC-type activity, namely, TCBP-I, 3,3' ,4,4' ,5- pentachlorobiphenyl and 3,3' ,4,4' ,5,5'-hexachlorobiphenyl, are only minor consti tuents of the commercial PCB mixtures [35] .
Recently, we reported that HpCBP is a mixed-type inducer of hepatic microsomal drug-metabolizing activity at the 150 pmol-kg- ' dosage level [20] . This particular isomer is only a trace component of the commercial PCB mixtures and clearly cannot be a major contr ibutor to their microsomal enzyme-induction activity. However, the properties of HpCBP suggested that congeners which contain the 3,3',4,4'-, 3,3' ,4,4' ,5,5' substitution (i.e. the structural prerequisite for MC-type induction} pattern plus one additional ortho chloro substi tuent may also be mixed-type inducers. Thus, all the remaining PCBs which fit these structural requirements, namely PCBP-I, PCBP-II, HCBP-I, HCBP-II and HCBP-III, were synthesized for this study. In addition, Aust and coworkers have reported that the brominated analogue of HCBP-I, which was isolated and purified (but not synthesized} from a com- mercial PBB mixture, was an inducer of both PB- and MC-type activity [ 17] . The results obtained for the synthetic PCB isomers and congeners were com- pared to parallel studies using PB-type inducers (PB and TCBP-II) and MC- type inducers (MC and TCBP-I}. The effects of a mixed-type inducer were simulated by coadministering either PB with MC or TCBP-I with TCBP-II. Like HpCBP, PCBP-I, PCBP-II, HCBP-II and HCBP-III exhibited mixed-type induction activity at the high (150 p m o l - k g -1) dosage level. However, HCBP-I resembled a PB-type inducer and differed significantly, therefore, from both the structurally-related PCB congeners and its PBB analogue. Further investigations into the reasons for this exceptional behaviour of HCBP-I are currently in progress.
The mixed activity of the four PCB isomers and congeners confirmed that in certain cases the addition of one ortho chloro substi tuent to MC-type inducers does not eliminate this induction activity. The results shown for HCBP-IV, a congener which contains two ortho substituents, suggest that this structural feature does eliminate MC-type activity and the properties of HCBP-IV induced microsomes were consistent with a PB-type induction pat- tern as previously reported [34] . The properties of PCBP I-II, HCBP I-III and HpCBP also suggest that two para chloro substi tuent {4 and 4'} and only one ortho substi tuent are sufficient structural prerequisites for PB-type induction activity for these penta, hexa and heptachlorinated biphenyl

287
isomers and congeners. The data also illustrated that the position of the ortho chloro substituent greatly influenced the MC-type activity of the PCB in- ducer. Comparison of the effects of PCBP-I versus PCBP-II and HCBP-I versus HCBP-II at low doses suggested that the 2,4,5-trichloro substitution results in a greater diminution of the MC-type specific activity of the PCB than does the 2,3,4-trichloro substitution pattern.
Since PCBP-I and PCBP-II have been identified as relatively major com- ponents of Clophen A50, Clophen A60 [35] and Aroclor 1254 [36] and HCBP-III as a minor component of Aroclor 1254, the results suggest that the mixed-induction properties of these individual isomers may play a significant role in the overall activity of these commercial mixtures.
It has been suggested that some of the toxic properties of several classes of halogenated aromatics may be related to their induction of microsomal cyto- chrome P-448 and its associated enzyme activities [14]. An enhanced activity of the cytochrome P-448-dependent monooxygenases may also be associated with an increased susceptibility to the cytotoxic effects of certain classes of chemical carcinogens such as the polynuclear aryl hydrocarbons [37--39]. Therefore, the biological effects, environmental occurrence and toxicity of the mixed-ty,~e inducers warrants further investigation.
ACKNOWLEDGEMENTS
The financial assistance of the Research Programs Directorate Health and Welfare Canada, (606-1444-X), the National Cancer Institute {U.S.A.), DHEW, Grant Number 1 BO1 CA21814-01 and the National Research Council of Canada is gratefully acknowledged.
REFERENCES
1 A.H. Conney, Pharmacological implications of microsomal enzyme induction, Phar- macol. Rev., 19 (1967) 317.
2 A.Y.H. Lu, R. Kuntzman and A.H. Conney, The liver microsomal hydroxylation enzyme system. Induction and properties of the functional components, Front. Gastrointest. Res., 2 (1976) 1.
3 N.E. Sladek and G.J. Mannering, Evidence for a new P-450 hemoprotein in hepatic microsomes from methylcholanthrene treated rats, Biochem. Biophys. Res. Commun., 24 (1966) 668.
4 Y. Gnasspelius, H. Thor and S. Orrhenius, A comparative study on the effects of phenobarbital and 3,4-benzpyrene on the hydroxylating system of rat-liver micro- somes, Chem.-Biol. Interact., 1 (1969/70) 125.
5 A.P. Alvares, G. Schilling, W. Levin and R. Kuntzman, Studies on the induction of CO-binding pigments in liver microsomes by phenobarbital and 3°methylcholanthrene, Biochem. Biophys. Res. Commun., 29 (1967) 521.
6 A.Y.H. Lu, R. Kuntzman, S. West, M. Jacobson and A.H. Conney, Reconstituted liver microsomal enzyme system that hYdroxylates drugs, other foreign compounds, and endogenous substrates. II. Role of the cytochrome P-450 and P-448 fractions in drug and steroid hydroxylations, J. Biol. Chem., 247 (1972) 1727.
7 C.L. Litterist, T.M. Farber, A.M. Baker and E.J. Van Loon, Effect of polychlorinated biphenyls on hepatic microsomal enzymes in the rat, Toxicol. Appl. Pharmacol., 23 (1972) 112.

288
8 D.J. Ecobichon and A.M. Comeau, Comparative effects of commercial aroclors on rat liver enzyme activities, Chem.-Biol. Interact., 9 (1974) 341.
9 A.P. Alvares and A. Kappas, The inducing properties of polychlorinated biphenyls on hepatic monooxygenases, Clin. Pharmacol. Ther., 22 (1977) 809.
10 A.P. Alvares, D.R. Bickers and A. Kappas, Potychlorinated biphenyls: A new type of inducer of cytochrome P-448 in the liver, Proc. Natl. Acad. Sci. U.S.A., 70 (1973) 1321.
11 A.P. Alvares and A. Kappas, Heterogeneity of cytochrome P-450s induced by poly- chlorinated biphenyls, J. Biol. Chem., 252 (1977) 6373.
12 D.E. Ryan, P.E. Thomas and W. Levin, Properties of purified liver microsomal cyto- chrome P-450 from rats treated with the polychlorinated biphenyl mixture Aroclor 1254, Mol. Pharmacol., 13 (1977) 521.
13 D.E. Ryan, P.E. Thomas, D. Korzeniowski and W. Levin, Separation and characteri- zation of highly purified forms of liver microsomal cytochrome P-450 from rats treated with polychlorinated biphenyls, phenobarbital, and 3-methylcholanthrene, J. Biol. Chem., 254 (1979) 1365.
14 A. Poland and E. Glover, Chlorinated biphenyl induction of aryl hydrocarbon hy- droxylase activity: A study of the structure-activity relationship, Mol. Pharmacol., 13 (1977) 924.
15 J.A. Goldstein, P. Hickman, H. Bergman, J.D. McKinney and M.P. Walker, Separation of pure polychlorinated biphenyl isomers into two types of inducers on the basis of induction of cytochrome P-450 or P-448, Chem.-Biol. Interact., 17 (1977) 69.
16 H. Yoshimura, N. Ozawa and S. Saeki, Inductive effects of polychlorinated biphenyls mixture and individual isomers on the hepatic microsomal enzymes, Chem. Pharm. Bull., 26 (1978) 1215.
17 G.A. Dannan, R.W. Moore, L.C. Besaw and S.D. Aust, 2,4,5,3',4',5'-Hexabromobi- phenyl is both a 3-methylcholanthrene- and a phenobarbital-type inducer of micro- somal drug metabolizing enzymes, Biochem. Biophys. Res. Commun., 85 (1978) 450.
18 M.D. Stonard, Mixed type hepatic microsomal enzyme induction by hexachloro- benzene, Biochem. Pharmacol., 24 (1975) 1959.
19 J.A. Goldstein, M. Friesen, T.M. Scotti, P. Hickman, J.R. Hass and H. Bergman, Assessment of the contribution of chlorinated dibenzo-p-dioxins and dibenzofurans to hexachlorobenzene-induced toxicity, porphyria, changes in mixed function oxygen- ases, and histopathological changes, Toxicol. Appl. Pharmacol., 46 (1978) 633.
20 A. Parkinson, R. Cockerline and S. Safe, Induction of both 3-methylcholanthrene and phenobarbitone-type activity by a single polyehlorinated biphenyl isomer, Biochem. Pharmacol., in press.
21 J.I.G. Cadogan, A convenient new method of aromatic arylation, J. Chem. Soc. (Lond.), (1962) 4257.
22 O. Hutzinger, S. Safe and V. Zitko, P~lychlorinated biphenyls: synthesis of some individual isomers, Bull. Environ. Contam. Toxicol., 6 (1971) 209.
23 O.H. Lowry, N.J. Rosebrough, A.L. Farr and R.J. Randall, Protein measurement with Folin phenol reagent, J. Biol. Chem., 193 (1951) 265.
24 T. Omura and R. Sato, The carbon monoxide-binding pigment of liver microsomes. I. Evidence for its haemoprotein nature, J. Biol. Chem., 239 (1964) 2370.
25 H. Nishibayashi, T. Omura and R. Sato, The~binding of ethyl isocyanide by hepatic microsomai hemoprotein, Biochim. Biophys. Acta., 118 (1966) 651.
26 J.B. Schenkman, H. Returner and R.W. Estabrook, Spectral studies of drug interaction with hepatic microsomal cytochrome, Mol. Pharmacoi., 3 (1967) 113.
27 I. Raw and H.R. Mahler, Studies of electron transport enzymes. III. Cytochrome b~ of pig liver mitochondria, J. Biol. Chem., 234 (1959) 1867.
28 A. Parkinson and S. Safe, The detection of enzyme induction by rat liver microsomes prepared by isoelectric precipitation, J. Pharm. Pharmacol., 31 (1979) 444.

289
29 T. Nash, The colorimetric estimation of formaldehyde by means of the Hantzsh reaction, Biochem. J., 55 (1953) 416.
30 J.W. DePierre, M.S. Morson, K.A.M. Johannesen and L. Ernster, A reliable, sensitive and convenient radioactive assay for benzpyrene monooxygenase, Anal. Biochem., 63 (1975) 470.
31 S. Nesnow, W.E. Fahl and C.R. Je fcoa te , An improved radiochemical assay for benzo[a ]pyrene monooxygenase, Anal. Biochem., 80 (1977) 258.
32 C.S. Yang, F.A. Strickhart and L.P. Kicha, Analysis of the aryl hydrocarbon hydroxy- lase assay, Biochem. Pharmacol., 27 (1978) 2321.
33 C.H. Williams and H. Kamin, Microsomal t r iphosphopyridine nucleotide-cytochrome c reductase of liver, J. Biol. Chem., 237 (1962) 587.
34 J.A. Goldstein, J.R. I-Iass, P. Linko and D.J. Harvan, 2,3,7,8-Tetrachlorodibenzofuran in a commercially available 99% pure polychlorinated biphenyl isomer identified as the inducer of hepatic cytochrome P-448 and aryl hydroxylase in the rat, Drug Metab. Dlspos., 6 (1978) 258.
35 S. Jensen and G. Sundstrom, Structures and levels of most chlorobiphenyls in two technical PCB products and in human adipose tissue, Ambio, 3 (1974) 70.
36 D. Sissons and D. Welti, Structural identification of polychlorinated biphenyls in commercial mixtures by gas-liquid chromatography nuclear magnetic resonance and mass spectrometry, J. Chromatogr., 60 (1971) 15.
37 D.L. Busbee, J. Guyden, T. Kingston, F.L. Rose and E.T. Cantrell, Metabolism of benzo[a ]pyrene in animals with high aryl hydrocarbon hydroxylase levels and high rates of spontaneous cancer, Cancer Lett. , 4 (1978) 61.
38 D.W. Nebert and S.A. Atlas, The Ah locus: Aromatic hydrocarbon responsiveness of mice and men, Human Genet., Suppl. 1 (1978) 149.
39 A.R. Boobis, D.W. Nebert and O. Pelkonen, Effects of microsomal enzyme inducers in vivo and inhibitors in vitro on the covalent binding of benzo[a ]pyrene metaboli tes to DNA catalyzed by liver microsomes from genetically responsive and non-responsive mice, Biochem. Pharmacol., 28 (1979) 111.
Related Documents











