12 Springer Dear Author: Please find attached the final pdf file of your contribution, which can be viewed using the Acrobat Reader, version 3.0 or higher. We would kindly like to draw your attention to the fact that copyright law is also valid for electronic products. This means especially that: • You may print the file and distribute it amongst your colleagues in the scientific community for scientific and/or personal use. • You may make your article published by Springer-Verlag available on your personal home page provided the source of the published article is cited and Springer-Verlag and/or other owner is mentioned as copyright holder. You are requested to create a link to the published article in Springer's internet service. The link must be accompanied by the following text: "The original publication is available at springerlink.com". Please use the appropriate DOI for the article. Articles disseminated via SpringerLink are indexed, abstracted and referenced by many abstracting and information services, bibliographic networks, subscription agencies, library networks and consortia. • Without having asked Springer-Verlag for a separate permission your institute/your company is not allowed to place this file on its homepage. • You may not alter the pdf file, as changes to the published contribution are prohibited by copyright law. • Please address any queries to the production editor of the journal in question, giving your name, the journal title, volume and first page number. Yours sincerely, Springer-Verlag

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

12 Springer
Dear Author: Please find attached the final pdf file of your contribution, which can be viewed using the Acrobat Reader, version 3.0 or higher. We would kindly like to draw your attention to the fact that copyright law is also valid for electronic products. This means especially that:
• You may print the file and distribute it amongst your colleagues in the scientific community for scientific and/or personal use.
• You may make your article published by Springer-Verlag available on your personal home page provided the source of the published article is cited and Springer-Verlag and/or other owner is mentioned as copyright holder. You are requested to create a link to the published article in Springer's internet service. The link must be accompanied by the following text: "The original publication is available at springerlink.com". Please use the appropriate DOI for the article. Articles disseminated via SpringerLink are indexed, abstracted and referenced by many abstracting and information services, bibliographic networks, subscription agencies, library networks and consortia.
• Without having asked Springer-Verlag for a separate permission your institute/your company is not allowed to place this file on its homepage.
• You may not alter the pdf file, as changes to the published contribution are prohibited by copyright law.
• Please address any queries to the production editor of the journal in question, giving your name, the journal title, volume and first page number.
Yours sincerely, Springer-Verlag
Verwendete Distiller 5.0.x Joboptions
Dieser Report wurde automatisch mit Hilfe der Adobe Acrobat Distiller Erweiterung "Distiller Secrets v1.0.5" der IMPRESSED GmbH erstellt. Sie koennen diese Startup-Datei für die Distiller Versionen 4.0.5 und 5.0.x kostenlos unter http://www.impressed.de herunterladen. ALLGEMEIN ---------------------------------------- Dateioptionen: Kompatibilität: PDF 1.3 Für schnelle Web-Anzeige optimieren: Ja Piktogramme einbetten: Ja Seiten automatisch drehen: Nein Seiten von: 1 Seiten bis: Alle Seiten Bund: Links Auflösung: [ 300 300 ] dpi Papierformat: [ 595 842 ] Punkt KOMPRIMIERUNG ---------------------------------------- Farbbilder: Downsampling: Ja Berechnungsmethode: Bikubische Neuberechnung Downsample-Auflösung: 150 dpi Downsampling für Bilder über: 225 dpi Komprimieren: Ja Automatische Bestimmung der Komprimierungsart: Ja JPEG-Qualität: Mittel Bitanzahl pro Pixel: Wie Original Bit Graustufenbilder: Downsampling: Ja Berechnungsmethode: Bikubische Neuberechnung Downsample-Auflösung: 150 dpi Downsampling für Bilder über: 225 dpi Komprimieren: Ja Automatische Bestimmung der Komprimierungsart: Ja JPEG-Qualität: Mittel Bitanzahl pro Pixel: Wie Original Bit Schwarzweiß-Bilder: Downsampling: Ja Berechnungsmethode: Bikubische Neuberechnung Downsample-Auflösung: 600 dpi Downsampling für Bilder über: 900 dpi Komprimieren: Ja Komprimierungsart: CCITT CCITT-Gruppe: 4 Graustufen glätten: Nein Text und Vektorgrafiken komprimieren: Ja SCHRIFTEN ---------------------------------------- Alle Schriften einbetten: Ja Untergruppen aller eingebetteten Schriften: Nein Wenn Einbetten fehlschlägt: Warnen und weiter Einbetten: Immer einbetten: [ ] Nie einbetten: [ ] FARBE(N) ---------------------------------------- Farbmanagement: Farbumrechnungsmethode: Alle Farben zu sRGB konvertieren Methode: Standard Arbeitsbereiche: Graustufen ICC-Profil: RGB ICC-Profil: sRGB IEC61966-2.1 CMYK ICC-Profil: U.S. Web Coated (SWOP) v2 Geräteabhängige Daten: Einstellungen für Überdrucken beibehalten: Ja Unterfarbreduktion und Schwarzaufbau beibehalten: Ja Transferfunktionen: Anwenden Rastereinstellungen beibehalten: Ja ERWEITERT ---------------------------------------- Optionen: Prolog/Epilog verwenden: Nein PostScript-Datei darf Einstellungen überschreiben: Ja Level 2 copypage-Semantik beibehalten: Ja Portable Job Ticket in PDF-Datei speichern: Nein Illustrator-Überdruckmodus: Ja Farbverläufe zu weichen Nuancen konvertieren: Nein ASCII-Format: Nein Document Structuring Conventions (DSC): DSC-Kommentare verarbeiten: Nein ANDERE ---------------------------------------- Distiller-Kern Version: 5000 ZIP-Komprimierung verwenden: Ja Optimierungen deaktivieren: Nein Bildspeicher: 524288 Byte Farbbilder glätten: Nein Graustufenbilder glätten: Nein Bilder (< 257 Farben) in indizierten Farbraum konvertieren: Ja sRGB ICC-Profil: sRGB IEC61966-2.1 ENDE DES REPORTS ---------------------------------------- IMPRESSED GmbH Bahrenfelder Chaussee 49 22761 Hamburg, Germany Tel. +49 40 897189-0 Fax +49 40 897189-71 Email: [email protected] Web: www.impressed.de
Adobe Acrobat Distiller 5.0.x Joboption Datei
<< /ColorSettingsFile () /AntiAliasMonoImages false /CannotEmbedFontPolicy /Warning /ParseDSCComments false /DoThumbnails true /CompressPages true /CalRGBProfile (sRGB IEC61966-2.1) /MaxSubsetPct 100 /EncodeColorImages true /GrayImageFilter /DCTEncode /Optimize true /ParseDSCCommentsForDocInfo false /EmitDSCWarnings false /CalGrayProfile () /NeverEmbed [ ] /GrayImageDownsampleThreshold 1.5 /UsePrologue false /GrayImageDict << /QFactor 0.9 /Blend 1 /HSamples [ 2 1 1 2 ] /VSamples [ 2 1 1 2 ] >> /AutoFilterColorImages true /sRGBProfile (sRGB IEC61966-2.1) /ColorImageDepth -1 /PreserveOverprintSettings true /AutoRotatePages /None /UCRandBGInfo /Preserve /EmbedAllFonts true /CompatibilityLevel 1.3 /StartPage 1 /AntiAliasColorImages false /CreateJobTicket false /ConvertImagesToIndexed true /ColorImageDownsampleType /Bicubic /ColorImageDownsampleThreshold 1.5 /MonoImageDownsampleType /Bicubic /DetectBlends false /GrayImageDownsampleType /Bicubic /PreserveEPSInfo false /GrayACSImageDict << /VSamples [ 2 1 1 2 ] /QFactor 0.76 /Blend 1 /HSamples [ 2 1 1 2 ] /ColorTransform 1 >> /ColorACSImageDict << /VSamples [ 2 1 1 2 ] /QFactor 0.76 /Blend 1 /HSamples [ 2 1 1 2 ] /ColorTransform 1 >> /PreserveCopyPage true /EncodeMonoImages true /ColorConversionStrategy /sRGB /PreserveOPIComments false /AntiAliasGrayImages false /GrayImageDepth -1 /ColorImageResolution 150 /EndPage -1 /AutoPositionEPSFiles false /MonoImageDepth -1 /TransferFunctionInfo /Apply /EncodeGrayImages true /DownsampleGrayImages true /DownsampleMonoImages true /DownsampleColorImages true /MonoImageDownsampleThreshold 1.5 /MonoImageDict << /K -1 >> /Binding /Left /CalCMYKProfile (U.S. Web Coated (SWOP) v2) /MonoImageResolution 600 /AutoFilterGrayImages true /AlwaysEmbed [ ] /ImageMemory 524288 /SubsetFonts false /DefaultRenderingIntent /Default /OPM 1 /MonoImageFilter /CCITTFaxEncode /GrayImageResolution 150 /ColorImageFilter /DCTEncode /PreserveHalftoneInfo true /ColorImageDict << /QFactor 0.9 /Blend 1 /HSamples [ 2 1 1 2 ] /VSamples [ 2 1 1 2 ] >> /ASCII85EncodePages false /LockDistillerParams false >> setdistillerparams << /PageSize [ 595.276 841.890 ] /HWResolution [ 300 300 ] >> setpagedevice

Polychaete assemblages along a rocky shore on the South Adriatic coast(Mediterranean Sea): patterns of spatial distribution
Received: 8 February 2003 / Accepted: 25 June 2003 / Published online: 6 September 2003� Springer-Verlag 2003
Abstract This study reports on the distribution of poly-chaete assemblages along a depth gradient from 5 to25 m, along a stretch of rocky coast near Otranto (SouthAdriatic sea, Italy). Three depths were sampled in Mayand November 2000. At each depth three sites (about100 m apart) were sampled by scraping off three repli-cate quadrats of 20·20 cm. The experimental designenabled identification of across- and along-shore spatialpatterns of variation of polychaete assemblages. A totalof 4,168 specimens, belonging to 152 taxa were collected.Multivariate analysis showed that the polychaeteassemblages differed significantly among depths with theclearest differences between the shallowest sites (5 m)and the deeper ones (15–25 m). A considerable source ofvariation among sites at each level of the shore was alsoexhibited by the analyses, with the greatest differencesamong sites within depths recorded at 5 m. The speciesmost contributing to the differentiation of assemblagesamong depths and sites within each depth were identi-fied. Some potential causes of the observed differencesare hypothesized and discussed. The importance ofquantitative observation to provide the context forstudying the underlying ecological processes is alsostressed.
Introduction
Most of the information on patterns of distribution ofMediterranean benthic assemblages has been based onclassic studies of zonation along depth gradients, and
changes in assemblage structure have been interpreted inrelation to environmental factors such as light intensityand hydrodynamic features (Riedl 1971; Peres 1982; Roset al. 1985). Thus, differences in the structure ofassemblages were explained in relation to only a fewenvironmental variables, often overlooking the potentialimportance of a suite of other sources of variability.However, it has been demonstrated as in many differenthabitats vertical gradients alone cannot explain all thevariability observed in patterns of spatial distribution(Benedetti-Cecchi 2001; Terlizzi et al. 2003).
Recently, it has been shown that a considerablehorizontal variability in abundances of organisms onrocky shores occurs on scales of 10s of meters or less(Kennelly and Underwood 1993; Chapman et al. 1995;Cornell and Karlson 1996; Roberts 1996; Underwoodand Chapman 1996; Guichard and Bourget 1998; Smithand Witman 1999; Benedetti-Cecchi 2001). Such pat-terns, however, are well documented only in rockyintertidal assemblages of temperate latitudes, and studieshave often been focused on large and conspicuous sessileorganisms (Caffey 1985; Bertness et al. 1991; Under-wood and Chapman, 1996; 1998a; 1998b; Menconi et al.1999; Benedetti-Cecchi 2000; Benedetti-Cecchi 2001).The information is limited to sessile and easily recog-nizable taxa also in the subtidal (Ferdeghini et al. 2000;Fraschetti et al. 2001). Spatial distributions of small andcryptic vagile invertebrates, abundant within algae onrocky shores, have been scarcely studied. In contrast, ithas been recently demonstrated that the biodiversity ofvagile invertebrates can vary significantly among differ-ent algal structures on a small scale (Chemello andMilazzo 2002), and on a scale of 100s of meters (Terlizziet al. 2003), or, on the same substrate, between sites on ascale of 10s of meters (Kelaher et al. 2001).
Among the vagile invertebrates, polychaetes are oneof the best-represented groups. Due to their abundanceand diversified feeding types, data on polychaetes couldprovide the rationale for investigating biological pro-cesses and physical factors responsible for structuringpatterns of biodiversity.
Marine Biology (2003) 143: 1109–1116DOI 10.1007/s00227-003-1162-0
A. Giangrande Æ A. L. Delos Æ S. FraschettiL. Musco Æ M. Licciano Æ A. Terlizzi
Communicated by R. Cattaneo-Vietti, Genova
A. Giangrande (&) Æ A. L. Delos Æ S. Fraschetti Æ L. MuscoM. Licciano Æ A. TerlizziDepartment of Biological and Environmental Science and Tech-nology, University of Lecce, CoNISMa, 73100 Lecce, ItalyE-mail: [email protected].: +39-832-298659Fax: +39-832-298626

In the Mediterranean Sea, hard-substrate polychaeteshave been well investigated (Bellan 1969; 1971; Katz-mann 1971; Somaschini 1988; Sarda 1991; Alos 1999;Lopez and Vietez 1999; Tena et al. 2000).
Abbiati et al. (1987) and Giangrande (1988) haveexamined the polychaete distributions along one verticaltransect, while Fresi et al. (1983; 1984) analysed patternsof distribution including also a horizontal gradient. Allof these studies, however, were based on poorly repli-cated sampling designs. In Fraschetti et al. (2002) the useof replicates showed that polychaete assemblages, atleast among the fringe of brown algae of the genusCystoseira (0.5–1 m depth), did not show a great deal ofvariability on a large scale (>100s km), or on the scaleof replicates (10s cm).
The aim of this study is to provide information aboutpolychaete assemblage distributions along a vertical cliffin the South Adriatic Sea (Mediterranean), quantifyingpossible differences in assemblage structure along adepth gradient and among different sites, located 100 mapart, at each of the considered depths.
Materials and methods
Study area
The investigated location is situated along the south-eastern coastof Apulia (Italy, 40�08¢N, 18�30¢E) (Fig. 1). The coast comprisesabout 60 km of steep rocky shores abruptly extending from theshore to about 30 m. It has been recently included in a list ofEuropean Marine Biodiversity Research Sites selected because ofits pristine status (BIOMARE, http://www.pml.ac.uk/biomare/si-te.htm). A dense fringe of the brown algae Cystoseira amentaceacharacterizes the shallow infralittoral. From the shore to 5–7 mdepth, the rocky substrate is steep and characterized by patches ofhydroids (Aglaophenia sp., Eudendrium sp.) and photophilicassemblages, mainly dominated by algae of the genera Dictyota sp.,Laurencia complex, Acetabularia acetabulum and Corallina sp.,encrusting calcified red algae, and encrusting (e.g. Crambe crambe)and globose sponges (e.g. Chondrilla nucula).
At 12–15 m the assemblage is mainly characterized by botherect (e.g. Flabellia petiolata, Padina pavonica, Peyssonneliasquamaria) and encrusting algae (Peyssonnelia sp., Lithophyllumsp., Mesophyllum sp.), and by sponges of the species Agelas oroides,Phorbas spp. Ircinia spp. and Sarcotragus spp.. Below 20 m, thesubstrate is composed of ‘‘coralligenous formations’’, a termbroadly defining the secondary hard substrate formed by encrust-ing calcified algae, bryozoans, serpulids and the complex biotainhabiting them (Sara 1969; Ros et al. 1985). Among sessileinvertebrates, the most characteristic species are large sponges(Axinella spp.), anthozoans (Leptosammia pruvoti, Parazoanthusaxinellae), bryozoans (Calpensia nobilis, Myriapora truncata cryp-tica, Pentapora fasciata) and ascidians (Microcosmus sp. andHalocynthia papillosa) (Terlizzi et al. 2003).
Experimental design and sampling methods
Samples were collected by SCUBA divers by scraping off the rockysubstrate on a quadrat of 20·20 cm. Three sites (about 100 m apart)were randomly chosen froma set of possible sites separatedbyat least50–60 m and providing comparable habitats in terms of biologicalassemblages, type and slope of the substrate and exposure of waves.
At each site, samplings were undertaken at three depths (5, 10,25 m) in three replicates at each depth. Sampling was repeatedtwice in May and November 2000 with a total of 54 samples. After
collection, samples were fixed in 8% formaldehyde solution, sortedunder a binocular microscope, preserved in 70% ethanol andtaxonomically identified. Nomenclature followed San Martin(1984), Castelli et al. (1995) and Licher (1999).
Data were organized in a taxon/sample matrix and analysed bymultivariate techniques. Two-way crossed analysis of similarities(ANOSIM, Clarke 1993) was used to test for differences inassemblage structures, among both depths and sites at each time ofsampling. Comparisons were based on Bray-Curtis similarity val-ues (SBC) calculated on all species within each quadrat (PRIMER,Plymouth Marine Laboratory) (Clarke 1993). Prior to analysis datawere square-root-transformed to arrange all taxa in the same rangeof abundance and prevent a few abundant species from dominatingthe analyses. Differences among depths as well as among sites ateach depth were represented by non-metric multidimensionalscaling ordinations (nMDS), considering all replicates and thecentroids (average) of assemblages from the three replicates col-lected at the three depths at each of the three sites. For each periodof sampling, the species (or taxa) contributing most to similaritieswithin each depth and, at each depth, to the dissimilarity (calcu-lated as 1-SBC) among sites were investigated using a similaritypercentage procedure (SIMPER, Clarke 1993). Such proceduresallowed identification, at both sampling times, of the species (ortaxa) most responsible for characterizing depths and/or for differ-entiating, at each depth, sites. Species (or taxa) were consideredimportant if they exceeded an arbitrarily chosen threshold value of5% of similarity within depths and/or 3% of dissimilarity amongsites at each depth.
Fig. 1 Sampling location and its positioning in the Mediterraneanregion
1110

Results
A total of 4,168 specimens belonging to 152 taxa ofpolychaetes were collected. Of these, 138 were identifiedto species level (Table 1). On the whole, 22 families wererecorded. The most speciose family was Syllidae, with 63taxa. Sabellidae accounted for 17 taxa, followed bySerpulidae (15), Eunicidae (11), Phyllodocidae (9), andNereididae (8).
Taxonomic considerations
Sixty species represent the first record of these taxa for theSouth Adriatic Sea (Table 1). Within the family Syllidae,the species Syllis bouvieri Gravier 1900, and S. truncatacryptica Ben-Ehliau 1977 sensu San Martin (1984) havebeen reported. Even though Licher (1999) considers theformer species synonymous with S. prolifera (Krhon1852), and the latter synonymous with S. gerlachi (Hart-mann-Schroder 1960), in our opinion the difference inchaetal and acicular shape in both species characterizesS. bouvieri and S. truncata cryptica. Among this family, anew taxon for the Mediterranean fauna was collected,Syllis alosae San Martin 1992, a species described in theGulf of Mexico, but also reported in the Caribbean area(Licher 1999). In addition, three new species from theItalian coast have been reported: (1) Syllis cruzi Nunezand San Martin 1991, (2) Pionosyllis serratisetosa Lopez,SanMartin and Jimenez 1996, previously known from theAtlantic Ocean and the Spanish Mediterranean coast(Nunez et al. 1992; Lopez et al. 1996), and (3) S. gerund-ensis (Alos and Campoy 1981), previously reported onlyfrom the Spanish Mediterranean coast. Other species,such as S. jorgei SanMartin and Lopez 2000, S. compactaGravier 1900, and S. pulvinata (Langerhans 1881),although not included on the checklist of the Italianpolychaetes (Castelli et al. 1995), must be consideredalready present along the Italian coast. This is due to theirrecent synonymizations with species already reported onthe checklist: S. jorgei with S. lutea San Martin 1984(Licher 1999), S. compacta with S. golfonovoensis (Hart-mann-Schroder 1962) (Lopez et al. 1996), andS. pulvinatawith S. mediterranea San Martin 1984 (Licher 1999).Lastly, the absence of S. vivipara Krohn, 1869 from thechecklist must be considered an omission.
Two new taxa of the family Sabellidae have been foundand are yet to be described: Hypsicomus sp. and Pseud-obranchiomma sp. The first genus, endemic of the Medi-terranean Sea, was up to now considered monotypic,being represented by only one species,H. stichophthalmus(Grube 1863). The genus Pseudobranchiomma sp. wasrecently found in the Mediterranean Sea with the speciesP. tarantoensis (Knight-Jones and Giangrande 2003).Until now, this genus has been represented in the Medi-terranean by two taxa found exclusively along thesouthern Italian coast.
Table 1 Taxonomic list of the species recorded. * New for theSouth Adriatic Coast, ** new for the Italian coast, *** new for theMediterranean Sea
Spionidae
*Laonice cirrata (Sars, 1851)*Polydora ciliata (Johnston, 1838)Prionospio cirrifera Wiren, 1883*Spio filicornis (Muller, 1766)CirratulidaeCaulleriella bioculata (Keferstein, 1862)Dodecaceria concharum Oersted, 1843OpheliidaePolyophthalmus pictus(Dujardin, 1839)Phyllodocidae*Notophyllum foliosum (Sars, 1835)*Phyllodoce lamelligera (Linnaeus, 1791)*Phyllodoce laminosa Savigny, 1818*Phyllodoce mucosa (Oersted, 1843)Phyllodoce pusilla(Claparede, 1868)*Phyllodoce vittata Ehlers, 1864*Pirakia punctifera (Grube, 1860)*Pterocirrus limbatus (Claparede, 1868)Pterocirrus macroceros (Grube, 1860)GlyceridaeGlycera tesselata Grube, 1863Goniadidae*Goniada maculata Oersted, 1843HesionidaeKefersteinia cirrata (Keferstein, 1862)Syllidae*Autolytus benazzii Cognetti, 1853*Autolytus brachycephalus (Marenzeller, 1874)*Autolytus convolutus Cognetti, 1953*Autolytus edwardsii Saint-Joseph, 1887*Autolytus prolifer (Muller, 1788)*Autolytus quindecemdentatus Langerhans, 1884Autolytus sp.*Branchiosyllis exilis (Gravier, 1900)Brania arminii Langerhans, 1880Brania pusilla (Dujardin, 1839)*Ehlersia ferrugina (Langerhans, 1881)Ehlersia sp.Eurysyllis tuberculata Ehlers, 1864*Eusyllis assimilis Marenzeller, 1875*Eusyllis lamelligera Marion and Bobretzky, 1875Eusyllissp.Exogone dispar Webster, 1879Exogone naidina Oersted, 1845*Exogone rostrata Naville, 1933*Exogone verugera (Claparede, 1868)Exogonesp.Grubeosyllis clavata (Claparede, 1863)*Grubeosyllis limbata (Claparede, 1868)Grubeosyllis vieitezi (San Martin, 1984)Haplosyllis spongicola (Grube, 1855)*Odontosyllis fulgurans (Audouin and M. Edwards, 1934)*Odontosyllis gibba Claparede, 1863*Parapionosyllis brevicirra Day, 1954*Parapionosyllis minuta (Pierantoni, 1903)*Pionosyllis lamelligera Saint-Joseph, 1856*Pionosyllis morenoae San Martin, 1984**Pionosyllis serratisetosa Lopez et.al, 1997*Procerea macrophthalma Marenzeller, 1975*Pseudosyllis brevipennis Grube, 1863*Sphaerosyllis cryptica Ben-Ehliau, 1977Sphaerosyllis hystrix Claparede, 1863Sphaerosyllis pirifera Claparede, 1868*Sphaerosyllis taylori Perkins, 1981Syllides fulvus Marion and Bobretzky, 1875
1111

Lastly, within the Eunicidae family, the speciesLysibranchia paucibranchiata Cantone 1983 was found.This genus, endemic of the Mediterranean Sea, is atpresent known only in the eastern Mediterranean(Cantone 1983).
Assemblage structure
ANOSIM revealed at both times of sampling significantdifferences among depths in the structure of assemblages(Table 2). Pairwise tests showed that, in both periods,the highest differences were between 5 m versus 15 and25 m, the lowest between 15 m versus 25 m (Table 2).
ANOSIM also detected (averaged across depths)significant differences among sites. The average values ofdissimilarity among sites calculated at each depth inboth periods are shown in Fig. 2 to indicate how theassemblages differed among sites at each depth. Thehighest values of dissimilarity among sites were recorded
Table 1 (Contd.)
Spionidae
***Syllis alosae San Martin, 1992Syllis armillaris Muller, 1771*Syllis beneliahuae Campoy and Alquezar, 1982Syllis bouvieri Gravier, 1900*Syllis compacta Gravier, 1900Syllis columbretensis (Campoy, 1982)**Syllis cruzi Nunez and San Martin, 1991*Syllis ferrani Alos and San Martin, 1987Syllis garciai Campoy, 1982**Syllis gerundensis (Alos and Campoy, 1981)Syllis gracilis Grube, 1840Syllis hyalina Grube, 1863*Syllis jorgei San Martin and Lopez, 2000Syllis krohniEhlers, 1864Syllis prolifera (Krohn, 1852)*Syllis pulvinata Langerhans, 1881*Syllis rosea Langerhans, 1879Syllis truncata cryptica Ben-Eliahu, 1977Syllis variegata Grube, 1860*Syllis vivipara Krohn, 1869*Syllis westheidei San Martin, 1984*Trypanosyllis coeliaca Claparede, 1868*Trypanosyllis zebra (Grube, 1860)Xenosyllis scabra (Ehlers, 1864)NereididaeCeratonereis costae (Grube, 1840)Micronereissp.*Nereis flavipes Ehlers, 1868Nereis rava Ehlers, 1868Nereis zonata Malmgren, 1867Nereissp.Perinereis cultrifera (Grube, 1840)Platynereis dumerilii (Audouin and M. Edwards, 1833)AphroditidaePontogenia chrysocoma (Baird, 1865)PolynoidaeHarmothoe sp.Lepidonotus clava(Montagu, 1808)ChrysopetalidaeChrysopetalum debile(Grube, 1855)Eunicidae*Eunice aphroditois (Pallas, 1788)Eunice harassiiAudouin and M. Edwards, 1834*Eunice oerstedii Stimpson, 1854Eunice pennata(Muller, 1778)Eunice torquataQuatrefages, 1865)Eunice vittata (Delle Chiaje, 1828)*Lysibranchia paucibranchiata Cantone, 1983*Lysidice collaris Grube, 1870Lysidice ninettaAudouin and M. Edwards, 1833Nematonereis unicornis Schmarda, 1861Palola siciliensis (Grube, 1840)Lumbrineridae*Lumbrineriopsis paradoxa (Saint-Joseph, 1888)Lumbrineris coccinea (Renier, 1804)Lumbrineris funchalensis (Kimberg, 1865)Lumbrineris gracilis (Ehlers, 1868)Lumbrineris tetraura (Schmarda, 1861)DorvilleidaeDorvillea rubrovittata (Grube, 1855)Flabelligeridae*Piromis eruca (Claparede, 1870)TrichobranchiidaeTerebellides sp.TerebellidaeAmphitrite cirrata Muller, 1771Neoamphitrite rubra (Risso, 1828)
Table 1 (Contd.)
Spionidae
Pista sp.Polycirrussp.*Streblosoma hesslei (Day, 1955)Thelepus triserialis (Grube, 1855)SabellariidaeSabellaria alveolata Linneus, 1767Sabellidae*Amphicorina grahamensis Giangrande et al. 1999Amphiglena mediterranea (Leydig, 1851)Branchiomma bombyx (Dalyell, 1853)Branchiomma lucullanum (Delle Chiaje, 1828)Chone collaris Langerhans, 1880Demonax brachychona (Claparede, 1870)Demonax langerhansi Knight-Jones, 1983Demonax tommasi Giangrande, 1994Fabricia stellaris (Muller, 1774)Hypsicomus stichophthalmos (Grube, 1863)Hypsicomussp.Jasmineira elegans Saint-Joseph, 1884*Laonome salmacidis Claparede, 1868Megalommasp.Perkinsiana rubra (Langerhans, 1880)Pseudobranchiommasp.Pseudopotamilla reniformis (Bruguiere, 1789)SerpulidaeFilogranasp.Hydroides elegans(Haswell, 1883)Hydroides helmatus(Iroso, 1921)Hydroides niger Zibrowius, 1971Hydroides pseudouncinatus Zibrowius, 1968*Pomatoceros lamarckii (Quatrefages, 1865)Pomatoceros triqueter (Linnaeus, 1767)Protula sp.*Semivermilia pomatostegoides (Zibrowius, 1969)Serpula concharum Langerhans, 1880Serpula vermicularis Linnaeus, 1767Spirobranchus polytrema (Philippi, 1844)Vermiliopsis infundibulum (Philippi, 1844)Vermiliopsis labiata (Costa, 1961)Vermiliopsis striaticeps (Grube, 1862)
1112
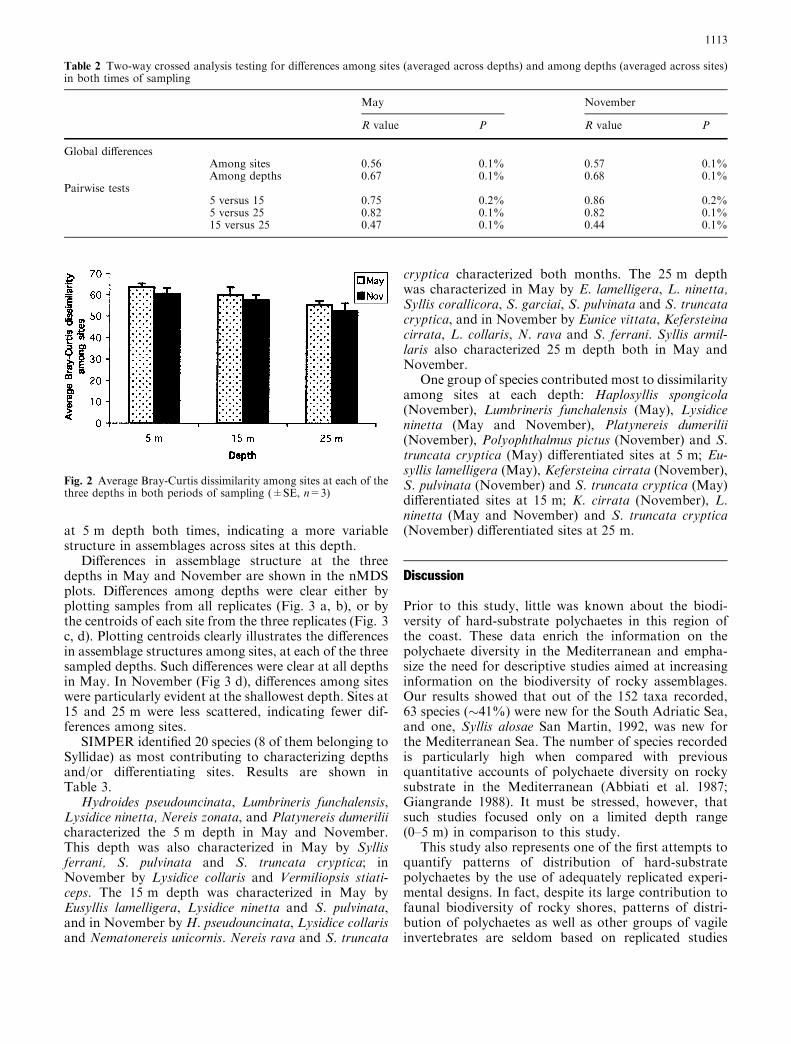
at 5 m depth both times, indicating a more variablestructure in assemblages across sites at this depth.
Differences in assemblage structure at the threedepths in May and November are shown in the nMDSplots. Differences among depths were clear either byplotting samples from all replicates (Fig. 3 a, b), or bythe centroids of each site from the three replicates (Fig. 3c, d). Plotting centroids clearly illustrates the differencesin assemblage structures among sites, at each of the threesampled depths. Such differences were clear at all depthsin May. In November (Fig 3 d), differences among siteswere particularly evident at the shallowest depth. Sites at15 and 25 m were less scattered, indicating fewer dif-ferences among sites.
SIMPER identified 20 species (8 of them belonging toSyllidae) as most contributing to characterizing depthsand/or differentiating sites. Results are shown inTable 3.
Hydroides pseudouncinata, Lumbrineris funchalensis,Lysidice ninetta, Nereis zonata, and Platynereis dumeriliicharacterized the 5 m depth in May and November.This depth was also characterized in May by Syllisferrani, S. pulvinata and S. truncata cryptica; inNovember by Lysidice collaris and Vermiliopsis stiati-ceps. The 15 m depth was characterized in May byEusyllis lamelligera, Lysidice ninetta and S. pulvinata,and in November by H. pseudouncinata, Lysidice collarisand Nematonereis unicornis. Nereis rava and S. truncata
cryptica characterized both months. The 25 m depthwas characterized in May by E. lamelligera, L. ninetta,Syllis corallicora, S. garciai, S. pulvinata and S. truncatacryptica, and in November by Eunice vittata, Kefersteinacirrata, L. collaris, N. rava and S. ferrani. Syllis armil-laris also characterized 25 m depth both in May andNovember.
One group of species contributed most to dissimilarityamong sites at each depth: Haplosyllis spongicola(November), Lumbrineris funchalensis (May), Lysidiceninetta (May and November), Platynereis dumerilii(November), Polyophthalmus pictus (November) and S.truncata cryptica (May) differentiated sites at 5 m; Eu-syllis lamelligera (May), Kefersteina cirrata (November),S. pulvinata (November) and S. truncata cryptica (May)differentiated sites at 15 m; K. cirrata (November), L.ninetta (May and November) and S. truncata cryptica(November) differentiated sites at 25 m.
Discussion
Prior to this study, little was known about the biodi-versity of hard-substrate polychaetes in this region ofthe coast. These data enrich the information on thepolychaete diversity in the Mediterranean and empha-size the need for descriptive studies aimed at increasinginformation on the biodiversity of rocky assemblages.Our results showed that out of the 152 taxa recorded,63 species (�41%) were new for the South Adriatic Sea,and one, Syllis alosae San Martin, 1992, was new forthe Mediterranean Sea. The number of species recordedis particularly high when compared with previousquantitative accounts of polychaete diversity on rockysubstrate in the Mediterranean (Abbiati et al. 1987;Giangrande 1988). It must be stressed, however, thatsuch studies focused only on a limited depth range(0–5 m) in comparison to this study.
This study also represents one of the first attempts toquantify patterns of distribution of hard-substratepolychaetes by the use of adequately replicated experi-mental designs. In fact, despite its large contribution tofaunal biodiversity of rocky shores, patterns of distri-bution of polychaetes as well as other groups of vagileinvertebrates are seldom based on replicated studies
Table 2 Two-way crossed analysis testing for differences among sites (averaged across depths) and among depths (averaged across sites)in both times of sampling
May November
R value P R value P
Global differencesAmong sites 0.56 0.1% 0.57 0.1%Among depths 0.67 0.1% 0.68 0.1%
Pairwise tests5 versus 15 0.75 0.2% 0.86 0.2%5 versus 25 0.82 0.1% 0.82 0.1%15 versus 25 0.47 0.1% 0.44 0.1%
Fig. 2 Average Bray-Curtis dissimilarity among sites at each of thethree depths in both periods of sampling (±SE, n=3)
1113

(Kelaher et al. 2001; Olabarria and Chapman 2001;Fraschetti et al. 2002; Terlizzi et al. 2003). A possibleexplanation is that working with vagile invertebratesrequires time-consuming studies in terms of sortingoperations and species identification. Moreover, theexpertise required for the identification of organisms atthe species level is becoming rarer in ecological studies(Boero 2001; Giangrande 2003). This results in moststudies on rocky substrate assemblages being under-taken on sessile organisms, easily recognizable in situand/or by photographic records.
A main result from the present study was that poly-chaete assemblages differed between depths, with a clearpattern of zonation from the shallower to the deepeststations. The greatest differences were, however,observed between the shallowest sites (5 m depth) andthe remaining ones (15 and 25 m depths). Changes inpolychaete assemblages with depth were mainly due toquantitative changes in the assemblage structure alongthe vertical gradient, with increasing abundance of somespecies such as Eusyllis lamelligera, Kefersteinia cirrata,Nereis rava, and Syllis armillaris, and decreasing num-bers of some others, such as Hydroides pseudouncinata,Lumbrineris funchalensis, Lysidice ninetta, and Nereiszonata. Among the characterizing species, onlyPlatynereis dumerilii was found exclusively at 5 m depth,whilst Eunice vittata, Syllis corallicola and S. garciaiwere not present at this depth. This pattern was con-sistent with time despite differences in abundance valuesobserved between the two times of sampling.
Overall, the vast majority of the species collectedbelow 5 m are known as typical of sciaphilous envi-ronments (Sarda 1991). Conversely, the finding of P.dumerilii as strongly characterizing the shallow sites(5 m) confirms the species as typical of shallow infra-littoral photophilic environments (Giangrande 1988;Giangrande et al. 2002).
The knowledge of Mediterranean polychaete assem-blages of coralligenous and sciaphilous environments isvery scant and mainly limited to qualitative studies(Bellan 1961; Laubier 1966; Sarda 1991). As a conse-quence, there is no information available from previousstudies to allow comparisons with our results.
In addition to changes occurring along the depthgradient, multivariate analyses also revealed significantdifferences among sites at each depth, indicating thatbesides differences along the depth gradient, a horizontalvariability in the structure of the polychaete assemblagesalso exists at each level of the subtidal, particularly at5 m depth. Therefore, as already observed in similarstudies conducted in different habitats, there are othersources of variability, including differences among siteswithin any particular level of the subtidal and temporalchanges in patterns of distribution, that need to beproperly quantified before any conclusion about thevertical distribution of hard-substrate invertebrate vagileassemblages can be made (Benedetti-Cecchi et al. 1996;Underwood 2000).
Within the framework of the same monitoringprogram, along the same stretch of coast and at thesame sampling occasion, Terlizzi et al. (2003) observedcomparable patterns of along and across subtidalspatial distribution for molluscs. Molluscan assem-blages differed among depths, with a clear pattern ofzonation from the shallower to the deepest stations,and considerable differences in the structure of theassemblages were also observed among sites at eachdepth range.
It is obviously not possible to interpret the results interms of causal mechanisms leading to the observedpattern, because data are merely descriptive. Moreover,such mechanisms could vary depending on the differenttaxonomic groups (e.g. molluscs vs polychaetes). How-ever, the quantitative description of patterns is the first,
Fig. 3 Two-factor nMDS plotshowing differences among sitesand among depths in bothperiods of sampling (a, c May,b, d November). In a and b eachpoint represents the assemblagesfrom the three replicates. In cand d each point represents thecentroid of assemblages fromthe three replicates (white 5 m,grey 15 m, black 25 m)(s;u;M= sites 1, 2 and 3,respectively)
1114

crucial step before any model regarding the ecologicalfactors triggering the observed patterns can be proposedand tested (Underwood et al. 2000).
Some potential causes for the differences observed inpolychaete assemblages can be hypothesized.
One possible model is linked to the features of sub-strate covering by algae. The architectural complexity ofalgal assemblages has been demonstrated to influence thestructure of invertebrate vagile assemblages (Abbiatiet al. 1987; Giangrande 1988; Chemello and Milazzo2002). In our study location (see Materials and meth-ods), a shift from photophilic algal assemblages domi-nated by erected and fleshy species towards sciaphilousalgal assemblages dominated by encrusting calcifiedalgae is present at about 15 m depth. Such differences insubstrate complexity can be hypothesized as responsiblefor the differences observed and could justify the greatersimilarities between the 15 m and 25 m assemblages thanthose at 5 m. Similar causes could explain differencesamong sites within each depth. Patchiness in sessileassemblages (Terlizzi et al., unpublished data) on a scaleof 10s of meters, in particular, has been recently quan-tified and demonstrated to be higher at 5 m than below.
Clearly, the processes producing changes in poly-chaete assemblage on subtidal cliffs are likely to becomplex and require detailed experimental inspection.Other models (e.g. the influence of predation, competi-tive interaction among resident species, food resourcesand recruits supply) remain to be explored beforeinvoking algal complexities as the only possible factorscontrolling the structure of polychaete assemblages.Such models will be analysed in detail in future manip-
ulative experiments aimed at highlighting the processesinfluencing the structure of invertebrate vagile assem-blages on rocky substrates.
Acknowledgements Research funded by the Regione Puglia(INTERREG Albany-Italy project), and MURST (COFIN pro-ject). C. Vaglio provided invaluable assistance during field-work.
References
Abbiati M, Bianchi CN, Castelli A (1987) Polychaete verticalzonation along a littoral cliff in the west Mediterranean. PSZN:Mar Ecol 8:33–48
Alos C (1999) Anelidos poliquetos del Cabo de Creus (NE de Es-pana). Faces de Corallina elongata Ellis & Solander y de Cy-stoseira mediterranea (J. Feldmann). Misc Zool 14:17–27
Bellan G (1961) Polychetes de la campagne 1956 du ‘Gyf’ dansle Nord de la Corse (Corse). Rec Trav St Mar Endoume23:71–83
Bellan G (1969) Polychetes des horizon superieurs de l’etage in-fralittoral rocheux dans la region provencale. Tethys 1:349–366
Bellan G (1971) Etude sommaire des polychetes des horizonssuperieurs de l’etage infralittoral sur substrat dur dans la regionprovencale. Rapp Comm Int Mer Medit 20:217–219
Benedetti-Cecchi L (2000) Priority effects, taxonomic resolution,and the prediction of variable patterns of succession in littoralrock pools. Oecologia 123:265–274
Benedetti-Cecchi L (2001). Variability in abundance of algae andinvertebrates at different spatial scales on rocky sea shores. MarEcol Prog Ser 215:79–92
Benedetti-Cecchi L, Airoldi L, Abbiati M, Cinelli F (1996) Esti-mating the abundance of benthic invertebrates: a comparison ofprocedures and variability between observers. Mar Ecol ProgSer 138:93–101
Bertness MD, Gaines SD, Bermudez D, Sanford E (1991) Extremespatial variation in the growth and reproduction output of the
Table 3 Characterization of depths: species most contributing to the Bray-Curtis similarity within each depth range (averaged across sites).Values of similarity ‡5% in bold.Differentiation among sites: species most contributing to dissimilarity among sites at each depth.Dissimilarity ‡3% in bold
Species Characterization of depths Differentiation among sites
5 m 15 m 25 m 5 m 15 m 25 m
May Nov May Nov May Nov May Nov May Nov May Nov
Eunice vittata 0.0 0.0 0.3 0.0 0.4 8.8 0.0 0.0 0.9 1.9 1.8 2.2Eusyllis lamelligera 1.9 1.1 7.10 1.0 6.76 2.0 2.7 2.8 3.5 2.0 2.1 2.1Haplosyllis spongicola 3.3 3.0 2.0 1.6 2.2 2.2 2.2 4.1 1.9 2.5 1.7 2.2Hydroides pseudouncinata 8.39 7.2 3.6 5.3 3.4 2.6 2.0 2.1 2.1 1.8 2.0 1.7Kefersteinia cirrata 1.9 0.2 4.0 3.7 4.9 6.4 1.7 1.0 2.6 4.2 2.2 3.5Lumbrinereis funchalensis 7.51 8.7 3.8 1.0 0.1 4.7 3.7 2.6 1.5 1.8 1.0 1.4Lysidice collaris 0.2 9.6 1.2 13.0 0.3 6.8 1.1 2.1 2.5 1.7 1.9 2.7Lysidice ninetta 12.73 6.8 11.18 2.8 5.72 2.7 3.1 3.0 2.6 2.9 3.2 3.2Nematonereis unicornis 0.8 1.3 4.9 5.1 0.8 1.4 1.9 2.2 1.5 1.5 1.6 2.0Nereis rava 0.1 3.5 5.63 6.8 3.9 8.8 1.0 1.9 1.7 1.5 3.1 1.5Nereis zonata 11.18 6.1 1.9 2.3 1.5 0.5 1.4 2.5 2.6 2.8 1.7 1.7Platynereis dumerilii 6.61 11.2 0.0 0.0 0.0 0.0 2.5 3.6 0.7 0.4 0.8 0.5Polyophthalmus pictus 0.0 1.4 0.3 0.7 0.0 1.6 0.7 3.4 0.6 2.2 0.0 2.8Syllis armillaris 1.8 0.0 3.5 1.8 8.13 5.8 1.4 0.6 2.5 2.4 1.4 2.5Syllis golfonovoensis 0.0 0.0 1.8 2.6 5.72 3.8 0.5 0.0 1.2 1.9 1.7 1.9Syllis ferrani 5.76 3.6 1.8 3.8 2.2 5.5 1.7 1.7 1.8 2.0 2.3 1.1Syllis garciai 0.0 0.0 0.9 0.0 5.46 0.7 0.0 0.0 1.1 0.0 1.7 1.0Syllis pulvinata 5.0 4.5 7.56 3.4 5.92 1.6 2.9 1.9 1.6 3.0 1.8 2.5Syllis truncata cryptica 5.84 3.6 9.59 9.1 10.41 3.1 3.1 2.8 3.0 1.2 2.2 4.0Vermiliopsis striaticeps 2.5 5.1 1.5 1.5 4.8 1.0 2.5 2.9 1.4 2.1 0.9 0.0
1115

acorn barnacle Semibalanus balanoides. Mar Ecol Prog Ser75:91–100
Boero F (2001) Light after dark: the partnership for enhancingexpertise in taxonomy. Trends Ecol Evol 16:266
Caffey HM (1985) Spatial and temporal variation in settlement andrecruitment of intertidal barnacles. Ecol Monogr 55:313–332
Cantone G (1983) Un nuovo genere di Eunicidae (Annellida:Polycaeta) del golfo di Catania. Animalia 10:81–86
Castelli A, Abbiati M, Badalamenti F, Bianchi CN, Cantone G,Gambi MC, Giangrande A, Gravina MF, Lanera P, Lardicci C,Somaschini A, Sordino P (1995) Annelida Polychaeta, Pogono-phora, Echiura, Sipuncula. In: Minelli A, Ruffo S, La Posta S(eds) Checklist delle specie della fauna italiana 19. Calderini,Bologna
Chapman MG, Underwood AJ, Skilleter GA (1995) Variability atdifferent spatial scales between a subtidal assemblage exposedto the discharge of sewage and two control assemblages. J ExpMar Biol Ecol 189:103–122
Chemello R, Milazzo M (2002) Effect of algal architecture onassociated fauna: some evidence from phytal molluscs. MarBiol 140:981–990
Clarke KR (1993) Non-parametric multivariate analysis of changesin community structure. Aust J Ecol 18:117–143
Connell HV, Karlson RH (1996) Species richness of reef-buildingcorals determined by local and regional processes. J Anim Ecol65:233–241
Ferdeghini F, Acunto S, Cocito S, Cinelli F (2000) Variability atdifferent spatial scales of a coralligenous assemblage at Gian-nutri Island (Tuscan Archipelago, northwestern Mediterra-nean). Hydrobiologia 440:27–36
Fraschetti S, Bianchi CN, Terlizzi A, Fanelli G, Morri C, Boero F(2001) Spatial variability and human disturbance in shallowsubtidal hard substrate assemblages: a regional approach. MarEcol Prog Ser 212:1–12
Fraschetti S, Giangrande A, Terlizzi A, Della Tommasa L, Migli-etta MP, Boero F (2002) Spatio-temporal variation of hydroidsand polychaetes associated with Cystoseira amentacea (Fucales,Phaeophyceae). Mar Biol 140:949–957
Fresi E, Colognola R, Gambi MC, Giangrande A, Scardi M(1983) Ricerche sui popolamenti bentonici di substrato durodel Porto di Ischia. Infralitorale fotofilo: Policheti. Cah BiolMar 24:1–19
Fresi E, Colognola R, Gambi MC, Giangrande A, Scardi M (1984)Ricerche sui popolamenti bentonice di substrato duro del Portodi Ischia. Infralitorale fotofilo: Policheti (II). Cah Biol Mar25:33–47
Giangrande A (1988) Polychaete zonation and its relation to algaldistribution down a vertical cliff in the western Mediterranean(Italy): a structural analysis. J Exp Mar Biol Ecol 120:263–276
Giangrande A (2003) Biodiversity, conservation and the taxonomicimpediment. Aquatic Conserv: Mar Freshw Ecosyst (in press)
Giangrande A, Fraschetti S, Terlizzi A (2002) Local recruitmentdifferences in Platynereis dumerilii (Polychaeta, Nereididae) andtheir consequences for population structure. Ital J Zool 69:133–139
Guichard F, Bourget E (1998) Topographic heterogeneity, hydro-dynamics, and benthic community structure: a scale-dependentcascade. Mar Ecol Prog Ser 171:59–70
Katzmann W (1971) Polychaeten (Errantier, Sedentatier) ausnordadriatischen Cystoseira-Bestanden und derer Epiphyten.Oecologia 8:31–35
Kelaher BP, Chapman MG, Underwood AJ (2001). Spatial pat-terns of diverse macrofaunal assemblages in coralline turf andtheir associations with environmental variables. J Mar BiolAssoc UK 81:917–930
Kennelly SJ, Underwood AJ (1993) Geographic consistencies ofeffects of experimental physical disturbance on understoryspecies in sublittoral kelp forests in central New South Wales.J Exp Mar Biol Ecol 168:35–58
Knight-Jones P, Giangrande A (2003) Two new species of anatypical group of Pseudobranchiomma Jones (Polychaeta: Sa-bellidae). Hydrobiologia (in press)
Laubier L (1966) Le coralligene des Alberes. Monographie bio-cenotique Ann Inst Oceanogr Paris 43:137–316
Licher F (1999) Revision der Gattung Typosyllis Langherans, 1879(Polychaeta: Syllidae). Morphologie, taxonomic un Phylogenic.Abh Senckenb Naturforsch Ges 551:1–336
Lopez E, Vieitez JM (1999) Polychaete assemblages on non-encrusting infralittoral algae from the Chafarinas Islands (SWMediterranean). Cah Biol Mar 40:375–384
Lopez E, San Martin G, Jimenez M (1996) Sylline (Syllidae,Anellida, Polychaeta) from Chafarinas Island (Alboran Sea, WMediterranean). Misc Zool 19:105–118
Menconi M, Benedetti-Cecchi L, Cinelli F (1999) Spatial andtemporal variability in the distribution of algae and inverte-brates on rocky shores in the northwest Mediterranean. J ExpMar Biol Ecol 233:1–23
Nunez J, San Martin G, Acero C (1992) Syllinae (Polychaeta,Syllidae) de las Islas Canarias. Rev Acad Canr Cinc 4:109–129
Olabarria C, Chapman MG (2001) Comparison of patterns ofspatial variation of microgastropods between two contrastingintertidal habitats. Mar Ecol Prog Ser 220:201–211
Peres JM (1982) Major benthic assemblages. In: Kinne OJ (ed)Marine ecology, vol 5. Wiley, London, pp 373–508
Riedl R (1971) Water movement: animals. In: Kinne OJ (ed)Marine ecology, vol 5. Wiley, London, pp1123–1156
Roberts DE (1996) Patterns in subtidal marine assemblages asso-ciated with a deep-water sewage outfall. Mar Freshw Res 47:1–9
Ros JD, Romero J, Ballestreros E, Gili JM (1985) Diving in bluewater. The benthos. In: Margalef R (ed) Western Mediterra-nean. Pergamon , Oxford, pp. 233–295
San Martin G (1984) Estudio biogeografico, faunistico y sistema-tico de los poliquetos de la famiglia Silidos (Syllidae: Policha-eta) en Baleares. Pubbl Univ Coplutense de Madrid
Sara M (1969) Research on coralligenous formations: problemsand perspectives. PSZN: Mar Ecol 37:124–134
Sarda R (1991) Polychaete communities related to plant covering inthe mediolittoral and infralittoral zones of the Balearic Islands(Western Mediterranean). PSZN: Mar Ecol 12:341–360
Smith F, Witman JD (1999) Species diversity in subtidal land-scapes: maintenance by physical processes and larval recruit-ment. Ecology 80:51–69
Somaschini A (1988) Policheti della biocenosi ad alghe fotofile(facies a Corallina elongata) nel Lazio settentrionale. Atti SocTosc Sci Nat Mem Ser B 95:83–94
Tena J, Capaccioni-Azzati R, Torres-Gavila FJ, Garcia-CarrascosaAM (2000) Polychaetes associated with different facies of thephotophilic algal community in the Chafarinas archipelago (SWMediterranean). Bull Mar Sci 67:55–72
Terlizzi A, Scuderi D, Fraschetti S, Guidetti P, Boero F (2003)Molluscs on subtidal cliffs: patterns of spatial distribution.J Mar Biol Assoc UK 83:165–172
Underwood AJ (2000) Experimental ecology of rocky intertidalhabitats. What are we learning? J Exp Mar Biol Ecol 250:51–76
Underwood AJ, Chapman MG (1996) Scales of spatial patterns ofdistribution of intertidal invertebrates. Oecologia 107:212–224
Underwood AJ, Chapman MG (1998a) Spatial analyses of inter-tidal assemblages on sheltered rocky shores. Aust J Ecol23:138–157
Underwood AJ, Chapman MG (1998b) Variation in algal assem-blages on wave-exposed rocky shores in New South Wales. MarFreshw Res 49:241–254
Underwood AJ, Chapman MG, Connell SD (2000) Observations inecology: you can’t make progress on processes without under-standing the patterns. Exp Mar Biol Ecol 250:97–115
1116
Related Documents











