Plant bioassay to assess the effects of allelochemicals on the metabolome of the target species Aegilops geniculata by an NMR-based approach Brigida D’Abrosca a , Monica Scognamiglio a,⇑ , Vittorio Fiumano a , Assunta Esposito a , Young Hae Choi b , Robert Verpoorte b , Antonio Fiorentino a a Department of Environmental, Biological and Pharmaceutical Sciences and Technologies, Second University of Naples, via Vivaldi 43, I-81100 Caserta, Italy b Natural Products Laboratory, Institute of Biology Leiden, Leiden University, P.O. Box 9502, 2300 RA Leiden, The Netherlands article info Article history: Received 1 August 2012 Received in revised form 20 March 2013 Available online 27 April 2013 Keywords: Allelochemicals Aegilops geniculata Roth. Metabolome NMR-based metabolomics abstract A metabolomic-based approach for the study of allelopathic interactions in the Mediterranean area is proposed using Aegilops geniculata Roth (Poaceae), a Mediterranean herbaceous plant, as test species. Its metabolome has been elucidated by 1D and 2D NMR experiments. Hydroponic plant cultures of A. geniculata were treated with specific compounds of known allelopathic potential: catechol, coumarin, p-coumaric acid, p-hydroxybenzoic acid, ferulic acid and juglone. The met- abolic variations due to the presence of allelochemicals have been analyzed and measured. All of the compounds showed the strongest effects at the highest concentration, with coumarin and juglone as the most active compounds, causing an increase of several metabolites. The metabolome changes in test plants confirmed the allelochemicals’ reported modes of action. The results demonstrated that the proposed method is a promising tool. It can be applied to plant extracts, making it possible to evidence the metabolites responsible for the activity, as well as their mech- anisms of action. Ó 2013 Elsevier Ltd. All rights reserved. 1. Introduction Unravelling the allelopathic interaction between plants requires a systemic approach in which the input from several fields, such as plant physiology, ecology, biochemistry, agriculture and natural products chemistry can be used for a better comprehension of the inherent physiological and metabolic processes. The elucida- tion of allelopathic interactions may aid to explain their involve- ment in the ecosystem. Besides the fundamental questions to be resolved, it appears that allelopathy is a valuable source for natural herbicides discovery, as many allelochemicals exhibit phytotoxic activities. Unfortunately, despite the numerous previous studies, allelo- pathic interactions are far from being clarified and it remains a controversial topic (Gatti et al., 2010). Conclusive evidence for an allelopathic interaction is not a simple find, as this biological activ- ity is governed by a number of different factors such as: individual chemical structure, antagonistic, agonistic, additive, sequential, or synergistic effects (An et al., 2001), to name but a few. Up to now, notwithstanding the evident requirement of new methods to be used in allelopathy, few papers deal with a systems biology approach in this field (Rinu et al., 2005). This holistic approach, relying on ‘‘omics’’ sciences, studies the totality of a pro- cess or of an organism in an integrated way, at different levels of the biological organization (Keurentjes et al., 2011). In this context, metabolomics, with an ultimate aim of both the qualitative and quantitative analysis of all metabolites in an organism (Kim et al., 2011; Verpoorte et al., 2007), could be an important procedure in identifying compounds involved in allelopathic interactions. An interesting multidisciplinary approach has been used by Broz et al. (2008, 2010) in order to study the effects of neighbour- ing species and other organisms on plant metabolome: the chem- ical composition of plant extracts in response to specific environments and stimuli has been studied, while no allelochemi- cal involvement has been proven. To get more insight into the allelopathic process, an NMR based metabolomics approach can be applied to plant extracts, thanks to high reproducibility, easy quantification and fast analysis (Ver- poorte et al., 2007). Compared to the classical methods used to iso- late allelochemicals, metabolomics gives the possibility to identify them without purification steps: plant extracts’ composition is de- fined, and specific correlations between their growth inhibition activity and their metabolite content can give clues about the ac- tive compounds. Besides the application of metabolomics in replacement of bioassay guided fractionation, the analysis of ex- tract activity has the advantage to highlight synergistic or additive 0031-9422/$ - see front matter Ó 2013 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.phytochem.2013.03.016 ⇑ Corresponding author. Tel.: +39 0823 274507; fax: +39 0823 274571. E-mail address: [email protected] (M. Scognamiglio). Phytochemistry 93 (2013) 27–40 Contents lists available at SciVerse ScienceDirect Phytochemistry journal homepage: www.elsevier.com/locate/phytochem

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Phytochemistry 93 (2013) 27–40
Contents lists available at SciVerse ScienceDirect
Phytochemistry
journal homepage: www.elsevier .com/locate /phytochem
Plant bioassay to assess the effects of allelochemicals on the metabolome ofthe target species Aegilops geniculata by an NMR-based approach
Brigida D’Abrosca a, Monica Scognamiglio a,⇑, Vittorio Fiumano a, Assunta Esposito a, Young Hae Choi b,Robert Verpoorte b, Antonio Fiorentino a
a Department of Environmental, Biological and Pharmaceutical Sciences and Technologies, Second University of Naples, via Vivaldi 43, I-81100 Caserta, Italyb Natural Products Laboratory, Institute of Biology Leiden, Leiden University, P.O. Box 9502, 2300 RA Leiden, The Netherlands
a r t i c l e i n f o
Article history:Received 1 August 2012Received in revised form 20 March 2013Available online 27 April 2013
Keywords:AllelochemicalsAegilops geniculata Roth.MetabolomeNMR-based metabolomics
0031-9422/$ - see front matter � 2013 Elsevier Ltd. Ahttp://dx.doi.org/10.1016/j.phytochem.2013.03.016
⇑ Corresponding author. Tel.: +39 0823 274507; faxE-mail address: [email protected] (M
a b s t r a c t
A metabolomic-based approach for the study of allelopathic interactions in the Mediterranean area isproposed using Aegilops geniculata Roth (Poaceae), a Mediterranean herbaceous plant, as test species.Its metabolome has been elucidated by 1D and 2D NMR experiments.
Hydroponic plant cultures of A. geniculata were treated with specific compounds of known allelopathicpotential: catechol, coumarin, p-coumaric acid, p-hydroxybenzoic acid, ferulic acid and juglone. The met-abolic variations due to the presence of allelochemicals have been analyzed and measured. All of thecompounds showed the strongest effects at the highest concentration, with coumarin and juglone asthe most active compounds, causing an increase of several metabolites. The metabolome changes in testplants confirmed the allelochemicals’ reported modes of action.
The results demonstrated that the proposed method is a promising tool. It can be applied to plantextracts, making it possible to evidence the metabolites responsible for the activity, as well as their mech-anisms of action.
� 2013 Elsevier Ltd. All rights reserved.
1. Introduction
Unravelling the allelopathic interaction between plants requiresa systemic approach in which the input from several fields, such asplant physiology, ecology, biochemistry, agriculture and naturalproducts chemistry can be used for a better comprehension ofthe inherent physiological and metabolic processes. The elucida-tion of allelopathic interactions may aid to explain their involve-ment in the ecosystem. Besides the fundamental questions to beresolved, it appears that allelopathy is a valuable source for naturalherbicides discovery, as many allelochemicals exhibit phytotoxicactivities.
Unfortunately, despite the numerous previous studies, allelo-pathic interactions are far from being clarified and it remains acontroversial topic (Gatti et al., 2010). Conclusive evidence for anallelopathic interaction is not a simple find, as this biological activ-ity is governed by a number of different factors such as: individualchemical structure, antagonistic, agonistic, additive, sequential, orsynergistic effects (An et al., 2001), to name but a few.
Up to now, notwithstanding the evident requirement of newmethods to be used in allelopathy, few papers deal with a systems
ll rights reserved.
: +39 0823 274571.. Scognamiglio).
biology approach in this field (Rinu et al., 2005). This holisticapproach, relying on ‘‘omics’’ sciences, studies the totality of a pro-cess or of an organism in an integrated way, at different levels of thebiological organization (Keurentjes et al., 2011). In this context,metabolomics, with an ultimate aim of both the qualitative andquantitative analysis of all metabolites in an organism (Kim et al.,2011; Verpoorte et al., 2007), could be an important procedure inidentifying compounds involved in allelopathic interactions.
An interesting multidisciplinary approach has been used byBroz et al. (2008, 2010) in order to study the effects of neighbour-ing species and other organisms on plant metabolome: the chem-ical composition of plant extracts in response to specificenvironments and stimuli has been studied, while no allelochemi-cal involvement has been proven.
To get more insight into the allelopathic process, an NMR basedmetabolomics approach can be applied to plant extracts, thanks tohigh reproducibility, easy quantification and fast analysis (Ver-poorte et al., 2007). Compared to the classical methods used to iso-late allelochemicals, metabolomics gives the possibility to identifythem without purification steps: plant extracts’ composition is de-fined, and specific correlations between their growth inhibitionactivity and their metabolite content can give clues about the ac-tive compounds. Besides the application of metabolomics inreplacement of bioassay guided fractionation, the analysis of ex-tract activity has the advantage to highlight synergistic or additive

28 B. D’Abrosca et al. / Phytochemistry 93 (2013) 27–40
effects, which are often only supposed to exist, but very difficult todemonstrate. This approach is well standardized, as it has been ap-plied to the study of several biological activities (Yuliana et al.,2011), but, to the best of our knowledge, it is rarely used to studyallelopathy.
In the allelopathic phenomenon donor and receiving plants areinvolved. The above mentioned approach can be used in order tostudy the donor plant properties. Concerning the receiving plant,classical bioassays aim at the evaluation of seedling performance,by measuring some parameters like shoot/root growth, plant bio-mass or also physiological processes (Macías et al., 2000). Thesetests are usually enough to determine if a compound is toxic ornot, but monitoring the receiving plant metabolome can help inelucidating the mode of action and also to study the fate of theallelochemical (e.g. if it is translocated, metabolized, detoxified,etc.). Knowing the effects of a chemical on plant metabolism maybe very important both in order to suggest its role in a natural con-text, but in a herbicide development framework as well. Herewitha method to study the receiving plant response to allelochemicals,which takes under consideration the effects of allelochemicals onplant metabolism, is reported.
With the aim of standardization and validation, test plants weretreated with known allelochemicals. Metabolomic analysis wascarried out in order to measure the effects of the compounds onplant metabolism and to associate the known modes of action withmetabolic changes in the treated plants.
This approach is going to be applied to the elucidation of allelo-pathic interactions in the Mediterranean ecosystem, one of theworld’s biomes with the highest biodiversity (Naveh, 1989).
Our research group is studying this phenomenon (Espositoet al., 2008; Scognamiglio et al., 2012a,b), as Mediterranean vege-tation is characterized by a complex mosaic of high species andplant communities diversity, but it is not clear to what extent alle-lopathy plays a role in structuring these plant coenoses and theirdynamical processes.
Aegilops geniculata Roth. (Poaceae) was chosen as model plant inthis study. This is a steno-Mediterranean (areal distribution con-fined to Mediterranean coasts, characterized by arid summer per-iod) monocotyledon annual herbaceous plant, also known asTriticum ovatum L. or wild wheat (Pignatti, 1982). It has showngreat tolerance to drought and saline stress, obtained by the reduc-tion of leaf number and length, osmotic adjustment, decrease ofcell elasticity and the increase of apoplast water content (Mguiset al., 2012; Pradhan et al., 2012).
The reported method can be helpful in elucidating allelopathicinteractions giving the chance to identify in a single experimentalprocedure the active compounds, their effects on receiving plantmorphology and metabolism (and therefore their putative modeof action), and their fate.
Moreover, the detection of metabolic changes upon allelochem-ical treatment may offer an useful tool: a specific response patterncan be ascribed to a given mode of action. As a consequence, themechanisms of new allelochemicals could be hypothesized, nar-rowing down the number of experiments to determine and proveit.
2. Results and discussion
2.1. A. geniculata metabolites identification
With the aim to set up a new method to study allelopathicinteractions among Mediterranean species, and to overcome sev-eral limitations in allelopathy studies, some typical plants of Med-iterranean vegetation were grown in a hydroponic system made upby a tube filled with 5 mL of Hoagland solution and with a layer of
perlite, in order to assure mechanical support. Among them, A. gen-iculata was selected as model plant, based on several consider-ations: first of all, it is a Mediterranean species and it allowsstudying allelopathic interactions among species belonging to thesame habitat and chorological group. It is also fast growing andbig enough to let the treatments and analyses to be performed ina short time. Moreover it has a quite simple metabolome: animportant feature that allows a simple elucidation of what is goingon in the system.
In the first step, A. geniculata plants were grown using thehydroponic system and NMR metabolomic analysis was carriedout. Main metabolites (Fig. 1, Table 1) were identified on the basisof literature data (Ali et al., 2009; Liu et al., 2010; Kim et al., 2010;López-Gresa et al., 2010; Verpoorte et al., 2007), and their identi-ties were confirmed by HMBC correlations (Fig. 2, Table 1), alongwith other 2D-NMR experiments (HSQC and COSY). Apart fromeasily identified signals belonging to amino acids, organic acidsand sugars, two singlets, at d 6.73 and at d 1.76, along with a fewother signals, could not be attributed to any compound in the firstanalysis, so further NMR experiments were needed.
The singlet at d 6.73, attached to a carbon at d 134.6, showedcorrelations, in the HMBC spectrum with an olefinic carbon at d141.1, a carboxylic carbon at d 177.1 and with a methylene carbonat d 39.4, bound to the protons at d 3.56 and 3.99. The former pro-ton was in turn correlated to the carbons at d 134.6 and 141.1 andto two carboxylic carbons: the already mentioned signal at d 177.1and another one at d 180.1. The proton at d 3.99 showed correla-tions with the latter carbon and with a third carboxylic carbon sig-nal at d 176.0. These data are in good agreement with atricarboxylic acid: cis-aconitic acid, an important metabolicintermediate.
Besides all of these ubiquitous metabolites, a hemiterpene gly-coside was also found. A singlet at d 1.76, belonging to a methyl ona double bond, showed correlations with two olefinic carbons at d115.1 and 146.9 and with a carbinol carbon at d 76.9, bound,according to the HSQC experiment, to a proton at d 4.20. The car-bon at d 115.1 showed correlation with two protons in the HSQCexperiment: the one at d 4.98 (1H, s), correlated, in the HMBC spec-trum, with the methyl carbon at d 20.9 and with the other olefiniccarbon; the other at d 5.06 (1H, s), showed the same correlationsalong with a correlation with the carbon at d 76.9. The olefinic car-bon at d 146.9 correlated also with two diasterotopic protons at d3.31 and 4.32, bound to the C-4 (d 76.3) of the aglycone moiety.The former proton showed a correlation with the anomeric carbonof a glucose moiety at d 105.7. This one was bound to a proton at d4.26 (doublet, J = 7.8 Hz), whose values were in accordance with ab-glucose moiety. Further correlations due to the glucose moietywere also evident, but, as the sugar region in the extract spectrumwas very crowded, a purification of this compound was carried out,in order to confirm its identity. 1D and 2D NMR data of the puremetabolite were in good agreement with those reported by Ouy-ang et al. (2007) and allowed the compound to be identified asoblongaroside A, previously reported from Ilex oblonga.
2.2. Plant treatments
The next step was aimed at determining whether or not theplant metabolome was in any way affected by the addition of aspecific metabolite with known allelopathic potential to the cul-ture media.
Plants react to external stimuli turning on and off differentgenes, expressing different proteins and enzymes, producing a gi-ven metabolic state. Hence, as a perturbation (e.g. a physical,chemical or biotic stress) alters to some extent plant metabolism,measuring the metabolome of treated plants gives important infor-mation on what is happening at the biochemical level in the cells.
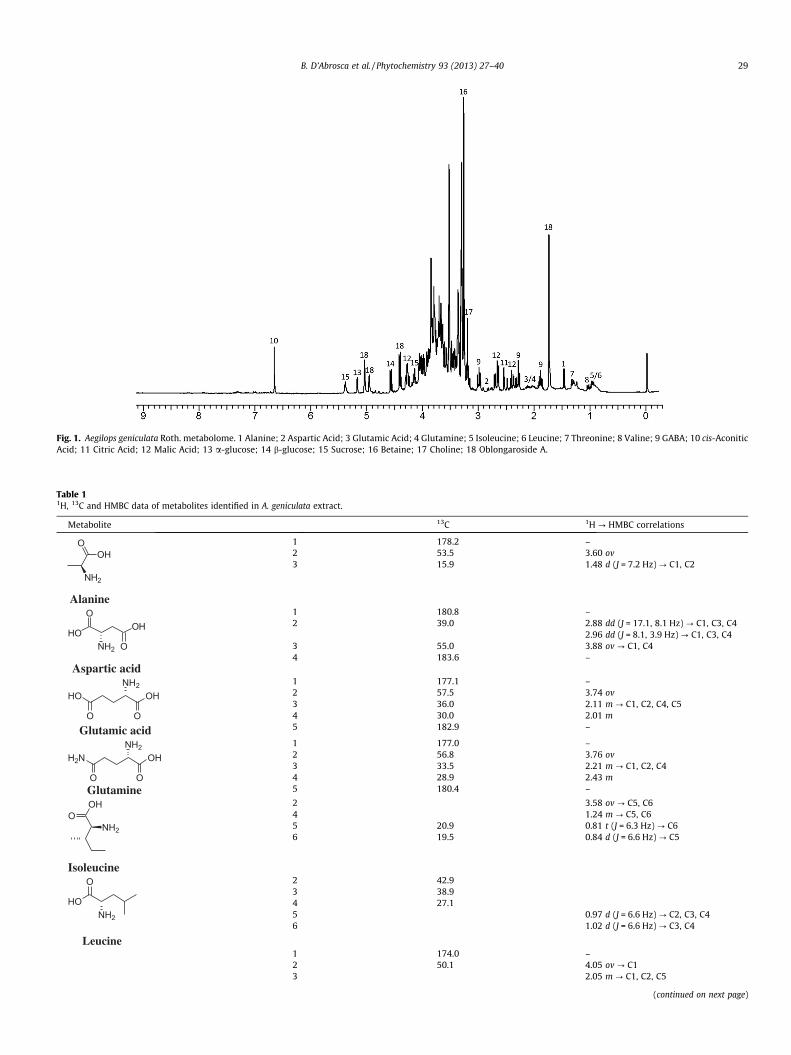
Fig. 1. Aegilops geniculata Roth. metabolome. 1 Alanine; 2 Aspartic Acid; 3 Glutamic Acid; 4 Glutamine; 5 Isoleucine; 6 Leucine; 7 Threonine; 8 Valine; 9 GABA; 10 cis-AconiticAcid; 11 Citric Acid; 12 Malic Acid; 13 a-glucose; 14 b-glucose; 15 Sucrose; 16 Betaine; 17 Choline; 18 Oblongaroside A.
Table 11H, 13C and HMBC data of metabolites identified in A. geniculata extract.
Metabolite 13C 1H ? HMBC correlations
NH2
OOH
Alanine
1 178.2 –2 53.5 3.60 ov3 15.9 1.48 d (J = 7.2 Hz) ? C1, C2
NH2 O
OHO
HO
Aspartic acid
1 180.8 –2 39.0 2.88 dd (J = 17.1, 8.1 Hz) ? C1, C3, C4
2.96 dd (J = 8.1, 3.9 Hz) ? C1, C3, C43 55.0 3.88 ov ? C1, C44 183.6 –
NH2
O
HO
O
OH
Glutamic acid
1 177.1 –2 57.5 3.74 ov3 36.0 2.11 m ? C1, C2, C4, C54 30.0 2.01 m5 182.9 –
NH2
O
H2N
O
OH
Glutamine
1 177.0 –2 56.8 3.76 ov3 33.5 2.21 m ? C1, C2, C44 28.9 2.43 m5 180.4 –
NH2O
OH
Isoleucine
2 3.58 ov ? C5, C64 1.24 m ? C5, C65 20.9 0.81 t (J = 6.3 Hz) ? C66 19.5 0.84 d (J = 6.6 Hz) ? C5
NH2
O
HO
Leucine
2 42.93 38.94 27.15 0.97 d (J = 6.6 Hz) ? C2, C3, C46 1.02 d (J = 6.6 Hz) ? C3, C4
1 174.0 –2 50.1 4.05 ov ? C13 2.05 m ? C1, C2, C5
(continued on next page)
B. D’Abrosca et al. / Phytochemistry 93 (2013) 27–40 29
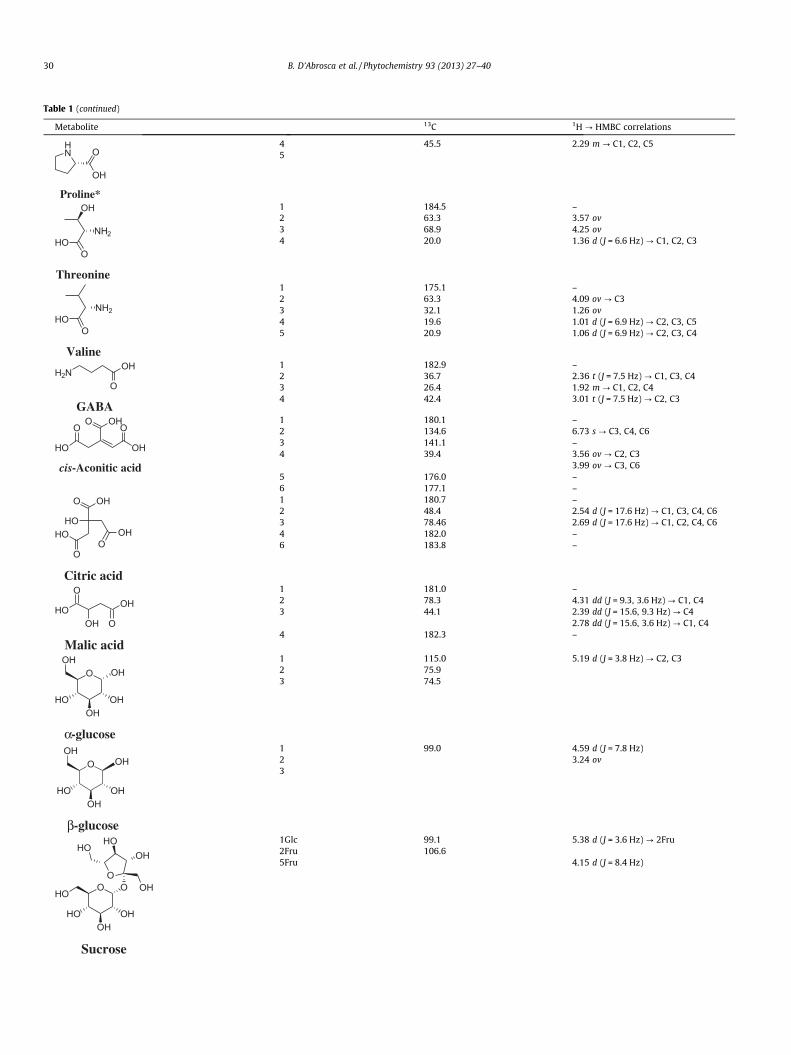
Table 1 (continued)
Metabolite 13C 1H ? HMBC correlations
HN O
OH
Proline*
4 45.5 2.29 m ? C1, C2, C55
NH2
OH
OHO
Threonine
1 184.5 –2 63.3 3.57 ov3 68.9 4.25 ov4 20.0 1.36 d (J = 6.6 Hz) ? C1, C2, C3
NH2
OHO
Valine
1 175.1 –2 63.3 4.09 ov ? C33 32.1 1.26 ov4 19.6 1.01 d (J = 6.9 Hz) ? C2, C3, C55 20.9 1.06 d (J = 6.9 Hz) ? C2, C3, C4
H2NO
OH
GABA
1 182.9 –2 36.7 2.36 t (J = 7.5 Hz) ? C1, C3, C43 26.4 1.92 m ? C1, C2, C44 42.4 3.01 t (J = 7.5 Hz) ? C2, C3
O
OH
O OHO
HO
cis-Aconitic acid
1 180.1 –2 134.6 6.73 s ? C3, C4, C63 141.1 –4 39.4 3.56 ov ? C2, C3
3.99 ov ? C3, C65 176.0 –6 177.1 –
HO
OOH
O OH
O
HO
Citric acid
1 180.7 –2 48.4 2.54 d (J = 17.6 Hz) ? C1, C3, C4, C63 78.46 2.69 d (J = 17.6 Hz) ? C1, C2, C4, C64 182.0 –6 183.8 –
OH
O
HOO
OH
Malic acid
1 181.0 –2 78.3 4.31 dd (J = 9.3, 3.6 Hz) ? C1, C43 44.1 2.39 dd (J = 15.6, 9.3 Hz) ? C4
2.78 dd (J = 15.6, 3.6 Hz) ? C1, C44 182.3 –
OHOH
HO
OOH
OH
α-glucose
1 115.0 5.19 d (J = 3.8 Hz) ? C2, C32 75.93 74.5
OHOH
HO
OOH
OH
β-glucose
1 99.0 4.59 d (J = 7.8 Hz)2 3.24 ov3
OH
OHHO
O
HO
O
OHOH
HO
HOO
Sucrose
1Glc 99.1 5.38 d (J = 3.6 Hz) ? 2Fru2Fru 106.65Fru 4.15 d (J = 8.4 Hz)
30 B. D’Abrosca et al. / Phytochemistry 93 (2013) 27–40
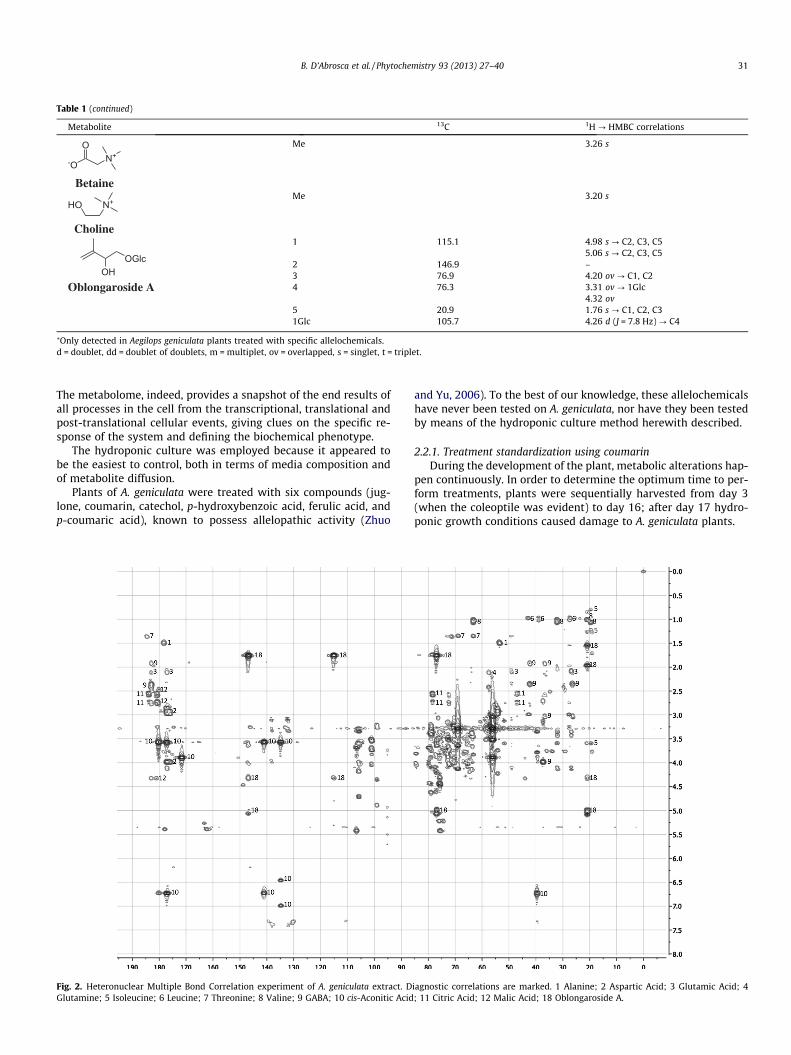
Table 1 (continued)
Metabolite 13C 1H ? HMBC correlations
-O
ON+
Betaine
Me 3.26 s
HO N+
Choline
Me 3.20 s
OGlcOH
Oblongaroside A
1 115.1 4.98 s ? C2, C3, C55.06 s ? C2, C3, C5
2 146.9 –3 76.9 4.20 ov ? C1, C24 76.3 3.31 ov ? 1Glc
4.32 ov5 20.9 1.76 s ? C1, C2, C31Glc 105.7 4.26 d (J = 7.8 Hz) ? C4
⁄Only detected in Aegilops geniculata plants treated with specific allelochemicals.d = doublet, dd = doublet of doublets, m = multiplet, ov = overlapped, s = singlet, t = triplet.
B. D’Abrosca et al. / Phytochemistry 93 (2013) 27–40 31
The metabolome, indeed, provides a snapshot of the end results ofall processes in the cell from the transcriptional, translational andpost-translational cellular events, giving clues on the specific re-sponse of the system and defining the biochemical phenotype.
The hydroponic culture was employed because it appeared tobe the easiest to control, both in terms of media composition andof metabolite diffusion.
Plants of A. geniculata were treated with six compounds (jug-lone, coumarin, catechol, p-hydroxybenzoic acid, ferulic acid, andp-coumaric acid), known to possess allelopathic activity (Zhuo
Fig. 2. Heteronuclear Multiple Bond Correlation experiment of A. geniculata extract. DGlutamine; 5 Isoleucine; 6 Leucine; 7 Threonine; 8 Valine; 9 GABA; 10 cis-Aconitic Acid
and Yu, 2006). To the best of our knowledge, these allelochemicalshave never been tested on A. geniculata, nor have they been testedby means of the hydroponic culture method herewith described.
2.2.1. Treatment standardization using coumarinDuring the development of the plant, metabolic alterations hap-
pen continuously. In order to determine the optimum time to per-form treatments, plants were sequentially harvested from day 3(when the coleoptile was evident) to day 16; after day 17 hydro-ponic growth conditions caused damage to A. geniculata plants.
iagnostic correlations are marked. 1 Alanine; 2 Aspartic Acid; 3 Glutamic Acid; 4; 11 Citric Acid; 12 Malic Acid; 18 Oblongaroside A.
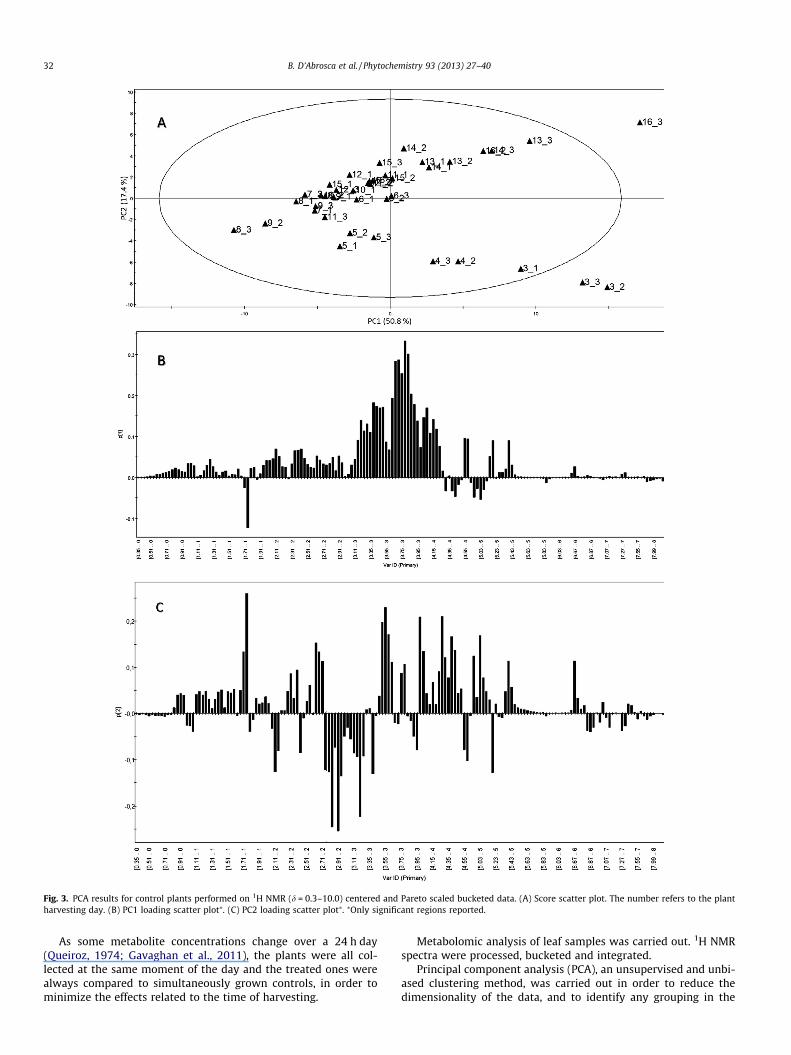
Fig. 3. PCA results for control plants performed on 1H NMR (d = 0.3–10.0) centered and Pareto scaled bucketed data. (A) Score scatter plot. The number refers to the plantharvesting day. (B) PC1 loading scatter plot⁄. (C) PC2 loading scatter plot⁄. ⁄Only significant regions reported.
32 B. D’Abrosca et al. / Phytochemistry 93 (2013) 27–40
As some metabolite concentrations change over a 24 h day(Queiroz, 1974; Gavaghan et al., 2011), the plants were all col-lected at the same moment of the day and the treated ones werealways compared to simultaneously grown controls, in order tominimize the effects related to the time of harvesting.
Metabolomic analysis of leaf samples was carried out. 1H NMRspectra were processed, bucketed and integrated.
Principal component analysis (PCA), an unsupervised and unbi-ased clustering method, was carried out in order to reduce thedimensionality of the data, and to identify any grouping in the
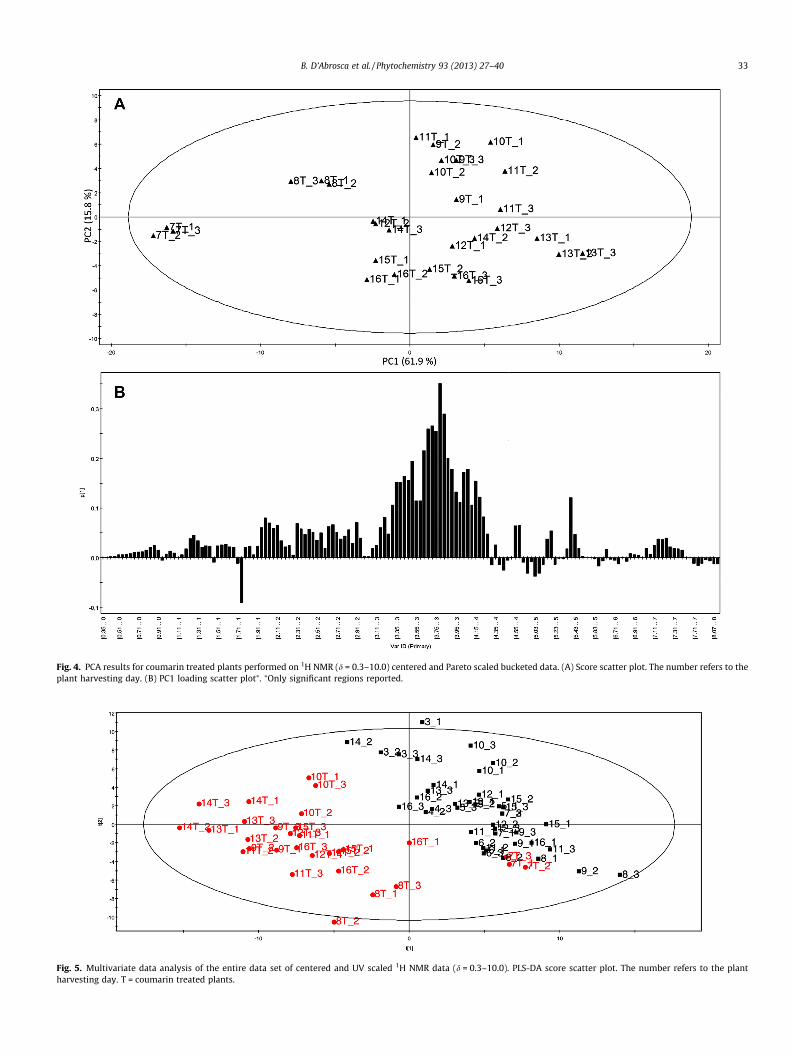
Fig. 4. PCA results for coumarin treated plants performed on 1H NMR (d = 0.3–10.0) centered and Pareto scaled bucketed data. (A) Score scatter plot. The number refers to theplant harvesting day. (B) PC1 loading scatter plot⁄. ⁄Only significant regions reported.
Fig. 5. Multivariate data analysis of the entire data set of centered and UV scaled 1H NMR data (d = 0.3–10.0). PLS-DA score scatter plot. The number refers to the plantharvesting day. T = coumarin treated plants.
B. D’Abrosca et al. / Phytochemistry 93 (2013) 27–40 33
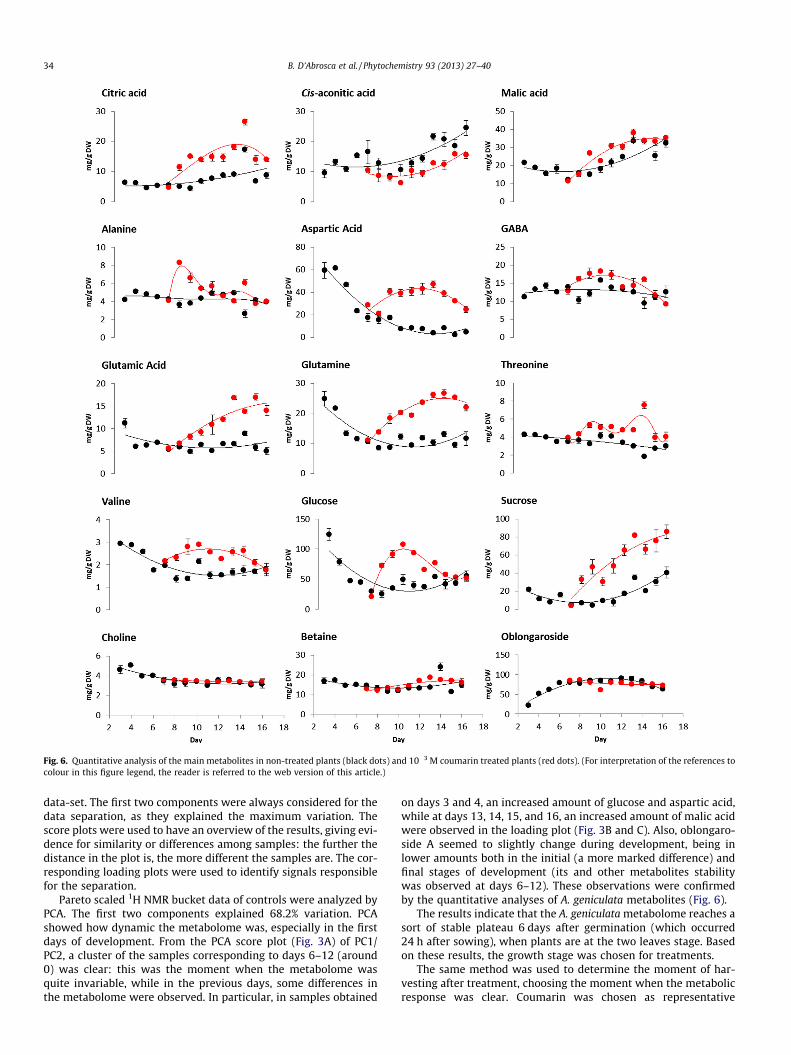
Fig. 6. Quantitative analysis of the main metabolites in non-treated plants (black dots) and 10�3 M coumarin treated plants (red dots). (For interpretation of the references tocolour in this figure legend, the reader is referred to the web version of this article.)
34 B. D’Abrosca et al. / Phytochemistry 93 (2013) 27–40
data-set. The first two components were always considered for thedata separation, as they explained the maximum variation. Thescore plots were used to have an overview of the results, giving evi-dence for similarity or differences among samples: the further thedistance in the plot is, the more different the samples are. The cor-responding loading plots were used to identify signals responsiblefor the separation.
Pareto scaled 1H NMR bucket data of controls were analyzed byPCA. The first two components explained 68.2% variation. PCAshowed how dynamic the metabolome was, especially in the firstdays of development. From the PCA score plot (Fig. 3A) of PC1/PC2, a cluster of the samples corresponding to days 6–12 (around0) was clear: this was the moment when the metabolome wasquite invariable, while in the previous days, some differences inthe metabolome were observed. In particular, in samples obtained
on days 3 and 4, an increased amount of glucose and aspartic acid,while at days 13, 14, 15, and 16, an increased amount of malic acidwere observed in the loading plot (Fig. 3B and C). Also, oblongaro-side A seemed to slightly change during development, being inlower amounts both in the initial (a more marked difference) andfinal stages of development (its and other metabolites stabilitywas observed at days 6–12). These observations were confirmedby the quantitative analyses of A. geniculata metabolites (Fig. 6).
The results indicate that the A. geniculata metabolome reaches asort of stable plateau 6 days after germination (which occurred24 h after sowing), when plants are at the two leaves stage. Basedon these results, the growth stage was chosen for treatments.
The same method was used to determine the moment of har-vesting after treatment, choosing the moment when the metabolicresponse was clear. Coumarin was chosen as representative
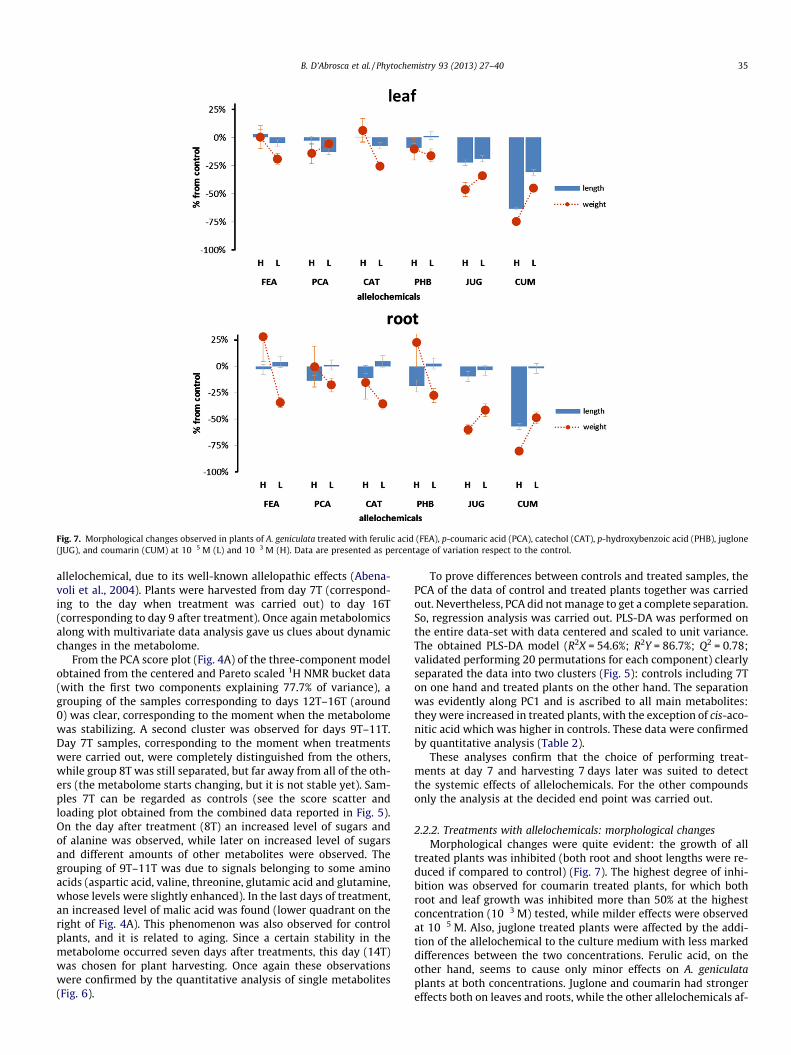
Fig. 7. Morphological changes observed in plants of A. geniculata treated with ferulic acid (FEA), p-coumaric acid (PCA), catechol (CAT), p-hydroxybenzoic acid (PHB), juglone(JUG), and coumarin (CUM) at 10�5 M (L) and 10�3 M (H). Data are presented as percentage of variation respect to the control.
B. D’Abrosca et al. / Phytochemistry 93 (2013) 27–40 35
allelochemical, due to its well-known allelopathic effects (Abena-voli et al., 2004). Plants were harvested from day 7T (correspond-ing to the day when treatment was carried out) to day 16T(corresponding to day 9 after treatment). Once again metabolomicsalong with multivariate data analysis gave us clues about dynamicchanges in the metabolome.
From the PCA score plot (Fig. 4A) of the three-component modelobtained from the centered and Pareto scaled 1H NMR bucket data(with the first two components explaining 77.7% of variance), agrouping of the samples corresponding to days 12T–16T (around0) was clear, corresponding to the moment when the metabolomewas stabilizing. A second cluster was observed for days 9T–11T.Day 7T samples, corresponding to the moment when treatmentswere carried out, were completely distinguished from the others,while group 8T was still separated, but far away from all of the oth-ers (the metabolome starts changing, but it is not stable yet). Sam-ples 7T can be regarded as controls (see the score scatter andloading plot obtained from the combined data reported in Fig. 5).On the day after treatment (8T) an increased level of sugars andof alanine was observed, while later on increased level of sugarsand different amounts of other metabolites were observed. Thegrouping of 9T–11T was due to signals belonging to some aminoacids (aspartic acid, valine, threonine, glutamic acid and glutamine,whose levels were slightly enhanced). In the last days of treatment,an increased level of malic acid was found (lower quadrant on theright of Fig. 4A). This phenomenon was also observed for controlplants, and it is related to aging. Since a certain stability in themetabolome occurred seven days after treatments, this day (14T)was chosen for plant harvesting. Once again these observationswere confirmed by the quantitative analysis of single metabolites(Fig. 6).
To prove differences between controls and treated samples, thePCA of the data of control and treated plants together was carriedout. Nevertheless, PCA did not manage to get a complete separation.So, regression analysis was carried out. PLS-DA was performed onthe entire data-set with data centered and scaled to unit variance.The obtained PLS-DA model (R2X = 54.6%; R2Y = 86.7%; Q2 = 0.78;validated performing 20 permutations for each component) clearlyseparated the data into two clusters (Fig. 5): controls including 7Ton one hand and treated plants on the other hand. The separationwas evidently along PC1 and is ascribed to all main metabolites:they were increased in treated plants, with the exception of cis-aco-nitic acid which was higher in controls. These data were confirmedby quantitative analysis (Table 2).
These analyses confirm that the choice of performing treat-ments at day 7 and harvesting 7 days later was suited to detectthe systemic effects of allelochemicals. For the other compoundsonly the analysis at the decided end point was carried out.
2.2.2. Treatments with allelochemicals: morphological changesMorphological changes were quite evident: the growth of all
treated plants was inhibited (both root and shoot lengths were re-duced if compared to control) (Fig. 7). The highest degree of inhi-bition was observed for coumarin treated plants, for which bothroot and leaf growth was inhibited more than 50% at the highestconcentration (10�3 M) tested, while milder effects were observedat 10�5 M. Also, juglone treated plants were affected by the addi-tion of the allelochemical to the culture medium with less markeddifferences between the two concentrations. Ferulic acid, on theother hand, seems to cause only minor effects on A. geniculataplants at both concentrations. Juglone and coumarin had strongereffects both on leaves and roots, while the other allelochemicals af-
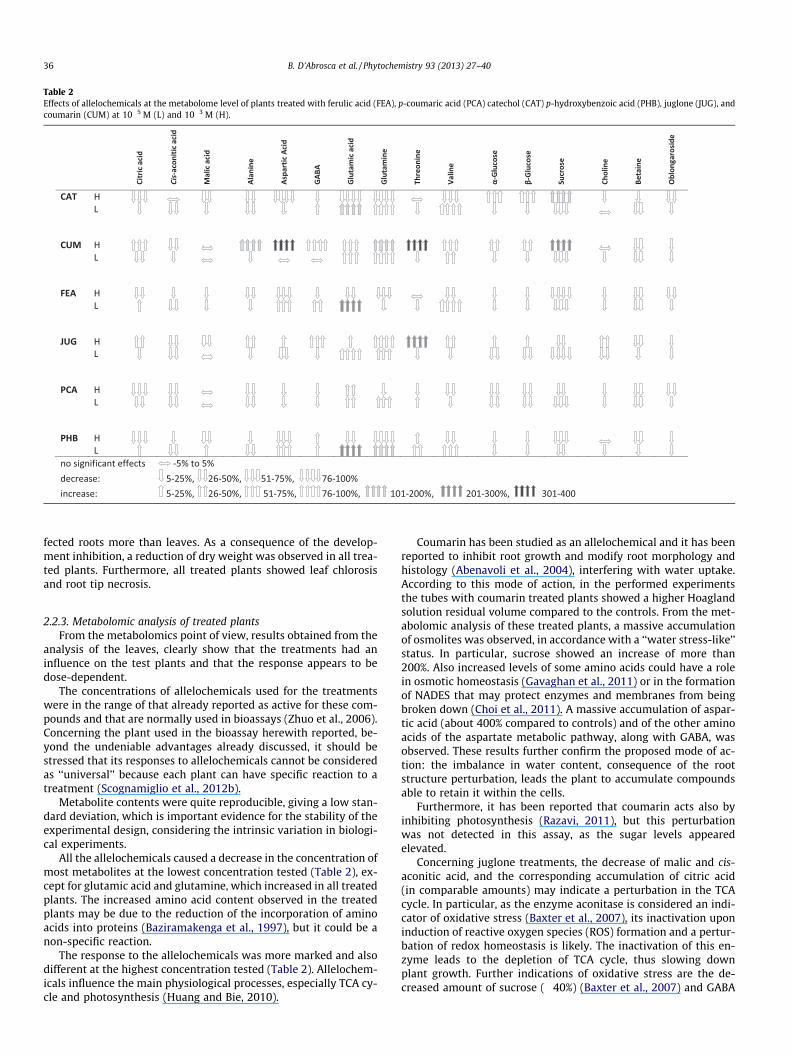
Table 2Effects of allelochemicals at the metabolome level of plants treated with ferulic acid (FEA), p-coumaric acid (PCA) catechol (CAT) p-hydroxybenzoic acid (PHB), juglone (JUG), andcoumarin (CUM) at 10�5 M (L) and 10�3 M (H).
36 B. D’Abrosca et al. / Phytochemistry 93 (2013) 27–40
fected roots more than leaves. As a consequence of the develop-ment inhibition, a reduction of dry weight was observed in all trea-ted plants. Furthermore, all treated plants showed leaf chlorosisand root tip necrosis.
2.2.3. Metabolomic analysis of treated plantsFrom the metabolomics point of view, results obtained from the
analysis of the leaves, clearly show that the treatments had aninfluence on the test plants and that the response appears to bedose-dependent.
The concentrations of allelochemicals used for the treatmentswere in the range of that already reported as active for these com-pounds and that are normally used in bioassays (Zhuo et al., 2006).Concerning the plant used in the bioassay herewith reported, be-yond the undeniable advantages already discussed, it should bestressed that its responses to allelochemicals cannot be consideredas ‘‘universal’’ because each plant can have specific reaction to atreatment (Scognamiglio et al., 2012b).
Metabolite contents were quite reproducible, giving a low stan-dard deviation, which is important evidence for the stability of theexperimental design, considering the intrinsic variation in biologi-cal experiments.
All the allelochemicals caused a decrease in the concentration ofmost metabolites at the lowest concentration tested (Table 2), ex-cept for glutamic acid and glutamine, which increased in all treatedplants. The increased amino acid content observed in the treatedplants may be due to the reduction of the incorporation of aminoacids into proteins (Baziramakenga et al., 1997), but it could be anon-specific reaction.
The response to the allelochemicals was more marked and alsodifferent at the highest concentration tested (Table 2). Allelochem-icals influence the main physiological processes, especially TCA cy-cle and photosynthesis (Huang and Bie, 2010).
Coumarin has been studied as an allelochemical and it has beenreported to inhibit root growth and modify root morphology andhistology (Abenavoli et al., 2004), interfering with water uptake.According to this mode of action, in the performed experimentsthe tubes with coumarin treated plants showed a higher Hoaglandsolution residual volume compared to the controls. From the met-abolomic analysis of these treated plants, a massive accumulationof osmolites was observed, in accordance with a ‘‘water stress-like’’status. In particular, sucrose showed an increase of more than200%. Also increased levels of some amino acids could have a rolein osmotic homeostasis (Gavaghan et al., 2011) or in the formationof NADES that may protect enzymes and membranes from beingbroken down (Choi et al., 2011). A massive accumulation of aspar-tic acid (about 400% compared to controls) and of the other aminoacids of the aspartate metabolic pathway, along with GABA, wasobserved. These results further confirm the proposed mode of ac-tion: the imbalance in water content, consequence of the rootstructure perturbation, leads the plant to accumulate compoundsable to retain it within the cells.
Furthermore, it has been reported that coumarin acts also byinhibiting photosynthesis (Razavi, 2011), but this perturbationwas not detected in this assay, as the sugar levels appearedelevated.
Concerning juglone treatments, the decrease of malic and cis-aconitic acid, and the corresponding accumulation of citric acid(in comparable amounts) may indicate a perturbation in the TCAcycle. In particular, as the enzyme aconitase is considered an indi-cator of oxidative stress (Baxter et al., 2007), its inactivation uponinduction of reactive oxygen species (ROS) formation and a pertur-bation of redox homeostasis is likely. The inactivation of this en-zyme leads to the depletion of TCA cycle, thus slowing downplant growth. Further indications of oxidative stress are the de-creased amount of sucrose (�40%) (Baxter et al., 2007) and GABA

B. D’Abrosca et al. / Phytochemistry 93 (2013) 27–40 37
accumulation (Bouché and Fromm, 2004). The latter phenomenonis a further prove of TCA cycle depletion, as it has been demon-strated that the GABA shunt operates as an alternative source ofmitochondrial succinate when necessary (Araújo et al., 2012).These observations are in good agreement with the reported modeof action of juglone. Indeed, Sytykiewicz (2011) provided strongmolecular evidence that the allelopathic effect of juglone ongrowth and development of plants can be ascribed to the inductionof oxidative stress, as treatments with juglone on maize seedlingsresulted in the over-expression of the glutathione transferase (themain enzyme involved in ROS detoxification) gene.
The accumulation of amino acids linked to TCA cycle, seems tobe inconsistent with this mechanism of action. They are usuallylowered in oxidative stress, with this mechanism being also a sur-vival strategy as the plant is trying to prevent the waste of energyderiving from the synthesis of new proteins that will be subjectedto immediate oxidative damage (Baxter et al., 2007). An explana-tion to this incongruity can be sought in protein degradation oftenoccurring after abiotic stress and resulting in increased levels offree amino acids (Cramer et al., 2011), more than in a de novosynthesis.
Concerning hydroxycinnamic acids, they seem to inhibit rootgrowth and to perform allelopathic interference by altering ion up-take and transport and reducing chlorophyll content. The contactof phenolic acids with the root cell membranes causes their depo-larization, an efflux of ions and a reduction of water and nutrientuptake (Hussain and Reigosa, 2011). Furthermore, many phenolicacids, like BOA, cinnamic acid, p-coumaric, caffeic, p-ferulic andsalicylic acids are reported to cause water stress in plants (Barko-sky and Einhelling, 2003). Cinnamic acids are also considered tohave an antiauxin activity (Gardner et al., 1994), therefore they ex-hibit inhibitory activity when applied exogenously, with the stron-gest effects exercised on roots (Jitareanu et al., 2011).
However the results of the present study did not show anymarked effect on plant growth for ferulic acid, even though somechanges were observed at the metabolome level. All the mainmetabolites showed more or less reduction compared to the con-trol. This is not surprising, as ferulic acid may influence the photo-synthesis rate, as reported for some cinnamic acids derivatives(Huang and Bie, 2010), with the slowing down of all the down-stream metabolic pathways. It exerts this action by decreasing
Fig. 8. PCA results for assays performed on 1H NMR (d = 0.3–10.0) centered and Paretocoumaric acid (PCA), catechol (CAT), p-hydroxybenzoic acid (PHB), juglone (JUG), and co
chlorophyll accumulation (Inderjit and Duke, 2003), as alreadystated.
An analogous mode of action can be ruled out for p-coumaricand p-hydroxybenzoic acid. Indeed, a similar metabolomic profilewas observed. Furthermore, p-coumaric acid has been reportedto act like ferulic acid by inhibiting chlorophyll accumulation andhence photosynthesis (Inderjit and Duke, 2003), while p-hydroxy-benzoic acid produces the same effect by decreasing stomatal con-ductance and inhibiting photosynthetic electron transport (Zhuoand Yu, 2006).
In catechol treated plants an accumulation of both glucose andsucrose was observed along with the decrease of the levels of allthe other metabolites. An inhibition of glycolysis and of the down-stream pathways or the inhibition of 2-oxoglutarate dehydroge-nase, whose effects are in agreement with those observed(Araújo et al., 2012), can be proposed.
It is worth to underline that even though each compound trig-gers different responses, there are also some similarities. The dif-ferent and specific responses of plants to various forms of stresswill use different metabolic pathways, but these are all part of alarge metabolic network in which everything is connected. As a re-sult there may be similarities in the response in certain centralparts of the network that needs to supply precursors for proteins,co-factors, energy and secondary metabolites (Cramer et al.,2011; Hirayama and Shinokazi, 2010).
For example, the decrease of malic acid levels observed in thejuglone, coumarin, catechol and ferulic acid treated plants maybe linked to the increased activity of NADP-dependent malic en-zyme observed in several abiotic stress (Fu et al., 2011). The roleof this C4 photosynthetic enzymes in C3 plants (as A. geniculata)has been recently reviewed (Doubnerová and Ryšlavá, 2011). Nonphotosynthetic phosphoenolpyruvate carboxylase (PEPC), NADP-malic enzyme (NADP-ME) and pyruvate phosphate dikinase(PPDK) play important roles in the maintenance of pH and replen-ishment of Krebs cycle intermediates, thereby contributing to thebiosynthesis of amino acids and other compounds, and providingNADPH for biosynthesis of damaged biomolecules. Enhanced activ-ity of these enzymes are found in C3 plants under various types ofstress.
Plants treated with p-coumaric acid and juglone showed anaccumulation of proline (not detected in controls), which was not
scaled bucketed data obtained from metabolomics analysis of ferulic acid (FEA), p-umarin (CUM) treated plants; B = control.

38 B. D’Abrosca et al. / Phytochemistry 93 (2013) 27–40
quantified because its signals were overlapped with others, but itwas identified thanks to HMBC correlations. Free proline accumu-lation seems to occur after exposure to a wide range of biotic andabiotic stresses, being a non-specific response in all kind of organ-isms (Choi et al., 2011; Hare and Cress, 1997), and it has been pro-ven to be involved in free radical scavenging processes (Yang et al.,2011).
Certainly, the results herewith discussed must be related tothe specific biological and analytical system: it is likely that otherplants react differently to each allelochemical. Hence, furtherstudies may take under consideration different target plants,with, for example, different life strategies. Moreover, the depen-dence of the response on the dose may be further explored usingdifferent concentrations but the aim of this study was the stan-dardization and validation of the use of metabolomics in combi-nation with the experimental design of the bioassay. It wasvery important to establish whether or not A. geniculata was sen-sitive to the allelochemicals and if the responses could be differ-entiated on the basis of the allelochemicals’ known mechanismsof action. Furthermore, it was important to determine if the met-abolomic changes were reproducible, and, more importantly,detectable by means of NMR.
Multivariate data analysis proved that the method is reliable. 1HNMR data were bucketed, and these data were centered and Paretoscaled and analyzed by PCA. The score plot of PC1/PC2 (with thetwo components explaining 72.1% of variation) of plants treatedwith the highest concentration of allelochemicals (Fig. 8A), con-firmed juglone and coumarin as the strongest allelopathic com-pounds, even though the other treated plants also differed fromthe control. The considerations drawn above were completed withthe data of the lowest concentration tested (Supplementary data),where no clear responses were observed.
The NMR-based metabolomics approach applied in this studyshowed promising results: allelopathy is studied not only at themorphological level but, most importantly, also at the biochemicallevel, giving insight in the metabolic pathways affected by allelo-chemicals, and thus in possible mechanisms of action on a sys-temic level.
3. Conclusions
Traditional approaches to study allelopathic interactions aredominated by bioassay-guided fractionation. Although this ap-proach has proven essential to uncover many chemically mediatedinteractions, it is time consuming and has other limitations.
The NMR-based metabolomic approach to study allelopathyherewith proposed offers the possibility to overcome several prob-lems correlated to this activity assessment. Metabolomic analysisof treated plants can be done in a relatively short time, allowinglarge number of samples to be analyzed and consequently morevariables can be studied at the same time. This is very useful bothin the identification of allelopathic compounds and in the elucida-tion of the mode of action of allelochemicals. Indeed the methodproved to be able to predict the mode of action of knownallelochemicals.
This approach may aid in the detection of certain modes of ac-tion by patterns of metabolic changes (biomarkers) and thus rap-idly seeing if active extracts represent a known mode of action ormay have a novel one. Also in case of synergy one may identifythe compounds in an extract that correlate with activity.
The last, but not least, important consideration, is that this ap-proach can be easily fitted to other plants, for example crop orweed species, and compare if similar metabolic changes occur.
All these aspects make the proposed approach, a powerful andversatile tool for studying allelopathic effects.
4. Experimental
4.1. Seed collection
Seeds of A. geniculata Roth. (Poaceae) were randomly collectedin June 2010 at ‘‘Castel Volturno’’ Nature Reserve, a flat coastal areain the north of Naples (Southern Italy). Seeds were stored at roomtemperature and in the darkness. Voucher specimens (CE0125)were deposited at the Herbarium of Department of Environmental,Biological and Pharmaceutical Sciences and Technologies of theSecond University of Naples. Yellow caryopses of A. geniculata wereselected according to Onnis et al. (1995) on the basis of their uni-formity, by observing them under a binocular microscope ZeissStemi 2000, by discarding the undersized and damaged ones. Fur-thermore, a preliminary test to evaluate the germinability (95% ofseeds) was carried out in a growth chamber Sanyo GallenkampMLR-350 at 28 �C in the dark.
4.2. Plant growth
Seeds were germinated in Petri dishes and transferred to thehydroponic system, after 24 h, made up by a tube filled with5 mL of Hoagland solution (KH2PO4 0.50 mM, K2HPO4 0.50 mM,K2SO4 1.25 mM, MgSO4 2.05 mM, CaCl2 1.00 mM, KNO3 5.00 mM,KCl 25.0 lM, H3BO3 12.5 lM, CuSO4 0.25 lM, ZnSO4 1.78 lM, Na2-
MoO4 82 nM, FeCl3 25 lM, Na2EDTA 28 lM) and with a 3 mm layerof perlite, in order to assure mechanical support. Plants wereplaced in a growth chamber with controlled temperature and rel-ative humidity (27 �C and 60% respectively), under photoperiod16:8 (light:darkness).
In order to choose treatment and harvesting times, untreatedplants were collected from day 3 to 16 (3–16), while treated plantswere collected from day 0 to 9 (7T–16T).
4.3. Plant treatments
Seven days after sowing, when plants were at the two leavesstage, they were treated with known allelochemicals: catechol,coumarin, p-coumaric acid, p-hydroxybenzoic acid, ferulic acidand juglone. They were added to the nutrient solution at 10�3
and 10�5 concentrations. Analyses were carried out in triplicates,each one consisting of ten seedlings. Blanks were performed, aswell. A week after treatment plants were harvested and root andshoot lengths were determined. Plants were immediately frozen,and then lyophilized. Metabolomic analysis of treated plant leaveswas carried out.
4.4. Extraction procedure for metabolomic analysis
Freeze-dried plant material (50 mg) was transferred to a 2 mLmicrotube. NMR samples were prepared in a mixture of phosphatebuffer (Fluka Chemika; 90 mM; pH 6.0) in D2O (Sigma–Aldrich)containing 0.1% w/w trimethylsilylpropionic-2,2,3,3-d4 acid so-dium salt (TMSP, Sigma–Aldrich) and methanol-d4 (Sigma–Al-drich). A volume of 1.5 mL of phosphate buffer in D2O andmethanol-d4 (1:1) was added to the plant samples. The mixturewas vortexed at room temperature for 1 min, ultrasonicated (Elma�
Transonic Digitals) for 40 min, and centrifuged (Beckman Allegra™64R) at 13,000 rpm for 10 min. An aliquot of 0.6 mL was transferredto an NMR tube and analyzed by NMR (Kim et al., 2010).
4.5. NMR experiments
NMR spectra were recorded at 25 �C on a 300.03 MHz for 1H and75.45 MHz for 13C on a Varian Mercury Plus 300 Fourier transformNMR. CD3OD was used as the internal lock.

B. D’Abrosca et al. / Phytochemistry 93 (2013) 27–40 39
Each 1H NMR spectrum consisted of 256 scans with the follow-ing parameters: 0.16 Hz/point, acquisition time (AQ) = 1.0 s, relax-ation delay (RD) = 1.5 s, 90� pulse width (PW) = 13.8 ls. Apresaturation sequence was used to suppress the residual H2O sig-nal. FIDs were Fourier transformed with LB = 0.3 Hz. The resultingspectra were manually phased and baseline-corrected and cali-brated to TMSP at 0.0 ppm.
1H–1H correlated spectroscopy (COSY), heteronuclear singlequantum coherence (HSQC) and heteronuclear multiple bond cor-relation (HMBC) spectra were recorded. COSY spectra were ac-quired with a 1.0 s relaxation delay and 2514 Hz spectral widthin both dimensions. The window function for COSY spectra wassine-bell (SSB = 0). HSQC and HMBC spectra were obtained with a1.0 s relaxation delay and 3140 Hz spectral width in f2 and18,116 Hz in f1. Qsine (SSB = 2.0) was used for the window func-tion of the HMBC. The optimized coupling constants were 140 Hzfor HSQC and 8 Hz for HMBC. FIDs were Fourier transformed andthe resulting spectra were manually phased and baseline-correctedand calibrated to TMSP at 0.0 ppm, using 1H NMR processor (ACDL-ABS 12.0).
In order to quantitate metabolites, specific signals (Table 1)were manually integrated and scaled to the internal standard sig-nal. The amount of each metabolite was calculated. Results wereexpressed as percentage from control.
4.6. Multivariate data analysis
1H NMR spectra were scaled to total intensity and the region be-tween d 0.31–9.99 was bucketed (Liland, 2011), reducing it to inte-gral segments with a width of 0.04 ppm with ACDLABS 12.0 1HNMR processor. The regions at d 4.59–4.91 and 3.30–3.38 were ex-cluded from the analysis because of solvents residual signals. Prin-cipal component analysis (PCA) and partial least-squares-discriminant analysis (PLS-DA) were performed with the SIMCA-P + software (version 12.0, Umetrics, Umeå Sweden) with scalingbased on Pareto or Unit Variance methods. PLS-DA models werevalidated by permutation test.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.phytochem.2013.03.016.
References
Abenavoli, M.R., Sorgonà, A., Albano, S., Cacco, G., 2004. Coumarin differentiallyaffects the morphology of different toot types of maize seedlings. J. Chem. Ecol.30, 1871–1883.
Ali, K., Maltese, F., Zyprian, E., Rex, M., Choi, Y.H., Verpoorte, R., 2009. NMRmetabolic fingerprinting based identification of grapevine metabolitesassociates with downy mildew resistance. J. Agric. Food Chem. 57, 9599–9606.
An, M., Prateley, J.E., Haig, T., 2001. Phytotoxicity of Vulpia residues. III: Biologicalactivity of identifies allelochemicals from Vulpia myuoros. J. Chem. Ecol. 27,383–394.
Araújo, W.L., Nunes-Nesi, A., Nikoloski, Z., Sweetlove, L.J., Fernie, A.R., 2012.Metabolic control and regulation of the tricarboxylic acid cycle inphotosynthetic and heterotrophic plant tissues. Plant, Cell Environ. 35, 1–21.
Barkosky, R.R., Einhelling, F.A., 2003. Allelopathic interference of plant–waterrelationships by para-hydroxybenzoic acid. Bot. Bull. Acad. Sin. 44, 53–58.
Baxter, C.J., Redestig, H., Schauer, N., Repsilber, D., Patil, K.R., Nielsen, J., Selbig, J., Liu,J., Fernie, A.R., Sweetlove, L., 2007. The metabolic response of heterotrophicArabidopsis cells to oxidative stress. Plant Physiol. 143, 312–325.
Baziramakenga, R., Leroux, G.D., Simard, R.R., Nadeau, P., 1997. Allelopathic effectsof phenolic acids on nucleic acid and protein levels in soybean seedlings. Can. J.Bot. 75, 445–450.
Bouché, N., Fromm, H., 2004. GABA in plants: just a metabolite? Trends Plant Sci. 9,110–115.
Broz, A.K., Corey, D.C., De-la-Peña, C., Lewis, M.R., Greene, E., Callaway, R.M.,Sumner, L., Vivanco, J.M., 2010. Plant neighbor identity influences plantbiochemistry and physiology relate to defense. BMC Plant Biol. 10, 115–129.
Broz, A.K., Manter, D.K., Callaway, R.M., Paschke, M.W., Vivanco, J.M., 2008. Amolecular approach to understanding plant–plant interactions in the context ofinvasion biology. Funct. Plant Biol. 35, 1123–1134.
Choi, Y.H., van Spronsen, J., Dai, Y., Verberne, M., Hollmann, F., Arends, I.W.C.E.,Witkamp, G.-J., Verpoorte, R., 2011. Are natural deep eutectic solvents themissing link in understanding cellular metabolism and physiology? PlantPhysiol. 156, 1701–1705.
Cramer, G.R., Urano, K., Delrot, S., Pezzotti, M., Shinozaki, K., 2011. Effects of abioticstress on plants: a systems biology perspective. Plant Biol. 11, 163–177.
Doubnerová, V., Ryšlavá, H., 2011. What can enzymes of C4 photosynthesis do for C3
plants under stress? Plant Sci. 180, 575–583.Esposito, A., Fiorentino, A., D’Abrosca, B., Izzo, A., Cefarelli, G., Golino, A., Monaco, P.,
2008. Potential allelopathic interference of Melilotus neapolitana on threecoexisting species of Mediterranean herbaceous plant community. J. PlantInteract. 3, 199–210.
Fu, Z.Y., Zhang, Z.B., Liu, Z.H., Xu, P., 2011. The effects of abiotic stresses on theNADP-dependent malic enzyme in the leaves of the hexaploid wheat. Biol.Plant. 55, 196–200.
Gardner, F.P., Pearce, R.B., Mitchell, R.L., 1994. Physiology of Crop Plants. Iowa StateUniversity Press, Ames.
Gavaghan, C.L., Li, J.V., Hadfield, S.T., Hole, S., Nicholson, J.K., Wilson, I.D., Howe,P.W.A., Stansley, P.D., Holmes, E., 2011. Application on NMR-basedmetabolomics to the investigation of salt stress in maize (Zea mays).Phytochem. Anal. 22, 214–224.
Gatti, A.B., Ferreira, A.G., Arduin, M., Gualtieri de Andrade Perez, S.C., 2010.Allelopathic effects of aqueous extracts of Artistolochia esperanzae O. Kuntze ondevelopment of Sesamum indicum L. seedlings. Acta Bot. Bras. 24, 454–461.
Hirayama, T., Shinokazi, K., 2010. Research on plant abiotic stress responses in thepost-genome era: past, present and future. Plant J. 61, 1041–1052.
Hare, P.D., Cress, W.A., 1997. Metabolic implications of stress-induced prolineaccumulation in plants. Plant Growth Regul. 21, 79–102.
Huang, X.X., Bie, Z.L., 2010. Cinnamic acid-inhibited ribulose-1,5-bisphosphatecarboxylase activity is mediated through decreased spermine and changes inthe ratio of polyamines in cowpea. J. Plant Physiol. 167, 47–53.
Hussain, M.I., Reigosa, M.J., 2011. Allelochemical stress inhibits growth, leaf waterrelations, PSII photochemistry, non-photochemical fluorescence quenching, andheat energy dissipation in three C3 perennial species. J. Exp. Bot. 62,4533–4545.
Inderjit, Duke, S.O., 2003. Ecophysiological aspects of allelopathy. Planta 217, 529–539.
Jitareanu, A., Tataringa, G., Zbancioc, A.-M., Stanescu, U., 2011. Toxicity of somecinnamic acid derivatives to common bean (Phaseolus vulgaris). Not. Bot. HortiAgrobo. 39, 130–134.
Keurentjes, J.J.B., Angenent, G.C., Dicke, M., Martins Dos Santos, V.A.P., Molenaar, J.,van der Putten, W.H., de Ruiter, P.C., Struik, P.C., Thomma, B.P.H.J., 2011.Redefining plant systems biology: from cell to ecosystem. Trends Plant Sci. 16,183–190.
Kim, H.K., Choi, Y.H., Verpoorte, R., 2010. NMR-based metabolomic analysis ofplants. Nat. Protoc. 5, 536–549.
Kim, H.K., Choi, Y.H., Verpoorte, R., 2011. NMR-based plant metabolomics: where dowe stand, where do we go? Trends Biotechnol. 29, 267–275.
Liland, K.H., 2011. Multivariate methods in metabolomics – from pre-processing todimension reduction and statistical analysis. Trends Anal. Chem. 30, 827–841.
Liu, N.Q., Cao, M., Frèdèrich, M., Choi, Y.H., Verpoorte, R., vad der Kooy, F., 2010.Metabolomic investigation of the ethnopharmacological use of Artemisia afrawith NMR spectroscopy and multivariate data analysis. J. Ethnopharmacol. 128,230–235.
López-Gresa, M.P., Maltese, F., Bellès, J.M., Conejero, V., Kin, H.K., Choi, Y.H.,Verpoorte, R., 2010. Metabolic response of tomato leaves upon different plant–pathogen interactions. Phytochem. Anal. 21, 89–94.
Macías, F.A., Castellano, D., Molinillo, J.M.G., 2000. Search for a standard phytotoxicbioassay for allelochemicals. Selection of standard target species. J. Agric. FoodChem. 48, 2512–2521.
Mguis, K., Albouchi, A., Khadhri, A., Abassi, M., Yakoubi-Tej, M., Mahjoub, A.,Ouerghi, Z., Brahim, N.B., 2012. Adjustments in leaf water relations of wildwheat relative Aegilops geniculata Roth. and wheat (Triticum durum Desf.) plantsgrown in a salinity gradient. Aust. J. Crop Sci. 6, 768–776.
Naveh, Z., 1989. Fire in the Mediterranean – a landscape ecological perspective. In:Goldammer, J.G., Jenkis, M.J. (Eds.), Fire in the Ecosystems Dynamics. SPBAcademic Publishing, The Hague, pp. 1–20.
Onnis, A., Bertacchi, A., Lombardi, T., Stefani, A., 1995. Morphology and germinationof yellow and brown caryopses of Aegilops geniculata Roth. (Graminae)population from Italy. Giorn. Bot. Ital. 129, 813–821.
Ouyang, M.-A., Wang, C.-Z., Wang, S.-B., 2007. Water soluble constituents of Ilexoblonga. J. Asian Nat. Prod. Res. 9, 399–405.
Pradhan, G.P., Prasad, P.V.V., Fritz, A.K., Kirkham, M.B., Gill, B.S., 2012. Response ofAegilops species to drought stress during reproductive stages of development.Funct. Plant Biol. 39, 51–59.
Pignatti, S., 1982. Flora d’Italia. Bologna Edagricole, vol. 1. p. 15.Queiroz, O., 1974. Circadian rhythms and metabolic patterns. Annu. Rev. Plant
Physiol. 25, 115–134.Rinu, G., Mur, L.A.J., Jones, D.G., 2005. Advancing our understanding of allelopathy
through metabolomics. In: Proceeding of the 4th World Congress onAllelopathy, ‘‘Establishing the Scientific Base’’, pp. 197–200.
Razavi, S.M., 2011. Plant coumarins as allelopathic agents. Int. J. Biol. Chem. 5, 86–90.

40 B. D’Abrosca et al. / Phytochemistry 93 (2013) 27–40
Scognamiglio, M., D’Abrosca, B., Fiumano, V., Chambery, A., Severino, V., Tsafantakis,N., Pacifico, S., Esposito, A., Fiorentino, A., 2012a. Oleanane saponins from Bellissylvestris Cyr. and evaluation of their phytotoxicity on Aegilops geniculata Roth.Phytochemistry 84, 125–134.
Scognamiglio, M., Esposito, A., D’Abrosca, B., Pacifico, S., Fiumano, V., Tsafantakis, N.,Monaco, P., Fiorentino, A., 2012b. Isolation, distribution and allelopathic effectof caffeic acid derivatives from Bellis perennis L. Biochem. Syst. Ecol. 43, 108–113.
Sytykiewicz, H., 2011. Expression patterns of Glutathione Transferase gene (GstI) inmaize seedlings under juglone-induced oxidative stress. Int. J. Mol. Sci. 12,7982–7995.
Verpoorte, R., Choi, Y.H., Kim, H.K., 2007. NMR-based metabolomics at work inphytochemistry. Phytochem. Rev. 6, 3–14.
Yang, Y., Zhang, Y., Wei, X., You, J., Wang, W., Lu, J., Shi, R., 2011. Comparativeantioxidative responses and proline metabolism in two wheat cultivars undershort term lead stress. Ecotoxicol. Environ. Saf. 74, 733–740.
Yuliana, N.D., Khatib, A., Choi, Y.H., Verpoorte, R., 2011. Metabolomics for bioactivityassessment of natural products. Phytother. Res. 25, 157–169.
Zhuo, Y.H., Yu, J.Q., 2006. Allelochemicals and photosynthesis. In: Reigosa, M.J.,Pedrol, N., González, L. (Eds.), Allelopathy: A Physiological Process withEcological Implications. Kluwer Academic Publishers, Netherlands, pp. 127–139.
Related Documents











