1 3 Plant Mol Biol DOI 10.1007/s11103-014-0209-6 Pisum sativum p68 DEAD‑box protein is ATP‑dependent RNA helicase and unique bipolar DNA helicase Narendra Tuteja · Mohammed Tarique · Mst. Sufara Akhter Banu · Moaz Ahmad · Renu Tuteja Received: 3 April 2014 / Accepted: 29 May 2014 © Springer Science+Business Media Dordrecht 2014 Keywords Bipolar unwinding · DEAD-box proteins · Dual helicase · p68 · Pisum sativum · RNA and DNA binding Introduction Nucleic acids metabolism is crucial for large num- ber of genetic processes which require various essential enzymes especially ATPases and helicases. Helicases are ubiquitous enzymes required for unwinding of energeti- cally stable duplex DNA (DNA helicases) or duplex RNA secondary structures (RNA helicases) (Tuteja and Tuteja 2004a, b; Tuteja and Pradhan 2006). These helicases play essential roles in DNA replication, repair, recombina- tion, transcription, translation and in RNA metabolism such as RNA synthesis, splicing, editing, rRNA process- ing, regulation of RNA stability, ribosome assembly and translation initiation and therefore are involved in basic cellular processes regulating plant growth and develop- ment (Tuteja 2003; Gong et al. 2005; Cordin et al. 2006; Owttrim 2006; Guan et al. 2013; Kammel et al. 2013). It seems therefore that helicases might be playing an important role in stabilizing growth in plants under stress by regulating stress-induced transcription and transla- tion. There are reports on the presence of helicases in the stress-induced open reading frames identified by cDNA microarray. These helicases act as an essential molecu- lar tool for cellular machinery and possess a set of highly conserved amino acid sequences called ‘helicase motifs’, which are clustered together for helicase function (Tuteja and Tuteja 2004a, b). Because of the presence of a DEAD (Asp-Glu-Ala-Asp) motif, the family of these proteins are also known as DEAD-box family of proteins (Tuteja and Tuteja 2004b; Vashisht and Tuteja 2006). Abstract DEAD-box helicases play essential role in DNA and RNA metabolism such as replication, repair, recombination, transcription, translation, ribosome biogen- esis and splicing which regulate plant growth and develop- ment. The presence of helicases in the stress-induced ORFs identified by cDNA microarray indicates that helicases might be playing an important role in stabilizing growth in plants under stress. p68 DEAD-box helicase has been iden- tified and characterized from animal systems but the prop- erties and functions of plant p68 are poorly understood. In this study, the identification, purification and characteriza- tion of recombinant p68 from Pisum sativum (Psp68) is presented. Psp68 possesses all the characteristic motifs like DEAD-box ATP-binding and helicase C terminal motifs and is structurally similar to human p68 homologue. Psp68 exhibits ATPase activity in the presence of both DNA and RNA and it binds to DNA as well as RNA. It contains the characteristic RNA helicase activity. Interestingly Psp68 also shows the unique DNA helicase activity, which is bipolar in nature (unwinds DNA in both the 5′–3′ and 3′–5′ directions). The Km values of Psp68 for ATPase are 0.5126 and 0.9142 mM in the presence of DNA and RNA, respec- tively. The Km values of Psp68 are 1.6129 and 1.14 nM for DNA helicase and RNA helicase, respectively. The unique properties of Psp68 suggest that it could be a multi- functional protein involved in different aspect of DNA and RNA metabolism. This discovery should make an impor- tant contribution to better understanding of nucleic acids metabolism plants. N. Tuteja (*) · M. Tarique · M. S. A. Banu · M. Ahmad · R. Tuteja International Centre for Genetic Engineering and Biotechnology, Aruna Asaf Ali Marg, New Delhi 110067, India e-mail: [email protected]

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1 3
Plant Mol BiolDOI 10.1007/s11103-014-0209-6
Pisum sativum p68 DEAD‑box protein is ATP‑dependent RNA helicase and unique bipolar DNA helicase
Narendra Tuteja · Mohammed Tarique · Mst. Sufara Akhter Banu · Moaz Ahmad · Renu Tuteja
Received: 3 April 2014 / Accepted: 29 May 2014 © Springer Science+Business Media Dordrecht 2014
Keywords Bipolar unwinding · DEAD-box proteins · Dual helicase · p68 · Pisum sativum · RNA and DNA binding
Introduction
Nucleic acids metabolism is crucial for large num-ber of genetic processes which require various essential enzymes especially ATPases and helicases. Helicases are ubiquitous enzymes required for unwinding of energeti-cally stable duplex DNA (DNA helicases) or duplex RNA secondary structures (RNA helicases) (Tuteja and Tuteja 2004a, b; Tuteja and Pradhan 2006). These helicases play essential roles in DNA replication, repair, recombina-tion, transcription, translation and in RNA metabolism such as RNA synthesis, splicing, editing, rRNA process-ing, regulation of RNA stability, ribosome assembly and translation initiation and therefore are involved in basic cellular processes regulating plant growth and develop-ment (Tuteja 2003; Gong et al. 2005; Cordin et al. 2006; Owttrim 2006; Guan et al. 2013; Kammel et al. 2013). It seems therefore that helicases might be playing an important role in stabilizing growth in plants under stress by regulating stress-induced transcription and transla-tion. There are reports on the presence of helicases in the stress-induced open reading frames identified by cDNA microarray. These helicases act as an essential molecu-lar tool for cellular machinery and possess a set of highly conserved amino acid sequences called ‘helicase motifs’, which are clustered together for helicase function (Tuteja and Tuteja 2004a, b). Because of the presence of a DEAD (Asp-Glu-Ala-Asp) motif, the family of these proteins are also known as DEAD-box family of proteins (Tuteja and Tuteja 2004b; Vashisht and Tuteja 2006).
Abstract DEAD-box helicases play essential role in DNA and RNA metabolism such as replication, repair, recombination, transcription, translation, ribosome biogen-esis and splicing which regulate plant growth and develop-ment. The presence of helicases in the stress-induced ORFs identified by cDNA microarray indicates that helicases might be playing an important role in stabilizing growth in plants under stress. p68 DEAD-box helicase has been iden-tified and characterized from animal systems but the prop-erties and functions of plant p68 are poorly understood. In this study, the identification, purification and characteriza-tion of recombinant p68 from Pisum sativum (Psp68) is presented. Psp68 possesses all the characteristic motifs like DEAD-box ATP-binding and helicase C terminal motifs and is structurally similar to human p68 homologue. Psp68 exhibits ATPase activity in the presence of both DNA and RNA and it binds to DNA as well as RNA. It contains the characteristic RNA helicase activity. Interestingly Psp68 also shows the unique DNA helicase activity, which is bipolar in nature (unwinds DNA in both the 5′–3′ and 3′–5′ directions). The Km values of Psp68 for ATPase are 0.5126 and 0.9142 mM in the presence of DNA and RNA, respec-tively. The Km values of Psp68 are 1.6129 and 1.14 nM for DNA helicase and RNA helicase, respectively. The unique properties of Psp68 suggest that it could be a multi-functional protein involved in different aspect of DNA and RNA metabolism. This discovery should make an impor-tant contribution to better understanding of nucleic acids metabolism plants.
N. Tuteja (*) · M. Tarique · M. S. A. Banu · M. Ahmad · R. Tuteja International Centre for Genetic Engineering and Biotechnology, Aruna Asaf Ali Marg, New Delhi 110067, Indiae-mail: [email protected]

Plant Mol Biol
1 3
The DEAD-box RNA helicases is the largest family of RNA helicases with conserved sequence D-E-A-D in motif II (Linder and Jankowsky 2011). In spite of the sequence resemblance of DEAD-box RNA helicases within the core helicase regions, each DEAD-box helicase is believed to play crucial roles in plant growth and development (Linder and Jankowsky 2011). p68 a member of the ‘DEAD-box’ protein family was identified because of its immunological relations to the SV40 large tumor antigen and it has well conserved orthologues from yeast to human. Initially p68 protein was identified as nuclear protein but further stud-ies revealed that Human p68 protein shuttles between the nucleus and the cytoplasm (Iggo and Lane 1989; Wang et al. 2009). Nucleocytoplasmic shuttling of p68 protein is medi-ated by two nuclear exporting signal (NES) and two nuclear localization signal (NLS) sequence via a classical RanGT-Pase dependent pathway (Wang et al. 2009) during the cell cycle. Earlier, the expression of p68 was shown to be growth and developmentally regulated and to correlate with organ maturation/differentiation (Stevenson et al. 1998). It was also reported that ATPase activity of recombinant p68 was stimulated by double-stranded RNA (Huang and Liu 2002). Recombinant p68 RNA helicase unwinds dsRNA in both 3′–5′ and 5′–3′ directions (Huang and Liu 2002). The characterization of p68 homologue from human malaria parasite Plasmodium falciparum revealed that it is a DNA helicase also, which unwinds duplex DNA in both 3′–5′ and 5′–3′ directions (Pradhan et al. 2005a, b). Previous studies have shown that p68 RNA helicase is phosphorylated on tyrosine, serine, and threonine residues and its helicase and ATPase activities are stimulated after phosphorylation with protein kinase C (Pradhan et al. 2005b). These observa-tions suggest that p68 regulates diverse signal transduction pathways in response to a variety of external stimuli (Yang et al. 2005). It is well established that DEAD-box proteins usually act as components of large multi-protein complexes and their interactions with other factors, via their variable N- and C-terminal extensions, in the complexes most likely is responsible for the many different functions attributed to these proteins (Fuller-Pace 2013).
In animal system p68 protein interacts with and acts as a transcriptional coactivator for the nuclear receptor estro-gen receptor alpha (ERα) (Endoh et al. 1999). It was also reported that p68 interacts with the transcriptional coacti-vators CBP/p300, HDAC1 and RNA Pol II, stimulating transcriptional activation mediated by CBP/p300 (Rossow and Janknecht 2003; Bates et al. 2005). The studies with Drosophila homologue, Dmp68, suggested a novel role of p68 in transcript clearance from transcription sites and in transcriptional deactivation (Buszczak and Spradling 2006). The p68 and its yeast homologue Dbp2 are known to be important for rRNA processing and nonsense medi-ated mRNA decay (Fuller-Pace 2006). Wang et al. (2013)
reported that p68 also interacts with Ca2+–CaM. In plant it is well established that Ca2+–CaM regulates diverse signalling pathways leading to stress tolerance (Yang and Poovaiah 2003). Overall, p68 is a multifunctional protein involved in RNA splicing, pre-rRNA processing, RNA-induced silencing, transcription initiation and transcrip-tional repression etc. However, very little is known about p68 protein in plant system. In a previous study it was reported that the AtDRH1 (Arabidopsis thaliana DEAD-box RNA helicase 1), the homologue of human nuclear protein p68 is ATP/dATP-dependent RNA helicase and pol-ynucleotide-dependent ATPase (Okanami et al. 1998).
In this study recombinant p68 protein from Pisum sati-vum (Psp68) was purified and detailed biochemical studies were performed. The structural modeling revealed its simi-larity with the human p68 template. ATPase assay in the presence of either DNA or RNA was performed and Psp68 exhibits ATP hydrolyzing properties. The Psp68 contains the DNA and RNA helicase and DNA and RNA binding activi-ties. The detailed Psp68 ATPase kinetics assays revealed that Km value in the presence of DNA is 0.5126 mM and in the presence of RNA is 0.9142 mM. Psp68 helicase kinet-ics was also performed and the Km values are 1.6129 and 1.14 nM for DNA helicase and RNA helicase, respectively. This study will be helpful in understanding the biochemical properties of Psp68 protein which in turn will be useful in developing new varieties of plants, which can grow at sub-optimal conditions to full-fill the need of humans.
Materials and methods
Molecular cloning, sequence analysis and structural modeling
The pea p68 cDNA was cloned by screening of pea cDNA library using 1.9 kb cDNA fragment of p68 DEAH-box protein from A. thaliana (kindly provided by Tetsuo Meshi of Kyoto University, Japan) as a probe. The complete ORF of Psp68 (1.8 kb) was cloned into pGEMT easy vector and sequenced (Accession number: AF271892.1). The com-plete sequence of Ps-P68 was submitted to the Swissmodel homology-modeling server (http://swissmodel.expasy.org/) (Arnold et al. 2006) for the purpose of molecular modeling. The structural modeling of Psp68 was therefore done using the known crystal structure of the human DEAD-box heli-case DDX3X as the template (Hogbom et al. 2007).
Overexpression and purification of recombinant pea p68 protein
Pea p68 gene was sub-cloned in pET14b using NdeI and BamHI restriction sites of the expression vector. Transformed

Plant Mol Biol
1 3
cells containing recombinant plasmids were identified by colony PCR and plasmid of colony PCR positive clone was further confirmed by restriction digestion analysis and finally confirmed by the DNA sequencing. The E. coli strain BL21 [DE3, (codon +)], containing the recombinant pea p68-pET14b plasmid, was grown overnight in 100 ml LB medium at 37 °C and used as primary culture. Secondary culture was grown in LB medium at 37 °C until the OD600 reached at 0.6–0.7. The induction was carried out with 1 mM IPTG and the culture was kept again at 18 °C for overnight. The cells were harvested by centrifugation (8,000 rpm, 30 min at 4 °C) and the cell pellet was used for the recom-binant Psp68 protein purification. The cell pellet was washed twice with native buffer [50 mM Tris–Hcl (pH 8.0), 100 mM NaCl and 10 % Glycerol)] and finally re-suspended in soni-cation buffer containing 50 mM Tris–HCl (pH 8.0), 100 mM NaCl, 1 mM each of lysozyme, PMSF and benzamidine. The cells were lysed by sonication and soluble fraction was used for the purification of the recombinant protein with Q-Sepha-rose resins (pre-equilibrated with native buffer). The column was washed with 15–20 volumes of wash buffer containing 50 mM Tris–HCl (pH 8.0), 100 mM NaCl and 10 % glyc-erol. The recombinant Psp68 protein was eluted with linear gradient of 200–800 mM NaCl in a buffer containing 50 mM Tris–HCl (pH 8.0) and protease inhibitors cocktail (1X).
This purified protein was further purified by using Ni2+–NTA-agarose chromatography. Protein was again incu-bated with Ni2+–NTA resins (pre-equilibrated with wash buffer) for 45 min at 4 °C in rotating shaker in the pres-ence of 10 mM of imidazole to avoid the binding of con-taminant proteins with less than six histidine residues and loaded onto a column. The column was washed with 15–20 volumes of wash buffer containing 50 mM Tris–HCl (pH 8.0), 100 mM NaCl and 10 % glycerol and 50 mM of imi-dazole. The wash fraction was also collected. Psp68 protein was eluted with linear gradient of 50–500 mM imidazole in a buffer containing 50 mM Tris–HCl (pH 8.0), 200 mM NaCl and protease inhibitors mix. The purified protein was further dialyzed against storage buffer (50 mM Tris–HCl pH 8.0, 100 mM NaCl) to avoid the excess salts. The puri-fied recombinant protein was analyzed by using standard protocol of SDS-PAGE, silver staining and western blotting as described in our previous reports (Ahmad et al. 2012; Ahmad and Tuteja 2013). This purified protein was used for further biochemical characterization.
DNA/RNA-dependent ATP hydrolysis assay (ATPase assay)
Standard protocol was adopted from previous report (Tarique et al. 2013). The hydrolysis of ATP catalyzed by pure Psp68 protein was assayed by measuring the formation of 32P from [γ-32P]ATP. The reaction (10 µl) was performed
in the buffer (20 mM Tris–HCl, pH 8.0, 8 mM DTT, 1.0 mM MgCl2, 20 mM KCl and 16 µg/ml BSA) for 1 h at 37 °C in the presence of purified Psp68 and 10 ng of RNA or M13 mp19ssDNA and a mixture of [γ-32P] ATP (~17 nM) and 1 mM cold ATP. The products were separated by thin layer chromatography (TLC) (buffer composition: 0.5 M LiCl2 and 1 M formic acid) and the plate was exposed to hyper film for autoradiography and scanned on phosphoimager. For the concentration curve analysis different concentra-tions of Psp68 protein were used. The time course analysis was performed with a fixed concentration of Psp68 protein and time duration ranging from 10 to 90 min. The quan-titation assay was done using IMAGE j/geldoc software (http://rsbweb.nih.gov/ij/) and percentage of ATP hydrolysis was plotted as the bar diagram using Microsoft excel 2010.
Preparation of DNA helicase substrate
Standard published protocol was used to prepare these helicase substrates (Ahmad et al. 2012; Ahmad and Tuteja 2013). The helicase assays were performed using a partially duplex substrate consisting of 32P-labelled 47-mer DNA oli-godeoxynucleotide annealed with M13mp19 phage ssDNA. At both the 5′ and 3′ ends, this oligodeoxynucleotide con-tained 15 base-pairs of non-complementary region. 10 ng of the oligodeoxynucleotide was labelled at 5′-end with T4 pol-ynucleotide kinase (PNK) (5U) (New England Biolabs) and 1.85 MBq of [γ-32P]ATP (specific activity 222 TBq/mmol) using the standard PNK buffer (New England Biolabs) at 37 °C for 1 h. The labeled oligodeoxynucleotide was then annealed with 100 ng of single-stranded circular M13mp19 (+) phage DNA by using standard annealing buffer (20 mM Tris–HCl, pH 7.5, 10 mM MgCl2,100 mM NaCl, 1 mM DTT) followed by heating at 95 °C for 1 min. It was then immediately transferred to 65 °C for 2 min and then cooled down slowly at room temperature. The non-hybridized oli-godeoxynucleotide was removed using gel filtration through a Sepharose 4B column (Pharmacia, Sweden).
Preparation of blunt end DNA helicase substrate
17 mer oligodeoxynucleotide with the sequence (5′-GTT TTCCCAGTCACGAC-3′) was used to prepare the blunt end duplex substrate. This was labeled at the 5′ end using the method described above and was annealed to its complemen-tary oligodeoxynucleotide with the sequence 5′-GTCGT-GACTGGGAAAAC-3′. The substrate was purified by using Sepharose 4B column (Pharmacia, Sweden). Similarly radi-olabeed 17 mer oligodeoxynucleotide was annealed with circular M13mp19 phage ssDNA to prepare another helicase substrate. Radiobelled 32 mer oligodeoxynucleotide was also used to anneal with circular M13mp19 phage ssDNA to prepare another DNA duplex helicase substrate.

Plant Mol Biol
1 3
Preparation of direction-specific DNA substrate
To construct the direction-specific substrate (5′–3′) a 32-mer oligodeoxynucleotide (5′-TTCGAGCTCGGTACC CGGGGATCCTCTAGAGT-3′) was first annealed with M13mp19 ssDNA using annealing buffer (20 mM Tris–HCl, pH 7.5, 10 mM MgCl2, 100 mM NaCl, 1 mM DTT) and then labeled at 3′-OH end in appropriate buffer with 50 µCurie [α-32P]dCTP and 5 units of DNA polymerase I (large fragment) at 23 °C for 20 min. The incubation was continued for further 20 min at 23 °C after increasing the dCTP to 50 mM using unlabelled dCTP. This resulting duplex substrate was digested with SmaI and purified by gel filtration through 1 ml Sepharose 4B. The substrate consisting of long linear M13mp19 ssDNA with short duplex ends for 3′–5′ unwinding was prepared by first 5′-end labelling of 32-mer oligodeoxynucleotide and then annealing with M13mp19 ssDNA as described above. The annealed substrate was digested with SmaI and purified by gel filtration through 1 ml of Sepharose 4B.
DNA unwinding assay
The helicase assay measures the unwinding of a 32P-labeled DNA fragment from a partially duplex DNA molecule, catalyzed by Psp68 protein. The 10 µl reaction mixture containing buffer (20 mM Tris–HCl, pH 8.0, 8 mM DTT, 1.0 mM MgCl2, 20 mM KCl and 16 µg/ml BSA), the 32P-labeled helicase substrate (1000 cpm/10 µl, different substrate used for different helicase assays) and the purified Psp68 protein was incubated at 37 °C for 60 min. Reaction was performed for both concentration curve analysis (dif-ferent concentrations of Psp68 protein) and time course analysis [fixed concentration of Psp68 protein for different time duration (10–90 min)]. The reaction was terminated by the addition of 0.3 % SDS, 10 mM EDTA, 5 % glycerol and 0.03 % bromophenol blue. The substrate and products were separated by electrophoresis on a nondenaturing 12 or 15 % (for the blunt end substrate) PAGE, dried, and the gel was exposed to hyper film for autoradiography or scanned on phosphoimager and both the substrate and unwound DNA bands were quantified. The quantitation was done using IMAGE j/geldoc software (http://rsbweb.nih.gov/ij/) and the percent unwinding was plotted as the bar diagram.
Preparation of RNA helicase substrate and unwinding assay
Standard protocol to prepare RNA helicase substrate (RNA duplex) was adopted from previous report (Tarique et al. 2013). For RNA helicase assay, the RNA helicase substrate was prepared by using the RNA oligonucleo-tides synthesized from Primm srl (Milan, Italy): 13 mer
5′-AUAGCCUCAACC-G-3′ and 39 mer 5′-GGGAG AAAUCACUCGGUUGAGGCUAUCCGUAAAG CACGC-3′. About 10 ng of the 13-mer oligonucleotide was labeled at the 5′-end with five units of bacteriophage T4 polynucleotide kinase (NEB, England). The labeled oligo-nucleotide was then annealed with the 39-mer oligonucleo-tide and purified as described in previous section. The reac-tion setup for RNA helicase assay (concentration and time course analysis) was similar as DNA helicase as described in the DNA helicase assay section.
In vitro RNA binding assay
The RNA binding assay was done by a method previously described (Cheng et al. 2005; Shankar et al. 2008). Differ-ent amounts of Psp68 and BSA (1 μg) were dot-blotted on pre-charged PVDF membrane and then incubated in block-ing buffer containing 25 mM NaCl, 10 mM MgCl2, 10 mM HEPES, 0.1 mM EDTA, 1 mM DTT and 3 % BSA. The 13 mer RNA oligonucleotide which was used for the prepa-ration of RNA helicase substrate was labeled at the 5′-end using the standard procedure and T4 polynucleotide kinase (NEB, England) with 1.85 MBq of [γ-32P] ATP (specific activity222 TBq/mmol) and purified using Sepharose 4B (Pharmacia, Sweden) column chromatography. After blocking, the membrane was incubated for 2 h in binding buffer containing 10 pmol of 32P-labeled RNA oligonucleo-tide. After binding, the membrane was washed thrice with binding buffer and exposed for autoradiography. To check for loading of proteins, increasing amounts of Psp68 pro-tein was dot-blotted on another pre-charged PVDF mem-brane. This membrane was blocked with blocking buffer [2 % BSA in Tris buffered saline (TBS)] for 1 h at room temperature and further probed for another 1 h with HRP conjugated anti-his-antibody (Sigma Chemical Co., St. Louis, MO, USA) in the same buffer. The blot was washed and developed using standard protocol.
In vitro DNA binding assay
The same method described for RNA binding assay was used for the DNA-binding assay also but end-labeled DNA oligodeoxynucleotide of 32 bases with the sequence 5′-TTCGAGCTCGGTACCCGGGGATCCTCTAGAGT-3′ was used. BSA (1 μg) and different amounts of Psp68 protein was dot-blotted on pre-charged PVDF membrane and the membrane was incubated in blocking buffer as described in previous section. After blocking the mem-brane was incubated for 2 h in binding buffer containing 10 pmol of 32P-labeled DNA oligodeoxynucleotide. After binding, the membrane was washed thrice with binding buffer and exposed for autoradiography. To check for load-ing of proteins, increasing amounts of Psp68 protein was

Plant Mol Biol
1 3
dot-blotted on another pre-charged PVDF membrane. This membrane was blocked with blocking buffer (2 % BSA in TBS) for 1 h at room temperature and probed for further 1 h with HRP conjugated anti-his antibody (Sigma Chemi-cal Co., St. Louis, MO,USA) in the same buffer. The blot was washed and developed by using the standard protocol.
Results
Psp68 homolog possess DEAD-box helicase motifs and similar structure to its template
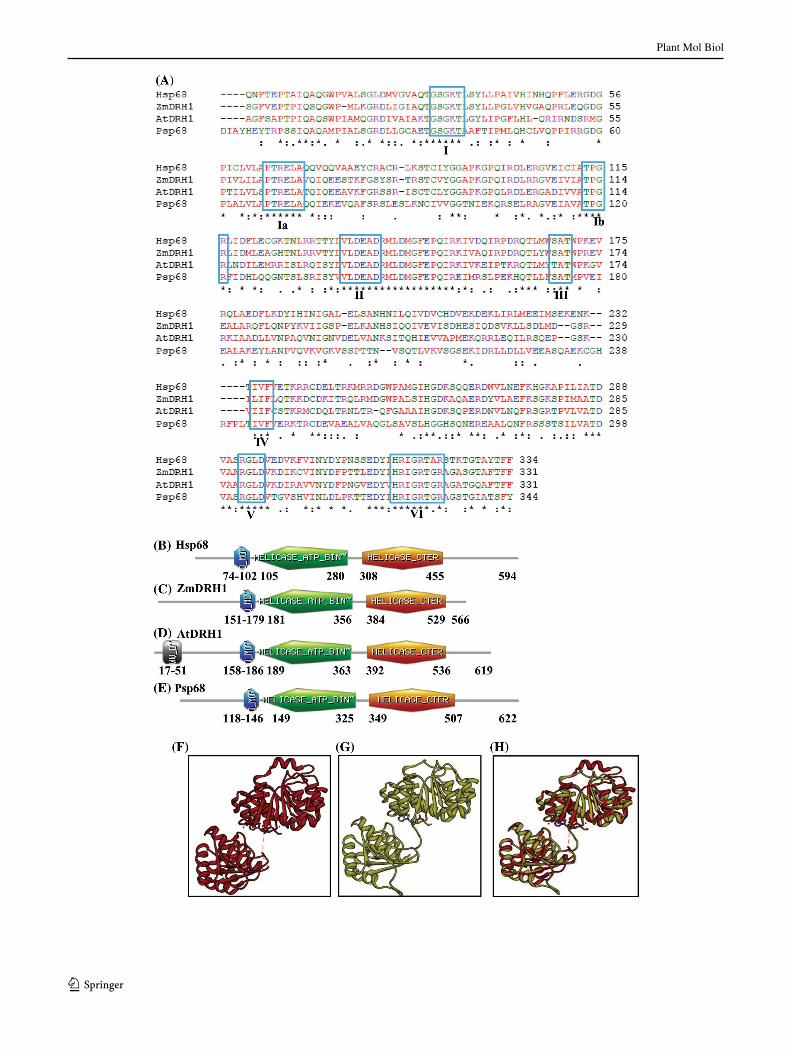
The sequence relationship of Psp68 to other DEAD-box helicases was studied by performing BLAST searches. It was observed that Psp68 is homologous to human p68 and some other plant helicases. The alignment of the core helicase domain showed that it showed ~47 % identity to human p68, AtDRH1 and ZmDRH1 (Zea mays DEAD-box RNA helicase 1) (Gendra et al. 2004) (Fig. 1a). It is interesting to note further that the terminal parts of the proteins are much less conserved but the SCAN prosite (www.expasy.org) analysis shows that all the four proteins contain the characteristic Q motif, helicase ATP-binding and helicase C-terminal domains (Fig. 1b–e). To get the insight of structure of Psp68, in silico approach was used. For structural modeling the sequence of full-length Psp68 was submitted to the Swissmodel homology-modeling server (http://swissmodel.expasy.org/). A total of four mod-els were obtained and one model which covered the areas ranging from 107 to 492 amino acids of Psp68 was stud-ied in detail. This model was built using human DEAD-box RNA helicase DDX3X as template (Hogbom et al. 2007). The structural modeling of the Psp68 was therefore done using the known crystal structure of this homologue as the template (PDB number 2i4i at http://www.rcsb.org/pdb/). The ribbon diagram of the template is shown in Fig. 1f and the predicted structure of Psp68 is shown in Fig. 1g. When the modeled structure of Psp68 and the template were superimposed, it is clear that these structures superimpose almost completely (Fig. 1h). Molecular graphics images were produced using the UCSF Chimera package (http://www.cgl.ucsf.edu/chimera) from the resource for biocomput-ing, visualization and informatics at the university of Cali-fornia, San Francisco (supported by NIH P41 RR-01081) (Pettersen et al. 2004).
Purification of recombinant p68 shows homogeneous preparation
The standard conditions for Psp68 expression are at 18 °C for 18 h following induction by 1.0 mM IPTG. Psp68
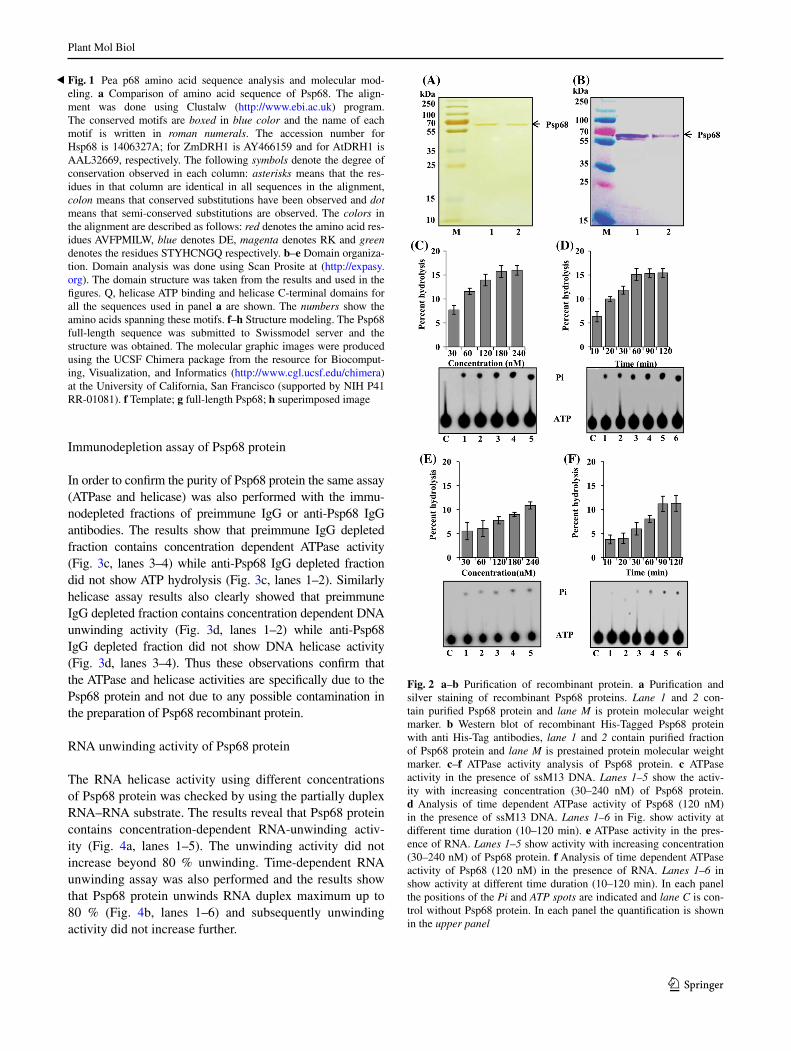
protein was purified under native conditions using Ni–NTA affinity chromatography. The SDS-PAGE analysis followed by silver staining of the purified proteins showed that Psp68 protein of ~68 kDa is pure and is a homogeneous preparation (Fig. 2a, lanes 1–2). The western blot analysis of purified Psp68 with anti his-tag antibody showed that the purified fractions contain only single bands (Fig. 2b, lanes 1–2) and this fraction was used for the biochemical characterizations.
DNA and RNA dependent ATPase activity of Psp68 protein
The ATPase activity of Psp68 protein was analyzed using standard assay conditions, purified Psp68 protein and the method described in the “Materials and methods” section. The percent release of radioactive phosphate (Pi) from [γ32P] ATP using different concentrations of Psp68 pro-tein is shown in Fig. 2c (lane 1–5). The results show that Psp68 protein contains concentration-dependent ATPase activity and hydrolyses up to a maximum of 15 % of ATP in the reaction (Fig. 2c, lanes 1–5). In another experiment steady state concentration (120 nM) of Psp68 protein was used for the time dependent ATPase activity analy-sis. The results of this analysis confirmed that Psp68 also shows characteristic time dependent ATPase activity up to 60 min while beyond this time the ATPase activity does not increase significantly (Fig. 2d, lanes 1–6). Thus it is clear from these observations that Psp68 is an ATPase. In addition to ATPase activity in the presence of DNA, Psp68 also shows slightly weak but concentration and time dependent ATPase activity (~10 %) in the presence of RNA (Fig. 2e, lanes 1–5 and Fig. 2f, lanes 1–6, respec-tively). Thus on the basis of results of ATPase activity of Psp68, it can be categorized among weak ATPase because it is able to hydrolyze maximum up to 15 % of ATP in the in vitro ATPase assay.
DNA unwinding activity of Psp68 protein
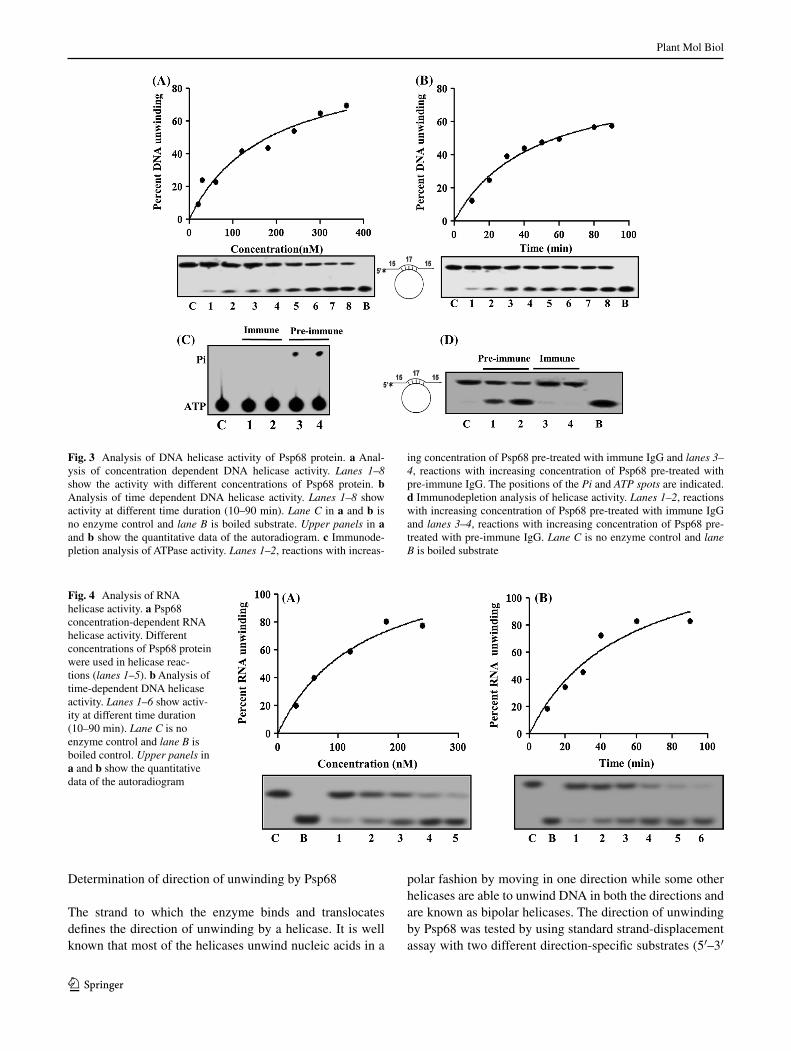
The helicase assay with purified Psp68 protein was per-formed to measure the unwinding of 32P labelled DNA fragment from a partially duplex DNA substrate. The reac-tion mixture containing helicase buffer, ATP, Psp68 protein and the substrate was incubated at 37 °C for 60 min. The substrate and products were separated by electrophoresis on non-denaturing polyacrylamide gel followed by auto-radiography. The helicase assay results with increasing Psp68 clearly show that it contains helicase activity and unwinds DNA maximum up to 60 % in vitro (Fig. 3a, lanes 1–8). The helicase activity from Psp68 is time dependent and optimum time for the helicase activity is ~80–90 min (Fig. 3b, lane 1–8).
Plant Mol Biol
1 3
Plant Mol Biol
1 3
Immunodepletion assay of Psp68 protein
In order to confirm the purity of Psp68 protein the same assay (ATPase and helicase) was also performed with the immu-nodepleted fractions of preimmune IgG or anti-Psp68 IgG antibodies. The results show that preimmune IgG depleted fraction contains concentration dependent ATPase activity (Fig. 3c, lanes 3–4) while anti-Psp68 IgG depleted fraction did not show ATP hydrolysis (Fig. 3c, lanes 1–2). Similarly helicase assay results also clearly showed that preimmune IgG depleted fraction contains concentration dependent DNA unwinding activity (Fig. 3d, lanes 1–2) while anti-Psp68 IgG depleted fraction did not show DNA helicase activity (Fig. 3d, lanes 3–4). Thus these observations confirm that the ATPase and helicase activities are specifically due to the Psp68 protein and not due to any possible contamination in the preparation of Psp68 recombinant protein.
RNA unwinding activity of Psp68 protein
The RNA helicase activity using different concentrations of Psp68 protein was checked by using the partially duplex RNA–RNA substrate. The results reveal that Psp68 protein contains concentration-dependent RNA-unwinding activ-ity (Fig. 4a, lanes 1–5). The unwinding activity did not increase beyond 80 % unwinding. Time-dependent RNA unwinding assay was also performed and the results show that Psp68 protein unwinds RNA duplex maximum up to 80 % (Fig. 4b, lanes 1–6) and subsequently unwinding activity did not increase further.
Fig. 1 Pea p68 amino acid sequence analysis and molecular mod-eling. a Comparison of amino acid sequence of Psp68. The align-ment was done using Clustalw (http://www.ebi.ac.uk) program. The conserved motifs are boxed in blue color and the name of each motif is written in roman numerals. The accession number for Hsp68 is 1406327A; for ZmDRH1 is AY466159 and for AtDRH1 is AAL32669, respectively. The following symbols denote the degree of conservation observed in each column: asterisks means that the res-idues in that column are identical in all sequences in the alignment, colon means that conserved substitutions have been observed and dot means that semi-conserved substitutions are observed. The colors in the alignment are described as follows: red denotes the amino acid res-idues AVFPMILW, blue denotes DE, magenta denotes RK and green denotes the residues STYHCNGQ respectively. b–e Domain organiza-tion. Domain analysis was done using Scan Prosite at (http://expasy.org). The domain structure was taken from the results and used in the figures. Q, helicase ATP binding and helicase C-terminal domains for all the sequences used in panel a are shown. The numbers show the amino acids spanning these motifs. f–h Structure modeling. The Psp68 full-length sequence was submitted to Swissmodel server and the structure was obtained. The molecular graphic images were produced using the UCSF Chimera package from the resource for Biocomput-ing, Visualization, and Informatics (http://www.cgl.ucsf.edu/chimera) at the University of California, San Francisco (supported by NIH P41 RR-01081). f Template; g full-length Psp68; h superimposed image
Fig. 2 a–b Purification of recombinant protein. a Purification and silver staining of recombinant Psp68 proteins. Lane 1 and 2 con-tain purified Psp68 protein and lane M is protein molecular weight marker. b Western blot of recombinant His-Tagged Psp68 protein with anti His-Tag antibodies, lane 1 and 2 contain purified fraction of Psp68 protein and lane M is prestained protein molecular weight marker. c–f ATPase activity analysis of Psp68 protein. c ATPase activity in the presence of ssM13 DNA. Lanes 1–5 show the activ-ity with increasing concentration (30–240 nM) of Psp68 protein. d Analysis of time dependent ATPase activity of Psp68 (120 nM) in the presence of ssM13 DNA. Lanes 1–6 in Fig. show activity at different time duration (10–120 min). e ATPase activity in the pres-ence of RNA. Lanes 1–5 show activity with increasing concentration (30–240 nM) of Psp68 protein. f Analysis of time dependent ATPase activity of Psp68 (120 nM) in the presence of RNA. Lanes 1–6 in show activity at different time duration (10–120 min). In each panel the positions of the Pi and ATP spots are indicated and lane C is con-trol without Psp68 protein. In each panel the quantification is shown in the upper panel
◂
Plant Mol Biol
1 3
Determination of direction of unwinding by Psp68
The strand to which the enzyme binds and translocates defines the direction of unwinding by a helicase. It is well known that most of the helicases unwind nucleic acids in a
polar fashion by moving in one direction while some other helicases are able to unwind DNA in both the directions and are known as bipolar helicases. The direction of unwinding by Psp68 was tested by using standard strand-displacement assay with two different direction-specific substrates (5′–3′
Fig. 3 Analysis of DNA helicase activity of Psp68 protein. a Anal-ysis of concentration dependent DNA helicase activity. Lanes 1–8 show the activity with different concentrations of Psp68 protein. b Analysis of time dependent DNA helicase activity. Lanes 1–8 show activity at different time duration (10–90 min). Lane C in a and b is no enzyme control and lane B is boiled substrate. Upper panels in a and b show the quantitative data of the autoradiogram. c Immunode-pletion analysis of ATPase activity. Lanes 1–2, reactions with increas-
ing concentration of Psp68 pre-treated with immune IgG and lanes 3–4, reactions with increasing concentration of Psp68 pre-treated with pre-immune IgG. The positions of the Pi and ATP spots are indicated. d Immunodepletion analysis of helicase activity. Lanes 1–2, reactions with increasing concentration of Psp68 pre-treated with immune IgG and lanes 3–4, reactions with increasing concentration of Psp68 pre-treated with pre-immune IgG. Lane C is no enzyme control and lane B is boiled substrate
Fig. 4 Analysis of RNA helicase activity. a Psp68 concentration-dependent RNA helicase activity. Different concentrations of Psp68 protein were used in helicase reac-tions (lanes 1–5). b Analysis of time-dependent DNA helicase activity. Lanes 1–6 show activ-ity at different time duration (10–90 min). Lane C is no enzyme control and lane B is boiled control. Upper panels in a and b show the quantitative data of the autoradiogram
Plant Mol Biol
1 3
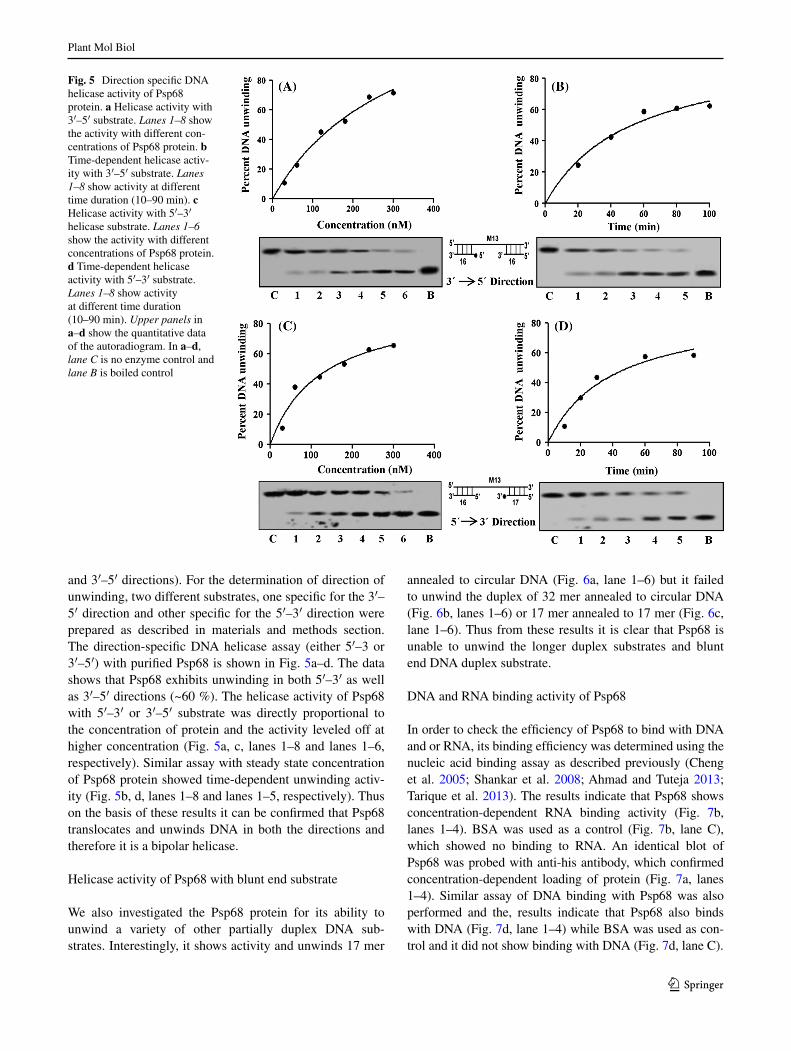
and 3′–5′ directions). For the determination of direction of unwinding, two different substrates, one specific for the 3′–5′ direction and other specific for the 5′–3′ direction were prepared as described in materials and methods section. The direction-specific DNA helicase assay (either 5′–3 or 3′–5′) with purified Psp68 is shown in Fig. 5a–d. The data shows that Psp68 exhibits unwinding in both 5′–3′ as well as 3′–5′ directions (~60 %). The helicase activity of Psp68 with 5′–3′ or 3′–5′ substrate was directly proportional to the concentration of protein and the activity leveled off at higher concentration (Fig. 5a, c, lanes 1–8 and lanes 1–6, respectively). Similar assay with steady state concentration of Psp68 protein showed time-dependent unwinding activ-ity (Fig. 5b, d, lanes 1–8 and lanes 1–5, respectively). Thus on the basis of these results it can be confirmed that Psp68 translocates and unwinds DNA in both the directions and therefore it is a bipolar helicase.
Helicase activity of Psp68 with blunt end substrate
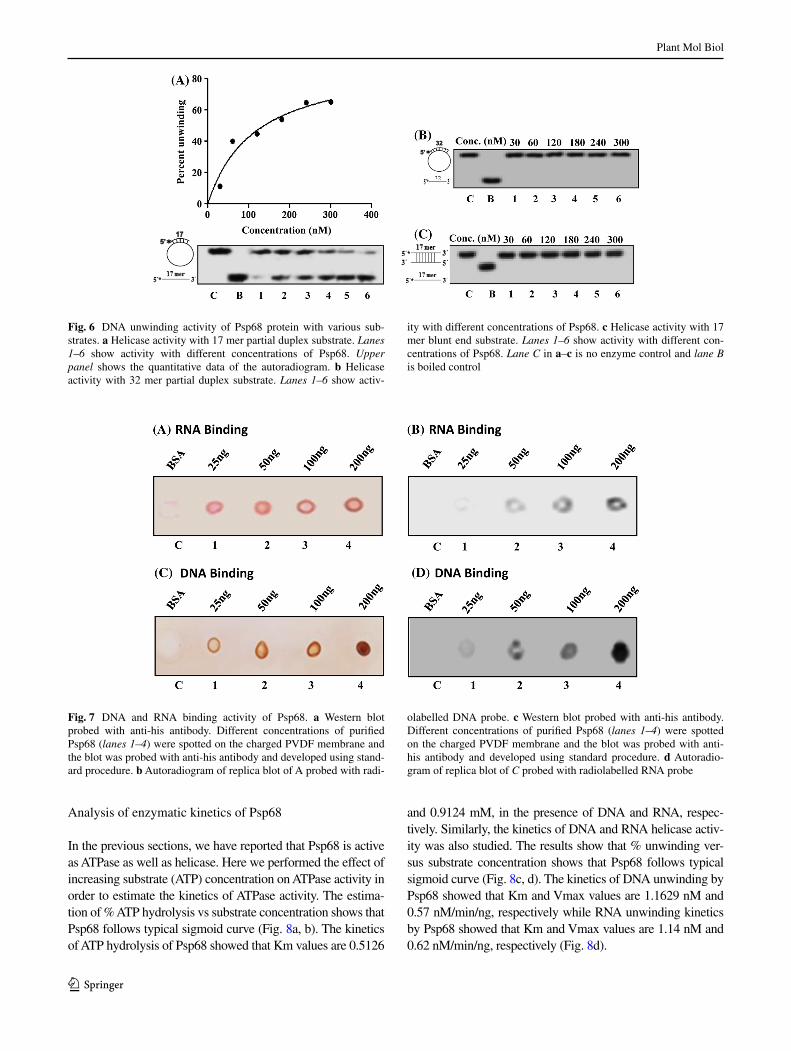
We also investigated the Psp68 protein for its ability to unwind a variety of other partially duplex DNA sub-strates. Interestingly, it shows activity and unwinds 17 mer
annealed to circular DNA (Fig. 6a, lane 1–6) but it failed to unwind the duplex of 32 mer annealed to circular DNA (Fig. 6b, lanes 1–6) or 17 mer annealed to 17 mer (Fig. 6c, lane 1–6). Thus from these results it is clear that Psp68 is unable to unwind the longer duplex substrates and blunt end DNA duplex substrate.
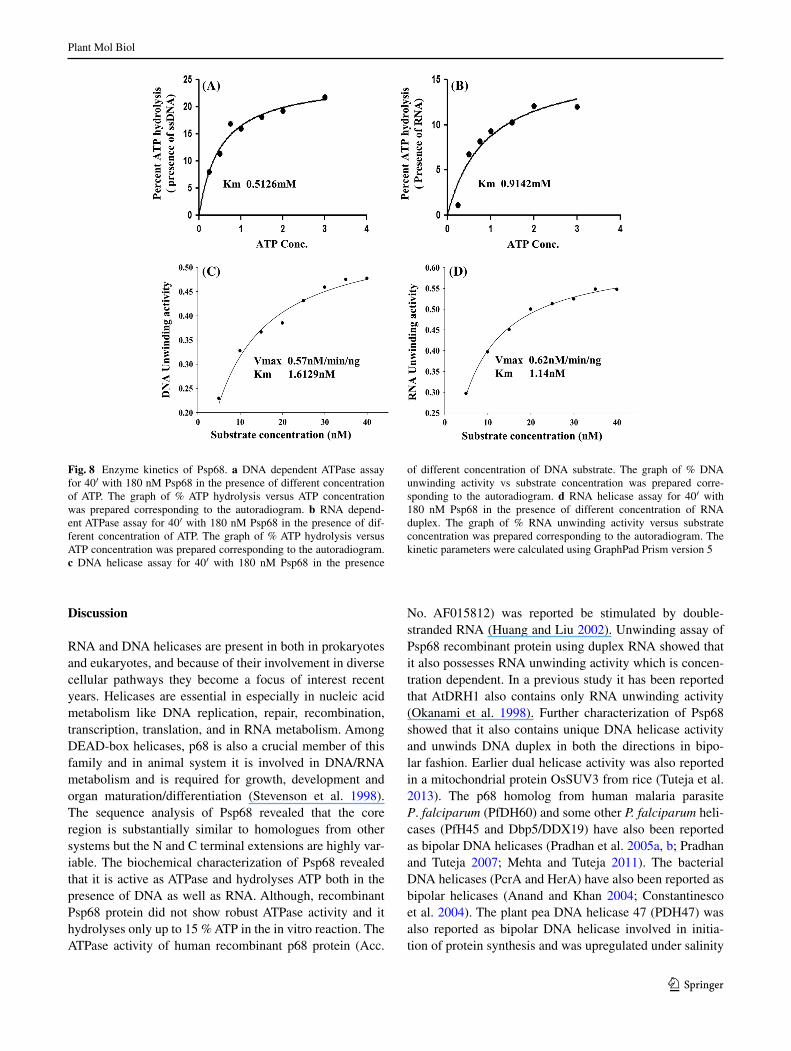
DNA and RNA binding activity of Psp68
In order to check the efficiency of Psp68 to bind with DNA and or RNA, its binding efficiency was determined using the nucleic acid binding assay as described previously (Cheng et al. 2005; Shankar et al. 2008; Ahmad and Tuteja 2013; Tarique et al. 2013). The results indicate that Psp68 shows concentration-dependent RNA binding activity (Fig. 7b, lanes 1–4). BSA was used as a control (Fig. 7b, lane C), which showed no binding to RNA. An identical blot of Psp68 was probed with anti-his antibody, which confirmed concentration-dependent loading of protein (Fig. 7a, lanes 1–4). Similar assay of DNA binding with Psp68 was also performed and the, results indicate that Psp68 also binds with DNA (Fig. 7d, lane 1–4) while BSA was used as con-trol and it did not show binding with DNA (Fig. 7d, lane C).
Fig. 5 Direction specific DNA helicase activity of Psp68 protein. a Helicase activity with 3′–5′ substrate. Lanes 1–8 show the activity with different con-centrations of Psp68 protein. b Time-dependent helicase activ-ity with 3′–5′ substrate. Lanes 1–8 show activity at different time duration (10–90 min). c Helicase activity with 5′–3′ helicase substrate. Lanes 1–6 show the activity with different concentrations of Psp68 protein. d Time-dependent helicase activity with 5′–3′ substrate. Lanes 1–8 show activity at different time duration (10–90 min). Upper panels in a–d show the quantitative data of the autoradiogram. In a–d, lane C is no enzyme control and lane B is boiled control
Plant Mol Biol
1 3
Analysis of enzymatic kinetics of Psp68
In the previous sections, we have reported that Psp68 is active as ATPase as well as helicase. Here we performed the effect of increasing substrate (ATP) concentration on ATPase activity in order to estimate the kinetics of ATPase activity. The estima-tion of % ATP hydrolysis vs substrate concentration shows that Psp68 follows typical sigmoid curve (Fig. 8a, b). The kinetics of ATP hydrolysis of Psp68 showed that Km values are 0.5126
and 0.9124 mM, in the presence of DNA and RNA, respec-tively. Similarly, the kinetics of DNA and RNA helicase activ-ity was also studied. The results show that % unwinding ver-sus substrate concentration shows that Psp68 follows typical sigmoid curve (Fig. 8c, d). The kinetics of DNA unwinding by Psp68 showed that Km and Vmax values are 1.1629 nM and 0.57 nM/min/ng, respectively while RNA unwinding kinetics by Psp68 showed that Km and Vmax values are 1.14 nM and 0.62 nM/min/ng, respectively (Fig. 8d).
Fig. 6 DNA unwinding activity of Psp68 protein with various sub-strates. a Helicase activity with 17 mer partial duplex substrate. Lanes 1–6 show activity with different concentrations of Psp68. Upper panel shows the quantitative data of the autoradiogram. b Helicase activity with 32 mer partial duplex substrate. Lanes 1–6 show activ-
ity with different concentrations of Psp68. c Helicase activity with 17 mer blunt end substrate. Lanes 1–6 show activity with different con-centrations of Psp68. Lane C in a–c is no enzyme control and lane B is boiled control
Fig. 7 DNA and RNA binding activity of Psp68. a Western blot probed with anti-his antibody. Different concentrations of purified Psp68 (lanes 1–4) were spotted on the charged PVDF membrane and the blot was probed with anti-his antibody and developed using stand-ard procedure. b Autoradiogram of replica blot of A probed with radi-
olabelled DNA probe. c Western blot probed with anti-his antibody. Different concentrations of purified Psp68 (lanes 1–4) were spotted on the charged PVDF membrane and the blot was probed with anti-his antibody and developed using standard procedure. d Autoradio-gram of replica blot of C probed with radiolabelled RNA probe
Plant Mol Biol
1 3
Discussion
RNA and DNA helicases are present in both in prokaryotes and eukaryotes, and because of their involvement in diverse cellular pathways they become a focus of interest recent years. Helicases are essential in especially in nucleic acid metabolism like DNA replication, repair, recombination, transcription, translation, and in RNA metabolism. Among DEAD-box helicases, p68 is also a crucial member of this family and in animal system it is involved in DNA/RNA metabolism and is required for growth, development and organ maturation/differentiation (Stevenson et al. 1998). The sequence analysis of Psp68 revealed that the core region is substantially similar to homologues from other systems but the N and C terminal extensions are highly var-iable. The biochemical characterization of Psp68 revealed that it is active as ATPase and hydrolyses ATP both in the presence of DNA as well as RNA. Although, recombinant Psp68 protein did not show robust ATPase activity and it hydrolyses only up to 15 % ATP in the in vitro reaction. The ATPase activity of human recombinant p68 protein (Acc.
No. AF015812) was reported be stimulated by double-stranded RNA (Huang and Liu 2002). Unwinding assay of Psp68 recombinant protein using duplex RNA showed that it also possesses RNA unwinding activity which is concen-tration dependent. In a previous study it has been reported that AtDRH1 also contains only RNA unwinding activity (Okanami et al. 1998). Further characterization of Psp68 showed that it also contains unique DNA helicase activity and unwinds DNA duplex in both the directions in bipo-lar fashion. Earlier dual helicase activity was also reported in a mitochondrial protein OsSUV3 from rice (Tuteja et al. 2013). The p68 homolog from human malaria parasite P. falciparum (PfDH60) and some other P. falciparum heli-cases (PfH45 and Dbp5/DDX19) have also been reported as bipolar DNA helicases (Pradhan et al. 2005a, b; Pradhan and Tuteja 2007; Mehta and Tuteja 2011). The bacterial DNA helicases (PcrA and HerA) have also been reported as bipolar helicases (Anand and Khan 2004; Constantinesco et al. 2004). The plant pea DNA helicase 47 (PDH47) was also reported as bipolar DNA helicase involved in initia-tion of protein synthesis and was upregulated under salinity
Fig. 8 Enzyme kinetics of Psp68. a DNA dependent ATPase assay for 40′ with 180 nM Psp68 in the presence of different concentration of ATP. The graph of % ATP hydrolysis versus ATP concentration was prepared corresponding to the autoradiogram. b RNA depend-ent ATPase assay for 40′ with 180 nM Psp68 in the presence of dif-ferent concentration of ATP. The graph of % ATP hydrolysis versus ATP concentration was prepared corresponding to the autoradiogram. c DNA helicase assay for 40′ with 180 nM Psp68 in the presence
of different concentration of DNA substrate. The graph of % DNA unwinding activity vs substrate concentration was prepared corre-sponding to the autoradiogram. d RNA helicase assay for 40′ with 180 nM Psp68 in the presence of different concentration of RNA duplex. The graph of % RNA unwinding activity versus substrate concentration was prepared corresponding to the autoradiogram. The kinetic parameters were calculated using GraphPad Prism version 5

Plant Mol Biol
1 3
and cold stress (Vashisht et al. 2005). The bipolar helicase activity of the stress-induced plant helicases including Psp68 might be involved in different cellular processes in various conditions. Further biochemical characterization of Psp68 revealed that it failed to show unwinding with longer (32 mer) duplex substrate and blunt end duplex substrate. Some biological roles of p68 protein have been explored in the animal system which includes its interaction with CBP/p300, etc., and acts as a transcriptional coactivator (Endoh et al. 1999; Rossow and Janknecht 2003; Wilson et al. 2004). The p68 homologue Dbp2 is known to be important for rRNA processing and nonsense-mediated mRNA decay (Fuller-Pace 2006).
As far as plant p68 protein is concerned, it was first reported in Nicotiana sylvestris and transcript of the gene was observed in different tissues like leaves, roots and flowers (Itadani et al. 1994). The transcript of Arabidopsis p68 (AtDRH1) and pea p68 (Psp68) was found to be almost equally distributed in every tissues of the Arabidopsis and pea plants (Okanami et al. 1998; Tuteja et al. 2014). Recent reports showed that the expression of Psp68 was up-regu-lated in response to salinity, cold, ABA and heat stress and the expression was more in aerial parts as compared to the roots (Tuteja et al. 2014; Banu et al. 2014a). Furthermore, the overexpression of Psp68 provides salinity stress tol-erance in tobacco and rice by improving photosynthesis machinery as well as by scavenging of ROS-mediated H2O2 and by interacting with argonaute (Tuteja et al. 2014; Banu et al. 2014a). The pea p68 has also been shown to self-interact (Tuteja et al. 2014) and interact with many other proteins including Calmodulin (CaM) and argonaute (Banu et al. 2014a). The interaction of Psp68 with Ca2+–CaM might be regulating diverse signalling pathways leading to stress tolerance (Banu et al. 2014a). The overexpression of Psp68 was also reported to regulate many transcripts involved in the abiotic and oxidative stress responses in rice and therefore it functions as a molecular switch in differ-ent signalling pathways leading to stress tolerance (Banu et al. 2014a). Recently, the stress-inducible nature of Psp68 promoter has been reported and the promoter contains salt, dehydration, cold, auxin ABA and MeJA related cis-elements, which drive the expression of the reporter gene in response to the stress and plant hormones (Banu et al. 2014b).
The variable N and C terminal extension of Psp68 sug-gest that this protein is endowed with specialized function. Thus it seems that p68 protein acts as a multifunctional protein. Since RNA molecules are more prone to forming stable non-functional secondary structures, therefore, their proper functioning requires RNA chaperones (Jones et al. 1996). DEAD-box RNA helicases are the best candidates for RNA chaperones because these proteins can use energy derived from ATP hydrolysis to actively disrupt misfolded
RNA structures so that correct folding can occur (Jones et al. 1996). Since Psp68 exhibits RNA helicase activity, therefore, it may act as RNA chaperones especially dur-ing stress condition in plants where secondary structures in RNA are common. We have shown that DEAD-box Psp68 protein exhibits unique properties of possessing dual heli-case and bipolar DNA helicase activities, which suggest that it could be a multifunctional protein involved in diverse processes including DNA and RNA metabolism. We specu-late that this study provides valuable information for better understanding of nucleic acids metabolism in plants.
Acknowledgments Work on plant helicases and abiotic stress toler-ance in NT’s laboratory is partially supported by Department of Bio-technology (DBT) and ICGEB, New Delhi. NT is thankful to Dr. Hoi Xuan Pham for his help in the initial stage of the work. We do not have any conflict of interest to declare.
References
Ahmad M, Tuteja R (2013) Plasmodium falciparum RuvB2 translo-cates in 5′–3′ direction, relocalizes during schizont stage and its enzymatic activities are up regulated by RuvB3 of the same com-plex. Biochim Biophys Acta 1834:2795–2811
Ahmad M, Ansari A, Tarique M, Satsangi AT, Tuteja R (2012) Plas-modium falciparum UvrD helicase translocates in 3′ to 5′ direc-tion, colocalizes with MLH and modulates its activity through physical interaction. PLoS One 7:e49385
Anand SP, Khan SA (2004) Structure-specific DNA binding and bipo-lar helicase activities of PcrA. Nucleic Acids Res 32:3190–3197
Arnold K, Bordoli L, Kopp J, Schwede T (2006) The SWISS-MODEL workspace: a web-based environment for protein struc-ture homology modelling. Bioinformatics 22:195–201
Banu MSA, Huda KMK, Shaoo RK, Garg B, Tula S, Islam SMS, Tuteja R, Tuteja N (2014a) Pea p68 imparts salinity stress tolerance in rice by scavenging of ROS-mediated H2O2 and interacts with argonaute. Plant Mol Biol Rep. doi:10.1007/s11105-014-0748-7
Banu MSA, Huda KMK, Tuteja N (2014b) Isolation and functional characterization of the promoter of a DEAD-box helicase Psp68 using Agrobacterium-mediated transient assay. Plant Signal Behav 9:e28992
Bates GJ, Nicol SM, Wilson BJ, Jacobs AM, Bourdon JC, Wardrop J, Gregory DJ, Lane DP, Perkins ND, Fuller-Pace FV (2005) The DEAD box protein p68: a novel transcriptional coactivator of the p53 tumour suppressor. EMBO J 24:543–553
Buszczak M, Spradling AC (2006) The Drosophila P68 RNA helicase regulates transcriptional deactivation by promoting RNA release from chromatin. Genes Dev 20:977–989
Cheng Z, Coller J, Parker R, Song H (2005) Crystal structure and functional analysis of DEAD-box protein Dhh1p. RNA 11:1258–1270
Constantinesco F, Forterre P, Koonin EV, Aravind L, Elie CA (2004) Bipolar DNA helicase gene, herA, clusters with rad50, mre11 and nurA genes in thermophilic archaea. Nucleic Acids Res 32:1439–1447
Cordin O, Banroques J, Tanner NK, Linder P (2006) The DEAD-box protein family of RNA helicases. Gene 367:17–37
Endoh H, Maruyama K, Masuhiro Y, Kobayashi Y, Goto M, Tai H, Yanagisawa J, Metzger D, Hashimoto S, Kato S (1999) Puri-fication and identification of p68 RNA helicase acting as a

Plant Mol Biol
1 3
transcriptional coactivator specific for the activation function 1 of human estrogen receptor alpha. Mol Cell Biol 19:5363–5372
Fuller-Pace FV (2006) DExD/H box RNA helicases multifunc-tional proteins with important roles in transcriptional regulation. Nucleic Acids Res 34:4206–4215
Fuller-Pace FV (2013) The DEAD box proteins DDX5 (p68) and DDX17 (p72): multi-tasking transcriptional regulators. Biochim Biophys Acta 1829:756–763
Gendra E, Moreno A, Alba MM, Pages M (2004) Interaction of the plant glycine-rich RNA-binding protein MA16 with a novel nucleolar DEAD box RNA helicase protein from Zea mays. Plant J 38:875–886
Gong Z, Dong CH, Lee H, Zhu J, Xiong L, Gong D, Stevenson B, Zhu JK (2005) A DEAD box RNA helicase is essential for mRNA export and important for development and stress responses in Arabidopsis. Plant Cell 17:256–267
Guan Q, Wu J, Zhang Y, Jiang C, Liu R, Chai C, Zhu J (2013) A DEAD box RNA helicase is critical for pre-mRNA splicing, cold-responsive gene regulation, and cold tolerance in Arabidop-sis. Plant Cell 25:342–356
Hogbom M, Collins R, van den Berg S, Jenvert RM, Karlberg T, Kotenyova T, Flores A, Karlsson Hedestam GB, Schiavone LH (2007) Crystal structure of conserved domains 1 and 2 of the human DEAD-box helicase DDX3X in complex with the mono-nucleotide AMP. J Mol Biol 372:150–159
Huang Y, Liu ZR (2002) The ATPase, RNA unwinding, and RNA binding activities of recombinant p68 RNA helicase. J Biol Chem 277:12810–12815
Iggo RD, Lane DP (1989) Nuclear protein p68 is an RNA-dependent ATPase. EMBO J 8:1827
Itadani H, Sugita M, Sugiura M (1994) Structure and expression of a cDNA encoding an RNA helicase-like protein in tobacco. Plant Mol Biol 24:249–252
Jones PG, Mitta M, Kim Y, Jiang W, Inouye M (1996) Cold shock induces a major ribosomal-associated protein that unwinds dou-ble stranded RNA in Escherichia coli. Proc Natl Acad Sci USA 93:76–80
Kammel C, Thomaier M, Sorensen BB, Schubert T, Langst G, Grasser M, Grasser KD (2013) Arabidopsis DEAD-box RNA helicase UAP56 interacts with both RNA and DNA as well as with mRNA export factors. PLoS One 8:e60644
Linder P, Jankowsky E (2011) From unwinding to clamping—the DEAD box RNA helicase family. Nat Rev Mol Cell Biol 12:505–516
Mehta J, Tuteja R (2011) A novel dual Dbp5/DDX19 homologue from Plasmodium falciparum requires Q motif for activity. Mol Biochem Parasitol 176:58–63
Okanami M, Meshi T, Iwabuchi M (1998) Characterization of a DEAD box ATPase/RNA helicase protein of Arabidopsis thali-ana. Nucleic Acids Res 26:2638–2643
Owttrim GW (2006) RNA helicases and abiotic stress. Nucleic Acids Res 34:3220–3230
Pettersen EF, Goddard TD, Huang CC, Couch GS, Greenblatt DM, Meng EC, Ferrin TE (2004) UCSF Chimera—a visualization system for exploratory research and analysis. J Comput Chem 25:1605–1612
Pradhan A, Tuteja R (2007) Bipolar, dual Plasmodium falciparum helicase 45 expressed in the intraerythrocytic developmental cycle is required for parasite growth. J Mol Biol 373:268–281
Pradhan A, Chauhan VS, Tuteja R (2005a) A novel ‘DEAD-box’ DNA helicase from Plasmodium falciparum is homologous to p68. Mol Biochem Parasitol 140:55–60
Pradhan A, Chauhan VS, Tuteja R (2005b) Plasmodium falciparum DNA helicase 60 is a schizont stage specific, bipolar and dual helicase stimulated by PKC phosphorylation. Mol Biochem Para-sitol 144:133–141
Rossow KL, Janknecht R (2003) Synergism between p68 RNA heli-case and the transcriptional coactivators CBP and p300. Onco-gene 22:151–156
Shankar J, Pradhan A, Tuteja R (2008) Isolation and characterization of Plasmodium falciparum UAP56 homolog evidence for the coupling of RNA binding and splicing activity by site-directed mutations. Arch Biochem Biophys 478:143–153
Stevenson RJ, Hamilton SJ, MacCallum DE, Hall PA, Fuller-Pace FV (1998) Expression of the ‘dead box’ RNA helicase p68 is devel-opmentally and growth regulated and correlates with organ dif-ferentiation/maturation in the fetus. J Pathol 184:351–359
Tarique M, Ahmad M, Ansari A, Tuteja R (2013) Plasmodium falcipa-rum DOZI, an RNA helicase interacts with eIF4E. Gene 522:46–59
Tuteja N (2003) Plant DNA helicases: the long unwinding road. J Exp Bot 54:2201–2214
Tuteja R, Pradhan A (2006) Unraveling the ‘DEAD-box’ helicases of Plasmodium falciparum. Gene 376:1–12
Tuteja N, Tuteja R (2004a) Prokaryotic and eukaryotic DNA heli-cases. Essential molecular motor proteins for cellular machinery. Eur J Biochem 271:1835–1848
Tuteja N, Tuteja R (2004b) Unraveling DNA helicases. Motif, struc-ture, mechanism and function. Eur J Biochem 271:1849–1863
Tuteja N, Sahoo RK, Garg B, Tuteja N (2013) OsSUV3 dual helicase functions in salinity stress tolerance by maintaining photosynthe-sis and antioxidant machinery in rice (Oryza sativa L. cv. IR64). Plant J 76:115–127. doi:10.1111/tpj.12277
Tuteja N, Banu MSA, Huda KMK, Gill SS, Jain P, Pham XH, Tuteja R (2014) Pea p68, a DEAD-box helicase, provides salinity stress tolerance in transgenic tobacco by reducing oxidative stress and improving photosynthesis machinery. PLOS One 9(5):e98287. doi:10.1371/journal.pone.0098287
Vashisht AA, Tuteja N (2006) Stress responsive DEAD-box helicases: a new pathway to engineer plant stress tolerance. J Photochem Photobiol B 84:150–160
Vashisht AA, Pradhan A, Tuteja R, Tuteja N (2005) Cold- and salinity stress-induced bipolar pea DNA helicase 47 is involved in protein synthesis and stimulated by phosphorylation with protein kinase C. Plant J 44:76–87
Wang H, Gao X, Huang Y, Yang J, Liu ZR (2009) P68 RNA helicase is a nucleocytoplasmic shuttling protein. Cell Res 19:1388–1400
Wang H, Gao X, Yang JJ, Liu ZR (2013) Interaction between p68 RNA helicase and Ca2+–calmodulin promotes cell migration and metastasis. Nat Commun 4:1354
Wilson BJ, Bates GJ, Nicol SM, Gregory DJ, Perkins ND, Fuller-Pace FV (2004) The p68 and p72 DEAD box RNA helicases interact with HDAC1 and repress transcription in a promoter-specific manner. BMC Mol Biol 5:11. doi:10.1186/1471-2199-5-11
Yang T, Poovaiah BW (2003) Calcium/calmodulin-mediated signal network in plants. Trends Plant Sci 8:505–512
Yang L, Lin C, Liu ZR (2005) Signaling to the DEAD box—regu-lation of DEAD-box p68 RNA helicase by protein phosphoryla-tions. Cell Signal 17:1495–1504
Related Documents











