* Corresponding author. Fax: 001-803-777-6610. E-mail address: thunell@geol.sc.edu (R.C. Thunell) Deep-Sea Research II 47 (2000) 1157}1176 Planktonic foraminiferal #uxes in the Santa Barbara Basin: response to seasonal and interannual hydrographic changes Eileen Kincaid!, Robert C. Thunell!,*, J. Le!, Carina B. Lange", Amy L. Weinheimer", Freda M.H. Reid" !Department of Geological Sciences, University of South Carolina, Columbia, SC 29208, USA "Scripps Institution of Oceanography, University of California, San Diego, 9500 Gilman Drive, La Jolla, CA 92093-0215, USA Received 16 June 1997; received in revised form 26 May 1998; accepted 16 June 1999 Abstract Results of a three-year time-series sediment trap experiment in the Santa Barbara Basin indicate linkages between water-column dynamics and variations in plankton #uxes on both seasonal and interannual time scales. The two-week collection interval utilized for this study allows evaluation of high-frequency changes in the production and #ux of planktonic foraminifera, which are compared to those of siliceous microorganisms (diatoms, radiolaria and silico#agellates). Annual hydrographic conditions in Santa Barbara Basin are in#uenced by seasonal changes in insolation, the wind "eld and source waters. This study includes periods of weak to moderate El Nin 8 o conditions followed by a transitional period to a non-El Nin 8 o state. Planktonic foraminiferal #uxes and species turnover are related to seasonal and interannual changes in source waters and water column conditions. Globigerina bulloides, G. quinqueloba and Neogloboquadrina pachyderma (dextral) dominate the planktonic foraminiferal population in this location and are present throughout the year. Globigerina bulloides and G. quinqueloba have similar #ux patterns, and highest #uxes occur in association with diatom blooms. The #ux patterns of G. bulloides and G. quinqueloba in general are inversely related to that of N. pachyderma (dextral). Globigerina quinqueloba seems to respond earlier to the onset of upwelling than G. bulloides. Three temperature-limited species, Neogloboquadrina dutertrei, Orbulina universa and Globigerinoides ruber, are present in lower numbers and show more restricted seasonal ranges. In the thermal regime of the Santa Barbara Basin, N. dutertrei, like O. universa, disappears during upwelling. Neogloboquadrina dutertrei occurs in highest abund- ance when the water column is thermally strati"ed and there is a well developed deep 0967-0645/00/$ - see front matter ( 2000 Elsevier Science Ltd. All rights reserved. PII: S 0 9 6 7 - 0 6 4 5 ( 9 9 ) 0 0 1 4 0 - X

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

*Corresponding author. Fax: 001-803-777-6610.E-mail address: [email protected] (R.C. Thunell)
Deep-Sea Research II 47 (2000) 1157}1176
Planktonic foraminiferal #uxes in theSanta Barbara Basin: response to seasonal
and interannual hydrographic changes
Eileen Kincaid!, Robert C. Thunell!,*, J. Le!, Carina B. Lange",Amy L. Weinheimer", Freda M.H. Reid"
!Department of Geological Sciences, University of South Carolina, Columbia, SC 29208, USA"Scripps Institution of Oceanography, University of California, San Diego, 9500 Gilman Drive,
La Jolla, CA 92093-0215, USA
Received 16 June 1997; received in revised form 26 May 1998; accepted 16 June 1999
Abstract
Results of a three-year time-series sediment trap experiment in the Santa Barbara Basinindicate linkages between water-column dynamics and variations in plankton #uxes on bothseasonal and interannual time scales. The two-week collection interval utilized for thisstudy allows evaluation of high-frequency changes in the production and #ux of planktonicforaminifera, which are compared to those of siliceous microorganisms (diatoms, radiolaria andsilico#agellates). Annual hydrographic conditions in Santa Barbara Basin are in#uenced byseasonal changes in insolation, the wind "eld and source waters. This study includes periods ofweak to moderate El Nin8 o conditions followed by a transitional period to a non-El Nin8 o state.Planktonic foraminiferal #uxes and species turnover are related to seasonal and interannualchanges in source waters and water column conditions. Globigerina bulloides, G. quinquelobaand Neogloboquadrina pachyderma (dextral) dominate the planktonic foraminiferal populationin this location and are present throughout the year. Globigerina bulloides and G. quinquelobahave similar #ux patterns, and highest #uxes occur in association with diatom blooms. The #uxpatterns of G. bulloides and G. quinqueloba in general are inversely related to that ofN. pachyderma (dextral). Globigerina quinqueloba seems to respond earlier to the onset ofupwelling than G. bulloides. Three temperature-limited species, Neogloboquadrina dutertrei,Orbulina universa and Globigerinoides ruber, are present in lower numbers and show morerestricted seasonal ranges. In the thermal regime of the Santa Barbara Basin, N. dutertrei, likeO. universa, disappears during upwelling. Neogloboquadrina dutertrei occurs in highest abund-ance when the water column is thermally strati"ed and there is a well developed deep
0967-0645/00/$ - see front matter ( 2000 Elsevier Science Ltd. All rights reserved.PII: S 0 9 6 7 - 0 6 4 5 ( 9 9 ) 0 0 1 4 0 - X

chlorophyll maximum. Increased #uxes of Globigerinoi ruber the most thermally restricted ofthe dominant species in the basin, are associated with the presence of warm oceanic diatomsand warm-water radiolarians. Total planktonic foraminiferal #uxes were lower in 1995 thanduring the previous two years, which were characterized by El Nin8 o conditions. ( 2000Elsevier Science Ltd. All rights reserved.
1. Introduction
Deciphering past oceanographic conditions is dependent on our understanding ofthe factors that drive changes in the proxies utilized in making these reconstructions.Species abundance data of marine plankton preserved in sediments are commonlyused to reconstruct water-column conditions through geologic time. For this reason,it is critical that we identify the basic ecology and environmental conditions asso-ciated with paleoceanographically important groups of plankton.
We can assume that a number of factors control the abundance and composition ofa plankton population at a given time or in a given area. For example, for a plank-tonic foraminiferal species to #ourish, conditions must fall within the salinity andtemperature range tolerated by the individual species, su$cient nutrition should beobtainable, and, in the case of those species containing photosynthetic endosym-bionts, su$cient light must be available, and predation should not be excessive.Laboratory culture studies designed to determine the e!ect of environmental controlson the growth of individual species of planktonic foraminifera (Bijma et al., 1990b;Hemleben et al., 1989; Spero, 1987; Spero and Angel, 1991; Spero and DeNiro, 1987)and "eld studies undertaken to ascertain their depth habitat and seasonal range(BeH , 1977; BeH and Hutson, 1977; Bouvier-Soumagnac and Duplessy, 1985; Deuser,1986,1987; Deuser and Ross, 1989; Fairbanks et al., 1982; Fairbanks and Wiebe, 1980;Ortiz et al., 1995; Sautter and Sancetta, 1992; Sautter and Thunell, 1991b; Thunell andSautter, 1992) have removed some of the uncertainty inherent in the informationobtained from fossil planktonic foraminiferal assemblages.
The Santa Barbara Basin provides an ideal location to assess the reliability ofmethods utilized in paleoclimate studies. Anoxic bottom waters exist due to highlevels of primary productivity and restricted bottom water circulation. The combina-tion of seasonally varying sediment inputs and the lack of bioturbation results in thedeposition of varved sediments in the basin (Hulsemann and Emery, 1961; Soutar andCrill, 1977; Thunell et al., 1995; Thunell, 1998). The shallow depth of the basin (600 mmaximum depth) combined with anoxic bottom waters eliminate selective dissolutionas a major factor in altering the planktonic foraminiferal assemblages accumulating inthe sediments (Berger and Soutar, 1970). These varved sediments contain a high-resolution record of changes in upper water-column conditions. Data from varvedsediments have been used to reconstruct climate change during the Holocene(Heusser, 1978; Kennedy and Brassell, 1992; Lange et al., 1987; Pisias, 1978; Schim-melmann et al., 1990) and late Pleistocene (Behl and Kennett, 1996; Kennett andIngram, 1995). On shorter time periods, changes in siliceous and calcareous plankton
1158 E. Kincaid et al. / Deep-Sea Research II 47 (2000) 1157}1176

have been used to study the history of El Nin8 o events in the eastern Paci"c Ocean(Lange et al., 1987; Weinheimer et al., 1986; Weinheimer and Cayan, 1997).
The Santa Barbara Basin is strongly a!ected by both the California Current, theeastern boundary current of the North Paci"c Ocean, and quasi-periodic changesin the eastern tropical Paci"c due to El Nin8 o/Southern Oscillation (ENSO). Forexample, well-documented hydrographic changes occurred in the basin during the1983 El Nin8 o, including a deepening of the thermocline, warming of surface waters,increased shoreward transport of California Current water, reduced nutrient concen-trations, and increased oxygenation of bottom water (McGowan, 1985). Determininghow plankton respond to such conditions may enable us to identify proxies that canbe used to extract such signals from the sediment record. An understanding ofwater-column processes and the manner in which this information is recorded in thesediments will allow an assessment of past changes in the eastern North Paci"c.
In this paper we present planktonic foraminiferal #ux data from a three-yeartime-series sediment trap experiment in the Santa Barbara Basin and compare themto the #ux changes in siliceous microorganisms (diatoms, radiolaria and silico#agel-lates) (Lange et al., 1997). Flux data were collected at two-week intervals andcorrelated with hydrographic data from CTD casts collected at the trap site in orderto document the plankton response to physical forcing.
2. Methods
Particle #ux samples were obtained using a McLean Mark VII-W automatedsediment trap deployed in the center of the basin (34314@N, 120302@W; &590 m waterdepth) and positioned approximately 60 m above the sea#oor (Fig. 1). The trap wasrecovered and redeployed every six months and samples were collected continuouslyfor 14-day intervals. Sample bottles were "lled with a bu!ered seawater-sodium azidesolution (sodium azide serves as a poison) prior to deployment. In this paper wereport on samples collected between August 1993 and December 1995 (siliceousplankton #ux) and August 1993 and September 1996 (planktonic foraminiferal #ux).Data are missing for the periods from 1 November 1994 through 20 February 1995and 16 May 1995 through 25 August 1995 due to trap failure. For the period from 26August 1995 through 22 February 1996, the sample cups were poisoned with insu$-ciently bu!ered formalin, which caused some dissolution of the foraminifera. There-fore, the foraminiferal #ux data for this period are not reported.
Hydrographic data (conductivity, temperature and density) were collected at thetrap site at two- to four-week intervals (weather dependent) using a Sea Bird Seacat 19Pro"ler "tted with an oxygen sensor and transmissometer.
All sediment trap samples were split using a McLean four-head, rotary samplesplitter. Foraminiferal #ux assessments were conducted on 1/16th splits of the totalmaterial for each collection interval. Samples were washed with bu!ered (sodiumborate) distilled water through a 125 lm sieve. Material from the "rst two trapdeployments was soaked for 1 h in a Calgon solution to disaggregate the sample,placed on "lter paper and dried before the foraminifera were picked from the sample.
E. Kincaid et al. / Deep-Sea Research II 47 (2000) 1157}1176 1159
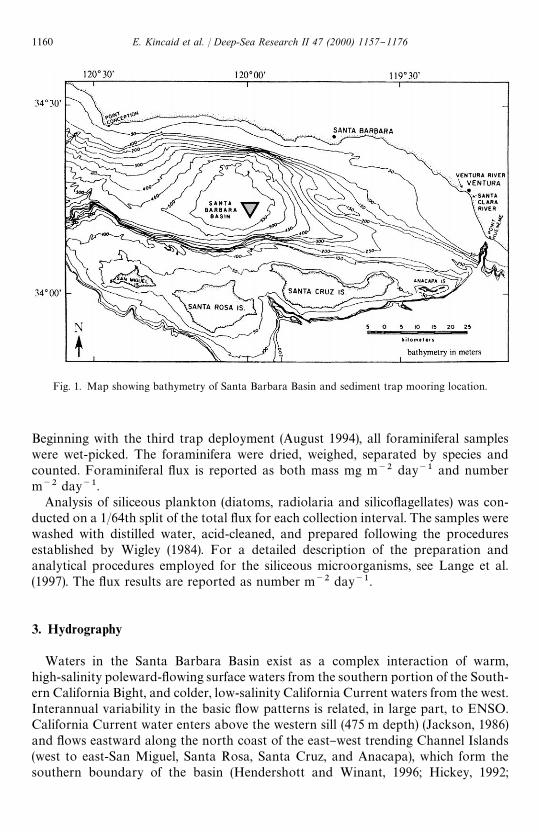
Fig. 1. Map showing bathymetry of Santa Barbara Basin and sediment trap mooring location.
Beginning with the third trap deployment (August 1994), all foraminiferal sampleswere wet-picked. The foraminifera were dried, weighed, separated by species andcounted. Foraminiferal #ux is reported as both mass mg m~2 day~1 and numberm~2 day~1.
Analysis of siliceous plankton (diatoms, radiolaria and silico#agellates) was con-ducted on a 1/64th split of the total #ux for each collection interval. The samples werewashed with distilled water, acid-cleaned, and prepared following the proceduresestablished by Wigley (1984). For a detailed description of the preparation andanalytical procedures employed for the siliceous microorganisms, see Lange et al.(1997). The #ux results are reported as number m~2 day~1.
3. Hydrography
Waters in the Santa Barbara Basin exist as a complex interaction of warm,high-salinity poleward-#owing surface waters from the southern portion of the South-ern California Bight, and colder, low-salinity California Current waters from the west.Interannual variability in the basic #ow patterns is related, in large part, to ENSO.California Current water enters above the western sill (475 m depth) (Jackson, 1986)and #ows eastward along the north coast of the east}west trending Channel Islands(west to east-San Miguel, Santa Rosa, Santa Cruz, and Anacapa), which form thesouthern boundary of the basin (Hendershott and Winant, 1996; Hickey, 1992;
1160 E. Kincaid et al. / Deep-Sea Research II 47 (2000) 1157}1176
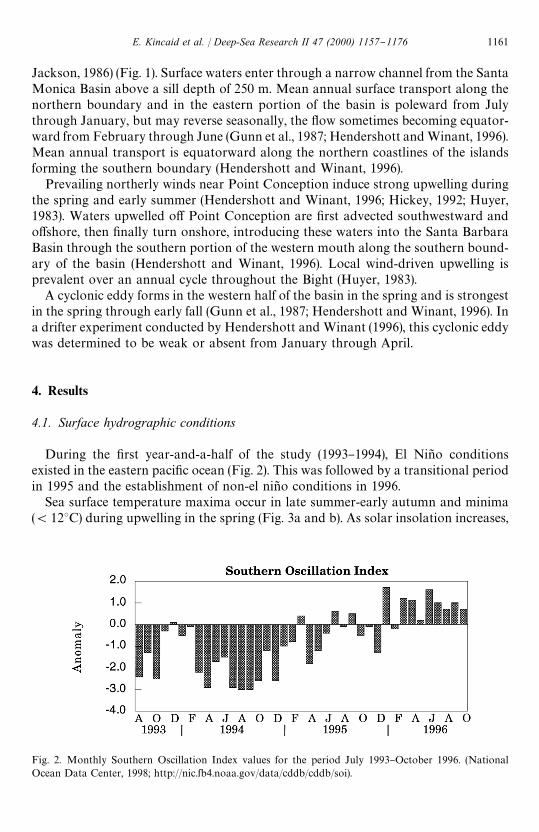
Fig. 2. Monthly Southern Oscillation Index values for the period July 1993}October 1996. (NationalOcean Data Center, 1998; http://nic.fb4.noaa.gov/data/cddb/cddb/soi).
Jackson, 1986) (Fig. 1). Surface waters enter through a narrow channel from the SantaMonica Basin above a sill depth of 250 m. Mean annual surface transport along thenorthern boundary and in the eastern portion of the basin is poleward from Julythrough January, but may reverse seasonally, the #ow sometimes becoming equator-ward from February through June (Gunn et al., 1987; Hendershott and Winant, 1996).Mean annual transport is equatorward along the northern coastlines of the islandsforming the southern boundary (Hendershott and Winant, 1996).
Prevailing northerly winds near Point Conception induce strong upwelling duringthe spring and early summer (Hendershott and Winant, 1996; Hickey, 1992; Huyer,1983). Waters upwelled o! Point Conception are "rst advected southwestward ando!shore, then "nally turn onshore, introducing these waters into the Santa BarbaraBasin through the southern portion of the western mouth along the southern bound-ary of the basin (Hendershott and Winant, 1996). Local wind-driven upwelling isprevalent over an annual cycle throughout the Bight (Huyer, 1983).
A cyclonic eddy forms in the western half of the basin in the spring and is strongestin the spring through early fall (Gunn et al., 1987; Hendershott and Winant, 1996). Ina drifter experiment conducted by Hendershott and Winant (1996), this cyclonic eddywas determined to be weak or absent from January through April.
4. Results
4.1. Surface hydrographic conditions
During the "rst year-and-a-half of the study (1993}1994), El Nin8 o conditionsexisted in the eastern paci"c ocean (Fig. 2). This was followed by a transitional periodin 1995 and the establishment of non-el nin8 o conditions in 1996.
Sea surface temperature maxima occur in late summer-early autumn and minima((123C) during upwelling in the spring (Fig. 3a and b). As solar insolation increases,
E. Kincaid et al. / Deep-Sea Research II 47 (2000) 1157}1176 1161
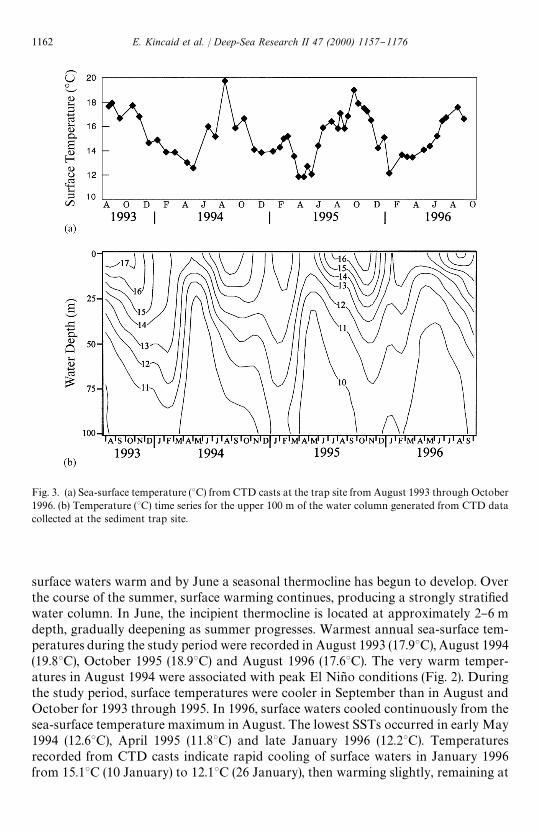
Fig. 3. (a) Sea-surface temperature (3C) from CTD casts at the trap site from August 1993 through October1996. (b) Temperature (3C) time series for the upper 100 m of the water column generated from CTD datacollected at the sediment trap site.
surface waters warm and by June a seasonal thermocline has begun to develop. Overthe course of the summer, surface warming continues, producing a strongly strati"edwater column. In June, the incipient thermocline is located at approximately 2}6 mdepth, gradually deepening as summer progresses. Warmest annual sea-surface tem-peratures during the study period were recorded in August 1993 (17.93C), August 1994(19.83C), October 1995 (18.93C) and August 1996 (17.63C). The very warm temper-atures in August 1994 were associated with peak El Nin8 o conditions (Fig. 2). Duringthe study period, surface temperatures were cooler in September than in August andOctober for 1993 through 1995. In 1996, surface waters cooled continuously from thesea-surface temperature maximum in August. The lowest SSTs occurred in early May1994 (12.63C), April 1995 (11.83C) and late January 1996 (12.23C). Temperaturesrecorded from CTD casts indicate rapid cooling of surface waters in January 1996from 15.13C (10 January) to 12.13C (26 January), then warming slightly, remaining at
1162 E. Kincaid et al. / Deep-Sea Research II 47 (2000) 1157}1176
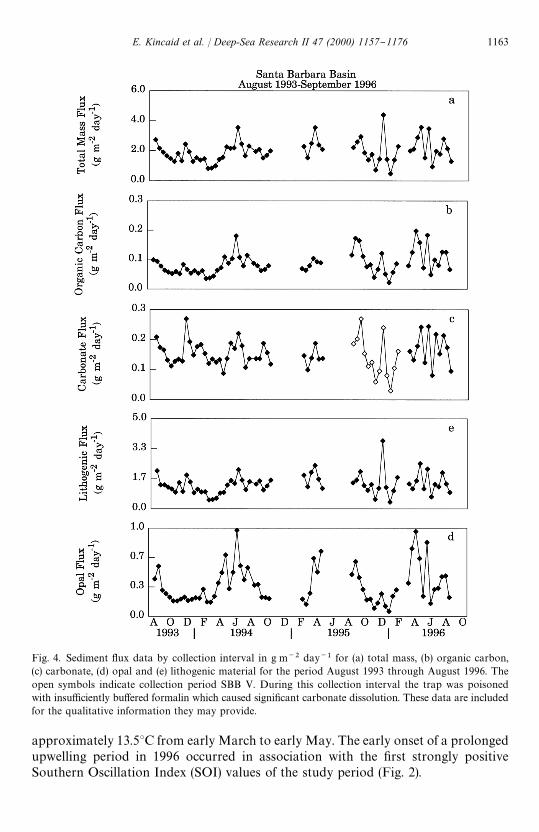
Fig. 4. Sediment #ux data by collection interval in g m~2 day~1 for (a) total mass, (b) organic carbon,(c) carbonate, (d) opal and (e) lithogenic material for the period August 1993 through August 1996. Theopen symbols indicate collection period SBB V. During this collection interval the trap was poisonedwith insu$ciently bu!ered formalin which caused signi"cant carbonate dissolution. These data are includedfor the qualitative information they may provide.
approximately 13.53C from early March to early May. The early onset of a prolongedupwelling period in 1996 occurred in association with the "rst strongly positiveSouthern Oscillation Index (SOI) values of the study period (Fig. 2).
E. Kincaid et al. / Deep-Sea Research II 47 (2000) 1157}1176 1163

4.2. Total particulate yux
The total particulate #ux in the Santa Barbara Basin during this three-year periodis discussed in detail by Thunell (1998). The patterns observed in particulate #ux areclosely related to seasonal hydrographic variability in the upper water column. Twodistinct #ux maxima occur over an annual cycle (Fig. 4a}e) and this pattern is closelymimicked by the lithogenic, organic carbon, and carbonate #uxes. The total mass #uxis dominated by the lithogenic component throughout the year (40}80%). Highterrigenous #ux during the fall and winter is due to increased riverine input associatedwith the rainy season (Soutar and Crill, 1977; Thunell, 1998; Thunell et al., 1995).Synchronous increases in the carbonate, lithogenic and organic carbon #uxes, espe-cially during the winter, may be related either to (1) riverine input of terrestrialcarbonate and organic matter, (2) increased productivity due to the input of dissolvednutrients in river discharge, or (3) increased productivity related to nutrients incorpor-ated from depth as thermal strati"cation breaks down and the increased wind stress ofwinter storms increases the depth of the mixed layer. The latter two processes alsoresult in increased scavenging of lithogenic material from the water column by organicaggregates. The second peak in both total #ux and the #ux of biogenic material occursin the spring with the onset of upwelling. This is probably due to the scavenging of "neparticles from the water column by organic aggregates during this period of highproductivity (Thunell et al., 1995). The biogenic opal #ux is highest in spring andsummer, low in fall and winter, and dominates the annual biogenic sediment #ux(up to 35% of the total #ux; Thunell, 1998).
4.3. Foraminiferal yux and species composition
The annual pattern of total planktonic foraminiferal #ux (Fig. 5a and b) re#ectsseasonal hydrographic variability, overprinted by the e!ects of both the reproductivecycle for individual species and interannual changes related to El Nin8 o. The reproduc-tive cycle for most planktonic foraminifera is semi-lunar (Globigerinoides ruber) orlunar (O. universa) (Bijma et al., 1990a; Hemleben et al., 1989). We speculate that thehigh-frequency variability in foraminiferal #ux is related to these reproductive cycles.During 1993}1994, maximum foraminiferal #uxes occurred during August whensea-surface temperatures were elevated and the water column was highly strati"ed. In1996, maximum #uxes occurred in the late spring during the latter part of the springupwelling event, followed by a smaller peak in September. The planktonicforaminiferal population was signi"cantly reduced during the 1995 transitional period(Table 1). Globigerina bulloides, N. pachyderma (dextral) and G. quinqueloba arepresent throughout the year and dominate the planktonic foraminiferal assemblage inthis area (Fig. 6a}c). Globigerina bulloides and G. quinqueloba (Fig. 6a and b) havesimilar seasonal #ux patterns but the magnitudes of the #uxes vary. Globigerinabulloides (Fig. 6a) #ux maxima occurred in August 1993 and May 1996, with addi-tional peaks in December 1993, June}August 1994 and a smaller peak in lateFebruary}March 1994. The percentage of the total foraminiferal #ux contributed by
1164 E. Kincaid et al. / Deep-Sea Research II 47 (2000) 1157}1176
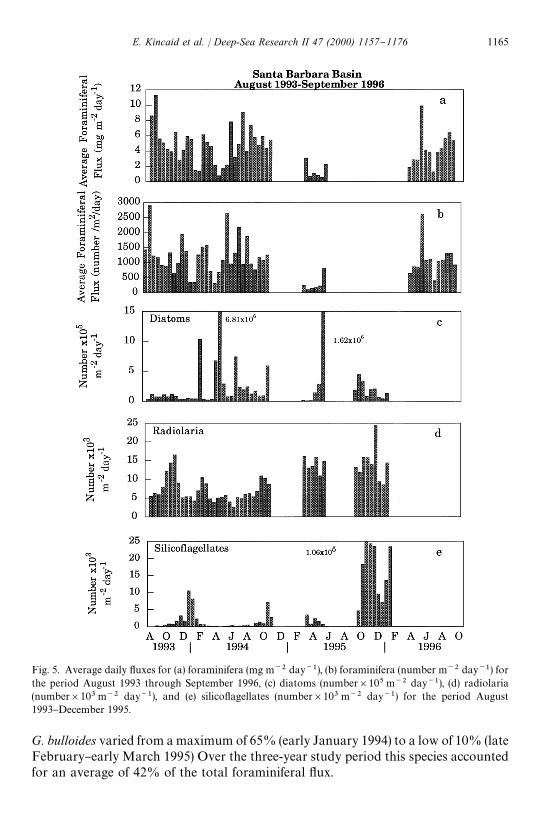
Fig. 5. Average daily #uxes for (a) foraminifera (mg m~2 day~1), (b) foraminifera (number m~2 day~1) forthe period August 1993 through September 1996, (c) diatoms (number]105 m~2 day~1), (d) radiolaria(number]103 m~2 day~1), and (e) silico#agellates (number]103 m~2 day~1) for the period August1993}December 1995.
G. bulloides varied from a maximum of 65% (early January 1994) to a low of 10% (lateFebruary}early March 1995) Over the three-year study period this species accountedfor an average of 42% of the total foraminiferal #ux.
E. Kincaid et al. / Deep-Sea Research II 47 (2000) 1157}1176 1165
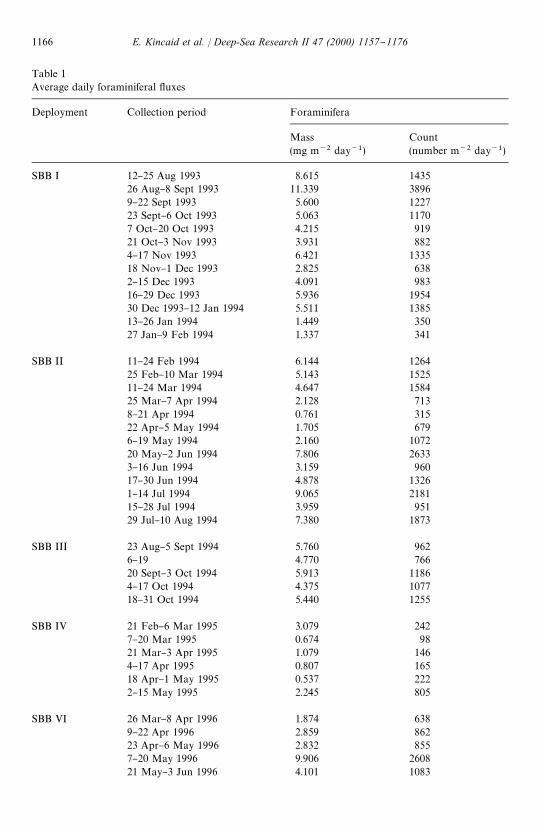
Table 1Average daily foraminiferal #uxes
Deployment Collection period Foraminifera
Mass(mg m~2 day~1)
Count(number m~2 day~1)
SBB I 12}25 Aug 1993 8.615 143526 Aug}8 Sept 1993 11.339 38969}22 Sept 1993 5.600 122723 Sept}6 Oct 1993 5.063 11707 Oct}20 Oct 1993 4.215 91921 Oct}3 Nov 1993 3.931 8824}17 Nov 1993 6.421 133518 Nov}1 Dec 1993 2.825 6382}15 Dec 1993 4.091 98316}29 Dec 1993 5.936 195430 Dec 1993}12 Jan 1994 5.511 138513}26 Jan 1994 1.449 35027 Jan}9 Feb 1994 1.337 341
SBB II 11}24 Feb 1994 6.144 126425 Feb}10 Mar 1994 5.143 152511}24 Mar 1994 4.647 158425 Mar}7 Apr 1994 2.128 7138}21 Apr 1994 0.761 31522 Apr}5 May 1994 1.705 6796}19 May 1994 2.160 107220 May}2 Jun 1994 7.806 26333}16 Jun 1994 3.159 96017}30 Jun 1994 4.878 13261}14 Jul 1994 9.065 218115}28 Jul 1994 3.959 95129 Jul}10 Aug 1994 7.380 1873
SBB III 23 Aug}5 Sept 1994 5.760 9626}19 4.770 76620 Sept}3 Oct 1994 5.913 11864}17 Oct 1994 4.375 107718}31 Oct 1994 5.440 1255
SBB IV 21 Feb}6 Mar 1995 3.079 2427}20 Mar 1995 0.674 9821 Mar}3 Apr 1995 1.079 1464}17 Apr 1995 0.807 16518 Apr}1 May 1995 0.537 2222}15 May 1995 2.245 805
SBB VI 26 Mar}8 Apr 1996 1.874 6389}22 Apr 1996 2.859 86223 Apr}6 May 1996 2.832 8557}20 May 1996 9.906 260821 May}3 Jun 1996 4.101 1083
1166 E. Kincaid et al. / Deep-Sea Research II 47 (2000) 1157}1176

Table 1 (continued)
Deployment Collection period Foraminifera
Mass(mg m~2 day~1)
Count(number m~2 day~1)
SBB VI 4}17 Jun 1996 3.787 111818 Jun}1 Jul 1996 1.255 4002}15 Jul 1996 3.778 104716}29 Jul 1996 4.311 108830 Jul}12 Aug 1996 5.701 130713}26 Aug 1996 6.375 132127 Aug}9 Sep 1996 5.342 936
Globigerina quinqueloba (Fig. 6b) dominated the assemblage in early March (52%)and mid-May 1994 (49%), but constituted less than 2% of the #ux in October 1993.Globigerina quinqueloba accounted for 22% of the total foraminiferal #ux fromAugust 1993 through September 1996. The N. pachyderma (Fig. 6c) population wassmaller than that of G. bulloides and slightly larger than that of G. quinqueloba. Duringlate September through December 1993 and October 1994 the #ux patterns ofG. bulloides and G. quinqueloba were inversely related to that of N. pachyderma (Fig.6c). Neogloboquadrina pachyderma contributed 60 and 64% of the foraminiferal #uxfor individual collection periods in October 1993 and October 1994, respectively, butonly 1% of the #ux in late February 1995. The average #ux for N. pachyderma was24% of the total foraminiferal #ux.
Orbulina universa, Neogloboquadrina dutertrei and Globigerinoides ruber, (Fig. 6d}f)are present seasonally with much lower #uxes than the three dominant species. The#ux patterns for these three species are similar, although G. ruber is the mostseasonally restricted of these species due to its narrower temperature tolerance range(Bijma et al., 1990b). Orbulina universa, N. dutertrei and G. ruber live in surface waters(Hemleben et al., 1989) and constitute a small percentage of the total #ux (mean"2.7,3.3 and 2.5%, respectively), when sea-surface temperatures are warm. Orbulina univer-sa (Fig. 6d) was present except during upwelling, from August 1993}October 1994 andlate February}early April 1995. Due to a partial trap failure we have no #ux data forthe intervening period. However, it would not be unreasonable to assume that thisspecies was present thoughout this period. The #ux for N. dutertrei (Fig. 6e) wasgreatest during the late summer and fall when the water column was well-strati"edand a deep chlorophyll maximum developed (Thunell and Sautter, 1992). During thesummer and fall of 1993 and summer 1994, N. dutertrei frequently was more abundantthan O. universa (Fig. 6d and e), a trend that reversed by December when the upperwater column cooled. Neogloboquadrina dutertrei disappeared with the onset ofupwelling, returning in low numbers approximately two months later. The El Nin8 o of1994}early 1995 (Fig. 2) had a pronounced e!ect on the foraminiferal population inthe Santa Barbara Basin. The total #ux for late February through early May 1995 wasonly 38% of the #ux measured for the same period of the previous year (Fig. 5a and b).
E. Kincaid et al. / Deep-Sea Research II 47 (2000) 1157}1176 1167
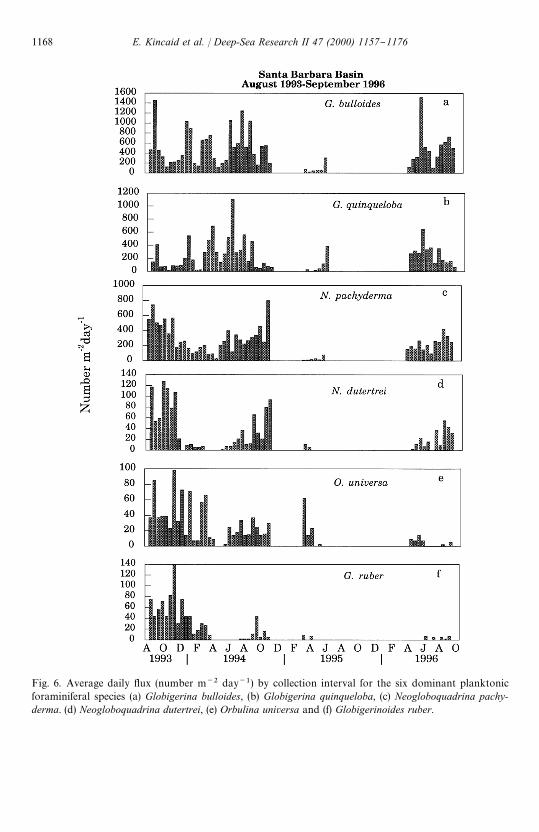
Fig. 6. Average daily #ux (number m~2 day~1) by collection interval for the six dominant planktonicforaminiferal species (a) Globigerina bulloides, (b) Globigerina quinqueloba, (c) Neogloboquadrina pachy-derma. (d) Neogloboquadrina dutertrei, (e) Orbulina universa and (f) Globigerinoides ruber.
1168 E. Kincaid et al. / Deep-Sea Research II 47 (2000) 1157}1176

Sea-surface temperatures initially cooled following the temperature maximum inAugust 1994 (19.83C) but then increased gradually from December 1994 (13.93C) toearly March (15.23C) (Fig. 3a). This was followed by an abrupt decrease in SST to13.53C in late March 1995 with the onset of upwelling and a further decrease to 11.83Cby mid-April 1995. During the "rst three collection intervals of trap deployment SBBIV (22 February}3 April 1994), G. bulloides and O. universa comprised 29 and 20%of the total foraminiferal #ux, respectively. This is within the expected range forG. bulloides but is high for O. universa. The increased contribution of O. universa to thetotal foraminiferal #ux is not due to increased production of this species, but rather tothe greatly reduced #uxes of other species normally present at this time.
4.4. Siliceous plankton yux
The siliceous plankton (diatom, radiolaria and silico#agellates) #ux data for theperiod August 1993}November 1994, as well as the compositional makeup of thesiliceous plankton groups, are presented in Lange et al. (1997). Herein, we extend thisrecord to December 1995. Diatoms dominated the siliceous biogenic #ux throughoutthe study and occurred in greatest numbers for brief periods during late winter andearly spring (Fig. 5c). Chaetoceros radicans resting spores were dominant duringupwelling conditions (spring), while representatives of temperate and warm o!shorewaters were common during non-upwelling conditions (Lange et al., 1997) (Fig. 7a}c).
Radiolarian #ux maxima occurred each fall (Fig. 5d). Although data are missing forpart of 1994 and 1995, the total radiolarian #ux was higher and more sustained during1995 than in the previous years. Increases in the #ux and relative abundance ofintermediate radiolarians were associated with upwelling events (Fig. 7e). Warm-water radiolarians had the highest relative abundance during late fall}early winter1993 and in late winter}early spring 1995 (Fig. 7f).
Flux maxima for silico#agellates occurred in winter 1993/1994 and early fall}winter1995 (Fig. 5e). The latter #ux peak was signi"cantly greater than any period in 1993and 1994.
5. Discussion
5.1. Foraminiferal and siliceous plankton yuxes
The planktonic foraminiferal and siliceous plankton #uxes show both seasonal andinterannual variability in response to environmental forcing. Production maxima ofdiatoms occur each year with the onset of upwelling in the early spring. Theforaminifera population increases quickly, approximately one month following theonset of upwelling, probably due to utilization of the diatoms as food. A similarrelationship between diatoms and planktonic foraminifera was previously reportedfor the San Pedro Basin by Sautter and Sancetta (1992). The maximum foraminiferal#ux occurs in late June}August when the surface waters are warm and there isa well-developed deep chlorophyll maximum.
E. Kincaid et al. / Deep-Sea Research II 47 (2000) 1157}1176 1169
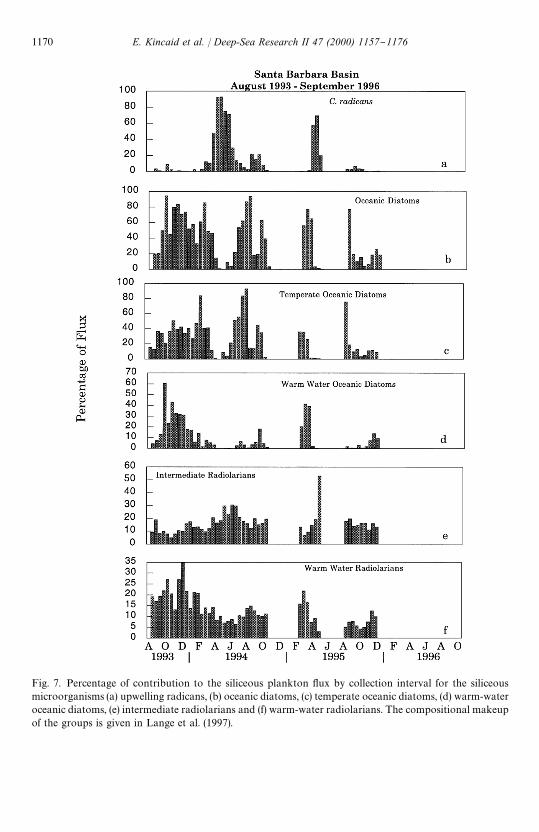
Fig. 7. Percentage of contribution to the siliceous plankton #ux by collection interval for the siliceousmicroorganisms (a) upwelling radicans, (b) oceanic diatoms, (c) temperate oceanic diatoms, (d) warm-wateroceanic diatoms, (e) intermediate radiolarians and (f) warm-water radiolarians. The compositional makeupof the groups is given in Lange et al. (1997).
1170 E. Kincaid et al. / Deep-Sea Research II 47 (2000) 1157}1176

Weak El Nin8 o conditions existed in 1993, when the trapping experiment began.These conditions strengthened in 1994 and persisted into early 1995 (Fig. 2) resultingin a reduction in the foraminiferal #ux. Unfortunately, trap failure during the fall andwinter of 1995}1996 precluded the determination of the precise timing of the return tomore normal population levels. However, the foraminiferal #ux during 1996 seems tohave returned to higher levels. Of the three groups of siliceous plankton studied, thesilico#agellates demonstrated the greatest interannual variability during our samplingperiod (Fig. 5e). Silico#agellate production was low from 1993 through early 1995,then increased dramatically in the second half of 1995 when non-El Nin8 o conditionsreturned (Fig. 5e).
5.2. Species yux pattern
The most abundant planktonic foraminiferal species in the Santa Barbara Basin areGlobigerina bulloides, Neogloboquadrina pachyderma and G. quinqueloba (Fig. 6a}c).The same three species dominate the planktonic foraminiferal fauna in other parts ofthe Southern California Bight (Sautter and Thunell, 1991b). However, the clear faunalsuccession present in the San Pedro Basin is not so obvious in the Santa BarbaraBasin. These species are present throughout the year in varying abundance in theSanta Barbara Basin. The maximum #uxes of G. bulloides and N. pachyderma tend tooccur synchronously in August, but during the remainder of the year their #uxpatterns show an inverse relationship. The pattern for G. quinqueloba is similar to thatof G. bulloides. Globigerina bulloides is a non-symbiont-bearing, predaceous speciesthat inhabits the transitional to polar zones (Hemleben et al., 1989). Ortiz et al. (1995),in a MOCNESS tow experiment, found G. bulloides living from the surface to100 m depth in the upwelling zone o! the Oregon coast. Although G. bulloides isfrequently considered to be an upwelling indicator, it is present and abundantthroughout the year in the Santa Barbara Basin. The periods of maximum #ux forG. bulloides include two upwelling events in May 1994 and 1996. However, other #uxmaxima occurred in September 1993, December 1993}January 1994, and severalintermittent periods in 1994 (February}March, early July and early August). The high#uxes of G. bulloides are not restricted to upwelling periods.
Globigerina quinqueloba harbors photosynthetic symbionts (Hemleben et al., 1989)and is therefore restricted to the upper water column (Hemleben et al., 1989; Ortizet al., 1995). In the Santa Barbara Basin, it was present in all collection intervals. Thehighest #uxes of this species are associated with spring upwelling and high diatomproduction. This observation is consistent with the results of an earlier study in theSan Pedro Basin, which reported that the highest #uxes of G. quinqueloba during Apriland May (Sautter and Thunell, 1991a). During this time, G. quinqueloba contributeda higher percentage of the total foraminiferal #ux than G. bulloides, the most abundantspecies in the Santa Barbara Basin (Fig. 8a and b), and increases in the #ux of thisspecies preceded those of G. bulloides following the diatom increase.
Neogloboquadrina dutertrei, O. universa and Globigerinoides ruber (Fig. 6d}f) arepresent in low numbers that vary seasonally. Orbulina universa, con"ned to the surfaceby the presence of photosynthetic endosymbionts (Spero and DeNiro, 1987), is present
E. Kincaid et al. / Deep-Sea Research II 47 (2000) 1157}1176 1171
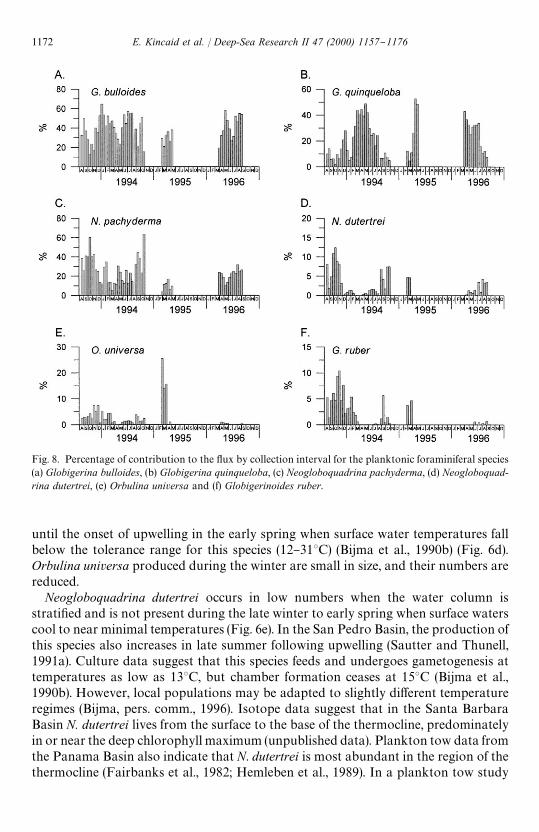
Fig. 8. Percentage of contribution to the #ux by collection interval for the planktonic foraminiferal species(a) Globigerina bulloides, (b) Globigerina quinqueloba, (c) Neogloboquadrina pachyderma, (d) Neogloboquad-rina dutertrei, (e) Orbulina universa and (f) Globigerinoides ruber.
until the onset of upwelling in the early spring when surface water temperatures fallbelow the tolerance range for this species (12}313C) (Bijma et al., 1990b) (Fig. 6d).Orbulina universa produced during the winter are small in size, and their numbers arereduced.
Neogloboquadrina dutertrei occurs in low numbers when the water column isstrati"ed and is not present during the late winter to early spring when surface waterscool to near minimal temperatures (Fig. 6e). In the San Pedro Basin, the production ofthis species also increases in late summer following upwelling (Sautter and Thunell,1991a). Culture data suggest that this species feeds and undergoes gametogenesis attemperatures as low as 133C, but chamber formation ceases at 153C (Bijma et al.,1990b). However, local populations may be adapted to slightly di!erent temperatureregimes (Bijma, pers. comm., 1996). Isotope data suggest that in the Santa BarbaraBasin N. dutertrei lives from the surface to the base of the thermocline, predominatelyin or near the deep chlorophyll maximum (unpublished data). Plankton tow data fromthe Panama Basin also indicate that N. dutertrei is most abundant in the region of thethermocline (Fairbanks et al., 1982; Hemleben et al., 1989). In a plankton tow study
1172 E. Kincaid et al. / Deep-Sea Research II 47 (2000) 1157}1176

conducted o! the Oregon coast, Ortiz and others (1995) found that this species hada subsurface maximum in the o!shore station but occurred in highest abundance inthe upper 50}70 m in the nearshore stations in the California Current.
Globigerinoides ruber, another symbiont-bearing species, is a tropical}subtropicalspecies that tolerates temperatures from 14}323C (Bijma et al., 1990b) and is present inthis area only when surface waters are warmest (from &14}19.83C), with #uxesincreasing when sea-surface temperatures increase (Fig. 6f). Increased #ux co-occurdwith increased siliceous plankton contributions from warm oceanic diatoms andwarm-water radiolarians. The seasonal range for G. ruber is more restricted than thatof O. universa and N. pachyderma.
The species composition and seasonal succession of siliceous microorganisms in theSanta Barbara Basin are linked to changes in circulation patterns and are diagnosticof water masses a!ecting the basin (Lange et al., 1997). The siliceous planktonexhibit distinct seasonal preferences. These seasonal successions may re#ect adapta-tion to water-column conditions and/or be related to source waters. Diatomsdominate the siliceous biogenic #ux (Fig. 5c) and are most abundant in late winterand spring. Upwelling occurs during this time, and the diatoms respond rapidly tothe increased nutrients in the surface water. Speci"cally, there is an increase in theabundance of C. radicans resting spores, which dominate the siliceous biogenic #ux,and moderate increases in intermediate radiolarians (Lithomelissa setosa), which liveat greater depths and are associated with colder, higher salinity waters (Fig. 7a)(Lange et al., 1997). Chaetoceros radicans, as well as Pseuudo-nitzschia delicatissima,are found under similar conditions in the San Pedro Basin (Sautter and Sancetta,1992).
As surface waters warm and thermal strati"cation of the water column begins todevelop in early summer, the spring upwelling assemblage (Fig. 7a and e) is replacedby moderate radiolarian #uxes containing a higher relative abundance of warm-waterradiolarians and increasing numbers of oceanic (predominately temperate) diatoms(Fig. 7b and e) During such events, surface waters enter the basin over the western silland #ow everywhere is eastward throughout the basin (Hendershott and Winant,1996). The radiolarian #uxes increased in 1995 relative to comparable periods in 1993and 1994. Silico#agellates, prior to 1995, had shown low, maximum #uxes in Decem-ber 1993 and August 1994. During 1995 there was a prolonged, signi"cant increase inthe #ux of silico#agellates from August through December (Fig. 5e).
The three-year time period represented by this sediment trap experiment containsa marked shift in hydrographic regimes in the Santa Barbara Basin. When the trapwas "rst deployed in August 1993, weak El Nin8 o conditions existed in this region(Fig. 2). The El Nin8 o intensi"ed in 1994 and continued into early 1995, whichapparently marked a transition to a di!erent regime. From spring through late fall1995 there was a persistent increase in the radiolarian #ux. Similarly, silico#agellate#uxes increased dramatically in the second half of 1995. Conversely, the diatom #uxesdecreased relative to the previous years of the study (Lange et al., 1997). Theforaminiferal #uxes in 1996, with the exception of the spike during May 1996, wereslightly reduced from that of the #uxes for comparable periods of 1993}1994 and alsoless variable. The contributions from those foraminiferal species that seem to be most
E. Kincaid et al. / Deep-Sea Research II 47 (2000) 1157}1176 1173

limited by temperature (i.e. O. universa, N. dutertrei) were sharply reduced in 1996relative to their 1993}1994 levels. In 1995, the foraminiferal #uxes were signi"cantlybelow the levels found during the remaining years of the study, the greatest reductionsoccurring in the species that are normally most abundant. These changes may re#ectinterannual di!erences in the hydrographic conditions present in the basin. During1994 and 1995 upwelling began in late March or early April (Fig. 3). In contrast, theonset of upwelling in 1996 appears to have occurred in late January and, althoughslightly weakened, continued into June, resulting in lower sea-surface temperatures inthe basin over this extended period of time. Water temperatures and the speciespresent in 1993 and 1994 suggest warmer conditions and stronger oceanic in#uencesthan in 1996 when non-El Nin8 o conditions existed.
6. Conclusions
The plankton distribution patterns de"ned by the sediment-trap #ux data suggestthat these trends are driven by both seasonal and interannual changes in the upperwater column. Seasonal succession of diatoms and radiolaria are apparently related tochanges in source waters and surface-water temperatures. Data on species turnovermay be utilized to examine changes in circulation in the basin (i.e., upwelling versusnon-upwelling conditions). We observed a signi"cant, prolonged increase in thesilico#agellate #ux in 1995 when sea-surface temperatures were generally cooler thanin the previous two years of the study.
Changes in planktonic #ux and the association of individual species with di!erenttemperature thresholds provide additional means of determining past changes inwater temperatures from fossil assemblages in sediments. Changes in assemblagecomposition may be utilized to ascertain seasonal to interannual hydrographicchanges in the overlying surface waters. By employing information obtained fromboth siliceous microorganisms and foraminifera, it should be possible to recoverinformation preserved in the varved sediments of the Santa Barbara Basin concerningchanges in the hydrographic regimes through time.
Highest planktonic foraminiferal #uxes occurred approximately one month follow-ing the onset of upwelling to early fall. Foraminiferal #uxes declined during El Nin8 oconditions and returned to moderate levels in 1996. Globigerina bulloides andG. quinqueloba had similar #ux patterns, which were, in general, inversely related tothat of N. pachyderma. Temperature-limited species present seasonally in the basinmay prove to be useful tools for determining seasonal variability in upper water-column processes and thermal boundaries in glacial}interglacial climate systems.
The three-year time period of the sediment trap study encompasses El Nin8 o tonon-El-Nin8 o conditions. The observed changes in both siliceous and calcareousplankton represent a clear response to these short-term climate variations. It isimportant to characterize thoroughly the modern plankton response to physical andclimatic forcing and to apply that knowledge to the sedimentary assemblage toreconstruct past climate systems.
1174 E. Kincaid et al. / Deep-Sea Research II 47 (2000) 1157}1176

Acknowledgements
This research was supported by NOAA Grants NA36GP0239 and NA56GPO243.We thank E. Tappa for coordinating the sediment trapping program and sampleprocessing.
References
BeH , A.W.H., 1977. An ecological, zoogeographic and taxonomic review of recent planktonic foraminifera.In: Ramsay, A.T.S. (Ed.), Oceanic Micropaleontology. Academic Press, London, pp. 1}100.
BeH , A.W.H., Hutson, W.H., 1977. Ecology of planktonic foraminifera and biogeographic patterns of life andfossil assemblages in the Indian Ocean. Micropaleontology 23, 369}414.
Behl, R.J., Kennett, P.J., 1996. Brief interstadial events in the Santa Barbara basin, NE Paci"c, during thepast 60 kyr. Nature 379, 243}246.
Berger, W.H., Soutar, A., 1970. Preservation of plankton shells in an anaerobic basin o! California.Geological Society of America Bulletin 81, 275}282.
Bijma, J., Erez, J., Hemleben, C., 1990a. Lunar and semi-lunar reproductive cycles in some spinoseplanktonic foraminifers. Journal of Foraminiferal Research 20, 117}127.
Bijma, J., Faber Jr., W.W., Hemleben, C., 1990b. Temperature and salinity limits for growth andsurvival of some planktonic foraminifers in laboratory cultures. Journal of Foraminiferal Research 20,95}116.
Bouvier-Soumagnac, Y., Duplessy, J.-C., 1985. Carbon and oxygen isotopic composition of planktonicforaminifera from laboratory culture, plankton tows and Recent sediment: implications for the recon-struction of paleoclimatic conditions and of the global carbon cycle. Journal of Foraminiferal Research15, 302}320.
Deuser, W.G., 1986. Seasonal and interannual variations in deep-water particle #uxes in the Sargasso Seaand their relation to surface hydrography. Deep-Sea Research 33, 225}246.
Deuser, W.G., 1987. Seasonal variations in isotopic composition and deep-water #uxes of the tests ofperennially abundant planktonic foraminifera of the Sargasso Sea: results from sediment-trap collec-tions and their paleoceanographic signi"cance. Journal of Foraminiferal Research 17, 14}27.
Deuser, W.G., Ross, E.H., 1989. Seasonally abundant planktonic foraminifera of the Sargasso Sea:succession, deep-water #uxes, isotopic compositions, and paleoceanographic implications. Journal ofForaminiferal Research 19, 268}293.
Fairbanks, R.G., Sverdlove, M., Free, R., Wiebe, P.H., BeH , A.W.H., 1982. Vertical distribution and isotopicfractionation of living planktonic foraminifera from the Panama Basin. Nature 298, 841}844.
Fairbanks, R.G., Wiebe, P.H., 1980. Foraminifera and chlorophyll maximum: vertical distribution, seasonalsuccession, and paleoceanographic signi"cance. Science 209, 1524}1526.
Gunn, J.T., Hamilton, P., Herring, H.J., Kantha, L.H., Lagerloef, G.S.E., Mellor, G.L., Muench, R.D.,Stegen, G.R., 1987. Santa Barbara Channel Circulation Model and Field Study, Vols. 1 and 2. Dyanlysisof Princeton, Princeton.
Hemleben, C., Spindler, M., Anderson, O.R., 1989. Modern Planktonic Foraminifera. Springer, New York,pp. 363.
Hendershott, M.C., Winant, C.D., 1996. Surface circulation in the Santa Barbara Channel. Oceanography9, 114}121.
Heusser, L., 1978. Pollen in Santa Barbara Basin, California: a 12,000 record. Geological Society of AmericaBulletin 89, 673}678.
Hickey, B.M., 1992. Circulation over the Santa Monica-San Pedro Basin and Shelf. Progress in Oceano-graphy 30, 37}115.
Hulsemann, J., Emery, K.O., 1961. Strati"cation in recent sediments of Santa Barbara Basin as controlledby organisms and water character. Journal of Geology 69, 279}290.
Huyer, A., 1983. Coastal upwelling in the California current system. Progress in Oceanography 12, 259}284.
E. Kincaid et al. / Deep-Sea Research II 47 (2000) 1157}1176 1175

Jackson, G.A., 1986. Physical oceanography of the Southern California Bight. In: Eppley, R.W. (Ed.),Plankton Dynamics of the Southern California Bight: lecture Notes on Coastal and Estuarine Studies.Springer, Berlin, pp. 13}52.
Kennedy, J.A., Brassell, S.C., 1992. Molecular records of twentieth-century El Nin8 o events in laminatedsediments from the Santa Barbara basin. Nature 357, 62}64.
Kennett, J.P., Ingram, B.L., 1995. A 20,000-year record of ocean circulation and climate change from theSanta Barbara Basin. Nature 377, 510}514.
Lange, C.B., Berger, W.H., Burke, S.K., Casey, R.E., Schimmelmann, A., Soutar, A., Weinheimer, A.L., 1987.El Nin8 o in Santa Barbara Basin: Diatom, radiolarian and foraminiferan responses to the `1983El Nin8 oa event. Marine Geology 78, 153}160.
Lange, C.B., Weinheimer, A.L., Reid, F.M.H., Thunell, R.C., 1997. Temporal sedimentation patterns ofdiatoms, radiolarians and silico#agellates in Santa Barbara Basin, California.. California CooperativeFisheries Investigations 38, 161}170.
McGowan, J.A., 1985. El Nin8 o 1983 in the Southern California Bight. In: Wooster, W.S. Fluharty,D.L (Eds.) El Nin8 o North. University of Washington, Seattle, pp. 166}184.
Ortiz, J.D., Mix, A.C., Collier, R.W., 1995. Environmental control of living symbiotic and asymbioticforaminifera of the California current. Paleoceanography 10 (6), 987}1009.
Pisias, N.G., 1978. Paleoceanography of the Santa Barbara Basin during the last 8000 years. QuaternaryResearch 10, 366}384.
Sautter, L.R., Sancetta, C., 1992. Seasonal associations of phytoplankton and planktic foraminifera in anupwelling region and their contribution to the sea#oor. Marine Micropaleontology 18, 263}278.
Sautter, L.R., Thunell, R.C., 1991a. Planktonic foraminiferal response to upwelling and seasonal hydro-graphic conditions: sediment trap results from San Pedro Basin, Southern California Bight. Journal ofForaminiferal Research 21, 347}363.
Sautter, L.R., Thunell, R.C., 1991b. Seasonal variability in the d18O and d13C of planktonic foraminiferafrom an upwelling environment: sediment trap results from the San Pedro Basin, Southern CaliforniaBight. Paleoceanography 6 (3), 307}334.
Schimmelmann, A., Lange, C.B., Berger, W.H., 1990. Climatically controlled marker layers in SantaBarbara Basin sediments and "ne-scale core-to-core correlation. Limnology and Oceanography 35,165}173.
Soutar, A., Crill, P.A., 1977. Sedimentation and climatic patterns in the Santa Barbara Basin during the19th and 20th centuries. Geological Society of America Bulletin 88, 1161}1172.
Spero, H.J., 1987. Symbiosis in the planktonic foraminifer, Orbulina universa, and the isolation of itssymbiotic dino#agellate Gymnodinium be& ii Sp. Nov. Journal of Phycology 23, 307}317.
Spero, H.J., Angel, D.L., 1991. Planktonic sarcodines: microhabitat for oceanic dino#agellates. Journal ofPhycology 27, 187}195.
Spero, H.J., DeNiro, M.J., 1987. The in#uence of symbiont photosynthesis on the d18O and d13C values ofplanktonic foraminiferal shell calcite. Symbiosis 4, 213}228.
Thunell, R.C., 1998. Particle #uxes in a coastal upwelling zone: sediment trap results from Santa BarbaraBasin, California. Deep-Sea Research 45, 1863}1884.
Thunell, R.C., Sautter, L.R., 1992. Planktonic foraminiferal faunal and stable isotopic indices of upwelling:a sediment trap study in the San Pedro Basin, Southern California Bight. In: Summerhayes, C.P., Prell,W.L., Emeis, K.C. (Eds.), Upwelling Systems: Evolution Since the Early Miocene. Geological Society,London, pp. 77}91.
Thunell, R.C., Tappa, E., Anderson, D.M., 1995. Sediment #uxes and varve formation in Santa BarbaraBasin, o!shore California. Geology 23, 1083}1086.
Weinheimer, A.L., Carson, T.L., Wigley, C.R., Casey, R.E., 1986. Radiolarian responses to recent andNeogene California El Nin8 o and Anti-El Nin8 o events. Palaeogeography, Palaeoclimatology,Palaeoecology 53, 3}35.
Weinheimer, A.L., Cayan, D.R., 1997. Radiolarian assemblages from Santa Barbara Basin sediments:Recent interdecadal variability. Paleoceanography 12 (5), 658}670.
Wigley, C.R., 1984. A radiolarian analysis of the Monterey formation: paleoceanographic reconstructions ofthe Neogene California Current System. Ph.D. Dissertation, Houston, Rice University, 467pp.
1176 E. Kincaid et al. / Deep-Sea Research II 47 (2000) 1157}1176
Related Documents











