European Journal of PROTISTOLOGY European Journal of Protistology 43 (2007) 193–204 Planktonic dinoflagellates of the northern Levantine Basin, northeastern Mediterranean Sea Sevim Polat a, , Tufan Koray b a University of C ¸ ukurova, Faculty of Fisheries, 01330, Balcalı, Adana, Turkey b University of Ege, Faculty of Fisheries, 35100, Bornova, I ˙ zmir, Turkey Received 21 December 2006; received in revised form 27 February 2007; accepted 11 March 2007 Abstract In the present study, a checklist of planktonic dinoflagellates was prepared from studies conducted on the northeastern Mediterranean coast of Turkey and by evaluating previously conducted studies. The sampling areas included I ˙ skenderun Bay, the coast near Karatas - at the west end of the Bay, and Babadıllimanı Bight, situated at the west of Mersin Bay, and its surroundings. The data of the present study derives from over 90 locations visited between 1994 and 2004. A total of 174 taxa were reported. Ceratium (57 taxa) and Protoperidinium (28 taxa) were the most species-rich genera. The most common taxa were Ceratium kofoidii, Ceratium trichoceros, Ceratium tripos var. atlanticum, Protoperidinium divergens, Protoperidinium steinii, Ceratocorys horrida, Goniodoma acuminatum and Gonyaulax polygramma. Toxic species such as Dinophysis caudata, Dinophysis tripos were reported, and Scrippsiella trochoidea and Prorocentrum micans, which are known as red tide species elsewhere, were also common. The number of species is similar to those along other coasts of the Levantine Basin. However, some species were observed that had not previously been reported for the Levantine Basin, which can be attributed to the fact that the number of studies conducted in the region has increased and that new species may have entered the environment. r 2007 Elsevier GmbH. All rights reserved. Keywords: Checklist; Dinoflagellates; Levantine basin; Northeastern Mediterranean Introduction In marine ecosystems, dinoflagellates, along with diatoms, are important components of the phytoplank- ton. Approximately 90% of all dinoflagellate species are marine, most of them distributed in temperate waters, and most prevalent in summer months (Taylor 1987). Since dinoflagellates display very diverse ecophysiologi- cal characteristics, generalizations about their roles in ecosystems are not easily made. Photosynthetic species play a role in marine primary production, but most species are heterotrophic (Loeblich III 1984), and may be important grazers on the larger phytoplankton. Some species may cause red tides and some species produce potent neuro-toxins which may accumulate in the food chain (Hallegraeff 1993). The Mediterranean is one of the less productive seas in the world. The concentrations of nutrients decline from the west towards the east (Azov 1991; Krom et al. 1991). The Levantine Basin, located in the northeastern corner of the eastern Mediterranean, is the most oligotrophic part of the Sea. The cyclonic circulation of waters poor in nutrients, the hot and dry climate, and ARTICLE IN PRESS www.elsevier.de/ejop 0932-4739/$ - see front matter r 2007 Elsevier GmbH. All rights reserved. doi:10.1016/j.ejop.2007.03.003 Corresponding author. Fax: +90 322 338 64 39. E-mail address: [email protected] (S. Polat).

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ARTICLE IN PRESS
European Journal of
PROTISTOLOGY
0932-4739/$ - se
doi:10.1016/j.ej
�CorrespondE-mail addr
European Journal of Protistology 43 (2007) 193–204
www.elsevier.de/ejop
Planktonic dinoflagellates of the northern Levantine Basin, northeastern
Mediterranean Sea
Sevim Polata,�, Tufan Korayb
aUniversity of Cukurova, Faculty of Fisheries, 01330, Balcalı, Adana, TurkeybUniversity of Ege, Faculty of Fisheries, 35100, Bornova, Izmir, Turkey
Received 21 December 2006; received in revised form 27 February 2007; accepted 11 March 2007
Abstract
In the present study, a checklist of planktonic dinoflagellates was prepared from studies conducted on thenortheastern Mediterranean coast of Turkey and by evaluating previously conducted studies. The sampling areasincluded Iskenderun Bay, the coast near Karatas- at the west end of the Bay, and Babadıllimanı Bight, situated at thewest of Mersin Bay, and its surroundings. The data of the present study derives from over 90 locations visited between1994 and 2004. A total of 174 taxa were reported. Ceratium (57 taxa) and Protoperidinium (28 taxa) were the mostspecies-rich genera. The most common taxa were Ceratium kofoidii, Ceratium trichoceros, Ceratium tripos var.atlanticum, Protoperidinium divergens, Protoperidinium steinii, Ceratocorys horrida, Goniodoma acuminatum andGonyaulax polygramma. Toxic species such as Dinophysis caudata, Dinophysis tripos were reported, and Scrippsiella
trochoidea and Prorocentrum micans, which are known as red tide species elsewhere, were also common. The number ofspecies is similar to those along other coasts of the Levantine Basin. However, some species were observed that had notpreviously been reported for the Levantine Basin, which can be attributed to the fact that the number of studiesconducted in the region has increased and that new species may have entered the environment.r 2007 Elsevier GmbH. All rights reserved.
Keywords: Checklist; Dinoflagellates; Levantine basin; Northeastern Mediterranean
Introduction
In marine ecosystems, dinoflagellates, along withdiatoms, are important components of the phytoplank-ton. Approximately 90% of all dinoflagellate species aremarine, most of them distributed in temperate waters,and most prevalent in summer months (Taylor 1987).Since dinoflagellates display very diverse ecophysiologi-cal characteristics, generalizations about their roles inecosystems are not easily made. Photosynthetic species
e front matter r 2007 Elsevier GmbH. All rights reserved.
op.2007.03.003
ing author. Fax: +90322 338 64 39.
ess: [email protected] (S. Polat).
play a role in marine primary production, but mostspecies are heterotrophic (Loeblich III 1984), and maybe important grazers on the larger phytoplankton. Somespecies may cause red tides and some species producepotent neuro-toxins which may accumulate in the foodchain (Hallegraeff 1993).
The Mediterranean is one of the less productive seasin the world. The concentrations of nutrients declinefrom the west towards the east (Azov 1991; Krom et al.1991). The Levantine Basin, located in the northeasterncorner of the eastern Mediterranean, is the mostoligotrophic part of the Sea. The cyclonic circulationof waters poor in nutrients, the hot and dry climate, and

ARTICLE IN PRESSS. Polat, T. Koray / European Journal of Protistology 43 (2007) 193–204194
the low terrestrial input lead to low plankton biomassand primary production (Turley et al. 2000).
The oligotrophic conditions result in a high diversityof dinoflagellates in the Mediterranean (Gomez 2003).Thus, the Mediterranean Sea contains nearly 43% of theworld’s known marine dinoflagellate species (Gomez2005). Numerous studies have been conducted onplankton in various parts of the Mediterranean byHalim (1963), El-Maghraby and Halim (1965), Dowidar(1974), Rampi and Bernhard (1980), Lakkis andLakkis (1980), Delgado and Fortuna (1991), Ignatiadeset al. (1995), Gomez and Gorsky (2003) and Gomez andClaustre (2003). However, although Turkey is oneof the countries with the longest coastline in theMediterranean, phytoplankton studies were few up tothe 1990s. From the 1990s onwards, in studies con-ducted by Polat et al. (2000), Polat (2002), Polat and Is-ık(2002) and Polat and Piner (2002a) in the northeasternMediterranean, the Iskenderun Bay phytoplanktonespecially was investigated. The phytoplankton inMersin Bay was investigated by Kıdeys- et al. (1989)and Eker and Kıdeys- (2000), and the western part ofMersin Bay by Polat and Piner (2002b), Polat andKoray (2002, 2003) and Polat et al. (2004). On the otherhand, qualitative properties of microplankton in Turk-ish Seas (Black Sea, Aegean Sea and NortheasternMediterranean Sea) were compared by Koray et al.(2000). In most of these studies, the dinoflagellates wereevaluated along with the other phytoplankton groups.The Mediterranean Sea is exposed to the migration ofRed Sea species through the Suez Canal and to thepassage of species from the Atlantic Ocean throughthe Strait of Gibraltar. On the other hand, humanpressure and climate change are expected to causewidespread changes in the physical and chemicalcharacteristics of the Mediterranean Sea (Goffart et al.2002). Phytoplankton diversity is being affected by allthese factors. Thus, investigation of the diversity ofphytoplankton in this changing environment becomestimely and important.
This study aimed at forming an up-to-date species listof the dinoflagellates by gathering the most recentsampling studies conducted in Iskenderun Bay and itssurroundings in the northern Levantine Basin. Resultsof previously published studies conducted in the north-eastern Mediterranean coast of Turkey were alsoevaluated.
Material and methods
Study area
The Levantine Basin is one of the largest basins of theeastern Mediterranean. The areas investigated in this
study include Iskenderun Bay, located on the northernLevantine coasts of Turkey, the region offshore fromKaratas-, located at the northern side of the exit fromIskenderun Bay, and Babadıllimanı Bight, situated atthe west of Mersin Bay, and its surroundings (lat.361070N–361540N and long. 331240E–36111.50E) (Fig. 1).The vicinity of Iskenderun Bay including the area tillMersin Bay covers the largest continental shelf in theeastern Mediterranean except for the Nile Delta.Iskenderun Bay is affected by current systems prevailingin the northeastern Mediterranean because of itsconnection through the wide opening and by theregional and local winds (Yılmaz et al. 1992). The seasurface temperature varies from 15–16 1C to 29–30 1Cand salinity in the region is around 39%. Salinitydecreases to lower values (37–38%) near freshwaterinputs (Yılmaz et al. 1992). The coastline betweenMersin and Iskenderun Bay is intensely industrializedwith sites including artificial fertilizer, iron–steel andpetroleum industries. The heavy marine traffic alsocontributes oil pollution in this region. The Seyhan andCeyhan Rivers, which are the most important rivers ofthe region, drain into the sea from an area betweenIskenderun and Mersin Bay.
Methods
The present study, which focuses on an investigationof the planktonic dinoflagellates of the northeasternMediterranean, provides an evaluation of the findingsobtained through sampling studies between 1994 and2004. The information about sampling studies con-ducted for this research and of the published literatureused as references are provided in Table 1. Sampling forthis study was carried out on the northwest part ofIskenderun Bay in 2001–2002, on the east part of thebay offshore from Iskenderun and Karatas- in 2003–2004(Table 1). The other sampling data are related to thepreviously published studies conducted in the regionsince 1994 (Table 1).
When the sampling performed in this study and in thepreviously conducted studies were gathered together,samples from a total of over 90 locations wereinvestigated, over 70 of which were sites of studiesconducted in Iskenderun Bay and its vicinity and over20 at the west of Mersin Bay.
Surface phytoplankton samples were collected bymeans of 20 and 45 mm mesh plankton nets. Verticalsamplings were conducted using a Nansen water sampler.After the plankton samples were obtained, they werefixed in 2% buffered formalin. For the identification ofthe dinoflagellates, samples were treated with sodiumhypochlorite to separate the thecal plates. A fluorescentdye, Calcofluor White was used to stain the dinoflagel-late plates (Fritz and Triemer 1985). The samples were
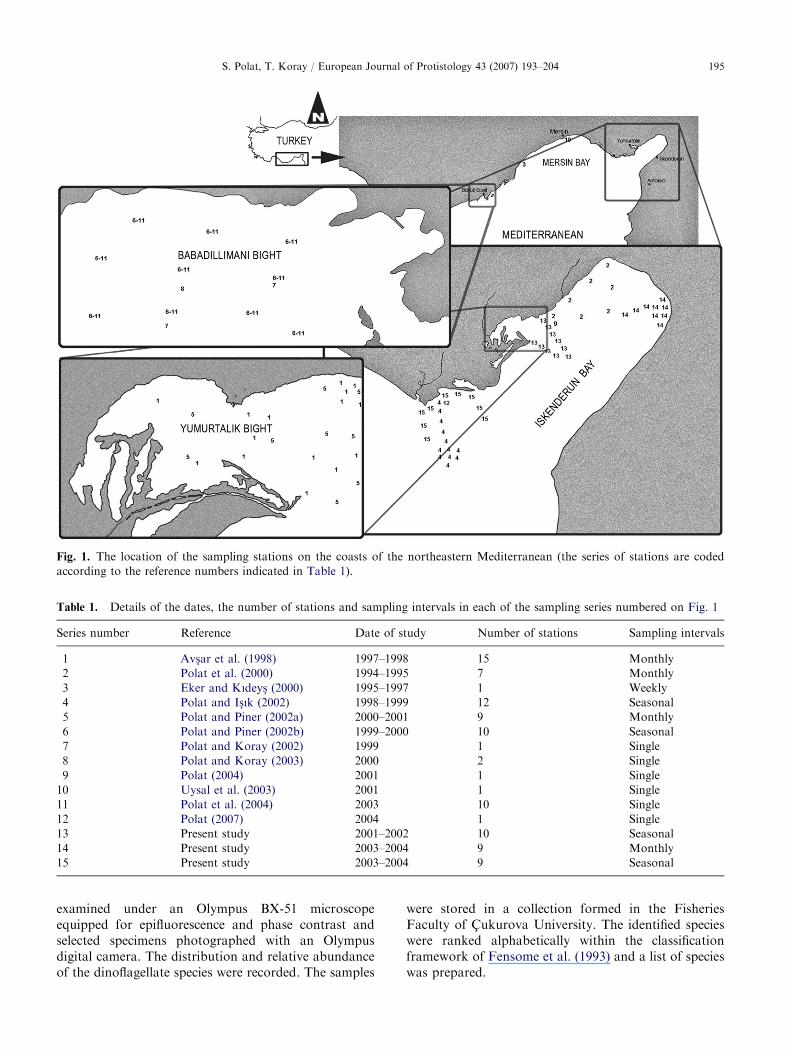
ARTICLE IN PRESS
Fig. 1. The location of the sampling stations on the coasts of the northeastern Mediterranean (the series of stations are coded
according to the reference numbers indicated in Table 1).
Table 1. Details of the dates, the number of stations and sampling intervals in each of the sampling series numbered on Fig. 1
Series number Reference Date of study Number of stations Sampling intervals
1 Avs-ar et al. (1998) 1997–1998 15 Monthly
2 Polat et al. (2000) 1994–1995 7 Monthly
3 Eker and Kıdeys- (2000) 1995–1997 1 Weekly
4 Polat and Is-ık (2002) 1998–1999 12 Seasonal
5 Polat and Piner (2002a) 2000–2001 9 Monthly
6 Polat and Piner (2002b) 1999–2000 10 Seasonal
7 Polat and Koray (2002) 1999 1 Single
8 Polat and Koray (2003) 2000 2 Single
9 Polat (2004) 2001 1 Single
10 Uysal et al. (2003) 2001 1 Single
11 Polat et al. (2004) 2003 10 Single
12 Polat (2007) 2004 1 Single
13 Present study 2001–2002 10 Seasonal
14 Present study 2003–2004 9 Monthly
15 Present study 2003–2004 9 Seasonal
S. Polat, T. Koray / European Journal of Protistology 43 (2007) 193–204 195
examined under an Olympus BX-51 microscopeequipped for epifluorescence and phase contrast andselected specimens photographed with an Olympusdigital camera. The distribution and relative abundanceof the dinoflagellate species were recorded. The samples
were stored in a collection formed in the FisheriesFaculty of Cukurova University. The identified specieswere ranked alphabetically within the classificationframework of Fensome et al. (1993) and a list of specieswas prepared.

ARTICLE IN PRESSS. Polat, T. Koray / European Journal of Protistology 43 (2007) 193–204196
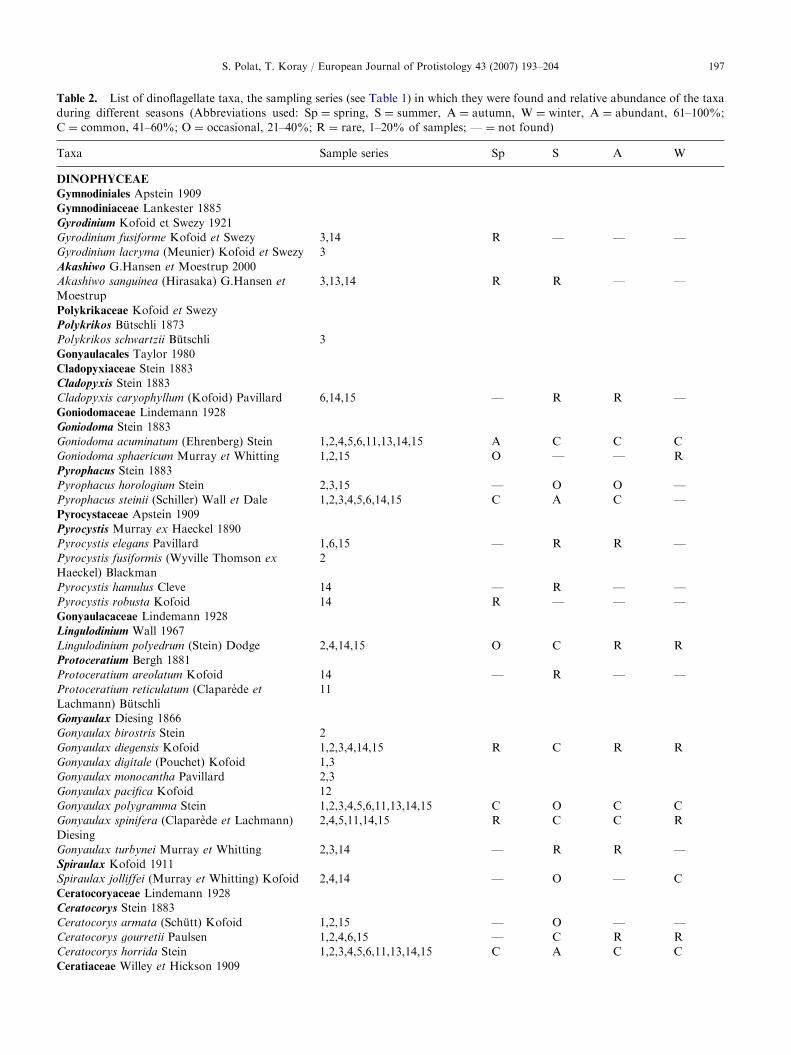
Results
A total of 30 genera of the planktonic dinoflagellatesand 174 taxa belonging to these genera were recordedduring the sampling studies (Table 2). The mostnumerous genera were Ceratium, with 57 taxa andProtoperidinium, with 28 taxa, accounting for 33% and16% of the total number of species, respectively. Insome previous studies, several species of Ceratium werenamed at the species level whereas in other studies thosespecies were identified to variety level. Thus, in thepresent study both the species and variety names ofthose species are given separately in the list along withthe numbers of the collections in which they were found(Table 2). The most common species of these genera areCeratium candelabrum, Ceratium furca, Ceratium fusus,
Ceratium kofoidii, Ceratium trichoceros, Ceratium tripos
var. atlanticum, Protoperidinium divergens and Proto-
peridinium steinii. The abundance of these speciesincreased in summer; nonetheless, the species belongingto these genera were observed to be in low numbers(mostly below 30� 102 cells l�1).
Ceratocorys horrida, Prorocentrum micans, Dinophysis
caudata and Gonyaulax polygramma were also found tobe frequent species. However, the rare detection of suchspecies as Ceratium longissimum, Ceratium platycorne,Ornithocercus carolinae, Ornithocercus splendidus, Am-
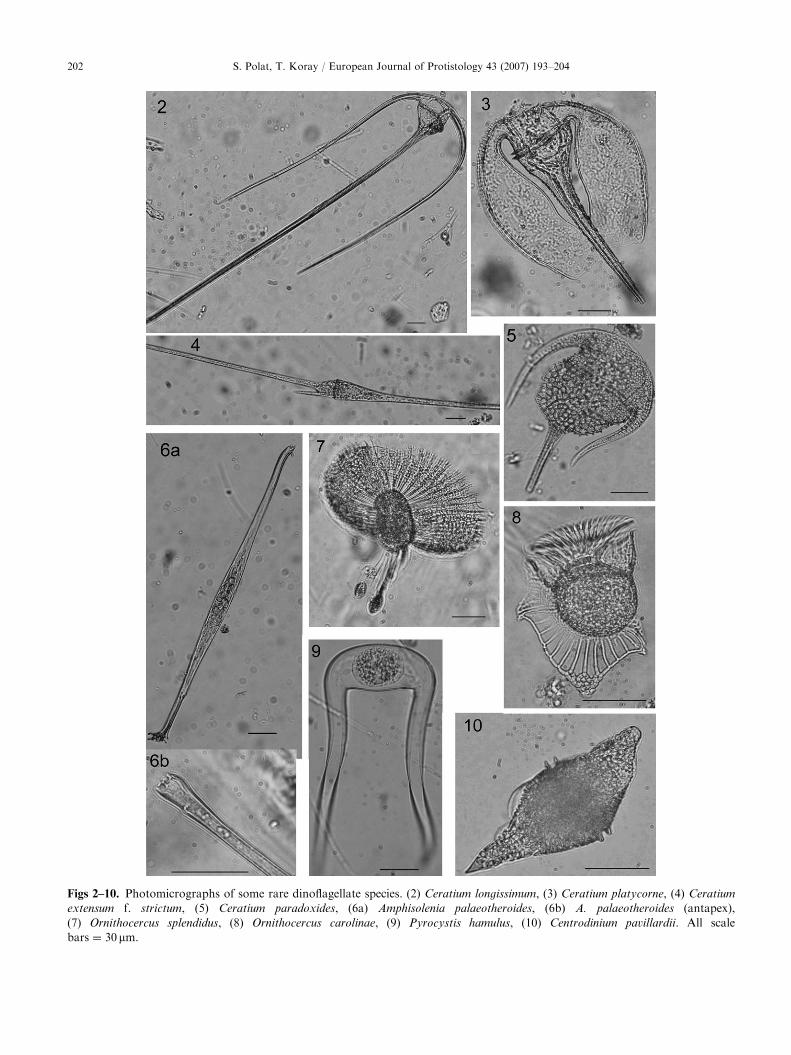
phisolenia palaeotheroides and Histioneis elongata in netsamples indicated the heterogeneity of the community.Figs 2–12 illustrate some of the rare species in the area.
The records of unarmoured species are few due to theuse of preserved samples. Such unarmoured species asGyrodinium fusiforme, Gyrodinium lacryma, Akashiwo
sanguinea and Polykrikos schwartzii also showed a poordistribution pattern.
Of the toxic dinoflagellates, D. caudata, Dinophysis
tripos and Dinophysis rotundata, and of the red tidespecies, Prorocentrum micans, Scrippsiella trochoidea
and Noctiluca scintillans were identified. Of these, whileD. tripos was a rare species, the others displayed arelatively wide distribution. However, no evidence isavailable either in the present study or in previouslyconducted studies in the region for any recent increase inthe toxic or harmful species.
Dinoflagellates were generally found in low abun-dances; mostly below 103 cells l�1. Between 1994 and1995, Prorocentrum micans was recorded as the mostnumerous species in Iskenderun Bay, with64.6� 102 cells l�1 (Polat et al. 2000). In the Karatas-offshore between 1998 and 1999, with the increase in theS. trochoidea, the highest dinoflagellate abundance wasreported to be 118.3� 102 cells l�1, whereas the lowestwas 0.27� 102 cells l�1 (Polat and Is-ık 2002). The mostabundant species identified in the bay between the years2000 and 2001 were Goniodoma acuminatum (formerlyGoniodoma polyedricum or Heteraulacus polyedricus)
with 72.7� 102 cells l�1 and Prorocentrum micans with25.9� 102 cells l�1 (Polat and Piner 2002a). However,the highest concentration of dinoflagellates, whichamounted to 870� 102 cells l�1, was caused on the eastof the bay between 2003 and 2004 by S. trochoidea.
Discussion
The results of this study bear some similarities withthose conducted in other countries with Mediterraneancoasts. In the eastern Mediterranean, Kimor and Wood(1975) identified 142, Lakkis and Lakkis (1980) 157, andDowidar (1974) 182 taxa. Furthermore, the species listformed in this study is similar in content to the check-listprepared for Turkish seas by Koray (2001), in which 247dinoflagellate taxa were reported. The number of speciesin the list prepared by Koray (2001) was higher becauseit encompassed all of the seas surrounding Turkey, butsuch species as Ceratium azoricum, Ceratium platycorne,
Amphisolenia palaeotheroides and Pyrocystis hamulus
(Figs. 3,6,9) listed in the present study were not reportedin that list.
Gomez (2003) reported that 268 dinoflagellate specieshad been recorded in the Levantine Basin in hisliterature study comparing checklists of free-livingdinoflagellates in the Mediterranean sub basins.Although this was the highest number of speciesreported for the dinoflagellates in the Levantine Basin,some species (over 20 species) recorded in the presentstudy have not been observed previously in theLevantine Basin, and such species as Heterodinium
angulatum and Histioneis elongata have not beenreported before in the Mediterranean Sea according tothe list presented by Gomez (2003). While 604 dino-flagellate species have been reported in the westernMediterranean, this number is 480 for the easternMediterranean (Gomez 2003), probably reflecting thescarcity of studies conducted in the Levantine Basin.However, it should be remembered that species areintroduced to the Mediterranean both through the Straitof Gibraltar from the Atlantic Ocean and through theSuez Canal from the Red Sea. In addition to Red Seaand Atlantic migrants, species are accidentally intro-duced into this environment via ship fouling and ballastwaters. The high temperature and salinity in theMediterranean could facilitate the adaptation of intro-duced tropical species. All these processes have affectedthe biodiversity of this environment. In recent years, thefinding of the Indo-Pacific Histioneis elongata andtropical Pacific Heterodinium angulatum in northernLevantine waters supports the views above about theintroduction of exotic species.
Dinoflagellate abundance was generally low in thearea. Data showed that dinoflagellate abundance anddistribution are related with hydrographic properties
ARTICLE IN PRESS
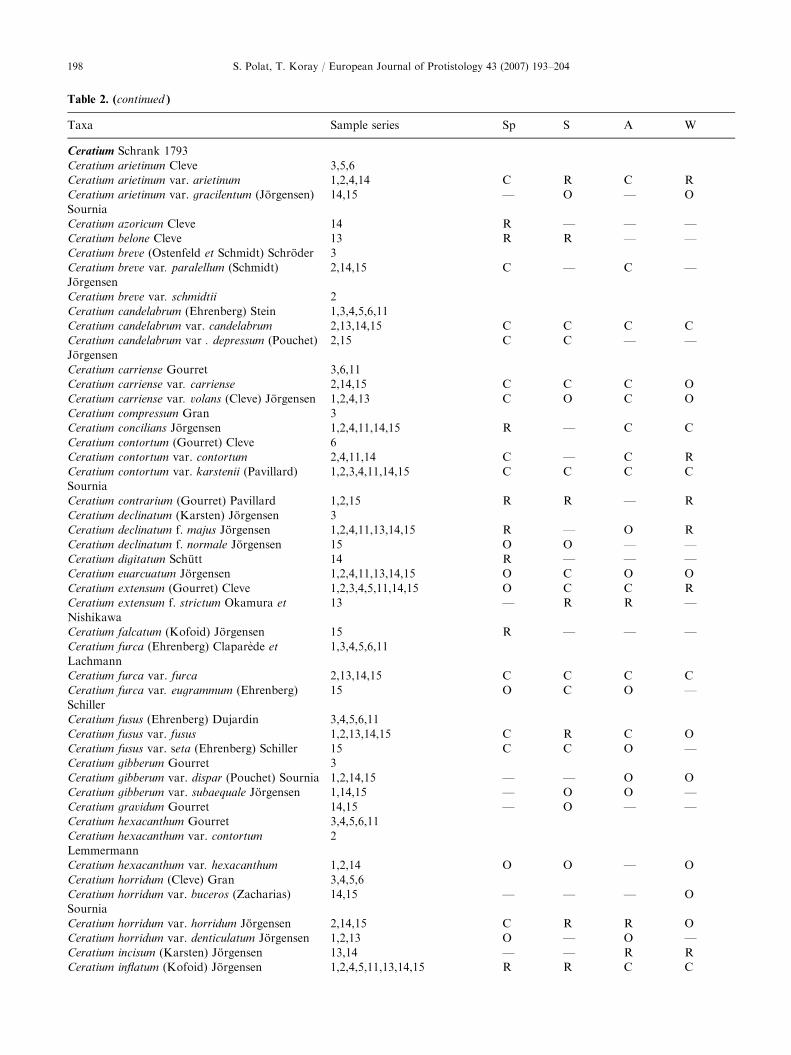
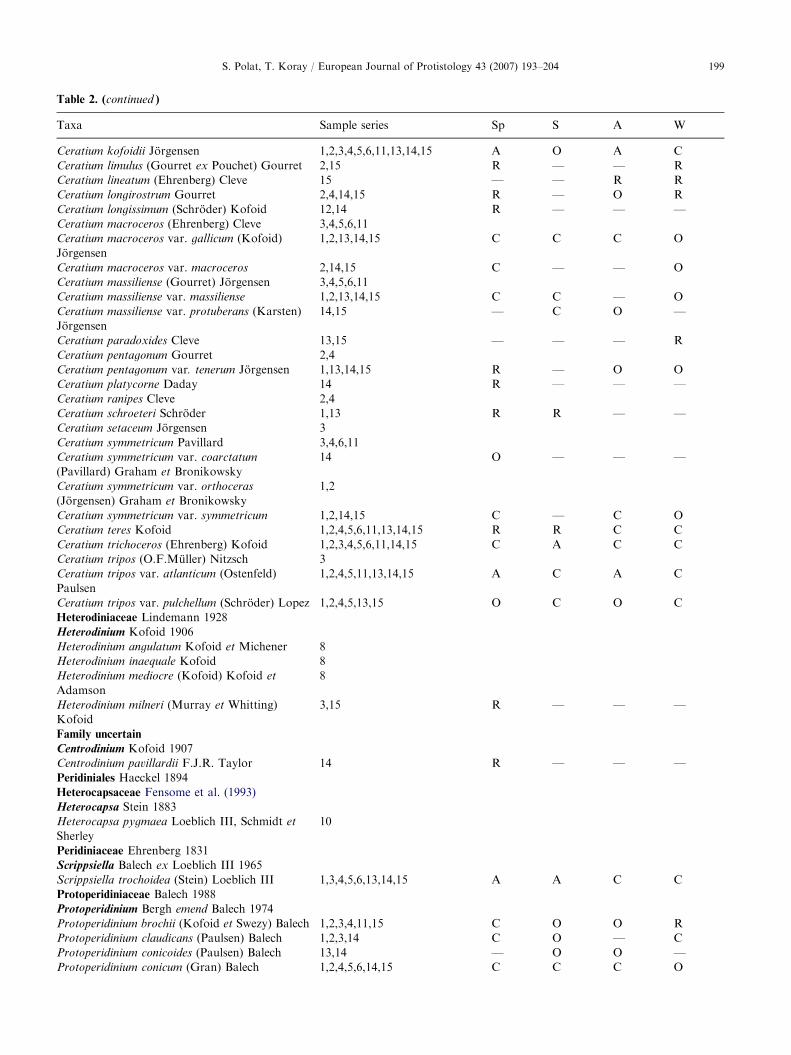
Table 2. List of dinoflagellate taxa, the sampling series (see Table 1) in which they were found and relative abundance of the taxa
during different seasons (Abbreviations used: Sp ¼ spring, S ¼ summer, A ¼ autumn, W ¼ winter, A ¼ abundant, 61–100%;
C ¼ common, 41–60%; O ¼ occasional, 21–40%; R ¼ rare, 1–20% of samples; — ¼ not found)�
Taxa Sample series Sp S A W
DINOPHYCEAE
Gymnodiniales Apstein 1909
Gymnodiniaceae Lankester 1885
Gyrodinium Kofoid et Swezy 1921
Gyrodinium fusiforme Kofoid et Swezy 3,14 R — — —
Gyrodinium lacryma (Meunier) Kofoid et Swezy 3
Akashiwo G.Hansen et Moestrup 2000
Akashiwo sanguinea (Hirasaka) G.Hansen et
Moestrup
3,13,14 R R — —
Polykrikaceae Kofoid et Swezy
Polykrikos Butschli 1873
Polykrikos schwartzii Butschli 3
Gonyaulacales Taylor 1980
Cladopyxiaceae Stein 1883
Cladopyxis Stein 1883
Cladopyxis caryophyllum (Kofoid) Pavillard 6,14,15 — R R —
Goniodomaceae Lindemann 1928
Goniodoma Stein 1883
Goniodoma acuminatum (Ehrenberg) Stein 1,2,4,5,6,11,13,14,15 A C C C
Goniodoma sphaericum Murray et Whitting 1,2,15 O — — R
Pyrophacus Stein 1883
Pyrophacus horologium Stein 2,3,15 — O O —
Pyrophacus steinii (Schiller) Wall et Dale 1,2,3,4,5,6,14,15 C A C —
Pyrocystaceae Apstein 1909
Pyrocystis Murray ex Haeckel 1890
Pyrocystis elegans Pavillard 1,6,15 — R R —
Pyrocystis fusiformis (Wyville Thomson ex
Haeckel) Blackman
2
Pyrocystis hamulus Cleve 14 — R — —
Pyrocystis robusta Kofoid 14 R — — —
Gonyaulacaceae Lindemann 1928
Lingulodinium Wall 1967
Lingulodinium polyedrum (Stein) Dodge 2,4,14,15 O C R R
Protoceratium Bergh 1881
Protoceratium areolatum Kofoid 14 — R — —
Protoceratium reticulatum (Claparede et
Lachmann) Butschli
11
Gonyaulax Diesing 1866
Gonyaulax birostris Stein 2
Gonyaulax diegensis Kofoid 1,2,3,4,14,15 R C R R
Gonyaulax digitale (Pouchet) Kofoid 1,3
Gonyaulax monocantha Pavillard 2,3
Gonyaulax pacifica Kofoid 12
Gonyaulax polygramma Stein 1,2,3,4,5,6,11,13,14,15 C O C C
Gonyaulax spinifera (Claparede et Lachmann)
Diesing
2,4,5,11,14,15 R C C R
Gonyaulax turbynei Murray et Whitting 2,3,14 — R R —
Spiraulax Kofoid 1911
Spiraulax jolliffei (Murray et Whitting) Kofoid 2,4,14 — O — C
Ceratocoryaceae Lindemann 1928
Ceratocorys Stein 1883
Ceratocorys armata (Schutt) Kofoid 1,2,15 — O — —
Ceratocorys gourretii Paulsen 1,2,4,6,15 — C R R
Ceratocorys horrida Stein 1,2,3,4,5,6,11,13,14,15 C A C C
Ceratiaceae Willey et Hickson 1909
S. Polat, T. Koray / European Journal of Protistology 43 (2007) 193–204 197
ARTICLE IN PRESS
Table 2. (continued )
Taxa Sample series Sp S A W
Ceratium Schrank 1793
Ceratium arietinum Cleve 3,5,6
Ceratium arietinum var. arietinum 1,2,4,14 C R C R
Ceratium arietinum var. gracilentum (Jorgensen)
Sournia
14,15 — O — O
Ceratium azoricum Cleve 14 R — — —
Ceratium belone Cleve 13 R R — —
Ceratium breve (Ostenfeld et Schmidt) Schroder 3
Ceratium breve var. paralellum (Schmidt)
Jorgensen
2,14,15 C — C —
Ceratium breve var. schmidtii 2
Ceratium candelabrum (Ehrenberg) Stein 1,3,4,5,6,11
Ceratium candelabrum var. candelabrum 2,13,14,15 C C C C
Ceratium candelabrum var . depressum (Pouchet)
Jorgensen
2,15 C C — —
Ceratium carriense Gourret 3,6,11
Ceratium carriense var. carriense 2,14,15 C C C O
Ceratium carriense var. volans (Cleve) Jorgensen 1,2,4,13 C O C O
Ceratium compressum Gran 3
Ceratium concilians Jorgensen 1,2,4,11,14,15 R — C C
Ceratium contortum (Gourret) Cleve 6
Ceratium contortum var. contortum 2,4,11,14 C — C R
Ceratium contortum var. karstenii (Pavillard)
Sournia
1,2,3,4,11,14,15 C C C C
Ceratium contrarium (Gourret) Pavillard 1,2,15 R R — R
Ceratium declinatum (Karsten) Jorgensen 3
Ceratium declinatum f. majus Jorgensen 1,2,4,11,13,14,15 R — O R
Ceratium declinatum f. normale Jorgensen 15 O O — —
Ceratium digitatum Schutt 14 R — — —
Ceratium euarcuatum Jorgensen 1,2,4,11,13,14,15 O C O O
Ceratium extensum (Gourret) Cleve 1,2,3,4,5,11,14,15 O C C R
Ceratium extensum f. strictum Okamura et
Nishikawa
13 — R R —
Ceratium falcatum (Kofoid) Jorgensen 15 R — — —
Ceratium furca (Ehrenberg) Claparede et
Lachmann
1,3,4,5,6,11
Ceratium furca var. furca 2,13,14,15 C C C C
Ceratium furca var. eugrammum (Ehrenberg)
Schiller
15 O C O —
Ceratium fusus (Ehrenberg) Dujardin 3,4,5,6,11
Ceratium fusus var. fusus 1,2,13,14,15 C R C O
Ceratium fusus var. seta (Ehrenberg) Schiller 15 C C O —
Ceratium gibberum Gourret 3
Ceratium gibberum var. dispar (Pouchet) Sournia 1,2,14,15 — — O O
Ceratium gibberum var. subaequale Jorgensen 1,14,15 — O O —
Ceratium gravidum Gourret 14,15 — O — —
Ceratium hexacanthum Gourret 3,4,5,6,11
Ceratium hexacanthum var. contortum
Lemmermann
2
Ceratium hexacanthum var. hexacanthum 1,2,14 O O — O
Ceratium horridum (Cleve) Gran 3,4,5,6
Ceratium horridum var. buceros (Zacharias)
Sournia
14,15 — — — O
Ceratium horridum var. horridum Jorgensen 2,14,15 C R R O
Ceratium horridum var. denticulatum Jorgensen 1,2,13 O — O —
Ceratium incisum (Karsten) Jorgensen 13,14 — — R R
Ceratium inflatum (Kofoid) Jorgensen 1,2,4,5,11,13,14,15 R R C C
S. Polat, T. Koray / European Journal of Protistology 43 (2007) 193–204198
ARTICLE IN PRESS
Table 2. (continued )
Taxa Sample series Sp S A W
Ceratium kofoidii Jorgensen 1,2,3,4,5,6,11,13,14,15 A O A C
Ceratium limulus (Gourret ex Pouchet) Gourret 2,15 R — — R
Ceratium lineatum (Ehrenberg) Cleve 15 — — R R
Ceratium longirostrum Gourret 2,4,14,15 R — O R
Ceratium longissimum (Schroder) Kofoid 12,14 R — — —
Ceratium macroceros (Ehrenberg) Cleve 3,4,5,6,11
Ceratium macroceros var. gallicum (Kofoid)
Jorgensen
1,2,13,14,15 C C C O
Ceratium macroceros var. macroceros 2,14,15 C — — O
Ceratium massiliense (Gourret) Jorgensen 3,4,5,6,11
Ceratium massiliense var. massiliense 1,2,13,14,15 C C — O
Ceratium massiliense var. protuberans (Karsten)
Jorgensen
14,15 — C O —
Ceratium paradoxides Cleve 13,15 — — — R
Ceratium pentagonum Gourret 2,4
Ceratium pentagonum var. tenerum Jorgensen 1,13,14,15 R — O O
Ceratium platycorne Daday 14 R — — —
Ceratium ranipes Cleve 2,4
Ceratium schroeteri Schroder 1,13 R R — —
Ceratium setaceum Jorgensen 3
Ceratium symmetricum Pavillard 3,4,6,11
Ceratium symmetricum var. coarctatum
(Pavillard) Graham et Bronikowsky
14 O — — —
Ceratium symmetricum var. orthoceras
(Jorgensen) Graham et Bronikowsky
1,2
Ceratium symmetricum var. symmetricum 1,2,14,15 C — C O
Ceratium teres Kofoid 1,2,4,5,6,11,13,14,15 R R C C
Ceratium trichoceros (Ehrenberg) Kofoid 1,2,3,4,5,6,11,14,15 C A C C
Ceratium tripos (O.F.Muller) Nitzsch 3
Ceratium tripos var. atlanticum (Ostenfeld)
Paulsen
1,2,4,5,11,13,14,15 A C A C
Ceratium tripos var. pulchellum (Schroder) Lopez 1,2,4,5,13,15 O C O C
Heterodiniaceae Lindemann 1928
Heterodinium Kofoid 1906
Heterodinium angulatum Kofoid et Michener 8
Heterodinium inaequale Kofoid 8
Heterodinium mediocre (Kofoid) Kofoid et
Adamson
8
Heterodinium milneri (Murray et Whitting)
Kofoid
3,15 R — — —
Family uncertain
Centrodinium Kofoid 1907
Centrodinium pavillardii F.J.R. Taylor 14 R — — —
Peridiniales Haeckel 1894
Heterocapsaceae Fensome et al. (1993)
Heterocapsa Stein 1883
Heterocapsa pygmaea Loeblich III, Schmidt et
Sherley
10
Peridiniaceae Ehrenberg 1831
Scrippsiella Balech ex Loeblich III 1965
Scrippsiella trochoidea (Stein) Loeblich III 1,3,4,5,6,13,14,15 A A C C
Protoperidiniaceae Balech 1988
Protoperidinium Bergh emend Balech 1974
Protoperidinium brochii (Kofoid et Swezy) Balech 1,2,3,4,11,15 C O O R
Protoperidinium claudicans (Paulsen) Balech 1,2,3,14 C O — C
Protoperidinium conicoides (Paulsen) Balech 13,14 — O O —
Protoperidinium conicum (Gran) Balech 1,2,4,5,6,14,15 C C C O
S. Polat, T. Koray / European Journal of Protistology 43 (2007) 193–204 199
ARTICLE IN PRESS
Table 2. (continued )
Taxa Sample series Sp S A W
Protoperidinium crassipes (Kofoid) Balech 1,3,13,14 O O — —
Protoperidinium depressum (Bailey) Balech 1,2,3,4,5,14,15 C C C —
Protoperidinium diabolus (Cleve) Balech 11,14,15 — O O —
Protoperidinium divergens (Ehrenberg) Balech 1,2,3,4,5,11,13,14,15 C A C C
Protoperidinium globulus (Stein) Balech 2,3,4,5,11,13 — — O C
Protoperidinium grande (Kofoid) Balech 2,3,15 — O O —
Protoperidinium granii (Ostenfeld) Balech 3,14,15 C C — —
Protoperidinium leonis (Pavillard) Balech 2,3
Protoperidinium mediterraneum (Kofoid) Balech 1,2,4,5,13,14 C C O C
Protoperidinium mite (Pavillard) Balech 14,15 — R O —
Protoperidinium oblongum (Aurivillius) Parke et
Dodge
3,15 — O O —
Protoperidinium oceanicum (Vanhoffen) Balech 1,2,3,4,5,6,11,15 C — C C
Protoperidinium oviforme (Dangeard) Balech 3,14,15 C C — —
Protoperidinium ovum (Schiller) Balech 14 — O — —
Protoperidinium pallidum (Ostenfeld) Balech 1,2,11,14,15 — C C O
Protoperidinium pedunculatum (Schutt) Balech 1,2,3,4,5,11,14,15 — C C O
Protoperidinium pellucidum Bergh 1,3,4,5,6,14 — C C O
Protoperidinium pentagonum (Gran) Balech 3,14,15 — O — —
Protoperidinium punctulatum (Paulsen)Balech 13,14 — O O —
Protoperidinium pyriforme (Paulsen) Balech 2,4,13,14 C C — A
Protoperidinium quarnerense (Schroder) Balech 2,3,4,13,14,15 O O C A
Protoperidinium solidicorne (Mangin)Balech 3
Protoperidinium steinii (Jorgensen) Balech 1,2,3,4,5,6,11,13,14,15 C A C R
Protoperidinium subinerme (Paulsen) Loeblich III 1,4,14,15 O O C O
Diplopelta Stein ex Jorgensen
Diplopelta bomba Stein ex Jorgensen 14,15 R R — —
Diplopsalis Bergh 1881
Diplopsalis lenticula Bergh 1,2,3,4,14,15 O C C R
Podolampaceae Lindemann 1928
Podolampas Stein 1883
Podolampas bipes Stein 1,2,4,5,6,13,14,15 R C C —
Podolampas elegans Schutt 11,14,15 — O C —
Podolampas palmipes Stein 14 — O O —
Podolampas spinifera Okamura 2,4,5,11,14 — C R R
Order uncertain
Oxytoxaceae Lindemann 1928
Oxytoxum Stein 1883
Oxytoxum compressum Kofoid 13,14 — O — —
Oxytoxum constrictum (Stein) Butschli 11,15 — O — —
Oxytoxum longiceps Schiller 14,15 — R R —
Oxytoxum milneri Murray et Whitting 13 — R R —
Oxytoxum reticulatum (Stein) Schutt 5,13,14 — O — —
Oxytoxum scolopax Stein 2,4,5,6,11,13,14,15 R C C R
Dinophysiales Kofoid 1926
Dinophysiaceae Stein 1883
Citharistes Stein 1883
Citharistes regius Stein 9
Dinophysis Ehrenberg 1839
Dinophysis acuta Ehrenberg 3
Dinophysis amandula Sournia 14 — O R —
Dinophysis argus (Stein) Abe 15 — R O O
Dinophysis caudata Saville-Kent 1,2,3,4,5,11,13,14,15 C C O C
Dinophysis diegensis Kofoid 3
Dinophysis doryphorum (Stein) Abe 1,2,14 — — — R
Dinophysis elongata (Jorgensen) Abe 13 R R — —
Dinophysis favus (Kofoid et Mich.) Abe 14,15 — O — —
S. Polat, T. Koray / European Journal of Protistology 43 (2007) 193–204200
ARTICLE IN PRESS
Table 2. (continued )
Taxa Sample series Sp S A W
Dinophysis hastata Stein 1,2,11,14 C C — —
Dinophysis mitra (Schutt) Abe 2,5,14 — — O O
Dinophysis odiosa (Pavillard) Tai et Skogsberg 4
Dinophysis ovum Schutt 1,11
Dinophysis parvula (Schutt) Balech 1,4
Dinophysis recurva Kofoid et Skogsberg 4
Dinophysis rapa (Stein) Abe 1,11,14 O O R —
Dinophysis rotundata Claparede et Lachmann 5,6,11,14 O O R —
Dinophysis schuettii Murray et Whitting 13,14 — O O —
Dinophysis sphaerica Stein 13,14 O O — —
Dinophysis tripos Gourret 3,14 O R — —
Histioneis Stein 1883
Histioneis depressa Schiller 7
Histioneis elongata Kofoid et Michener 7,15 R — — —
Histioneis expansa Rampi 7
Histioneis marchesonii Rampi 7
Histioneis oxypteris Schiller 14 — R — —
Histioneis striata Kofoid et Michener 7
Histioneis para Murray et Whitting 7
Ornithocercus Stein 1883
Ornithocercus carolinae Kofoid 14 — R — —
Ornithocercus heteroporus Kofoid 11,13, 14 O O R —
Ornithocercus magnificus Stein 1,4,5,6,13,14 — R C C
Ornithocercus quadratus Schutt 1,2,3,4,11,14,15 — C C O
Ornithocercus splendidus Schutt 13,14 R — — —
Ornithocercus steinii Schutt emend.Kofoid et
Skogsberg
14 C C — —
Ornithocercus thumii (Schmidt) Kofoid et
Skogsberg
15 — R — —
Amphisoleniaceae Lindemann 1928
Amphisolenia Stein 1883
Amphisolenia bidentata Schroder 1,2,4,6,11,14 C — C O
Amphisolenia palaeotheroides Kofoid 15 — — R —
Amphisolenia truncata Kofoid et Michener 2
Prorocentrales Lemmermann 1910
Prorocentraceae Stein 1883
Prorocentrum Ehrenberg 1834
Prorocentrum compressum (Bailey) Abe ex Dodge 1,2,4,14 C O — —
Prorocentrum cordatum (Ostenfeld) Dodge 3
Prorocentrum micans Ehrenberg 1,2,3,4,5,11,13,14,15 A C C C
NOCTILUCIPHYCEAE
Noctilucales Haeckel 1894
Noctilucaceae Saville-Kent 1881
Noctiluca Suriray in Lamarck 1816
Noctiluca scintillans (Macartney) Kofoid 13,14,15 R C — C
Kofoidiniaceae Taylor 1976
Kofoidinium Pavillard 1928
Kofoidinium velelloides Pavillard 2,4,5,11,14,15 O C — O
�The relative abundance of the taxa was determined by using sample series of 13–15.
S. Polat, T. Koray / European Journal of Protistology 43 (2007) 193–204 201
and specific species requirements. In the present study,the most abundant species was S. trochoidea and thehighest abundance of this species was recorded in theeastern parts of Iskenderun Bay where there were heavyland-based nutrient inputs. This suggests that nutrient
richness contributes to the increase of this small species.Similarly, Licea et al. (2004) concluded that nutrientscontributed to the bloom of S. trochoidea in the Gulf ofMexico. On the other hand, large species which arecharacteristic of nutrient-poor waters (Vila and Maso
ARTICLE IN PRESS
Figs 2–10. Photomicrographs of some rare dinoflagellate species. (2) Ceratium longissimum, (3) Ceratium platycorne, (4) Ceratium
extensum f. strictum, (5) Ceratium paradoxides, (6a) Amphisolenia palaeotheroides, (6b) A. palaeotheroides (antapex),
(7) Ornithocercus splendidus, (8) Ornithocercus carolinae, (9) Pyrocystis hamulus, (10) Centrodinium pavillardii. All scale
bars ¼ 30mm.
S. Polat, T. Koray / European Journal of Protistology 43 (2007) 193–204202

ARTICLE IN PRESS
Figs 11 and 12. Photomicrographs of two further rare
dinoflagellate species. (11) Heterodinium angulatum, (12)
Histioneis elongata. Scale bars ¼ 30mm.
S. Polat, T. Koray / European Journal of Protistology 43 (2007) 193–204 203
2005) were widely distributed, but they were in lowabundances even in nutrient rich coastal waters. More-over, despite the presence of certain species that hadbeen reported to be responsible for red tides or otherharmful blooms in other areas, no red tides wererecorded in the region through the time periodinvestigated.
Comparison of the results of the present study withother studies and those conducted in the other parts ofthe Levantine Basin, leads to the conclusion that thespecies diversity in the region has displayed changeseven if these are modest. It was reported that water
temperature in the Mediterranean has increased due toglobal warming and local influences (Turley 1999) andthat this increase could have resulted in alterations inthe marine biodiversity (Gomez and Claustre 2003). Insuch conditions, while some species perish, some warmwater species entering the Basin might increase. Changesin phytoplankton biodiversity are also likely to be infavour of the noxious species. Most of the studiesconducted on the changes in biodiversity in theMediterranean have focused on macroscopic organisms(see Gomez and Claustre 2003). Comprehensive taxo-nomic studies conducted with the aim of determiningMediterranean phytoplankton biodiversity will alsocontribute to the identification of community changestaking place with the passage of time.
References
Avs-ar, D., Polat, S., Is-ık, O., 1998. An investigation of summer
phytoplankton in Yumurtalık Bight (Adana). XVI Na-
tional Biology Congress, Proceedings, vol. II, pp. 227–239
(in Turkish).
Azov, Y., 1991. Eastern Mediterranean – a marine desert?
Mar. Pollut. Bull. 23, 225–232.
Delgado, M., Fortuna, J.M., 1991. Atlas de Fitoplancton del
Mar Mediterraneo. Sci. Mar. 55, 1–133.
Dowidar, N.M., 1974. The phytoplankton of the Mediter-
anean waters of Egypt. I. A checklist of the species
recorded. Bull. Inst. Oceanogr. Fish. Cairo 4, 319–344.
Eker, E., Kıdeys-, A.E., 2000. Weekly variations in phyto-
plankton structure of a harbour in Mersin Bay (north-
eastern Mediterranean). Turk. J. Bot. 24, 13–24.
El-Maghraby, A.M., Halim, Y., 1965. A quantitative and
qualitative study of phytoplankton of Alexandria waters.
Hydrobiologia 25, 221–238.
Fensome, R.A., Taylor, F.J.R., Norris, G., Sarjeant, W.A.S.,
Wharton, D.I., Williams, G.L., 1993. A Classification of
Living and Fossil Dinoflagellates. Sheridan Press, Hanover.
Fritz, L., Triemer, R.E., 1985. A rapid simple technique
utilizing calcofluor white M2R for the visualisation of
dinoflagellate thecal plates. J. Phycol. 21, 662–664.
Goffart, A., Hecq, J-H., Legendre, L., 2002. Changes in the
development of the winter-spring phytoplankton bloom in
the Bay of Calvi (NW Mediterranean) over the last two
decades: a response to changing climate? Mar. Ecol. Prog.
Ser. 236, 45–60.
Gomez, F., 2003. Checklist of Mediterranean free-living
dinoflagellates. Bot. Mar. 46, 215–242.
Gomez, F., 2005. Endemic and Indo-Pacific plankton in the
Mediterranean Sea: a study based on dinoflagellate records.
J. Biogeogr. 33, 261–270.
Gomez, F., Claustre, H., 2003. The genus Asterodinium
(Dinophyceae) as a possible biological indicator of warm-
ing in the western Mediterranean. J. Mar. Biol. Assoc. UK
83, 173–174.
Gomez, F., Gorsky, J.R.V., 2003. Microplankton annual
cycles in the Bay of Villefranche, Ligurian Sea, NW
Mediterranean Sea. J. Plankton Res. 25, 323–339.

ARTICLE IN PRESSS. Polat, T. Koray / European Journal of Protistology 43 (2007) 193–204204
Halim, Y., 1963. Microplancton des eaux egyptiennes, Le
genre Ceratium Schrank (Dinoflagellates). P.-v. Reun.
Commn. int. Explor. Mer. Medit. 17, 495–502.
Hallegraeff, G.M., 1993. A review of harmful algal blooms and
their apparent global increase. Phycologia 32, 79–99.
Ignatiades, L., Georgopoulos, D., Karydis, M., 1995. Descrip-
tion of the phytoplanktonic community of the oligotrophic
waters of the SE Aegean Sea (Mediterranean). P.S.Z.N.I.
Mar. Ecol. 16, 13–26.
Kıdeys-, A.E., Unsal, M., Bingel, F., 1989. Seasonal changes in
net phytoplankton off Erdemli, Northeastern Mediterra-
nean. Doga, Turk. J. Bot. 13, 45–54.
Kimor, B., Wood, B., 1975. A plankton study in the eastern
Mediterranean Sea. Mar. Biol. 29, 321–333.
Koray, T., 2001. A checklist for phytoplankton of Turkish
Seas. Ege.Univ. J. Fisheries Aquat. Sci. 18, 1–23.
Koray, T., Gokpınar, S- ., Polat, S., Turkoglu, M., Yurga, L.,
Colak, F., Benli, H.A., Sarıhan, E., 2000. Comparison of
the qualitative characteristics of microplankton (Protista)
of Turkish Seas (Black Sea, Aegean Sea, Northeastern
Mediterranean). Ege.Univ. J. Fisheries Aquat. Sci. 17,
231–247.
Krom, M.D., Kress, N., Brenner, N., Gordon, L.I., 1991.
Phosphorus limitation of primary productivity in the
eastern Mediterranean Sea. Limnol. Oceanogr. 36,
424–432.
Lakkis, S., Lakkis, V.N., 1980. Composition annual cycle and
species diversity of the phytoplankton in Lebanese coastal
water. J. Plankton Res. 3, 123–136.
Licea, S., Zamudio, M.E., Luna, R., Soto, J., 2004. Free-living
dinoflagellates in the southern Gulf of Mexico: report of
data (1979–2002). Phycol. Res. 52, 419–428.
Loeblich III, A.R., 1984. Dinoflagellate physiology and
biochemistry. In: Spector, D.L. (Ed.), Dinoflagellates.
Academic Press, New York, pp. 299–342.
Polat, S., 2002. Nutrients, chlorophyll a and phytoplankton in
the Iskenderun Bay (northeastern Mediterranean).
P.S.Z.N. Mar. Ecol. 23, 115–126.
Polat, S., 2004. New record for a dinoflagellate species
(Citharistes regius Stein) in the northern Levantine Basin
(eastern Mediterranean). Turk. J. Bot. 28, 507–509.
Polat, S., 2007. New record for a dinoflagellate species
(Gonyaulax pacifica Kofoid) from Turkish coastal waters
(northeastern Mediterranean Sea). Turk. J. Bot. 31, 67–70.
Polat, S., Is-ık, O., 2002. Phytoplankton distribution, diversity
and nutrients at the NE Mediterranean coast of Turkey
(Karatas--Adana). Turk. J. Bot. 26, 77–86.
Polat, S., Koray, T., 2002. New records of the genus Histioneis
F.R.Von Stein (Dinophyceae) from Turkish coastal waters.
Turk. J. Bot. 26, 481–484.
Polat, S., Koray, T., 2003. New records for the genus
Heterodinium Kofoid (Dinophyceae) from Turkish coastal
waters (north-eastern Mediterranean). Turk. J. Bot. 27,
427–430.
Polat, S., Piner, M.P., 2002a. Seasonal variations in biomass,
abundance and species diversity of phytoplankton in the
Iskenderun Bay (northeastern Mediterranean). Pak. J. Bot.
34, 101–112.
Polat, S., Piner, M.P., 2002b. Nutrients and phytoplankton in
the Babadıllimanı Bight, northeastern Mediterranean coast
of Turkey. Indian J. Mar. Sci. 31, 188–194.
Polat, S., Sarıhan, E., Koray, T., 2000. Seasonal changes in the
phytoplankton of the northeastern Mediterranean (Bay of
Iskenderun). Turk. J. Bot. 24, 1–12.
Polat, S., Akiz, A., Piner, M.P., 2004. Phytoplankton
composition of the Babadıllimanı Bight and offshore
waters (northeastern Mediterranean). V National Confer-
ence on Coastal and Marine Areas of Turkey, Proceedings,
vol. I, pp. 43–51 (in Turkish).
Rampi, L., Bernhard, M., 1980. Chiave per la Determinazione
delle Peridinee Pelagiche Mediterranee. CNEN-RT/BIO
(80)8.
Taylor, F.J.R., 1987. The Biology of Dinoflagellates. Botanical
Monographs. Blackwell Scientific Publications, Oxford.
Turley, C.M., 1999. The changing Mediterranean Sea – a
sensitive ecosystem? Prog. Oceanogr. 44, 387–400.
Turley, C.M., Bianchi, M., Christaki, U., Conan, P., Harris,
J.R.V., Psarra, S., Ruddy, G., Stutt, E.D., Tselepides, A.,
Wambeke, F.W., 2000. Relationship between primary
producers and bacteria in an oligotrophic sea – the
Mediterranean and biochemical implications. Mar. Ecol.
Prog. Ser. 193, 11–18.
Uysal, Z., Iwataki, M., Koray, T., 2003. On the presence of
Heterocapsa pygmaea A.R. Loebl. (Peridiniales, Dinophy-
ceae) in the northern Levantine Basin (Eastern Mediterra-
nean). Turk. J. Bot. 27, 149–152.
Vila, M., Maso, M., 2005. Phytoplankton functional groups
and harmful algal species in anthropogenically impac-
ted waters of the NW Mediterranean Sea. Sci. Mar. 69,
31–45.
Yılmaz, A., Bas-turk, O., Saydam, C., Ediger, D., Yılmaz, K.,
Hatipoglu, E., 1992. Eutrophication in Iskenderun Bay,
northeastern Mediterranean. Sci. Total Environ. (Suppl.),
705–771.
Related Documents











