GASTROENTEROLOGY 1995;108:554-563 SPECIAL REPORTS AND REVIEWS Plurichemical Transmission and Chemical Coding of Neurons in the Digestive Tract JOHN B. FURNESS, HEATHER M. YOUNG, SUELI POMPOLO, JOEL C. BORNSTEIN, WOLFGANG A. A. KUNZE, and KAREN MCCONALOGUE Department of Anatomy and Cell Biology, University of Melbourne, Parkville, Australia The enteric nervous system contains neurons with well- defined functions. However, when neurons of the same function are examined in different regions or species, they are found to show subtle differences in their phar- macologies of transmission and different chemical cod- ing. Individual enteric neurons use more than one trans- mitter, i.e., transmission is plurichemical. For example, enteric inhibitory neurons have three or more primary transmitters, including nitric oxide, vasoactive intesti- nal peptide, and possibly adenosine triphosphate and pituitary adenylyl cyclase activating peptide. Primary transmitters are highly conserved, although their rela- tive roles vary considerably between gut regions. Multi- ple substances, including transmitters and their syn- thesizing enzymes and nontransmitters (such as neurofilament proteins), provide neurons with a chemi- cal coding through which their functions and projec- tions can be identified. Although equivalent neurons in different regions have the same primary transmitters, other chemical markers differ substantially. Caution must be taken in extrapolating pharmacological and neurochemical observations between species or even between regions in the one species. On the other hand, careful interregion and interspecies comparisons lead to an understanding of the features of enteric neurons that are highly conserved and can be used in valid extrapolation. S ometimes in the course of scientific exploration, ob- servations can accumulate so rapidly that we fumble to assimilate them into a coherent view. We encounter such a period now; the number of possible neurotransmit- ters in peripheral neurons seems to exceed necessity by a large margin. Although it might be hypothesized that only two transmitters are necessary, one for excitation and one for inhibition, or perhaps that even one transmit- ter and two receptors is enough, about 30 transmitter candidates have been discovered in the enteric nervous system.* It has been difficult to make sense of the experi- mental data. Daniel et al. 2'~ and Fox-Threlkeld,4 for ex- ample, have drawn attention to the variations in re- sponses of intestinal preparations from differing regions and species to the same agonists and the substantial dif- ferences in responses to the same agonists delivered by different routes. Others have pointed out that the pres- ence of possible neurotransmitters in enteric neurons is inconsistent between species and regions) For example, neurons that are immunoreactive for substance P inner- vate the mucosa in some species but not in others. 6 An- other example is the directions that neurons project; so- matostatin immunoreactivity (IR) is in neurons with anally projecting axons in the guinea pig small intestine and in orally projecting neurons in the large intestine of the same species/ A number of ideas that may assist in interpretation of the pharmacological and localization data have been developed. In summary, these are as follows. (1) The plurichemical transmission hypothesis. This proposition is that each neuron communicates by the release and action of combinations of messenger substances. Those familiar with blood clotting or the immune system (e.g., complement fixation) will be attuned to cell communica- tion involving multiple chemical interactions. Until re- cently, neurotransmitters usually have been thought to act alone. (2) The chemical coding hypothesis. The hy- pothesis is that combinations of chemical attributes, principally related to the transmitters used by neurons, correlate with the functions and locations of neurons. The codes of functionally equivalent neurons have some chemical features in common that generally relate to their primary transmitters (see below). (3) Primary trans- mitters and neuromodulators. It is suggested that differ- ent substances released from the one neuron serve differ- ent purposes. Among transmitter roles are those of primary transmitters, a primary transmitter being the Abbreviations used in this paper: ACh, acetylcholine; CHAT, choline acetyltransferase; IR, immunoreactivRy; PACAP, pituitary adenylyl cyclase activating peptide; TK, tachykinin(s). © 3.995 by the American Gastroenterological Association 0016-5085/95/$3.00

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

GASTROENTEROLOGY 1995;108:554-563
SPECIAL REPORTS AND REVIEWS
Plurichemical Transmission and Chemical Coding of Neurons in the Digestive Tract
JOHN B. FURNESS, HEATHER M. YOUNG, SUELI POMPOLO, JOEL C. BORNSTEIN, WOLFGANG A. A. KUNZE, and KAREN MCCONALOGUE Department of Anatomy and Cell Biology, University of Melbourne, Parkville, Australia
The enteric nervous system contains neurons with well- defined functions. However, when neurons of the same function are examined in different regions or species, they are found to show subtle differences in their phar- macologies of transmission and different chemical cod- ing. Individual enteric neurons use more than one trans- mitter, i.e., transmission is plurichemical. For example, enteric inhibitory neurons have three or more primary transmitters, including nitric oxide, vasoactive intesti- nal peptide, and possibly adenosine triphosphate and pituitary adenylyl cyclase activating peptide. Primary transmitters are highly conserved, although their rela- tive roles vary considerably between gut regions. Multi- ple substances, including transmitters and their syn- thesizing enzymes and nontransmitters (such as neurofilament proteins), provide neurons with a chemi- cal coding through which their functions and projec- tions can be identified. Although equivalent neurons in different regions have the same primary transmitters, other chemical markers differ substantially. Caution must be taken in extrapolating pharmacological and neurochemical observations between species or even between regions in the one species. On the other hand, careful interregion and interspecies comparisons lead to an understanding of the features of enteric neurons that are highly conserved and can be used in valid extrapolation.
S ometimes in the course of scientific exploration, ob- servations can accumulate so rapidly that we fumble
to assimilate them into a coherent view. We encounter such a period now; the number of possible neurotransmit- ters in peripheral neurons seems to exceed necessity by a large margin. Although it might be hypothesized that only two transmitters are necessary, one for excitation and one for inhibition, or perhaps that even one transmit- ter and two receptors is enough, about 30 transmitter candidates have been discovered in the enteric nervous system.* It has been difficult to make sense of the experi- mental data. Daniel et al. 2'~ and Fox-Threlkeld, 4 for ex- ample, have drawn attention to the variations in re-
sponses of intestinal preparations from differing regions and species to the same agonists and the substantial dif- ferences in responses to the same agonists delivered by different routes. Others have pointed out that the pres- ence of possible neurotransmitters in enteric neurons is inconsistent between species and regions) For example, neurons that are immunoreactive for substance P inner- vate the mucosa in some species but not in others. 6 An- other example is the directions that neurons project; so- matostatin immunoreactivity (IR) is in neurons with anally projecting axons in the guinea pig small intestine
and in orally projecting neurons in the large intestine of the same species/
A number of ideas that may assist in interpretation of the pharmacological and localization data have been developed. In summary, these are as follows. (1) The plurichemical transmission hypothesis. This proposition is that each neuron communicates by the release and action of combinations of messenger substances. Those familiar with blood clotting or the immune system (e.g., complement fixation) will be attuned to cell communica- tion involving multiple chemical interactions. Until re-
cently, neurotransmitters usually have been thought to act alone. (2) The chemical coding hypothesis. The hy- pothesis is that combinations of chemical attributes, principally related to the transmitters used by neurons, correlate with the functions and locations of neurons. The codes of functionally equivalent neurons have some chemical features in common that generally relate to their primary transmitters (see below). (3) Primary trans- mitters and neuromodulators. It is suggested that differ- ent substances released from the one neuron serve differ- ent purposes. Among transmitter roles are those of primary transmitters, a primary transmitter being the
Abbreviations used in this paper: ACh, acetylcholine; CHAT, choline acetyltransferase; IR, immunoreactivRy; PACAP, pituitary adenylyl cyclase activating peptide; TK, tachykinin(s).
© 3.995 by the American Gastroenterological Association 0016-5085/95/$3.00

February 1995 ENTERIC NEUROTRANSMITTERS 555
substance (or substances) that has the major role in
changing excitability of effector cells, and those of neuro- modulators, substances that modify transmission, for ex- ample, by adjusting transmitter release or changing the
efficacy of primary transmitters. The ideas that are expressed in this report have evolved
over a period of t ime 8-12 and are almost certain to change
as new discoveries are made. Meanwhile, they provide a
framework for considering the significance of existing data in relation to neural function in the enteric nervous
system and elsewhere.
In 1877, Du Bois Reymond reported that transmission from neurons was likely to be either electrical or chemi-
cal. 13 It took many years before adequate experiments
could be performed, but by the early 1950s, it became accepted that mammalian neurons communicate chemi-
cally. 14 At that time, only two peripheral transmitters,
norepinephrine (noradrenaline) and acetylcholine (ACh), were identified, and the consensus seemed to be that
transmission was either adrenergic or cholinergic. This view remained so strong that the defining acronym for an undetermined transmitter has become nonadrenergic,
noncholinergic. The realization that nonadrenergic, non- cholinergic transmission occurred in the periphery came
substantially from studies of transmission to gastrointes- tinal muscle. 15-17 In the period from the mid-1960s up to the present, substantial effort has been expended to identify nonadrenergic, noncholinergic transmitters in
the enteric nervous system, in most cases without consen- sus being reached. Most recently, it has been recognized that transmission from each neuron involves several chemicals. Such plurichemical transmission, explored for the inhibitory motor neurons, helps explain the difficulty in establishing transmitter identification.
Whereas pharmacological studies, to mimic or inter- fere with transmission, remain the most powerful means
to identify transmitters, histochemical methods have been instrumental in showing that the transmitters are produced by the relevant neurons and that the same neurons contain several potential transmitters. Moreover, the chemical coding shown by histochemical studies has been crucial for the analysis of the organization of nerve circuits.
To show the ideas that are presented in this review, we have concentrated on three groups of neurons: excitatory neurons to the muscle, inhibitory neurons to the muscle, and secretomotor neurons. Possible transmitters of other enteric neurons have been reviewed elsewhere. ~
Defining Transmitters and the Transmission Process
Chemical transmission is the process by which substances are released from nerve terminals and act on
effector cells to produce changes in the physiological or
biochemical state of the effector. Each substance that is
released from the nerve ending to affect cells that it innervates is a transmitter; the responses of the postjunc-
tional cells depend on the ways that transmitters or their
effects interact. This concept says nothing about the t ime
course of the effect; long-term effects, even lasting a day
or more, must be considered part of the transmission
process. However, only short-term effects, lasting milli- seconds to seconds, are considered in this review.
The transmitters with which we are most familiar
cause transient changes in the excitability of the inner-
vated cells. A good example is the primary transmitter,
ACh, released at the skeletal neuromuscular junction. As
is explained below, most enteric neurons use more than
one primary transmitter. In some cases, substances re-
leased from axons seem to have minor influences on
effector cells. For example, some blood vessels are
innervated by vasoconstrictor neurons that contain neuro-
peptide Y but use norepinephrine and adenosine triphos- phate (ATP) as primary transmitters, re-2° At some of
these neuroeffector junctions, neuropeptide Y has mini-
mal postjunctional effect and can be regarded as a second-
ary transmitter. Neurotransmitters may act to modify
the release or action of the primary transmitter; when
they act in this way, they are called neuromodulators.
Neuromodulators can act postsynaptically, as vasoactive
intestinal polypeptide (VIP) does on the salivary gland
cells of the cat, where it substantially increases the effec-
tiveness of ACh, the primary transmitter, in promoting secretion. 21'22 Presynaptic modulation is also common.
In this case, a neuromodulator acts on the nerve terminal
to modify transmitter release, usually reducing transmit-
ter output. These concepts of primary transmitters, sec-
ondary transmitters, and neuromodulators are still evolv-
ing. The distinction between the classes is not rigid, as
will be clear from the discussion below. Furthermore, a
transmitter can have more than one role at a neuroeffector
junction as norepinephrine does at many junctions where it is a primary transmitter and also has presynaptic, mod- ulatory effects. 12
A rather surprising outcome of the investigation of possible neurotransmitters is that, in some instances, no
function can be attributed to a biologically active sub- stance in a particular neuron. An example is substance P in preganglionic fibers supplying the amphibian heart. 23
Also surprising are cases in which a potential secondary transmitter or neuromodulator has an action opposite that of the primary transmitter, t° B o w e r s 24 has argued
that inactive substances should be expected due to imper- fect control of gene expression. He calls neurotransmit- ters that fall into this class superfluous transmitters.

556 FURNESS ET AL. GASTROENTEROLOGY Vol. 108, No. 2
The Use of Chemical Coding
The observation that neurons can be distinguished by their chemical code has been helpful in analysis of neural circuits. For example, the coding of vasomotor neurons allowed Gibbins and Morris 25 to show that suc-
cessive segments of the arterial tree are separately inner- vated. Chemical coding also showed that noradrenergic neurons that control intestinal secretion are separate from those that control motili ty or arterial resistance. 8 Immu-
noreactivity for calbindin in the guinea pig small intes- tine has allowed the projections of enteric sensory neurons to be deduced, even though the reason why calbindin is
specifically contained in these neurons is unknown.
Transmitters of Inhibitory Motor Neurons
The identity of the transmitter of these neurons has been hotly debated. Burnstock et al. 16 produced evi-
dence to suggest that ATP is the transmitter; a few years later, evidence for VIP was reported. 26'27 More recent
evidence for the involvement of nitric oxide in this trans- mission is compelling. 28 Contribution to transmission by
pituitary adenylyl cyclase activating peptide (PACAP) is also possible, 29'3° as is the participation of carbon monox- ide. 31
An explanation for the differences encountered by in- vestigators is that there are at least two, and in many regions three, primary transmitters of these neurons and that the relative roles of the primary transmitters differ between regions and species. Stimulation of the inhibi- tory neurons causes relaxation and hyperpolarization of the muscle as shown by mechanical recording, electrical recording, and pharmacological analysis. Mechanical re- cording, by virtue of its simplicity, has allowed a broad range of preparations to be surveyed, but it lacks the temporal resolution that can be achieved with intracellu- lar microelectrodes.
Electrophysiological evidence for more than one inhib- itory transmitter was obtained by Niel et al., 32 who dis-
tinguished two responses to inhibitory nerve stimulation in the guinea pig small intestine: a fast hyperpolarization that is blocked by apamin, which blocks a class of small conductance potassium channels, and a slower hyperpo- larization that is resistant to this drug. This observation was confirmed in the same tissue by Crist et al., 33 who reported the fast component to be partly reduced by desensitization with o~,~-methylene ATP and the slow component to be blocked by the antagonist of VIP recep- tors, VIP 10-28 . Crist et al. 33 concluded that ATP and VIP are cotransmitters and that other substances may be involved because pharmacological antagonism was in-
complete. It was subsequently reported, also in guinea pig small intestine, that the slow component is blocked by inhibition of N O synthase. 34'35 Because the effect of
VIP was also reduced by N O synthase inhibition, He and Goya134 concluded that VIP acts indirectly, i.e., VIP
causes N O to be produced in the tissue. Further evidence for an indirect action of VIP being mediated by NO release was provided by Grider et al. 36 However, in some
gut muscle, VIP seems to act directly in that its action is not inhibited by blocking NO synthase. 37 Grider et al. 36 also showed that NO amplifies VIP release.
The conclusion that N O contributes only to slow com- ponents of transmission cannot be extrapolated; the in- hibitory nerves of canine proximal colon cause fast hyper- polarizing potentials that are blocked by NO synthase inhibition. 38 N O could be acting directly on the muscle
of the canine colon or could be evoking the release of another substance. Although apamin seems to be selec- tive for the non-NO component of transmission in the
guinea pig intestine, this is not the case in the dog small intestine, where apamin antagonizes both the action of NO and the NO synthase-sensitive component of inhib- itory transmission. 39 Conversely, in the circular muscle
of the opossum esophagus, apamin does not affect the fast hyperpolarization caused by nerve stimulation. 4°
Pharmacological analysis of relaxations also provides evidence for plurichemical transmission and confirms that the relative roles of transmitters differ substantially between regions. 8'41-45 Variable contributions of different
transmitters are suggested by variations in the extent by which a component of transmission from the inhibitory
motor neurons is blocked by apamin between gut regions in the guinea pig. 4I'46 For example, inhibitory transmis-
sion to the taeniae coli is almost completely blocked by apamin, whereas transmission to the longitudinal muscle of the proximal colon of the same species is almost fully resistant. 41'46 Investigations of transmission in the rat
intestine indicated that ATP could contribute signifi- cantly to transmission in the duodenum but not in the ileum. 42 The degree of contribution of N O to transmis-
sion also varies; antagonism of NO-mediated transmis- sion is effective in reducing inhibitory transmission in many gut regions but is poorly effective in the taenia. 36'4v
Similarly, Suthamnatpong et al. 4~ found that inhibition of NO-mediated transmission is effective in the proximal colon but not in the distal colon of the rat. Conversely, VIP receptor antagonism is effective in the distal but not in the proximal colon. In another region, the circular muscle of the lower esophagus of the opossum, ATP does not seem to contribute to transmission, whereas VIP and N O probably do. 27'4°'48 Pharmacological studies in the gastric fundus of the rat also point to N O and VIP being

February 1995 ENTERIC NEUROTRANSMITTERS 557
cotransmitters. 4v'49 A third component of the inhibitory
response in the same muscle may be mediated by ATP because the response is reduced by desensitization with oq~-methylene ATP. 5° Three pharmacologically distin- guishable components of inhibitory transmission to the circular muscle of the guinea pig colon have also been
reported; one sensitive to NO synthase inhibition, one sensitive to apamin, and one insensitive to combined apamin and NO synthase inhibition. 45
Although VIP has been referred to as a possible trans- mitter, the closely related peptide, peptide histidine iso- leucine, is produced by the same gene; it is localized histochemically in the same neurons, and it also relaxes the muscle. Thus, peptide histidine isoleucine and VIP should be considered together as possible transmitters. 51
A further compound, PACAP, needs to be included as a possible inhibitory transmitter. This peptide is found in axons that innervate the muscle throughout the gastro- intestinal tract. 52 In the taeniae coli, these have been
shown to be the axons of inhibitory motor neurons to the muscle. 3° The direct action of PACAP on gastrointestinal muscle is to cause relaxation. However, electrophysiolog-
ical evidence for PACAP being responsible for the mem- brane hyperpolarization associated with inhibitory trans- mission is lacking. A neurotransmitter role also needs to be considered for CO that relaxes the internal anal sphincter in the opossum; moreover, when the inhibitory nerves are stimulated, the levels of the enzyme for CO synthesis, heine oxygenase, increases. 3. Inhibitory trans- mission was antagonized when heme oxygenase was in- hibited with zinc protoporphyrin IX. It is puzzling that
the inhibitory action of VIP was also antagonized; this may suggest that the action of zinc protoporphyrin is not specific for heme oxygenase.
In conclusion, the enteric inhibitory neurons do not have a single transmitter. It is probable that each of ATP, NO, peptide histidine isoleucine/VIP, and PACAP contributes to inhibitory transmission to gastrointestinal muscle but to differing extents depending on region and species. Moreover, significant interplay occurs between the transmitters in that they can influence each other's release and ac t ion . 34'36'53
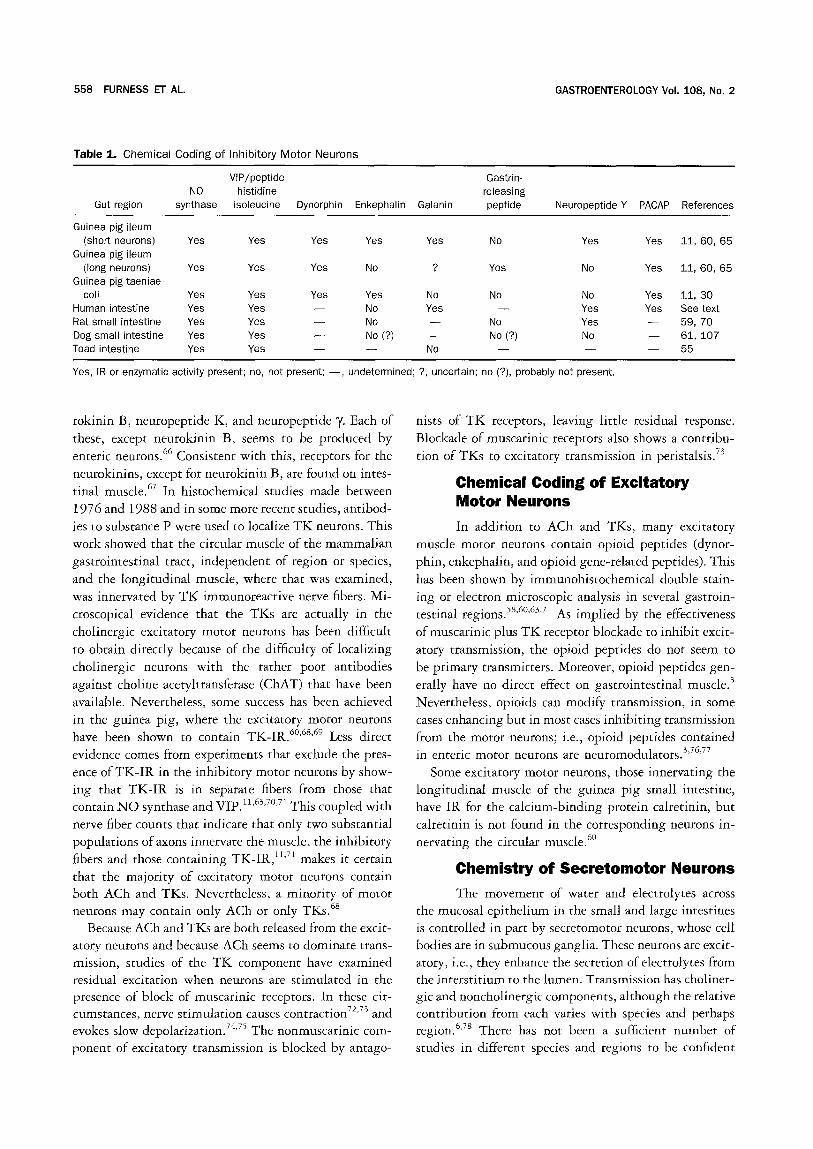
Chemical Coding of Inhibitory Motor Neurons
In this context, aspects of the chemistry that can be revealed histochemically are considered. In a wide range of vertebrates, IR for VIP is in nerve fibers in- nervating the muscle (we are unaware of any situation in which it has been investigated and not been found), and N O synthase, revealed either by immunohistochem- istry or by histochemical localization of reduced nicotin-
amide adenine dinucleotide phosphate diaphorase, is also present.54 56 That these two inhibitory transmitters are
in the same fibers innervating the muscle can be deduced from double labeling experiments in guinea pig small intestine and t aen iae coli, 11'57'58 rat small intestine, 59 and
toad intestine) 5 Both pharmacological evidence and sin- gle labeling histochemistry suggests that this will prove
to be the case elsewhere. In contrast, other chemical attributes of the inhibitory neurons are highly variable (Table 1). In the small intestine of the guinea pig, inhibi- tory neurons with short projections along the intestine
have IR for enkephalin and neuropeptide Y, whereas neurons with long projections have gastrin-releasing pep- tide and dynorphin-IR. .1'6° However, in the taenia of the
guinea pig colon, the neurons contain both dynorphin and enkephalin-IR but do not show gastrin-releasing peptide or neuropeptide Y - I R . 5s In the large intestine
of the guinea pig and in the canine small intestine, neuro- peptide Y - I R nerve fibers project orally to supply the circular muscle; it is thus likely that the excitatory rather
than the inhibitory innervation is neuropeptide Y - I R in these regions. 7'61 In the rat and human intestine, neu-
ropeptide Y is colocalized with VIP in presumed inhibi- tory neurons that supply the muscle. 62'63 In the human
taeniae coli, galanin coexists with VIP in the inhibitory 64 neurons.
IR for PACAP is encountered in fibers innervating gastrointestinal muscle throughout the digestive tract of several species) 2'65 In the taenia of the guinea pig cecum,
PACAP-IR is in the N O synthase/VIP inhibitory nerve
fibers and in the small intestine and colon; the projections of the PACAP-IR neurons correspond to the projections of the inhibitory motor neurons. 3°'65 Thus, PACAP-IR
may be a marker of some inhibitory motor neurons. In conclusion, the chemical code of enteric inhibitory
motor neurons always seems to include N O synthase and peptide histidine isoleucine/VIP, whereas other markers that occur in these neurons do so in an apparently capri-
cious way and cannot be regarded as useful in predicting the function of any immunoreactive neurons without fur- ther data. PACAP is possibly a third marker of the inhib- itory motor neurons.
Transmitters of Excitatory Motor Neurons
There is incontrovertible evidence, extensively re- viewed in the past, that ACh is a primary transmitter of excitatory neurons that innervate gastrointestinal mus- cle. 2'41 However, these neurons also release tachykinins (TKs), which are direct excitants of the muscle and also contribute to transmission. The TKs are a group of closely related peptides: substance P, neurokinin A, neu-
558 FURNESS ET AL, GASTROENTEROLOGY Vol. 108 , No. 2
Table 1. Chemical Coding o f Inhibi tory Mo to r Neurons
VIP/peptide Gastrin- NO histidine releasing
Gut region synthase isoleucine Dynorphin Enkephalin Galanin peptide Neuropeptide Y PACAP References
Guinea pig ileum (short neurons) Yes Yes Yes Yes Yes No
Guinea pig ileum (long neurons) Yes Yes Yes No ? Yes
Guinea pig taeniae coil Yes Yes Yes Yes No No
Human intestine Yes Yes - - No Yes - - Rat small intestine Yes Yes - - No - - No Dog small intestine Yes Yes - - No (?) - - No (?) Toad intestine Yes Yes - - - - No - -
Yes Yes 11, 60, 65
No Yes 1 1 , 6 0 , 6 5
No Yes 11, 30 Yes Yes See t e~ Yes - - 59, 70 No - - 6 1 , 1 0 7 - - - - 55
Yes, IR or enzymatic activity present; no, not present; - - , undetermined; ?, uncertain; no (?), probably not present.
rokinin B, neuropeptide K, and neuropeptide 7. Each of these, except neurokinin B, seems to be produced by enteric neurons. 66 Consistent with this, receptors for the
neurokinins, except for neurokinin B, are found on intes- tinal muscle. 6v In histochemical studies made between 1976 and 1988 and in some more recent studies, antibod- ies to substance P were used to localize T K neurons. This work showed that the circular muscle of the mammalian gastrointestinal tract, independent of region or species,
and the longitudinal muscle, where that was examined, was innervated by T K immunoreactive nerve fibers. Mi- croscopical evidence that the TKs are actually in the cholinergic excitatory motor neurons has been difficult
to obtain directly because of the difficulty of localizing cholinergic neurons with the rather poor antibodies against choline acetyltransferase (CHAT) that have been available. Nevertheless, some success has been achieved in the guinea pig, where the excitatory motor neurons have been shown to contain TK-IR. 6°'68'69 Less direct
evidence comes from experiments that exclude the pres- ence of TK-IR in the inhibitory motor neurons by show- ing that TK-IR is in separate fibers from those that contain NO synthase and VIP. 11'63'v°'vl This coupled with
nerve fiber counts that indicate that only two substantial populations of axons innervate the muscle, the inhibitory fibers and those containing TK-IR, 11'vl makes it certain
that the majority of excitatory motor neurons contain both ACh and TKs. Nevertheless, a minority of motor neurons may contain only ACh or only TKs. 68
Because ACh and TKs are both released from the excit- atory neurons and because ACh seems to dominate trans- mission, studies of the TK component have examined residual excitation when neurons are stimulated in the presence of block of muscarinic receptors. In these cir- cumstances, nerve stimulation c a u s e s c o n t r a c t i o n 72'73 and evokes slow depolarization. 74'v5 The nonmuscarinic com-
ponent of excitatory transmission is blocked by antago-
nists of TK receptors, leaving little residual response. Blockade of muscarinic receptors also shows a contribu- tion of TKs to excitatory transmission in peristalsis, v3
Chemical Coding of Excitatory Motor Neurons
In addition to ACh and TKs, many excitatory muscle motor neurons contain opioid peptides (dynor- phin, enkephalin, and opioid gene-related peptides). This
has been shown by immunohistochemical double stain- ing or electron microscopic analysis in several gastroin- testinal regions. 5s'6°'63'71 As implied by the effectiveness
of muscarinic plus TK receptor blockade to inhibit excit- atory transmission, the opioid peptides do not seem to be primary transmitters. Moreover, opioid peptides gen- erally have no direct effect on gastrointestinal muscle) Nevertheless, opioids can modify transmission, in some cases enhancing but in most cases inhibiting transmission from the motor neurons; i.e., opioid peptides contained in enteric motor neurons are neuromodulators. 3'76'77
Some excitatory motor neurons, those innervating the longitudinal muscle of the guinea pig small intestine, have IR for the calcium-binding protein calretinin, but calretinin is not found in the corresponding neurons in- nervating the circular muscle. 6°
Chemistry of Secretomotor Neurons
The movement of water and electrolytes across the mucosal epithelium in the small and large intestines is controlled in part by secretomotor neurons, whose cell bodies are in submucous ganglia. These neurons are excit- atory, i.e., they enhance the secretion of electrolytes from the interstitium to the lumen. Transmission has choliner- gic and noncholinergic components, although the relative contribution from each varies with species and perhaps region. <v8 There has not been a sufficient number of studies in different species and regions to be confident

February 1995 ENTERIC NEUROTRANSMITTERS 559
Table 2. Chemical Coding of Submucous Neurons
Proportion Presumed Species Code (%) function
Guinea pig s±'84'8~'Sr Dynorphin/galanin/neuromedin U/peptide histidine isoleucine/VIP 45 Secretomotor ChAT/cholecystokinin/calcitonin gene-related peptide/dynorphin/_+galanin/
neuromedin U/neuropeptide Y/somatostatin 30 ChAT/(calbindin)/dynorphin/neuromedin U/substance P 10 ChAT/calretinin/dynorphin 15 Calbindin/neuropeptide Y/peptide histidine isoleucine/VIP 40 Calcitonin gene-related peptide/somatostatin/±substance P 18 Calcitonin gene-related peptide/substance P/neurofiiament protein 25 Neurofilament protein _+ substance P 15 Galanin 2
Rat88,89
Secretomotor Sensory Unknown Secretomotor Unknown Unknown Unknown Unknown
that all components of transmission are known or whether the cholinergic and noncholinergic neurons are separate. However, studies of transmitter presence, trans- mitter release, and pharmacological mimicry strongly implicate VIP as a primary noncholinergic transmitter, v9
The neurons of the submucosal plexus ganglia in guinea pig and rat small intestine provide an interesting study of differences of chemical coding between species. In these species, in contrast to larger animals such as pigs and humans, the submucosal plexus ganglia proba- bly only contain cell bodies of sensory neurons and neu- rons involved in secretomotor control. 8° In the guinea
pig, there are only four types of nerve cells in submucous ganglia (Table 2). 81'82 Neurons that contain ChAT and
substance P and react with some antibodies to calbindin are thought to be primary sensory neurons. 82-84 Choliner-
gic secretomotor neurons have the chemical code CHAT/
cholecystokinin/calcitonin gene-related peptide/dynor- phin/neuromedin U/neuropeptide Y/somatostatin (about 70% of these contain galanin), whereas the noncholiner- gic secretomotor neurons are VIP/dynorphin/galanin/ neuromedin U. 4t'81,sS-sv The fourth group has the coding
ChAT/calretinin/dynorphin. s1'85 In the rat small intes-
tine, neurons subserving the same functions are presumed to exist, but the chemical coding of the nerve cells is quite different 62'88'89 (Table 2).
Human Enter ic Neurons
The organization of the human and other mam- malian enteric nervous systems and the physiological control that enteric neurons exert over motility, secre- tion, and other digestive processes in different species are similar. 9°-92 In particular, drugs that modify neuro- transmission have similar effects in humans and other mammals. For example, muscarinic antagonists reduce acid secretion and suppress motili ty in humans and labo- ratory animals. Furthermore, the recent discovery that N O is an enteric inhibitory transmitter has been con- firmed pharmacologically in preparations of human gas-
trointestinal tract. 93 96 Thus, it can be anticipated that
the primary transmitters of human enteric neurons are the same as in other mammals. Based on comparative
studies (reviewed above), it might also be expected that substances that contribute to the chemical codes of hu- man enteric neurons will have different patterns of colo- calization from those of other species.
Muscle Motor Neurons
The two markers of motor neurons to gut muscle in other mammals, TKs and VIP, are also contained in neurons innervating the muscle throughout the human gastrointestinal tract. 54 As in other species, VIP-IR nerve
fibers project anally in human in tes t ines T K and VIP- IR are in separate fibers as is also the case in other mam- mals. 63 VIP relaxes human gastrointestinal muscle, whereas TKs cause it to contract.98-loo These observations
imply that TKs mark the excitatory neurons and VIP marks the inhibitory neurons. 63'97 The effectiveness of
muscarinic antagonists in inhibiting excitation and of
N O synthase antagonism in reducing inhibitory trans- mission indicates that the excitatory neurons are cholin- ergic and that the inhibitory neurons contain NO syn-
thase. The presence of N O synthase in nerve fibers innervating the muscle has been confirmed histochemi- cally. %'*°1-1°3 Double labeling has shown that the TK-
IR nerve fibers are immunoreactive for enkephalin and that many of the VIP-IR nerve fibers are immunoreactive for neuropeptide Y and galanin. 63'64 It has been recently
shown that PACAP-IR is also colocalized with VIP-IR in fibers innervating the muscle of the human intes- tine. t°4 Thus, the coding of excitatory muscle motor neu- rons in human intestine can be deduced to be CHAT/ TK/enkephalin and of inhibitory muscle motor neurons to be NO synthase/PACAP/VIP/+galanin/+neuropep - tide Y. As in other species, there may be regional differ- ences in chemical coding; such differences, if they occur in humans, are yet to be identified. It seems that the coding N O synthase/PACAP/VIP for inhibitory motor

560 FURNESS ET AL. GASTROENTEROLOGY Vol. 108, No. 2
neurons and ChAT/TK for excitatory motor neurons is
conserved among mammalian species.
Secretomotor Neurons
The only consistent marker of secretomotor neu-
rons that has been identified in mammals is VIP. VIP-
IR fibers, which are presumed to have a secretomotor
function, provide a rich innervation of the mucosa of the human small and large intestine. 1°5 By analogy with
other species, these would be expected to be noncholiner-
gic secretomotor neurons. However, there are also cholin-
ergic secretomotor neurons supplying the human mu- cosa, 1°6 but the chemical coding of these neurons is
unknown.
Conclusions
A small part of the extensive literature on the
enteric nervous system has been selected for review to
show that substantial variations in the chemistries of
neurons that have the same functions are encountered
when neurons of different regions and species are com-
pared, even though the primary transmitters are well
conserved between species. It is concluded that transmis-
sion from each enteric neuron involves the release of
several substances, although the relative roles of these
mult iple transmitters vary with region and species. Some
of the substances whose presence distinguishes popula-
tions of neurons are neuromodulators, not primary trans- mitters, or may even have no role in transmission. The
recognition and exploitation of chemical coding of neu-
rons has been instrumental in deciphering neural cir-
cuitry within the intestine and elsewhere in the auto- nomic nervous system. ~2'4I'6°'83
Refe rences
1. McConalogue K, Furness JB. Gastrointestinal neurotransmit- ters. In: Fuller P, Shulkes A, eds. The gut as an endocrine organ. London: Bailli~re Tindall, 1994 :51-76 .
2. Daniel EE, Collins SM, Fox JET, Huizinga JD. Pharmacology of drugs acting on gastrointestinal motility. In: Wood JD, ed. Hand- book of physiology, the gastrointestinal system. Washington: American Physiological Society, 1989 :715-757 .
3. Daniel EE, Collins SM, Fox JET, Huizinga JD. Pharmacology of neuroendocrine peptides. In: Wood JD, ed. Handbook of physiol- ogy, the gastrointestinal system. Washington: American Physio- logical Society, 1989 :759-816 .
4. Fox-Threlkeld JET. Neuropeptide motor actions vary between in vivo and in vitro experimental conditions. In: Daniel EE, ed. Neuropeptide function in the gastrointestinal tract. Boca Raton, FL: CRC, 1991 :181-191 .
5. Ekblad E, H~kanson R, Sundler F. Microanatomy and chemical coding of peptide-containing neurons in the digestive tract. In: Daniel EE, ed. Neuropeptide function in the gastrointestinal tract. Boca Raton: CRC, 1991 :131-179 .
6. Keast JR. Mucosal innervation and control of water and ion transport in the intestine. Rev Physiol Biochem Pharmacol 1987; 109 :1 -59 .
7. Messenger JP, Furness JB. Projections of chemically-specified neurons in the guinea-pig colon. Arch Histol Cytol 1990; 53 :467-495 .
8. Costa M, Furness JB. Somatostatin is present in a subpopula- tion of noradrenergic nerve fibres supplying the intestine. Neuro- science 1984; 13 :911-919 .
9. Furness JB, Costa M, Morris JL, Gibbins IL. Novel neuro-trans- mitters and the chemical coding of neurons. In: McLennan H. ed. Advances in physiological research. New York: Plenum, 1987 :143-165 .
10. Furness JB, Morris JL, Gibbins IL, Costa M. Chemical coding of neurons and plurichemical transmission. Annu Rev Pharmacol Toxicol 1989 ;29 :289-306 .
11. Furness JB, Bornstein JC, Murphy R, Pompolo S. Roles of pep- tides in transmission in the enteric nervous system. Trends Neurosci 1992 ;15 :66 -71 .
12. Morris JL, Gibbins IL. Co-transmission and neuromodulation. In: Burnstock G, Hoyle CHV, eds. Autonomic neuroeffector mechanisms. Reading: Harwood Academic, 1992 :33-119 .
13. Du Bois Reymond E. Gesammelte Abhandlung der allgemeinen Muskel und Nervenphysik. Nervenphysik 1877;2:700.
14. Bacq ZM. Chemical transmission of nerve impulses. A historical sketch. Oxford: Pergamon, 1975.
15. Burnstock G, Campbell G, Bennett M, Holman ME. Innervation of the guinea-pig taenia coil: are there intrinsic inhibitory nerves which are distinct from sympathetic nerves? Int J Neuropharma- col 1964 ;3 :163-166 .
16. Burnstock G, Campbell G, Satchell D, Smythe A. Evidence that adenosine triphosphate or a related nucleotide is the transmit- ter substance released by non-adrenergic nerves in the gut. Br J Pharmacol 1970 ;40 :668-688 .
17. Campbell G. Autonomic nervous supply to effector tissues. In: BOibring E, Brading A, Jones A, Tomita T, eds. Smooth muscle. London: Arnold, 1970 :451-495 .
18. Pernow J. Co-release and functional interactions of neuropep- tide Y and noradrenaline in peripheral vascular control. Acta Physiol Scand 1988;133 (Suppl 568) :1 -56.
19. Potter EK. Neuropeptide Y as an autonomic neurotransmitter. Pharmacol Ther 1988; 37 :251-273 .
20. Kennedy C. ATP as a cotransmitter with noradrenaline in sympa- thetic perivascular nerves. In: Edvinsson L, Uddman R, eds. Vascular innervation and receptor mechanisms. New perspec- tives. New York: Academic, 1993; 187 -199 .
21. Lundberg JM, Anggard A, Fahrenkrug J. Complementary role of vasoactive intestinal polypeptide (ViP) and acetylcholine for cat submandibular gland blood flow and secretion. II1. Effects of local infusions. Acta Physiol Scand 1982 ;114 :329-337 .
22. Lundberg JM, Hedlund B, Bartfai T. Vasoactive intestinal poly- peptide enhances muscarinic ligand binding in cat submandibu- lar salivary gland. Nature 1982 ;295 :147-149 .
23. Bowers CW, Jan LY, Jan YN. A substance P-l ike peptide in bullfrog autonomic nerve terminals: anatomy biochemistry and physiology. Neuroscience 1986 ;19 :343-356 .
24. Bowers CW. Superfluous neurotransmitters? Trends Neurosci 1994 ;17 :315-320 .
25. Gibbins IL, Morris JL. Sympathetic noradrenergic neurons con- taining dynorphin but not neuropeptide Y innervate small cuta- neous blood vessels of guinea-pigs. J Auton Nerv Syst 1990 ;29 :137-150 .
26. Fahrenkrug J, Haglund U, Jodal M, Lundgren O, Olbe L, Schaffa- litzky de Muckadell O. Nervous release of vasoactive intestinal polypeptide in the gastrointestinal tract of cats: possible physio- logical implications. J Physiol (Lond) 1978 ;284 :291-305 .
27. Goyal RK, Rattan S, Said SI. VlP as a possible neurotransmitter of non-cholinergic non-adrenergic inhibitory neurons. Nature 1980; 288 :378-380 .
28. Stark ME, Szurszewski JH. Role of nitric oxide in gastrointestinal

February 1995 ENTERIC NEUROTRANSMITTERS 561
and hepatic function and disease. Gastroenterology 1992; 103 :1928-1949 .
29. SchwSrer H, Katsoulis S, Creutzfeldt W, Schmidt WE. Pituitary adenyiate cyclase activating peptide, a novel VIP-like gut-brain peptide, relaxes the guinea-pig taenia caeci via apamin-sensi- tive potassium channels. Naunyn Schmiedebergs Arch Pharma- col 1992 ;346 :511 -514 .
30. McConalogue K, Furness JB, Vremec MA, Hoist J J, Tornoe K, Marley PD. Histochemical, pharmacological, biochemical and chromatographic evidence that pituitary adenylyl cyclase acti- vating peptide is involved in inhibitory neurotransmission in the taenia of the guinea-pig caecum. J Auton Nerv Syst (in press).
31. Rattan S, Chakder S. Inhibitory effect of CO on internal anal sphincter: heme oxygenase inhibitor inhibits NANC relaxation. Am J Physiol 1993;265:G799-G804.
32. Niel JP, Bywater RAR, Taylor GS. Apamin-resistant post-stimulus hyperpoiarization in the circular muscle of the guinea-pig ileum. J Auton Nerv Syst 1983 ;9 :565-569 .
33. Crist JR, He XD, Goyal RK. Both ATP and the peptide VIP are inhibitory neurotransmitters in guinea-pig ileum circular muscle. J Physiol (Lond) 1992 ;447 :119 -131 .
34. He XD, Goyal RK. Nitric oxide involvement in the peptide VIP- associated inhibitory junction potential in the guinea-pig ileum. J Physiol (Lond) 1993 ;461 :485 -499 .
35. Lyster D J, Bywater RAR, Taylor GS, Watson MJ. Effects of a nitric oxide synthase inhibitor on non-cholinergicjunction potentials in the circular muscle of the guinea pig ileum. J Auton Nerv Syst 1992 ;41 :187 -196 .
36. Grider JR, Murthy KS, Jin J-G, Makhlouf GM. Stimulation of nitric oxide from muscle cells by VIP: prejunctional enhancement of VIP release. Am J Physiol 1992;262:G774-G778.
37. Tottrup A, Svane D, Forman A. Nitric oxide mediating NANC inhibition in opossum lower esophageal sphincter. Am J Physiol 1991; 260:G385-G389.
38. Dalziel HH, Thornbury KD, Ward SM, Sanders KM. Involvement of nitric oxide synthetic pathway in inhibitory junction potentials in canine proximal colon. Am J Physio11991; 260:G789-G792.
39. Bayguinov O, Vogalis F, Morris B, Sanders KM. Patterns of electrical activity and neural responses in canine proximal co- lon. Am J Physiol 1992; 193:G887-G894.
40. Daniel EE, Helmy-Elkholy A, Jager LP, Kannan MS. Neither a purine nor VIP is the mediator of inhibitory nerves of opossum esophageal smooth muscle. J Physiol (Lond) 1983 ;336 :243 - 260.
41. Furness JB, Costa M. The enteric nervous system. Edinburgh: Churchill Livingstone, 1987.
42. Manzini S, Maggi CA, Meli A. Pharmacological evidence that at least two different non-adrenergic non-cholinergic inhibitory systems are present in the rat small intestine. Eur J Pharmacol 1986; 123 :229-236 .
43. Grundy D, Gharib-Naseri MK, Hutson D. Role of nitric oxide and vasoactive intestinal polypeptide in vagally mediated relaxation of the gastric corpus in the anaesthetized ferret. J Auton Nerv Syst 1993 ;43 :241 -246 .
44. Suthamnatpong N, Hata F, Kanada A, Takeuchi T, Yagasaki O. Mediators of nonadrenergic, noncholinergic inhibition in the proximal, middle and distal regions of rat colon. Br J Pharmacol 1993; 108 :348-355 .
45. Maggi CA, Giuliani S. Multiple inhibitory mechanisms mediate non-adrenergic non-cholinergic relaxation in the circular muscle of the guinea-pig colon. Naunyn Schmiedebergs Arch Pharmacol 1993 ;347 :630 -634 .
46. Costa M, Furness JB, Humphreys CMS. Apamin distinguishes two types of relaxation mediated by enteric nerves in the guinea- pig gastrointestinal tract. Naunyn Schmiedebergs Arch Pharma- col 1986; 332 :79 -88 .
47. Li CG, Rand MJ. Nitric oxide and vasoactive intestinal polypep-
tide mediate non-adrenergic, non-cholinergic inhibitory trans- mission to smooth muscle of the rat gastric fundus. Eur J Phar- macol 1990; 191 :303-309 .
48. Yamato S, Spechler S J, Goyal RK. Role of nitric oxide in esopha- geal peristalsis in the opossum. Gastroenterology 1992; 103 :197-204 .
49. D'Amato M, Curro D, Montuschi P. Evidence for dual compo- nents in the non-adrenergic non-cholinergic relaxation in the rat gastric fundus: role of endogenous nitric oxide and vasoactive intestinal polypeptide. J Auton Nerv Syst 1992 ;37 :175-186 .
50. Belai A, Lefebvre RA, Burnstock G. Motor activity and neuro- transmitter release in the gastric fundus of streptozotocin-dia- betic rats. Eur J Pharmacol 1991; 194 :225-234 .
51. Grider JR, Makhlouf GM. Prejunctional inhibition of vasoactive intestinal peptide release. Am J Physiol 1987;253:G7-G12.
52. Sundler F, Ekblad E, Absood A, H&kanson R, K6ves K, Arimura A. Pituitary adenylate cyclase activating peptide: a novel vaso- active intestinal peptide-like neuropeptide in the gut. Neurosci- ence 1992 ;46 :439-454 .
53. Grider JR. Interplay of VlP and nitric oxide in regulation of the descending relaxation phase of peristalsis. Am J Physiol 1993; 264:G334-G340.
54. Furness JB, Lleweliyn-Smith I J, Bornstein JC, Costa M. Chemical neuroanatomy and the analysis of neuronal circuitw in the en- teric nervous system. In: Bjorklund A, H5kfelt T, Owman C, eds. Handbook of chemical neuroanatomy. Volume 6. Amsterdam: Elsevier, 1988 :161-218 .
55. Li ZS, Murphy S, Furness JB, Young HM, Campbell G. Relation- ships between nitric oxide synthase, vasoactive intestinal pep- tide and substance P immunoreactivities in neurons of the am- phibian intestine. J Auton Nerv Syst 1993 ;44 :197-206 .
56. Timmermans J-P, Barbiers M, Scheuermann DW, Bogers J J, Adriaensen D, Fekete E, Mayer B, Van Marck EA, De Groodt- Lasseel MHA. Nitric oxide synthase immunoreactivity in the en- teric nervous system of the developing human digestive tract. Cell Tissue Res 1994 ;275 :235-245 .
57. Costa M, Furness JB, Pompolo S, Brookes SJH, Bornstein JC, Bredt DS, Snyder SH. Projections and chemical coding of neu- rons with immunereactivity for nitric oxide synthase in the guinea-pig small intestine. Neurosci Lett 1992; 148 :121-125 .
58. Furness JB, Pompole S, Shuttleworth CWR, Burleigh DE. Light- and electron-microscopic immunochemical analysis of nerve fi- bre types innervating the taenia of the guinea-pig caecum. Cell Tissue Res 1992 ;270 :125-137 .
59. Aimi Y, Kimura H, Kinoshita T, Minami Y, Fujimura M, Vincent SR. Histochemical localization of nitric oxide synthase in rat enteric nervous system. Neuroscience 1993 ;53 :553-560 .
60. Costa M, Brookes SA, Waterman S, Mayo R. Enteric neuronal circuitry and transmitters controlling intestinal motor function. In: Holle GE, ed. Advances in the innervation of the gastrointesti- nal tract. Amsterdam: Elsevier, 1992 :115-121 .
61. Daniel EE, Fumess JB, Costa M, Belbeck L. The projections of chemically identified nerve fibres in canine ileum. Cell Tissue Res 1987 ;247 :377-384 .
62. Ekblad E, Hakanson R, Sundler F. VIP and PHI coexist with an NPY-like peptide in intramural neurones of the small intestine. Regul Pept 1984 ;10 :47 -55 .
63. Wattchow DA, Furness JB, Costa M. Distribution and coexis- tence of peptides in nerve fibers of the external muscle of the human gastrointestinal tract. Gastroenterology 1988 ;95 :32 - 41.
64. Burleigh DE, Furness JB. Distribution and actions of galanin and vasoactive intestinal peptide in the human colon. Neuro- peptides 1990; 16 :77-82 .
65. Portbury AL, McConalogue K, Furness JB, Young HM. Distribu- tion of pituitary adenylyl cyclase activating peptide (PACAP) im- munoreactivity in neurons of the guinea-pig digestive tract and

562 FURNESS ET AL. GASTROENTEROLOGY Vol. 108, No. 2
their projections in the ileum and colon. Cell Tissue Res (in press).
66. Sternini C, Anderson K, Frantz G, Krause JE, Brecha N. Expres- sion of substance P/neurokinin A-encoding preprotachykinin messenger ribonucleic acids in the rat enteric nervous system. Gastroenterology 1989; 97 :348 - 356.
67. Mantyh PW, Mantyh CR, Gates T, Vigna SR, Maggie JE. Receptor binding sites for substance P and substance K in the canine gastrointestinal tract and their possible role in inflammatory bowel disease. Neuroscience 1988 ;25 :817-837 .
68. Brookes SJH, Steele PA, Costa M. Identification and immunohis- tochemistry of cholinergic and non-cholinergic circular muscle motor neurons in the guinea-pig small intestine. Neuroscience 1991 ;42 :863 -878 .
69. Steele PA, Brookes SJH, Costa M. Immunohistochemical identi- fication of cholinergic neurons in the myenteric plexus of guinea- pig small intestine. Neuroscience 1991 ;45 :227-239 .
70. Ekblad E, Winther C, Ekman R, Hakanson R, Sundler F. Projec- tions of peptide-containing neurons in rat small intestine. Neu- roscience 1987; 20 :169-188 .
71. Lieweilyn-Smith I J, Furness JB, Gibbins iL, Costa M. Quantitative ultrastructural analysis of enkephalin-, substance P-, and VIP- immunoreactive nerve fibers in the circular muscle of the guinea-pig small intestine. J Comp Neurol 1988 ;272 :139 - 148.
72. France R, Costa M, Furness JB. Evidence for the release of endogenous substance P from intestinal nerves. Naunyn Schmiedebergs Arch Pharmacol 1979 ;306 :195-201 .
73. Barth6 L, Holzer P. Search for a physiological role of substance P in gastrointestinal motility. Neuroscience 1985 ;16 :1 -32 .
74. Niel JP, Bywater RAR, Taylor GS. Effect of substance P on non- cholinergic fast and slow post-stimulus depolarization in the guinea-pig ileum. J Auton Nerv Syst 1983 ;9 :573-584 .
75. Taylor GS, Bywater RAR. Antagonism of non-cholinergic excit- atory junction potentials in the guinea-pig ileum by a substance P analogue antagonist. Neurosci Lett 1986 ;63 :23 -26 .
76. Blanquet F, Bouvier M, Gonella J. Effects of enkephalins and morphine on spontaneous electrical activity and on junction potentials elicited by parasympathetic nerve stimulation in cat and rabbit colon. Br J Pharmacol 1982 ;77 :419-429 .
77. Tonini M, Waterman SA, Candura SM, Coccini T, Costa M. Sites of action of morphine on the ascending excitatory reflex in the guinea-pig small intestine. Neurosci Lett 1992; 144 :195-198 .
78. Brown DR, Miller RJ. Neurohormonal control of fluid and electro- lyte transport in intestinal mucosa. In: Field M, Frizzell RA, eds. Handbook of physiology--the gastrointestinal system IV. Wash- ington: American Physiological Society, 1991 :527-589 .
79. Brunsson I, Fahrenkrug J, Jodal M, Sj6qvist A, Theodorsson E, Lundgren O. On the role of vasoactive intestinal poiypeptide and tachykinins in the secretory reflex elicited by chemical peri- tonitis in the cat small intestine. Acta Physiol Scand 1990; 139 :63-75 .
80. Timmermans J-P, Scheuermann DW, Stach W, Adriaensen D, De Groodt-Lasseel MHA. Functional morphology of the enteric nervous system with special reference to large mammals. Eur J Morphol 1992 ;30 :113-122 .
81. Furness JB, Costa M, Keast JR. Choline acetyltransferase and peptide immunoreactivity of submucous neurons in the small intestine of the guinea-pig. Cell Tissue Res 1984 ;237 :329 - 336.
82. Evans R J, Jiang M-M, Surprenant A. Morphological properties and projections of electrophysiologically characterized neurons in the guinea-pig submucosal plexus. Neuroscience 1994; 59 :1093-1110 .
83. Bornstein JC, Furness JB. Correlated electrophysiological and histochemical studies of submucous neurons and their contri-
bution to understanding enteric neural circuits. J Auton Nerv Syst 1988; 25 :1 -13 .
84. Kirchgessner AL, Tamir H, Gershon MD. Identification and stim- ulation by serotonin of intrinsic sensory neurons of the aubmu- cosal plexus of the guinea pig gut: activity-induced expression of Fos immunoreactivity. J Neurosci 1992 ;12 :235-248 .
85. Steele PA, Costa M. Opioid-like immunoreactive neurons in se- cretomotor pathways of the guinea-pig ileum. Neuroscience 1990 ;38 :771-786 .
86. Song Z-M, Brookes SJH, Steele PA, Costa M. Projections and pathways of submucous neurons to the mucosa of the guinea- pig small intestine. Cell Tissue Res 1992 ;269 :87 -98 .
87. Furness JB, Pompolo S, Murphy R, Giraud A. Projections of neurons with neuromedin U-l ike immunoreactivity in the small intestine of the guinea-pig. Cell Tissue Res 1989 ;257 :415 - 422.
88. Pataky DM, Curtis SB, Buchan AMJ. The co-localization of neuro- peptides in the submucosa of the small intestine of normal Wistar and non-diabetic BB rats. Neuroscience 1990 ;36 :247 - 254.
89. Buchan AMJ. Neurofilament M and calbindin D28 K are present in mutually exclusive subpopulations of enteric neurons in the rat submucous plexus. Brain Res 1991 ;538 :171-175 .
90. Lundgren O, Svanvik J, Jiveg~rd L. Enteric nervous system. I. Physiology and pathophysiology of the intestinal tract. Dig Dis Sci 1989; 34 :264-283 .
91. Davenport HW. Gastrointestinal physiology, 1895 -1975 : motil- ity. In: Wood JD, ed. Handbook of physiology, the gastrointesti- nal system. Washington: American Physiological Society, 1989 :1 -101 .
92. Daugherty DF, Lucey MR, Yamada T. Gastric secretion. In: Ya- mada T, ed. Textbook of gastroenterology. Philadelphia: Lippin- cott, 1991 :233-264 .
93. Maggi CA, Barbanti G, Turini D, Giuliani S. Effect of NG-mono - methyl L-arginine (L-NMMA) and NG-nitro L-arginine (L-NOARG) on non-adrenergic non-cholinergic relaxation in the circular muscle of the human ileum. Br J Pharmacol 1991 ;103 :1970-1972 .
94. Burleigh DE. Ng-nitro-L-arginine reduces non-adrenergic, non- cholinergic relaxations of human gut. Gastroenterology 1992; 102 :679-683 .
95. Tam FS, Hillier K. The role of nitric oxide in mediating non- adrenergic non-cholinergic relaxation in longitudinal muscle of human taenia coll. Life Sci 1992 ;51 :1277-1284 .
96. Stark ME, Bauer A J, Sarr MG, Szurszewski JH. Nitric oxide medi- ates inhibitory nerve input in human and canine jejunum. Gas- troenterology 1993; 104 :398-409 .
97. Demote T, Bishop AE, Oki M, Polak JM. An in vitro study of the projections of enteric vasoactive intestinal polypeptide-immuno- reactive neurons in the human colon. Gastroenterology 1990 ;98 :819-827 .
98. Maggi CA, Patacchini R, Santicioli P, Giuliani S, Turini D, Bar- banti G, Giachetti A, Meli A. Human isolated ileum: motor re- sponses of the circular muscle to electrical field stimulation and exogenous neuropeptides. Naunyn Schmiedebergs Arch Pharmacol 1990; 341 :256-261 .
99. Couture R, Mizrahi J, Regoli D, Devroede G. Peptides and the human colon: an in vitro pharmacological study. Can J Physiol Pharmacol 1981; 59 :957-964 .
100. Burleigh DE. Motor responsiveness of proximal and distal hu- man colonic muscle layers to acetylcholine, noradrenaline, and vasoactive intestinal peptide. Dig Dis Sci 1990 ;35 :617-621 .
101. Springall DR, Riveros-Moreno V, Buttery L, Suburo A, Bishop AE, Merrett M, Moncada S. Immunological detection of nitric oxide synthase(s) in human tissues using heterologous antibod- ies suggesting different isoforms. Histochemistry 1992;98: 259 -266 .
102. De Giorgio R, Parodi JE, Brecha NC, Brunicardi FC, Becker JM,

February 1995 ENTERIC NEUROTRANSMITTERS 563
Go VLW, Sternini C. Nitric oxide producing neurons in the mon- key and human digestive system. J Comp Neurol 1994; 342 :619-627 .
103. Bogers J J, Timmermans J-P, Scheuermann DW, Pelckmans PA, Mayer B, Van Marck EA. Localization of nitric oxide synthase in enteric neurons of the porcine and human ileocaecal junction. Ann Anat 1994; 176 :131-135 .
104. Shen Z, Larsson LT, Malmfors G, Absood A, H~kanson R, Sun- dler F. A novel neuropeptide, pituitary adenylate cyclase-activat- ing polypeptide (PACAP), in human intestine: evidence for re- duced content in Hirschsprung's disease. Cell Tissue Res 1992 ;269 :369 -374 .
105. Ferri G, Adrian TE, Ghatei MA, O'Shaughnessy D J, Probert D J, Lee YC, Buchan AM J, Polak JM, Bloom SR. Tissue localization and relative distribution of regulatory peptides in separated lay- ers from the human bowel. Gastroenterology 1983 ;84 :777- 786.
106. Hubel KA, Renquist KS, Shirazi S. Intramural cholinergic nerves affect mucosal ion transport by the left colon of man (abstr). Gastroenterology 1983; 64:1192.
107. Furness JB, Lloyd KCK, Sternini C, Walsh JH. Projections of substance P, vasoactive intestinal peptide and tyrosine hydroxy- lase immunoreactive nerve fibres in the canine intestine, with special reference to the innervation of the circular muscle. Arch Histol Cytol 1990 ;53 :129-140 .
Received May 20, 1994. Accepted August 30, 1994. Address requests for reprints to: John B. Furness, Ph.D., Depart-
ment of Anatomy and Cell Biology, University of Melbourne, Parkville, Victoria 3052, Australia. Fax: (61) 3-347-5219.
Supported by grants from the National Health and Medical Re- search Council of Australia and U.S. Public Health Service grant NS 23816.
Related Documents











