Abstract The study of plant functional types (PFTs) has been widely emphasized when anal- ysing plant community changes in relation to variations in climate and disturbance regime. In this study, we search for PFTs of woody species near forestgrassland boundaries in South Brazil where, due to climate, forests tend to expand over grassland but are being restricted by fre- quent res. We aimed at answering the ques- tions: (i) which plant functional types of forest woody species can establish in adjacent grassland subject to re disturbance and (ii) which plant functional types of forest and grassland woody species are related to short-term community dynamics in frequently burned grassland. Traits were assessed in woody plants in 156 plots (6.75 m 2 ) arranged in 12 transects across forest grassland boundaries with different re history in their grassland part. The analysis used a recur- sive algorithm to search for traits and PFTs maximally associated to spatial distance from forest limit in one analysis, and elapsed time since last re in another. As a result, nine PFTs of forest woody species were identied that best described community patterns associated to dis- tance from forest. Resprouting ability charac- terized forest plants able to colonize grasslands. PFT diversity was higher in border plots than inside forest or grassland. Four PFTs of forest and grassland woody species best described woody species community patterns in the grass- land associated to elapsed time since re. Taller individuals of single-stemmed shrubs predomi- nated in late post-re recovery (34 years), while shorter multi-stemmed shrubs in recently burned areas (3 months to 1 year). PFTs of forest trees occurred in border plots or, as established adults, in grassland, remaining unaffected by re. We conclude that easily measurable structural plant traits, such as those used in our study, are suf- cient to evaluate post-re community dynamics. Forest PFTs in burned grassland are restricted to those with resprouting ability to survive recur- rent re events. Establishment success is highest on protected sites with lesser or low-intensity re. Keywords Forest expansion˘ Resprouter ˘ Shrubs ˘ South Brazil ˘ Tree establishment ˘ Vegetation dynamics S. C. Mu¤ller (& ) ˘V. D. Pillar Laborato · rio de Ecologia Quantitativa, Departamento de Ecologia, Universidade Federal do Rio Grande do Sul, Bento Gonc ‚alves, 9500, 91540-000 Porto Alegre, Brazil e-mail: [email protected] G. E. Overbeck ˘ J. Pfadenhauer Vegetation Ecology, Technische Universita¤t Mu¤nchen, Freising-Weihenstephan, Germany Plant Ecol (2007) 189:114 DOI 10.1007/s11258-006-9162-z 123 ORIGINAL PAPER Plant functional types of woody species related to re disturbance in forest—grassland ecotones Sandra C. Mu ‹ller ˘ Gerhard E. Overbeck ˘ Jo ‹rg Pfadenhauer ˘ Vale«rio D. Pillar Received: 1 September 2005 / Accepted: 8 May 2006 / Published online: 9 June 2006 Springer Science+Business Media B.V. 2006

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Abstract The study of plant functional types(PFTs) has been widely emphasized when anal-ysing plant community changes in relation tovariations in climate and disturbance regime. Inthis study, we search for PFTs of woody speciesnear forest–grassland boundaries in South Brazilwhere, due to climate, forests tend to expandover grassland but are being restricted by fre-quent �res. We aimed at answering the ques-tions: (i) which plant functional types of forestwoody species can establish in adjacent grasslandsubject to �re disturbance and (ii) which plantfunctional types of forest and grassland woodyspecies are related to short-term communitydynamics in frequently burned grassland. Traitswere assessed in woody plants in 156 plots(6.75 m2) arranged in 12 transects across forest–grassland boundaries with different �re history intheir grassland part. The analysis used a recur-sive algorithm to search for traits and PFTsmaximally associated to spatial distance from
forest limit in one analysis, and elapsed timesince last �re in another. As a result, nine PFTsof forest woody species were identi�ed that bestdescribed community patterns associated to dis-tance from forest. Resprouting ability charac-terized forest plants able to colonize grasslands.PFT diversity was higher in border plots thaninside forest or grassland. Four PFTs of forestand grassland woody species best describedwoody species community patterns in the grass-land associated to elapsed time since �re. Tallerindividuals of single-stemmed shrubs predomi-nated in late post-�re recovery (3–4 years), whileshorter multi-stemmed shrubs in recently burnedareas (3 months to 1 year). PFTs of forest treesoccurred in border plots or, as established adults,in grassland, remaining unaffected by �re. Weconclude that easily measurable structural planttraits, such as those used in our study, are suf-�cient to evaluate post-�re community dynamics.Forest PFTs in burned grassland are restricted tothose with resprouting ability to survive recur-rent �re events. Establishment success is higheston protected sites with lesser or low-intensity�re.
Keywords Forest expansionÆResprouter ÆShrubs ÆSouth Brazil ÆTree establishment ÆVegetation dynamics
S. C. Muller ( & ) ÆV. D. PillarLaboratorio de Ecologia Quantitativa, Departamentode Ecologia, Universidade Federal do Rio Grande doSul, Bento Goncalves, 9500, 91540-000 Porto Alegre,Brazile-mail: [email protected]
G. E. Overbeck ÆJ. PfadenhauerVegetation Ecology, Technische UniversitatMunchen, Freising-Weihenstephan, Germany
Plant Ecol (2007) 189:1–14DOI 10.1007/s11258-006-9162-z
123
ORIGINAL PAPER
Plant functional types of woody species related to Þredisturbance in forestÐgrassland ecotones
Sandra C. Mu¬ller ÆGerhard E. Overbeck ÆJo¬rg Pfadenhauer ÆVale«rio D. Pillar
Received: 1 September 2005 / Accepted: 8 May 2006 / Published online: 9 June 2006� Springer Science+Business Media B.V. 2006

Introduction
Assessing plant functional types has been widelyemphasized as an alternative to determiningspecies composition for the evaluation of thein�uence of climatic changes, differences in theatmospheric composition (e.g. rising CO2 con-centration) and land-use changes over present-day plant communities around the world (Box1996; Woodward and Cramer 1996; D�´az andCabido 1997; Pausas 1999).
In spite of some conceptual differences, it isagreed in most de�nitions that plant functionaltypes (PFTs) are groups of plants similar in termsof traits and similar in their responses to certainenvironmental conditions (e.g. soil conditions,temperature, moisture, disturbance regimes) and/or in their effects on ecosystem processes (e.g.biomass production, litter decomposition) (Pillarand Orloci 1993a, b; Box 1996; Noble and Gitay1996; Woodward and Cramer 1996; Lavorel andGarnier 2002; Pillar and Sosinski 2003). Based onthe assumption of a relationship between formand function (Barkman 1988), the structural–functional approach permits the use of visiblestructural attributes as substitutes for functionalpatterns. Thus, these have been widely used asbasic features for PFT classi�cations and, whenpossible, further complemented by physiologicaltraits (Box 1996; Weiher et al. 1999).
The many methods to elaborate PFT classi�-cations follow different approaches. Groups ofplants may be derived by numerical analysis, witha selection of traits related to environmental (ordisturbance) variables, or may be deduceda pri-
ori based on features that are assumed to have acertain functionality in the system (Bugmann1996; Noble and Gitay 1996; Woodward andCramer 1996). Even though an appealing thought,it has been acknowledged that a universal func-tional classi�cation of plants is unlikely to beaccomplished. In contrast, classi�cations becomemore useful when plant types are determined fora particular purpose. For example, phenologicaldata and life-cycle information are useful to studylandscape change in relation to disturbance(Noble and Gitay 1996; Campbell et al. 1999), orlife forms can be applied to study environmentalgradients in contrasting �oras of regions differing
greatly in biogeography (Pillar and Orlo ci 1993b;D� az et al. 2004).
In ecotones of grassland–forest formations insouthern Brazil, studies have focused on thecauses of their continued existence, since presentclimate conditions are suitable for the develop-ment of forest formations (Rambo 1956; Pillarand Quadros 1997) and no restrictions by soilproperties for this forest expansion have beenobserved (e.g. Garcia Martinez 2005). The pro-cess of forest expansion that started after midHolocene has been well documented by palyno-logical studies in peat pro�les in southern Brazil(e.g. Behling 2001; Behling et al. 2004). In thesestudies, charcoal particles from much earlier inthe Holocene have also been recorded. Fire thushas been present in the region for a long time as afactor preventing successful forest tree speciesestablishment within grassland (Hoffmann et al.2003; Pillar 2003). In spite of that, many (mostlyzoochoric) forest tree species can be observedcolonizing grassland areas in forest–grassland–mosaics, close to the border or forming ratherdense patches of woody species in the grasslandmatrix (Pillar 2003; Mu ller and Forneck 2004;Duarte et al. in press). The latter is often relatedto the presence of boulders or rock outcrops,which locally prevent high intensity �res and al-low the survival of established woody plants(Pillar 2003; Muller and Forneck 2004). Addi-tionally, shrub encroachment occurs in thesegrasslands, mainly by non-forest, anemochoricspecies.
In this paper, we are interested in identifyingPFTs that synthesize the general features andspatial pattern of established woody species (bothforest trees and grassland shrubs) in forest–grassland–mosaics, thus being able to discussprocesses independent of species identity. Wewish to clarify the in�uence of �re and distancefrom the forest–grassland border on PFT com-position. To de�ne PFT, structural traits wereselected and sampled in the �eld on individualplants. This allowed us to explore variabilitywithin species (Cornelissen et al. 2003), that mayresult from genetic variation, environmentalconditions, and/or disturbance restrictions. Usingthis approach we de�ne a classi�cation underwhich individuals of a single species may belong
2 Plant Ecol (2007) 189:1–14
123

to different PFTs (Pillar and Sosinski 2003;Sosinski and Pillar 2004).
This study has two speci�c objectives: First, weanalyse only distribution and characteristics offorest woody plants and, presuming a forestexpansion process into grassland, we search forPFTs related to spatial distance from the forestborder both into forest and grassland. Second, weanalyse only the grassland portion in the ecotonesituation, searching for PFTs characterizing short-term responses to �re disturbance, for which weconsidered all woody plants (forest and grasslandspecies) present in grassland.
Methods
Study area
The study was carried out on Morro Santana(Santana hill; 30� 03¢S, 51� 07¢W) part of a chainof granitic hills in Porto Alegre, Rio Grande doSul, Brazil. Natural vegetation cover in the regionis a mosaic of grassland and forest, the latterpreferentially on sites with southern and south-eastern aspect, with many patches of woodyplants in the grassland matrix and riparian forestalong creeks or rivers (Rambo 1956; Teixeiraet al. 1986). Grassland predominates on the top ofthe hill (altitude max. 311 m a.s.l.) and on thenorthern slopes, comprising approx. 220 ha. Fireis a frequent disturbance in the grassland, usuallycaused by residents in the neighbourhood, withreturn intervals of 3–5 years. The present mosaicof vegetation is maintained by �re, since the areais not grazed and climate conditions are suitableto forest formation (Overbeck et al. 2005). Forestis too moist to burn and thus grassland �res stopat the border, causing abrupt and rather stabletransitions.
The climate is subtropical humid withoutdrought season, corresponding to the Cfa’s typeaccording to Koppen’s classi�cation, with meanannual precipitation of 1348 mm and averageannual temperature of 19.5� C (Nimer 1990). Thesoil types are typical dystrophic red-yellow argi-sols, developed from granite, corresponding toacrisols, alisols and umbrisols in the FAO-classi-
�cation (Garcia Martinez 2005). Boulders androck outcrops are abundant on the hill.
Experimental setting and data collection
Data of woody plants were collected in 1.5 by1.5 m plots arranged in 12 transects located ontransitional zones of forest–grassland vegetation.Transect location was de�ned based on differ-ences in the woody vegetation physiognomy ofthe grassland matrix. Transects were arranged inpairs and each transect in the pair, as much aspossible similar to one another, separated by �vemeters. Two of these pairs were on sites withnorthern aspect, two on sites on the top andsouthern aspect, and two on the top and south-western aspect. Each transect consisted of 39contiguous plots, with 21 being in grassland and18 in forest. The limit between grassland andforest was de�ned by the last adult forest tree.
In October 2002, before sampling, we sub-jected the grassland portion of one randomly se-lected transect in each pair to a controlled burn.Vegetation in the transects located on top/south-western position had last burned in January 2002and by the time of the experimental burn had notaccumulated suf�cient �ammable biomass tosupport spreading of a �re. Therefore, theexperimental burn was successful only in fourtransects located in grassland that had been un-burned for 3 years or more (two on sites withnorthern and two on sites on the top and withsouthern aspect). The survey of the grasslandplots was conducted in January/February 2003.Such pattern of burned and unburned transectsallowed us to de�ne three groups of plots, basedon time elapsed since last grassland �re event:group 1, 3 months (plots of the transects experi-mentally burned in October 2002); group 2,1 year (plots of the transect pairs last burned inJanuary 2002); group 3, 3–4 years (unburned plotsin the transect pairs subject to experimentalburns). The �rst three grassland plots (closest tothe forest) in each transect were taken as forminga distinct group of border plots (group 4) for theanalysis involving the in�uence of �re, as they didnot burn in any transect due to lack of �ammablebiomass. In the interpretation of results we usedspace for time-substitution by taking plots from
Plant Ecol (2007) 189:1–14 3
123

groups 1 to 4 as a chronosequence. We assumedthat transects in a pair had similar plant compo-sition before the experimental burn; differencesfound between groups 1 and 3 should thus be dueto in�uence of the single experimental �re event.The survey of the forest plots was carried outfrom July to September 2003.
For the surveys every woody plant above10 cm high was sampled in all plots. We recordedspecies identity, plant height, stem shaft height(height to the �rst branch) and basal stem diam-eter. Other traits were recorded at individual,population or species level (Table 1). Habit, dis-persal syndrome, resprouting ability and leaftexture were assigned at the species level, basedon �eld observations and literature. Resproutingwas considered as the ability to regenerate after a�re by basal sprouting (Bellingham and Sparrow2000; Bond and Midgley 2001). Therefore, specieswith a general sprouting capacity that occurredexclusively inside forest areas, where �re does notspread in the study site, were not considered asresprouters. All other traits were measured di-rectly in the �eld on individual plants andextrapolated for a population. For this study wede�ne ‘‘population’’ as a group of individuals ofthe same species occurring in the same plot and
being acceptably homogenous for the traits beingconsidered (Table 1). Later, in data handling,individuals belonging to the same species andsharing the same states forplant phase and stem
base in a plot were grouped as a different popu-lation. The other traits were then averaged ineach of these populations and categorized intoclasses of states. For example, the state forplant
height in a population was the trait mean value ofindividuals in the same plant phase and stem base
state belonging to the same species in a plot. Theintent with this choice was to avoid excessive traitvariability within a population, e.g. among seed-lings and adult trees (Cornelissen et al. 2003).
Data analysis
For the analysis we used data de�ned for larger(4.5 · 1.5 m) plots generated by pooling sets ofthree adjacent 1.5· 1.5 m plots, resulting in 13plots per transect. To identify PFTs we followedthe method for trait-based community analysisdescribed in Pillar and Sosinski (2003), imple-mented in the SYNCSA software (Pillar 2004).The method allowed us to identify, in two inde-pendent analyses, PFTs maximally associated tothe forest–grassland transition (position of plots
Nr Attribute(Trait)
Code States
1 habit ha 1: shrub, 2: tree1 Habit * ha 1: shrub, 2: treelets, 3: low-size tree, 4: mid-size trees, 5: large trees2 dispersal syndrome ds 1: autochory, 2: anemochory, 3: zoochory3 resprouting ability ra 1: non-sprouter, 2: resprouter4 plant phase pp 1: below 30 cm high, 2: 30–79, 3: >80 cm high, but less than 5 cm of basal diameter,
4: above state 35 plant height ph 1: < 0.2 m; 2: 0.2–0.5; 3: 0.5–1.5; 4: 1.5–3; 5: 3–6; 6: 6–9; 7: >9 m6 shaft height sh 0: unbranched; 1: < 0.2 m; 2: 0.2–0.5; 3: 0.5–1.5; 4: 1.5–3; 5: 3–6; 6: 6–9; 7: >9 m7 stem base sb 1: single-stemmed, 2: multi-stemmed8 stem consistency sc 1: semi-woody, 2: woody9 stem structure se 1: unbranched stem, 2: branched stem
10 leaf shape ls class of width leaf:length leaf ratio—0: no leaf; 1: < 0.1; 2: 0.1–0.3; 3: 0.3–0.6; 4: 0.6–0.8;5: 0.8–1; 6: >1
11 leaf area la 0: no leaf; 1: < 1 cm2; 2: 1–5; 3: 5–15; 4: 15–25; 5: 25–35; 6: 35–50; 7: 50–80; 8: >80 cm2
12 leaf texture lt 0: no leaf, 1: coriaceous, 2: intermediate between 1 and 3, 3: membranous
*Groups according to Brack et al. 1998 referring to a species’ maximum height (shrubs: less than 2.5 m; treelets: 2.5–5 m,low-size trees: 5–9 m, mid-size trees: 9–15 m, large trees: above 15 m)
Table 1 Traits used for description of woody plants ingrassland, border and forest plots on Morro Santana, RS,Brazil. Alternative states for habit (*) were used for treesand understory shrubs in forest–grassland transition
analysis. For the analysis, all traits were considered to bequantitative or semi-quantitative according to stateslogically ordered in the table
4 Plant Ecol (2007) 189:1–14
123

on the gradient) and PFTs maximally associatedto the effect of a recent �re disturbance. In short,the algorithm described by Pillar and Sosinski(2003) involves three data matrices: one matrix(B) of plant populations described by the states oftraits, a second matrix (W) of populations de-scribed by their performance (e.g. density orcover) in the communities (our pooled plots), anda third matrix ( E) of community sites describedby environmental variable(s) (in our case, dis-tance from forest limit or time since last �re). Therecursive algorithm searches, in matrix B, for asubset of traits and groups of populations basedon cluster analysis with these traits (PFTs) in sucha way to maximize the congruence between veg-etation described by the PFTs so de�ned and thechosen environmental variable. Congruence ismeasured by the matrix correlation q (D, D),where D contains community dissimilarities(chord distances) based on the PFT performances(population performances in matrix W rear-ranged and pooled in each plot according to thede�ned PFTs), and D the community sites’ dis-similarities (Euclidean distances) based on thechosen environmental variables (E).
For the analysis we categorized the woodyspecies into typical grassland species (shrubs,never present in forest) and forest species (trees,typical of forest, but sometimes present in grass-land as pioneers). The following analyses wereconducted:
(1) With the objective of identifying PFTsassociated to a presumed forest expansionprocess into grassland (forest–grasslandtransition analysis) we considered only for-est species, excluding plots where no forestspecies were found, and used, in matrixE,the distance to the forest limit as environ-mental variable. In this analysis matrix Wwas de�ned by population density per plot.
(2) With the objective of identifying PFTsassociated to �re disturbance in grassland,we considered all woody species present ingrassland plots and used, as a proxy for post-�re recovery stage, time elapsed since thelast �re event as the environmental variablein matrix E (previously mentioned groups ofplots ordered 1–4). Matrix W was de�ned by
population basal area per plot in the �rstsurvey.
Each analysis resulted in the identi�cation ofan optimal trait subset and PFTs de�ned by thesetraits, as well as theq (D, D) congruence valuesindicating the correlation between the de�nedPFTs and the environmental variable used. For asynthetic view of results we used ordination byPrincipal Coordinates Analysis (PCoA) of plots,using the matrix of plots by de�ned PFTs, basedon chord distances (Podani 2000). In order tocomplement the interpretation of the PFTsaccording to the traits that maximized congru-ence, a PCoA ordination analysis of PFTs basedon the mean value of states in the optimal traitsubset was performed using Gower’s index asresemblance measure. Additionally, all traits(optimal and suboptimal) were depicted on thediagram accordingly to their correlation valuewith the two �rst ordination scores (Pillar andSosinski 2003). Groups of plots based on timeelapsed since last �re were compared for PFTcomposition and PFT diversity, by analysis ofvariance with randomization testing, using 1000random permutations (Pillar and Orlo ci 1996).Diversity was measured by the Shannon indexbased on the natural logarithm (nats).
Results
PFTs of forest species in forest–grasslandtransition
This trait-based analysis involved 1344 popula-tions, belonging to 80 species of 40 families.Myrtaceae (14 spp.), Lauraceae (7 spp.) andRubiaceae (6 spp.) were the most species richfamilies, followed by Sapindaceae, Euphorbia-ceae and Salicaceae, each one with four species.These are forest trees, except for a few Rubiaceaeshrubs, and some species (Dodonaea viscosa (L.)Jacq., Agarista eucalyptoides (Cham. et Schl.) G.Don, Campomanesia aurea O. Berg, Opuntia
monacantha Haw. and Cereus hildmannianus
K. Schum.) that are uncommon inside the forestitself but were considered in this analysis due totheir importance in border plots. Appendix 1
Plant Ecol (2007) 189:1–14 5
123
contains the complete list of species and theirinclusion or not in both trait-based analyses.
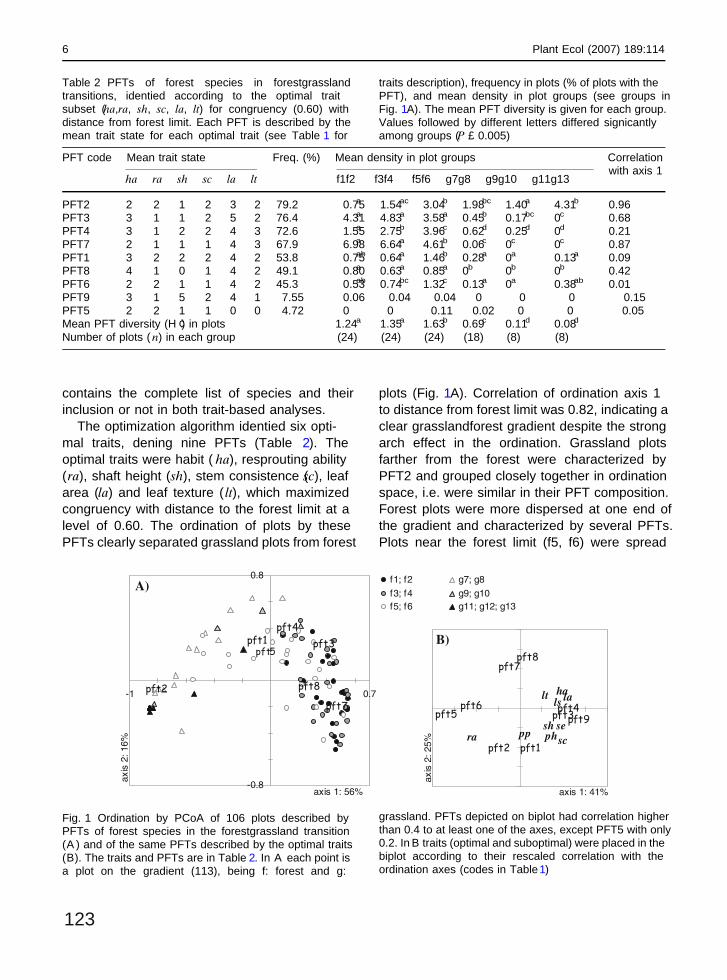
The optimization algorithm identi�ed six opti-mal traits, de�ning nine PFTs (Table 2). Theoptimal traits were habit ( ha), resprouting ability(ra), shaft height (sh), stem consistence (sc), leafarea (la) and leaf texture ( lt), which maximizedcongruency with distance to the forest limit at alevel of 0.60. The ordination of plots by thesePFTs clearly separated grassland plots from forest
plots (Fig. 1A). Correlation of ordination axis 1to distance from forest limit was 0.82, indicating aclear grassland–forest gradient despite the strongarch effect in the ordination. Grassland plotsfarther from the forest were characterized byPFT2 and grouped closely together in ordinationspace, i.e. were similar in their PFT composition.Forest plots were more dispersed at one end ofthe gradient and characterized by several PFTs.Plots near the forest limit (f5, f6) were spread
PFT code Mean trait state Freq. (%) Mean density in plot groups Correlationwith axis 1
ha ra sh sc la lt f1–f2 f3–f4 f5–f6 g7–g8 g9–g10 g11–g13
PFT2 2 2 1 2 3 2 79.2 0.75a 1.54ac 3.04b 1.98bc 1.40a 4.31b –0.96PFT3 3 1 1 2 5 2 76.4 4.31a 4.83a 3.58a 0.45b 0.17bc 0c 0.68PFT4 3 1 2 2 4 3 72.6 1.55a 2.75b 3.96c 0.62d 0.25d 0d 0.21PFT7 2 1 1 1 4 3 67.9 6.98a 6.64a 4.61b 0.06c 0c 0c 0.87PFT1 3 2 2 2 4 2 53.8 0.75ab 0.64a 1.46b 0.28a 0a 0.13a –0.09PFT8 4 1 0 1 4 2 49.1 0.80a 0.63a 0.85a 0b 0b 0b 0.42PFT6 2 2 1 1 4 2 45.3 0.53ab 0.74bc 1.32c 0.13a 0a 0.38ab 0.01PFT9 3 1 5 2 4 1 7.55 0.06 0.04 0.04 0 0 0 0.15PFT5 2 2 1 1 0 0 4.72 0 0 0.11 0.02 0 0 –0.05Mean PFT diversity (H ¢) in plots 1.24a 1.35a 1.63b 0.69c 0.11d 0.08d
Number of plots ( n) in each group (24) (24) (24) (18) (8) (8)
Table 2 PFTs of forest species in forest–grasslandtransitions, identi�ed according to the optimal traitsubset (ha,ra, sh, sc, la, lt) for congruency (0.60) withdistance from forest limit. Each PFT is described by themean trait state for each optimal trait (see Table 1 for
traits’ description), frequency in plots (% of plots with thePFT), and mean density in plot groups (see groups inFig. 1A). The mean PFT diversity is given for each group.Values followed by different letters differed signi�cantlyamong groups (P £ 0.005)
-0.8
0.8
-1 0.7
f1; f2 g7; g8
f3; f4 g9; g10f5; f6 g11; g12; g13
axis
2: 1
6%
axis 1: 56% axis 1: 41%
axis
2: 2
5%
A)
B)
Fig. 1 Ordination by PCoA of 106 plots described byPFTs of forest species in the forest–grassland transition(A ) and of the same PFTs described by the optimal traits(B). The traits and PFTs are in Table 2. In A each point isa plot on the gradient (1–13), being f: forest and g:
grassland. PFTs depicted on biplot had correlation higherthan 0.4 to at least one of the axes, except PFT5 with only0.2. In B traits (optimal and suboptimal) were placed in thebiplot according to their rescaled correlation with theordination axes (codes in Table1)
6 Plant Ecol (2007) 189:1–14
123
over the diagram, indicating higher diversity ofPFT composition and a transitional situation toborder and grassland plots. The ordination ofPFTs by the optimal traits (Fig. 1B) depicts theidenti�ed PFT and their trait similarities. The �rstordination axis denotes variation in resproutingability (from left to right, resprouter to non-sprouter), stem consistence (semi-woody to woo-dy stems), leaf area and shape (larger and broaderleaves to the right), leaf texture (from coriaceousto membranous) and habit (shrubs/treelets tomid-size trees). The second axis mainly indicatedvariation of resprouting ability, stem consistence,and plant height. Redundancy among suboptimaland optimal traits was evident in this ordinationanalysis, possibly indicating similar functionalityin chosen traits.
Resprouting ability was the trait that best de-�ned forest PFTs in grassland plots (compareboth ordination diagrams of Fig. 1 and the den-sities of PFT2 in grassland plots in Table2).PFT2, characterizing grassland plots, was formedmainly by treelets and low-size trees with res-prouting ability and leaves with less than 15 cm2
in area and intermediate texture (most specieshad coriaceous leaves, but also intermediate andmembranous leaves). PFT1 was formed mostly bylow-size trees with the ability to resprout, withlarger leaves than PFT2 (as all other PFTs aswell), while PFT4 lacked resprouting ability andhad membranous leaves. Both PFTs 1 and 4 weresigni�cantly more abundant at border plots. PFT7was comprised mostly of understory shrubs andyoung treelets/low-size trees (Appendix 2) with
mid-size membranous leaves, being more abun-dant inside the forest, while PFT8 were saplingsof mid-size trees with unbranched (sh) and semi-woody (sc) stems with large leaves of intermedi-ary texture. PFT8 had similar densities in allforest plots and was completely absent in grass-land plots. PFT5 comprised only cactus species atthe border plots. PFT diversity was signi�cantlyhigher at the border than inside forest or in thegrassland (Table2).
PFTs of forest and grassland woody speciesand time elapsed since �re in grassland
This analysis of grassland and border plots onlyinvolved 605 populations belonging to 58 speciesof 25 families. Asteraceae was the family richestin species (17 species), followed by Myrtaceae (5),Euphorbiaceae (4), Sapindaceae (3), Fabaceae(3) and Rubiaceae (3). About 24 species occurredonly at border plots. Border plots differed fromother grassland plots due to proximity of forestand, consequently, absence of direct �re in�uenceon plant composition, but had no adult forest treespecies.
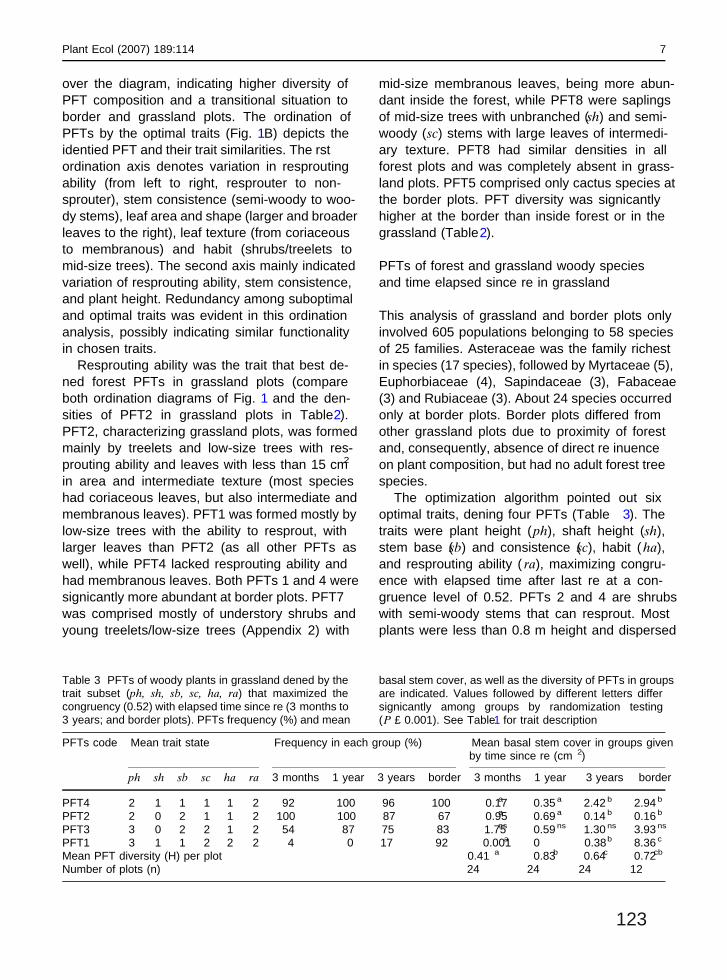
The optimization algorithm pointed out sixoptimal traits, de�ning four PFTs (Table 3). Thetraits were plant height (ph), shaft height (sh),stem base (sb) and consistence (sc), habit (ha),and resprouting ability ( ra), maximizing congru-ence with elapsed time after last �re at a con-gruence level of 0.52. PFTs 2 and 4 are shrubswith semi-woody stems that can resprout. Mostplants were less than 0.8 m height and dispersed
PFTs code Mean trait state Frequency in each group (%) Mean basal stem cover in groups givenby time since �re (cm 2)
ph sh sb sc ha ra 3 months 1 year 3 years border 3 months 1 year 3 years border
PFT4 2 1 1 1 1 2 92 100 96 100 0.17a 0.35 a 2.42 b 2.94 b
PFT2 2 0 2 1 1 2 100 100 87 67 0.95a 0.69 a 0.14 b 0.16 b
PFT3 3 0 2 2 1 2 54 87 75 83 1.75ns 0.59 ns 1.30 ns 3.93 ns
PFT1 3 1 1 2 2 2 4 0 17 92 0.001a 0 0.38b 8.36 c
Mean PFT diversity (H’) per plot 0.41 a 0.83b 0.64c 0.72cb
Number of plots (n) 24 24 24 12
Table 3 PFTs of woody plants in grassland de�ned by thetrait subset (ph, sh, sb, sc, ha, ra) that maximized thecongruency (0.52) with elapsed time since �re (3 months to3 years; and border plots). PFTs’ frequency (%) and mean
basal stem cover, as well as the diversity of PFTs in groupsare indicated. Values followed by different letters differsigni�cantly among groups by randomization testing(P £ 0.001). See Table1 for trait description
Plant Ecol (2007) 189:1–14 7
123

by wind, but these PFTs differed at the stem base.Multi-stemmed shrubs were in PFT2, character-izing recently burned plots (Table 3) while PFT4,encompassing single-stemmed shrubs with a meanbasal cover higher than PFT2, was typical oflonger unburned areas (3 years and border plots).Plots 1 year after �re had an intermediate com-position, but, considering performance of PFTs 2and 4, they resembled more recently burned plots.PFT3, with woody multi-stemmed plants (shrubsand trees, see Appendix 3) between 0.5 and 1.5 mhigh, did not present any difference in basal coverbetween plot groups. PFT1 characterized borderplots (group 4) and was mainly consisted of sin-gle-stemmed tree individuals without resproutingability, with large leaves and zoochorous dispersalsyndrome (the last two traits were not selected,though). This PFT did not occur much in burnedareas. Regarding PFT diversity (Table3), differ-ences were signi�cant (P < 0.001) between re-cently burned plots and the other groups.
Discussion
The results suggest that easily measurable traitsare suf�cient for the identi�cation of PFTs ofwoody plants in grassland related to presumedforest expansion (distance to the forest limit) andto post-�re dynamics (elapsed time since last �re).Forest species able to expand within grasslandvegetation subject to burns were almost all lim-ited to one PFT (PFT2 in analysis 1), with res-prouting ability indicating an ef�cient strategy tosurvive recurrent �re events once established onprotected sites with less-intensity �re (e.g. nearboulders).
The selected traits are substitutes for functions,in a similar manner as in other studies that haveused more physiological trait sets. Functionalrobustness of traits frequently is related to leafand stem features, biomass, maximal plant heightand seed mass (D�´az et al. 1999; Weiher et al.1999; Cornelissen et al. 2003).Habit (maximalspecies height),plant height, stem base, resprout-
ing ability, leaf area and leaf texture of woodyplants in grassland–forest ecotones were relevantfor de�ning PFTs in relation to selected envi-
ronmental variables (forest limit distance andelapsed time since �re). Delimiting traits in morehomogeneous populations according to plant
phase and stem base within the same species washelpful to reveal consistent vegetation patterns inforest–grassland ecotones and burned grassland,seeing that trait-state shifts within a species mayoccur from seedlings to adults, principally fortrees (Cornelissen et al. 2003), and plants of thesame species may have different responses toenvironmental factors when growing in differenthabitats (Bellingham and Sparrow 2000; Vesk andWestoby 2004).
Traits generally used to distinguish canopyversus understory trees and deciduous versusevergreen species in studies identifying functionaltypes in forests (Condit et al. 1996) should be ofless importance to understand spatial patterns inforest–grassland ecotones, since understory tre-elets (e.g. Myrciaria cuspidata O. Berg, Eugenia
hyemalis Camb.) as well as canopy species (e.g.
Myrsine umbellata Mart., Matayba elaeagnoides
Radlk.) were found colonizing grassland areasand practically all sampled plants were evergreenspecies. This underlines the importance ofobserving traits locally and de�ning PFTs inrelation to local disturbance pressure or envi-ronmental conditions, which has been proposedas an advantage of the present method (Pillar andSosinski 2003), allowing for a context-dependentclassi�cation of PFTs (Lavorel et al. 1997).
PFTs of forest species in forest–grasslandtransitions
Among the PFTs de�ned in the analysis of forest–grassland transition, the traits that characterizedtypes occurring in grassland provided advantagesfor persistence under the current �re regime, suchas resprouting ability. Resprouting trees invest instorage organs, with generally slower growth andhigh persistence in time (D�az and Cabido 1997),in contrast to pioneer trees. However, in our case,trees playing the role of pioneers (we say ‘‘play-ing’’ because most of the sampled species may befound inside mature forest as well, in canopy orunderstory strata) clearly were from species thatcould resprout. Despite the apparent incoher-ence, we should take into account that for an
8 Plant Ecol (2007) 189:1–14
123

individual tree colonizing a grassland submittedto frequent burns, not only dispersal, establish-ment and recruitment are critical, but its successis also (if not principally) restricted by thecapacity to survive recurrent �re events. There-fore, the generally known ‘‘typical’’ pioneer treeswith fast (above-ground) growth but low invest-ment in storage organs were absent in these areassubject to frequent burns.
There is also a stochastic component for suc-cessful persistence in grassland, which can be re-lated to patchiness in �re events, giving betterchances of survival to trees growing on siteswhere �re is less intense (Bond and Midgley2001). Furthermore, other attributes of PFT2 (thetype of forest species present in grassland plots)such as mid-size leaves with no membranoustexture, low-high branched woody stems andshorter plants re�ect strategies for ef�cient use ofwater in sunny habitats (by diminishing leaf areaand increasing cuticle). These species invest inunderground storage organs, but not in height(Bellingham and Sparrow 2000). The fact thatforest plots closest to the forest limit presentedhigher PFT-diversity may suggest that in absenceof �re other PFTs tend to advance further inadjacent grassland plots, even if slowly. Probablyonly PFT7 and 8 (most understory shrubs andshade-tolerant treelets) are always restricted tooccur under a dense canopy.
PFTs of woody species in burned grassland
Considering forest and non-forest woody plantsestablished in grassland, our results revealed verysimple PFTs associated to vegetation dynamicssince the last �re event. We have con�dence onthe assumption that groups 1 through 4 in ourexperimental setup correspond to correctly or-dered post-�re recovery stages. Comparing re-cently burned plots (last �re taking place3 months before) with plots with longer timewithout �re (3–4 years), differences in PFT com-position demonstrate important features of woo-dy plants in frequently burned grassland and inadjacent forest border sites (less frequent �re).Two types representing trees with establishedindividuals on grassland were distinguished. Onetype (with resprouting ability and multi-stemmed
base) occupies grassland and border plots inde-pendent of post-�re recovery stage; its persistencein grassland plots seems to be linked to stochasticevents of establishment in sites with less �reseverity, considering the constancy in basal coveracross post-�re recovery stages. Another type(with resprouting ability, single-stemmed baseand potentially taller than the �rst one) was moreabundant in plots close to the forest border and ingrassland plots at late post-�re recovery stage,being practically absent in recently burned plots(3 months to 1 year), except for a few individualsthat were established under adult tree plants(�eld observation). In the present study, speciessuch asMyrciaria cuspidata and Myrcia palustris
(DC.) Kausel displayed a wide range of growthforms, corresponding to different PFTs (bothtypes cited before). These are examples of speciesthat are able to assume different structural growthforms under contrasting environmental condi-tions (e.g. forest and grassland) or under distinctdisturbance pressures (e.g. �re and herbivory)(Archibald and Bond 2003).
Taller shrubs occurred in all post-�re recoverystages, but with lower cover in recently burnedareas. Some examples wereD. viscosa, Hetero-
thalamus psiadioides Less. and Baccharis dra-
cunculifolia DC., which are fast growing specieswith a large amount of seeds dispersed by windbut unable to resprout after �re events. In con-trast, multi-stemmed resprouting grasslandshrubs were less frequent in border plots anddenser in recently burned plots. Although mostspecies of this type also have the capacity of fastgrowth, they invest in underground storage or-gans, which result in shorter individuals if com-pared to single-stemmed shrubs (Bellingham andSparrow 2000). Some important species of thisresprouting type were Baccharis cognata DC., B.
trimera (Less.) DC., B. ochracea Spreng., B.
sessilifolia DC., Vernonia nudiflora Less.,Schinus
weinmanniaefolius Engl. and Eupatorium ligula-
efolium Hook. et Arn.. The presence of only fourPFTs of woody plants in grassland at differentpost-�re recovery stages suggests that a verysimple classi�cation is suf�cient to evaluate thedynamics of communities under �re disturbance,as supported by Noble and Gitay (1996) andPausas (1999). Traits de�ning these PFTs were
Plant Ecol (2007) 189:1–14 9
123

plant height, stem base, stem consistency, habit andresprouting ability.
Woody PFTs dynamics of adjacent transectplots (groups 1 and 3) showed that �re effect wassigni�cant in reducing cover of tall single-stem-med shrubs although increasing cover of multi-stemmed shrubs, which should be local persistent(persistence niche—Bond and Midgley 2001).Woody species abundance in grassland vegetationis frequently related to disturbances, including�re, grazing and CO 2 concentration changes, andmay increase in a complex trade-off with grassdominance (Bond and Midgley 2000; Roqueset al. 2001; Langevelde et al. 2003), often for somespeci�c types only, such as resprouter species(Pendergrass et al. 1998). On the other hand, �recan decrease diversity of woody species regener-ative types due to the elimination of those withouteffective mechanisms for post-�re regeneration(Lloret and Vila` 2003). In the present study, whenconsidering composition of all woody plants(grassland and forest species), PFT diversity ingrassland was low only in plots which had beenvery recently burned, which suggests spatial sta-bility of the woody plants community under thepresent �re regime. However, when consideringPFT diversity for forest species along the forest–grassland transition, a drastic decrease occurredfrom the border to the grassland plots, indicating�re in�uence on the maintenance of this pattern.
The analyses revealed that not only commonspecies were important to characterize commu-nity changes, as had been supposed before (D�´azet al. 2004), but that, in contrast, all species maybe necessary, even (or especially) the less fre-quent ones, as these may form speci�c PFTs forspecial situations in the system. PFTs formed byinfrequent species with similar attributes, andthus with similar function or responses to theenvironment, could, for example, indicate plantsor groups of plants that are more endangered inthe community under the present environmentalconditions or disturbance regimes (it is the case offorest species that colonize grassland areas). Onthe other hand, PFTs formed by frequent speciesin communities susceptible to disturbance, such asthe multi-stemmed grassland shrubs in the pres-ent study, may be responsible for ecosystemresilience, seing that they remain practicallyunaffected under recurrent disturbance events(Eriksson 2000).
Acknowledgments We would like to thank EnioSosinski Jr. and Carolina Blanco for discussion andtechnical assistance and Joa˜o A. Jarenkow, Marcos So-bral, Ilsi I. Boldrini, Nelson Matzenbacher, and PauloBrack for their help with species identi�cation. S.M. wassupported by a CAPES PhD scholarship and G.O. by aPhD grant from the German National Academic Foun-dation. V.P. received support from CNPq. The projecthas been supported by CAPES (Brazil) and DAAD(Germany) under ProBral.
Family Species Code Habit Analysis
Acanthaceae Justicia brasiliana Roth Jubr 1 aAnacardiaceae Schinus molle L. Scmo 4 a
Lithrea brasiliensis March. Libr 4 a, bSchinus weinmanniaefolius Engl. Scwe 1 b
Annonaceae Rollinia sylvatica (A.St.Hil.) Mart. Rosy 3 aAquifoliaceae Ilex dumosa Reiss. Ildu 2 aArecaceae Syagrus romanzoffiana (Cham.) Glassm. Syro 4 a
Butia capitata (Mart.) Becc. Buca 4 a, b
Appendix
Appendix 1 List of families and species, their code, habitand inclusion in the PFTs-based analyses (a: forest–grassland transition; b: border + grassland plots (groups1–4); c: only plots of groups 1 and 3 in both survey years.
See methods for more explanations). For trait habit = 1:shrub, 2: treelets, 3: low-size tree, 4: mid-size trees, 5: largetrees
10 Plant Ecol (2007) 189:1–14
123
Appendix 1 continued
Family Species Code Habit Analysis
Asteraceae Baccharidastrum triplinervium (Less.) Cabrera BAtr 1 bBaccharis articulata (Cham.) Pers. Baar 1 bBaccharis cognata DC. Baco 1 bBaccharis dracunculifolia DC. Badr 1 bBaccharis leucopappa DC. Bale 1 bBaccharis ochracea Spreng. Baoc 1 bBaccharis patens Baker Bapa 1 bBaccharis sessilifolia DC. Base 1 bBaccharis trimera (Less.) DC. Batr 1 bEupatorium intermedium DC. Euin 1 bEupatorium ligulaefolium Hook. et Arn. Euli 1 bEupatorium pedunculosum Hook. et Arn. Eupe 1 bEupatorium tweendianum Hook. et Arn. Eutw 1 bHeterothalamus psiadioides Less. Heps 1 bPorophyllum lanceolatum DC. Pola 1 bVerbesina subcordata DC. Vesu 1 bVernonia nudiflora Less. Venu 1 b
Cactaceae Cereus hildmannianus K. Schum. Cehi 2 aOpuntia monacantha Haw. Opmo 1 a,b
Celastraceae Maytenus cassineformis Reiss. Maca 2 a, bClusiaceae Garcinia gardneriana (Pl. et Tr.) Zappi Gaga 2 aEbenaceae Diospyros inconstans Jacq. Diin 3 a, bEricaceae Agarista eucalyptoides (Cham. et Schl.) G. Don Ageu 2 a, bErythroxylaceae Erythroxylum argentinum O.E. Schulz Erar 3 a
Erythroxylum microphyllum A.St.Hil. Ermi 1 bEuphorbiaceae Gymnanthes concolor Spreng. Gyco 2 a
Pachystroma longifolium (Nees) Johnst. Palo 4 aSebastiania brasiliensis Spreng. Sebr 3 a, bSebastiania serrata (M.Arg.) M.Arg. Sese 3 a, bCroton cf.nitrariaefolius Baill. Crni 1 bCroton cf.thermarum Mu ll.Arg. Crth 1 b
Fabaceae Calliandra tweediei Benth. Catw 2 a, bCollaea stenophylla Benth. Cost 1 bMimosa parvipinna Benth. Mipa 1 b
Icacinaceae Citronella paniculata (Mart.) R.A.Howard Cipa 2 aLamiaceae Hyptis mirabilis Briq. Hymi 1 bLauraceae Aiouea saligna Meis. Aisa 4 a
Endlicheria paniculata (Spreng.) Macbr. Enpa 2 aNectandra megapotamica (Spreng.) Mez Neme 5 aOcotea indecora Schott ex Meissn. Ocin 4 aOcotea pulchella Mart. OCpu 4 aOcotea silvestris Vattimo-Gil Ocsi 4 aOcotea puberula Nees Ocpu 5 a, b
Loganiaceae Strychnos brasiliensis (Spreng.) Mart. Stbr 2 aMalvaceae Triumfetta semitriloba Jack. Trse 1 a
Pavonia hastata Cav. Paha 1 bSida rhombifolia L. Sirh 1 b
Melastomataceae Miconia hyemalis A.St.Hil. et Naud. Mihy 1 aMeliaceae Cabralea canjerana (Vell.) Mart. Caca 5 a
Trichilia claussenii C.DC. Trcl 4 a, bTrichilia elegans A.Juss. Trel 2 a
Monnimiaceae Mollinedia elegans Tul. Moel 1 a,bMoraceae Sorocea bonplandii (Bail.) Burg. Lanj. et Boer Sobo 2 a
Plant Ecol (2007) 189:1–14 11
123
Appendix 1 continued
Family Species Code Habit Analysis
Myrsinaceae Myrsine guianensis (Aubl.) Kuntze Mygu 3 aMyrsine coriacea (Sw.) R. Br. Myco 4 a, bMyrsine umbellata Mart. Myum 4 a, b
Myrtaceae Blepharocalyx salicifolius (Kunth) O.Berg Blsa 3 aCampomanesia aurea O.Berg Caau 2 a, bCampomanesia xanthocarpa O.Berg Caxa 4 aEugenia involucrata DC. EUin 3 aEugenia hyemalis Camb. Euhy 2 a, bEugenia rostrifolia D.Legrand Euro 4 aEugenia schuechiana O.Berg Eusc 2 aEugenia uniflora L. Euun 2 aEugenia uruguayensis Camb. Euur 2 aMyrcia glabra (Berg) Legr. Mygl 2 aMyrcianthes gigantea (Legr.) Legr. Mygi 3 aMyrcia palustris (DC.) Kausel Mypa 2 a, bMyrciaria cuspidata O.Berg Mycu 2 a, bPsidium sp. Pssp 1 b
Nyctaginaceae Guapira opposita (Vell.) Reitz Guop 4 a, bOpiliaceae Agonandra brasiliensis Benth. & Hook.f. Agbr 3 aPiperaceae Piper gaudichaudianum Kunth Piga 1 aProteaceae Roupala brasiliensis Klotz. Robr 4 aQuillajaceae Quillaia brasiliensis (A.St.Hil. et Tul.) Mart. Qubr 4 a, bRosaceae Prunus myrtifolia Koehne Prse 4 aRubiaceae Faramea montevidensis Cham. Fama 2 a
Psychotria brachyceras M.Arg. Psbr 1 aRudgea parquioides (Cham.) M.Arg. Rupa 1 aPsychotria carthagenensis Jacq. Psca 1 a, bPsychotria leiocarpa Cham. et Schtdl. Psle 1 a, bGuettarda uruguensis Cham. et Schtdl. Guur 2 a, b
Rutaceae Zanthoxylum rhoifolium Lam. Zarh 4 a, bSalicaceae Casearia decandra Jacq. Cade 2 a
Casearia sylvestris Sw. Casi 3 aXylosma pseudosalzmannii Sleumer Xyps 4 aBanara parviflora (Gray) Benth. Bapa 4 a
Sapindaceae Allophylus edulis (A.St.Hil.) Radlk. Aled 3 aCupania vernalis Cambess Cuve 4 a, bMatayba elaeagnoides Radlk. Mael 4 a, bDodonaea viscosa (L.) Jacq. Dovi 2 a, b
Sapotaceae Chrysophyllum marginatum (Hook. et Arn.) Radlk. Chma 2 aSolanaceae Cestrum strigillatum Ruiz et Pav. Cest 1 a, b
Solanum sanctaecatharinae Dunal Sosa 2 aStyracaceae Styrax leprosum Hook. et Arn. Stle 3 a, bSymplocaceae Symplocos tetrandra Mart. ex Miq. Syte 2 a, b
Symplocos uniflora (Pohl) Benth. Syun 2 a, bThymelaeaceae Daphnopsis racemosa Griseb. Dara 2 aUrticaceae Coussapoa microcarpa (Schott) Rizzini Comi 5 a
12 Plant Ecol (2007) 189:1–14
123

References
Archibald S, Bond WJ (2003) Growing tall vs growingwide: tree architecture and allometry of Acacia Kar-roo in forest, savanna, and arid environments. Oikos102:3–14
Barkman JJ (1988) New systems of plant growth forms andphenological plant types. In: Werger MJA, van derAart PJM, During HJ, Verhoeven JTA (eds) Plantform and vegetation structure. SPB Academic Pub-lishing, pp 9–44
Behling H (2001) South and southeast Brazilian grasslandsduring Late Quaternary times: a synthesis. Palaege-ogr, Palaeclimatol, Palaeoecol 2710:1–9
Behling H, Pillar VD, Orlo ´ci L, Bauermann SG (2004)Late Quaternary Araucaria forest, grassland (Cam-pos), �re and climate dynamics, studied by high-res-olution pollen, charcoal and multivariate analysis ofthe Cambara do Sul core in southern Brazil. Palae-geogr, Palaeclimatol, Palaeoecol 203:277–297
Bellingham PJ, Sparrow AD (2000) Resprouting as a lifehistory strategy in woody plant communities. Oikos89:409–416
Bond WJ, Midgley GF (2000) A proposed CO2-controlledmechanism of woody plant invasion in grassland andsavannas. Global Change Biol 6:865–869
Bond WJ, Midgley GF (2001) Ecology of sprouting inwoody plants: the persistence niche. Trends Ecol Evol16:45–51
Box EO (1996) Plant functional types and climate at theglobal scale. J Veg Sci 7:309–320
Brack P, Rodrigues RS, Sobral M, Leite SLC (1998)A rvores e arbustos na vegetac¸ao natural de PortoAlegre, Rio Grande do Sul, Brasil, Iheringia. Ser Bot51:139–166
Bugmann H (1996) Functional types of trees in temperateand boreal forest: classi�cation and testing. J Veg Sci7:359–370
Campbell BD, Stafford Smith DM, Ash AJ (1999) A rule-based model for the functional analysis of vegetationchange in Australasian grasslands. J Veg Sci 10:723–730
Condit R, Hubbell SP, Foster RB (1996) Assessing theresponse of plant functional types to climatic changein tropical forest. J Veg Sci 7:405–416
Cornelissen JHC, Cerabolini B, Castro-D�ey P, Villar-Salvador P, Montserrat-Mart� G, Puyravaud JP,
Appendix 2 Species belonging to the de�ned PFTs in the forest–grassland transition analysis (Table2). Understory shrubsare underlined. See species code in Appendix 1
PFT no.sp.
Species code
PFT 1 11 Mael, Zarh, Stle, Guur, Casi, Libr, Sese, Xips, Blsa, Mygu, BapaPFT 2 17 Syun, Ageu, Catw, Mycu, Syte, Caau, Guur, Euhy, Mypa,Psca, Euur, Cest, Maca,Mihi, Dara, Stbr, IlduPFT 3 29 Myco, Myum, Ocin, Guop, Rupa, Mygi, Ocpu, Euro, Trcl, Aisa, Sobo, Gaga, Mygl, OCpu, Buca, Fama,
Syro, Caca, Gyco, Euun, Scmo, Qubr, Neme, Ocsi, Cipa, Robr, Euin, Palo, ComiPFT 4 20 Dovi, Chma, Stle, Aled, Trel, Eusc,Moel, Diin, Cuve, Erar, Psle, Guur, Aisa, Sebr, Rosy, Prse, Caca,
Cade,Jubr, EnpaPFT 5 2 Opmo, CehiPFT 6 18 Mael, Syun, Zarh, Catw, Mycu, Syte, Casi, Euhy,Psca, Sosa, Xips, Euur, Cest, Maca,Mihi, Mygu, Trse, BapaPFT 7 25 Dovi, Chma, Stle, Aled, Trel, Eusc, Rupa,Moel, Diin, Cuve, Psbr, Erar,Psle, Sobo, Gaga, Sebr, Fama,
Prse, Caca, Cade,Piga, Gyco,Jubr, Caxa, AgbrPFT 8 14 Myco, Myum, Ocin, Guop, Mygi, Ocpu, Euro, Trcl, Aisa, OCpu, Syro, Neme, Ocsi, RobrPFT 9 4 Guop, Mygi, Mygl, Euun
Appendix 3 Species belonging to the de�ned PFTs in border and grassland plots in relation to elapsed time since last �re(Table 3). Treelets or tree species have underline code (Appendix 1)
PFT no.sp.
Species code
PFT 1 21 Mael,Syun,Zarh,Myco,Myum,Ageu,Mycu,Mypa,Maca,Syte,Guop,Cuve,Sebr,Ocpu,Trcl,Diin, Stle,Euhy,Sese,Qubr,Buca
PFT 2 21 Venu, Baco, Scwe,Mycu, Hymi, Baoc, Euli, Batr, Baar, Euin, Pola, Bapa, Base, Paha, Eupe,Maca,Eutw, Vesu, Bale,Euhy, Cost
PFT 3 27 Syun,Myco,Ageu, Baco, Scwe,Mycu,Libr, Baoc, Euli, Euin, Caau, Bapa, Base, Paha,Mypa, Crni,Cest,Guur,Syte, Mipa, Psca, Catw,Stle,Euhy,Sese, Cost, BAtr
PFT 4 30 Venu, Pssp,Dovi, Baco, Scwe, Hymi, Baoc, Euli, Batr, Baar, Euin, Pola, Caau, Badr, Bapa, Base,Heps, Crth, Paha, Eupe, Crni, Psle, Moel, Mipa, Psca, Eutw, Sirh,Opmo, Cost, Ermi
Plant Ecol (2007) 189:1–14 13
123

Maestro M, Werger MJA, et al (2003) Functionaltraits of woody plants: correspondence of speciesrankings between �eld adults and laboratory-grownseedlings? J Veg Sci 14:311–322
D� az S, Cabido M (1997) Plant functional types and eco-system function in relation to global change. J Veg Sci8:463–474
D� az S, Cabido M, Zak M, Carretero EM, Aran�´bar J(1999) Plant functional traits, ecosystem structure andland-use history along a climatic gradient in central-western Argentina. J Veg Sci 10:651–660
D� az S, Hodgson JG, Thompson K, Cabido M, Corne-lissen JHC, Jalili A, Montserrat-Mart� ´ G, Grime JP,et al (2004) The plant traits that drive ecosystems:Evidence from three continents. J Veg Sci 15:295–304
Duarte LS, Dos-Santos MG, Hartz SM, Pillar VD (inpress). The role of nurse plants onAraucaria forestexpansion over grassland in south Brazil. Australecology
Eriksson O (2000) Functional roles of remnant plantpopulations in communities and ecosystems. GlobalEcol Biogeogr 9:443–449
Garcia Martinez P (2005) Caracterizacion qu�mica y f�sicade los suelos del Morro Santana (Porto Alegre, RioGrande do Sul, Brasil). Chair of Soil Sciences, Tech-nische Universitat Mu nchen, Freising-Weihenste-phan, Germany
Hoffmann WA, Orthen B, Nascimento PKVd (2003)Comparative �re ecology of tropical savanna andforest trees. Funct Ecol 17:720–726
Langevelde FV, Van de Vijver CADM, Kumar L, KoppelJvd, Ridder Nd, Andel JV, Skidmore AK, HearneJW, et al (2003) Effects of �re and herbivory on thestability of savanna ecosystems. Ecology 84:337–350
Lavorel S, Garnier E (2002) Predicting changes in com-munity composition and ecosystem functioning fromplant traits: revisiting the Holy Grail. Funct Ecol16:545–556
Lavorel S, McIntyre S, Landsberg J, Forbes TDA (1997)Plant functional classi�cation, from general groups tospeci�c groups based on response to disturbance.Trends Ecol Evol 12:474–478
Lloret F, Vila` M (2003) Diversity patterns of plant func-tional types in relation to �re regime and previousland use in Mediterranean woodlands. J Veg Sci14:387–398
Mu ller SC, Forneck ED (2004) Forest–grassland mosaicsin the hills of Porto Alegre city: a study case of forestexpansion patterns in Santana hill, Rio Grande doSul, Brazil. In: MLP (ed) Workshop-Protec ao emanejo da vegetacao natural da regiao de Porto Al-egre, com base em pesquisas de padro˜es e dinamica davegetacao. -Programa de Pos-Graduacao em Ecolo-gia/UFRGS, pp 29–37
Nimer E (1990) Clima. In: Geogra�a do Brasil: Regia o Sul(ed IBGE), pp 151–(187) IBGE
Noble IR, Gitay H (1996) A functional classi�cation forpredicting the dynamics of landscapes. J Veg Sci7:329–336
Overbeck GE, Mu ller SC, Pfadenhauer J, Pillar VD (2005)Fine-scale post-�re dynamics in southern Braziliansubtropical grassland. J Veg Sci 16:655–664
Pausas JG (1999) Response of plant functional types tochanges in the �re regime in Mediterranean ecosys-tems: a simulation approach. J Veg Sci 10:717–722
Pendergrass KL, Miller PM, Kauffman JB (1998) Pre-scribed �re and the response of woody species inWillamette Valley wetland prairies. Restor Ecol6:303–311
Pillar VD (2003) Dinaˆmica de expansao �orestal em mo-saicos de �oresta e campos no sul do Brasil. In: Ca-bido M (ed) Ecossistemas brasileiros: manejo econservacao. pp 209–216. Expressa˜o Gra�ca e Editora
Pillar VD (2004) SYNCSA software for character-basedcommunity analysis, v. 2.2.4. -Departamento de Eco-logia, UFRGS
Pillar VD, Orlo´ci L (1993a) Character-Based CommunityAnalysis; the Theory and an Application Program.SPB Academic Publishing
Pillar VD, Orlo´ci L (1993b) Taxonomy and perception invegetation analysis. Coenoses 8:53–66
Pillar VD, Orlo´ci L (1996) On randomization testing invegetation science:multifactor comparisons of relevegroups. J Veg Sci 7:582–592
Pillar VD, Quadros FLF (1997) Grassland–forest bound-aries in southern Brazil. Coenoses 12:119–126
Pillar VD, Sosinski EE, Jr (2003) An improved method forsearching plant functional types by numerical analysis.J Veg Sci 14:323–332
Podani J (2000) Introduction to the exploration of multi-variate biological data. Backhuys Publishers
Rambo B (1956) A �sionomia do Rio Grande do Sul.Selbach
Roques KG, O’Connor TG, Watkinson AR (2001)Dynamics of shrub encroachment in an African sa-vanna: relative in�uences of �re, herbivory, rainfalland density dependence. J Appl Ecol 38:268–280
Sosinski EE, Jr, Pillar VD (2004) Respostas de tipos fun-cionais de plantas aintensidade de pastejo em vege-tacao campestre. Pesquisa Agropecua´ria Brasileira39:1–9
Teixeira MB, Coura-Neto AB, Pastore U, Rangel FilhoALR (1986) Vegetac ao. In: Levantamento de recur-sos naturais. (ed IBGE) pp 541–632. IBGE
Vesk PA, Westoby M (2004) Sprouting ability across di-verse disturbances and vegetation types worldwide. JEcol 92:310–320
Weiher E, van der Werf A, Thompson K, Roderick M,Garnier E, Eriksson O (1999) Challenging Theo-phrastus: a common core list of plant traits for func-tional ecology. J Veg Sci 10:609–620
Woodward FI, Cramer W (1996) Plant functional typesand climatic changes: introduction. J Veg Sci 7:306–308
14 Plant Ecol (2007) 189:1–14
123
Related Documents











