Plasmid-Mediated Tolerance Toward Environmental Pollutants ANA SEGURA, 1 LÁZARO MOLINA, 2 and JUAN LUIS RAMOS 1 1 Estación Experimental del Zaidin (CSIC), Environmental Protection Department, Profesor Albareda, 1, 18008 Granada, Spain; 2 CIDERTA, Laboratorio de Investigación y Control Agroalimentario (LICAH), Parque Huelva Empresarial, 21007 Huelva, Spain ABSTRACT The survival capacity of microorganisms in a contaminated environment is limited by the concentration and/or toxicity of the pollutant. Through evolutionary processes, some bacteria have developed or acquired mechanisms to cope with the deleterious effects of toxic compounds, a phenomenon known as tolerance. Common mechanisms of tolerance include the extrusion of contaminants to the outer media and, when concentrations of pollutants are low, the degradation of the toxic compound. For both of these approaches, plasmids that encode genes for the degradation of contaminants such as toluene, naphthalene, phenol, nitrobenzene, and triazine or are involved in tolerance toward organic solvents and heavy metals, play an important role in the evolution and dissemination of these catabolic pathways and efflux pumps. Environmental plasmids are often conjugative and can transfer their genes between different strains; furthermore, many catabolic or efflux pump genes are often associated with transposable elements, making them one of the major players in bacterial evolution. In this review, we will briefly describe catabolic and tolerance plasmids and advances in the knowledge and biotechnological applications of these plasmids. INTRODUCTION Since the Industrial Revolution, there has been an in- creasing pace in the production of environmentally haz- ardous compounds that deliberately or accidentally have reached waters and soils, polluting them. The survival capacity of microorganisms in a contaminated environ- ment is limited by the concentration and/or toxicity of the pollutant. Some contaminants are able to disrupt the normal development of the cell, others induce mutations, and some of these can kill cells at very low concen- trations. Through evolutionary processes, some bacteria have developed or acquired mechanisms to cope with the deleterious effects of toxic compounds, permitting normal cellular subsistence in polluted environments— a phenomenon known as tolerance. Common mecha- nisms of tolerance include the extrusion of contaminants to the outer media and, when concentrations of pol- lutants are low, the degradation of the toxic compound. For both of these approaches, plasmids play an impor- tant role in the evolution and dissemination of the cata- bolic pathways and efflux pumps. In this article, we will briefly describe catabolic and tolerance plasmids and advances in the knowledge and biotechnological applications of tolerance plasmids. ENVIRONMENTAL CATABOLIC PLASMIDS The mineralization of pollutants decreases their con- centration in the environment and therefore allows better survival of the organism; at the same time, pollutants are used to obtain energy for growth. This mechanism of resistance is only useful when the con- taminant concentration is moderate or low enough to allow normal bacterial metabolism; for example, while Pseudomonas putida mt-2 is able to mineralize toluene through the TOL pathway, the strain fails to thrive in high concentrations of toluene ( 1). Received: 4 February 2014, Accepted: 5 February 2014, Published: •••••• Editors: Marcelo E. Tolmasky, California State University, Fullerton, CA, and Juan Carlos Alonso, Centro Nacional de Biotecnología, Cantoblanco, Madrid, Spain Citation: Segura A, Molina L, Ramos JL. 2014. Plasmid-Mediated tolerance toward environmental pollutants. Microbiol Spectrum 2(4):PLAS-0013-2013. doi:10.1128/microbiolspec.PLAS-0013-2013. Correspondence: Ana Segura, [email protected] © 2014 American Society for Microbiology. All rights reserved. ASMscience.org/MicrobiolSpectrum 1

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Plasmid-Mediated ToleranceToward Environmental Pollutants
ANA SEGURA,1 LÁZARO MOLINA,2 and JUAN LUIS RAMOS11Estación Experimental del Zaidin (CSIC), Environmental Protection Department, Profesor Albareda,
1, 18008 Granada, Spain; 2CIDERTA, Laboratorio de Investigación y Control Agroalimentario(LICAH), Parque Huelva Empresarial, 21007 Huelva, Spain
ABSTRACT The survival capacity of microorganisms in acontaminated environment is limited by the concentrationand/or toxicity of the pollutant. Through evolutionary processes,some bacteria have developed or acquired mechanisms to copewith the deleterious effects of toxic compounds, a phenomenonknown as tolerance. Common mechanisms of tolerance includethe extrusion of contaminants to the outer media and, whenconcentrations of pollutants are low, the degradation of thetoxic compound. For both of these approaches, plasmids thatencode genes for the degradation of contaminants such astoluene, naphthalene, phenol, nitrobenzene, and triazine or areinvolved in tolerance toward organic solvents and heavy metals,play an important role in the evolution and dissemination ofthese catabolic pathways and efflux pumps. Environmentalplasmids are often conjugative and can transfer their genesbetween different strains; furthermore, many catabolic or effluxpump genes are often associated with transposable elements,making them one of the major players in bacterial evolution.In this review, we will briefly describe catabolic and toleranceplasmids and advances in the knowledge and biotechnologicalapplications of these plasmids.
INTRODUCTIONSince the Industrial Revolution, there has been an in-creasing pace in the production of environmentally haz-ardous compounds that deliberately or accidentally havereached waters and soils, polluting them. The survivalcapacity of microorganisms in a contaminated environ-ment is limited by the concentration and/or toxicity of thepollutant. Some contaminants are able to disrupt thenormal development of the cell, others induce mutations,and some of these can kill cells at very low concen-trations. Through evolutionary processes, some bacteriahave developed or acquired mechanisms to cope withthe deleterious effects of toxic compounds, permitting
normal cellular subsistence in polluted environments—a phenomenon known as tolerance. Common mecha-nisms of tolerance include the extrusion of contaminantsto the outer media and, when concentrations of pol-lutants are low, the degradation of the toxic compound.For both of these approaches, plasmids play an impor-tant role in the evolution and dissemination of the cata-bolic pathways and efflux pumps.
In this article, we will briefly describe catabolic andtolerance plasmids and advances in the knowledge andbiotechnological applications of tolerance plasmids.
ENVIRONMENTAL CATABOLIC PLASMIDSThe mineralization of pollutants decreases their con-centration in the environment and therefore allowsbetter survival of the organism; at the same time,pollutants are used to obtain energy for growth. Thismechanism of resistance is only useful when the con-taminant concentration is moderate or low enough toallow normal bacterial metabolism; for example, whilePseudomonas putida mt-2 is able to mineralize toluenethrough the TOL pathway, the strain fails to thrive inhigh concentrations of toluene (1).
Received: 4 February 2014, Accepted: 5 February 2014,Published: ••••••Editors: Marcelo E. Tolmasky, California State University, Fullerton,CA, and Juan Carlos Alonso, Centro Nacional de Biotecnología,Cantoblanco, Madrid, Spain
Citation: Segura A, Molina L, Ramos JL. 2014. Plasmid-Mediatedtolerance toward environmental pollutants. Microbiol Spectrum2(4):PLAS-0013-2013. doi:10.1128/microbiolspec.PLAS-0013-2013.
Correspondence: Ana Segura, [email protected]
© 2014 American Society for Microbiology. All rights reserved.
ASMscience.org/MicrobiolSpectrum 1

Environmental plasmids are often conjugative andcan transfer their genes between different strains; fur-thermore, many catabolic genes are often associatedwith transposable elements, making them one of themajor players in bacterial evolution. Genes for the deg-radation of contaminants such as toluene, xylenes, al-kanes, naphthalene, phenol, nitrobenzene, triazine, andothers have been reported to be encoded in plasmids(2, 3, 4). Catabolic plasmids are generally large in size(60 to 200 kb), making them difficult to isolate andcomplicating the ascription of genes to these extra-chromosomal elements. Recent advances in sequencingtechnologies have resulted in a growing number ofcompletely sequenced catabolic plasmids. Metagenomictechniques have significantly advanced our knowledgeof genes that are present in noncultivatable bacteria;however, it has been suggested that the presence ofplasmid-encoded genes in metagenomic libraries is oftenunderrepresented (5), leaving a knowledge gap regard-ing the role and importance of plasmids harbored bynoncultivable bacteria.
It should be noted that the presence of catabolic geneshas not only been identified in plasmids, but also in thechromosome. Here, we describe plasmid-based systemsthat have been studied in detail.
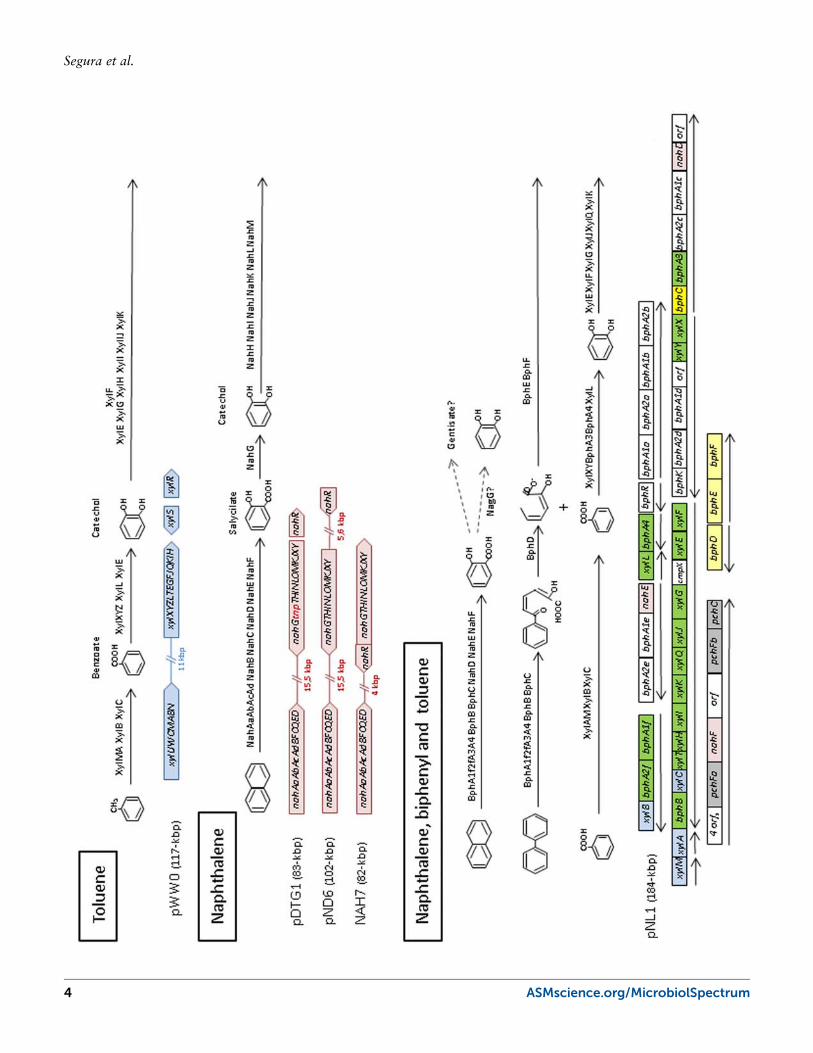
Plasmids That Encode Pathways for theDegradation of Monoaromatic CompoundsToluene is a natural product and, although aerobic bac-teria have evolved several different pathways to degradeit, they all share a common strategy. In many cases,an upper pathway is able to activate the aromatic ringthroughout mono- or dioxygenases, or by oxidation ofthe alkylic substituent; subsequently, the lower pathwaybreaks down the activated aromatic ring to produceintermediates that can enter the tricarboxylic acid cycle(6, 7). The most studied plasmid for the degradation ofmonoaromatic compounds is the TOL plasmid (pWW0),which encodes the genes necessary for the degradationof toluene, m- and p-xylene. This plasmid was origi-nally identified in P. putida mt-2 and has since been thesubject of intensive research (8). pWW0, a 117-kbp self-transmissible plasmid belonging to the IncP-9 incom-patibility group, was completely sequenced in 2002(9). Together with toluene catabolic genes, the plasmidencodes all the functions necessary for its replication,stable inheritance, and conjugation. The catabolic geneswithin the pWW0 plasmid are organized in two operons,the “upper pathway” (xylUWCMABN), which convertstoluene and xylenes into benzoate and methyl benzo-
ates, and the “meta pathway” (xylXYZLTEGFJQKIH),which transforms these intermediates into Krebs cycleintermediates (Fig. 1). Two regulatory proteins, XylRand XylS, control the expression of these two operonsthrough a fine-tuned regulatory circuit that has also beenstudied in great detail (10, 11). In fact, this regulatorysystem represents a paradigm for signal integration ingene regulatory networks (12). These two operons, to-gether with the 14 open reading frames between them,are located between two identical repeats of 1275 bp.This complete region has been named IS1246 becauseof its insertion sequence characteristics. Spontaneousdeletion of a 39-kbp region, comprising the catabolicgenes, has been observed, probably as a consequence ofrecombination between the two repeated sequences (13,14). Two transposable elements, with characteristics ofclass II transposons, have been identified in the pWW0plasmid (Tn4651 and Tn4653) (15, 16); and anotherinsertion sequence (ISPpu12) is also present and activein this plasmid (17). The presence of multiple insertionsequences, transposases, and recombinases is a commonelement inmost catabolic plasmids, and it is thought to berelated to the acquisition of new genes and microbialevolution.
Plasmids that carry xyl genes homologous to thoseof the pWW0 plasmids are normally referred to as TOLplasmids and have been isolated from different locations(18, 19, 20, 21). They differ in the organization of thecatabolic pathways, in size, and even in replicon type;e.g., plasmids pWW53 and pDK1 belong to the IncP-7plasmid family (22, 23). Comparison between multipleTOL plasmids indicate that transposition-related genesand sites have contributed to the diversification of plas-mid structures and to the dissemination of common geneclusters to various plasmids (22).
Plasmid pTOM (latter designated pBV1E04) of Burk-holderia vietnamensis G4 (24; CP_000620) encodesthe toluene 2-monooxygenase (T2MO) that catalyzesthe two initial oxidation steps in toluene degradation,transforming toluene into 3-methylcatechol via o-cresol(25). Ring cleavage proceeds via a meta cleavage path-way that is also encoded in the pBV1E04 plasmid. Al-though it is known that T2MO activity is inducible, theregulation of this pathway has not been studied; how-ever, a gene with homology to the XylR/NtrC family oftranscriptional regulators that is located upstream of theT2MO gene cluster has been hypothesized to be theregulatory protein of the operon (6). B. vietnamensisG4 was initially isolated because it oxidized trichloro-ethylene (TCE), but it was latter demonstrated that itcan grow in the presence of toluene, phenol, o- ,m-, and
2 ASMscience.org/MicrobiolSpectrum
Segura et al.

p-cresol, and benzoate (26). This strain and the pTOMplasmid have been used extensively for different bio-technological approaches, including TCE and toluenerhizoremediation (see below). Interestingly, the back-bone of the pBV1E04 plasmid is quite similar to thatof pGRT1 from the solvent tolerant P. putidaDOT-T1Estrain (27). Several transposases, insertion sequences,and integrases are encoded in the pBVIE04 plasmid,many of them surrounding the area where the catabolicgenes are located, suggesting transfer of catabolic genesto/from other strains.
Plasmids That Encode Pathways forthe Degradation of PolycyclicAromatic Hydrocarbons andHeteroaromatic CompoundsSince the first report of a plasmid that encoded the ca-tabolism of camphor, the CAM plasmid (28, 29), vari-ous plasmids that confer the ability to grow in polycyclicaromatic compounds have been isolated. Naphthalenehas served as a model compound for the study of poly-cyclic aromatic hydrocarbons (PAH) degradation andthe associated plasmids have been named as NAH plas-mids. Naphthalene degradation enzymes are encodedwithin two operons; the upper pathway encodes func-tions for the conversion of naphthalene to salicylate(Fig. 1), and the lower pathway encodes genes for themeta-ring fission pathway and a predicted methyl-accepting chemotaxis protein, NahY (30, 31, 32, 33, 34,35). The nahR gene encodes the regulatory protein ofthe system, which belongs to the LysR family (36) andinduces the two operons in the presence of salicylate.
Plasmids pDTG1 from P. putida strain NCIB 9816-4,pND6-1 from Pseudomonas sp. strain ND6, and NAH7from P. putida G7 have been completely sequenced (37,38, 39). These three plasmids encode the nahR gene, aswell as two catabolic operons: the nah operon, whichencodes de upper pathway, and the lower operon forsalicylate degradation (Fig. 1). Proteins for naphthalenedegradation are almost identical in the three strains(99% to 100% identity in amino acid sequences), withthe exception of two duplicated genes in pND6. Fur-thermore, nucleotide identity in the catabolic regionof plasmid pND6 and pDTG1 is >99%, including a15-kbp region between the two operons that contain anumber of functionally unrelated genes (38). Despitethis homology, P. putida strain NCIB 9816-4 degradesnaphthalene through a chromosomally encoded ortho-pathway due to the presence of an ISPre1 between thenahG (encoding the salicylate hydroxylase) and nahT
genes of the pDGT1 plasmid (37). In pND6 and pDTG1,the two operons are transcribed in opposite directions,while, in NAH7, both operons are transcribed in thesame direction. nahR in NAH7 is not located upstreamof the lower operon, but in between the two operons(38). In this plasmid, the naphthalene degradation genesare located within a class II transposon (Tn4655), whichlacks a tnpA gene, but that can be mobilized by theaction of the TnpA from pWW0 Tn4653 suppliedin trans (40). Within these three plasmids, numeroustransposases, resolvases, and integrases are located inthe vicinity of the catabolic genes, and, in many cases,they are similar to elements found in catabolic plas-mids pWW0 and pCAR1. The “backbone” of thepDTG1 and NAH7 plasmids is homologous to that ofthe pWW0 plasmid, suggesting that the exchange ofcatabolic genes by means of horizontal gene transfermay have occurred between these plasmids.
Plasmids involved in the degradation of polycyclicaromatic hydrocarbons with three aromatic rings havealso been described (e.g., the NAH-plasmid-encodedpathway is able to mineralize phenanthrene and an-thracene [41]). Not only pseudomonads, but many otherbacteria are able to degrade naphthalene and phenan-threne through similar biochemical steps. The enzymesof the naphthalene pathway are also able to degradephenanthrene and anthracene via the 1-hydroxy-2-naphthoic acid intermediate, which is subsequently ox-idized through salicylate and catechol (in other cases,this intermediate can be channeled to o-phthalate andprotocatechuate [42]). The Burkholderia sp. strainRP007 contains a plasmid that encodes the upper path-way for the degradation of naphthalene and phenan-threne (43). The Sphingomonas aromaticivorans F199plasmid known as pNL1 encodes genes associated withthe degradation of biphenyl, naphthalene, m-xylene,and p-cresol. Interestingly, although the related genesare organized in operons under a commonly occurringpseudomonad regulatory scheme, in sphingomonadsthe genes are scattered across the plasmids (44, 45). Inthe pNL1 plasmid, genes associated with aromatic deg-radation are distributed among at least 11 transcrip-tional units (Fig. 1). The unusual coclustering of genesassociated with different catabolic pathways (biphenyl,toluene, xylenes, and naphthalene) observed in thisplasmid is likely due to evolutionary modifications ap-plied to similar biochemical mechanisms for the degra-dation of intermediates in the different pathways (46).As in many other catabolic plasmids, the presence ofa recombinase, an excisionase, a phage-type integrase-recombinase, and two transposons have been identified.
ASMscience.org/MicrobiolSpectrum 3
Plasmid-Mediated Tolerance Toward Environmental Pollutants

The genus Sphingomonas is able to degrade a widevariety of xenobiotics (biphenyl, PAHs, and substitutedPAHs, carbazole, diphenyl ethers, furans, dibenzo-p-dioxins, and others) and many of the catabolic genes forthese compounds are located on plasmids that can bemobilized to other Sphingomonas strains (45, 47, 48).Plasmid pBN6 of S. xenophaga BN6 encodes the path-way involved the degradation of naphthalene sulfonate,and the related genes show high similarity to those en-coded in the pNL1 plasmid of S. aromaticivorans F199and also share the structural organization of the naph-thalene, biphenyl, and phenanthrene genes within threetranscriptional units. Not surprisingly, these genes areflanked with genes that encode putative mobile geneticelements (49).
Another group of plasmids that have been extensivelystudied are the pCAR plasmids that encode genes forthe degradation of carbazole/dioxin. pCAR plasmidsthat have been fully sequenced include pCAR1 fromPseudomonas resinovorans CA10 (50), pCAR2 fromP. putida HS01 (51), and pCAR3 from Sphingomonassp. KA1 (52). Enzymes encoded by the car operontransform carbazole to anthranilate, and the proteinsencoded by the ant operon are responsible for the con-version of anthranilate to catechol (Fig. 2). Carbazoledegradation starts with the angular deoxygenation of thecompound by the CARDO complex, a three-componentdioxygenase that has been extensively characterized (53,54, 55). In pCAR1, the car and ant operons are foundwithin a 72.8-kbp transposon named Tn4676 whosetransposon-related genes show homology with Tn4651of the TOL plasmid pWW0. After the introduction ofplasmid pCAR1 plasmid into P. fluorescens Pf0-1 manyrearrangements were observed; one of these is the inser-tion of Tn4676 into the chromosome (56). Numerousmobile genetic elements are found within the Tn4676transposon and its flanking region (50). Insertion se-quences identical to IS1162 (57), with homology toISEc8 (58), IS1491 (59), and ISPre (60) are also encodedin pCAR1. The pCAR3 plasmid is bigger than pCAR1(254,797 versus 199,035 bp); encoded in pCAR3 arethe car and and operons, as well as a second cluster ofcar genes (car-II) and putative genes for the degradation
of catechol, protocatechuate, and phthalate. Five differ-ent types of insertion sequences and transposons wereidentified in pCAR3. The backbone of this plasmid issimilar to that of pNL1 of Novosphingobium aroma-ticivorans F1999, and, although it encoded all the func-tions necessary for conjugation, attempts to transfer it toa pCAR3-cured strain have been unsuccessful (52).
Plasmid pARUE113 (pAL1) from the ActinobacteriaArthrobacter sp. Rue61a encodes genes for the degra-dation of quinaldine (61). Quinaldine is transformedto anthranilate through the enzymes encoded by themeqABC (qoxLMS) operon that encodes quinaldine4-oxidase and the divergently transcribed meqDEF(formerly named moq, hod, and amp) (62, 63) (Fig. 2).The plasmid also contains a lower pathway for thedegradation of anthranilate through CoA intermediatesto produce 2-amino-5-oxo-cyclohex-1-ene-carbonyl-CoA. This product is then thought to be degraded via aβ-oxidation-like pathway encoded in the chromosome(63, 64). The enzymes of the pathway are induced inthe presence of the substrate, and two putative tran-scriptional regulators, belonging to the GntR super-family, are located near the catabolic cluster involvedin quinaldine degradation. The function of MeqR1 iscurrently unknown, while MeqR2 is able to bind thepromoter region of mepC, meqD, orf1, and to its ownpromoter and shows high binding specificity foranthraniloyl-CoA. These results suggest that MeqR2may be involved in the regulation of the pathway, al-though other regulatory systems exist that are known togovern the expression of the meq genes (65). Althoughconserved gene clustering has been observed in pAL1,suggesting a modular structure, only one insertion se-quence has been detected (63). Interestingly, pAL1 is aconjugative linear plasmid (61). It has been suggestedthat the replication of linear plasmids proceeds bidirec-tionally from an internal origin toward the telomeresgenerating replicative intermediates that contain 3′-strand overhangs (66). The left and right ends of pAL1contain palindromic sequences that could be importantfor telomere patching; furthermore, pAL1 encodes pu-tative proteins that could also be associated with thetelomere patching.
FIGURE 1 Degradation pathways of mono- and biaromatic compounds. Major inter-mediates of the pathways are depicted. Genes or operons in different plasmids are coloredto indicate their role: blue for toluene degradation genes, pink for naphthalene degra-dation genes, and yellow for biphenyl degradation genes. In green are the genes thatcan function in different degradation pathways. Genes and operons are not drawn atscale. Operon organization in some cases has not been experimentally demonstrated.doi: 10.1128/microbiolspec.PLAS-0013-2013.f1
ASMscience.org/MicrobiolSpectrum 5
Plasmid-Mediated Tolerance Toward Environmental Pollutants
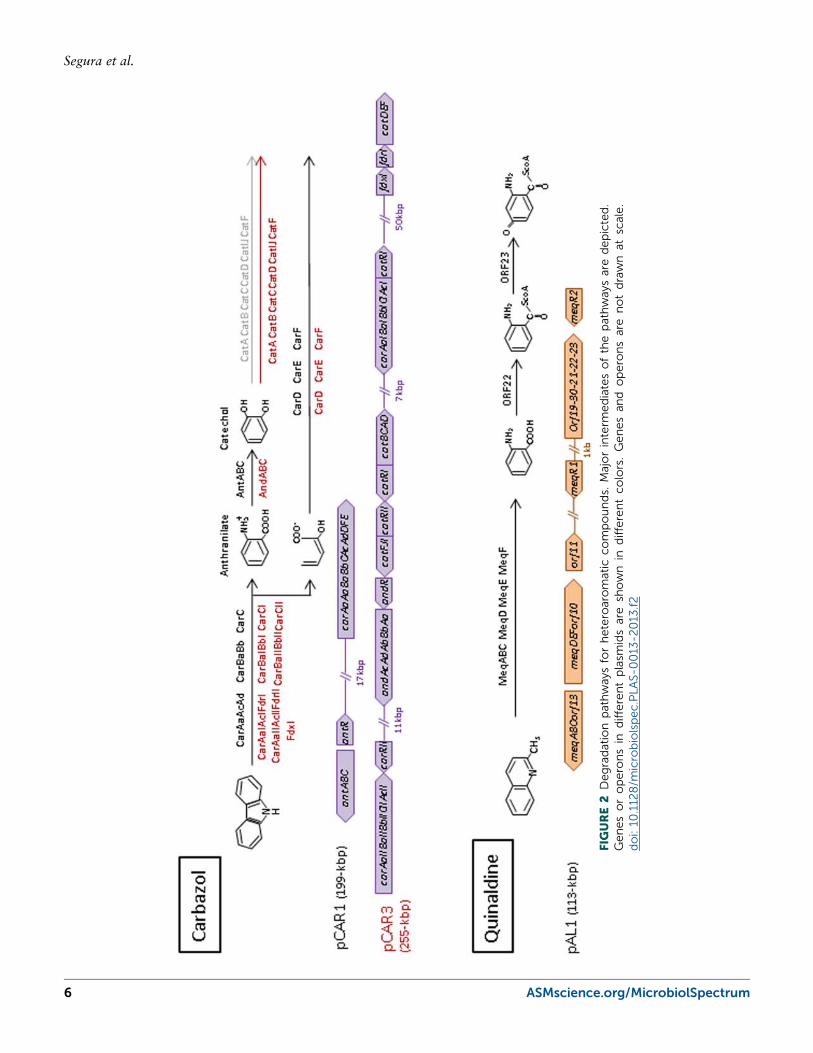
FIGURE2
Degradationpathwaysforheteroaromatic
compounds.
Majorinterm
ediatesofthepathwaysaredepicted.
Genesoroperonsin
differentplasm
idsare
shown
indifferentco
lors.Genesan
doperonsare
notdrawn
atscale.
doi:10
.112
8/m
icrobiolspec.PLA
S-0013
-2013
.f2
6 ASMscience.org/MicrobiolSpectrum
Segura et al.

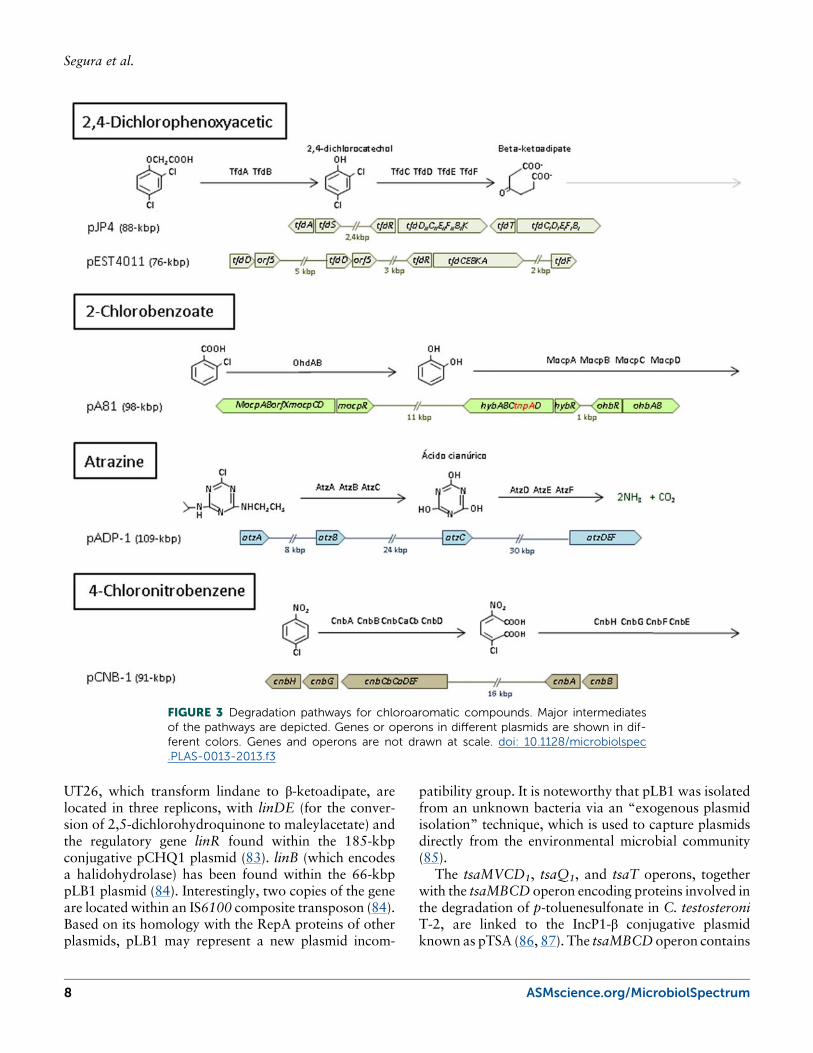
Plasmids That Encode Pathways for theDegradation of Chlorinated CompoundsCatabolic pathways for the degradation of 2,4-dichlo-rophenoxyacetic acid (2,4-D), a xenobiotic herbicide,have been studied for almost 30 years. One of thebest characterized pathways for the degradation of thisherbicide is the pathway encoded by the pJP4 plasmidfrom Wautersia eutropha JMP134 (formerly Ralstoniaeutropha), a strain that is able to mineralize 2,4-D and 3-chlorobenzoic acid (67) (Fig. 2). Two chlorocatechol-degrading gene clusters (tfdI and tfdII) are required forefficient degradation of the compounds (68, 69). TheIncP1-β conjugative plasmid pJP4 was sequenced in2004 (70) and was reported to contain inactive trans-posons and remnants of lateral gene transfer events. Aputative ISJP4-based transposon encompassing the tfdIIgene cluster and a putative IS1071-based transposonthat flanks all the catabolic genes are found in the pJP4sequence. However, it has been suggested that theseputative transposons are not active (70). The pEST4011plasmid from Achromobacter xylosoxidans subsp.denitrificans also carries genes for the degradation of2,4-D (71). Based on differences among backboneproteins of the IncP1 group, it has been hypothesizedthat this plasmid belongs to a new category, known asthe δ subgroup. Within this group, degradative genes areinserted into the backbone at a 48-kbp catabolic trans-poson that is very similar to Tn5530, which was iden-tified in plasmid pIJB1 from Burkholderia cepacia 2a(72). The tfd operon is quite similar to tfdII of pJP4,suggesting a common origin, although tfdD and tfdFhave been deleted in the pEST4011 operon (Fig. 2).These genes could have been recruited during evolutionto replace the genes lost from the original operon.
Chlorobenzoates are intermediates within the bacterialdegradation of polychlorinated biphenyls. Plasmid pA81,from A. xylosoxidans A8 is an IncP1-β plasmid thatharbors genes for the degradation of ortho-substitutedchlorobenzoates (ohbRAB, mocpRABCD) in a class Itransposon named TnAxI (73). This transposon alsocarries an operon predicted to function in salicylate deg-radation, known as hybRABCD (74) (Fig. 3). The pA81plasmid carries another transposon, known as TnAxII,that is involved in heavy metal resistance.
Atrazine is another widespread herbicide that manybacteria are able to degrade (75). Genes for the degra-dation of atrazine in Pseudomonas sp. ADP are locatedin the 109-kbp pADP-1 plasmid, which was sequencedin 2001 (76, 77). atzA, atzB, and atzC encode the en-zymes for the transformation of atrazine to cyanuricacid. Each of these genes is flanked by elements with
homology to IS1071, suggesting that they may havebeen acquired at different times by independent trans-position events. The atzDEF operon encodes enzymesthat convert cyanuric acid to ammonia and carbon di-oxide (Fig. 3). The plasmid belongs to the IncPβ plasmidand has full capabilities for replication, stable mainte-nance, and conjugation (76, 77).
Chloroaniline degradation is linked to the presence ofplasmids pWDL7::rfp (a derivative of plasmid pWDL7of Comamonas testosteroni strain WDL7) and pNB8c(fromDelftia acidovorans strain B8c). Contained withinthese plasmids are genes for the upper pathway(dcaQTA1A2BR) of chloroaniline degradation, whichmediates its transformation to chlorocatechol. Thisproduct is then converted to tricarboxylic acid (TCA)intermediates by modified ortho- or meta-pathways en-coded by the chromosomes. Both plasmids belong tothe IncP-1β incompatibility group and their sequencesare very similar, although pWDL7::rfp contains twoTn6063 transposons that carry dcaQTA1A2BR genes(78). These plasmids can be transferred between Beta-proteobacteria and Gammaproteobacteria indicatinga broad host range. Transfer of plasmid pNB8c intoCupriavidus pinatubonensis JMP228 confers the strainthe ability to degrade anilines but not chloroanilines.This deficiency is due to the lack of induction of the3-CA pathway in this strain. Sequencing of the twoplasmids allowed the identification of a difference inthe dca promoter region responsible for this lack of in-duction (78).
Chlorinated nitroaromatic compounds are amongthe most difficult compounds to degrade because theelectron-withdrawing properties of the nitro and chlorogroups. However, some genes associated with chloro-nitrobenzene degradation are encoded in the pCNB-1plasmid of Comamonas sp. CNB-1 (79, 80, 81). Thedegradation pathway converts chloronitrobenzene into2-amino-5-chloromuconate, which is then degraded toTCA intermediates (Fig. 3). The cnb genes, together withan operon that encodes for resistance to chromate andarsenate are contained within a large transposon be-longing to the class I transposon named TnCNB1 (82).pCNB-1 contains 45 open reading frames (ORFs) thatprovide capabilities for replication, transfer and stabili-ty, and similarly to other catabolic plasmids, belongs tothe IncP1-β incompatibility group.
Other Catabolic Genes Located in PlasmidsA number of genes associated with degradation oflindane (γ-hexachlorocyclohexane) have been found onplasmids. The lin genes of Sphingobium japonicum
ASMscience.org/MicrobiolSpectrum 7
Plasmid-Mediated Tolerance Toward Environmental Pollutants
UT26, which transform lindane to β-ketoadipate, arelocated in three replicons, with linDE (for the conver-sion of 2,5-dichlorohydroquinone to maleylacetate) andthe regulatory gene linR found within the 185-kbpconjugative pCHQ1 plasmid (83). linB (which encodesa halidohydrolase) has been found within the 66-kbppLB1 plasmid (84). Interestingly, two copies of the geneare located within an IS6100 composite transposon (84).Based on its homology with the RepA proteins of otherplasmids, pLB1 may represent a new plasmid incom-
patibility group. It is noteworthy that pLB1 was isolatedfrom an unknown bacteria via an “exogenous plasmidisolation” technique, which is used to capture plasmidsdirectly from the environmental microbial community(85).
The tsaMVCD1, tsaQ1, and tsaT operons, togetherwith the tsaMBCD operon encoding proteins involved inthe degradation of p-toluenesulfonate in C. testosteroniT-2, are linked to the IncP1-β conjugative plasmidknown as pTSA (86, 87). The tsaMBCD operon contains
FIGURE 3 Degradation pathways for chloroaromatic compounds. Major intermediatesof the pathways are depicted. Genes or operons in different plasmids are shown in dif-ferent colors. Genes and operons are not drawn at scale. doi: 10.1128/microbiolspec.PLAS-0013-2013.f3
8 ASMscience.org/MicrobiolSpectrum
Segura et al.

no promoter-like sequence and is not expressed. The tsaregion forms part of a composite transposon that isflanked by two IS1071 elements. This transposon hasbeen found in several bacterial strains that were isolatedfrom samples enriched in toluenesulfonate on three dif-ferent continents and was found to be inserted withinplasmids or chromosomes (87).
Degradation of phenol has been linked with thepresence of plasmids in Pseudomonas sp. EST1001 andPseudomonas sp. CF600. The Phe+ plasmid of strainEST1001 allows it to grow on phenol (88, 89), and,although it has not been completely sequenced, it isknown that the pheAB operon is flanked by two ISelements that are involved in the activation of phenolgenes (90) and in horizontal gene transfer (91). Pseu-domonas sp. CF600 harbors the pVI150 megaplasmidthat belongs to the IncP-2 incompatibility group andencodes all the genetic information for the degradationof phenol, cresols, and 3,4-dimethylphenol (92, 93).
The alkane degradation pathway encoded in the OCTplasmid ofP. putidaGPo1 is organized into two operons,alkBFGHJKL and alkST. The first operon encodes all,except for one, of the enzymes required for the terminaloxidation of alkanes to their corresponding fatty acids.The alkST cluster codes for the transcriptional regulatorAlkS and for a rubredoxin reductase (AlkT ) that is partof the alkane hydroxylase complex (94, 95). The OCTplasmid was first described in the 1970s, and its operonexpression has served as a model for the study of globalregulation responses (96). The two alk operons areflanked by ISPpu4, forming a class I transposon.
TOLERANCE PLASMIDS INBIODEGRADATIONAlthough catabolic genes can contribute to the survivalof the bacteria in the presence of contaminants, whenthese contaminants are present at high concentrations,additional mechanisms—in addition to the degradationof the compounds—are required to avoid toxicity.This is the case with organic solvents, which are highlyhydrophobic and accumulate in and disrupt cell mem-branes causing cellular death (97). If the toxic com-pounds are not degradable (i.e., heavy metals), survivalis provided by the action of efflux pumps that eliminatethe compounds from the cytoplasm or the membranes ofthe microorganisms. These mechanisms are normallyknown as tolerance or resistance mechanisms and manyof them are also encoded in plasmids, although, as issometimes the case with catabolic genes, they can also belocated on the chromosome.
Plasmids Involved in Solvent/AromaticCompound ToleranceP. putida DOT-T1E, a strain isolated from a seawatertreatment plant in Granada, provides a well-studied ex-ample of survival in extreme conditions. This strain isable to degrade different aromatic compounds includingtoluene (98), and to survive at solvent concentrations upto 90% (vol/vol) (99). The genes that encode the toluenedegradation pathway are located on the chromosome(100, 101), while its ability to survive in the presenceof high concentrations of toluene is enabled by a 133-kbp plasmid named pGRT1 (102). Solvent tolerance isa multifactorial process that involves a wide range ofphysiological changes to overcome solvent damage (103);however, mutational experiments have demonstratedthat the TtgGHI efflux pump is the most important de-terminant of solvent tolerance (102, 104). The ttgGHIoperon of the pGRT1 plasmid encodes an efflux pump oftheRNDfamily (resistance, nodulation, cell division) thatextrudes a wide variety of compounds, including anti-biotics, although not all of them with the same efficiency(104). This family of efflux pumps has been extensivelystudied because they confer multidrug resistance to somerelevant clinical strains (105, 106). The efflux system isformed by three components: an effluxpump transporter,located in the cytoplasmic membrane that recognizessubstrates in the periplasm or in the cytoplasmic mem-brane (107, 108); an outer membrane protein that formsa trimeric channel capable of penetrating into the peri-plasm and contacting directly with the efflux pumptransporter (109); and a lipoprotein anchored to the innermembrane that expands into the periplasmic space andmay serve as a bracket for the other two components(110, 111). In the pGRT1 system, the ttgH gene encodesfor the efflux pump, ttgI for the outer membrane protein,and ttgG for the periplasmic adaptor protein. Two otherRND efflux pumps, chromosomally encoded, have beenshown to participate in solvent tolerance in P. putidastrain DOT-T1E, TtgABC (112), and TtgDEF (113).Despite the contribution of TtgABC and TtgDEF to thesolvent-tolerant phenotype, the loss of the TtgGHI effluxpump renders the cells unable to survive in the presenceof high concentrations of toluene (102, 104).
The pGRT1 plasmid sequence confirms that it is nota catabolic plasmid (27); however, it shares many fea-tures typical of catabolic plasmids. First, the toluenetolerance genes are located within a Tn4653-like trans-poson (Fig. 4A), which is similar to those found in TOLand pCAR plasmids. This transposon also contains ttgV,which encodes the regulatory protein that controls theexpression of the efflux pumps (114), two genes that
ASMscience.org/MicrobiolSpectrum 9
Plasmid-Mediated Tolerance Toward Environmental Pollutants

encode methyl-accepting chemotaxis proteins (mcpT1and mcpT2), and a gene that is homologous to uvrD.The two MCP proteins are almost identical and allowthis bacterium to move toward a broad variety of aro-matic compounds and crude oil (115). Monocopies ofMcpT homologues (99% sequence identity) have beenfound on other catabolic plasmids of hydrocarbon-degrading strains like pCAR1 of P. resinovorans (116),the TOL plasmid pWW53 of P. putida (22) and thepMAQU02 plasmid of Marinobacter aquaeolei VT8(NC_008739). All of these strains showed a chemo-attractive response to toluene. Other examples of MCPslocated on plasmids are the NahY protein located onthe NAH7 catabolic plasmid of P. putida that respondsto naphthalene (35) and NbaY from P. fluorescensKU-7that responds to 2-nitrobenzoate (117). UvrD is a DNAhelicase that regulates the activity of the ruvA and ruvBgenes, which are present on the pGRT1 plasmid (Fig. 4),and which is involved in the repair of DNA cross-linksproduced by exposure to UV light and by exposure totoluene (118). UV-resistance genes are located within abroad set of catabolic plasmids. Examples of catabolicplasmids bearing close homologues to the uvrD, ruvA,and ruvB genes from pGRT1 are the naphthalene-degradative pND6 plasmid and the toluene-degradativeplasmids pWW0 and pWW53 from P. putida (9, 22,38).
As well as these traits, other characteristics of pGRT1that are shared with catabolic plasmids include the factthat it encodes proteins related to DNA replication andmodification, plasmid maintenance, mobilization, andtransfer (27) with high similarity to that of the pBVIE04plasmid of B. vietnamensis G4. Inserted within thepGRT1 backbone, in addition to the Tn4653-like
transposon, is a second DNA segment that contains in-vertases, transposases, and recombinase (Fig. 4B). Thissegment contains ruvA, ruvB, and other genes related tostress responses. As well, the protein encoded by orf32is a new toluene tolerance modulator in DOT-T1E. Thisprotein contains a domain identified in several SdiA-regulated proteins, and it is known that chromosomallyencoded SdiA proteins in Escherichia coli and Salmo-nella enterica were able to modulate AcrAB activity(119, 120). Knockout mutants of ORF32 showed a re-duction of the expression of the ttgV repressor gene thatin turn enhances the expression of the ttgGHI effluxpump and concomitantly increases solvent tolerance(27). ORF32 is highly homologous to a protein of un-known function (p032) from the pBS228 antibiotic re-sistance plasmid of Pseudomonas aeruginosa (121), andproteins encoded by naphthalene-degradative plasmidspND071, pDTG1, and pND6-1 (37, 38). One homo-logue to the universal stress protein (UspA) of E. coli(122, 123) is also encoded in this pGRT1 fragment. Itwas demonstrated that a pGRT1 mutant in UspA wasmore sensitive to UV light than wild type, indicatingthat UspA has a role in the cells’ response to UV stress,possibly in cooperation with UvrD, RuvA, and RuvBhomologues (27). The presence of the pGRT1 plasmidwas also required for the release of siderophores intothe media in response to iron deficiency. This pheno-type was linked to the presence of ORF35, which is alsoencoded by pGRT1. Surprisingly, a BLAST search usingORF35 returns a putative and chromosomally encodedsulfate permease (sulP) found within Pseudomonas andBurkholderia strains (27), Q1although one homologue ofthis protein has been identified in the antibiotic resis-tance plasmid pOZ176 of P. aeruginosa 96 (124).
FIGURE 4 Schematic representation of Tn4653-like region (A) and second region en-coding stress resistance genes in pGRT1 (B). In red are indicated transposition-relatedgenes, in green are stress-related functions, and in blue are putative recombinases orintegrases. doi: 10.1128/microbiolspec.PLAS-0013-2013.f4
10 ASMscience.org/MicrobiolSpectrum
Segura et al.

pGRT1 was the first plasmid that was identified toconfer cells with solvent tolerant traits. Genes with highsequence homology to those of the ttgGHI operon havebeen found in other Pseudomonas strains but they arelocated either within the chromosome (L. Molina et al.,unpublished results)Q2 or are at undefined locations (125).Because the spectrum of substrates transported by theefflux pump is so wide and extrusion is so effective,plasmid pGRT1 has been considered a paradigm anda model for solvent tolerance. Furthermore, pGRT1encodes determinants for solvent tolerance, strongchemotaxis toward pollutants, survival under differentstress conditions, and genetic transfer to other Pseudo-monas strains, thus providing degrader bacteria theability to survive in heterogeneously polluted environ-ments and to achieve more efficient in situ bioremedia-tion (126). The localization of the efflux pump genes in atransposon and the multiple residues of transposases,recombinases, and integrases found in pGRT1 suggestthat genes associated with stress endurance have beenrecruited during evolution from different origins.
Although TtgGHI is an extreme example of the con-tributions of an efflux pump toward the resistanceto high concentrations of aromatic compounds, manyother bacteria have developed systems to cope withtoxic compounds present in their niche. An exampleis the nitrogen-fixing rhizobia bacteria, which are welladapted to live in polyphenol-rich environments. Poly-phenols are produced by plants under biotic and abioticstresses such as water stress, bacterial or fungal in-fections, UV radiation, and others (127). Some of thepolyphenols exuded by legume seeds and roots arenecessary to establish a symbiotic association betweenrhizobia with their host plant (reviewed in reference128). However, rhizobia have also developed severalmechanisms to thrive in the presence of otherwise toxicconcentrations of these compounds, including degrada-tion (129) and tolerance (130). One of the tolerancemechanisms in Rhizobium etli CFN42 is mediated bythe RmrAB efflux pump that is encoded by one of thestrain’s seven plasmids (131). The rhizobium multi-resistance genes (rmrAB) of R. etli are located withinthe 184-kbp plasmid known as p42b. This operon isinduced by bean exudates and is able to extrude narin-genin (a flavonoid inductor of nodulation), coumaric,and salicylic acid, a well-known inductor of plant de-fense mechanisms that is important for the survival ofR. etli in rhizospheric environments (130).
While extrusion of the toxic compounds forms partof the defensive mechanisms against pollutants, stressresponsemechanisms also play a role.Heat shock proteins
(HSPs), ubiquitous proteins that are the key players in thegeneral stress response system, act by mediating the fold-ing and transport of proteins. Organic solvents causeprotein damage leading to misfolding. It has been dem-onstrated that overexpression of heat shock proteins,such as the GroESL system inClostridium acetobutylicum(a butanol-producing bacteria), results in improved sol-vent tolerance and solvent production (132). HSP genescan be found in a broad range of antibiotic-resistantplasmids from the Enterobacteriaceae family and in plas-mids involved in rhizobia-plant symbiosis. The ORFsknown as groES and groEL encode small heat shockproteins that in Rhizobium sp. NGR234 are locatedwithin the pNGR234b plasmid (133). Interestingly, genesinvolved in the degradation of protocatechuate, opine,and naphthalene are also encoded in this plasmid as wellas 26 integrases and recombinases. Fragments with dif-ferent G+C content have been identified, suggesting thatlateral transfer of genetic material may have occurred.The byi_1p (BY123_D) plasmid of Burkholderia sp. YI23also contains genes for heat shock proteins. This strainwas isolated because it was able to degrade the organo-phosphorus insecticide fenitrothion. With three chro-mosomes and three plasmids, the strain has degradativegenes located in plasmid byi_2p (BY123_E) and byi_3p(BY123_F) (134). Whether or not the heat shock proteinsidentified within this strain are involved in tolerance hasnot yet been investigated.
Plasmids Involved in Heavy Metal ToleranceMetal-containing minerals are abundant on Earth.Natural events such as volcanic emissions, forest fires,deep-sea vents and geysers, together with anthropogenicactivities (mining, smelting, the creation of industrialchemical waste, etc.) have contributed to the distributionof these metals across the world. Some metals are nec-essary to sustain life (calcium and sodium are essentialmicronutrients; cobalt, copper, nickel, and zinc are vitalcofactors for enzymes and metalloproteins); however,at high concentrations they can have toxic effects onthe organisms. Other nonessential heavy metals, such asmercury, lead, and cadmium are considered toxic at anyconcentration (135). Most organisms have developedhomeostasis systems in order to maintain optimal in-tracellular concentrations of metals. This is achievedthrough the control of the processes of transport (importand export) and intracellular trafficking to prevent cel-lular damage and ensure cellular bioavailability. Metaltransporters move metal ions or chelates through mem-branes, metallochaperones transfer the metal to appro-priate cellular compartments or acceptor proteins, and
ASMscience.org/MicrobiolSpectrum 11
Plasmid-Mediated Tolerance Toward Environmental Pollutants

efflux pumps can eliminate excess and unwanted metals.Regulatory proteins serve to control the expression ofall these proteins in response to metal deprivation oroverload (136). While normal heavy metal concen-trations in soils are generally nontoxic, there are specificlocations where metal concentrations are high enough toprevent “normal” organisms from existing (137) andwhere only organisms with special mechanisms of tol-erance can thrive. Efflux pumps are one of the mainmechanisms for metal resistance in several bacteria, butmetal complexation and metal reduction can also con-tribute to tolerance (135).
Cupriavidus metallidurans CH34 has served as amodel microorganism for heavy metal resistance. Thebacterium was isolated from the sludge from a zincdecantation tank in Belgium (138) and tolerates highconcentrations of heavy metal ions, including but notnecessarily limited to Cu+, Cu2+, Ni2+, Zn2+, Co2+, Cd2+,CrO4
2−, Pb2+, Ag+, Au+, Au3+, HAsO42−, AsO2
−, Hg2+,Cs+, Bi3+, Tl+, SeO3
2−, SeO42−, and Sr2+. This strain is a
facultative chemolithoautotroph that is able to growusing toluene, benzene, or xylene as sole carbon source.This ability is conferred by a number of chromosomallyencoded genes as well as uncharacterized dioxygenaseslocated on plasmids. The genome of C. metalliduransCH34 is composed by four replicons: two main chro-mosomes and two megaplasmids named pMOL28 andpMOL30 (135, 139). This strain accomplishes metaldetoxification via the action of a wide variety of effluxsystems of the RND, P-type ATPase, and Cation Diffu-sion Facilitation (CDF) families. It also contains trans-porters that are specific to certain metal ions, wherebyefflux may be followed by metal sequestration or com-plexation. Many of these systems are localized intwo large plasmids known as pMOL28 and pMOL30(135).
The pMOL28 plasmid (139) has been associatedwith tolerance to Ni(II), Co(II), CrO2−
4 , and Hg(II). Thenickel/cobalt tolerance determinant present in pMOL28is the cnr operon, which is composed of the RND effluxpump CnrABC, and a regulatory complex formed bythe regulatory proteins CnrXY and the sigma factor,CnrH. Additionally, CnrT, a cation diffusion facilitator,is encoded by this region. Cation diffusion facilitatorsare transport systems driven by a chemiosmotic gradient(140) and represent another line of defense againstexcess metal cations. These proteins form a family ofmembrane-bound secondary transport systems for di-valent transition metal cations (141). The CupriavidusCH34 strain contains two additional CDF proteins,known as DmeF and FieF (140), that are chromosomally
encoded (142). Chromate tolerance is mediated by theefflux pump ChrA, which belongs to the chromate-efflux-related protein family. These systems are drivenby proton-motive force and remove chromate by effluxin cooperation with other proteins (143). Proteins re-lated to this efflux pump are the regulatory proteinsChrF, ChrI, and ChrB; ChrE, which is involved in theprocessing of chromium-glutathione complexes; andChrC, an iron superoxide dismutase (Fe-SOD). Homo-logues of the chr operon have also been found in aplasmid of the potential PAH degrader Arthrobacter sp.FB24 (144); in pCNB1, which provides ComamonasCNB-1 the ability to degrade chloronitrobenzenes (82);and in the pRA4000 plasmid, which allows P. putidaNCIMB 9866 to degrade 2,4-xylenol and p-cresol (145).Mercury resistance is widespread in plasmids. Althoughmercury concentration in soils and waters is normallyvery low, since the start of the industrial revolution,the amount of mercury mobilized and released intothe biosphere has increased, and, in some places, localmercury levels have increased by several thousandfoldabove background. The heightened levels of mercuryin the environment may be responsible for the wide-spread occurrence of the mer operon in nature (146).In pMOL28, mercury resistance is associated with thepresence of the merRTPADE operon (147). MerR actsas the key regulator of this operon (148). The highlytoxic Hg2+ cation is bound in the periplasm by MerP,imported into the cytoplasm by MerT, and reducedto metallic mercury by the MerA NADPH-dependentflavoprotein. While these enzymes are essential for mer-cury detoxification, in some organisms the operon con-tains additional genes includingmerB, which encodes anorganomercurial lyase; merC, a mercury transporter;and MerD, which is involved in regulation of the meroperon (reviewed in reference 149). The presence ofthe mer operon in other plasmids has been associatedwith other degradative pathways. Examples include thepW2 plasmid from P. putida W2 (NC_013176), in-volved in bisphenol A degradation; the byi_1p plasmidfrom Burkholderia sp. YI23, that degrades fenitrothion(134); the pI2 plasmid from Comamonadaceae for thedegradation of aniline (150); and in the pJP4 plasmidfrom R. eutropha JMP134, which allows degradation ofchloroaromatics (70).
The backbone genes of the pMOL28 plasmid arehighly homologous with the pHG1 plasmid core genes,which contains essential genes required for the faculta-tive lithoautotrophic and facultative anaerobic lifestylesof R. eutropha H16 (151) and pSym of Rhizobiumtaiwanensis LMG19424 (152). The presence of one
12 ASMscience.org/MicrobiolSpectrum
Segura et al.

genomic island in pMOL28 that contains all the deter-minants for heavy metal resistance has been described(139). This region is flanked by IS1071 and a partial ISfrom the Tn3 family.
The pMOL30 plasmid (234-kbp) encodes determi-nants for tolerance to Ag(I), Cd(II), Co(II), Cu(II),Hg(II), Pb(II), and Zn(II) (139). In addition to containinga mer operon that is similar what is found in pMOL28,pMOL30 contains the czcABC operon (one of the beststudied zinc-tolerance systems), which encodes an RNDefflux pump for cobalt, zinc, and cadmium resistancein C. metallidurans (140). The czcABC operon includesthree components of the proton antiporter efflux sys-tem: czcD, which encodes a cation diffusion facilitator;and czcSR, which encodes a two-component regulatorysystem. These regulators are homologues to those in-volved in copper resistance, and it is likely that this op-eron mediates copper extrusion (139). Three additionalproteins, CzcN, CzcI, and CzCE, are putative compo-nents of the resistance mechanisms, although their func-tion is currently unknown. Extrusion systems that arehomologous to czc have been found in catabolic plas-mids such pCAR3 of Sphingomonas sp. KA1, whichdegrades chlorinated dibenzo-p-dioxins (52), byi_1pof Burkholderia sp. YI23 (134), and pCMU01 from thechloromethane degrader Methylobacterium extorquens(153). Belonging to the same RND subgroup, thenccCBA efflux system (154) is also encoded by pMOL30.The cnr (of pMOL28) and ncc systems are related tothe NccA protein of the nickel-cobalt-cadmium deter-minant of the pTOM9 plasmid from Achromobacter sp.31A (155). The sil/cus system encoded by pMOL30 maybe involved in the efflux of silver and copper ions. ThesilABC operon encodes an efflux pump for the transportof these ions (140). Other factors that are not foundon the pMOL30 plasmid include: SilE, a protein thatacts as an extracellular metal-binding protein; and silRS,which encodes a two-component sensor and transcrip-tional responder. The silRS operon is widespread inplasmids originating from clinical strains probably be-cause silver compounds are used as antimicrobial agentsagainst bacterial infections (156).
Two of the 13 predicted P-type ATPases of C. metal-lidurans CH34 are present in pMOL30. P-type ATPasesconstitute a ubiquitous superfamily of transport proteinsthat are driven by ATP hydrolysis. Among their sub-strates are inorganic cations such as H+, Na+, K+, Mg2+,Ca2+, Cu+, Ag+, Zn2+, and Cd2+. In contrast to RNDpumps, these ATPases are also able to detoxify heavymetal cations bound to thiols. One of the P-typeATPases of pMOL30 is encoded by the large cluster
copVTMKNS1R1A1B1C1D1IJGFLQHE inwhichCopFis the P-type ATPase involved in Cu(II) efflux from thecytoplasm. CopT is believed to be involved in metaltransport from the periplasm to the cytoplasm; CopRand CopS are members of the two-component family ofsensor regulators; CopA is a multicopper oxidase; CopB isthought to be involved in Cu(I) fixation; CopC containsbinding sites for Cu(I) and Cu(II) and may be involvedin detoxification of both ions; copD encodes a putativechannel involved in loading CopA with Cu(II); CopI is aputative oxidoreductase; CopJ showed similarity withcytochrome c proteins; CopL may have a role in regula-tion; and CopH has homology with czcE. No function hasbeen assigned to CopV, CopK, CopM, CopN, CopG,CopQ, and CopE (157). The cop system is responsible forcopper and silver resistance of E. coli (158). Homologouscop genes have been identified in the metal toleranceplasmid pSPHCH01 of Sphingobium chlorophenolicumL-1 (159), and in the plasmid pISP0 from Sphingomonassp. MM-1, which also harbors genes involved in the deg-radation of γ-hexachlorocyclohexane (160). Homologuesof copA,mco (multicopper oxidase gene), cadD, and cadX(cadmium resistance) have been also recently identifiedin multiresistance plasmids from Staphylococcus aureusST398 (161). The colocalization of antimicrobial resis-tance genes and genes that confer tolerance to heavymetalscan facilitate the persistence and dissemination of thesetraits and could pose a serious problem for human health.Because of the widespread use of copper bactericides tocontrol bacterial infections in crop plants, the copABCDsystem (present in the chromosome of C. metalliduransCH34 and sharing low sequence homology with the copoperon of plasmid pMOL30) and the cusCBA operon arefound in plasmids of P. syringae pv. tomato (162) and pv.syringae UMAF0081 (163). The second P-type ATPasesystem present in pMOL30 is the lead resistance operon,pbr, which contains the following structural genes: pbrT,which encodes a Pb(II) uptake protein; pbrA, which en-codes a P-type Pb(II) effluxATPase; pbrB, which encodes apredicted integral membrane protein of unknown func-tion; and pbrC, which encodes a predicted prolipoproteinsignal peptidase. Downstream of pbrC, the pbrD geneencodes a Pb(II)-binding protein. The PpbrA promoter isregulated by PbrR, which belongs to the MerR family ofmetal ion-sensing regulatory proteins. This operon is alsofound in antibiotic/heavy metal tolerance plasmids ofEnterobacter cloacae and Klebsiella pneumoniae strainKCTC 2242 (164).
pMOL30 parAB genes are very similar to those inB. vietnamensis G4, and belong to a different incom-patibility group than pMOL28, as they are stably
ASMscience.org/MicrobiolSpectrum 13
Plasmid-Mediated Tolerance Toward Environmental Pollutants

maintained. pMOL30 was found to be able to transferat very low frequency. Many recombinases, IS, andtruncated IS have been identified in pMOL30, mostof them close to tolerance determinants. The czc andpbr clusters are flanked by the mercury transposonTn4380 on one side and by three mer genes that mightbe remnants of former rearrangements. The region con-taining the cop, sil, and mre-ncc clusters is flanked by acomplete ISRme10 element and a remnant of anothernamed ISRme10 (139).
Resistance to the toxic divalent heavy metal cations ofcobalt, nickel, cadmium, and zinc is widespread amongbacteria, although in many of them the genetic deter-minants of this characteristic are associated with chromo-somal genes (165). As mentioned above, C. metalliduransCH34also hasmany tolerance determinants located on thechromosome.
Resistance to arsenite (As[III], As(OH)3), arsenate(As[V], AsO4
3−), and antimony Sb(III) is widely foundamong Gram-negative and Gram-positive bacteria.Usually this resistance is determined by the presence ofan ars operon with a minimum of three cotranscribedgenes that include arsR (a regulatory repressor), arsB(a membrane transport pump), and arsC (a small in-tracellular arsenate reductase). Additionally, in this op-eron other proteins are encoded, including: ArsA, whichis an ATPase coupled to ArsB (together, these increasearsenite resistance); ArsD, which acts as an arsenitechaperon; and ArsP, which is a putative membranepermease (166). Although the ars operon is mostly lo-cated chromosomally, components are also distributedin plasmids, especially those encoding antibiotic re-sistance (167). One example of ars operon within aplasmid is found in Arthrobacter sp. Rue61, whichdegrades quinaldine through a pathway encoded bythe pARUE113 (pAL1) linear plasmid. This strain alsohas a circular plasmid, named pARUE232, which har-bors ars genes (64). Another example of a plasmid thathas the ars operon is the metal tolerance plasmidpOC167 from Oligotropha carboxidovorans OM5(168).
GENOMIC BIOAUGMENTATION AS ABIOTECHNOLOGICAL APPLICATIONThe existence of the described solvent, heavy metal tol-erance, and catabolic plasmids provides a useful batteryof biotechnological tools for remediation of pollutedenvironments. Contaminants are normally present incomplex mixtures that make degradation difficult. Someof these compounds, as described earlier, are toxic and
are able to avoid the action of the catabolic pathwaysof pollutant degrader microorganisms. Tolerance plas-mids and genes, once introduced into degrader strains,will allow the survival and the metabolic activity ofbacteria living in polluted environments. One of thestrategies to bioremediate contaminated environmentsis bioaugmentation with strains that have the capacityto degrade pollutants. However, it has been observedthat laboratory strains, on many occasions, were notable to thrive in the new environment, leading to un-successful bioremediation (169). Given the possibilitiesof plasmid exchange among strains, bioaugmentation incatabolic genes, not in bacteria, has taken relevance inthe past decade (170). This approach is called “geneticbioaugmentation” and involves the introduction ofbacteria harboring a relevant catabolic self-transmissibleplasmid that stimulates the horizontal gene transfer ofthe plasmids into indigenous microorganisms with bet-ter fitness for survival in the corresponding niche (171).To design an optimal genetic bioaugmentation is im-portant to choose the appropriate type of plasmid, tostudy the transfer capacity of the microorganism andthe stability of the plasmid in the new bacteria, as wellas to study the expression of the catabolic genes (171,172). Plasmid stability in the recipient strains may im-pact the effectiveness of bioremediation; highly stableplasmids could be necessary to clean up sites that con-tinuously receive contaminant input (171). Horizontalgene transfer from donor to indigenous bacteria fol-lowing the deliberate release of phenol-degrading labo-ratory bacteria was shown to be important in thedegradation of phenol in river waters continuouslypolluted by phenolic compounds (91).
It is well documented that the TOL plasmid can betransferred from P. putida to other Pseudomonas andErwinia strains (173), and to Enterobacteriaceae (174).Ikuma et al. (175) showed that the soil organic carbonpresent in sterilized soil slurries was sufficient fortransfer of TOL plasmid using different mixtures of re-cipient bacteria (including E. coli, E. cloacae, Serratiamarcescens, P. fluorescens, and P. putida BBC443).However, they also observed that addition of glucosesometimes improved the specific toluene degradationrates of Enterobacteriaceae transconjugants. The gua-nine and cytosine (G+C) genomic content of the recipi-ent strains have a clear influence on the expression of thetoluene degradation genes in TOL transconjugants andthe presence of alternative carbon sources (such as glu-cose) was shown to alleviate the limitations of the ex-pression of the acquired genes in some transconjugants(176). Observations made under laboratory conditions
14 ASMscience.org/MicrobiolSpectrum
Segura et al.

(177) indicated the importance of the initial recipient-to-donor cell density; however, in the experiments con-ducted by Ikuma et al. (175) this factor had only a minorimpact on plasmid conjugation. The spatial separationbetween donor and recipient strains can pose a problemfor the plasmid transfer and efficiency of biodegrada-tion; this problem has been tackled with the use of dif-ferent approaches. The use of earthworms has beenreported to increase the dispersal of donor (A. eutrophus[pJP4]) and recipient bacteria (P. fluorescens) and toincrease the frequency of transconjugants in soil micro-cosms (178). Similarly, biofilm structures, with openchannels and pores, allow the efficient transport ofdonor cells facilitating collision between bacteria andenhancing the introduction of mobile elements intoan existing microbial community (179). Interestingly,transfer of the TOL plasmid in sequencing batch biofilmreactors (SBBR) used for treating synthetic wastewatercontaining benzyl alcohol was observed on a laboratoryscale but not in the pilot scale bioreactor (180). Thisfailure can be attributed to different operational con-ditions in the two bioreactors and to the lack of selectivepressure in the pilot scale bioreactor, because completebenzyl alcohol removal was achieved during the first60 minutes of operation. While the presence of con-taminant has been reported to exert a positive effecton the efficiency of the pJP4 plasmid transfer undernonsterile conditions (181), the presence of toluene didnot influence the TOL transfer frequency when tested infilter matings (177) or in slurry soils at environmentallyrelevant concentrations (176).
Other approaches to bring together donor and re-cipient bacteria take advantage of the high number ofbacteria living near the plant roots; roots additionallyprovide a solid surface for conjugation (172). Althoughthe rhizosphere can improve the survival of micro-organisms in soil (182) and thus promote the eliminationof contaminants (183), it is also a complex environmentwhere plant and microorganisms established differentkinds of relationships. Because the rhizosphere is highlypopulated, microbes have to compete for niches andnutrients (184); furthermore, plants secrete harmfulcompounds such as phytoalexins, phenolic derivatives,and others. Transcriptional experiments performedwith the rhizospheric strain P. putida KT2440 showedthat they sense the rhizosphere as a stressful environ-ment (185). Some genetic traits involved in coping withthis high-stress environment have been shown to beencoded on plasmids; the best characterized examplesare the symbiotic plasmids from Rhizobium strains.These plasmids encode chemotaxis systems that recog-
nize rhizosphere nutrients, genes involved in catabolismof these nutrients, nutrient uptake, attachment to roots,and detoxification of harmful molecules produced byplants (reviewed in reference 186). To avoid competitionwith other rhizospheric microorganisms Barac et al.(187) used a new approach; it involved the introductionof the pTOM plasmid into B. cepacia L.S.2.4, a naturalendophyte of the legume,Lupinus luteus (yellow lupine).This approach improved the biodegradation of toluene.Inoculation of poplar trees with another yellow lupineendophyte, B. cepacia VM1468 containing the pTOMplasmid, also resulted in decreased toluene toxicity to-ward the plant. Although B. cepacia VM1468 was notable to successfully establish in the plant at high levels, itwas able to transfer the pTOM plasmid to other indig-enous endophytes, demonstrating the horizontal transferof the plasmid (188).
In recent years some “rhizospheric” plasmids havebeen identified. One of them is plasmid pQBR103 fromP. fluorescens SBW25. It belongs to a large group ofplasmids known to persist in the sugar beet phytosphere(rhizosphere and phyllosphere) and to be confinedwithin the Pseudomonas group. The pQBR103 is a 425-kbp plasmid that encodes one mercury resistance operonlocated in Tn5042 type II transposon, and also encodesthe RulAB proteins that confer UV light resistance. Fieldrelease trials have shown that pQBR103 confers a sig-nificant advantage to the SBW25 strain, 3 to 5 monthsafter planting out (189). Preliminary studies demon-strated that the advantages provided by this plasmid areimposed by the modulation of plant responses, althoughthe mechanism of the adaptive advantage remains un-clear (190). It has been shown that pQBR103 regulatesup to 48 proteins encoded on the chromosome of its hoststrain.
PERSPECTIVESEven though the decreasing costs of sequencing haveallowed more information to be obtained about cata-bolic or tolerance plasmids in the environment, there isstill a considerable lack of knowledge regarding plas-mids from noncultivable bacteria. Several approacheshave been followed to fill this gap. The most commonlyused technique has probably been “exogenous plasmidisolation.” In this technique, a cultivatable recipientstrain is used to acquire plasmids from different en-vironments through conjugation (191, 192, 193, 194,195). However, transfer is limited by the in/compati-bility of the indigenous bacteria for establishing in-teractions with the recipient bacteria and the capacity of
ASMscience.org/MicrobiolSpectrum 15
Plasmid-Mediated Tolerance Toward Environmental Pollutants

the plasmid to be transferred and replicate in the newstrain. To avoid the dependence on the plasmid-encodedtraits two different techniques have been described. Oneof them is called transposon-aided capture (TRACA)of plasmids. Genomic extracts from environmental sam-ples are treated with plasmid-safe DNase that digestlinear but not circular DNA, then circular plasmids aresubjected to transposition with insertions that containedan E. coli origin of replication and a selectable markerand latter transformed into E. coli strains (5). Thistechnique has been successfully used for the identifica-tion of plasmids resident in the human gut meta-genomebut, to date, we have not found any report describingthe utilization of this technique in the isolation of cata-bolic or tolerance plasmids. We can envisage certaindrawbacks for TRACA utilization in the identification ofcatabolic or tolerance plasmids; because of the large sizeof the catabolic plasmids, the DNA can be physicallybroken or damaged during the extraction process andthus later on digested by the DNase; also because of thislarge size transformation into E. coli strains would bedifficult. The approach followed by Kav and colleagues(196) is also based on the treatment of genomic envi-ronmental DNA with a plasmid-safe DNase, but theresultant circular plasmid DNA is subjected to amplifi-cation with DNA polymerase from phage φ29 and se-quenced. With the use of this technique, the plasmidomeof the bovine rumen has been identified (196). Imple-mentation and improvements in both protocols couldcircumvent the plasmid size issue of catabolic and tol-erance plasmids and will allow the analysis of theplasmidome of environmentally relevant niches.
The impact of catabolic plasmids on host cell physi-ology is another area of research that has to be furtherdeveloped (review in reference 197), because this infor-mation is important for improving the biotechnologicalutilization of bacteria carrying catabolic genes. It isknown that plasmid carriage generally leads to loss ofhost fitness (198), but catabolic and tolerance plasmidsare quite stable in their corresponding natural hosteven in the absence of selective pressure. The pCAR1plasmid is stably maintained not only in its naturalhost, P. resinovorans, but also in P. putida KT2440. Thetranscriptome of P. putida KT2440 harboring pCAR1was compared with that of the plasmid-free strainwhen growing on succinate (116). Interestingly, plas-mid pCAR1 did not significantly interfere with the hosttranscriptional patterns, and only parI, a homologueof the ParA family of plasmid partitioning proteins froma cryptic genomic island, showed a significant induc-tion in expression. It was latter demonstrated that ParI
interferes with the IncP-7 plasmid partitioning system(199), so the reason why pCAR1 is stable in KT2440remains unclear. Transcriptional analysis of pCAR1-carrying and pCAR1-free strains of P. putida KT2440,P. aeruginosa PAO1, and P. fluorescens Pf0-1 was per-formed by using high-density tiling arrays in order toidentify changes in transcriptional patterns. Although 70to 100 genes (depending on the strain) were found to bealtered in their expression levels in the pCAR1-carryingstrains compared with the pCAR1-free strains, only fourgenes were found to be upregulated in the three strains.Three of the genes form an operon that contained a Fur-associated gene (Fur is a global regulator in response toiron limitation); the fourth gene (phuR) is known to beupregulated under iron-limited conditions in P. aeru-ginosa. Among the commonly regulated genes in twoof the hosts, most of them were related to iron acquisi-tion and transport systems. Notably, pCAR1-carryingP. putida KT2440 and P. aeruginosa PAO1 have higherlevels of pyoverdine than the plasmid-free cells, whichsuggests that iron concentration may be key factor in themaintenance of pCAR1 in host cells (200). Backbone-related functions are also important in plasmid survivalin different strains; more than 90% of ORFs on thepCAR1 backbone were transcribed in six different hoststrains (201).The influence of the NAH7 plasmid on theP. putidaKT2440 transcriptome has also been analyzed.Despite the fact that the presence of the plasmid relievedthe stress caused by the presence of naphthalene in thehost strain, few genes were differentially expressed be-tween the containing and noncontaining strains whennaphthalene was not provided (202). The effect thatnucleoid-associated proteins (NAPs) have on the ex-pression of plasmids and host chromosomal genes hasto be further explored. Disruption of Pmr, a histone-likeprotein H1(H-NS) encoded on pCAR1, significantlyalters the expression levels of several genes in P. putidaKT2440 (203). The knockout inactivation of TurA, achromosomally encoded protein with structural simi-larity to H-NS proteins, resulted in enhanced trans-cription initiation from the Pu promoter, suggesting anegative regulatory role of TurA on Pu expression (204).
Many different biotechnological applications that relyon plasmid activities in a community are currently beingdeveloped for the elimination of contaminants (205).Although plasmid stability is usually high, segregantscan become a majority if plasmid-free cells have fastergrowing rates (206). To monitor the abundance ofplasmids, new rapid, noninvasive, in situ monitoringtechniques will have to be improved to detect the main-tenance of plasmids within the reactors. Traditional
16 ASMscience.org/MicrobiolSpectrum
Segura et al.

monitoring techniques, such as replica plating, selectivemarkers, and PCR detection (207) have, in general,low sensitivity and, in some cases, they can disrupt thefunction of the system. Although techniques based onthe introduction of gfp-labeled plasmids have been pro-posed as a noninvasive, in situmonitoring solution (208),the use of recombinant plasmids may carry some legalrestrictions in real-life applications.
CONCLUSIONSAlthough many catabolic pathways and resistance op-erons are located on plasmids, these genes can also onmany occasions be found on the chromosome. Never-theless, catabolic and tolerance plasmids that are generallyself-transmissible, are valuable tools for bioremediationapplications. Although in the 1980s there was avid in-terest in the discovery of contaminant degradation path-ways and their regulatory circuits, it was at the beginningof the 21st century that the simplification of sequencingtechniques allowed a better understanding of the plasmidsthat encode these pathways. It is now known that mostof the catabolic and tolerance genes encoded on plasmidsare associated with mobile elements, and that transfer toor from the host bacteria has occurred not only as a con-sequence of plasmid transfer, but also because of trans-position and recombination. The backbones of many ofthese plasmids are related to the IncP incompatibilitygroup, and, in many cases, plasmids share a similarbackbone but are loaded with different catabolic or tol-erance genes (i.e., the backbone of pBVIE04 that carriesthe toluene monoxygenase genes and the backbone ofpGRT1 that carries the toluene tolerance operon).
There is a lack of information about plasmids inthe environment that are harbored by noncultivatablestrains; this knowledge gap needs to be filled. Awarenessof the abundance and significance of these plasmids inthe environment will allow us to better understand thefunction of ecosystems. Understanding the mechanismsbehind plasmid stability will improve the outcomes ofthe biotechnological application of these plasmids.
ACKNOWLEDGMENTSConflicts of interest: We declare that we have no conflicts.
REFERENCES1. Segura A, Rojas A, Hurtado A, Huertas MJ, Ramos JL. 2003. Com-parative genomic analysis of solvent extrusion pumps in Pseudomonasstrains exhibiting different degrees of solvent tolerance. Extremophiles7:371–376.2. Williams PA, Jones RM, Zysltra GJ. 2004. Genomics of catabolicplasmids, p 165–195. In Ramos JL (ed), Pseudomonas, vol 1. KluwerAcademic, Plenum Publishers, New York.
3. Ogawa N, Chackrabarty AM, Zaborina O. 2004. Degradative plas-mids, p 341–392. In Funnell BE, Phillips GJ (ed), Plasmid Biology. ASMPress, Washington, DC.4. Springael D, Top EM. 2004. Horizontal gene transfer and microbialadaptation to xenobiotics: new types of mobile genetic elements andlessons from ecological studies. Trends Microbiol 12:53–58.5. Jones BV, Marchesi JR. 2007. Transposon-aided capture (TRACA) ofplasmids resident in the human gut mobile metagenome. Nat Methods4:55–61.6. Parales RE, Parales JV, Pelletier DA, Ditty JL. 2008. Diversity of mi-crobial toluene degradation pathways. Adv Appl Microbiol 64:1–73.7. Bertini L, Calafaro V, Proietti S, Caporale C, Capasso P, Caruso C,Di Donato A. 2013. Deepening TOL and TAU catabolic pathways ofPseudomonas sp. OX1: Cloning, sequencing and characterization of thelower pathways. Biochimie 95:241–250.
8. Williams PA, Murray K. 1974. Metabolism of benzoate and themethylbenzoates by Pseudomonas putida (arvilla) mt-2: evidence for theexistence of a TOL plasmid. J Bacteriol 120:416–423.
9. Greated A, Lambertsen L, Williams PA, Thomas CM. 2002. Completesequence of the IncP-9 TOL plasmid pWW0 from Pseudomonas putida.Environ Microbiol 4:856–871.
10. Gallegos MT, Williams PA, Ramos JL. 1997. Transcriptional controlof the multiple catabolic pathways encoded on the TOL plasmid pWW53of Pseudomonas putida MT53. J Bacteriol 179:5024–5029.
11. Ramos JL, Marques S, Timmis KN. 1997. Transcriptional controlof the Pseudomonas TOL plasmid catabolic operons is achieved throughan interplay of host factors and plasmid-encoded regulators. Annu RevMicrobiol 51:341–371.
12. Silva-Roche R, de Lorenzo V. 2013. The TOL network of Pseudo-monas putidamt-2 processes multiple environmental inputs into a narrowresponse space. Env Microbiol 15:271–286.
13. Bayley SA, Duggleby CJ, Worsey MJ, Williams PA, Hardy KB,Broda P. 1977. Two modes of loss of the Tol function from Pseudomonasputida mt-2. Mol Gen Genet 154:203–204.
14. Muñoz R, Hernández M, Segura A, Gouveia J, Rojas A, Ramos JL,Villaverde S. 2009. Continuous cultures of Pseudomonas putida mt-2overcome catabolic function loss under real case operating conditions.Appl Microbiol Biotech 83:189–198.
15. Tsuda M, Iino T. 1987. Genetic analysis of a transposon carryingtoluene degrading genes on a TOL plasmid pWW0. Mol Gen Genet 210:270–276.
16. TsudaM, Iino T. 1988. Identification and characterization of Tn4653,a transposon covering the toluene transposon Tn4651 on TOL plasmidpWW0. Mol Gen Genet 213:72–77.
17. Williams PA, Jones RM, Shaw LE. 2002. A third transposableelement, ISPpu12, from the toluene-xylene catabolic plasmid pWW0of Pseudomonas putida mt-2. J Bacteriol 184:6572–6580.
18. Williams PA, Worsey MJ. 1976. Ubiquity of plasmids in coding fortoluene and xylene metabolism in soil bacteria: evidence for the existenceof new TOL plasmids. J Bacteriol 125:818–828.
19. Keil H, Keil S, Pickup W, Williams PA. 1985. Evolutionary conser-vation of genes coding for meta pathway enzymes within TOL plasmidspWW0 and pWW53. J Bacteriol 164:887–895.
20. Chatfield LK, Williams PA. 1986. Naturally occurring TOL plasmidsin Pseudomonas strains carry either two homologous or two nonhomol-ogous catechol 2,3-oxygenase genes. J Bacteriol 168:878–885.
21. Sentchilo VS, Perebituk AN, Zehnder AJ, van der Meer JR. 2000.Molecular diversity of plasmids bearing genes that encode toluene andxylene metabolism in Pseudomonas strains isolated from different con-taminated sites in Belarus. Appl Environ Microbiol 66:2842–2852.
22. Yano H, Garruto CE, Sota M, Ohtsubo Y, Nagata Y, Zylstra GJ,Williams PA, Tsuda M. 2007. Complete sequence determination com-
ASMscience.org/MicrobiolSpectrum 17
Plasmid-Mediated Tolerance Toward Environmental Pollutants

bined with analysis of transposition/site-specific recombination events toexplain genetic organization of IncP-7 TOL plasmid pWW53 and relatedmobile genetic elements. J Mol Biol 369:11–26.23. Yano H,Miyakoshi M, Ohshima K, TabataM, Nagata Y, Hattori M,Tsuda M. 2010. Complete nucleotide sequence of TOL plasmid pDK1provides evidence for evolutionary history of IncP-7 catabolic plasmids.J Bacteriol 192:4337–4347.24. Shields MS, Reagin MJ, Gerger RR, Campbell R, Somerville C. 1995.TOM, a new aromatic degradative plasmid from Burkholderia (Pseudo-monas) cepacia G4. Appl Environ Microbiol 61:1352–1356.25. Shields MS, Montgomery SO, Cuskey SM, Chapman PJ, PritchardPH. 1991. Mutants of Pseudomonas cepacia G4 defective in catabolismof aromatic compounds and trichloroethylene. Appl Environ Microbiol57:1935–1941.26. Nelson MJK, Montgomery SO, Mahaffey WR, Pritchard PH. 1987.Biodegradation of trichloroethylene and involvement of an aromaticbiodegradative pathway. Appl Environ Microbiol 53:949–954.27. Molina L, Duque E, Gómez MJ, Krell T, Lacal J, García-Puente A,García,V,MatillaMA, Ramos JL, Segura A. 2011. The pGRT1 plasmid ofPseudomonas putida DOT-T1E encodes functions relevant for survivalunder harsh conditions in the environment. Environ Microbiol 13:2315–2327.28. Rheinwald JG, Chakrabarty AM, Gunsalus IC. 1973. A transmissibleplasmid controlling camphor oxidation in Pseudomonas putida. Proc NatlAcad Sci USA 70:885–889.29. Dunn NW, Gunsalus IC. 1973. Transmissible plasmid coding earlyenzymes of naphthalene oxidation in Pseudomonas putida. J Bacteriol114:974–979.30. Yen KM, Gunsalus IC. 1982. Plasmid gene organization: naphthalene/salicylate oxidation. Proc Natl Acad Sci USA 79:874–879.31. Yen KM, Serdar CM. 1988. Genetics of naphthalene catabolism inpseudomonads. Crit Rev Microbiol 15:247–268.32. Denome SA, Stanley DC, Olson ES, Young KD. 1993. Metabolismof dibenzothiophene and naphthalene in Pseudomonas strains: completeDNA sequence of upper naphthalene catabolic pathway. J Bacteriol 175:6890–6901.33. SimonMJ, Osslund TD, Saunders R, Ensley BD, Suggs S, Harcourt A,Suen WC, Cruden DL, Gibson DT, Zylstra GJ. 1993. Sequences of genesencoding naphthalene dioxygenase in Pseudomonas putida strains G7 andNCIB 9816-4. Gene 127:31–37.34. Eaton RW. 1994. Organization and evolution of naphthalene cata-bolic pathways: sequence of the DNA encoding 2-hydroxy-chromene-2-carboxylate isomerase and trans-o-hydroxybenzylidenepyruvate hydratase-aldolase from the NAH7 plasmid. J Bacteriol 176:7757–7762.35. Grimm AC, Harwood CS. 1999. NahY, a catabolic plasmid-encodedreceptor required for chemotaxis of Pseudomonas putida to the aromatichydrocarbon naphthalene. J Bacteriol 181:3310–3316.36. Schell MA, Sukordhaman M. 1989. Evidence that the transcriptionactivator encoded by the Pseudomonas putida nahR gene is evolutionarilyrelated to the transcription activators encoded by the Rhizobium nodDgenes. J Bacteriol 171:1952–1959.37. Dennis JJ, Zylstra GJ. 2004. Complete sequence and genetic orga-nization of pDTG1, the 83 kilobase naphthalene degradation plasmidfrom Pseudomonas putida strain NCIB 9816-4. J Mol Biol 341:753–768.38. Li W, Shi J, Wang X, Han Y, Tong W, Ma L, Liu B, Cai B. 2004.Complete nucleotide sequence and organization of the naphthalene cata-bolic plasmid pND6-1 from Pseudomonas sp. strain ND6.Gene 336:231–240.39. Sota M, Yano H, Ono A, Miyazaki R, Ishii H, Genka H, TopEM, Tsuda M. 2006. Genomic and functional analysis of the IncP-9naphthalene-catabolic plasmid NAH7 and its transposon Tn4655 sug-gests catabolic gene spread by a tyrosine recombinase. J Bacteriol 188:4057–4067.
40. Tsuda M, Iino T. 1990. Naphthalene degrading genes on plasmidNAH7 are on a defective transposon. Mol Gen Genet 223:33–39.41. Sanseverino J, Applegate BM, King JMH, Sayler GS. 1993. Plasmid-mediated mineralization of naphthalene, phenanthrene and anthracene.Appl Environ Microbiol 59:1931–1937.42. Kiyohara H, Nagao K. 1978. The catabolism of phenanthrene andnaphthalene in bacteria. J Gen Microbiol 105:69–75.43. Laurie AD, Lloyd-Jones G. 1999. The phn genes of Burkholderia sp.strain RP007 constitute a divergent gene cluster for polycyclic aromatichydrocarbon catabolism. J Bacteriol 181:531–540.
44. PinyakongO, HabeH, Omori T. 2003. The unique aromatic catabolicgenes in sphingomonads degrading polycyclic aromatic hydrocarbons(PAHs) J Gen Appl Microbiol 49:1–19.
45. Basta T, Keck A, Klein J, Stolz A. 2004. Detection and characteriza-tion of conjugative degradative plasmids in xenobiotic-degrading Sphin-gomonas strains. J Bacteriol 186:3862–3872.
46. RomineMF, Stillwell LC, Wong K-K, Thurston SJ, Sisk EC, Sensen C,Gaasterland T, Fredrickson JK, Saffer JD. 1999. Complete sequence of a184-kilobase catabolic plasmid from Sphingomonas aromaticivoransF199. J Bacteriol 181:1585–1602.
47. Feng X, Ou X-L, Ogram A. 1997. Plasmid-mediated mineralizationof carbofuran by Sphingomonas sp. CF-06. Appl Environ Microbiol 63:1332–1337.
48. Basta I, Buerger S, Stolz A. 2005. Structural and replicative diversity oflarge plasmids from sphingomonads that degrade polycyclic aromaticcompounds and xenobiotics. Microbiology 151:2025–2037.
49. Keck A, Conradt D, Mahler A, Stolz A, Mattes R, Klein J. 2006.Identification and functional analysis of the genes for naphthalenesulfonatecatabolism by Sphingomonas xenophaga BN6. Microbiology 152:1929–1940.
50. Maeda K, Nojiri H, Shintani M, Yoshida T, Habe H, Omori T. 2003.Complete nucleotide sequence of carbazole/dioxin-degrading plasmidpCAR1 in Pseudomonas resinovorans strain CA10 indicates its mosaicityand the presence of large catabolic transposon Tn4676. J Mol Biol 326:21–33.
51. Takahasi Y, Shintani M, Yamane H, Nojiri H. 2009. The completenucleotide sequence of pCAR2: pCAR2 and pCAR1 were structurallyidentical IncP-7 carbazole degradative plasmids. Biosci Biotechnol Bio-chem 73:744–746.
52. Shintani M, Urata M, Inoue K, Eto K, Habe H, Omori T, Yamane H,Nojiri H. 2007. The Sphingomonas plasmid pCAR3 is involved in com-plete mineralization of carbazole. J Bacteriol 189:2007–2020.
53. Nam J-W, Nojiri H, Noguchi H, Uchimura H, Yoshida T. Habe H,Yamane H, Omori T. 2002. Purification and characterization of carbazole1,9a-dioxigenase, a three-component dioxygenase system of Pseudomo-nas resinovorans strain CA10. Appl Environ Microbiol 68:5882–5890.
54. Nojiri H. 2012. Structural and molecular genetic analyses of thebacterial carbazole degradation system. Biosci Biotechnol Biochem 76:1–18.
55. Ashikawa Y, Fujimoto Z, Usami Y, Inoue K, Noguchi H, Yamane H,Nojiri H. 2012. Structural insight into the substrate- and dioxygen-binding manner in the catalytic cycle of Rieske nonheme iron oxygenasesystem, carbazole 1,9a-dioxygenase. BMC Struct Biol 12:15. doi: 10.1186/1472-6807-12-15.
56. Shintani M, Matsumoto T, Yoshikawa H, Yamane H, Ohkuma M,Nojiri H. 2011. DNA rearrangement has occurred in the carbazole-degradative plasmid pCAR1 and the chromosome of its unsuitable host,Pseudomonas fluorescens PF0-1. Microbiology 157:3405–3416.
57. Solinas F, Marconi AM, Ruzzi M, Zennaro E. 1995. Characterizationand sequence of a novel insertion sequence, IS1162, from Pseudomonasfluorescens. Gene 155:77–82.
58. Schneiker S, Kosier B, Puhler A, Selbitschka W. 1999. TheSinorhizobium meliloti insertion sequence (IS) element ISRm14 is related
18 ASMscience.org/MicrobiolSpectrum
Segura et al.

to a previously unrecognized IS element located adjacent to the Esche-richia coli locus of enterocyte effacement (LEE) pathogenicity island. CurrMicrobiol 39:274–281.59. Yao CC, Wong DTS, Poh CL. 1998. IS1491 from Pseudomonasalcaligenes NCIB 9867: characterization and distribution among Pseu-domonas species. Plasmid 39:187–195.60. Nojiri H, Sekiguchi H, Maeda K, Urata M, Nakai S, Yoshida T,Habe H, Omori T. 2001. Genetic characterization and evolutionaryimplications of a car gene cluster in carbazole degrader Pseudomonas sp.strain CA10. J Bacteriol 183:3663–3679.61. Overhage J, Sielker S, Hombrug S, Parschat K, Fetzner S. 2005.Identification of large linear plasmids in Arthrobacter spp. encoding thedegradation of quinaldine to anthranilate. Microbiology 151:491–500.62. Parschat K, Hauer B, Kappl R, Kraft R, Hüttermann J, Fetzner S.2003. Gene cluster of Arthrobacter ilicis Rü61a involved in the degrada-tion of quinaldine to anthranilate. Characterization and functional ex-pression of the quinaldine 4-oxidase qoxLMS genes. J Biol Chem 278:27483–27494.63. Parschat K, Overhage J, Strittmatter AW, Henne A, Gottschalk G,Fetzner S. 2007. Complete nucleotide sequence of the 113-kilobase linearcatabolic plasmid pAL1 of Arthrobacter nitroguajacolicus Rü61a andtranscriptional analysis of genes involved in quinaldine degradation.J Bacteriol 189:3855–3867.64. Niewerth H, Schuldes J, Parschat K, Kiefer P, Vorholt JA, Daniel R,Fetzner S. 2012. Complete genome sequence and metabolic potential ofthe quinaldine-degrading bacterium Arthrobacter sp. Rue61a. BMCGenomics 13:534. doi: 10.1186/1471-2164-13-534.65. Niewerth H, Parschat K, RauschenbergM, Ravoo BJ, Fetzner S. 2013.The PaaX-type repressor MeqR2 of Arthrobacter sp. strain Rue61a, in-volved in the regulation of quinaldine catabolism, binds to its own pro-moter and to catabolic promoters and specifically responds to anthraniloylcoenzyme A. J Bacteriol. 195:1068–1080.
66. Wu W, Leblanc SKD, Piktel J, Jensen SE, Roy KL. 2006. Predictionand functional analysis of the replication origin of the linear plasmidpSCL2 in Streptomyces clavuligerus. Can J Microbiol 52:293–300.
67. Don RH, Pemberton JM. 1981. Properties of six pesticide degradationplasmids isolated from Alcaligenes paradoxus and Alcaligenes eutrophus.J Bacteriol 145:681–686.
68. Laemmli CM, Leveau JHJ, Zehnder AJB, van der Meer JR. 2000.Characterization of a second tfd gene cluster for chlorophenol andchlorocatechol metabolism on plasmid pJP4 in Ralstonia eutrophaJMP134 (pJP4). J Bacteriol 182:4165–4172.
69. Pérez-Pantoja D, Guzmán L, Manzano M, Pieper DH, González B.2000. Role of tfdCIDIEIFI and tfdDIICIIEIIFII gene modules in catabolismof 3-chlorobenzoate by Ralstonia eutropha JMP134 (pJP4). Appl EnvironMicrobiol 66:1602–1608.
70. Trefault N, De la Iglesia R,Molina AM,ManzanoM, Ledger T, Perez-Pantoja D, Sánchez MA, Stuardo M, Gonzalez B. 2004. Genetic organi-zation of the catabolic plasmid pJP4 from Ralstonia eutropha JMP134(pJP4) reveals mechanisms of adaptation to chloroaromatic pollutants andevolution of specialized chloroaromatic degradation pathways. EnvironMicrobiol 6:655–668.
71. Vedler E, Vahter M, Heinaru A. 2004. The completely sequencedplasmid pEST4011 contains a novel IncP1 backbone and a catabolictransposon harboring tfd genes for 2,4-dichlorophenoxyacetic acid deg-radation. J Bacteriol 186:7161–7174.
72. Poh RP-C, Smith ARW, Bruce IJ. 2002. Complete characterization ofTn5530 from Burkholderia cepacia strain 2a (pIJB1) and studies of 2,4-dichlorophenoxyacetate uptake by the organism. Plasmid 48:1–12.
73. Tsoi TV, Plotnikova EG, Cole JR, Guerin WF, Bagdasarian M,Tiedje JM. 1999. Cloning, expression and nucleotide sequence of thePseudomonas aeruginosa 142 ohb genes coding for oxygenolytic orthodehalogenation of halobenzoates. Appl Environ Microbiol 65:2151–2162.
74. Hickey WJ, Sabat G, Yuroff AS, Arment AR, Perez-Lesher J. 2001.Cloning, nucleotide sequencing and functional analysis of a novel, mobilecluster of biodegradation genes from Pseudomonas aeruginosa strain JB2.Appl Environ Microbiol 67:4603–4609.75. de Souza ML, Sadowsky MJ, Seffernick J, Martinez B, Wackett LP.1998. The atrazine catabolism genes are widespread and highly conserved.J Bacteriol 180:1951–1954.76. de Souza ML, Wackett LP, Sadowsky MJ. 1998. The atzABC genesencoding atrazine catabolism are located on a self-transmissible plasmid inPseudomonas sp. strain ADP. Appl Environ Microbiol 64:2323–2326.77. Martinez B, Tomkins J, Wackett LP, Wing R, Sadowsky MJ. 2001.Complete nucleotide sequence and organization of the atrazine catabolicplasmid pADP-1 from Pseudomonas sp. strain ADP. J Bacteriol 183:5684–5697.78. Król JE, Penrod JT, McCaslin H, Rogers LM, Yano H, Stancik AD,Dejonghe W, Brown CJ, Parales RE, Wuertz S, Top EM. 2011. Role ofIncP-1ß plasmids pWDL7::rfp and pNB8c in chloroaniline catabolism asdetermined by genomic and functional analyses. Appl Environ Microbiol78:828–838.79.WuJF, SunCW, JiangCY, LiuZP, Liu SJ.2005.Anovel 2-aminophenol1,6-dioxygenase involved in the degradation of p-chloronitrobenzene byComamonas strain CNB-1 purification, properties, genetic cloning andexpression in Escherichia coli. Arch Microbiol 183:1–8.80. Wu JF, Jiang CY, Wang BJ, Ma YF, Liu ZP, Liu SJ. 2006. Novelpartial reductive pathway for 4-chloronitrobenzene and nitrobenzenedegradation in Comamonas sp. strain CNB-1. Appl Environ Microbiol72:1759–1765.81. Liu L, Wu JF, Ma YF, Wang SY, Zhao GP, Liu SJ. 2007. A noveldeaminase involved in chloronitrobenzene and nitrobenzene degradationwith Comamonas sp. strain CNB-1. J Bacteriol 189:2677–2682.82. Ma YF, Wu JF, Wang SY, Jiang CY, Zhang Y, Qi SW, Liu L,Zhao GP, Liu SJ. 2007. Nucleotide sequence of plasmid pCNB1 fromComamonas strain CNB-1 reveals novel genetic organization and evolu-tion for 4-chloronitrobenzene degradation. Appl Environ Microbiol 73:4477–4483.83. Nagata Y, Kamakura M, Endo R, Miyazaki R, Ohtsubo Y, Tsuda M.2006. Distribution of γ-hexachlorocyclohexane-degrading genes on threereplicons in Sphingobium japonicum UT26. FEMS Microbiol Lett256:112–118.84. Miyazaki R, Sato Y, Ito M, Ohtsubo Y, Nagata Y, Tsuda M.2006. Complete nucleotide sequence of an exogenously isolated plasmidpLB1, involved in γ-hexachlorocyclohexane degradation. Appl EnvironMicrobiol 72:6923–6933.85. Smalla K, Osborn AM, Wellington EMH. 2000. Isolation and char-acterization of plasmids from bacteria, p 207–248. In Thomas CM(ed), The Horizontal Gene Pool–Bacterial Plasmids and Gene Spread.Harwood Academic Publishers, Amsterdam, The Netherlands.86. Junker F, Cook AM. 1997. Conjugative plasmids and the degradationof arylsulfonates in Comamonas testosteroni. Appl Environ Microbiol63:2403–2410.
87. Tralau T, Cook AM, Ruff J. 2001. Map of the IncP1ß plasmidpTSA encoding the widespread genes (tsa) for p-toluenesulfonate degra-dation inComamonas testosteroni T-2.Appl EnvironMicrobiol 67:1508–1516.
88. Kivisaar MA, Habicht JK, Heinaru AL. 1989. Degradation of phenoland m-toluate in Pseudomonas sp. strain EST1001 and its Pseudomonasputida transconjugants is determined by a multiplasmid system. J Bac-teriol 171:5111–5116.
89. Kivisaar M, Hõrak R, Kasak L, Heinaru A, Habicht J. 1990. Selectionof independent plasmids determining phenol degradation in Pseudomonasputida and the cloning and expression of genes encoding phenol mono-oxygenase and catechol 1,2-dioxygenase. Plasmid 24:25–36.
90. Kallastu A, Hõrak R, Kivisaar M. 1998. Identification and charac-terization of IS1411, a new insertion sequencewhich causes transcriptional
ASMscience.org/MicrobiolSpectrum 19
Plasmid-Mediated Tolerance Toward Environmental Pollutants

activation of the phenol degradation genes in Pseudomonas putida.J Bacteriol 180:5306–5312.91. Peters M, Heinaru E, Talpsep E, Wand H, Stottmeister U, Heinaru A,Nurk A. 1997. Acquisition of a deliberately introduced phenol degrada-tion operon, pheBA, by different indigenous Pseudomonas species. ApplEnviron Microbiol 63:4899–4906.92. Shingler V, Franklin FC, TsudaM, Holroyd D, Bagdasarian M. 1989.Molecular analysis of a plasmid-encoded phenol hydroxylase fromPseudomonas CF600. J Gen Microbiol. 135:1083–1092.93. Powlowski J, Shingler V. 1994. Genetics and biochemistry of phenoldegradation by Pseudomonas sp. CF600. Biodegradation 5:219–236.94. van Beilen JB, Wubbolts MG, Witholt B. 1994. Genetics of alkaneoxidation by Pseudomonas oleovorans. Biodegradation 5:161–174.95. van Beilen JB, Panke S, Lucchini S, Franchini AG, Röthlisberger M,Witholt B. 2001. Analysis of Pseudomonas putida alkane degradationgene clusters and flanking insertion sequences: evolution and regulation ofthe alk genes. Microbiology 147:1621–1630.96. DinamarcaMA, Aranda-Olmedo I, Puyet A, Rojo F. 2003. Expressionof the Pseudomonas putida OCT plasmid alkane degradation pathway ismodulated by two different global control signals: evidence from contin-uous cultures. J Bacteriol. 185:4772–4778.97. Sikkema J, de Bont JA, Poolman B. 1995. Mechanisms of membranetoxicity of hydrocarbons. Microbiol Rev 59:201–222.98. Udaondo Z, Molina L, Daniels C, Gómez MJ, Molina-Henares MA,Matilla MA, Roca A, Fernández M, Duque E, Segura A, Ramos JL. 2013.Metabolic potential of the organic-solvent tolerant Pseudomonas putidaDOT-T1E deduced from its annotated genome.Microb Biotechnol 6:598–611.99. Ramos JL, Duque E, Huertas MJ, Haïdour A. 1995. Isolation andexpansion of the catabolic potential of a Pseudomonas putida strain ableto grow in the presence of high concentrations of aromatic hydrocarbons.J Bacteriol 177:3911–3916.100. Mosqueda G, Ramos-Gonzalez MI, Ramos JL. 1999. Toluene me-tabolism by the solvent tolerant Pseudomonas putidaDOT-T1 strain, andits role in solvent impermeabilization. Gene 232:69–76.101. Udaondo Z, Duque E. Fernández M, Molina L, de la Torre J,Bernal P, Niqui JL, Pini C, Roca A, Matilla MA, Molina-Henares MA,Silva-Jiménez H, Navarro-Avilés G, Busch, A, Lacal J, Krell T, Segura A,Ramos JL. 2012. Analysis of solvent tolerance in Pseudomonas putidaDOT-T1E based on its genome sequence and a collection of mutants.FEBS Lett 586:2932–2938.102. Rodríguez-Herva JJ, García V, Hurtado A, Segura A, Ramos JL.2007. The ttgGHI solvent efflux pump operon of Pseudomonas putidaDOT-T1E is located on a large self-transmissible plasmid. EnvironMicrobiol 9:1550–1561.103. Segura A, Molina L, Fillet S, Krell T, Bernal P, Muñoz-Rojas J,Ramos JL. 2012. Solvent tolerance in Gram-negative bacteria. Curr OpinBiotechnol 23:415–421.104. Rojas A, Duque E, Mosqueda G, Golden G, Hurtado A, Ramos JL,Segura A. 2001. Three efflux pumps are required to provide efficienttolerance to toluene to toluene in Pseudomonas putida DOT-TIE. J Bac-teriol 183:3967–3973.
105. Blair JM, Piddock LJ. 2009 Structure, function and inhibitionof RND efflux pumps in Gram-negative bacteria: an update. Curr OpinMicrobiol 5:512–519.
106. Hinchliffe P, Symmons MF, Hughes C, Koronakis V. 2013. Structureandoperationof bacterial tripartite pumps.AnnuRevMicrobiol67:221–242.
107. Murakami S, Nakashima R, Yamashita E, Yamaguchi A. 2002.Crystal structure of bacterial multidrug efflux transporter AcrB. Nature419:587–593.
108. Sennhauser G, BukowskaMA, Briand C, Grütter MG. 2009. Crystalstructure of the multidrug exporter MexB from Pseudomonas aeruginosa.J Mol Biol 389:134–145.
109. Koronakis V, Sharff A, Koronakis E, Luisi B, Hughes C. 2000.Crystal structure of the bacterial membrane protein TolC central tomultidrug efflux and protein export. Nature 405:914–919.110. Nikaido H, Zgurskaya HI. 2001 AcrAB and related multidrug effluxpumps of Escherichia coli. J Mol Microbiol Biotechnol 3:215–218.111. Symmons MF, Bokma E, Koronakis E, Hughes C, Koronakis V.2009. The assembled structure of a complete tripartite bacterial multidrugefflux pump. Proc Natl Acad SciUSA 106:7173–7178.112. Ramos J L, Duque E, Godoy P, Segura A. 1998. Efflux pumps in-volved in toluene tolerance in Pseudomonas putida DOT-T1E. J Bacteriol180:3323–3329.113. Mosqueda G, Ramos JL. 2000. A set of genes encoding a secondtoluene efflux system in Pseudomonas putida DOT-T1E is linked to thetod genes for toluene metabolism. J Bacteriol 182:937–943.114. Rojas A, Segura A, Guazzaroni ME, Terán W, Hurtado A, GallegosMT, Ramos JL. 2003. In vivo and in vitro evidence that TtgV is thespecific regulator of the TtgGHI multidrug and solvent efflux pump ofPseudomonas putida. J Bacteriol 185:4755–4763.115. Lacal J, Muñoz-Martínez F, Reyes-Darías JA, Duque E, Matilla M,Segura A, Calvo JJ, Jímenez-Sánchez C, Krell T, Ramos JL. 2011. Bac-terial chemotaxis towards aromatic hydrocarbons in Pseudomonas. En-viron Microbiol 13:1733–1744.116. Miyakoshi M, Shintani M, Terabayashi T, Kai S, Yamane H,Nojiri H. 2007. Transcriptome analysis of Pseudomonas putida KT2440harboring the completely sequenced IncP-7 plasmid pCAR1. J Bacteriol189:6849–6860.117. Iwaki H, Muraki T, Ishihara S, Hasegawa Y, Rankin KN,Sulea T, Boyd J, Lau PC. 2007. Characterization of a pseudomonad2-nitrobenzoate nitroreductase and its catabolic pathway-associated 2-hydroxylaminobenzoate mutase and a chemoreceptor involved in 2-nitrobenzoate chemotaxis. J Bacteriol 189:3502–3514.118. Yoakum GH, Cole RS. 1977. Role of ATP in removal of psoralencross-links from DNA of Escherichia coli permeabilized by treatment withtoluene. J Biol Chem 252:7023–7030.119. Rahmati S, Yang S, Davidson AL, Zechiedrich EL. 2002. Controlof the AcrAB multidrug efflux pump by quorum-sensing regulator SdiA.Mol Microbiol 43:677–685.120. Nikaido E, Yamaguchi A, Nishino K. 2008. AcrAB multidrug effluxpump regulation in Salmonella enterica serovar Typhimurium by RamA inresponse to environmental signals. J Biol Chem 283:24245–24253.121. Haines AS, Jones K, Batt SM, Kosheleva IA, Thomas CM. 2007.Sequence of plasmid pBS228 and reconstruction of the IncP-1a phylogeny.Plasmid 58:76–83.122. Kvint K, Nachin L, Diez A, Nyström T. 2003. The bacterial universalstress protein: function and regulation. Curr Opin Microbiol 6:140–145.123. Nachin L, Nannmark U, Nyström T. 2005. Differential roles of theuniversal stress proteins of Escherichia coli in oxidative stress resistance,adhesion, and motility. J Bacteriol 187:6265–6272.124. Xiong J, Alexander DC, Ma JH, Déraspe M, Low DE, Jamieson FB,Roy PH. 2013. Complete sequence of pOZ176, a 500-kilobase IncP-2 plas-mid encoding IMP-9-mediated carbapenem resistance, from outbreak isolatePseudomonas aeruginosa 96.Antimicrob Agents Chemother 57:3775–3782.125. Kieboom J, Dennis JJ, de Bont JA, Zylstra GJ. 1998. Identificationand molecular characterization of an efflux pump involved in Pseudo-monas putida S12 solvent tolerance. J Biol Chem 273:85–91.126. Lacal J, Reyes-Darias JA, García-Fontana C, Ramos JL, Krell T.2013. Tactic responses to pollutants and their potential to increase bio-degradation efficiency. J Appl Microbiol 114:923–933.127. Kuc J. 1995. Phytoalexins, stress metabolism, and disease resistancein plants. Annu Rev Phytopathol 33:275–297.128. Spaink HP. 1995. The molecular basis of infection and nodulation byrhizobia: the ins and outs of sympathogenesis. Annu Rev Phytopathol33:345–368.
20 ASMscience.org/MicrobiolSpectrum
Segura et al.

129. Rao JR, Cooper JE. 1994. Rhizobia catabolize nod gene-inducingflavonoids via C-ring fission mechanisms. J Bacteriol 176:5409–5413.130. González-Pasayo R, Martínez-Romero E. 2000. Multiresistancegenes of Rhizobium etli CFN42.Mol Plant Microbe Interact 13:572–577.131. GonzalezV, SantamariaRI, Bustos P,Hernandez-Gonzalez I,Medrano-Soto A, Moreno-Hagelsieb G, Janga, SC, Ramirez MA, Jimenez-Jacinto V,Collado-Vides J, Davila G. 2006. The partitioned Rhizobium etli genome:genetic and metabolic redundancy in seven interacting replicons. Proc NatlAcad Sci USA 103:3834–3839.132. Mann MS, Dragovic Z, Schirrmacher G, Lütke-Eversloh T. 2012.Over-expression of stress protein-encoding genes helps Clostridiumacetobutylicum to rapidly adapt to butanol stress. Biotechnol Lett 34:1643–1649.133. Streit WR, Schmitz RA, Perret X, Staehelin C, Deakin WJ, Raasch C,Liesegang H, Broughton WJ. 2004. An evolutionary hot spot: thepNGR234b replicon of Rhizobium sp. strain NGR234. J Bacteriol 186:535–542.134. Lim JS, Choi BS, Choi AY, Kim KD, Kim DI, Choi IY, Ka JO. 2012.Complete genome sequence of the fenitrothion-degrading Burkholderiasp. strain YI23. J Bacteriol 194:896. doi: 10.1128/JB.06479-11.135. Janssen PJ, Van Houdt R, Moors H, Monsieurs P, Morin N,Michaux A, Benotmane MA, Leys N, Vallaeys T. Lapidus A, Monchy S,Médigue C, Taghavi S, McCorkle S, Dunn J, van der Lelie D, MergeayM.2010. The complete genome sequence of Cupriavidus metallidurans strainCH34, a master survivalist in harsh and anthropogenic environments.PLoS One 5:e10433. doi: 10.1371/journal.pone.0010433.136. Ma Z, Jacobsen FE, Giedroc DP. 2009. Metal transporters and metalsensors: How coordination chemistry controls bacterial metal homeosta-sis. Chem Rev 13:4644–4681.137. Adriano DC. 2001. Trace Elements in Terrestrial Environments:Biogeochemistry, Bioavailability, and Risks of Metals. Springer-Verlag,New York, NY.138. Mergeay M, Nies D, Schlegel HG, Gerits J, Charles P, Van GijsegemF. 1985. Alcaligenes eutrophus CH34 is a facultative chemolithotrophwith plasmid-bound resistance to heavy metals. J Bacteriol 162:328–334.139. Monchy S, Benotmane MA, Janssen P, Vallaeys T, Taghavi S, vander Lelie D, Mergeay M. 2007. Plasmids pMOL28 and pMOL30 ofCupriavidus metallidurans are specialized in the maximal viable responseto heavy metals. J Bacteriol 189:7417–7425.140. Nies DH. 2003. Efflux-mediated heavy metal resistance in pro-karyotes. FEMS Microbiol Rev 27:313–339.141. Paulsen IT, Saier MH, Jr. 1997. A novel family of ubiquitous heavymetal ion transport proteins. J Membr Biol 156:99–103.142. Munkelt D, Grass G, Nies DH. 2004. The chromosomally encodedcation diffusion facilitator proteins DmeF and FieF from WautersiametalliduransCH34 are transporters of broadmetal specificity. J Bacteriol186:8036–8043.143. Ramírez-Díaz MI, Díaz-Pérez C, Vargas E, Riveros-Rosas H,Campos-García J, Cervantes C. 2008. Mechanisms of bacterial resistanceto chromium compounds. Biometals 21:321–332.
144. Henne KL, Nakatsu CH, Thompson DK, Konopka AE. 2009. High-level chromate resistance inArthrobacter sp. strain FB24 requires previouslyuncharacterized accessory genes. BMC Microbiol 16:199. doi: 10.1186/1471-2180-9-199.
145. Chen YF, Chao H, Zhou NY. 2014. The catabolism of 2,4-xylenoland p-cresol share the enzymes for the oxidation of para-methyl groupin Pseudomonas putida NCIMB 9866. Appl Microbiol Biotechnol 98:1349–1356.
146. Misra TK. 1992. Bacterial resistances to inorganic mercury salts andorganomercurials. Plasmid 27:4–16.
147. Diels L, Faelen M, Mergeay M, Nies D. 1985. Mercury transposonsfrom plasmids governing multiple resistance to heavymetals inAlcaligeneseutrophus CH34. Arch Intern Physiol Biochem 93:27–28.
148. Silver S, Phung LT. 1996. Bacterial heavy metal resistance: newsurprises. Annu Rev Microbiol 50:753–789.149. Nascimento AM, Chartone-Souza E. 2003. Operon mer: bacterialresistance to mercury and potential for bioremediation of contaminatedenvironments. Genet Mol Res 2:92–101.150. Boon N, Goris J, De Vos P, Verstraete W, Top EM. 2001. Geneticdiversity among 3-chloroaniline- and aniline-degrading strains of theComamonadaceae. Appl Environ Microbiol 67:1107–1115.151. Schwartz E, Henne A, Cramm R, Eitinger T, Friedrich B, GottschalkG. 2003. Complete nucleotide sequence of pHG1: a Rastonia eutrophaH16 megaplasmid encoding key enzymes of H(2)-based lithoautotrophyand anaerobiosis. J Mol Biol 332:369–383.152. Chen WM, Moulin L, Bontemps C, Vandamme P, Bena G,Boivin-Masson C. 2003. Legume symbiotic nitrogen fixation by beta-proteobacteria is widespread in nature. J Bacteriol 185:7266–7272.
153. Roselli S, Nadalig T, Vuilleumier S, Bringel F. 2013. The 380 KbppCMU01 plasmid encodes chloromethane utilization genes and redun-dant genes for vitamin B12- and tetrahydrofolate-dependent chloro-methane metabolism inMethylobacterium extorquens CM4: A proteomicand bioinformatics study. PLoS One 8:e56598. doi: 10.1371/journal.pone.0056598.
154. Liesegang H, Lemke K, Siddiqui RA, Schlegel HG. 1993. Charac-terization of the inducible nickel and cobalt resistance determinantcnr from pMOL28 of Alcaligenes eutrophus CH34. J Bacteriol 175:767–778.
155. Schmidt T, Schlegel HG. 1994. Combined nickel-cobalt-cadmiumresistance encoded by the ncc locus of Alcaligenes xylosoxidans 31A.J Bacteriol 176:7045–7054.
156. Silver S, Gupta A,Matsui K, Lo JF. 1999. Resistance to Ag(I) Cationsin bacteria: environments, genes and proteins. Met Based Drugs 6:315–320.
157. Monchy S, Benotmane MA, Wattiez R, van Aelst S, Auquier V,Borremans B, Mergeay M, Taghavi S, van der Lelie D, Vallaeys T. 2006.Transcriptomic and proteomic analyses of the pMOL30-encoded copperresistance in Cupriavidus metallidurans strain CH34. Microbiology152:1765–1776.
158. Mealman TD, Blackburn NJ, McEvoy MM. 2012. Metal export byCusCFBA, the periplasmic Cu(I)/Ag(I) transport system of Escherichiacoli. Curr Top Membr 69:163–196.
159. Copley SD, Rokicki J, Turner P, Daligault H, Nolan M, Land M.2012. The whole genome sequence of Sphingobium chlorophenolicum L-1: insights into the evolution of the pentachlorophenol degradationpathway. Genome Biol Evol 4:184–198.
160. Tabata M, Ohtsubo Y, Ohhata S, Tsuda M, Nagata Y. 2013.Complete genome sequence of the γ-hexachlorocyclohexane-degradingbacterium Sphingomonas sp. strain MM-1. Genome Announc 1:piie00247-13. doi: 10.1128/genomeA.00247-13.
161. Gómez-Sanz E, Kadlec K, Feßler AT, Zaragoza M, Torre, C,Schwarz S. 2013. Novel erm(T)-carrying multiresistance plasmids fromporcine and human isolates of methicillin-resistant Staphylococcus aureusST398 that also harbor cadmium and copper resistance determinants.Antimicrob Agents Chemother 57:3275–3282.
162. Bender CL, Cooksey DA. 1987. Molecular cloning of copper resis-tance genes from Pseudomonas syringae pv. tomato. J Bacteriol 169:470–474.
163. Gutiérrez-Barranquero JA, de Vicente A, Carrión VJ, Sundin GW,Cazorla FM. 2013. Recruitment and rearrangement of three differentgenetic determinants into a conjugative plasmid increase copper resistancein Pseudomonas syringae. Appl Environ Microbiol 79:1028–1033.
164. Shin SH, Kim S, Kim JY, Lee S, UmY, Oh MK, Kim YR, Lee J,Yang KS. 2012. Complete genome sequence of the 2,3-butanediol-producing Klebsiella pneumoniae strain KCTC 2242. J Bacteriol 194:2736–2737.
ASMscience.org/MicrobiolSpectrum 21
Plasmid-Mediated Tolerance Toward Environmental Pollutants

165. Diels L, Mergeay M. 1990. DNA probe-mediated detection of re-sistance bacteria from soils highly polluted by heavy metals. Appl EnvironMicrobiol 56:1485–1491.166. Slyemi D, Bonnefoy V. 2012. How prokaryotes deal with arsenic.Environ Microbiol Reports 4:571–586.167. Dhuldhaj UP, Yadav IC, Singh S, Sharma NK. 2013. Microbialinteractions in the arsenic cycle: adoptive strategies and applications inenvironmental management. Rev Environ Cont Toxicol 224:1–38.168. Volland S, Rachinger M, Strittmatter A, Daniel R, Gottschalk G,Meyer O. 2011. Complete genome sequences of the chemolithoauto-trophicOligotropha carboxidovorans strains OM4 and OM5. J Bacteriol193:5043. doi: 10.1128/JB.05619-11.169. Vogel TM. 1996. Bioaugmentation as a soil bioremediation ap-proach. Curr Opin Biotechnol 7:311–316.170. Top EM, Springael D, Boon N. 2002. Catabolic mobile geneticelements and their potential use in bioaugmentation of polluted soils andwaters. FEMS Microbiol Ecol 42:199–208.171. Ikuma K, Gunsch CK. 2012. Genetic bioaugmentation as an effectivemethod for in situ bioremediation. Functionality of catabolic plasmidsfollowing conjugal transfers. Bioengineered 3:236–241.172. Jussila MM, Zhao J, Souminen L, Lindström K. 2007. TOL plasmidtransfer during bacterial conjugation in vitro and rhizoremediation of oilcompounds in vivo. Environ Pollut 146:510–524.173. Ramos-Gonzalez MI, Duque E, Ramos JL. 1991. Conjugationaltransfer of recombinant DNA in cultures and in soils: host range of Pseu-domonasputida TOL plasmids. Appl Environ Microbiol 57:3020–3027.174. Molbak L, Licht TR, Kvist T, Kroer N, Andersen SR. 2003. Plasmidtransfer from Pseudomonas putida to the indigenous bacteria on alfalfasprouts: characterization, direct quantification and in situ location oftransconjugant cells. Appl Environ Microbiol 69:5536–5542.175. Ikuma K, Holzem RM, Gunsch CK. 2012. Impacts of organic carbonavailability and recipient bacteria characteristics on the potential for TOLplasmid genetic bioaugmentationon soil slurries.Chemosphere89:158–163.176. Ikuma K, Gunsch CK. 2013. Functionality of the TOL plasmid undervarying environmental conditions following conjugal transfer. ApplMicrobiol Biotechnol 97:395–408.177. Pinedo CA, Smets BF. 2005. Conjugal TOL transfer from Pseudo-monas putida to Pseudomonas aeruginosa: effects of restriction proficien-cy, toxicant exposure, cell density ratios, and conjugation detectionmethodon observed transfer efficiencies. Appl Environ Microbiol 71:51–57.178. Daane LL, Molina J, Sadowsky MJ. 1997. Plasmid transfer betweenspatially separated donor and recipient bacteria en earthworm-containingsoil microcosms. Appl Environ Microbiol 63:679–686.179. Wuertz S, Okabe S, Hausner M. 2004. Microbial communities andtheir interactions in biofilm systems: an overview. Water Sci Technol49:327–336.180. Mohan SV, Falkentoft C, Nancharaiah YV, McSwain Sturm BS,Wattiau P, Wilderer PA,Wuertz S, HausnerM. 2009. Bioaugmentation ofmicrobial communities in laboratory and pilot scale sequencing batchbiofilm reactors using the TOL plasmid. Bioresour Technol 100:1746–1753.181. Neilson JW, Josephson KL, Pepper IL, Arnold RB, Di Giovanni GD,Sinclair NA. 1994. Frequency of horizontal gene transfer of a large cat-abolic plasmid (pJP4) in soil. Appl Environ Microbiol 60:4053–4058.182. Molina L, Ramos C, Duque E, Ronchel MC, Garcöla JM, Wyke L,Ramos JL. 2000. Survival of Pseudomonas putida KT2440 in soil and inthe rhizosphere of plants under greenhouse and environmental conditions.Soil Biol Biochem 32:315–321.183. Mackova M, Dowling D, Macek T (ed). 2006. Phytoremediationand Rhizoremediation. Theoretical Background. Series: Focus on Bio-technology. Springer, New York, NY.184. Walker TS, Bais HP, Grotewold E, Vivanco JM. 2003. Root exu-dation and rhizosphere biology. Plant Physiol 132:44–51.
185. Matilla MA, Espinosa-Urgel M, Rodriguez-Hervá JJ, Ramos JL,Ramos-González MI. 2007. Genomic analysis reveals the major drivingforces of bacterial life in the rhizosphere.GenomeBiol8:R179. doi:10.1186/gb-2007-8-9-r179.186. López-Guerrero MG, Ormeño-Orrillo E, Acosta JL, Mendoza-Vargas A, Rogel MA, Ramírez MA, Rosenblueth M, Martínez-Romero J, Martínez-Romero E. 2012. Rhizobial extrachromosomalreplicon variability, stability and expression in natural niches. Plasmid 68:149–158.187. Barac T, Taghavi S, Borremans B, Provoost A, Oeyen L, ColpaertJV, Vangronsveld J, van der Lelie D. 2004. Engineered endophytic bac-teria improve phytoremediation of water-soluble, volatile, organicpollutants. Nat Biotech 22:583–588.188. Taghavi S, Barac T, Greenberg B, Borremans B, Vangronsveld J, vander Lelie D. 2005. Horizontal gene transfer to endogenous endophyticbacteria from poplar improves phytoremediation of toluene. Appl Envi-ron Microbiol 71:8500–8505.189. Lilley AK, Bailey MJ. 1997. Impact of plasmid pQBR103 acquisitionand carriage on the phytosphere fitness of Pseudomonas fluorescensSBW25: burden and benefit. Appl Environ Microbiol 63:1584–1587.
190. Tett A, Spiers AJ, Crossman LC, Ager D, Ciric L, Dow, JM, Fry JC,Harris D, Lilley A, Oliver A, Parkhill J, Quail MA, Rainey PB, SaundersNJ, Seeger K, Snyder LA, Squares R, Thomas CM, Turner SL, Zhang XX,Field D, Bailey MJ. 2007. Sequence-based analysis of pQBR103; a rep-resentative of a unique, transfer-proficient mega plasmid resident in themicrobial community of sugar beet. ISME J 1:331–340.
191. Bale MJ, Fry JC, Day MJ. 1987. Plasmid transfer between strains ofPseudomonas aeruginosa on membrane filters attached to river stones.J Gen Microbiol 133:3099–3107.
192. Top EM, Holben WE, Forney LJ. 1995. Characterization of diverse2,4-dichlorophenoxyacetic acid-degradative plasmids isolated from soilby complementation. Appl Environ Microbiol 61:1691–1698.
193. Lilley AK, Bailey MJ. 1997. The acquisition of indigenous plasmidsby a generically marked pseudomonad population colonizing the sugarbeet phytosphere is related to local environmental conditions. Appl En-viron Microbiol 63:1577–1583.
194. Dahlberg C, Linberg C, Torsvik VL, Hermansson M. 1997. Con-jugative plasmids isolated from bacteria in marine environments showvarious degrees of homology to each other and are not closely related towell characterized plasmids. Appl Environ Microbiol 63:4692–4697.
195. Ono A, Miyazaki R, Sota M, Ohtsubo Y, Nagata Y, Tsuda M.2007. Isolation and characterization of naphthalene-catabolic genes andplasmids from oil-contaminated soil by using two cultivation-independentapproaches. Appl Microbiol Biotechnol 74:501–510.
196. Kav AB, Sasson G, Jami E, Doron-Faigenboim A, Benhar I,Mizrahi I. 2012. Insights into the bovine rumen plasmidome. Proc NatlAcad Sci USA 109:5452–5457.
197. Nojiri H. 2013. Impact of catabolic plasmids on host cell physiology.Curr Opin Biotech 24:423–430.
198. Diaz-Ricci JC, Hernández ME. 2000. Plasmid effects on Escherichiacoli metabolism. Crit Rev Biotechnol 20:79–108.
199. Miyakoshi M, Shitani M, Inoue K, Terabayashi T, Sai F, OhkumaM, Nojiri H, Nagata Y. Tsuda M. 2012. ParI, an orphan ParA familyprotein from Pseudomonas putida KT2440-specific genomic island,interferes with the partition system of IncP-7 plasmids. Environ Microbiol14:2946–2959.
200. Shintani M, Takahashi Y, Tokumaru H, Kadota K, Hara H,Miyakoshi M, Naito K, Yamane H, Nishida H, Nojiri H. 2010. Res-ponse of the Pseudomonas host chromosomal transcriptome to carriageof the IncP-7 plasmid pCAR1. Environ Microbiol 12:1413–1426.
201. Shintani M, Tokumaru H, Takahasi Y, Miyakoshi M, Yamane H,Nishida H, Nojiri H. 2011. Alterations of RNA maps of IncP-7 plasmidpCAR1 in various Pseudomonas bacteria. Plasmid 66:85–92.
22 ASMscience.org/MicrobiolSpectrum
Segura et al.

202. Fernández M, Niqui-Arroyo JL, Conde S, Ramos JL, Duque E.2012. Enhanced tolerance to naphthalene and enhanced rhizoremediationperformance for Pseudomonas putida KT2440 via the NAH7 catabolicplasmid. Appl Environ Microbiol 78:5104–5110.
203. Yun CS, Suzuki C, Naito K, Takeda T, Takahashi Y, Sai F,Terabayashi T, Miyakoshi M, Shintani M, Nishida H, Yamane H, NojiriH. 2010. Pmr, a histone-like protein H1 (H-NS) family protein encoded bythe IncP-7 plasmid pCAR1, is a key global regulator that alters hostfunction. J Bacteriol 192:4720–4731.
204. Rescalli E, Saini S, Bartocci C, Rychlewski L, de Lorenzo V, BertoniG. 2004. Novel physiological modulation of the Pu promoter of TOLplasmid: negative regulatory role of the TurA protein of Pseudomonasputida in the response to suboptimal growth temperatures. J Biol Chem279:7777–7784.
205. Bathe S, Mohan TV, Wuertz S, Hausner M. 2004. Bioaugmentationof a sequencing batch biofilm reactor by horizontal gene transfer. WaterSci Technol 49:337–344.206. De Gelder L, Vandecasteele FP, Brown CJ, Forney LJ, Top EM.2005. Plasmid donor affects host range of promiscuous IncP-1 betaplasmid pB10 in an activated-sludge microbial community. Appl EnvironMicrobiol 71:5309–5317.207. Smalla K, Sobecky PA. 2002. The prevalence and diversity of mobilegenetic elements in bacterial communities of different environmentalhabitats: insights gained from different methodological approaches. FEMSMicrobiol Ecol 42:165–175.208. Ma H, Katzenmeyer KN, Bryers JD. 2013. Non-invasive in situmonitoring and quantification of TOL plasmid segregational loss withinPseudomonas putida biofilms. Biotech Bioeng 110:2949–2958.
ASMscience.org/MicrobiolSpectrum 23
Plasmid-Mediated Tolerance Toward Environmental Pollutants

Author Queries
Q1: Please verify that reference 27 is correct here for the citation of Molina et al, 2011.Q2: Please provide other authors and their initials for these unpublished results.
Related Documents











