This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Toxicology Letters 193 (2010) 209–216
Contents lists available at ScienceDirect
Toxicology Letters
journa l homepage: www.e lsev ier .com/ locate / tox le t
PM10 impairs the antioxidant defense system and exacerbates oxidative stressdriven cell death
Yolanda I. Chirinoa,e,1, Yesennia Sánchez-Péreza,1, Álvaro R. Osornio-Vargasa,Rocío Morales-Bárcenasa, María Concepción Gutiérrez-Ruízb, Yazmin Segura-Garcíaa,Irma Rosasc, José Pedraza-Chaverrid, Claudia M. García-Cuellara,∗
a Instituto Nacional de Cancerología (INCan), Subdirección de Investigación Básica, San Fernando No. 22, Tlalpan, 14080 Mexico D.F., Mexicob Departamento de Ciencias de la Salud, División de Ciencias Biológicas y de la Salud (DCBS), Universidad Autónoma Metropolitana-Iztapalapa (UAM-Iztapalapa),09340 Mexico D.F., Mexicoc Centro de Ciencias de la Atmósfera, Universidad Nacional Autónoma de México (UNAM), 04510 Mexico D.F., Mexicod Facultad de Química, Departamento de Biología, Universidad Nacional Autónoma de México (UNAM), 04510 Mexico D.F., Mexicoe Unidad de Biomedicina, Facultad de Estudios Superiores Iztacala, Av. De Los Barrios No.1, Los Reyes Iztacala, 54090, Estado de México, Mexico
a r t i c l e i n f o
Article history:Received 20 October 2009Received in revised form 11 January 2010Accepted 13 January 2010Available online 22 January 2010
Keywords:PM10
Antioxidant enzymatic defenseOxidative stress
a b s t r a c t
The aim of this study was to investigate the effect of airborne particulate matter with a mean aerody-namic diameter of ≤10 �m (PM10) on oxidative stress markers and antioxidant enzymatic activity andits relevance in the face of acute oxidative challenge in a human lung epithelial cell line (A549). PM10-induced reactive oxygen species (ROS) generation and oxidative damage with no changes in cellularviability. In addition, PM10 decreased glutathione (GSH) levels (54.9%) and the activity of the antioxidantenzymes superoxide dismutase (65%), catalase (31.2%), glutathione reductase (61.5%) and glutathione-S-transferase (42.39%). Trolox, a scavenger of reactive species, prevented the increase of ROS generationand the decrease in GSH levels but partially prevented PM10-induced oxidative damage. Interestingly, itwas unable to avoid the decrease in the activity of antioxidant enzymes. Finally, the survival of the cellspreviously exposed to PM10 and challenged with hydrogen peroxide was significantly lower. We con-clude that the impairment in the antioxidant defense system induced by PM10 weaken ROS detoxificationwhich exacerbates cell death when these cells are exposed to an acute oxidative challenge.
© 2010 Elsevier Ireland Ltd. All rights reserved.
1. Introduction
The epidemiological evidence of potential adverse health effectsassociated with exposure to airborne particulate matter with amean aerodynamic diameter of ≤10 �m (PM10) is increasing (Popeet al., 2002). In fact it has been associated with cardiovascular(Nurkiewicz et al., 2006; Miller et al., 2009; Wang et al., in press)and pulmonary diseases (Liu et al., 2009) chronic obstructive pul-monary disease (Churg et al., 2003; Lagorio et al., 2006; Sint etal., 2008) and cancer (Pope et al., 2002; Brunekreef et al., 2009).Interestingly, the oxidative stress seems to be a common featureof PM10 exposure (Nurkiewicz et al., 2006; Churg et al., 2003;Sanchez-Perez et al., 2009) and free radical generation by PM10is responsible from the important inflammatory effects in vitro
∗ Corresponding author. Tel.: +52 55 5628 0462; fax: +52 55 5628 0432.E-mail addresses: [email protected], [email protected]
(C.M. García-Cuellar).1 These authors contributed equally to this work.
and in vivo models (Li et al., 1996). Some oxidative stress markershave been used, including: reactive oxygen species (ROS) (Sanchez-Perez et al., 2009), myeloperoxidase expression (Nurkiewicz etal., 2006), TNF-� release (Brown et al., 2004), and DNA adducts(Valavanidis et al., 2005), among others, in order to explain theoxidative stress mechanism induced by PM10. The oxidative stresshas been attributed to PM10 components including diesel exhaustparticles (DEP) (Li et al., 2008) and ambient ultrafine particles (UFP)(Araujo et al., 2008). The observed effects are attributed, at least inpart, to the large reactive surface area of PM10 and its deposition inthe lung and to some organic compounds able to diffuse from par-ticle surface into cells (Calderon-Garciduenas et al., 2001; Nemmaret al., 2004; Oberdorster et al., 2004), for the reason that not all thePM10 components are internalized into the cell, such as endotox-ins, which execute their effects through the stimulation of Toll-likereceptor 4 (Becker et al., 2002). In addition, the ROS formation hasbeen attributed to metals contained in PM10, including iron. Iron isa transition metal that catalyzes the production of hydroxyl radi-cal (OH•) by the Fenton reaction contributing to lipid peroxidation(Winterbourn, 1995; Aust et al., 2002). On the other hand, PM10
0378-4274/$ – see front matter © 2010 Elsevier Ireland Ltd. All rights reserved.doi:10.1016/j.toxlet.2010.01.009

Author's personal copy
210 Y.I. Chirino et al. / Toxicology Letters 193 (2010) 209–216
directly interacts with antioxidant enzymes causing lost of enzy-matic activity in a cell-free system (Hatzis et al., 2006) but remainsunclear if the same effects will be observed in a cellular system andif ROS is directly involved in enzymatic activity loss. The aim of thisstudy was to investigate the effect of PM10 on oxidative stress mark-ers and antioxidant enzymatic activity under sub-lethal conditionsand search its relevance in the face of acute oxidative challenge ina human lung epithelial cell line. In addition, we investigated if thePM10-induced changes were related to the decrease in the antioxi-dant defense; in this study trolox (Tlx) was used, as an antioxidantcompound. This work clearly shows that PM10 induces oxidativestress without affecting cell viability, but the cells were unable tomaintain its integrity when they were exposed to a second oxida-tive challenge using hydrogen peroxide (H2O2), which is a centralmolecule in the inflammatory lung development (Cook-Mills, 2006;Dekhuijzen et al., 1996; Morissette et al., 2008; Ryter et al., 2007;Song et al., 2009).
2. Materials and methods
2.1. PM10 sampling
PM10 was collected in a commercial zone (with predominately traffic sources) ofMexico City using a high-volume particle collector with a flux of 1.13 m3/min (GMWmodel 1 200 VFC HVPM10; Sierra Andersen, Smyrna, GA, USA). PM10 was recover on3 �m pore size cellulose nitrate filters (Sartorius AG, Goettingen, Germany), 3 daysa week. Particles were scraped from the filters using an electric toothbrush (Oral-B,AdvancePower 400, Braun, Mexico) as has been previously reported (Alfaro-Morenoet al., 2009). The particles obtained from one year were stored in endotoxin-free glassvials in a dryer at 4 ◦C until their use as previously done by our group (Alfaro-Morenoet al., 2009). Filters were maintained in the darkness at 4 ◦C in a desiccator prior toparticle removal. Particles were gently scraped off the membranes with a surgicalblade into endotoxin-free glass vials, maintained in the dark in a dryer at 4 ◦C untiluse (Alfaro-Moreno et al., 2009).
2.2. Cell culture and PM10/Tlx treatment
Lung epithelial cell line (A549) was obtained from the American Type CultureCollection (ATCC, Rockville, MD, USA). Cells were cultured in F12K medium (Invit-rogen, Carlsbard, CA, USA) supplemented with 10% fetal bovine serum in a 5% CO2
atmosphere at 37 ◦C. All cell exposure experiments were performed at 80% of cellconfluence. Once the cell reached confluence, the culture medium was replaced withserum-free medium and cells were exposed 24 h to PM10 or PM10 plus 10 �M of Tlx.In order to investigate if the PM10-induced changes were related to the decrease inthe antioxidant defense, we used Tlx, as an antioxidant compound, in this study.One milligram of PM10 were suspended in 1 mL of F12K medium and added to1 × 106 A549 cells which were seeded in 10 cm2 well, and then the final PM10 con-centration was 10 �g/cm2 (Sanchez-Perez et al., 2009; Osornio-Vargas et al., 2003;Alfaro-Moreno et al., 2002). Free-serum medium was used in order to avoid possiblemetal chelation from albumin contained in the serum, affecting results due to PM10
metal content (Moriya et al., 2008).
2.3. ROS production
2′ ,7′-Dichlorohydrofluorescein diacetate (DCFH-DA) (Invitrogen, Carlsbard, CA,USA) was used as an intracellular ROS indicator (Lawler et al., 2003). Cells weregrown to 80% confluence on 96-well tissue culture plates and exposed to PM10
(5 �g/cm2, 10 �g/cm2 and 20 �g/cm2) or PM10 plus Tlx in free-serum medium. Greenfluorescence (oxidized DCFH) by oxidant presence was quantified at 1 h, 3 h, 6 h, 12 hand 24 h of incubation at 37 ◦C using a microplate fluorometer with a 480 nm excita-tion and 515 nm emission filters cell-free wells containing only particles and DCFHwere used to assess non-specific particle-induced DCFH oxidation. Fluorescence wasreported as fluorescence units (FU) per million cells (1 × 106).
2.4. Cytotoxicity assay
Cytotoxicity was measured employing the crystal violet method of Kueng et al.(1989). To evaluate cytotoxicity, proliferating A549 cells were exposed to 5 �g/cm2,10 �g/cm2 and 20 �g/cm2 of PM10 and/or antioxidant (10 �M Tlx). After 24 h ofexposure the cytotoxicity was determined by counting residual cell numbers withcrystal violet staining using a plate reader at an optical density at 570 nm. The resultwas reported as viability with respect to control. After exposure, the cells wereobserved under a microscope to verify even PM10 distribution on cells. Images wereobtained with a Nikon Coolpix 4500 camera.
2.5. Glutathione determination
The reduced glutathione (GSH) was evaluated with Glutathione Assay Kit II (Cal-biochem Inc., San Diego, CA) according to details provided by the manufacturer. ThePM10 exposed cells were homogenized in cold buffer (50 mM MES, pH 7.0, contain-ing 1 mM EDTA) and the samples were deproteinated. GSH was quantified using amicroplate fluorometer with filters at 405 nm based on the GSH standard curve. GSHwas reported as �mol of GSH/1 × 106 cells.
2.6. Lipid peroxidation
Lipid peroxidation was measure using Lipid Peroxidase Assay Kit (CalbiochemInc., San Diego, CA), was used to evaluate malondialdehyde (MDA). After exposure,the cells were lysed in a buffer containing 10 mM Tris, 10 mM phenylmethane-sulphonylfluoride, and 150 mM NaCl, pH 7.4. The cell lysate was reacted withN-methy-2-phenylindole and this mixture was incubated with methanesulfonicacid at 45 ◦C for 60 min. MDA content was measured at 586 nm by comparison withthe predetermined MDA standard curve. The results are the average of three inde-pendents experiments and the results are reported as �mol of MDA per milligramof protein.
2.7. Protein oxidation
Protein oxidation was determined by quantifying protein carbonyl groups usingthe Oxyblot kit (Chemicon, Temecula, CA, USA). The cells were lysed in a buffer con-taining 10 mM Tris, 10 mM PMSF, and 150 mM NaCl, pH 7.4. Fifteen micrograms ofprotein were derivatized with dinitrophenylhydrazine (DNPH). After neutralization,the proteins derivates were electrophoresed in 12% (w/v) Bis–Tris gels and trans-ferred onto nitrocellulose. The membrane was blocked in 1% (w/v) bovine serumalbumin (BSA) in PBS-Tween (PBS-T) for 1 h, followed by incubation with anti-DNPantibody [1:100 dilution in PBS-T containing 1% (w/v) BSA], after one hour, the mem-brane was incubated with secondary antibody (horseradish peroxidase-conjugatedanti-rabbit, 1:300 dilution in PBS-T). Blots were developed with a chemilumines-cence detection system (Millipore Corporation, Billerica, MA, USA). Protein oxidationlevel was evaluated through densitometry using the Image J freeware program. Con-trol was normalized to 1 unit and the changes in densitometry were expressed asfold increase relative to control.
2.8. Determination of 8-Oxoguanine
The 8-Oxoguanine (8-OxoG) adducts were measured using the OxyDNA AssayKit, Fluorogenic (Calbiochem Inc, San Diego, CA). The procedure was performedaccording to manufacturer’s instructions. After exposure, the cells were fixed in4% (v/v) formaldehyde at 4 ◦C and dehydrated in 70% (v/v) methanol followed by95% (v/v) methanol. The cells were permeabilized and rehydrated with 95% (v/v)and then 70% (v/v) methanol. Non-specific binding sites were blocked and incubatedwith FITC-conjugated overnight at 4 ◦C. 8-Oxoguanine adducts were observed undera fluorescent microscope (Olympus BX40) with FITC filter. Images were obtainedwith a Nikon Coolpix 4500 camera. Fluorescence in the control was normalized to1 unit and the changes in fluorescence were expressed as fold increase relative tocontrol.
2.9. Superoxide dismutase (SOD) and catalase activity
The total SOD activity was determined using Superoxide Dismutase Assay Kit(Calbiochem Inc., San Diego, CA). The assay measures the rate of oxidation of5,6,6a,11b-tetrahydroxylbenzo[c]fluorene in aqueous alkaline solution to yield achromophore that absorbs at 525 nm. One unit is determined from the ratio of veloc-ity sample: velocity control compared to the ratio provided by the manufacturer.Results are reported as U/mg of protein. Catalase activity was quantified spectropho-tometrically by evaluating the decrease in (H2O2) absorbance at 240 nm in presenceof the enzyme (Beers and Sizer, 1952). One unit of catalase is defined as 1 mmol ofH2O2 consumed per minute. Data are expressed as U/mg of protein.
2.10. Glutathione reductase activity
The glutathione reductase (GR) activity assay was performed as originallydescribed by Carlberg and Mannervik (1975). After the cells were exposed to differ-ent treatments, they were lysed with Tris 0.01 M buffer at pH 8.0 and 0.01% of TritonX-100, and centrifuged 10,000 × g for 15 min to 4 ◦C. Samples were incubated in0.1 M PBS buffer at pH 7.6 using 0.5 mM EDTA, 0.5 mM oxidized glutathione (GSSG)and 0.1 mM NADPH. The GR activity was measured by the NADPH consumptionat 340 nm. One unit is defined as 1 �mol of NADPH oxidized per minute. Data areexpressed as U/mg of protein.
2.11. Glutathione-S-transferase activity
The glutathione-S-transferase (GST) activity assay was performed as originallydescribed by Habig et al. (1974). After different treatment, the cells were lysed withTris 0.01 M pH 8 and 0.01% of Triton X-100, and centrifuged 10,000 × g by 15 minto 4 ◦C. Samples were incubated in the reaction mixture consisted of 0.05 M PBS
Author's personal copy
Y.I. Chirino et al. / Toxicology Letters 193 (2010) 209–216 211
(pH 6.5), 1 mM GSH, 1 mM 1-chloro-2,4-dinitro benzene (CDNB) and the changesin the absorbance were recorded at 340 nm. One unit is defined as 1 �mol of CDNBconjugated with GSH per minute at pH 6.5 using a molar extinction coefficient of9.6 × 103/M/cm. Data are expressed as U/mg of protein.
2.12. Western blot of Mn-SOD, Cu/Zn-SOD, catalase and CCS
Cells were lysed with 20 mM Tris, 1% NP-40, and 150 mM NaCl, pH 8.0, witha protease inhibitor cocktail (Sigma Chemical Co. St. Louis, MO, USA). The proteinof samples was determined through the bicinchoninic acid protein assay (Smith etal., 1985). Twenty five micrograms of protein were separated using 15% SDS-PAGE.The proteins were transferred onto a nitrocellulose membrane in a Semi-Dry Trans-fer System. The membrane was blocked with 5% (w/v) dry milk in PBS with 0.05%(v/v) Tween-20 (PBS-T) and incubated with the primary antibody anti-SOD-Mn(Stressgen, Ontario Canada, SOD-111) 1:5000 dilution, anti-Cu/Zn-SOD (Stress-gen, Ontario Canada, SOD-101) 1:1000 dilution, antibody anti-CCS (Santa Cruz,sc-55569) 1:1000 dilution, and antibody anti-catalase (Calbiochem Inc., San DiegoCA, USA, 219010) 1:2000 dilution. Actin was utilized as loading control (kindlyprovided by Dr. Manuel Hernández, Department of Cellular Biology, CINVESTAV-IPN, Mexico City, Mexico). To detect Mn-SOD and Cu/Zn-SOD, we used horseradishperoxidase-conjugated anti-rabbit IgG (Amersham GE, Pittsburgh, PA, NA934V)1:2,000 dilution, as secondary antibody, whereas, for catalase, CCS and actin, weemployed a horseradish peroxidase-conjugated anti-mouse IgG (Amersham GE,Pittsburgh, PA, NXA931) 1:2000 dilution. Immunoreactivity was visualized by anenhanced chemiluminescence detection kit (Millipore Corporation, Billerica, MA,USA). Protein levels were evaluated through densitometry using the Image J free-ware program (http://rsb.info.nih.gov/ij) (Girish and Vijayalakshmi, 2004).
2.13. Exposure to PM10 and challenge with H2O2
A549 cells were seeded at a density of 2 × 104 cells/cm2 onto 96-well plasticplates and used for experiments on the following day. Then, cells were exposedto 10 �g/cm2 of PM10 for 24 h with fresh F12K medium. After this time, cells wereextensively washed with PBS and exposure for 24 h in F12K medium supplementedwith 10% fetal bovine serum to the following H2O2 concentrations: 500 �M, 750 �Mand 1000 �M. At the end of this incubation, the cells were washed with PBS and cellviability was measured by the crystal violet method (Kueng et al., 1989).
2.14. Statistical analysis
Results obtained are expressed as mean ± SD with exception of 8-oxoG adductswhich are expressed as densitometry of fluorescence on fifty cells from three inde-pendent experiments, median, 25th and 75th percentiles, and maximal and minimalvalues. Data were analyzed with the software Prism v. 5a (GraphPad, San Diego, CA,USA) by one-way analysis of variance followed by multiple comparisons accordingto Bonferroni. A value of p < 0.05 was considered as statistically significant.
3. Results
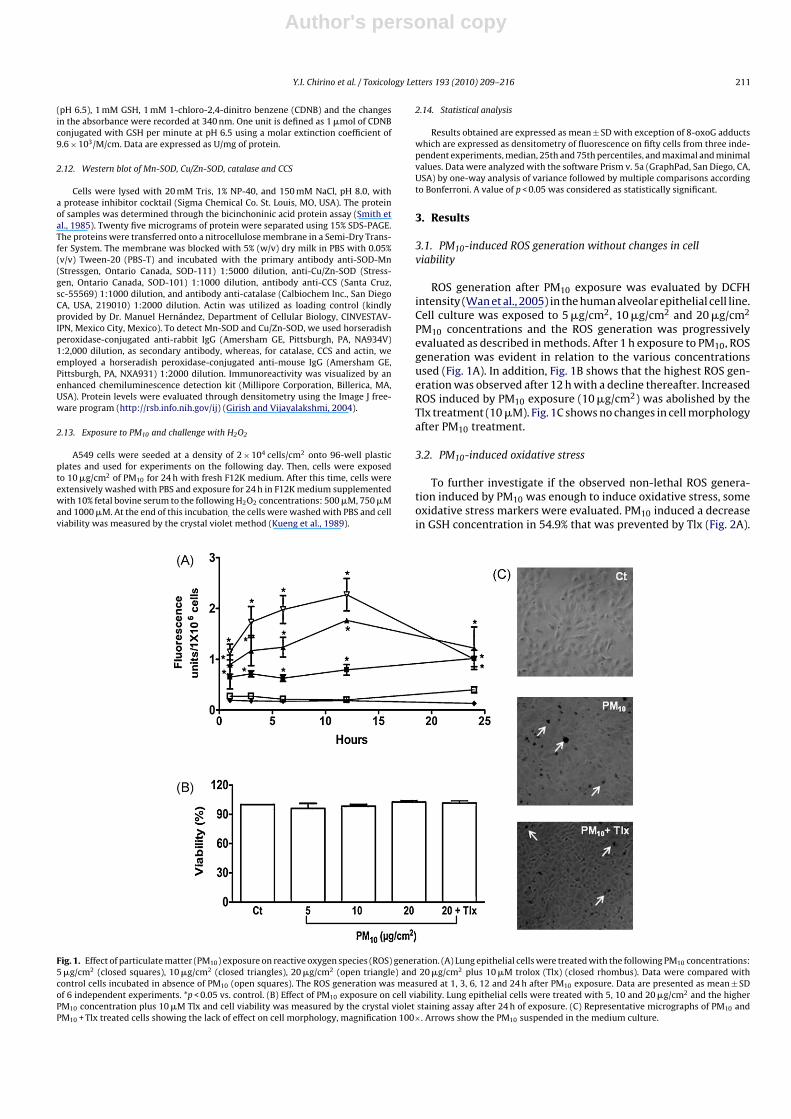
3.1. PM10-induced ROS generation without changes in cellviability
ROS generation after PM10 exposure was evaluated by DCFHintensity (Wan et al., 2005) in the human alveolar epithelial cell line.Cell culture was exposed to 5 �g/cm2, 10 �g/cm2 and 20 �g/cm2
PM10 concentrations and the ROS generation was progressivelyevaluated as described in methods. After 1 h exposure to PM10, ROSgeneration was evident in relation to the various concentrationsused (Fig. 1A). In addition, Fig. 1B shows that the highest ROS gen-eration was observed after 12 h with a decline thereafter. IncreasedROS induced by PM10 exposure (10 �g/cm2) was abolished by theTlx treatment (10 �M). Fig. 1C shows no changes in cell morphologyafter PM10 treatment.
3.2. PM10-induced oxidative stress
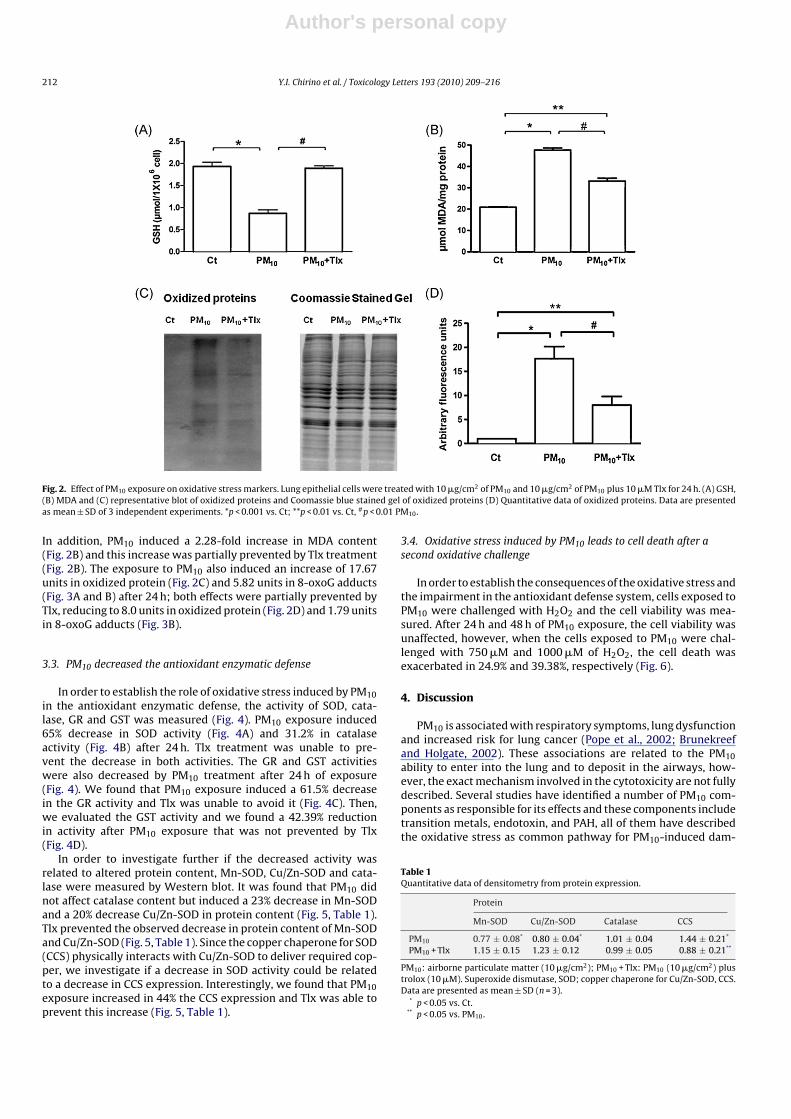
To further investigate if the observed non-lethal ROS genera-tion induced by PM10 was enough to induce oxidative stress, someoxidative stress markers were evaluated. PM10 induced a decreasein GSH concentration in 54.9% that was prevented by Tlx (Fig. 2A).
Fig. 1. Effect of particulate matter (PM10) exposure on reactive oxygen species (ROS) generation. (A) Lung epithelial cells were treated with the following PM10 concentrations:5 �g/cm2 (closed squares), 10 �g/cm2 (closed triangles), 20 �g/cm2 (open triangle) and 20 �g/cm2 plus 10 �M trolox (Tlx) (closed rhombus). Data were compared withcontrol cells incubated in absence of PM10 (open squares). The ROS generation was measured at 1, 3, 6, 12 and 24 h after PM10 exposure. Data are presented as mean ± SDof 6 independent experiments. *p < 0.05 vs. control. (B) Effect of PM10 exposure on cell viability. Lung epithelial cells were treated with 5, 10 and 20 �g/cm2 and the higherPM10 concentration plus 10 �M Tlx and cell viability was measured by the crystal violet staining assay after 24 h of exposure. (C) Representative micrographs of PM10 andPM10 + Tlx treated cells showing the lack of effect on cell morphology, magnification 100×. Arrows show the PM10 suspended in the medium culture.
Author's personal copy
212 Y.I. Chirino et al. / Toxicology Letters 193 (2010) 209–216
Fig. 2. Effect of PM10 exposure on oxidative stress markers. Lung epithelial cells were treated with 10 �g/cm2 of PM10 and 10 �g/cm2 of PM10 plus 10 �M Tlx for 24 h. (A) GSH,(B) MDA and (C) representative blot of oxidized proteins and Coomassie blue stained gel of oxidized proteins (D) Quantitative data of oxidized proteins. Data are presentedas mean ± SD of 3 independent experiments. *p < 0.001 vs. Ct; **p < 0.01 vs. Ct, #p < 0.01 PM10.
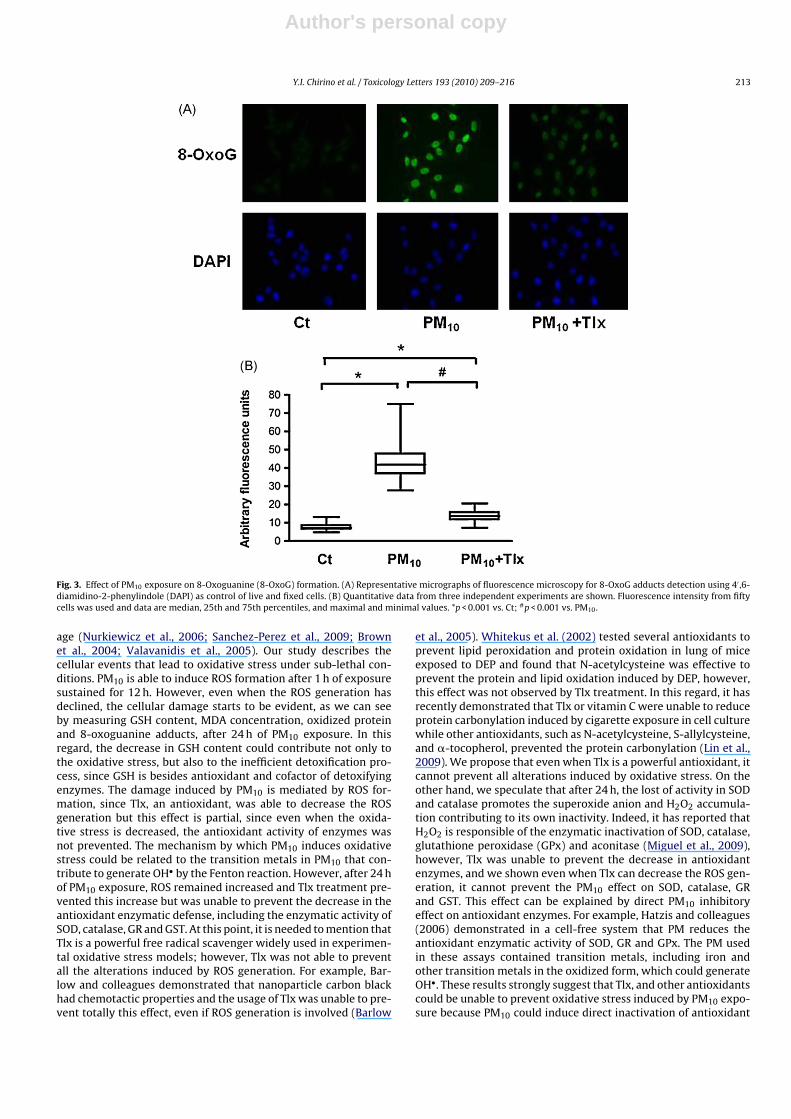
In addition, PM10 induced a 2.28-fold increase in MDA content(Fig. 2B) and this increase was partially prevented by Tlx treatment(Fig. 2B). The exposure to PM10 also induced an increase of 17.67units in oxidized protein (Fig. 2C) and 5.82 units in 8-oxoG adducts(Fig. 3A and B) after 24 h; both effects were partially prevented byTlx, reducing to 8.0 units in oxidized protein (Fig. 2D) and 1.79 unitsin 8-oxoG adducts (Fig. 3B).
3.3. PM10 decreased the antioxidant enzymatic defense
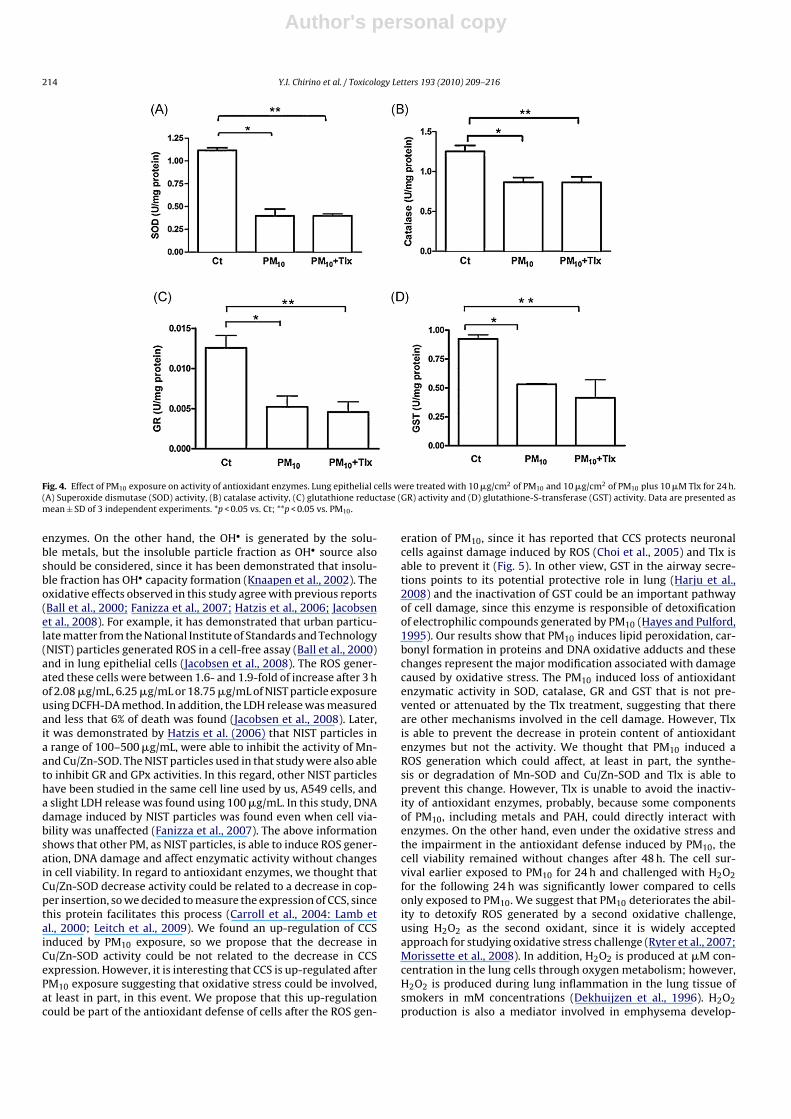
In order to establish the role of oxidative stress induced by PM10in the antioxidant enzymatic defense, the activity of SOD, cata-lase, GR and GST was measured (Fig. 4). PM10 exposure induced65% decrease in SOD activity (Fig. 4A) and 31.2% in catalaseactivity (Fig. 4B) after 24 h. Tlx treatment was unable to pre-vent the decrease in both activities. The GR and GST activitieswere also decreased by PM10 treatment after 24 h of exposure(Fig. 4). We found that PM10 exposure induced a 61.5% decreasein the GR activity and Tlx was unable to avoid it (Fig. 4C). Then,we evaluated the GST activity and we found a 42.39% reductionin activity after PM10 exposure that was not prevented by Tlx(Fig. 4D).
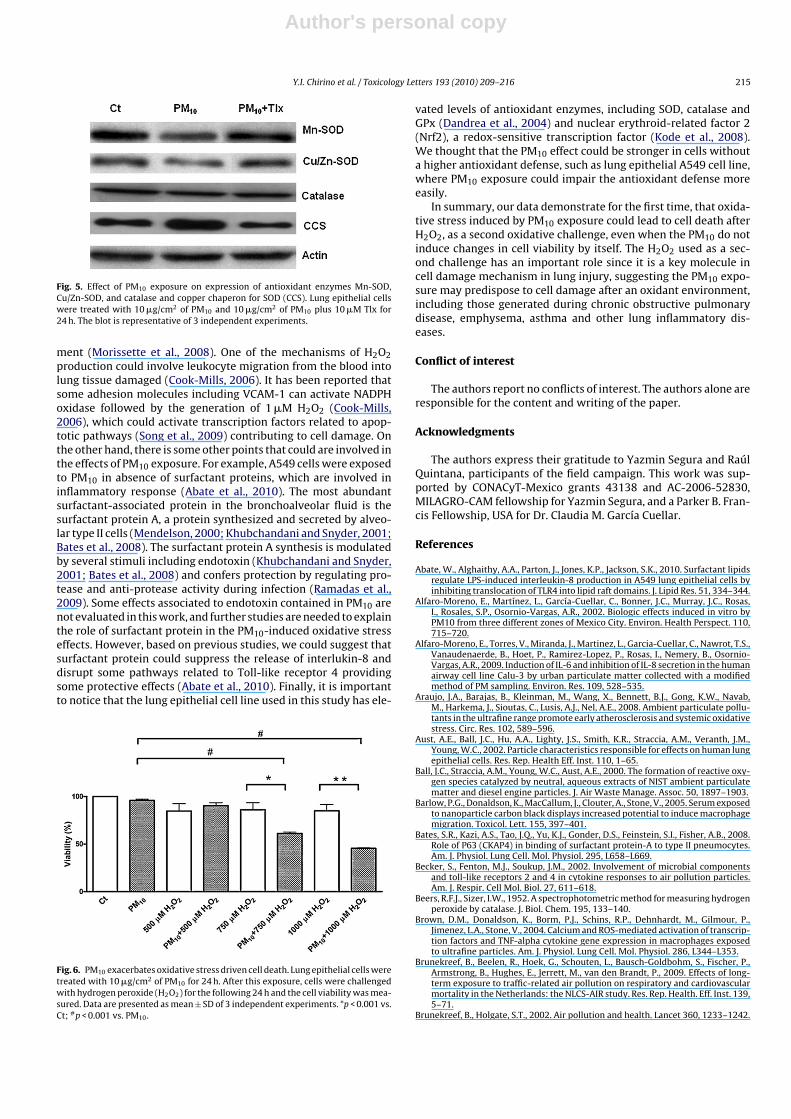
In order to investigate further if the decreased activity wasrelated to altered protein content, Mn-SOD, Cu/Zn-SOD and cata-lase were measured by Western blot. It was found that PM10 didnot affect catalase content but induced a 23% decrease in Mn-SODand a 20% decrease Cu/Zn-SOD in protein content (Fig. 5, Table 1).Tlx prevented the observed decrease in protein content of Mn-SODand Cu/Zn-SOD (Fig. 5, Table 1). Since the copper chaperone for SOD(CCS) physically interacts with Cu/Zn-SOD to deliver required cop-per, we investigate if a decrease in SOD activity could be relatedto a decrease in CCS expression. Interestingly, we found that PM10exposure increased in 44% the CCS expression and Tlx was able toprevent this increase (Fig. 5, Table 1).
3.4. Oxidative stress induced by PM10 leads to cell death after asecond oxidative challenge
In order to establish the consequences of the oxidative stress andthe impairment in the antioxidant defense system, cells exposed toPM10 were challenged with H2O2 and the cell viability was mea-sured. After 24 h and 48 h of PM10 exposure, the cell viability wasunaffected, however, when the cells exposed to PM10 were chal-lenged with 750 �M and 1000 �M of H2O2, the cell death wasexacerbated in 24.9% and 39.38%, respectively (Fig. 6).
4. Discussion
PM10 is associated with respiratory symptoms, lung dysfunctionand increased risk for lung cancer (Pope et al., 2002; Brunekreefand Holgate, 2002). These associations are related to the PM10ability to enter into the lung and to deposit in the airways, how-ever, the exact mechanism involved in the cytotoxicity are not fullydescribed. Several studies have identified a number of PM10 com-ponents as responsible for its effects and these components includetransition metals, endotoxin, and PAH, all of them have describedthe oxidative stress as common pathway for PM10-induced dam-
Table 1Quantitative data of densitometry from protein expression.
Protein
Mn-SOD Cu/Zn-SOD Catalase CCS
PM10 0.77 ± 0.08* 0.80 ± 0.04* 1.01 ± 0.04 1.44 ± 0.21*
PM10 + Tlx 1.15 ± 0.15 1.23 ± 0.12 0.99 ± 0.05 0.88 ± 0.21**
PM10: airborne particulate matter (10 �g/cm2); PM10 + Tlx: PM10 (10 �g/cm2) plustrolox (10 �M). Superoxide dismutase, SOD; copper chaperone for Cu/Zn-SOD, CCS.Data are presented as mean ± SD (n = 3).
* p < 0.05 vs. Ct.** p < 0.05 vs. PM10.
Author's personal copy
Y.I. Chirino et al. / Toxicology Letters 193 (2010) 209–216 213
Fig. 3. Effect of PM10 exposure on 8-Oxoguanine (8-OxoG) formation. (A) Representative micrographs of fluorescence microscopy for 8-OxoG adducts detection using 4′ ,6-diamidino-2-phenylindole (DAPI) as control of live and fixed cells. (B) Quantitative data from three independent experiments are shown. Fluorescence intensity from fiftycells was used and data are median, 25th and 75th percentiles, and maximal and minimal values. *p < 0.001 vs. Ct; #p < 0.001 vs. PM10.
age (Nurkiewicz et al., 2006; Sanchez-Perez et al., 2009; Brownet al., 2004; Valavanidis et al., 2005). Our study describes thecellular events that lead to oxidative stress under sub-lethal con-ditions. PM10 is able to induce ROS formation after 1 h of exposuresustained for 12 h. However, even when the ROS generation hasdeclined, the cellular damage starts to be evident, as we can seeby measuring GSH content, MDA concentration, oxidized proteinand 8-oxoguanine adducts, after 24 h of PM10 exposure. In thisregard, the decrease in GSH content could contribute not only tothe oxidative stress, but also to the inefficient detoxification pro-cess, since GSH is besides antioxidant and cofactor of detoxifyingenzymes. The damage induced by PM10 is mediated by ROS for-mation, since Tlx, an antioxidant, was able to decrease the ROSgeneration but this effect is partial, since even when the oxida-tive stress is decreased, the antioxidant activity of enzymes wasnot prevented. The mechanism by which PM10 induces oxidativestress could be related to the transition metals in PM10 that con-tribute to generate OH• by the Fenton reaction. However, after 24 hof PM10 exposure, ROS remained increased and Tlx treatment pre-vented this increase but was unable to prevent the decrease in theantioxidant enzymatic defense, including the enzymatic activity ofSOD, catalase, GR and GST. At this point, it is needed to mention thatTlx is a powerful free radical scavenger widely used in experimen-tal oxidative stress models; however, Tlx was not able to preventall the alterations induced by ROS generation. For example, Bar-low and colleagues demonstrated that nanoparticle carbon blackhad chemotactic properties and the usage of Tlx was unable to pre-vent totally this effect, even if ROS generation is involved (Barlow
et al., 2005). Whitekus et al. (2002) tested several antioxidants toprevent lipid peroxidation and protein oxidation in lung of miceexposed to DEP and found that N-acetylcysteine was effective toprevent the protein and lipid oxidation induced by DEP, however,this effect was not observed by Tlx treatment. In this regard, it hasrecently demonstrated that Tlx or vitamin C were unable to reduceprotein carbonylation induced by cigarette exposure in cell culturewhile other antioxidants, such as N-acetylcysteine, S-allylcysteine,and �-tocopherol, prevented the protein carbonylation (Lin et al.,2009). We propose that even when Tlx is a powerful antioxidant, itcannot prevent all alterations induced by oxidative stress. On theother hand, we speculate that after 24 h, the lost of activity in SODand catalase promotes the superoxide anion and H2O2 accumula-tion contributing to its own inactivity. Indeed, it has reported thatH2O2 is responsible of the enzymatic inactivation of SOD, catalase,glutathione peroxidase (GPx) and aconitase (Miguel et al., 2009),however, Tlx was unable to prevent the decrease in antioxidantenzymes, and we shown even when Tlx can decrease the ROS gen-eration, it cannot prevent the PM10 effect on SOD, catalase, GRand GST. This effect can be explained by direct PM10 inhibitoryeffect on antioxidant enzymes. For example, Hatzis and colleagues(2006) demonstrated in a cell-free system that PM reduces theantioxidant enzymatic activity of SOD, GR and GPx. The PM usedin these assays contained transition metals, including iron andother transition metals in the oxidized form, which could generateOH•. These results strongly suggest that Tlx, and other antioxidantscould be unable to prevent oxidative stress induced by PM10 expo-sure because PM10 could induce direct inactivation of antioxidant
Author's personal copy
214 Y.I. Chirino et al. / Toxicology Letters 193 (2010) 209–216
Fig. 4. Effect of PM10 exposure on activity of antioxidant enzymes. Lung epithelial cells were treated with 10 �g/cm2 of PM10 and 10 �g/cm2 of PM10 plus 10 �M Tlx for 24 h.(A) Superoxide dismutase (SOD) activity, (B) catalase activity, (C) glutathione reductase (GR) activity and (D) glutathione-S-transferase (GST) activity. Data are presented asmean ± SD of 3 independent experiments. *p < 0.05 vs. Ct; **p < 0.05 vs. PM10.
enzymes. On the other hand, the OH• is generated by the solu-ble metals, but the insoluble particle fraction as OH• source alsoshould be considered, since it has been demonstrated that insolu-ble fraction has OH• capacity formation (Knaapen et al., 2002). Theoxidative effects observed in this study agree with previous reports(Ball et al., 2000; Fanizza et al., 2007; Hatzis et al., 2006; Jacobsenet al., 2008). For example, it has demonstrated that urban particu-late matter from the National Institute of Standards and Technology(NIST) particles generated ROS in a cell-free assay (Ball et al., 2000)and in lung epithelial cells (Jacobsen et al., 2008). The ROS gener-ated these cells were between 1.6- and 1.9-fold of increase after 3 hof 2.08 �g/mL, 6.25 �g/mL or 18.75 �g/mL of NIST particle exposureusing DCFH-DA method. In addition, the LDH release was measuredand less that 6% of death was found (Jacobsen et al., 2008). Later,it was demonstrated by Hatzis et al. (2006) that NIST particles ina range of 100–500 �g/mL, were able to inhibit the activity of Mn-and Cu/Zn-SOD. The NIST particles used in that study were also ableto inhibit GR and GPx activities. In this regard, other NIST particleshave been studied in the same cell line used by us, A549 cells, anda slight LDH release was found using 100 �g/mL. In this study, DNAdamage induced by NIST particles was found even when cell via-bility was unaffected (Fanizza et al., 2007). The above informationshows that other PM, as NIST particles, is able to induce ROS gener-ation, DNA damage and affect enzymatic activity without changesin cell viability. In regard to antioxidant enzymes, we thought thatCu/Zn-SOD decrease activity could be related to a decrease in cop-per insertion, so we decided to measure the expression of CCS, sincethis protein facilitates this process (Carroll et al., 2004: Lamb etal., 2000; Leitch et al., 2009). We found an up-regulation of CCSinduced by PM10 exposure, so we propose that the decrease inCu/Zn-SOD activity could be not related to the decrease in CCSexpression. However, it is interesting that CCS is up-regulated afterPM10 exposure suggesting that oxidative stress could be involved,at least in part, in this event. We propose that this up-regulationcould be part of the antioxidant defense of cells after the ROS gen-
eration of PM10, since it has reported that CCS protects neuronalcells against damage induced by ROS (Choi et al., 2005) and Tlx isable to prevent it (Fig. 5). In other view, GST in the airway secre-tions points to its potential protective role in lung (Harju et al.,2008) and the inactivation of GST could be an important pathwayof cell damage, since this enzyme is responsible of detoxificationof electrophilic compounds generated by PM10 (Hayes and Pulford,1995). Our results show that PM10 induces lipid peroxidation, car-bonyl formation in proteins and DNA oxidative adducts and thesechanges represent the major modification associated with damagecaused by oxidative stress. The PM10 induced loss of antioxidantenzymatic activity in SOD, catalase, GR and GST that is not pre-vented or attenuated by the Tlx treatment, suggesting that thereare other mechanisms involved in the cell damage. However, Tlxis able to prevent the decrease in protein content of antioxidantenzymes but not the activity. We thought that PM10 induced aROS generation which could affect, at least in part, the synthe-sis or degradation of Mn-SOD and Cu/Zn-SOD and Tlx is able toprevent this change. However, Tlx is unable to avoid the inactiv-ity of antioxidant enzymes, probably, because some componentsof PM10, including metals and PAH, could directly interact withenzymes. On the other hand, even under the oxidative stress andthe impairment in the antioxidant defense induced by PM10, thecell viability remained without changes after 48 h. The cell sur-vival earlier exposed to PM10 for 24 h and challenged with H2O2for the following 24 h was significantly lower compared to cellsonly exposed to PM10. We suggest that PM10 deteriorates the abil-ity to detoxify ROS generated by a second oxidative challenge,using H2O2 as the second oxidant, since it is widely acceptedapproach for studying oxidative stress challenge (Ryter et al., 2007;Morissette et al., 2008). In addition, H2O2 is produced at �M con-centration in the lung cells through oxygen metabolism; however,H2O2 is produced during lung inflammation in the lung tissue ofsmokers in mM concentrations (Dekhuijzen et al., 1996). H2O2production is also a mediator involved in emphysema develop-
Author's personal copy
Y.I. Chirino et al. / Toxicology Letters 193 (2010) 209–216 215
Fig. 5. Effect of PM10 exposure on expression of antioxidant enzymes Mn-SOD,Cu/Zn-SOD, and catalase and copper chaperon for SOD (CCS). Lung epithelial cellswere treated with 10 �g/cm2 of PM10 and 10 �g/cm2 of PM10 plus 10 �M Tlx for24 h. The blot is representative of 3 independent experiments.
ment (Morissette et al., 2008). One of the mechanisms of H2O2production could involve leukocyte migration from the blood intolung tissue damaged (Cook-Mills, 2006). It has been reported thatsome adhesion molecules including VCAM-1 can activate NADPHoxidase followed by the generation of 1 �M H2O2 (Cook-Mills,2006), which could activate transcription factors related to apop-totic pathways (Song et al., 2009) contributing to cell damage. Onthe other hand, there is some other points that could are involved inthe effects of PM10 exposure. For example, A549 cells were exposedto PM10 in absence of surfactant proteins, which are involved ininflammatory response (Abate et al., 2010). The most abundantsurfactant-associated protein in the bronchoalveolar fluid is thesurfactant protein A, a protein synthesized and secreted by alveo-lar type II cells (Mendelson, 2000; Khubchandani and Snyder, 2001;Bates et al., 2008). The surfactant protein A synthesis is modulatedby several stimuli including endotoxin (Khubchandani and Snyder,2001; Bates et al., 2008) and confers protection by regulating pro-tease and anti-protease activity during infection (Ramadas et al.,2009). Some effects associated to endotoxin contained in PM10 arenot evaluated in this work, and further studies are needed to explainthe role of surfactant protein in the PM10-induced oxidative stresseffects. However, based on previous studies, we could suggest thatsurfactant protein could suppress the release of interlukin-8 anddisrupt some pathways related to Toll-like receptor 4 providingsome protective effects (Abate et al., 2010). Finally, it is importantto notice that the lung epithelial cell line used in this study has ele-
Fig. 6. PM10 exacerbates oxidative stress driven cell death. Lung epithelial cells weretreated with 10 �g/cm2 of PM10 for 24 h. After this exposure, cells were challengedwith hydrogen peroxide (H2O2) for the following 24 h and the cell viability was mea-sured. Data are presented as mean ± SD of 3 independent experiments. *p < 0.001 vs.Ct; #p < 0.001 vs. PM10.
vated levels of antioxidant enzymes, including SOD, catalase andGPx (Dandrea et al., 2004) and nuclear erythroid-related factor 2(Nrf2), a redox-sensitive transcription factor (Kode et al., 2008).We thought that the PM10 effect could be stronger in cells withouta higher antioxidant defense, such as lung epithelial A549 cell line,where PM10 exposure could impair the antioxidant defense moreeasily.
In summary, our data demonstrate for the first time, that oxida-tive stress induced by PM10 exposure could lead to cell death afterH2O2, as a second oxidative challenge, even when the PM10 do notinduce changes in cell viability by itself. The H2O2 used as a sec-ond challenge has an important role since it is a key molecule incell damage mechanism in lung injury, suggesting the PM10 expo-sure may predispose to cell damage after an oxidant environment,including those generated during chronic obstructive pulmonarydisease, emphysema, asthma and other lung inflammatory dis-eases.
Conflict of interest
The authors report no conflicts of interest. The authors alone areresponsible for the content and writing of the paper.
Acknowledgments
The authors express their gratitude to Yazmin Segura and RaúlQuintana, participants of the field campaign. This work was sup-ported by CONACyT-Mexico grants 43138 and AC-2006-52830,MILAGRO-CAM fellowship for Yazmin Segura, and a Parker B. Fran-cis Fellowship, USA for Dr. Claudia M. García Cuellar.
References
Abate, W., Alghaithy, A.A., Parton, J., Jones, K.P., Jackson, S.K., 2010. Surfactant lipidsregulate LPS-induced interleukin-8 production in A549 lung epithelial cells byinhibiting translocation of TLR4 into lipid raft domains. J. Lipid Res. 51, 334–344.
Alfaro-Moreno, E., Martínez, L., García-Cuellar, C., Bonner, J.C., Murray, J.C., Rosas,I., Rosales, S.P., Osornio-Vargas, A.R., 2002. Biologic effects induced in vitro byPM10 from three different zones of Mexico City. Environ. Health Perspect. 110,715–720.
Alfaro-Moreno, E., Torres, V., Miranda, J., Martinez, L., Garcia-Cuellar, C., Nawrot, T.S.,Vanaudenaerde, B., Hoet, P., Ramirez-Lopez, P., Rosas, I., Nemery, B., Osornio-Vargas, A.R., 2009. Induction of IL-6 and inhibition of IL-8 secretion in the humanairway cell line Calu-3 by urban particulate matter collected with a modifiedmethod of PM sampling. Environ. Res. 109, 528–535.
Araujo, J.A., Barajas, B., Kleinman, M., Wang, X., Bennett, B.J., Gong, K.W., Navab,M., Harkema, J., Sioutas, C., Lusis, A.J., Nel, A.E., 2008. Ambient particulate pollu-tants in the ultrafine range promote early atherosclerosis and systemic oxidativestress. Circ. Res. 102, 589–596.
Aust, A.E., Ball, J.C., Hu, A.A., Lighty, J.S., Smith, K.R., Straccia, A.M., Veranth, J.M.,Young, W.C., 2002. Particle characteristics responsible for effects on human lungepithelial cells. Res. Rep. Health Eff. Inst. 110, 1–65.
Ball, J.C., Straccia, A.M., Young, W.C., Aust, A.E., 2000. The formation of reactive oxy-gen species catalyzed by neutral, aqueous extracts of NIST ambient particulatematter and diesel engine particles. J. Air Waste Manage. Assoc. 50, 1897–1903.
Barlow, P.G., Donaldson, K., MacCallum, J., Clouter, A., Stone, V., 2005. Serum exposedto nanoparticle carbon black displays increased potential to induce macrophagemigration. Toxicol. Lett. 155, 397–401.
Bates, S.R., Kazi, A.S., Tao, J.Q., Yu, K.J., Gonder, D.S., Feinstein, S.I., Fisher, A.B., 2008.Role of P63 (CKAP4) in binding of surfactant protein-A to type II pneumocytes.Am. J. Physiol. Lung Cell. Mol. Physiol. 295, L658–L669.
Becker, S., Fenton, M.J., Soukup, J.M., 2002. Involvement of microbial componentsand toll-like receptors 2 and 4 in cytokine responses to air pollution particles.Am. J. Respir. Cell Mol. Biol. 27, 611–618.
Beers, R.F.J., Sizer, I.W., 1952. A spectrophotometric method for measuring hydrogenperoxide by catalase. J. Biol. Chem. 195, 133–140.
Brown, D.M., Donaldson, K., Borm, P.J., Schins, R.P., Dehnhardt, M., Gilmour, P.,Jimenez, L.A., Stone, V., 2004. Calcium and ROS-mediated activation of transcrip-tion factors and TNF-alpha cytokine gene expression in macrophages exposedto ultrafine particles. Am. J. Physiol. Lung Cell. Mol. Physiol. 286, L344–L353.
Brunekreef, B., Beelen, R., Hoek, G., Schouten, L., Bausch-Goldbohm, S., Fischer, P.,Armstrong, B., Hughes, E., Jerrett, M., van den Brandt, P., 2009. Effects of long-term exposure to traffic-related air pollution on respiratory and cardiovascularmortality in the Netherlands: the NLCS-AIR study. Res. Rep. Health. Eff. Inst. 139,5–71.
Brunekreef, B., Holgate, S.T., 2002. Air pollution and health. Lancet 360, 1233–1242.

Author's personal copy
216 Y.I. Chirino et al. / Toxicology Letters 193 (2010) 209–216
Calderon-Garciduenas, L., Gambling, T.M., Acuna, H., Garcia, R., Osnaya, N., Monroy,S., Villarreal-Calderon, A., Carson, J., Koren, H.S., Devlin, R.B., 2001. Canines assentinel species for assessing chronic exposures to air pollutants. Part 2. Cardiacpathology. Toxicol. Sci. 61, 356–367.
Carlberg, I., Mannervik, B., 1975. Purification and characterization of the flavoen-zyme glutathione reductase from rat liver. J. Biol. Chem. 250, 5475–5480.
Carroll, M.C., Girouard, J.B., Ulloa, J.L., Subramaniam, J.R., Wong, P.C., Valentine, J.S.,Culotta, V.C., 2004. Mechanisms for activating Cu- and Zn-containing superoxidedismutase in the absence of the CCS Cu chaperone. Proc. Natl. Acad. Sci. U.S.A.101, 5964–5969.
Choi, S.H., Kim, D.W., Kim, S.Y., An, J.J., Lee, S.H., Choi, H.S., Sohn, E.J., Hwang, S.I.,Won, M.H., Kang, T.C., Kwon, H.J., Kang, J.H., Cho, S.W., Park, J., Eum, W.S., Choi,S.Y., 2005. Transduced human copper chaperone for Cu. Zn-SOD (PEP-1-CCS)protects against neuronal cell death. Mol. Cells 20, 401–408.
Churg, A., Brauer, M., del Carmen Avila-Casado, M., Fortoul, T.I., Wright, J.L., 2003.Chronic exposure to high levels of particulate air pollution and small airwayremodeling. Environ. Health Perspect. 111, 714–718.
Cook-Mills, J.M., 2006. Hydrogen peroxide activation of endothelial cell-associatedMMPs during VCAM-1-dependent leukocyte migration. Cell. Mol. Biol. 52, 8–16.
Dandrea, T., Hellmold, H., Jonsson, C., Zhivotovsky, B., Hofer, T., Wärngård, L., Cot-greave, I., 2004. The transcriptosomal response of human A549 lung cells to ahydrogen peroxide-generating system: relationship to DNA damage, cell cyclearrest, and caspase activation. Free Radic. Biol. Med. 36, 881–896.
Dekhuijzen, P.N., Aben, K.K., Dekker, I., Aarts, L.P., Wielders, P.L., van Herwaarden,C.L., Bast, A., 1996. Increased exhalation of hydrogen peroxide in patients withstable and unstable chronic obstructive pulmonary disease. Am. J. Respir. Crit.Care Med. 154, 813–816.
Fanizza, C., Ursini, C.L., Paba, E., Ciervo, A., Di Francesco, A., Maiello, R., De Simone, P.,Cavallo, D., 2007. Cytotoxicity and DNA-damage in human lung epithelial cellsexposed to respirable alpha-quartz. Toxicol. In Vitro 21, 586–594.
Girish, V., Vijayalakshmi, A., 2004. Affordable image analysis using NIHImage/ImageJ. Indian J. Cancer 41, 47.
Habig, W.H., Pabst, M.J., Fleischner, G., Gatmaitan, Z., Arias, I.M., Jakoby, W.B., 1974.The identity of glutathione S-transferase B with ligandin, a major binding proteinof liver. Proc. Natl. Acad. Sci. U.S.A. 71, 3879–3882.
Harju, T., Mazur, W., Merikallio, H., Soini, Y., Kinnula, V.L., 2008. Glutathione-S-transferases in lung and sputum specimens, effects of smoking and COPDseverity. Respir. Res. 9, 80.
Hatzis, C., Godleski, J.J., Gonzalez-Flecha, B., Wolfson, J.M., Koutrakis, P., 2006. Ambi-ent particulate matter exhibits direct inhibitory effects on oxidative stressenzymes. Environ. Sci. Technol. 40, 2805–2811.
Hayes, J.D., Pulford, D.J., 1995. The glutathione S-transferase supergene family: regu-lation of GST and the contribution of the isoenzymes to cancer chemoprotectionand drug resistance. Crit. Rev. Biochem. Mol. Biol. 30, 445–600.
Jacobsen, N.R., Møller, P., Cohn, C.A., Loft, S., Vogel, U., Wallin, H., 2008. Diesel exhaustparticles are mutagenic in FE1-MutaMouse lung epithelial cells. Mutat. Res. 641,54–57.
Khubchandani, K.R., Snyder, J.M., 2001. Surfactant protein A (SP-A): the alveolus andbeyond. FASEB J. 15, 59–69.
Knaapen, A.M., Shi, T., Borm, P.J., Schins, R.P., 2002. Soluble metals as well as the insol-uble particle fraction are involved in cellular DNA damage induced by particulatematter. Mol. Cell. Biochem. 234–235, 317–326.
Kode, A., Rajendrasozhan, S., Caito, S., Yang, S.R., Megson, I.L., Rahman, I., 2008.Resveratrol induces glutathione synthesis by activation of Nrf2 and protectsagainst cigarette smoke-mediated oxidative stress in human lung epithelialcells. Am. J. Physiol. Lung Cell. Mol. Physiol. 94, L478–L488.
Kueng, W., Silber, E., Eppenberger, U., 1989. Quantification of cells cultured on 96-well plates. Anal. Biochem. 182, 16–19.
Lagorio, S., Forastiere, F., Pistelli, R., Iavarone, I., Michelozzi, P., Fano, V., Marconi, A.,Ziemacki, G., Ostro, B.D., 2006. Air pollution and lung function among susceptibleadult subjects: a panel study. Environ. Health 5, 11.
Lamb, A.L., Torres, A.S., O’Halloran, T.V., Rosenzweig, A.C., 2000. Heterodimer for-mation between superoxide dismutase and its copper chaperone. Biochemistry39, 14720–14727.
Lawler, J.M., Song, W., Demaree, S.R., 2003. Hindlimb unloading increases oxidativestress and disrupts antioxidant capacity in skeletal muscle. Free Radic. Biol. Med.35, 9–16.
Leitch, J.M., Yick, P.C., Culotta, V.C., 2009. The right to choose: multiple pathways foractivating copper, zinc superoxide dismutase. J. Biol. Chem. 37, 24679–24683.
Li, Y.J., Takizawa, H., Azuma, A., Kohyama, T., Yamauchi, Y., Takahashi, S., Yamamoto,M., Kawada, T., Kudoh, S., Sugawara, I., 2008. Disruption of Nrf2 enhances suscep-tibility to airway inflammatory responses induced by low-dose diesel exhaustparticles in mice. Clin. Immunol. 128, 366–373.
Lin, C.C., Su, T.H., Wang, T.S., 2009. Protein carbonylation in THP-1 cells induced bycigarette smoke extract via a copper-catalyzed pathway. Chem. Res. Toxicol. 22,1232–1238.
Liu, L., Poon, R., Chen, L., Frescura, A.M., Montuschi, P., Ciabattoni, G., Wheeler,A., Dales, R., 2009. Acute effects of air pollution on pulmonary function, air-way inflammation, and oxidative stress in asthmatic children. Environ. HealthPerspect. 117, 668–674.
Li, X.Y., Gilmour, P.S., Donaldson, K., MacNee, W., 1996. Free radical activity andpro-inflammatory effects of particulate air pollution (PM10) in vivo and in vitro.Thorax 51, 1216–1222.
Mendelson, C.R., 2000. Role of transcription factors in fetal lung development andsurfactant protein gene expression. Annu. Rev. Physiol. 62, 875–915.
Miguel, F., Augusto, A.C., Gurgueira, S.A., 2009. Effect of acute vs. chronic H2O2-induced oxidative stress on antioxidant enzyme activities. Free Radic. Res. 43,340–347.
Miller, M.R., Borthwick, S.J., Shaw, C.A., McLean, S.G., McClure, D., Mills, N.L., Duf-fin, R., Donaldson, K., Megson, I.L., Hadoke, P.W., Newby, D.E., 2009. Directimpairment of vascular function by diesel exhaust particulate through reducedbioavailability of endothelium-derived nitric oxide induced by superoxide freeradicals. Environ. Health Perspect. 117, 611–616.
Morissette, M.C., Vachon-Beaudoin, G., Parent, J., Chakir, J., Milot, J., 2008. Increasedp53 level, Bax/Bcl-x(L) ratio, and TRAIL receptor expression in human emphy-sema. Am. J. Respir. Crit. Care Med. 178, 240–247.
Moriya, M., Ho, Y.H., Grana, A., Nguyen, L., Alvarez, A., Jamil, R., Ackland, M.L., Michal-czyk, A., Hamer, P., Ramos, D., Kim, S., Mercer, J.F., Linder, M.C., 2008. Copper istaken up efficiently from albumin and alpha2-macroglobulin by cultured humancells by more than one mechanism. Am. J. Physiol. Cell. Physiol. 295, C708–C721.
Nemmar, A., Hoylaerts, M.F., Hoet, P.H., Nemery, B., 2004. Possible mechanismsof the cardiovascular effects of inhaled particles: systemic translocation andprothrombotic effects. Toxicol. Lett. 149, 243–253.
Nurkiewicz, T.R., Porter, D.W., Barger, M., Millecchia, L., Rao, K.M., Marvar, P.J., Hubbs,A.F., Castranova, V., Boegehold, M.A., 2006. Systemic microvascular dysfunctionand inflammation after pulmonary particulate matter exposure. Environ. HealthPerspect. 114, 412–419.
Oberdorster, G., Sharp, Z., Atudorei, V., Elder, A., Gelein, R., Kreyling, W., Cox, C.,2004. Translocation of inhaled ultrafine particles to the brain. Inhal. Toxicol. 16,437–445.
Osornio-Vargas, A.R., Bonner, J.C., Alfaro-Moreno, E., Martínez, L., García-Cuellar,C., Ponce-de-León Rosales, S., Miranda, J., Rosas, I., 2003. Proinflammatory andcytotoxic effects of Mexico City air pollution particulate matter in vitro aredependent on particle size and composition. Environ. Health Perspect. 111,1289–1293.
Pope III, C.A., Burnett, R.T., Thun, M.J., Calle, E.E., Krewski, D., Ito, K., Thurston, G.D.,2002. Lung cancer, cardiopulmonary mortality, and long-term exposure to fineparticulate air pollution. JAMA 287, 1132–1141.
Ramadas, R.A., Wu, L., LeVine, A.M., 2009. Surfactant protein A enhances productionof secretory leukoprotease inhibitor and protects it from cleavage by matrixmetalloproteinases. J. Immunol. 182, 1560–1567.
Ryter, S.W., Kim, H.P., Hoetzel, A., Park, J.W., Nakahira, K., Wang, X., Choi, A.M., 2007.Mechanisms of cell death in oxidative stress. Antioxid. Redox Signal. 9, 49–89.
Sanchez-Perez, Y., Chirino, Y.I., Osornio-Vargas, A.R., Morales-Barcenas, R.,Gutierrez-Ruiz, C., Vazquez-Lopez, I., Garcia-Cuellar, C.M., 2009. DNA damageresponse of A549 cells treated with particulate matter (PM10) of urban air pol-lutants. Cancer Lett. 278, 192–200.
Sint, T., Donohue, J.F., Ghio, A.J., 2008. Ambient air pollution particles and the acuteexacerbation of chronic obstructive pulmonary disease. Inhal. Toxicol. 20, 25–29.
Smith, P.K., Krohn, R.I., Hermanson, G.T., Mallia, A.K., Gartner, F.H., Provenzano, M.D.,Fujimoto, E.K., Goeke, N.M., Olson, B.J., Klenk, D.C., 1985. Measurement of proteinusing bicinchoninic acid. Anal. Biochem. 150, 76–85.
Song, L., Zhang, B., Feng, Y., Luo, X., Wei, X., Xiao, X., 2009. A role for forkhead boxA1 in acute lung injury. Inflammation 32, 322–332.
Valavanidis, A., Vlahoyianni, T., Fiotakis, K., 2005. Comparative study of the for-mation of oxidative damage marker 8-hydroxy-2′-deoxyguanosine (8-OHdG)adduct from the nucleoside 2′-deoxyguanosine by transition metals and sus-pensions of particulate matter in relation to metal content and redox reactivity.Free Radic. Res. 39, 1071–1081.
Wan, X.S., Zhou, Z., Ware, J.H., Kennedy, A.R., 2005. Standardization of a fluoro-metric assay for measuring oxidative stress in irradiated cells. Radiat. Res. 163,232–240.
Wang, T., Chiang, E.T., Moreno-Vinasco, L., Lang, G.D., Pendyala, S., Samet, J.M.,Geyh, A.S., Breysse, P.N., Chillrud, S.N., Natarajan, V., Garcia, J.G., 2009. Partic-ulate matter disrupts human lung endothelial barrier integrity via ROS- andp38 MAPK-dependent pathways. Am. J. Respir. Cell Mol. in press.
Whitekus, M.J., Li, N., Zhang, M., Wang, M., Horwitz, M.A., Nelson, S.K., Horwitz,L.D., Brechun, N., Diaz-Sanchez, D., Nel, A.E., 2002. Thiol antioxidants inhibit theadjuvant effects of aerosolized diesel exhaust particles in a murine model forovalbumin sensitization. J. Immunol. 168, 2560–2567.
Winterbourn, C.C., 1995. Toxicity of iron and hydrogen peroxide: the Fenton reac-tion. Toxicol. Lett. 82–83, 969–974.
Related Documents











