Pituitary adenylyl cyclase-activating polypeptide controls the proliferation of retinal progenitor cells through downregulation of cyclin D1 Brian Njaine, Rodrigo A. P. Martins, Marcelo F. Santiago, Rafael Linden and Mariana S. Silveira Instituto de Biofisica Carlos Chagas Filho – UFRJ, Av. Carlos Chagas Filho, 373, Edifı ´cio do Centro de Ciencias da Saude, Bloco G, Cidade Universitaria, Ilha do Funda ˜ o, 21941-902 Rio de Janeiro, Brazil Keywords: cell cycle, development, PACAP, rat, retina Abstract During retinal development, cell proliferation and exit from the cell cycle must be precisely regulated to ensure the generation of the appropriate numbers and proportions of the various retinal cell types. Previously, we showed that pituitary adenylyl cyclase-activating polypeptide (PACAP) exerts a neuroprotective effect in the developing retina of rats, through the cAMP–cAMP-dependent protein kinase (protein kinase A) (PKA) pathway. Here, we show that PACAP also regulates the proliferation of retinal progenitor cells. PACAP, PACAP-specific receptor (PAC1), and the receptors activated by both PACAP and vasoactive intestinal peptide (VIP), VPAC1 and VPAC2, are expressed during embryonic and postnatal development of the rat retina. Treatment of retinal explants with PACAP38 reduced the incorporation of [ 3 H]thymidine as well as the number of 5-bromo-2¢-deoxyuridine-positive and cyclin D1-positive cells. Pharmacological experiments indicated that PACAP triggers this antiproliferative effect through the activation of both PAC1 and VPACs, and the cAMP–PKA pathway. In addition, PACAP receptor activation decreased both cyclin D1 mRNA and protein content. Altogether, the data support the hypothesis that PACAP is a cell-extrinsic regulator with multiple roles during retinal development, including the regulation of proliferation in a subpopulation of retinal progenitor cells. Introduction The vertebrate retina is composed of seven major cell types produced from multipotent retinal progenitor cells. These progenitor cells are located in the neuroblastic layer (NBL) of the developing retina, which corresponds to the ventricular zone (Turner & Cepko, 1987; Dyer & Cepko, 2001a). During development, retinal progenitors expand through cell proliferation, commit to distinct cell types and exit the cell cycle to generate either retinal neurons or the Mu ¨ller glia. In consequence, depending on the stage of development, increasing numbers of postmitotic cells are present in the neuroblastic layer on their way to acquiring their final identity. As early as the the first postnatal day in the rat, the ganglion cell layer (GCL) and a forming inner cell layer (INL) are observed in addition to the neuroblastic layer. The GCL contains ganglion cells (the first cell type generated in retinal tissue), which will be joined by displaced amacrine cells in the mature retina. The latter cells are the first to constitute the INL during retinal development. In the mature retina, this cell layer contains, besides amacrine cells, horizontal, bipolar and Mu ¨ller glia. Photore- ceptors (cones and rods) constitute the outer nuclear layer in the mature retina. Therefore, exit from the cell cycle must be carefully coordinated to ensure the generation of the correct numbers and proportions of the various cell types of the adult retina (Donovan & Dyer, 2005). Loss of control of these processes during development severely compromises retinal histogenesis (Sicinski et al., 1995; Burmeister et al., 1996; Levine et al., 2000; Dyer & Cepko, 2001a). The proliferation of retinal progenitors is controlled by both cell- intrinsic and cell-extrinsic factors. Multiple classes of secretable factors, such as sonic hedgehog (Shh), fibroblast growth factors, epidermal growth factor, transforming growth factor-a and transform- ing growth factor-b, as well as neurotransmitters, regulate the proliferation of retinal progenitors during development (Anchan et al., 1991; Lillien & Cepko, 1992; Anchan & Reh, 1995; Jensen & Wallace, 1997; Young & Cepko, 2004; Close et al., 2005; Martins et al., 2006; Martins & Pearson, 2007). Although compelling evidence has been provided for most of the cell-extrinsic factors listed above, little is known of how retinal progenitor cells integrate signals from these multiple regulators. Among the above, Shh-induced cell proliferation is antagonized by the intracellular messenger cAMP (Epstein et al., 1996; Hammerschmidt et al., 1996; Waschek et al., 2006). Therefore, cell-extrinsic factors that signal through cAMP- dependent pathways may contribute to control the proliferation of retinal progenitor cells, as previously suggested (Taylor & Reh, 1990). We have previously shown that pituitary adenylyl cyclase-activating polypeptide (PACAP) decreases the sensitivity to programmed cell death in the developing retina in a cAMP-dependent manner (Silveira et al., 2002). An mRNA-encoded PACAP precursor is processed to Correspondence: Mariana S. Silveira, as above. E-mail: [email protected] Received 22 July 2008, revised 8 April 2010, accepted 18 April 2010 European Journal of Neuroscience, pp. 1–10, 2010 doi:10.1111/j.1460-9568.2010.07286.x ª The Authors (2010). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing Ltd European Journal of Neuroscience

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Pituitary adenylyl cyclase-activating polypeptide controlsthe proliferation of retinal progenitor cells throughdownregulation of cyclin D1
Brian Njaine, Rodrigo A. P. Martins, Marcelo F. Santiago, Rafael Linden and Mariana S. SilveiraInstituto de Biofisica Carlos Chagas Filho – UFRJ, Av. Carlos Chagas Filho, 373, Edifıcio do Centro de Ciencias da Saude, Bloco G,Cidade Universitaria, Ilha do Fundao, 21941-902 Rio de Janeiro, Brazil
Keywords: cell cycle, development, PACAP, rat, retina
Abstract
During retinal development, cell proliferation and exit from the cell cycle must be precisely regulated to ensure the generation of theappropriate numbers and proportions of the various retinal cell types. Previously, we showed that pituitary adenylyl cyclase-activatingpolypeptide (PACAP) exerts a neuroprotective effect in the developing retina of rats, through the cAMP–cAMP-dependent proteinkinase (protein kinase A) (PKA) pathway. Here, we show that PACAP also regulates the proliferation of retinal progenitor cells.PACAP, PACAP-specific receptor (PAC1), and the receptors activated by both PACAP and vasoactive intestinal peptide (VIP),VPAC1 and VPAC2, are expressed during embryonic and postnatal development of the rat retina. Treatment of retinal explants withPACAP38 reduced the incorporation of [3H]thymidine as well as the number of 5-bromo-2¢-deoxyuridine-positive and cyclinD1-positive cells. Pharmacological experiments indicated that PACAP triggers this antiproliferative effect through the activation ofboth PAC1 and VPACs, and the cAMP–PKA pathway. In addition, PACAP receptor activation decreased both cyclin D1 mRNA andprotein content. Altogether, the data support the hypothesis that PACAP is a cell-extrinsic regulator with multiple roles during retinaldevelopment, including the regulation of proliferation in a subpopulation of retinal progenitor cells.
Introduction
The vertebrate retina is composed of seven major cell types producedfrom multipotent retinal progenitor cells. These progenitor cells arelocated in the neuroblastic layer (NBL) of the developing retina, whichcorresponds to the ventricular zone (Turner & Cepko, 1987; Dyer &Cepko, 2001a). During development, retinal progenitors expandthrough cell proliferation, commit to distinct cell types and exit thecell cycle to generate either retinal neurons or the Muller glia. Inconsequence, depending on the stage of development, increasingnumbers of postmitotic cells are present in the neuroblastic layer ontheir way to acquiring their final identity. As early as the the firstpostnatal day in the rat, the ganglion cell layer (GCL) and a forminginner cell layer (INL) are observed in addition to the neuroblasticlayer. The GCL contains ganglion cells (the first cell type generated inretinal tissue), which will be joined by displaced amacrine cells in themature retina. The latter cells are the first to constitute the INL duringretinal development. In the mature retina, this cell layer contains,besides amacrine cells, horizontal, bipolar and Muller glia. Photore-ceptors (cones and rods) constitute the outer nuclear layer in themature retina. Therefore, exit from the cell cycle must be carefullycoordinated to ensure the generation of the correct numbers and
proportions of the various cell types of the adult retina (Donovan &Dyer, 2005). Loss of control of these processes during developmentseverely compromises retinal histogenesis (Sicinski et al., 1995;Burmeister et al., 1996; Levine et al., 2000; Dyer & Cepko, 2001a).The proliferation of retinal progenitors is controlled by both cell-
intrinsic and cell-extrinsic factors. Multiple classes of secretablefactors, such as sonic hedgehog (Shh), fibroblast growth factors,epidermal growth factor, transforming growth factor-a and transform-ing growth factor-b, as well as neurotransmitters, regulate theproliferation of retinal progenitors during development (Anchan et al.,1991; Lillien & Cepko, 1992; Anchan & Reh, 1995; Jensen &Wallace, 1997; Young & Cepko, 2004; Close et al., 2005; Martinset al., 2006; Martins & Pearson, 2007). Although compelling evidencehas been provided for most of the cell-extrinsic factors listed above,little is known of how retinal progenitor cells integrate signals fromthese multiple regulators. Among the above, Shh-induced cellproliferation is antagonized by the intracellular messenger cAMP(Epstein et al., 1996; Hammerschmidt et al., 1996; Waschek et al.,2006). Therefore, cell-extrinsic factors that signal through cAMP-dependent pathways may contribute to control the proliferation ofretinal progenitor cells, as previously suggested (Taylor & Reh, 1990).We have previously shown that pituitary adenylyl cyclase-activating
polypeptide (PACAP) decreases the sensitivity to programmed celldeath in the developing retina in a cAMP-dependent manner (Silveiraet al., 2002). An mRNA-encoded PACAP precursor is processed to
Correspondence: Mariana S. Silveira, as above.E-mail: [email protected]
Received 22 July 2008, revised 8 April 2010, accepted 18 April 2010
European Journal of Neuroscience, pp. 1–10, 2010 doi:10.1111/j.1460-9568.2010.07286.x
ª The Authors (2010). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing Ltd
European Journal of Neuroscience

generate the two isoforms of PACAP, with either 38 or 27 amino acids,of which PACAP38 seems to be the most abundant in various tissues.(Arimura et al., 1991; Vaudry et al., 2000, 2009). PACAP peptidessignal through three distinct receptors. PAC1 is selective for PACAP,and VPAC1 and VPAC2 are activated by both PACAP and vasoactiveintestinal peptide (VIP). The sequence of human PACAP27 shareshigh identity (68%) with VIP, identifying PACAP as a member of theVIP–secretin–growth hormone releasing hormone–glucagon superfam-ily (Vaudry et al., 2000, 2009). VIP was first isolated as a vasoactiveagent in the small intestine (Said & Mutt, 1970). However, moreextensive studies have shown this peptide to be widely distributed,especially throughout the central nervous system (CNS) and peripheralnervous system, in a variety of species (Dickson & Finlayson, 2009).Multiple roles are attributed to PACAP peptides in the CNS,
including the control of cell proliferation during development (Wascheket al., 1998; DiCicco-Bloom et al., 2000; Silveira et al., 2002; Zhouet al., 2002; Fahrenkrug & Hannibal, 2004; Borba et al., 2005; Falluel-Morel et al., 2005). Opposing effects upon cell proliferation were,nonetheless, described for PACAP for sympathetic and cerebrocorticalprecursors. These correlated with the activation of differing signalingpathways by distinct isoforms of PAC1. In the peripheral nervoussystem, activation of phospholipase C led to increased cell proliferation,whereas in the CNS, activation of adenylyl cyclase and consequentcAMP production resulted in an antiproliferative effect (Lu & DiCicco-Bloom, 1997; Lu et al., 1998; Nicot & DiCicco-Bloom, 2001; Suhet al., 2001; Nicot et al., 2002).The present study was designed to examine the role of PACAP and
the signaling pathways involved in the control of proliferation ofretinal precursors.
Materials and methods
Materials
Dulbecco’s modified Eagle’s medium, 5% fetal bovine serum andTrizol were from Invitrogen (Calsbad,CA, USA). PACAP38 andVIP were from Bachem (Torrance, CA, USA). Anti-5-bromo-2¢-deoxyuridine (BrdU) monoclonal antibody, the first-strand cDNAkit and ECL-plus were from GE Healthcare (Little Chalfont, UK).The Apoptag Tunel kit was from Oncogene (Cambridge, MA,USA). Primary antibodies for cyclin D1, p27kip and phospho-cAMPresponse element-binding protein (pCREB), and secondary antibod-ies linked to horseradish peroxidase (HRP), were from CellSignaling (Beverly, MA, USA). Antibody for Erk2 was from SantaCruz Biotechnology (Santa Cruz, CA, USA). The monoclonalantibody anti-proliferating cell nuclear antigen (PCNA) was fromDako (Glostrup, Denmark). Fluorescent secondary antibodies usedwere Alexa Fluor 555-conjugated goat anti-rabbit and Alexa Fluor488-conjugated donkey anti-mouse (Molecular Probes, Eugene, OR,USA). Alternatively (Fig. 3) a biotinylated goat anti-mouse (Vec-tastain Elite ABC system; Vector Laboratories, Burlingame, CA,USA) was used with Cy3 tyramide amplification (Perkin Elmer LifeSciences, Waltham, MA, USA) and Sytox Green nuclear stain(Molecular Probes). PAC1 antibody was kindly provided by V. May(University of Vermont, USA). Maxadilan (Moro & Lerner, 1997),and M65 (Moro et al., 1999) were kindly provided by E. Lerner(Harvard Medical School, USA). IBMX, H89, poly(l-lysine), 4¢,6-diamidino-2-phenylindole and monoclonal antibody for a-tubulinwere from Sigma (Saint Louis, MO, USA). In addition,recombinant DNase I (Ambion, Austin, TX, USA) and PowerSYBR Green PCR Master Mix (Applied Biosystems, Foster City,CA, USA) were used.
Tissue culture and histology
All experimental procedures with animals were approved by theCommittee on Animal Experimentation of the Instituto de BiofısicaCarlos Chagas Filho, on the basis of the currently acceptedinternational rules. Retinal explants were maintained in vitro aspreviously described (Rehen et al., 1996; Varella et al., 1997, 1999;Silveira et al., 2002). In brief, eyes of P1 Lister hooded rat pupskilled by instantaneous decapitation were dissected, and retinas werecut into �1 mm2 pieces (total of eight explants per P1 retina). All ofthe explants in each experiment were pooled, and at least eightexplants were used for each replicate of each experimental group (oneor two replicates were used for each experimental group). Retinalexplants were maintained in Dulbecco’s modified Eagle’s mediumsupplemented with 5% fetal bovine serum, 2 mm glutamine,10 U ⁄ mL penicillin and 100 lg ⁄ mL streptomycin at pH 7.4 for24 h (except where noted) in Erlenmeyer flasks with orbital agitation.For histological and immunohistochemical analysis, at the end ofeach experiment the explants were fixed by immersion in 4%paraformaldehyde in sodium phosphate buffer (pH 7.4) for 2 h, andthen cryoprotected in 30% (w ⁄ v) sucrose in phosphate buffer.Transverse 10-lm-thick sections through the retinal tissue were cut ina cryostat.
Quantification of cell death and identification of apoptosis
Dead cells were recognized by their homogeneously condensed anddeeply stained chromatin among normal neighboring cells, afterstaining of explant sections with Neutral Red (Rehen et al., 1996).The apoptotic form of cell death was selectively examined by stainingdead cell profiles with the terminal deoxynucleotidyl transferase-mediated dUTP nick end-labeling (TUNEL) technique, using anApoptag kit. Counts of pyknotic profiles and TUNEL-positive cellswere made at ·1000 magnification under oil immersion in threerandom fields of 0.0148 mm2 within the neuroblastic layer for eachexplant, and at least three randomly selected explants were analyzedfor each group in each experiment. The data were plotted as number ofquantified events per mm2.
A
B
Fig. 1. PACAP and PACAP receptors are expressed during retinal develop-ment. Total RNA and protein were extracted from retinas of rats at variousstages of development. The expression of PACAP, PAC1, VPAC1 and VPAC2and b-actin was analyzed by RT-PCR (A). PAC1 protein content was analyzedby western blot (B). neg, negative control of the PCR reaction.
2 B. Njaine et al.
ª The Authors (2010). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 1–10

mRNA extraction, cDNA synthesis and RT-PCR
Total RNAwas prepared, with the use of Trizol, from rat retinas of thefollowing developmental stages: E16, E19, E21, P1, P3, P6, P10, P15,P22 and adult; the RNA was treated with recombinant DNase I, andcDNA was synthesized. For RT-PCR, amplification was performedwith a pair of primers that recognize a region from the C-terminalintracellular domain common to all isoforms of PAC1 (5¢-CAC-AGTATTCGCCTTCTCTCC-3¢ and 5¢-GCCTATCCCTATCTCTC-TCTT-3¢), and with the following pairs of primers for VPACs andPACAP: VPAC1, 5¢-CGGCCACCCGACATTGGGAAG-3¢ and 5¢-CTGCATGTGGCGCCGTTGCTG-3¢; VPAC2, 5¢-AAGCTAACTTC-TCCAGATGTT-3¢ and 5¢-CAGCTAAATGACTGAGGTCTC-3¢; andpro-PACAP, 5¢-ATGCCTCTCTGGTTGTGATTG-3¢ and 5¢-CGCTA-TTCGGTCCTTTGTT-3¢ (Braas et al., 1998; Braas & May, 1999). Forquantitative RT-PCR (Q-RT-PCR), RNA extraction and cDNAsynthesis were performed as described above.
Q-RT-PCR
For Q-RT-PCR, each replicate of each experimental group contained atleast eight explants from P1 retinas. Q-RT-PCR reactions wereperformed with Power SYBR Green PCR Master Mix, and theanalysis was performed with ABI7500 (Applied Biosystems). Thegene-specific primers for cyclin D1, p27kip1 and actin used for Q-RT-PCR analysis are listed in Table 1. Control without reverse transcrip-tase was included to ensure that the results were not caused byamplification of genomic DNA. PCR product identity was confirmedby electrophoresis and by melting-point analysis. The expressionvalue of each gene was normalized to the amount of actin gene(endogenous control), and a relative amount of transcript wascalculated with reference to control samples. The comparativeCt (DDCt) method was used to determine the target quantity insample as compared with endogenous control.
Immunohistochemistry
For immunohistochemistry of cyclin D1 and pCREB, retinal explantsfrom rats were maintained in vitro with culture medium for 1 h. Afterthis period, the explants were incubated for 5 or 15 min either with orwithout 10 nm PACAP38 (Silveira et al., 2002), and sections wereprocessed with rabbit anti-pCREB (1 : 100 rabbit antibody) andmouse anti-cyclin D1 (1 : 100 mouse antibody). For PCNA and PAC1immunohistochemistry, P1 sections were processed with anti-PAC1(1 : 100 rabbit antibody) and anti-PCNA (1 : 100 monoclonal mouseantibody). TO-PRO or 4¢,6-diamidino-2-phenylindole were used fornuclear staining. All immunohistochemical reactions were developedwith Alexa Fluor conjugate secondary antibodies (1 : 200 AlexaFluor, 555-conjugated goat anti-rabbit, and 1 : 200 Alexa Fluor 488-conjugated donkey anti-mouse). For quantitation of cyclin D1-positivecells, after PACAP treatment, retinal explants were incubated for5 min with 0.05% trypsin at 37�C and mechanically dissociated toform a single-cell suspension, as described previously (Martins et al.,
2008). The dissociated cells were allowed to adhere to slidespreviously covered with poly(l-lysine), and immunocytochemistryfor cyclin D1 (1 : 100 mouse antibody) was performed, with DABused for development. Five hundred cells were counted in each ofthree independent assays.
Incorporation of [3H]thymidine and BrdU pulse-labelingproliferative assays
Retinal explants were incubated for 24 h either with or without 10 nm
PACAP38 (Silveira et al., 2002), 10 nm maxadilan (Silveira et al.,2002), 50 nm M65 (Gower et al., 2003) or 100 lm 3-isobutyl-1-methylxanthine (IBMX) (Borba et al., 2005). Antagonists andinhibitors were added 30 min before PACAP38. [3H]thymidine(1 lCi ⁄ mL) was added in the last 2 h, and retinal explants werewashed with phosphate-buffered saline (pH 7.4) and homogenized in0.4 m NaOH. Aliquots of homogenates were precipitated with 10%trichloroacetic acid and filtered, and filters were washed with ethanol,dried at 100�C in an oven and counted in a Packard model 1600TRliquid scintillation analyzer. These measurements were performed induplicate in each experiment. Another aliquot of the homogenate wasused in triplicate for protein determination with the Lowry method.The results were then presented in disintegrations per minute (DPM)normalized for total protein. For BrdU pulse-labeling, retinal explantswere similarly treated for 24 h, and 10 lm BrdU was added in the last2 h. Retinal explants were then treated for 5 min with 0.05% trypsin at37�C, and mechanically dissociated to form a single-cell suspension.Cells were then plated on poly(l-lysine)-treated eight-well glass slidesbefore fixation, and counted after the detection of BrdU withimmunofluorescence (1 : 3 mouse monoclonal antibody). Five hun-dred cells were counted from each experimental group, and thenumber of BrdU-positive cells and BrdU labeling index (% BrdU-positive cells ⁄ total) were determined.
Image acquisition
Images were acquired with a confocal microscope (LSM 510 META;Zeiss GmbH, Jena, Germany), using the 488-nm band of an Ar laser andthe 543-nmbandof anHe ⁄ Ne laser for excitation ofAlexa Fluor 488 andAlexa Fluor 555, respectively. Proper filter sets were used for specificdetection of each fluorochrome at a confocal section width of 0.5–1.5 lm. Confocal images (Figs 2 and 5; Fig. S1) were acquired with aPlan-Neofluar ·40 ⁄ 1.3 Oil DIC objective, with voxel sizes (x, y and z innm) in the green channel of 46, 46 and 165, in the red channel of 52, 52and 184, and in the blue channel of 60, 60 and 214 (Fig. 2 only), and forFig. S1, 42, 42 and 113 the in green channel and 52, 52 and 184 in the redchannel. The fluorescent images in Fig. 3 were acquired with a Plan-Neofluar ·40 ⁄ 1.3 objective and AxioVision 3.1 software, with anAxioCamHRcmounted on anAxiophotmicroscope (Zeiss GmbH). Forthis, a secondary antibody (1 : 200 biotinylated goat anti-mouse) wasused with Cy3 tyramide amplification (1 : 150), and Sytox Greennuclear stain. The images in Fig. 7 were acquired with a Plan-Neofluar
Table 1. Quantitative RT-PCR primer sequences
Gene Accession no. Forward primer Reverse primer Product size (bp)
CyclinD1 NM_171992 GAGATTGTGCCATCCATGCGGAAA GAGACAAGAAACGGTCCAGGTAGT 121p27kip1 NM_031762 AGCTTGCCCGAGTTCTACTACAGA TTTGCCTGAGACCCAATTGAAGGC 125Actin NM_031144 TCATGAAGTGTGACGTTGACATCCGT CCTAGAAGCATTTGCGGTGCACGATG 285
PACAP is antiproliferative in the retina 3
ª The Authors (2010). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 1–10

A
E
B
F
C
G
D
H
I
Fig. 2. PAC1 is expressed in retinal progenitors. PAC1 expression in retinal progenitor cells was analyzed in sections from retinal explants from P1 rats that werefixed immediately after dissection. Confocal images were obtained after (B) PAC1 and (C) PCNA immunohistochemistry. (A) Nuclear and (D) overlay images. Forquantification of PAC1-expressing retinal progenitors, explants were mechanically dissociated to single cells and the cells were analyzed by immunocytochemistryfor PAC1 and PCNA (E–H). Five hundred cells were counted in each of three independent experiments, and the numbers of PAC1-positive and PCNA-positive cellswere determined (I). Arrows indicate double-positive cells, and insets show zoom images of the area indicated in the squares. Scale bar: 20 lm. GCL, ganglion celllayer; INL, inner nuclear layer; NBL, neuroblastic layer.
4 B. Njaine et al.
ª The Authors (2010). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 1–10

·40 ⁄ 0.75 objective with AxioVision 3.1 software, with an AxioCamHRc mounted on an Axiophot microscope, (Zeiss GmbH).
Western blots
Retinal explants were washed with phosphate-buffered saline, andtotal retinal protein was extracted (10 mm Tris–HCl, 150 mm NaCl,1% NP-40, 1% Triton X-100, 5 mm EDTA, 0.1% sodium dodecyl-sulfate, 1% sodium deoxycholate, 1 mm phenylmethylsulfonyl fluo-ride, 1 lg ⁄ mL aprotinin, 1 lg ⁄ mL pepstatin, 1 lg ⁄ mL leupeptin, 1%sodium orthonovadate and 50 mm sodium fluoride). The concentra-tions of lysates were determined with a Lowry assay (Lowry et al.,1951). Lysates (30 lg) were electrophoresed in polyacrylamide–sodium dodecylsulfate gels and transferred to nitrocellulose mem-branes. Membranes were blocked with either 5% bovine serumalbumin or 5% non-fat dry milk, and incubated with primaryantibodies for cyclin D1 (1 : 1000 mouse antibody), p27kip1
(1 : 1000 rabbit antibody), a-tubulin (1 : 50 000 mouse antibody) orErk2 (1 : 5000 rabbit antibody), and secondary antibodies (1 : 2000goat HRP-conjugated anti-rabbit and 1 : 2000 horse HRP-conjugatedanti-mouse), and developed with the ECL-Plus system according tothe manufacturer’s instructions.
Statistical analyses
Statistical comparisons between two experimental groups were madewith Student’s two-tailed unpaired t-test. For experiments with morethan two groups, either one-way or two-way anova tests were used, as
appropriate, followed by Tukey or Bonferoni multiple comparisonstests, respectively. Results are reported as means ± standard errors ofthe mean (SEMs), and a was set to 0.05. For all analyses, prism 4.0software (GraphPad Software, San Diego, CA, USA) was used.
Results
Expression of PACAP and PACAP receptors in the developingretina
To test whether PACAP and its receptors are expressed duringdevelopment of the rat retina, we carried out RT-PCR for mRNA thatencodes the PACAP precursor, the propeptide that is further processedpost-translationally, and for the mRNAs of PAC1, VPAC1 and VPAC2at various developmental stages. PAC1 was detected at the earlieststage studied (E16), and then throughout postnatal development and inadult tissue. The mRNA for the precursor of PACAP, VPAC1 andVPAC2 were detected at all stages, except for E16 (Fig. 1A). Weconfirmed protein expression of PAC1 by western blot. In agreementwith the RT-PCR analysis, PAC1 was detected at all ages tested(Fig. 1B).
Retinal progenitor cells express PAC1
Our previous study demonstrated the expression of PAC1 in boththe neuroblastic layer and in the GCL of P1 retinas of the rat,suggesting that PACAP receptors are expressed in retinal pro-genitors, as well as in postmitotic retinal cells (Silveira et al.,2002). To directly test whether PAC1 is expressed in retinalprogenitor cells, we immunostained retinal explants for PCNA(which labels the population of proliferating cells) and for PAC1.Double labeling demonstrated that retinal progenitor cells expressPAC1 (Fig. 2A–D). The same analysis was also performed withBrdU labeling of retinal progenitors (Fig. S1), which showed thatPAC1 is present in retinal progenitors as soon as E16. Moreover,after dissociation of retinal explants for quantification of progenitorcells that were positive for both PAC1 and PCNA (Fig. 2E–H), itwas demonstrated that the vast majority of PCNA-positive cellswere PAC1-positive (approximately 85% of all PCNA-positivecells) (Fig. 2I).
PACAP38 modulates cell proliferation in the developing retinaltissue
To determine whether PACAP38 modulates the proliferation ofprogenitor cells, retinal explants were cultured for 24 h in the pres-ence or absence of PACAP38. For the last 2 h of the experiment,[3H]thymidine was added to label cells in S phase of the cell cycle.Treatment with 10 nm PACAP38 decreased the incorporation of[3H]thymidine [Fig. 3A; t-test; n = 4; P = 0.01; degrees of freedom(d.f.) = 6]. We extended this finding by demonstrating that treatmentwith PACAP38 also reduced the proportion of retinal progenitor cellsthat incorporated BrdU added during the last 2 h of the culture period.Activation of PACAP receptors reduced by approximately 50% theproportion of BrdU-incorporating cells in P1 retinas cultured for 24 h(Fig. 3B–D) (Fig. 3D; t-test; n = 3; P = 0.037; d.f. = 4). Similarresults were obtained with E21 retinas (data not shown). This decreasein cell proliferation was not caused by the death of retinal progenitorsinduced by the peptide. We found no significant differences in thenumbers of either pyknotic nuclei (control, 330.7 ± 102.5 ⁄ mm2;PACAP, 260.7 ± 104.3 ⁄ mm2; t-test; n = 6; P = 0.11) or of TUNEL-
A
B
C
D
Fig. 3. PACAP inhibits DNA synthesis. Retinal explants from newborn P1rats were maintained in vitro either in the presence or in the absence of 10 nm
PACAP38 for 24 h. Proliferation was examined by incorporation of [3H]thy-midine (1 lCi ⁄ mL) present during the last 2 h, and liquid scintillation countswere normalized for protein content (A). For BrdU assay, similarly to the[3H]thymidine experiment, 10 lm BrdU was present during the last 2 h; retinalexplants were then mechanically dissociated, and the single cells were analyzedby immunocytochemistry for BrdU (B and C). Five hundred cells were countedin each of three independent experiments, and the BrdU labeling index isindicated as the percentage of BrdU-positive cells over the total cell number(D). The results in A are presented as means ± SEM pooled from fourindependent experiments. #P < 0.05 vs. control in t-test.
PACAP is antiproliferative in the retina 5
ª The Authors (2010). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 1–10
positive cells (control, 440.0 ± 140.3 ⁄ mm2; PACAP, 508.0 ±41.49 ⁄ mm2; t-test; n = 3; P = 0.88) following treatment of P1 retinalexplants with PACAP for 24 h.
Pharmacology of PACAP in the control of cell proliferationin the developing retina
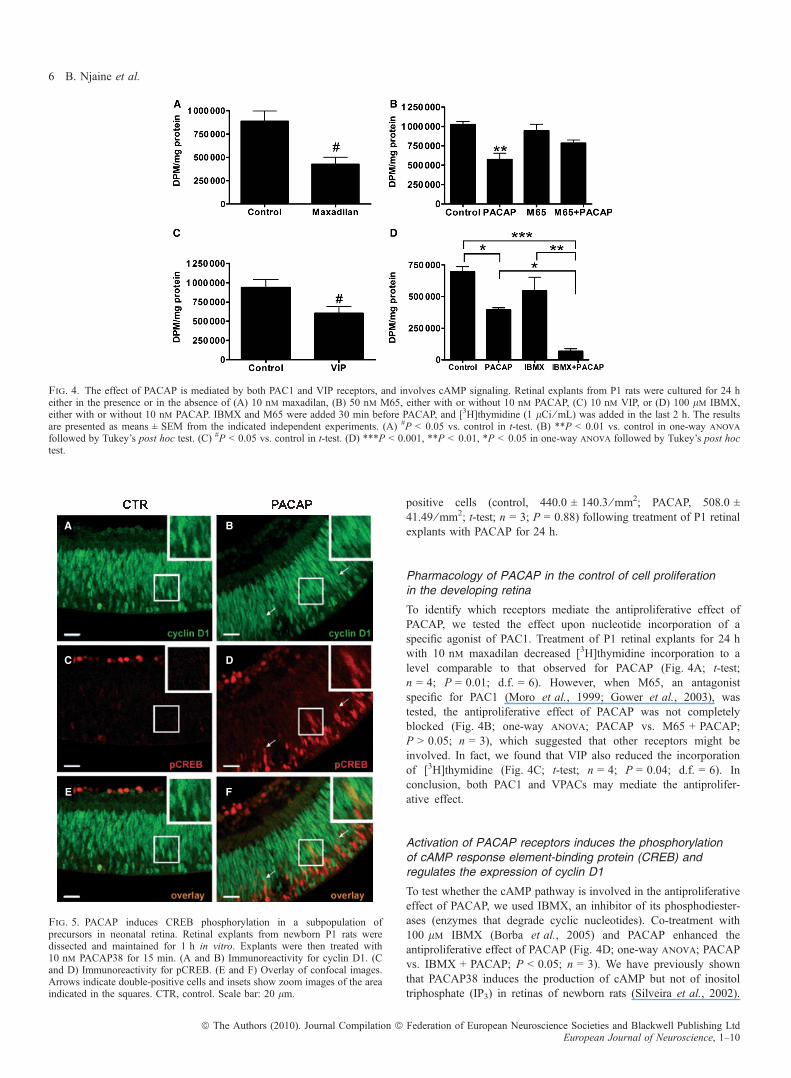
To identify which receptors mediate the antiproliferative effect ofPACAP, we tested the effect upon nucleotide incorporation of aspecific agonist of PAC1. Treatment of P1 retinal explants for 24 hwith 10 nm maxadilan decreased [3H]thymidine incorporation to alevel comparable to that observed for PACAP (Fig. 4A; t-test;n = 4; P = 0.01; d.f. = 6). However, when M65, an antagonistspecific for PAC1 (Moro et al., 1999; Gower et al., 2003), wastested, the antiproliferative effect of PACAP was not completelyblocked (Fig. 4B; one-way anova; PACAP vs. M65 + PACAP;P > 0.05; n = 3), which suggested that other receptors might beinvolved. In fact, we found that VIP also reduced the incorporationof [3H]thymidine (Fig. 4C; t-test; n = 4; P = 0.04; d.f. = 6). Inconclusion, both PAC1 and VPACs may mediate the antiprolifer-ative effect.
Activation of PACAP receptors induces the phosphorylationof cAMP response element-binding protein (CREB) andregulates the expression of cyclin D1
To test whether the cAMP pathway is involved in the antiproliferativeeffect of PACAP, we used IBMX, an inhibitor of its phosphodiester-ases (enzymes that degrade cyclic nucleotides). Co-treatment with100 lm IBMX (Borba et al., 2005) and PACAP enhanced theantiproliferative effect of PACAP (Fig. 4D; one-way anova; PACAPvs. IBMX + PACAP; P < 0.05; n = 3). We have previously shownthat PACAP38 induces the production of cAMP but not of inositoltriphosphate (IP3) in retinas of newborn rats (Silveira et al., 2002).
A
C
B
D
E F
Fig. 5. PACAP induces CREB phosphorylation in a subpopulation ofprecursors in neonatal retina. Retinal explants from newborn P1 rats weredissected and maintained for 1 h in vitro. Explants were then treated with10 nm PACAP38 for 15 min. (A and B) Immunoreactivity for cyclin D1. (Cand D) Immunoreactivity for pCREB. (E and F) Overlay of confocal images.Arrows indicate double-positive cells and insets show zoom images of the areaindicated in the squares. CTR, control. Scale bar: 20 lm.
Fig. 4. The effect of PACAP is mediated by both PAC1 and VIP receptors, and involves cAMP signaling. Retinal explants from P1 rats were cultured for 24 heither in the presence or in the absence of (A) 10 nm maxadilan, (B) 50 nm M65, either with or without 10 nm PACAP, (C) 10 nm VIP, or (D) 100 lm IBMX,either with or without 10 nm PACAP. IBMX and M65 were added 30 min before PACAP, and [3H]thymidine (1 lCi ⁄ mL) was added in the last 2 h. The resultsare presented as means ± SEM from the indicated independent experiments. (A) #P < 0.05 vs. control in t-test. (B) **P < 0.01 vs. control in one-way anova
followed by Tukey’s post hoc test. (C) #P < 0.05 vs. control in t-test. (D) ***P < 0.001, **P < 0.01, *P < 0.05 in one-way anova followed by Tukey’s post hoctest.
6 B. Njaine et al.
ª The Authors (2010). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 1–10

Altogether, these data suggest a role for cAMP in the antiproliferativeeffect of PACAP38.
The transcription factor CREB may be phosphorylated by cAMP-dependent protein kinase (protein kinase A; PKA) after cAMPproduction (Mayr & Montminy, 2001). To test for mechanismsdownstream of the activation of PAC1, we examined whetheractivation of PACAP receptors in retinal progenitor cells leads toCREB phosphorylation, which would be expected following thePACAP-induced increase in cAMP levels (Silveira et al., 2002).Retinal explants were cultured for 1 h, and then either PACAP38 orvehicle was added for 5 or 15 min. Sections from retinal explants werethen immunolabeled for pCREB (Fig. 5C and D) and cyclin D1, whichlabels most progenitor cells in developing retinal tissue (Fig. 5A andB). Treatment with PACAP resulted in pCREB immunostaining of cellnuclei located within the neuroblastic layer at both 5 min (data notshown) and 15 min after PACAP receptor activation (Fig. 5D). Incontrast, sections from explants maintained in control conditionsshowed almost no detectable labeling for pCREB within the outerportions of the neuroblastic layer (Fig. 5C). Importantly, we detectedpCREB immunoreactivity in cells within the neuroblastic layer thatwere also immunostained for cyclin D1 in PACAP-treated explants(Fig. 5F), which was almost undetectable in control retinas (Fig. 5E).
Cyclin D1 is a crucial regulator of cell cycle progression andproliferation of progenitor cells in the developing retina (Fantl et al.,1995; Sicinski et al., 1995; Ma et al., 1998), and acts in coordinationwith the cyclin-dependent kinase inhibitor (CKI) p27Kip1 to allowproper retinogenesis (Levine et al., 2000; Dyer & Cepko, 2001a; Tong& Pollard, 2001). To further test for molecular mechanisms involvedin the PACAP-induced antiproliferative effect, we analyzed whethertreatment with this neuropeptide changes the content of either cyclinD1 or p27kip1. mRNA levels for cyclin D1 as well as p27kip1 weremeasured by Q-RT-PCR after 1 and 6 h of treatment with PACAP.After 6 h of treatment, there was a decrease in cyclin D1 mRNAcontent (Fig. 6A, two-way anova; n = 6; P < 0.05), but no signif-icant alteration in p27kip1. mRNA content (Fig. 6B; two-way anova;n = 6; P > 0.05). When co-treatment with IBMX and PACAP was
used, the cyclin D1 mRNA level was decreased even more (Fig. 6C;one-way anova; PACAP vs. IBMX + PACAP; n = 4; P < 0.05).Moreover, treatment with H89 (a PKA inhibitor) blocked the effect ofPACAP on cyclin D1 mRNA content (Fig. 6D; one-way anova;PACAP vs. H89 + PACAP; n = 6; P < 0.05).We tested whether the effects on gene expression would translate to
protein content by performing western blots for cyclin D1 and p27kip1.Consistently, treatment of retinal explants for 8 h with PACAP38decreased cyclin D1 protein content when compared to controls(Fig. 7A and B) (Fig. 7B; two-way anova; n = 5; P < 0.05). Therewas no significant change in p27kip1 protein content after PACAPreceptor activation (Fig. 7C; two-way anova; n = 4; P > 0.05). Inaddition, we quantified the number of cyclin D1-positive cells after8 h of treatment. We showed that the number of cyclin D1-positivecells was reduced in response to treatment of retinal explants withPACAP (Fig. 7D–F) (Fig. 7F; t-test; n = 3; P = 0.027; d.f. = 4).
Discussion
In this study, we investigated the role of the neuroactive peptidePACAP in the proliferation of retinal progenitors. The data show thatboth PACAP and PACAP receptors are detected early in development,and that PAC1 is expressed in progenitor cells. PACAP reduced boththe incorporation of [3H]thymidine and the number of BrdU-positivecells. Both the PAC1-specific agonist maxadilan (Lerner et al., 1991;Moro & Lerner, 1997) and VIP had an antiproliferative effect similarto that of PACAP. The PAC1-specific antagonist M65 partiallyblocked the effect of PACAP, which reinforces the involvement ofboth PAC1 and VPACs. Activation of PACAP receptors inducedimmediate phosphorylation of CREB in a subpopulation of retinalprogenitors. We have also shown a decrease in cyclin D1 mRNA inresponse to PACAP, an effect that was enhanced by co-treatment withIBMX and blocked by H89. Finally, both cyclin D1 protein levels andthe number of cyclin D1-positive cells were reduced after PACAPtreatment.
A
C
B
D
Fig. 6. PACAP reduces the levels of cyclin D1 mRNA through the cAMP–PKA pathway. Real-time RT-PCR for cyclin D1 (A) and p27kip1 (B) from retinalexplants of P1 rats maintained in vitro for 1 or 6 h, in either the presence or the absence of 10 nm PACAP38. Treatment with 100 lm IBMX for 6 h increased theeffect of PACAP (C). Treatment with 1 lm H89 for 6 h blocked the effect of PACAP (D). Both IBMX and H89 were added 30 min before PACAP. The results arepresented as means ± SEM. *P < 0.05 in two-way anova followed by Bonferroni’s post hoc test (A). *P < 0.05 and ***P < 0.001 in one-way anova followed byTukey’s post hoc test (C and D).
PACAP is antiproliferative in the retina 7
ª The Authors (2010). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 1–10

The PACAP receptor PAC1 is expressed in postmitotic undifferen-tiated cells (Silveira et al., 2002) and in retinal progenitor cells in bothembryonic and postnatal rat retinas. Similarly, mRNAs for bothVPAC1 and VPAC2 were detected. In addition to the antiproliferativeeffects induced by PACAP treatment, we have also demonstrated thatVIP can regulate retinal cell proliferation. Together with the incom-plete reversal of PACAP effects by the antagonist M65, these resultssuggest that VPACs may also be involved in the effects of PACAP.Short-term activation of PACAP receptors (5–15 min) led to
phosphorylation of CREB in a subpopulation of retinal progenitorcells that express cyclin D1. Given the rapid activation of the CREBsignaling pathway by PACAP in retinal progenitors that are probablyin transition from G1 to S phase, it is likely that the antiproliferativeeffect of this neuropeptide results from direct activation of PACAPreceptors expressed by retinal progenitor cells; however, regulation ofcell proliferation through an indirect effect could not be ruled out. Infact, some postmitotic cells also present pCREB immunoreactivity inresponse to PACAP, and our group had previously demonstrated thatPACAP has a neuroprotective effect in this population (Silveira et al.,2002). Moreover, it is not known whether activation of the PACAP
receptors directly induces exit from the cell cycle or, alternatively,regulates the length of the cell cycle in retinal progenitors.Although preliminary data demonstrated that PAC1 isoforms that
induce IP3 production (Spengler et al., 1993) may be expressed in theretina (data not shown), our previous study showed that, in the retinasof newborn rats, PACAP receptor activation leads to cAMP, but notIP3, synthesis, and that the neuroprotective effect of PACAP dependson the cAMP–PKA pathway (Silveira et al., 2002). In this study, weshow that co-treatment of retinal tissue with PACAP and IBMX, adrug that prevents cAMP degradation, enhances the PACAP antipro-liferative effect and the induced decrease of cyclin D1 mRNAexpression, suggesting that the effect of PACAP upon retinalprogenitor cells is also mediated by the cAMP pathway. In addition,a PKA inhibitor (H89) blocked the reduction in cyclin D1 mRNAlevels induced by PACAP. The production of cAMP is modulated byvarious cell-extrinsic regulators, which suggests that this secondmessenger probably integrates multiple signaling pathways. Interest-ingly, cAMP signaling is well known to antagonize the Shh pathway(Epstein et al., 1996; Hammerschmidt et al., 1996; Waschek et al.,2006). Release of both Shh and PACAP by postmitotic cells has been
A
B C
D
E
F
Fig. 7. PACAP reduces the protein content of cyclin D1 and the number of cyclin D1-positive cells. For western blot analysis for cyclin D1, p27kip1 and a-tubulinretinal explants were treated for 3 h or 8 h. A representative western blot is shown in A. Densitometric analysis of cyclin D1 ⁄ a-tubulin from five independentexperiments (B) and p27kip1 ⁄ a-tubulin from four independent experiments (C). Results are presented as means ± SEM pooled from the indicated independentexperiments. *P < 0.05 in two-way anova followed by Bonferroni’s post hoc test (B). For the quantification of cyclin D1-positive cells, retinal explants weremechanically dissociated to single cells after treatment for 8 h with (E) or without (D) 10 nm PACAP, and the cells were analyzed by immunocytochemistry forcyclin D1. (F) Five hundred cells were counted in each experiment, and the number of cyclin D1-positive cells was determined. #P < 0.05 in t-test.
8 B. Njaine et al.
ª The Authors (2010). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 1–10

implicated in the regulation of proliferation in the cerebellum (Nicotet al., 2002), and recent studies have also shown that Shh is a potentmitogenic factor in the perinatal retina. It was proposed that Shhsecreted by ganglion cells controls the proliferation of progenitor cellsthrough modulation of the expression of cyclin D1 (Wang et al.,2005). Therefore, PACAP may regulate the retinal progenitor cellcycle in coordination with Shh. These findings indicate a potential areafor future research, as little is known about how retinal progenitor cellsintegrate signals from multiple cell-extrinsic factors.
The time course of expression and activity of certain proteins iscrucial to the control of cell cycle progression or cell cycle exit (Sherr& Roberts, 2004). In the developing retina, cyclin D1 is highlyexpressed by progenitor cells, and its misexpression severely impairsretinal development and normal photoreceptor formation (Fantl et al.,1995; Sicinski et al., 1995; Ma et al., 1998; Skapek et al., 2001). TheCKIs p19Ink4d, p27kip1 and p57kip2 are also known to control exit fromthe cell cycle in the retina (Levine et al., 2000; Dyer & Cepko, 2001b;Cunningham et al., 2002). Mice deficient for p27Kip1 exhibitedincreased cell proliferation throughout development, as well asextensive apoptosis, particularly during the later stages of retinalhistogenesis (Levine et al., 2000; Dyer & Cepko, 2001a). It has beenpreviously shown that the antiproliferative effect of PACAP incerebral cortical progenitors depends on the upregulation of p57kip2
and a decrease in cdk2-associated activity (Carey et al., 2002).However, we did not detect any change in p57kip2 following treatmentof retinal tissue with PACAP (data not shown). In mice, this CKI isexpressed in a small subset (�16%) of retinal progenitors (Dyer &Cepko, 2000). Besides indicating that distinct populations of retinalprogenitor cells may use different mechanisms to exit the cell cycleduring development (Dyer & Cepko, 2000, 2001a,b), the veryrestricted pattern of p57kip2 expression among retinal progenitor cellssuggests that more refined methods may be necessary to determinewhether p57kip2 expression is regulated by PACAP receptor activationin the developing retina. Altogether, the available data corroborate thatthe molecular mechanisms of cell cycle control triggered by PACAPmay differ in distinct areas of the developing CNS (Carey et al., 2002;Pagano & Jackson, 2004).
We showed that activation of PACAP receptors decreased the levelsof cyclin D1 mRNA and protein. Interestingly, when degradation ofcAMP was inhibited with IBMX, in combination with PACAPtreatment, the decrease in mRNA for cyclin D1 was enhanced. Thesefindings indicate that a cAMP–PKA signaling pathway may regulatecyclin D1 gene expression in the observed antiproliferative effect ofPACAP. Accordingly, inhibition of PKA blocked the reduction incyclin D1 levels induced by PACAP. This is the first cleardemonstration that the cAMP–PKA pathway controls cyclin D1expression in rodent retinal tissue. The link between the phosphor-ylation of CREB and the downregulation of cyclin D1 is, nonetheless,undetermined. Indeed, a disabling mutation of CREB leads toincreased hippocampal cell proliferation and neurogenesis (Gur et al.,2007), whereas activation of CREB has been shown to positivelyregulate transcription of the cyclin D1 gene in non-neuronal andneuroblastoma cells (Garcia et al., 2004; Garcıa-Silva & Aranda,2004; Sharma et al., 2004; Kim et al., 2006). It is possible that, inretinal progenitors, activation of CREB may regulate the expression ofcyclin D1 gene repressors, instead of directly regulating the cyclin D1promoter. This and other alternative molecular mechanisms of theregulation of cyclin D1 transcription by PACAP are now underinvestigation.
In conclusion, this study demonstrated that PACAP regulates theproliferation of a subpopulation of progenitor cells in the developingretinal tissue. Through the activation of PAC1 andVPACs and signaling
via the cAMP–PKA pathway, this peptide regulates both the phosphor-ylation of CREB and the expression of cyclin D1. The data suggest thatthe neuropeptide PACAP, which is present in the micro-environment ofretinal progenitor cells early in development, is a modulator of cellproliferation in the developing retinal tissue, andmay therefore affect thespecification and ⁄ or differentiation of retinal cell types.
Supporting Information
Additional supporting information may be found in the online versionof this article:Fig. S1. PAC1 is expressed in BrdU-positive retinal progenitors.Please note: As a service to our authors and readers, this journalprovides supporting information supplied by the authors. Suchmaterials are peer-reviewed and may be re-organized for onlinedelivery, but are not copy-edited or typeset by Wiley-Blackwell.Technical support issues arising from supporting information (otherthan missing files) should be addressed to the authors.
Acknowledgements
We thank Jose Nilson dos Santos, Jose Francisco Tiburcio, Gildo Brito deSouza, Talita Martins de Oliveira, Leonardo M. C. Pena, Maurıcio R. Martinsand Naiara C. Pimentel-Valim for help and technical support, Dr Victor May forthe PAC1 antibody and Dr Ethan Lerner for both maxadilan and M65. Thisinvestigation was supported by grants from PRONEX MCT ⁄ CNPq, FAPERJ,Fundacao Universitaria Jose Bonifacio and Prodoc-CAPES. M. F. Santiago issupported by grants from FAPERJ and Fundacao Universitaria Jose Bonifacio.
Abbreviations
BrdU, 5-bromo-2¢-deoxyuridine; CKI, cyclin-dependent kinase inhibitor; CNS,central nervous system; CREB, cAMP response element-binding protein; d.f.,degrees of freedom; DPM, disintegrations per minute; GCL, ganglion celllayer; HRP, horseradish peroxidase; IBMX, 3-isobutyl-1-methylxanthine; INL,inner cell layer; IP3, inositol triphosphate; PAC1, pituitary adenylyl cyclase-activating polypeptide-specific receptor; PACAP, pituitary adenylyl cyclase-activating polypeptide; PCNA, proliferating cell nuclear antigen; pCREB,phosphorylated cAMP response element-binding protein; PKA, cAMP-depen-dent protein kinase (protein kinase A); Q-RT-PCR, quantitative RT-PCR; SEM,standard error of the mean; Shh, Sonic hedgehog; TUNEL, terminaldeoxynucleotidyl transferase-mediated dUTP nick end-labeling; VIP, vasoac-tive intestinal peptide; VPAC, receptor activated by both pituitary adenylylcyclase-activating polypeptide and vasoactive intestinal peptide.
References
Anchan, R.M. & Reh, T.A. (1995) Transforming growth factor-beta-3 ismitogenic for rat retinal progenitor cells in vitro. J. Neurobiol., 28, 133–145.
Anchan, R.M., Reh, T.A., Angello, J., Balliet, A. & Walker, M. (1991) EGFand TGF-alpha stimulate retinal neuroepithelial cell proliferation in vitro.Neuron, 6, 923–936.
Arimura, A., Somogyvari-Vigh, A., Miyata, A., Mizuno, K., Coy, D.H. &Kitada, C. (1991) Tissue distribution of PACAP as determined by RIA:highly abundant in the rat brain and testes. Endocrinology, 129, 2787–2789.
Borba, J.C., Henze, I.P., Silveira, M.S., Kubrusly, R.C., Gardino, P.F., de Mello,M.C., Hokoc, J.N. & de Mello, F.G. (2005) Pituitary adenylate cyclase-activating polypeptide (PACAP) can act as determinant of the tyrosinehydroxylase phenotype of dopaminergic cells during retina development.Brain Res. Dev. Brain Res., 156, 193–201.
Braas, K.M. & May, V. (1999) Pituitary adenylate cyclase-activating polypep-tides directly stimulate sympathetic neuron neuropeptide Y release throughPac1 receptor isoform activation of specific intracellular signaling pathways.J. Biol. Chem., 274, 27702–27710.
Braas, K.M., May, V., Harakall, S.A., Hardwick, J.C. & Parsons, R.L. (1998)Pituitary adenylate cyclase-activating polypeptide expression and modula-tion of neuronal excitability in guinea pig cardiac ganglia. J. Neurosci., 18,9766–9779.
PACAP is antiproliferative in the retina 9
ª The Authors (2010). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 1–10

Burmeister, M., Novak, J., Liang, M.Y., Basu, S., Ploder, L., Hawes, N.L.,Vidgen, D., Hoover, F., Goldman, D., Kalnins, V.I., Roderick, T.H., Taylor,B.A., Hankin, M.H. & McInnes, R.R. (1996) Ocular retardation mousecaused by Chx10 homeobox null allele: impaired retinal progenitorproliferation and bipolar cell differentiation. Nat. Genet., 12, 376–384.
Carey, R.G., Li, B. & DiCicco-Bloom, E. (2002) Pituitary adenylate cyclaseactivating polypeptide anti-mitogenic signaling in cerebral cortical progen-itors is regulated by p57kip2-dependent Cdk2 activity. J. Neurosci., 22,1583–1591.
Close, J.L., Gumuscu, B. & Reh, T.A. (2005) Retinal neurons regulateproliferation of postnatal progenitors and Muller glia in the rat retina via TGFbeta signaling. Development, 132, 3015–3026.
Cunningham, J.J., Levine, E.M., Zindy, F., Goloubeva, O., Roussel, M.F. &Smeyne, R.J. (2002) The cyclin-dependent kinase inhibitors p19(Ink4d) andp27(Kip1) are coexpressed in select retinal cells and act cooperatively tocontrol cell cycle exit. Mol. Cell. Neurosci., 19, 359–374.
DiCicco-Bloom, E., Deutsch, P.J., Maltzman, J., Zhang, J., Pintar, J.E., Zheng,J., Friedman, W.F., Zhou, X. & Zaremba, T. (2000) Autocrine expression andontogenetic functions of the PACAP ligand ⁄ receptor system duringsympathetic development. Dev. Biol., 219, 197–213.
Dickson, L. & Finlayson, K. (2009) VPAC and PAC receptors: from ligands tofunction. Pharmacol. Ther., 121, 294–316.
Donovan, S.L. & Dyer, M.A. (2005) Regulation of proliferation during centralnervous system development. Semin. Cell Dev. Biol., 16, 407–421.
Dyer, M.A. & Cepko, C.L. (2000) p57 regulates progenitor cell proliferationand amacrine interneuron development in the mouse retina. Development,127, 3593–3605.
Dyer, M.A. & Cepko, C.L. (2001a) P27kip1 and P57kip2 regulate proliferationin distinct retinal progenitor cell populations. J. Neurosci., 21, 4259–4271.
Dyer, M.A. & Cepko, C.L. (2001b) The p57Kip2 cyclin kinase inhibitor isexpressed by a restricted set of amacrine cells in the rodent retina. J. Comp.Neurol., 429, 601–614.
Epstein, D.J., Marti, E., Scott, M.P. & McMahon, A.P. (1996) AntagonizingcAMP-dependent protein kinase A in the dorsal CNS activates a conservedSonic hedgehog signaling pathway. Development, 122, 2885–2894.
Fahrenkrug, J. & Hannibal, J. (2004) Neurotransmitters co-existing with VIP orPACAP. Peptides, 25, 393–401.
Falluel-Morel, A., Vaudry, D., Aubert, N., Galas, L., Benard, M., Basille, M.,Fontaine, M., Fournier, A., Vaudry, H. & Gonzalez, B.J. (2005) Pituitaryadenylate cyclase-activating polypeptide prevents the effects of ceramides onmigration, neurite outgrowth, and cytoskeleton remodeling. Proc. Natl Acad.Sci. USA, 102, 2637–2642.
Fantl, V., Stamp, G., Andrews, A., Rosewell, I. & Dickson, C. (1995) Micelacking cyclin D1 are small and show defects in eye and mammary glanddevelopment. Genes Dev., 9, 2364–2372.
Garcia, C., Paez, P., Davio, C., Soto, E.F. & Pasquini, J.M. (2004)Apotransferrin induces cAMP ⁄ CREB pathway and cell cycle exit inimmature oligodendroglial cells. J. Neurosci. Res., 78, 338–346.
Garcıa-Silva, S. & Aranda, A. (2004) The thyroid hormone receptor is asuppressor of ras-mediated transcription, proliferation, and transformation.Mol. Cell. Biol., 24, 7514–7523.
Gower, W.R. Jr, Dietz, J.R., McCuen, R.W., Fabri, P.J., Lerner, E.A. &Schubert, M.L. (2003) Regulation of atrial natriuretic peptide secretion bycholinergic and PACAP neurons of the gastric antrum. Am. J. Physiol.Gastrointest. Liver Physiol., 284, 68–74.
Gur, T.L., Conti, A.C., Holden, J., Bechtholt, A.J., Hill, T.E., Lucki, I.,Malberg, J.E. & Blendy, J.A. (2007) cAMP response element-bindingprotein deficiency allows for increased neurogenesis and a rapid onset ofantidepressant response. J. Neurosci., 27, 7860–7868.
Hammerschmidt, M., Bitgood, M.J. & McMahon, A.P. (1996) Protein kinase Ais a common negative regulator of Hedgehog signaling in the vertebrateembryo. Genes Dev., 10, 647–658.
Jensen, A. & Wallace, V. (1997) Expression of Sonic Hedgehog and its putativerole as a precursor cell mitogen in the developing mouse retina. Develop-ment, 124, 363–371.
Kim, M.J., Kang, J.H., Park, Y.G., Ryu, G.R., Ko, S.H., Jeong, I.K., Koh, K.H.,Rhie, D.J., Yoon, S.H., Hahn, S.J., Kim, M.S. & Jo, Y.H. (2006) Exendin-4induction of cyclin D1 expression in INS-1 beta-cells: involvement ofcAMP-responsive element. J. Endocrinol., 188, 623–633.
Lerner, E.A., Ribeiro, J.M., Nelson, R.J. & Lerner, M.R. (1991) Isolation ofmaxadilan, a potent vasodilatory peptide from the salivary glands of the sandfly Lutzomyia longipalpis. J. Biol. Chem., 266, 11234–11236.
Levine, E.M., Close, J., Fero, M., Ostrovsky, A. & Reh, T.A. (2000) p27(Kip1)regulates cell cycle withdrawal of late multipotent progenitor cells in themammalian retina. Dev. Biol., 219, 299–314.
Lillien, L. & Cepko, C. (1992) Control of proliferation in the retina: temporalchanges in responsiveness to FGF and TGF-alpha. Development, 115, 253–266.
Lowry, O.H., Rosebrough, N.J., Farr, A.L. & Randall, R.J. (1951) Proteinmeasurement with the folin phenol reagent. J. Biol. Chem., 193, 265–275.
Lu, N. & DiCicco-Bloom, E. (1997) Pituitary adenylate cyclase-activatingpolypeptide is an autocrine inhibitor of mitosis in cultured cortical precursorcells. Proc. Natl Acad. Sci. USA, 94, 3357–3362.
Lu, N., Zhou, R. & DiCicco-Bloom, E. (1998) Opposing mitogenic regulationby PACAP in sympathetic and cerebral cortical precursors correlates withdifferential expression of PACAP receptor (PAC1-R) isoforms. J. Neurosci.Res., 15, 651–662.
Ma, C., Papermaster, D. & Cepko, C.L. (1998) A unique pattern ofphotoreceptor degeneration in cyclin D1 mutant mice. Proc. Natl Acad.Sci. USA, 95, 9938–9943.
Martins, R.A.P. & Pearson, R.A. (2007) Control of cell proliferation byneurotransmitters in the developing vertebrate retina. Brain Res., 4, 37–60.
Martins, R.A.P., Linden, R. & Dyer, M.A. (2006) Glutamate regulates retinalprogenitor cell proliferation during development. Eur. J. Neurosci., 24,24969–24980.
Martins, R.A.P., Zindy, F., Donovan, S., Eisenman, R., Roussel, M. & Dyer, M.(2008) N-myc coordinates retinal growth with eye size during mousedevelopment. Genes Dev., 22, 179–193.
Mayr, B. & Montminy, M. (2001) Transcriptional regulation by the phosphor-ylation-dependent factor CREB. Nat. Rev. Mol. Cell Biol., 2, 599–609.
Moro, O. & Lerner, E.A. (1997) Maxadilan, the vasodilator from sand flies, is aspecific pituitary adenylate cyclase peptide type I receptor agonist. J. Biol.Chem., 272, 966–970.
Moro, O., Wakita, K., Ohnuma, M., Denda, S., Lerner, E. & Tajima, M.(1999) Characterization of structural alterations in the sequence of thevasodilatory peptide maxadilan yields a pituitary adenylate cyclase-activating peptide type 1 receptor-specific antagonist. J. Biol. Chem.,274, 23103–23110.
Nicot, A. & DiCicco-Bloom, E. (2001) Regulation of neuroblast mitosis isdetermined by PACAP receptor isoform expression. Proc. Natl Acad. Sci.USA, 98, 4758–4763.
Nicot, A., Lelievre, V., Tam, J., Waschek, J.A. & DiCicco-Bloom, E. (2002)Pituitary adenylate cyclase-activating polypeptide and sonic hedgehoginteract to control cerebellar granule precursor cell proliferation. Eur. J.Neurosci., 22, 9244–9254.
Pagano, M. & Jackson, P.K. (2004) Wagging the dogma: tissue specific cellcycle control in the mouse embryo. Cell, 118, 535–538.
Rehen, S.K., Varella, M.H., Freitas, F.G., Moraes, M.O. & Linden, R. (1996)Contrasting effects of protein synthesis inhibition and of cyclic AMP onapoptosis in the developing retina. Development, 122, 1439–1448.
Said, S.I. & Mutt, V. (1970) Polypeptide with broad biological activity:isolation from small intestine. Science, 169, 1217–1218.
Sharma, C., Pradeep, A., Pestell, R.G. & Rana, B. (2004) Peroxisomeproliferator-activated receptor gamma activation modulates cyclin D1transcription via beta-catenin-independent and cAMP-response element-binding protein-dependent pathways in mouse hepatocytes. J. Biol. Chem.,279, 16927–16938.
Sherr, C.J. & Roberts, J.M. (2004) Living with or without cyclins and cyclin-dependent kinases. Genes Dev., 18, 2699–2711.
Sicinski, P., Donaher, J.L., Parker, S.B., Li, T., Fazeli, A., Gardner, H., Haslam,S.Z., Bronson, R.T., Elledge, S.J. & Weinberg, R.A. (1995) Cyclin D1provides a link between development and oncogenesis in the retina andbreast. Cell, 82, 621–630.
Silveira, M.S., Costa, M.R., Bozza, M. & Linden, R. (2002) Pituitary adenylylcyclase-activating polypeptide prevents induced cell death in retinal tissuethrough activation of cyclic AMP-dependent protein kinase. J. Biol. Chem.,277, 16075–16080.
Skapek, S.X., Lin, S.C., Jablonski,M.M.,Mckeller, R.N., Tan,M., Hu, N.&Lee,E.Y. (2001) Persistent expression of cyclin D1 disrupts normal photoreceptordifferentiation and retina development. Oncogene, 20, 6742–6751.
Spengler, D., Waeber, C., Pantaloni, C., Holsboer, F., Bockaert, J., Seeburg,P.H. & Journot, L. (1993) Differential signal transduction by 5¢ splicevariants of the PACAP receptor. Nature, 365, 170–175.
Suh, J., Lu, N., Nicot, A., Tatsuno, I. & DiCicco-Bloom, E. (2001) PACAP isan anti-mitogenic signal in developing cerebral cortex. Nat. Neurosci., 4,123–124.
Taylor, M. & Reh, T.A. (1990) Induction of differentiation of rat retinal,germinal, neuroepithelial cells by dbcAMP. J. Neurobiol., 21, 470–481.
Tong, W. & Pollard, J.W. (2001) Genetic evidence for the interactions of cyclinD1 and p27(Kip1) in mice. Mol. Cell. Biol., 21, 1319–1328.
10 B. Njaine et al.
ª The Authors (2010). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 1–10

Turner, D.L. & Cepko, C.L. (1987) A common progenitor for neurons and gliapersists in rat retina late in development. Nature, 328, 131–136.
Varella, M.H., Correa, D.F., Campos, C.B.L., Chiarini, L.B. & Linden, R.(1997) Protein kinases selectively modulate apoptosis in the developingretina in vitro. Neurochem. Int., 31, 217–227.
Varella, M.H., de Mello, F.G. & Linden, R. (1999) Evidence for an antiapoptoticrole of dopamine in developing retinal tissue. J. Neurochem., 73, 485–492.
Vaudry, D., Gonzalez, B.J., Basille, M., Yon, L., Fournier, A. & Vaudry, H.(2000) Pituitary adenylate cyclase-activating polypeptide and its receptors:from structure to function. Pharmacol. Rev., 52, 269–324.
Vaudry, D., Falluel-Morel, A., Bourgault, S., Basille, M., Burel, D., Wurtz, O.,Fournier, A., Chow, B.K.C., Hashimoto, H., Galas, L. & Vaudry, H. (2009)Pituitary adenylate cyclase-activating polypeptide and its receptors: 20 yearsafter the discovery. Pharmacol. Rev., 61, 283–357.
Wang, Y., Dakubo, G.D., Thurig, S., Mazerolle, C.J. & Wallace, V.A. (2005)Retinal ganglion cell-derived sonic hedgehog locally controls proliferation
and the timing of RGC development in the embryonic mouse retina.Development, 132, 5103–5113.
Waschek, J.A., Casillas, R.A., Nguyen, T.B., DiCicco-Bloom, E.M.,Carpenter, E.M. & Rodriguez, W.I. (1998) Neural tube expression ofpituitary adenylate cyclase-activating peptide (PACAP) and receptor:potential role in patterning and neurogenesis. Proc. Natl Acad. Sci. USA,95, 9602–9607.
Waschek, J.A., Dicicco-Bloom, E., Nicot, A. & Lelievre, V. (2006) Hedgehogsignaling: new targets for GPCRs coupled to cAMP and protein kinase A.Ann. N Y Acad. Sci., 1070, 120–128.
Young, T.L. & Cepko, C.L. (2004) A role for ligand-gated ion channels in rodphotoreceptor development. Neuron, 41, 867–879.
Zhou, C.J., Shioda, S., Yada, T., Inagaki, N., Pleasure, S.J. & Kikuyama, S.(2002) PACAP and its receptors exert pleiotropic effects in the nervoussystem by activating multiple signaling pathways. Curr. Protein Pept. Sci., 3,423–439.
PACAP is antiproliferative in the retina 11
ª The Authors (2010). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 1–10
Related Documents











